Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 1978, p. 521-5270021-9193/78/0134-0521$02.00/0Copyright 0 197,8 American Society for Microbiology
Vol. 134, No. 2
Printed in U.S.A.
Protease Activity During Cell Differentiation of the CellularSlime Mold Dictyostelium discoideum
DUNNE FONG' * AND CHARLES L. RUTHERFORD2Department ofBiology, Princeton University, Princeton, New Jersey 08540,' and Department ofBiology,
Virginia Polytechnic Institute and State University, Blacksburg, Virginia 240612Received for publication 6 December 1977
Individual aggregates, migrating pseudoplasmodia, and sorocarps of Dictyoste-lium discoideum were assayed for proteolytic activities by colorimetric andfluorometric techniques. Cathepsin D-like and cathepsin B-like acid proteaseactivities were found to decrease throughout development, but the patterns ofdecrease were different for the two enzymes. A gradual decrease was found forcathepsin D, whereas a sharp decrease between aggregates and migrating pseu-doplasmodia was detected for cathepsin B. By using microdissection techniquesand fluorometric assays for amino acids and peptides, prestalk cells and presporecells exhibited no difference in cathepsin D activity, whereas cathepsin B activitywas higher in the prestalk cells. Similarly, stalk cells and spores in the sorocarpsshowed no difference in cathepsin D activity, but showed a fivefold highercathepsin B activity in the stalk cells. This finding suggests a possible role forcathepsin B in stalk cell differentiation.
During morphogenesis of the cellular slimemold Dictyostelium discoideum, amoebae ag-gregate to form a multicellular migrating pseu-doplasmodium, which later differentiates into asorocarp composed of stalk cells and spores (5,13). Aggregation and differentiation from oneinitial cell type to two final cell types are inducedby starvation, especially of amino acids (15, 16).Protein degradation occurs during this process,as measured by decreasing cellular protein con-tent from amoebae to sorocarps (6, 7, 27, 31).Thus, information on protease activities duringdevelopment ofDictyostelium would be of inter-est, especially with regard to stalk cell and sporedifferentiation, because the former undergoesaging and cell death, whereas the latter is astorehouse for future germination. Sussman andSussman (23) and Wiener and Ashworth (30)reported acid protease activity at pH optima of2.3 and 2, respectively, with minor changes inactivity during development (30). Recently, in arelated species, Polysphondyliun pallidum,O'Day (18) reported two peaks of acid proteaseactivity (pH optima 3.5 and 6.0) during micro-cyst germination.
Results of experiments described in the pres-ent report indicate that cathepsin-like proteaseactivities are present at different stages of de-velopment. (Reasons for attributing the pro-teases to be cathepsins will be discussed in alater section.) Single slime mold individuals (ag-gregates, migrating pseudoplasmodia, or soro-carps) were used for these assays. Moreover,
with microdissection and a fluorometric reactionfor amino acids and peptides, protease activitydifferences in differentiating cell types werestudied. No difference was detected in cathepsinD activity between prestalk and prespore cellsat the slug stage and between stalk cells andspores at the sorocarp stage. However, cathepsinB activity was higher in the pseudoplasmodialprestalk cells than in prespore cells. Similarly,cathepsin B activity in stalk cells was fivefoldhigher than in spores.
MATERIALS AND METHODSCulture conditions. LyophFlized individual pseu-
doplasmodia of D. discoideum (strain NC4) were pre-pared as described previously (11, 20). Briefly, vege-tative amoebae harvested from Escherichia coli B/rwere allowed to differentiate on filter paper disks.Samples at different time intervals were collected,lyophilized, and stored at -76°C. Axenic strains of D.discoideum (strains Ax2 and Ax3) were grown in HL-5 medium in liquid culture (26). Amoebae were har-vested at the exponential phase of growth and storedat -20°C until used.
