Embed Size (px)
Citation preview

University of Groningen
Alteration of Receptor/G-protein Interaction by Putative Endogenous Protein Kinase Activity inDictyostelium discoideum Membranesvan Haastert, Petrus
Published in:Default journal
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:1987
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Haastert, P. J. M. V. (1987). Alteration of Receptor/G-protein Interaction by Putative Endogenous ProteinKinase Activity in Dictyostelium discoideum Membranes. Default journal.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 11-02-2018

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1987 by The American Society of Biological Chemists, Inc
Vol .262, No. 7, Issue of ’ March 5, pp. 3239-3243,1987 Printed in U. S. A.
Alteration of Receptor/G-protein Interaction by Putative Endogenous Protein Kinase Activity in Dictyostelium discoideum Membranes*
(Received for publication, April 1, 1986)
Peter J. M. Van Haastert From the Cell Biology and Morphogenesis Unit, Zoological Laboratory, 2300 RA Leiden, The Netherlands
Membranes of Dictyostelium discoideum cells were incubated under phosphorylation conditions and washed, and the effects on cAMP binding to chemotac- tic receptors in the absence and presence of guanosine 5’-0-(3-thiotriphosphate) (GTPrS) were investigated. Most experiments were done with adenosine 5’-0-(3- thiotriphosphate) (ATPrS), which is a good substrate for many kinases, but the product, protein phosphoro- thioate, is not easily hydrolyzed by phosphatases.
Pretreatment of membranes under phosphorylating conditions with MgATPrS alters the site heterogeneity of the CAMP-binding forms, without a significant ef- fect on the total number of binding sites. A similar effect was induced by GTPrS under nonphosphoryla- tion conditions.
The effects of MgATPrS were rapid (tlk = 1 min), irreversible, and not induced by M$+ or ATPrS alone or by magnesium adenylyl imidodiphosphate and mag- nesium adenylyl (8,r-methy1ene)diphosphate.
MgATP induced a smaller inhibition than MgATPrS, which was potentiated by the addition of exogenous CAMP-dependent protein kinase. The effect of MgATP was rapidly reversible; reversibility was reduced by the phosphatase inhibitor NaF. These results suggest that the effects of MgATPrS are mediated by an en- dogenous protein kinase.
The major 35S-thiophosphorylated band detected by sodium dodecyl sulfate-polyacrylamide gel electropho- resis was a protein with M, = 36,000. The phospho- rylation of a protein with the molecular weight of the cAMP receptor (Mr = 40,000-45,000) was not ob- served.
Extracellular cAMP functions as a signal molecule in Dic- tyostelium discoideum during chemotaxis (I), morphogenesis (2), and cell differentiation (3). cAMP is detected by highly specific surface receptors, which results in several cellular responses such as the activation of adenylate and guanylate cyclase (these and other responses have been reviewed (4-6)). The stimulation of guanylate and adenylate cyclase termi- nates within a few seconds and a few minutes, respectively, even when cAMP remains present a t constant levels (7-10). Desensitization of adenylate cyclase stimulation has been studied extensively by Dinauer et al. (11, 12).
Recent results (13, 14) suggest the presence of two subpop- ulations of cell surface cAMP receptors, A and B sites, that
* This work was supported by the C. and C. Huygens Fund which is subsidized by the Netherlands Organization for the Advancement of Pure Scientific Research (Zuiver Wetenschappelijk Onderzoek). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
” - _____
have been implicated in the stimulation of adenylate and guanylate cyclase, respectively. Binding of cAMP to both subpopulations is complex, showing interconversions of bind- ing states in vivo (15, 16) which are promoted by guanine nucleotides i n uitro (16-19). This may suggest the involve- ment of guanine nucleotide regulatory proteins in the trans- duction pathways of adenylate and guanylate cyclase. This view is further supported by the recent observations (20, 21) that guanosine triphosphates stimulate adenylate cyclase i n uitro.
Desensitization of CAMP-stimulated guanylate cyclase oc- curs within a few seconds after cAMP addition (9) and may be caused by an impairment of receptor/G-protein interaction (16). Desensitization of adenylate cyclase is much slower and requires the presence of constant cAMP concentrations dur- ing several minutes (12). This desensitization process may be caused by a receptor modification, presumably by phospho- rylation (22-24). cAMP also induces the loss of detectable CAMP-binding sites (down-regulation) which occurs with sim- ilar kinetics as desensitization of adenylate cyclase, but which requires 10-fold higher cAMP concentrations and recovers more slowly (25).’
The role of phosphorylation of signal transducing compo- nents in desensitization has been extensively studied in ver- tebrates (26-33). These studies make use of the well-charac- terized functional coupling between receptors, G-proteins, and adenylate cyclase in native membranes or after reconstitution of purified components in artificial membranes. It has been shown that phosphorylation of receptor or G-protein dimin- ishes their functional interaction (29-31).
