Embed Size (px)
Citation preview

21 August 1959, Volume 130, Number 3373
CURRENT PROBLEMS IN RESEARCH
Affectional Responsein the Infant Monke'
Orphaned baby monkeys develop a strong and persisterattachment to inanimate surrogate mother
Harry F. Harlow and Robert R. Zimmerman
Investigators from diverse behavioralfields have long recognized the strongattachment of the neonatal and infantileanimal to its mother. Although this af-fectional behavior has been commonlyobserved, there is, outside the field ofethology, scant experimental evidencepermitting identification of the factorscritical to the formation of this bond.Lorenz (1) and others have stressed theimportance of innate visual and auditorymechanisms which, through the processof imprinting, give rise to persisting fol-lowing responses in the infant bird andfish. Imprinting behavior has been dem-onstrated successfully in a variety. ofavian species under controlled labora-tory conditions, and this phenomenonhas been investigated systematically inorder to identify those variables whichcontribute to its development and main-tenance [see, for example, Hinde,Thorpe, and Vince (2), Fabricius (3),Hess (4), Jaynes (5), and Moltz andRosenblum (6) ]. These studies representthe largest body of existent experimen-tal evidence measuring the tie betweeninfant and mother. At the mammalianlevel there is little or no systematic ex-perimental evidence of this nature.
Observations on monkeys by Carpen-ter (7), Nolte (8), and Zuckermann(9) and on chimpanzees by Kohler (10)and by Yerkes and Tomilin (11) show
The authors are on the staff of the primatelaboratory, department of psychology, Universityof Wisconsin, Madison.
21 AUGUST 1959
that monkey and chimpanmvelop strong ties to theirthat these affectional attapersist for years. It is, ofmon knowledge that hiform strong and persistenmothers.
Although students fromtific fields recognize this ament, there is considerableabout the nature of its devits fundamental underlying-A-common theory amongsociologists, and anthropoof learning based on driThis theory proposes thaattachment to the motherthe association of the motlform with the alleviation (
mary drive states, partici,and thirst. Thus, throughfection becomes a self-surived drive (12). Psychoarother hand, have stressed tiof various innate needs, suto suck and orally posse(2), or needs relating to cment, temperature (13),to the mother (14).The paucity of experime
concerning the developmtional responses has led tto derive their basic hypdeductions and intuitionsservation and analysis ofreports. As a result, the asvational evidence is often
SCIENCEpreconceived theoretical framework. Anexception to the above generalization isseen in the recent attempt by Bowlby(14) to analyze and integrate the avail-
S able observational and experimental evi-dence derived from both human andsubhuman infants. Bowlby has concludedthat a theory of component instinctualresponses, species specific, can best ac-count for the infant's tie to the mother.
it He suggests that the species-specific re-sponses for human beings (some of these
S. responses are not strictly limited to hu-man beings) include contact, clinging,
In sucking, crying, smiling, and following.He further emphasizes that these re-sponses are manifested independently ofprimary drive reduction in human andsubhuman infants.
zee infants de- The absence of experimental datamothers and which would allow a critical evaluation
ichments may of any theory of affectional developmentcourse, com- can be attributed to several causes. Theuman infants use of human infants as subjects has seri-t ties to their ous limitations, since it is not feasible to
employ all the experimental controlsdiverse scien- which would permit a completely ade-.biding attach- quate analysis of the proposed variables.disagreement In addition, the limited response reper-relopment and toire of the human neonate severely re-mechanisms. stricts the number of discrete or precisepsychologists, response categories that can be meas-logists is that ured until a considerable age has beenive reduction. attained. Thus, critical variables go un-t the infant's measured and become lost or confoundedr results from among the complex physiological, psy-her's face and chological, and cultural factors whichof certain pri- influence the developing human infant.ularly hunger Moreover, the use of common labora-learning, af- tory animals also has serious limitations,
pporting, de- for most of these animals have behav-ialysts, on the ioral repertoires very different fromhe importance those of the human being, and in manyich as a need species these systems mature so rapidlyss the breast that it is difficult to measure and assess,ontact, move- their orderly development. On the otherand clinging hand, subhuman primates, including
the macaque monkey, are born at a stateental evidence of maturity which makes it possible toent of aff&e begin precise measurements within thehese theorists first few days of life. Furthermore, their?otheses front postnatal maturational rate is slowbased on ob- enough to permit precise assessment ofadult verbal affectional variables and development.
iailable obser- Over a 3-year period prior to the be-forced into a ginning of the research program reported
, 421
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Fig. 1. Wire and cloth mother surrogates.
here (15), some 60 infant macaquemonkeys were separated from theirmothers 6 to 12 hours after birth andraised at the primate laboratory of theUniversity of Wisconsin. The success ofthe procedures developed to care forthese neonates was demonstrated by thelow mortality and by a gain in weightwhich was approximately 25 percentgreater than that of infants raised bytheir own mothers. All credit for thesuccess of this program belongs to vanWagenen (16), who had described theessential procedures in detail.