Protease characterization studies. To avoidpossible bacterial contamination, axenically grownamoebae were used in all of the enzyme inhibitorassays. Dictyostelium extracts were made by sonictreatment of the amoebae (Branson Sonifier, setting5, 1 min) and centrifuging to obtain the supernatant(15,000 rpm, 10 min, in a Sorvall SS-34 rotor). Fordiazoacetyl norleucine methyl ester inhibition of ca-thepsin D, the method of Keilova (12) was foilowed,with inhibition by diazoacetyl norleucine methyl esterand cupric acetate at pH 5 and subsequent assay for
521
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

522 FONG AND RUTHERFORD
hemoglobin hydrolysis at pH 2.5. For pepstatin inhi-bition of cathepsin D, the method of Umezawa andAoyagi (25) was adopted. Pepstatin was dissolved inmethanol before dilution in water. Chloroquine inhi-bition of cathepsin B was performed by the method ofWibo and Poole (28), except that benzoylarginine ni-troanilide was used as substrate. For iodoacetate, io-doacetamide, and tosyl lysyl chloromethyl ketone in-hibitions of cathepsin B, the methods of Barrett (2)and Snellman (22) were used. Enzyme extracts were
first incubated with inhibitors before benzoylargininenitroanilide addition at pH 5.5. Leupeptin inhibitionof cathepsin B was similar except that benzyl argininenaphthylamide was used together with an improvedcolor reagent (3).
Microtechniques. Microdissection with handmademicroscalpels on lyophilized tissues had been de-scribed (11, 20). Samples were weighed on a quartzfiber balance (with a sensitivity of 1.46,tg per deflec-tion); and then transferred to Kimble disposable bo-rosilicate glass culture tubes (6 by 50 mm). Buffer andsubstrate were added, and the tubes were then coveredwith Parafilm and incubated at 37°C. Constrictionmicropipettes (14) were used for the transfer of liquids.
Colorimetric protease assay. For cathepsin D,the buffer used was 0.05 M citrate, pH 2.75. Forcathepsin B, the buffer was 0.05 M acetate, pH 5.5,with added activators 4 mM Na2-ethylenediaminetet-raacetic acid and 10 mM dithiothreitol. In colorimetricassays, 94.9 !d of buffer was added to the tube, togetherwith 19.3 ,ul of 50-mg/ml azoalbumin (Sigma ChemicalCo.), pH 7.2. Incubation at 37°C was 15 h for cathepsinD and 20 h for cathepsin B. The reaction was stoppedby adding 111 ,ul of cold 10% trichloroacetic acid. Low-speed centrifugation (3,000 rpm, 15 min, Internationalcentrifuge, refrigerated model PR-J) gave a trichloro-acetic acid-soluble fraction. A 111-pl amount of thetrichloroacetic acid-soluble fraction was added with anequal volume of 0.5 N NaOH, and readings were takenat 440 nm (model 240 Gilford spectrophotometer) bythe method of Tomareili et al. (24).
Fluorometric protease assay. Fluorometric as-
says were similar to those of Hirsch and Parks (8),except that o-phthalaldehyde (Eastman OrganicChemical Division of Eastman Kodak Co.) and ,B-mercaptoethanol (Sigma) were used instead of flu-orescamine. For cathepsin D assay, 1% (wt/vol) he-moglobin (Worthington Biochemicals Corp.) was
added to 0.05 M citrate buffer at pH 2.75. Either 58.3or 100 id was used per tube, with a 10-h incubation at37°C. For cathepsin B assay, 1% (wt/vol) bovine serumalbumin (Sigma) was added to 0.05 M acetate buffer(pH 5.5) and activators (4 mM Na2-ethylenediamine-tetraacetic acid and 10 mM dithiothreitol). Either 51.4or 58.3 id was used per tube, with a 20-h incubation at37°C. Reactions were stopped with an equal volumeof cold 10% trichloroacetic acid, and centrifuged fortrichloroacetic acid-soluble fractions. Portions of thetrichloroacetic acid-soluble fractions, e.g., 27.2 and 46.5p1, were assayed fluorometrically for released aminoacids and peptides, by the method of Roth (19), byusing unincubated controls as blanks. Each trichloro-acetic acid-soluble portion was added to 1 ml of o-phthalaldehyde buffer (50 ml of 0.4 M borate adjustedto pH 10.4 with KOH, with 7.5 mg ofo-phthalaldehyde
in 0.75 ml of methanol, and with 50 1d of ,B-mercapto-ethanol). This was Vortex mixed, incubated in thedark for 30 min at room temperature, and finally readin a Farrand fluorometer model A4, with appropriatefilters for o-phthalaldehyde (excitation at 340 nm,emission at 455 nm).