Conditions for the functional reconstitution of isolated receptor, G-protein(s), and adenylate cyclase have not yet been found for the components from D. discoideum. There- fore, a study of the role of phosphorylation in transmembrane signaling in this organism is limited to experiments i n uiuo (22-24) or to receptor/G-protein/adenylate cyclase interac- tion in isolated membranes. In this study, the receptor/G- protein interaction was investigated by detecting the effects of guanine nucleotides on cAMP binding to membranes that were preincubated under conditions where protein kinases are active. The results show that phosphorylation conditions alter the receptor/G-protein interaction, and evidence is presented which suggests that this is due to an endogenous protein kinase.
EXPERIMENTAL PROCEDURES
M~terzals-[2,8-~H]cAMP (1.5 TBq/mmol), [y-32P]ATP (115 TBq/mmol), [y-35S]ATPyS2 (24 TBq/mmol), and carrier-free or-
’ P. J. M. Van Haastert, manuscript in preparation. The abbreviations used are: ATP+$ adenosine 5’-0-(3-thiotri-
phosphate); GTPyS, guanosine 5’-0-(3-thiotriphosphate); AppNHp, adenylyl imidodiphosphate; AppCHp, adenylyl @,y-methy1ene)di- phosphate; HEPES, N-2-hydroxyethylpiperazine-N’-2-ethanesul- fonic acid SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
3239
at University of G
roningen on August 23, 2007
ww
w.jbc.org
Dow
nloaded from

3240 Protein Kinase and ReceptorlG-protein Interaction in D. discoideum
th~[~'P]phosphate were obtained from Amersham Cop. CAMP, ATP, ATPyS, AppNHp, AppCHp, and GTPyS were from Boehringer Mannheim. Dithiothreitol was purchased from Sigma; CAMP-de- pendent protein kinase type I was isolated from bovine muscle by DEAE-cellulose chromatography as described (34).
Culture Conditions and Membrane Isolation-D. discoideum cells (strain NC-4) was grown on a buffered glucose/peptone medium (9). Cells were harvested in 10 mM KH,PO,/Na,HPO,, pH 6.5 (buffer A), freed from bacteria by repeated centrifugations at 100 X g for 2 min, and starved in buffer A by shaking at a density of lo7 cells/ml. After 5-6 h, cells were collected by centrifugation and washed twice with buffer A, and the pellet was resuspended in buffer B (40 mM HEPES/ NaOH, 0.5 mM EDTA, 250 mM sucrose, pH 7.7) to a density of 10' cells/ml. Cells were lysed by pressing them through 3-pm pores of a Nuclepore filter (35). The lysate was centrifuged at 10,000 X g for 5 min, the pellet was washed once with buffer B, and the final pellet was resuspended in buffer B to the equivalent of 2 X 10' cells/ml.
Incubation with ATP and Analogs-Membranes (300 pl) were incubated for 5 min at 20 "C with 3 pl of 100 X concentrated solutions yielding final concentrations of 10 mM NaF, 5 mM MgC12, and 1 mM ATP or one of its analogs. In one experiment, membranes were incubated in the presence of CAMP-dependent protein kinase. This enzyme (1 pmol of holoenzyme in 10 pl) was preactivated with 3 p~ (S,)-CAMPS at 20 "C for 15 min; the amount of enzyme was estimated from its cAMP binding activity as described (34).
Incubations were terminated by addition of 1200 pl of ice-cold buffer A and immediately centrifuged at 0 "C for 2 min at 10,000 X g. The pellet was washed once with 1 ml of buffer A, and the final pellet was resuspended in 600 p1 of buffer A.
CAMP Binding Assay-CAMP binding was measured in a total volume of 100 p1 containing buffer A, 5 nM t3H]cAMP, 10 mM dithiothreitol, GTPyS (30 p~ when present), and 80-pl membranes. The incubation period was 5 min a t 0 "C. Samples were centrifuged for 3 min at 10,000 X g, the supernatant was aspirated, and the pellet was dissolved in 75 p1 of 1 M acetic acid. Radioactivity was determined by liquid scintillation counting after the addition of 1.4 ml of Emul- sifier (Packard Instrument Co.). Nonspecific binding was determined in the presence of 1 mM cAMP and was subtracted from all data shown. Typical binding data for control membranes at 5 nM [3H] cAMP are input = 18,000 cpm, specific binding = 3,058 +- 112 cpm, and nonspecific binding = 186 * 21 cpm (both n = 3).
Scatchard analysis of CAMP-binding (Fig. 1) was determined by incubating membranes with different cAMP concentrations at 20 "C. The binding reaction was terminated by centrifugation of membranes through silicon oil (16).
"S-Thiophosphorylation and 32P-Phosphorylation in Vitro-"em- branes (100 pl) were incubated for 5 min at 20 "C in buffer B with 5 mM MgC12, 10 mM NaF, and 1 p~ [y-35S]ATPyS or [y-32P]ATP (4 TBq/mmol, 10 pCi/incubation). The reaction was terminated by the addition of 25 p1 of a mixture yielding 2% SDS, 2.5% dithiothreitol, and 10% glycine (w/v, final concentrations; sample buffer); SDS- PAGE was carried out as described (22) on a 10% acrylamide and 0.05% bisacrylamide gel. Gels were run at 50 V for 16 h; protein was stained with Coomassie Blue; and the gel was incubated with EN- HANCE (New England Nuclear), dried, and autoradiographed.