RESULTSCharacterization of acid protease activi-
ties. A preliminary study of inhibitors of Dic-tyostelium acid proteases is shown in Table 1.Cathepsin D activity was inhibited by diazoace-tyl norleucine methyl ester and partially in-hibited by pepstatin. The latter finding sug-gested that other acid proteases may also beinvolved because crude cell extract was used inthis enzyme assay. Cathepsin B activity wasinhibited by iodoacetate, iodoacetamide, tosyllysyl chloromethyl ketone, chloroquine, and leu-peptin. In conclusion, besides the pepstatin re-sult, all properties of enzyme inhibition corre-lated well with known cathepsin D and cathep-sin B activities (4).Cathepsin activity of whole individuals.
Single slime mold pseudoplasmodia were usedto assay for acid protease activities by usingazoalbumin as a substrate. The colorimetric pro-tease assay results are shown in Table 2. Pro-tease activity per unit (dry weight) decreasedthroughout development. At any one stage of
TABLE 1. Effects of inhibitors on acidproteaseactivities in crude D. discoideum extracte
Dtrina- Inhibitor Final concn bitiontion btoCathepsin D DNME 1 mM 26.6assay 10 mM 68.5
50 mM 81.5
Pepstatin 0.16 jg/ml 31.20.8 pg/ml 35.54 pg/ml 37.8
Cathepsin B Iodoacetate 0.01 mM 94.8assay 1 mM 100
lodoacetamide 1 mM 100
TLCK 1 pM 73.310 pM 93.5
100 pM 96.3
Chloroquine 5 mM 32.325 mM 74.150 mM 84.4
Leupeptin 0.32 Ag/ml 53.01.6 pg/ml 80.78 pg/ml 89.7
Amoebae extracts were tested for acid protease activitiesin the presence and absence of inhibitors. DNME, Diazoacetylnorleucine methyl ester; TLCK, tosyl lysyl chloromethyl ke-tone.
J. BACTERIOL.
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
PROTEASE IN D. DISCOIDEUM
development, cathepsin D activity was alwayshigher than cathepsin B activity. This last resultis similar to that reported by O'Day (18), whosepH 3.5 protease activity was higher than a pH 6protease activity in a related slime mold species.To achieve greater sensitivity, a fluorometric
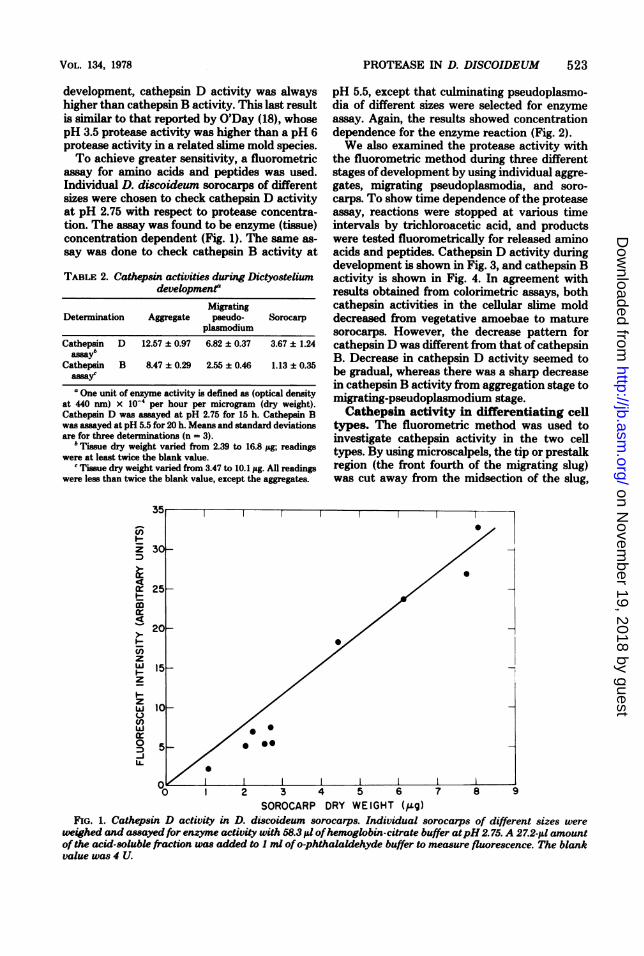
assay for amino acids and peptides was used.Individual D. discoideum sorocarps of differentsizes were chosen to check cathepsin D activityat pH 2.75 with respect to protease concentra-tion. The assay was found to be enzyme (tissue)concentration dependent (Fig. 1). The same as-say was done to check cathepsin B activity at
TABLE 2. Cathepsin activities during Dictyosteliumdevelopmenr
MigratingDetermination Aggregate pseudo- Sorocarp
plasmodium
Cathepsin D 12.57 ± 0.97 6.82 ± 0.37 3.67 ± 1.24assayb
Cathepsin B 8.47 ± 0.29 2.55 ± 0.46 1.13 ± 0.35assayca One unit of enzyme activity is defined as (optical density
at 440 nm) x 10-4 per hour per microgram (dry weight).Cathepsin D was assayed at pH 2.75 for 15 h. Cathepsin Bwas assayed at pH 5.5 for 20 h. Means and standard deviationsare for three determinations (n = 3).