Phosphorylation of Receptor in Viuo-Aggregative cells were har- vested in 15 mM Tris/HCI, pH 7.0, washed twice, and resuspended in this buffer at a density of 10' cells/ml. Cells (200 p l ) were incubated for 30 min with 1 mCi/ml orth~[~~P]phosphate and 5 mM caffeine. Then, the suspension was divided into two equal portions. One portion was stimulated with 10 mM dithiothreitol and 1 p M CAMP; the other portion was not stimulated. After 15 min, the reactions were terminated by the addition of 1 ml of 95% saturated ammonium sulfate, and the suspension was incubated for 5 min at 0 "C and centrifuged for 5 min at 10,000 X g. The pellet was washed once with 1 ml of 95% saturated ammonium sulfate, and the final pellet was resuspended in 1 ml of receptor buffer (22), which induced cell lysis. Membranes were centrifuged for 15 min at 10,000 X g, and the pellet was resuspended in 100 pl of sample buffer. SDS-PAGE was carried out as described in the previous section.
RESULTS
The aim of the present study was to investigate the effects of endogenous protein kinase activity on cell-surface CAMP receptors and on the interaction between receptor and puta- tive G-protein(s). D. discoideum cells contain high levels of
protein phosphophosphatase activity (36). To by-pass this phosphatase activity, a derivative of ATP was used, ATPyS, which is a relatively good substrate of many protein kinases. However, the product, protein phosphorothioate, is a poor substrate for protein phosphophosphatases (reviewed in Ref. 37). As a result, the phosphorylation reaction becomes irre- versible.
Scatchard Analysis of CAMP Binding-D. discoideum cells contain different forms of CAMP-binding sites. A form des- ignated as Bss has high affinity for cAMP (Kd - 13 nM) and shows a very slow rate of dissociation (h,? = 150 s). BS has the same high affinity for CAMP, but the complex dissociates 10- fold faster (tu = 15 s). The major part of the receptors are fast dissociating (Fast, tLh -1 s) and are composed of a t least two forms, AH and AL, with high (Kd = 60 nM) and low (& = 450 nM) affinity, respectively (15, 16). These binding forms are also present in membranes from D. discoideum cells; their proportioning is affected by guanine nucleotides, resulting in an increase of AL and a decrease of the other forms.
The influence of pretreatment, of membranes with MgATPyS on the binding heterogeneity was investigated. Membranes were incubated with 1 nM ATPyS and 5 mM M$+ for 5 min at 20 "C and then extensively washed at 0 "C. Control and MgATPy S-treated membranes were incubated with different concentrations [3H]cAMP in the absence and presence of 30 p~ GTPyS. Binding of [3H]cAMP was detected at equilibrium (b(O)), a t 10 s after the onset of dissociation (b(lO)), and at 2 min after the onset of dissociation (b(120)). Binding to the different receptor forms was calculated with the following set of equations (see Ref. 16).
b(0) = Fast + BS + Bss ( la)
b(10) = 0.65 BS + 0.95 Bss (1b)
b(120) = 0.56 Bss (IC)
The results are shown in Fig. 1. The binding of cAMP to BS or Bss is apparently homogeneous, whereas binding to Fast is clearly composed of at least two components. The kinetic data of BS and Bss were calculated by linear regression analysis, whereas the data of the fast dissociating components were calculated by a hyperbolic regression analysis.
The obtained kinetic data are presented in Table I. The site distribution of control membranes is 26% AH with Kd =
and 5% Bss with Kd = 7.7 nM. GTPyS strongly alters this distribution: about 98% of the sites have the properties of AL; A" and Bss are reduced to 1%, whereas BS is virtually absent ( ~ 0 . 2 % ) . It should be noted that the low abundance of AH complicates the determination of its affinity.
cAMP binding to MgATPyS-treated membranes in the absence of GTPyS strongly resembles the binding to control membranes in the presence of GTPyS. Binding of cAMP to A", BS, and Bss is strongly decreased with a concomitant increase of binding to AL. It should be noted that binding to BS is still detectable in MgATPyS-treated membranes. As in control membranes, this form is undetectable after the addi- tion of GTPyS.
The results of Fig. 1 and Table I indicate that GTPyS or pretreatment with MgATPyS predominantly alters the pro- portioning of binding forms, whereas the total number of binding sites and the affinities are not strongly altered. The effect of MgATPyS is a shift of high to low affinity binding forms, which is most easily detected by equilibrium binding of low cAMP concentrations.