b Tissue dry weight varied from 2.39 to 16.8 j&g; readingswere at least twice the blank value.
c Tissue dry weight varied from 3.47 to 10.1 jg. All readingswere less than twice the blank value, except the aggregates.
pH 5.5, except that culminating pseudoplasmo-dia of different sizes were selected for enzymeassay. Again, the results showed concentrationdependence for the enzyme reaction (Fig. 2).We also examined the protease activity with
the fluorometric method during three differentstages ofdevelopment by using individual aggre-gates, migrating pseudoplasmodia, and soro-carps. To show time dependence of the proteaseassay, reactions were stopped at various timeintervals by trichloroacetic acid, and productswere tested fluorometrically for released aminoacids and peptides. Cathepsin D activity duringdevelopment is shown in Fig. 3, and cathepsin Bactivity is shown in Fig. 4. In agreement withresults obtained from colorimetric assays, bothcathepsin activities in the cellular slime molddecreased from vegetative amoebae to maturesorocarps. However, the decrease pattern forcathepsin D was different from that of cathepsinB. Decrease in cathepsin D activity seemed tobe gradual, whereas there was a sharp decreasein cathepsin B activity from aggregation stage tomigrating-pseudoplasmodium stage.Cathepsin activity in differentiating cell
types. The fluorometric method was used toinvestigate cathepsin activity in the two celltypes. By using microscalpels, the tip or prestalkregion (the front fourth of the migrating slug)was cut away from the midsection of the slug,
I.-
I-
zwzI-
zw
UJ)w0-JLL
30[
25F
20O
15~
10[
5
-O 1 2 3 4 5 6 7 8 9
SOROCARP DRY WEIGHT (Q9g)FIG. 1. Cathepsin D activity in D. discoideum sorocarps. Individual sorocarps of different sizes were
weighed and assayed for enzyme activity with 58.3 1d ofhemoglobin-citrate buffer atpH2.75. A 27.2-i,u amountof the acid-solubk fraction was added to I ml ofo-phthalaldehyde buffer to measure fluorescence. The blankvalue was 4 U.
I0
00I
I I
VOL. 134, 1978 523
--I
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

524 FONG AND RUTHERFORD
ID
- 20
z~I
Z4
Q~ 0 2 0 12 1 6 1
ID
"CULMINATE" DRY WEIGHT I(sgI
FIG. 2. Cathepsin B activity in D. discoideum dur-ing early culmination. Individual culminatingmasses of different sizes were weighed and assayedfor enzyme activity with 51.4 ,ul of albumin-acetatebuffer at pH 5.5. A 51.4-,.tl amount of acid-solublefraction was added to)1 ml ofo-phthalaldehyde bufferto measure fluorescence. The blank value was 6.27 U.
I-
w
3:
c)
zwC-)
zL)w
0D
LL.
3C
20
15
10
5
0
0 2.5 5.0 7.5 10.0TIME (hours)
FIG. 3. Cathepsin D activity during D. discoideumdevelopment. Individuals were weighed and assayedfor enzyme activity in 100 ,ul of hemoglobin-citrate(pH2.75) at 37°C. A 88.4-1ul amount ofthe acid-solublefraction was added to I ml ofo-phthalaldehyde bufferto measure fluorescence. Unincubated individualsserved as blank values. Fluorescent intensities are inarbitrary units. Tissue dry weight variedfrom 1.40 to5.99 pg; the average value was 2.91 iLg. Aggregates(0), migrating pseudoplasmodia (C), and sorocarps
(A).
the prespore region. The last fourth of the slugwas not used because this includes potentialbasal disk cells, similar to prestalk cells. Mature
stalks were similarly dissected out from thespore masses in fruiting bodies. However, due tosensitivity problems in the protease assay, it wasnecessary to pool together 20 isolated stalks(after cutting away their basal disks) to getenough dry weight for one datum point. Anequivalent weight of spore material was assayedfor comparison in a separate tube. Moreover, theo-phthalaldehyde reagent used had significant
background fluorescence, so efforts were madeto obtain readings at least twice the blank values.Because of possible day-to-day variation in flu-orescence readings, the higher value of one celltype was taken to be 100, and the other valuewas calculated as a percentage of the highervalue, in that way showing the relative fluores-
TIME (hours)FIG. 4. Cathepsin B activity during D. discoideum
development. Individuals were weighed and assayedfor enzyme activity in 58.3 il of albumin-acetate (pH5.5) at 37°C. A 46.5-1l amount of the acid-solublefraction was added to 1 ml ofo-phthalaldehyde bufferto measure fluorescence. Unincubated individualsserved as blank values. Fluorescent intensities are inarbitrary units. Tissue dry weight varied from 0.85 to5.96 ,.ug; the average value was 2.31 pig. Aggregates(0), migrating pseudoplasmodia (E), and sorocarps(A).
T ~
)/ Aggregate_
Pseudoplasmodium
i~~~~~~0
i/A/O @t Sorocorp
II
J. BACTERIOL.
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PROTEASE IN D. DISCOIDEUM 525
cence between the two cell types. Within boththe slug and sorocarp stages of development,cathepsin D activity was similar between devel-oping stalk cells and spores (Tables 3 and 4).Thus, this enzyme activity did not change duringcell type differentiation at any stage of devel-opment.However, cathepsin B activity gave a totally
different picture. In the migrating pseudoplas-modium, cathepsin B activity was 1.7-fold higherin prestalk cells than prespore cells (Table 3).Also, in the sorocarp, cathepsin B activity aver-
aged 5.6-fold higher in the stalk cells in thespores (Table 4).Some values in Table 4 for spore-specific ac-
tivity were less than twice the blank values, so
it was necesary to determine whether this rep-
resented actual cathepsin B activity in thespores. We assayed for cathepsin B activity in
isolated spore tissue of increasing dry weights.As spore dry weight increased, cathepsin B ac-
tivity also increased (data not shown), indicating
TABLE 3. Cathepsin activities in D. discoideummigratingpseudoplasmodia
Determination Prestalk Preporecells cells
Cathepsin D assay"(1)b 100 78.9(2) 97.1 100(3) 100 94.2
Cathepsin B asay'(1)d 100 66.3(2) 100 60.6(3) 100 60.0(4) 100 66.1(5) 100 56.3(6) 100 51.0
'A 100-pl amount of hemoglobin-citrate buffer atpH 2.75 was used (10 h of incubation at 370C), and thereaction was terminated by trichloroacetic acid. Acid-soluble product was assayed fluorometrically with o-
phthalsldehyde buffer. The fluorescent reading minusblank was then divided by dry weight. Relative unitswere achieved by assuming the higher reading to be100, so the other reading became a percent value. Allfluorescent readings were at least twice the blankvalues.
b Prestalk and prespore cells were dissected fromthe pseudoplasmodia. Dry weights in micrograms (pre-stalk/prespore): (1) 6.86:7.18; (2) 6.15:6.19; and (3)6.06:6.06.
c A 58.3-pl amount of albumin-acetate buffer at pH5.5 was used for (1) to (4), and 51.4 yi was used for (5)and (6), with a 20-h incubation at 370C, and othertreatments same as footnote a.
d Prestalk/prespore dry weights in micrograms: (1)6.95:7.37; (2) 7.02:7.88; (3) 8.19:7.64; (4) 8.57:7.96; (5)7.64:7.65; and (6) 7.50:7.40.