Kinetics of Modification of CAMP Binding by MgATPyS- Membranes were preincubated for various time periods with MgATPyS at 20 "C and washed, and cAMP binding was
50 nM, 59% AL with K d = 545 nM, 10% BS with K d = 6.4 nM,
at University of G
roningen on August 23, 2007
ww
w.jbc.org
Dow
nloaded from
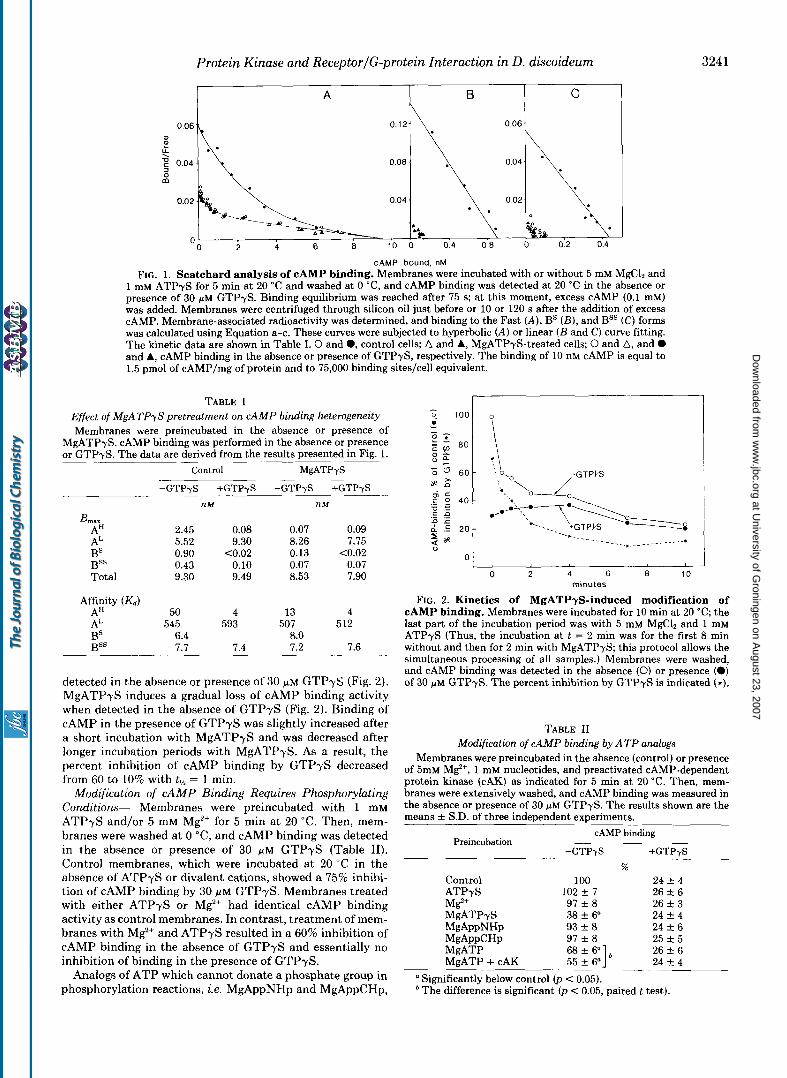
Protein Kinase and ReceptorlG-protein Interaction in D. discoideum 3241
FIG. 1. Scatchard analysis of cAMP binding. Membranes were incubated with or without 5 mM MgC1, and 1 mM ATPyS for 5 min at 20 "C and washed at 0 "C, and cAMP binding was detected at 20 "C in the absence or presence of 30 y~ GTPyS. Binding equilibrium was reached after 75 s; at this moment, excess cAMP (0.1 mM) was added. Membranes were centrifuged through silicon oil just before or 10 or 120 s after the addition of excess CAMP. Membrane-associated radioactivity was determined, and binding to the Fast (A), BS ( B ) , and Bss (C) forms was calculated using Equation a-c. These curves were subjected to hyperbolic (A) or linear ( B and C) curve fitting. The kinetic data are shown in Table I. 0 and 0, control cells; A and A, MgATPyS-treated cells; 0 and A, and 0 and A, cAMP binding in the absence or presence of GTPyS, respectively. The binding of 10 nM cAMP is equal to 1.5 pmol of cAMP/mg of protein and to 75,000 binding sites/cell equivalent.
cAMP bound, nM
TABLE I Effect of MgATPyS pretreatment on CAMP binding heterogeneity Membranes were preincubated in the absence or presence of
MgATPyS. cAMP binding was performed in the absence or presence or GTPyS. The data are derived from the results presented in Fig. 1. -
Control MgATPyS
-GTPyS +GTPyS -GTPyS +GTPyS nM n M
AH 2.45 0.08 0.07 0.09 AL 5.52 9.30 8.26 7.75 BS B S S
0.90 <0.02 0.13 <0.02 0.43 0.10 0.07 0.07
Total 9.30 9.49 8.53 7.90
Affinity (Kd) AH 50 4 13 4 A" 545 593 507 512 BS 6.4 BSS
8.0 7.7 7.4 7.2 7.6 -
detected in the absence or presence of 30 ~ L M GTPyS (Fig. 2). MgATPyS induces a gradual loss of cAMP binding activity when detected in the absence of GTPyS (Fig. 2). Binding of cAMP in the presence of GTPyS was slightly increased after a short incubation with MgATPyS and was decreased after longer incubation periods with MgATPyS. As a result, the percent inhibition of cAMP binding by GTPyS decreased from 60 to 10% with tL,> = 1 min.
Modification of cAMP Binding Requires Phosphorylating Conditions- Membranes were preincubated with 1 mM ATPyS and/or 5 mM Mg2+ for 5 min at 20 "C. Then, mem- branes were washed at 0 "C, and cAMP binding was detected in the absence or presence of 30 PM GTPyS (Table 11). Control membranes, which were incubated at 20 "C in the absence of ATPyS or divalent cations, showed a 75% inhibi- tion of cAMP binding by 30 FM GTPyS. Membranes treated with either ATPyS or M$+ had identical CAMP binding activity as control membranes. In contrast, treatment of mem- branes with M$+ and ATPyS resulted in a 60% inhibition of cAMP binding in the absence of GTPyS and essentially no inhibition of binding in the presence of GTPyS.
Analogs of ATP which cannot donate a phosphate group in phosphorylation reactions, i.e. MgAppNHp and MgAppCHp,
0 2 6 8 10 minutes
FIG. 2. Kinetics of MgATPyS-induced modification of cAMP binding. Membranes were incubated for 10 min at 20 "C; the last part of the incubation period was with 5 mM MgC1, and 1 mM ATPyS (Thus, the incubation at t = 2 min was for the first 8 min without and then for 2 min with MgATPyS; this protocol allows the simultaneous processing of all samples.) Membranes were washed, and cAMP binding was detected in the absence (0) or presence (0) of 30 ~ L M GTPyS. The percent inhibition by GTPyS is indicated (*).
TABLE I1 Modification of CAMP binding by ATP analogs
Membranes were preincubated in the absence (control) or presence of 5 m ~ M P , 1 mM nucleotides, and preactivated CAMP-dependent protein kinase (cAK) as indicated for 5 min at 20 "C. Then, mem- branes were extensively washed, and cAMP binding was measured in the absence or presence of 30 y~ GTPyS. The results shown are the means f S.D. of three independent experiments.
Preincubation cAMP binding
-GTPrS +GTP+ %
Control 100 24 +- 4 ATPyS 102 f 7 26 f 6
MgATPyS 38 f 6" 24 f 4
MgATP 25 f 5
MgATP + cAK 26 f 6
55 k 6' 24 f 4
M e 97 f 8 26 ? 3
MgAppNHp 93 -+ 8 24 f 6 MgAppCHp
Significantly below control (p < 0.05). * The difference is significant (p < 0.05, paired t test).
at University of G
roningen on August 23, 2007
ww
w.jbc.org
Dow
nloaded from
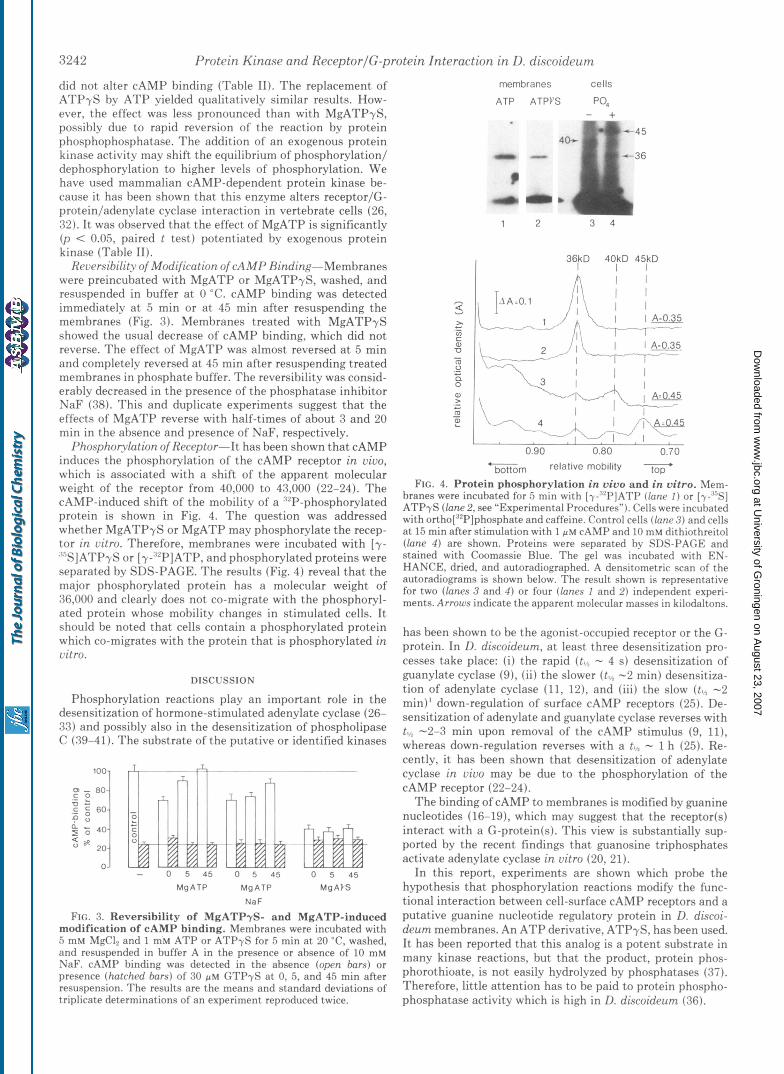
3242 Protein Kinase and ReceptorlG-protein Interaction in D. discoideum
did not alter CAMP binding (Table 11). The replacement of ATPyS by ATP yielded qualitatively similar results. How- ever, the effect was less pronounced than with MgATPyS, possibly due to rapid reversion of the reaction by protein phosphophosphatase. The addition of an exogenous protein kinase act.ivity may shift the equilibrium of phosphorylation/ dephosphorylation to higher levels of phosphorylation. We have used mammalian CAMP-dependent protein kinase be- cause it has been shown that this enzyme alters receptor/(;- protein/adenylate cyclase interaction in vertebrate cells (26, 32). It was observed that t.he effect of MgATP is significantly (p < 0.05, paired t test) potentiated by exogenous protein kinase (Table 11).