TABLE 4. Cathepsin activities in D. discoideumsorocarps
Determination Stalk cells Spores
Cathepsin D assaya(1)b 99.4 100(2) 100 89.1(3) 72.5 100(4) 100 75.1(5) 77.2 100
Cathepsin B assayc(1)d 100 20.3(2) 100 17.0(3) 100 24.3(4) 100 13.5(5) 100 10.3(6) 100 27.9(7) 100 11.1
a Same as Table 3. A 58.3-pl amount of hemoglobin-citrate was used in (1); in all others, 100 pil was used.All readings were at least twice the blank values.Relative units achieved by assuming the higher valueto be 100.
b Dry weights in micrograms: (1) Stalks with basaldisks, 3.17 from 6 sorocarps; spores, 3.08; (2) 8 stalkswith bases, 7.3; spores, 7.46; (3) 8 stalks with bases, 6.1;spores, 5.99; (4) 20 dissected stalks, 6.73; spores, 6.83;(5) 21 dissected stalks, 5.99; spores, 6.37.'Same as Table 3. A 51.4-!d amount of albumin-
acetate was used for (1), (2), (3), and (7); 58.3 yd wasused for (4), (5), and (6). In (1) to (3), readings were atleast twice the blank values; in (4) to (7) the sporevalues were less than twice the blank values.
d Dry weights in micrograms: (1) stalks with basaldisks, 8.54 from 10 sorocarps; spores 8.26; (2) 10 stalkswith bases, 6.41; spores, 6.60; (3) 16 dissected stalks,5.93; spores, 5.97; (4) 13 stalks, 7.39; spores, 7.34; (5) 15stalks, 6.64; spores, 6.80; (6) 20 stalks, 7.96; spores, 8.02;(7) 25 stalks, 7.53; spores, 7.74.
that spores did contain cathepsin B activity,although at much lower levels than stalk cells.
DISCUSSIONDifferences in cathepsin activity during Dic-
tyostelium development have been demon-strated by using microtechniques and fluoro-metric amino acid and peptide assays. Althoughcathepsin D activity was similar in prestalk andprespore cells, and in stalk cells and spores,cathepsin B activity was higher in prestalk thanprespore cells and again much higher in stalkcells than spores.We have used the terms cathepsin D and
cathepsin B because our enzymes showed manyof the characteristics of these two cathepsins(Table 1). Studies involving activators, inhibi-tors, substrates, and pH optima were performedto distinguish the two cathepsins. The terms arechosen because cathepsins D and B are themajor known lysosomal acid proteases, since
VOiL. 134, 1978
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

526 FONG AND RUTHERFORD
distribution of related enzymes, cathepsins Hand L, and cathepsin E has only been confirmedin rat liver and rabbit bone marrow, respectively(4).There may be other proteases in the cellular
slime mold. Partial inhibition by pepstatin indi-cated the possibility of additional acid proteases.Native hemoglobin-acrylamide gels showedthree protease bands with amoebae extract atpH 2.75, and at least three peptidase bands hadbeen detected by using dipeptides as substrates(D. Fong, unpublished data).Our finding of fivefold higher cathepsin B
activity in stalk cells than spores is consistentwith earlier results of Gregg et al. (6), who re-ported that spores retained higher protein con-tent than stalk cells. Because cathepsin D activ-ity was similar in the two cell types, it is possiblethat cathepsin B was responsible for higher stalkcell proteolysis.The evidence suggests that cathepsins and
other proteases may play roles during develop-ment ofan organism (9). Cathepsin D was shownto be needed for amphibian metamorphosis, be-cause its inhibitor pepstatin selectively blockedhormone-induced tadpole tail fin regression invitro (21). Protease activities change during de-velopment. Marks et al. (17) reported an initialdecrease and a later increase for cathepsin Dand neutral protease activity during maturationof the rat brain. A somewhat analogous caseconcerning cellular protein degradation was pre-sented by Amenta et al. (1) in cycloheximide-treated fibroblasts, which showed decreased spe-cific activities of the lysosomal proteases, withthe pattern of decrease for cathepsin D differentfrom that for cathepsin B. Our data also showedcathepsin D and cathepsin B activity decreasesduring differentiation from amoebae to soro-carps in D. discoideum. The pattern of decreasewas different for the two enzyme activities.