Rwersibility of Modification of cAMP Rinding-Membranes were preincubated with MgATP or MgATPyS, washed, and resuspended in buffer a t 0 "C. CAMP binding was detected immediately at 5 min or at 45 min after resuspending the membranes (Fig. 8). Membranes treat.ed with MgATPyS showed the usual decrease of CAMP binding, which did not reverse. The effect of MgATP was almost reversed a t 5 min and completely reversed a t 45 min after resuspending treated membranes in phosphat,e buffer. The reversibility was consid- erably decreased in the presence of the phosphatase inhibitor NaF (38). This and duplicate experiments suggest that the effects of MgATP reverse with half-times of about 3 and 20 min in the absence and presence of NaF, respectively.
Phosphoplation of Receptor-It has been shown that CAMP induces the phosphorylation of the CAMP receptor in vivo, which is associated with a shift of the apparent molecular weight of the receptor from 40,000 to 43,000 (22-24). The CAMP-induced shift of the mobility of a '"P-phosphorylated protein is shown in Fig. 4. The question was addressed whether MgATPyS or MgATP may phosphorylate the recep- tor in vitro. Therefore, membranes were incubated with [y- %]ATP$3 or [y-""PIATP, and phosphorylated proteins were separated by SDS-PAGE. The results (Fig. 4) reveal that the major phosphorylated protein has a molecular weight of 36,000 and clearly does not co-migrate with the phosphoryl- ated protein whose mobility changes in stimulated cells. I t should be noted that cells contain a phosphorylated protein which co-migrates with the protein that is phosphorylated in vitro.
. .
DISCUSSION
Phosphorylation reactions play an important role in the desensitization of hormone-stimulated adenylate cyclase (26- 33) and possibly also in the desensitization of phospholipase C (39-41). The substrate of the putative or identified kinases
MgATP
h
0 5 45 0 5 45
Mg ATP MgAIS NaF
FIG. 3. Reversibility of MgATPyS- and MgATP-induced modification of cAMP binding. Membranes were incubated with 5 mM MgCl? and 1 mM ATP or ATPyS for 5 min a t 20 "C, washed, and resuspended in buffer A in the presence or absence of 10 mM NaF. cAMP binding was detected in the absence (open bars) or presence (hatched bars) of 30 p~ GTPyS a t 0, 5, and 45 min after resuspension. The results are the means and standard deviations of triplicate determinations of an experiment reproduced twice.
membranes cells
ATP ATPVS PO4
1 2
36kD 40kD 45kD I I I h
I I I T
I
r I I I -
4 I "L-v, 'L -P
0.90 0.80 0.70 bottom relative mobility
FIG. 4. Protein phosphorylation in vivo and in vitro. Mem- branes were incubated for 5 rnin with [y-"P]ATP (lane I ) or [y-,'"S] ATPyS (lane 2, see "Experimental Procedures"). Cells were incubated with ortho["'P]phosphate and caffeine. Control cells (lane 3) and cells a t 15 min after stimulation with 1 p M cAMP and 10 mM dithiothreitol (lane 4 ) are shown. Proteins were separated by SDS-PAGE and stained with Coomassie Blue. The gel was incubated with EN- HANCE, dried, and autoradiographed. A densitometric scan of the autoradiograms is shown below. The result shown is representative for two (lanes 3 and 4 ) or four (lanes 1 and 2) independent experi- ments. Arrows indicate the apparent molecular masses in kilodaltons.
has been shown to be the agonist-occupied receptor or the G- protein. In D. discoideurn, a t least three desensitization pro- cesses take place: (i) the rapid (t,., - 4 s) desensitization of guanylate cyclase (9), (ii) the slower (tt,: -2 min) desensitiza- tion of adenylate cyclase (11, 12), and (iii) the slow (t1+ -2 rnin)' down-regulation of surface cAMP receptors (25). De- sensitization of adenylate and guanylate cyclase reverses with tt/> -2-3 min upon removal of the CAMP stimulus (9, 11), whereas down-regulation reverses with a tt,? - 1 h (25). Re- cently, it has been shown that desensitization of adenylate cyclase in vivo may be due to t,he phosphorylation of the CAMP receptor (22-24).
The binding of CAMP to membranes is modified by guanine nucleotides (16-19), which may suggest that the receptor(s) interact with a G-protein(s). This view is substantially sup- ported by the recent findings that guanosine triphosphates activate adenylate cyclase in vitro (20, 21).
In this report, experiments are shown which probe the hypothesis that phosphorylation reactions modify the func- tional interaction between cell-surface CAMP receptors and a putative guanine nucleotide regulatory protein in D. discoi- deum membranes. An ATP derivative, ATPyS, has been used. It has been reported that this analog is a potent substrate in many kinase reactions, but that the product, protein phos- phorothioate, is not easily hydrolyzed by phosphatases (37). Therefore, little attention has to be paid to protein phospho- phosphatase activity which is high in D. discoidam (36).
at University of G
roningen on August 23, 2007
ww
w.jbc.org
Dow
nloaded from

Protein Kinase and ReceptorlG-protein Interaction in D. discoideum 3243
The main results of this study are the following. 1) MgATPyS rapidly (t1,% -1 min) and irreversibly alters cAMP binding. The number of binding sites is slightly (-15%) reduced. In control membranes, GTP@ induces a transition of high affinity binding to a low affinity form. This low affinity binding form is induced by pretreatment of membranes with MgATPyS, and GTPyS does not strongly affect cAMP bind- ing any further. 2) These effects are not induced by Mg' or ATPyS alone or by MgAppNHp or MgAppCHzp. 3) MgATP induces qualitatively similar results as MgATPyS, which are, however, less pronounced. 4) The effect of MgATP is poten- tiated by exogenous CAMP-dependent protein kinase and is readily reversible. Reversibility is slowed down by NaF, a phosphatase inhibitor (38). 5 ) The major phosphorylated pro- tein in membranes is a protein with M, = 36,000.
These results suggest that the effects of MgATPyS and MgATP on cAMP binding are mediated by endogenous pro- tein kinase activity. The substrate of this reaction could be the cAMP receptor, the G-protein, or any other component that affects the receptor/(;-protein interaction. The major phosphorylated protein in isolated membranes has an appar- ent molecular weight of 36,000 and does not co-migrate on SDS-PAGE with the phosphorylated receptor. This suggests that the receptor is not phosphorylated in vitro or that phos- phorylation is below the detection limit. A definite proof whether phosphorylation of the receptor, G-protein, or an- other component is responsible for the observed effects can be given only after a functional analysis of purified phospho- rylated and nonphosphorylated components that are recon- stituted in artificial membranes. Unfortunately, conditions for the isolation and reconstitution of signal transducing components that have been used in vertebrates (31) are not effective in D. discoideum, and no alternative conditions have been found yet.3
Recently, it has been shown that guanosine triphosphate stimulates adenylate cyclase of D. discoideum in vitro (20,21). We observed only a modest degree of stimulation (70%). Interestingly, however, this stimulation was lost in mem- branes derived from desensitized cells (adenylate and guany- late cyclases were desensitized, and receptors were partly down-regulated). In addition, when membranes from control cells were preincubated with MgATPyS, the 70% stimulation of adenylate cyclase by GTPyS was converted to a 30% inhibition. These observations suggest that MgATPyS alters the receptor/G-protein as well as the G-protein/adenylate cyclase interactions. MgATPyS does not alter the number of binding sites. Interestingly, 40% of the binding sites are lost in a reversible manner upon further incubation of MgATPyS- treated membranes with cAMP and micromolar concentra- tions of Ca2'.4 These observations may suggest that the action of MgATPyS could be related to the mechanism of down- regulation of cAMP receptors.
The present and previous (42) results that a putative en- dogenous protein kinase activity modifies receptor/G-protein interactions allow the investigation of many potentially inter- esting questions such as whether occupation of the receptor
P. M. W. Janssens and R. Van Driel , personal communicat ion. P. J. M. V a n Haastert, unpublished results.
or G-protein affects the rate of phosphorylation and dephos- phorylation and concerning the identities of the kinase, the phosphatase, and their substrates. The notion that ATPyS irreversibly and ATP reversibly alter the cAMP binding ac- tivity could facilitate these experiments.
Acknowledgments-I gratefully acknowledge Ronald Van Eyk for exper t ass i s tance dur ing many exper iments and The0 Kon i jn , Pe t e r Devreotes , Ewa Snaar-Jagalska, Fanja Kesbeke, Reni De Wi t , and P i m Janssens for stimulating discussions.
REFERENCES 1. Konijn, T. M., Van de Meene, J. G. C., Bonner, J. T., and Barkley, D. S.