Cathepsin B may play a more important rolein cellular regulatory phenomena than cathepsinD. For instance, in a study of muscles of dys-trophic chickens, an increase in cathepsin Bactivity correlated well with muscle autolysisand preceded any increase in cathepsin D activ-ity (10). Cathepsin B and related thiol proteasesindicated preferred digestion of young cytosol,whereas cathepsin D showed no preference in astudy comparing protease digestion of youngversus old rat liver cytosol proteins (29). Ourdata for D. discoideum also suggested a regula-tory role for cathepsin B during development.There was a marked difference of cathepsin Bactivity between stalk cells and spores, whereasthere was no difference for cathepsin D activity.Hence, cathepsin B may play some develop-
mental function in the cellular slime molds. Per-haps the localization of cathepsin B activity inthe stalk cells is a reflection of cell-specific pro-tein degradation. The protease activity differ-ence between stalk cells and spores may be dueto selective synthesis or degradation of the en-zyme, activators or inhibitors of the enzyme, orsecretion of enzyme into the medium. O'Day(18) reported that inhibitors were probably ab-sent and proteases were being secreted, at leastduring microcyst germination in Polysphondy-lium. Obviously, the role of proteases in Dic-tyostelium needs further study.
ACKNOWLEDGMENTESDunne Fong is indebted to John Tyler Bonner, his advisor,
for advice, encouragement, and guidance. He also wishes tothank Alan Barrett for 4-amino-2',3-dimethylazobenzene, andHanao Umezawa for gifts of pepstatin and leupeptin. Wethank Michael Hanna, Will Kopachik, Austin Newton, andPauline Pan for reading the manuscript.
This study was supported by research grant PCM77-00201from the National Science Foundation, Public Health Servicegrant GM 17856 from the National Institute of General Med-ical Sciences to John Tyler Bonner, and grant AG 00677 fromNational Institutes of Health to Charles L. Rutherford.
LITERATURE CrMD
1. Amenta, J. S., M. J. Sargus, and F. M. Baccino. 1977.Control of cell protein degradation, changes in activitiesof lysosomal proteases. Biochim. Biophys. Acta476:253-261.
2. Barrett, A. J. 1973. Human cathepsin Bi, purificationand some properties of the enzyme. Biochem. J.131:809-822.
3. Batrett, A. J. 1976. An improved color reagent for use inBarrett's assay of cathepsin B. Anal. Biochem.76:374-376.
4. Barrett, A. J., and M. F. Heath. 1977. Lysosomal en-zymes, p. 19-145. In J. T. Dingle (ed.), Lysosomes, alaboratory handbook, 2nd ed. North-Holland, Amster-dam.
5. Bonner, J. T. 1967. The cellular slime molds, 2nd ed., p.1-205. Princeton University, Princeton.
6. Gregg, J. H., A. L Hackney, and J. 0. Krivanek.1954. Nitrogen metabolism of the slime mold Dictyo-stelium discoideum during growth and morphogenesis.Biol. Bull. (Woods Hole, Mass.) 107:226-235.
7. Hames, B. D., and J. M. Ashworth. 1974. The metab-olism of macromolecules during the differentiation ofmyxamoebae of the cellular slime mould Dictyosteliumdiscoideum containing different amounts of glycogen.Biochem. J. 142:301-315.
8. Hirsch, H. E., and M. E. Parks. 1973. The quantitativehistochemistry of acid proteinase in the nervous system:localization in neurons. J. Neurochem. 21:453-458.
9. Holzer, H., H. Betz, and E. Ebner. 1975. Intracellularproteinases in microorganisms, p. 103-156. In B. L.Horecker and E. R. Stadtman (eds.), Current topics incellular regulation, vol. 9. Academic Press Inc., NewYork.
10. Iodice, A. A., J. Chin, S. Perker, and I. M. Weinstock.1972. Cathepsins A, B, C, D and autolysis during devel-opment of breast muscle of normal and dystrophicchickens. Arch. Biochem. Biophys. 152:166-174.
11. Jefferson, B. L, and C. L Rutherford. 1976. A stalk-
J. BACTIERIOL.
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PROTEASE IN D. DISCOIDEUM 527
specific localization of trehalase activity in Dictyoste-lium discoideum. Exp. Cell Res. 103:127-134.