2. Schaap, P., Konijn, T. M., and Van Haastert, P. J. M. (1984) Proc. Nutl.
3. Kay, R. R. (1982) Proc. Nutl. Acad. Sci. U. S. A. 79,3228-3231 4. Gerisch, G . (1982) Annu. Reu. Physiol. 44 , 535-552 5. Van Haastert, P. J. M., and Konijn, T. M. (1982) Mol. Cell. Endocr. 2 6 , l -
6. Devreotes, P. N. (1983) Adu. Cyclic Nucleotide Res. 15,55-96
8. Rossier, C., Eitle, E., Van Driel, R., and Gerisch, G. (1980) SOC. Gen. 7. Devreotes, P. N., and Steck, T. L. (1979) J. Cell Bid . 80,300-309
9. Van Haastert, P. J. M., and Van der Heijden, P. R. (1983) J. Cell B i d . 9 6 ,
(1967) Proc. Natl. Acud. Sci. U. S. A. 5 8 , 1152-1154
Acud. Sci. U. S. A . 81,2122-2126
17
Microbiol. Symp. 30,405-427
10. Wurster, B., and Butz, U. (1983) J. Cell Biol. 9 6 , 1566-1570 11. Dinauer, M. C., Steck, T. L., and Devreotes, P. N. (1980) J. Cell B i d . 8 6 ,
347-353
SAS-SS'l 12. Dinauer, M. C., Steck, T. L., and Devreotes, P. N. (1980) J. Cell Biol. 8 6 ,
13. Van Haastert, P. J. M. (1985) Biochim. Biophys. Acta 846,324-333 14. Kesbeke, F., and Van Haastert, P. J. M. (1985) Biochim. Biophys. Acta
15. Van Haastert, P. J. M., and De Wit, R. J. W. (1984) J. Biol. Chem. 259 ,
16. Van Haastert, P. J. M., De Wit, R. J. W., Janssens, P. M. W., Kesbeke, F.,
17. Van Haastert, P. J. M. (1984) Biochem. Biophys. Res. Commun. 124,597-
18. Janssens, P. M. W., Van der Geer, P. L. J., Arents, J. C., and Van Driel,
19. Janssens, P. M. W., Arents, J. C., Van Haastert, P. J. M., and Van Driel,
20. Theibert, A,, and Devreotes, P. N. (1986) J . Biol. Chem. 261,15121-15125 21. Van Haastert, P. J. M., Snaar-Jagalska, B. E., and Janssens, P. M. W.
22. Klein, P., Theibert, A., Fontana, D., and Devreotes, P. N. (1985) J. Biol.
23. Devreotes, P. N., and Sherring, J. A. (1985) J. Biol. Chem. 260,6378-6384 24. Klein, C., Lubs-Haukeness, J., and Simons, S. (1985) J. Cell Biol. 100 ,
" V
554-561
847,33-39
13321-13328
and DeGoede, J. (1986) J. Biol. Chem. 261,6904-6911
604
R. (1985) Mol. Cell. Biochem. 6 7 , 119-124
R. (1986) Biochemistry 2 5 , 1316-1320
(1986) Eur. J . Biochem., in press
Chem. 2 6 0 , 1757-1764
715-720 25. Klein, C. (1979) J. Biol. Chem. 254,12573-12578 26. Stadel, J. M., Nambi, P., Shorr, R. G. L., Sawyer, D. R., Caron, M. G., and
Lefkowitz. R. J. (1983) Proc. Nutl. Acud. Sci. U. S. A. 80.3171-3177 27. Sibley, D. R:, Peters, J. R., Nambi, P., Caron, M. G., and Lefkowitz, R. J.
(1984) J. E d . Chem. 259.9742-9749 28. Kelleher, D. J., Pessin, J. E.', Ruoho, A. E., and Johnson, G. (1984) Proc.
Natl. Acud. Sci. U. S. A . 81,4316-4320 29. Nambi, P., Peters, J. R., Sibley, D. R., and Lefkowitz, R. J. (1985) J. Biol.
Chem. 260,2165-2171 30. Kirchik, H. J., Igengar, R., and Birnbaumer, L. (1983) Endocrinology 113 ,
163&1648 31. Benovic, J. L., Pike, L. J., Cerione, R. A,, Staniszewski, C., Yoshimasa, T.,
Codina, J., Caron, M. G., and Lefkowitz, R. J. (1985) J. Biol. Chem. 260 , ~ Q A - ~ I ~ I ."I ._"_
32. Sibley, D. R., and Lefkowitz, R. J. (1985) Nature 317 , 124-129 33. Katada, T.. Gilman. A. G.. Watanabe, Y.. Bauer. S.. and Jakobs. K. H.
(1985) Eur. J . Biochem. 151,431-437 34. Gilman, A. G. (1970) Proc. Nutl. Acud. Sci. U. S. A. 6 7 , 305-312 35. Das, 0. P., and Henderson, E. J. (1983) Biochim. Biophys. Acta 736 , 45-
36. Van Driel, R., Arents, J. C., Otte, A. P., and Bernstein, R. L. (1985)
37. Eckstein, F. (1985) Annu. Reu. Biochem. 54,367-402 38. Schaap, P., Van der Molen, L., and Konijn, T. M. (1981) Biol. Cell. 4 1 ,
39. Rittenhouse, S. E., and Sasson, J. P. (1985) J. Biol. Chem. 260,8657-8660 40. Vicentini, L. M., Di Virgilio, F., Ambrosini, A,, Pozzan, T., and Meldolesi,
41. Zavolco, G. B., Halenda, S. P., Sha'afi, R. I., and Feinstein, M. B. (1985)
42. Van Haastert, P. J. M., De Wit, R. J. W., and Van Lookeren Campagne,
56
Biochim. Biophys. Acta 847,122-127
133-142
J. (1985) Biochem. Biophys. Res. Commun. 127,310-317
Proc. Natl. Acud. Sci. U. S. A. 82,3859-3862
M. M. (1985) Biochem. Biophys. Res. Commun. 128, 185-192
at University of G
roningen on August 23, 2007
ww
w.jbc.org
Dow
nloaded from





