12. Keilova, H. 1970. Inhibition of cathepsin D by diazoace-tylnorleucine methyl ester. FEBS Lett. 6:312-314.
13. Loomis, W. F. 1975. Dictyostelium discoideum, a devel-opmental system, p. 1-214. Academic Press Inc., NewYork.
14. Lowry, 0. H., and J. V. Passonneau. 1972. A flexiblesystem of enzymatic analysis, p. 1-291. Academic PressInc., New York.
15. Marin, F. T. 1976. Regulation of development in Dictyo-stelium discoideum. I. Initiation of the growth to de-velopment transition by amino acid starvation. Dev.Biol. 48:110-117.
16. Marin, F. T. 1977. Regulation of development in Dictyo-stelium discoideum. II. Regulation of early cell differ-entiation by amino acid starvation and intercellularinteraction. Dev. Biol. 60:389-395.
17. Marks, N., F. Stern, and A. LaItha. 1975. Changes inproteolytic enzymes and proteins during maturation ofthe brain. Brain Res. 86:307-322.
18. O'Day, D. H. 1976. Acid protease activity during germi-nation of microcysts of the cellular slime mold Poly-sphondyliumpallidum. J. Bacteriol. 125:8-13.
19. Roth, M. 1971. Fluorescence reaction for amino acids.Anal. Chem. 43:880-882.
20. Rutherford, C. L, and J. F. Harris. 1976. Localizationof glycogen phosphorylase in specific cell types duringdifferentiation of Dictyosteluwm discoideum. Arch. Bio-chem. Biophys. 175:453-462.
21. Seshimo, H., M. Ryuzaki, and K. Yoshizato. 1977.Specific inhibition of triiodothyronine-induced tadpoletail-fin regression by cathepsin D-inhibitor pepstatin.Dev. Biol. 59:96-100.
22. Snellman, 0. 1970. A study of the reactivity of the thiolgroup of cathepsin B, p. 29-43. In A. J. Barrett and J.T. Dingle (ed.), Tissue proteinases, North-Holland, Am-
sterdam.23. Sussman, M., and R. Sussman. 1969. Patterns of RNA
synthesis and of enzyme accumulation and disappear-ance during cellular slime mould cytodifferentiation.Symp. Soc. Gen. Microbiol. 19:403-435.
24. TomareUi, R. M., J. Charney, and M. L. Harding.1949. Use of azoalbumin as substrate in the colorinmetricdetermination of peptic and tryptic activity. J. Lab.Clin. Med. 34:428-433.
25. Umezawa, H., and T. Aoyagi. 1977. Activities of pro-teinase inhibitors of microbial origin, p. 637-662. In A.J. Barrett (ed.), Proteinases in mammalian cells andtissues. North-Holland, Amsterdam.
26. Watts, D. J., and J. M. Ashworth. 1970. Growth ofmyxamoebae of the cellular slime mould Dictyosteliumdiscoideum in axenic culture. Biochem. J. 119:171-174.
27. White, G. J., and M. Sussman. 1961. Metabolism ofmajor cell components during slime mold morphogen-esis. Biochim. Biophys. Acta 53:285-293.
28. Wibo, M., and B. Poole. 1974. Protein degradation incultured cells. II. The uptake of chloroquine by ratfibroblasts and the inhibition of cellular protein degra-dation and cathepsin B1. J. Cell Biol. 63:430-440.
29. Wiederanders, B., S. Ansorge, P. Bohley, H.Kirschke, J. Langner, and EL Hanson. 1977. Theage dependence of intracellular proteolysis in the ratliver, p. 144-147. In V. Turk and N. Marks (ed.), Intra-cellular protein catabolism II. Plenum Publishing Corp.,New York.
30. Wiener, E., and J. M. Ashworth. 1970. The isolationand characterization of lysosomal particles from myxa-moebae of the cellular slime mould Dictyostelium dis-coideum. Biochem. J. 118:505-512.
31. Wright, B. E., and M. L. Anderson. 1960. Protein andamino acid turnover during differentiation in the slimemold. I. Utilization of endogenous amino acids andproteins. Biochim. Biophys. Acta 43:62-66.
VOL. 134, 1978
on Novem
ber 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from