These first 3 years were spent in de-vising measures to assess the multiple
capabilities of the neonatal and infantilemonkey. The studies which resulted haverevealed that the development of per-ception, learning, manipulation, explo-ration, frustration, and timidity in themacaque monkey follows a course andsequence which is very similar to that inthe human infant. The basic differencesbetween the two species appear to bethe advanced postnatal maturational sta-tus and the subsequent more rapidgrowth of the infant macaque. Probablythe most important similarities betweenthe two, in relation to the problem ofaffectional development, are character-istic responses that have been associated
with, and are considered basic to, affec-tion; these include nursing, clinging,and visual and auditory exploration.
In the course of raising these infantswe observed that they all showed astrong attachment to the cheeseclothblankets which were used to cover thewire floors of their cages. Removal ofthese cloth blankets resulted in violentemotional behavior. These responseswere not short-lived; indeed, the emo-tional disturbance lasted several days, aswas indicated by the infant's refusal towork on the standard learning tests thatwere being conducted at the time. Simi-lar observations had already been madeby Foley (17) and by van Wagenen(16), who stressed the importance ofadequate contact responses to the verysurvival of the neonatal macaque. Suchobservations suggested to us that con-tact was a true affectional variable andthat it should be possible to trace andmeasure the development and impor-tance of these responses. Indeed thereseemed to be every reason to believe thatone could manipulate all variables whichhave been considered critical to the de-velopment of the infant's attachment toa mother, or mother surrogate.To attain control over maternal vari-
ables, we took the calculated risk of con-structing and using inanimate mothersurrogates rather than real mothers. Thecloth mother that we used was a cylin-der of wood covered with a sheath ofterry cloth (18), and the wire motherwas a hardware-cloth cylinder. Initially,sponge rubber was placed underneaththe terry cloth sheath of the clothmother surrogate, and a light bulb be-hind each mother surrogate providedradiant heat. For reasons of sanitation
18
'5
UI)er0Ir
1 21
9
6
310
_ // ~~CLOTH FED_// A ~~CLOTH MOTHER
_g/ UN~~WIRE MOTHER _
/ ~~~WIRE FEo-o CLOTH MOTHERO---WIRE MOTHER
a_ = - = :_ _ - + -
145 1651-5 6- 11-Is 16-20 21-25 5 25 85 105 125MEAN AGE MEAN AGE
Time spent on cloth and wire mother surrogates. Fig. 2 (left). Short term. Fig. 3 (right). Long term.
24
21
18
15U)v12
Z 9
6
3
O
422 SCIENCE, VOL. 130
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

and safety these two factors were elimi-nated in construction of the standardmothers, with no observable effect onthe behavior of the infants. The twomothers were attached at a 45-degreeangle to aluminum bases and were givendifferent faces to assure uniqueness inthe various test situations (Fig. 1). Bot-tle holders were installed in the uppermiddle part of the bodies to permit nurs-ing. The mother was designed on thebasis of previous experience with infantmonkeys, which suggested that nursingin an upright or inclined position withsomething for the infant to clasp facili-tated successful nursing and resulted inhealthier infants (see 16). Thus, bothmothers provided the basic known re-quirements for adequate nursing, butthe cloth mother provided an additionalvariable of contact comfort. That bothof these surrogate mothers provided ade-quate nursing support is shown by thefact that the total ingestion of formulaand the weight gain was normal for allinfants fed on the surrogate mothers. Theonly consistent difference between thegroups lay in the softer stools of the in-fants fed on the wire mother.
Development of Affectional Responses
The initial experiments on the devel-opment of affectional responses havealready been reported (19) but will bebriefly reviewed here, since subsequentexperiments were derived from them. Inthe initial experiments, designed toevaluate the role of nursing on the de-velopment of affection, a cloth motherand a wire mother were placed in dif-ferent cubicles attached to the infant'sliving cage. Eight newborn monkeyswere placed in individual cages with thesurrogates; for four infant monkeys thecloth mother lactated and the wiremother did not, and for the other fourthis condition was reversed.The infants lived with their mother
surrogates for a minimum of 165 days,and during this time they were testedin a variety of situations designed tomeasure the development of affectionalresponsiveness. Differential affectionalresponsiveness was initially measured interms of mean hours per day spent onthe cloth and on the wire mothers undertwo conditions of feeding, as shown inFig. 2. Infants fed on the cloth motherand on the wire mother have highlysimilar scores after a short adaptationperiod (Fig. 3), and over a 165-dayperiod both groups show a distinct pref-21 AUGUST 1959
Fig. 4. Typical fear stimulus.
erence for the cloth mother. The per-
sistence of the differential responsive-ness to the mothers for both groups ofinfants is evident, and the over-all dif-ferences between the groups fall short ofstatistical significance.
These data make it obvious that con-
tact comfort is a variable of critical im-portance in the development of affec-tional responsiveness to the surrogatemother, and that nursing appears to playa negligible role. With increasing age
and opportunity to learn, an infant fedfrom a lactating wire mother -does not
become more responsive to her, as wouldbe predicted from a derived-drive the-ory, but instead becomes increasinglymore responsive to its nonlactating clothmother. These findings are at completevariance with a drive-reduction theoryof affectional development.The amount of time spent on the
mother does not necessarily indicate an
I100toz
CLso8(0)
LW 60
X. 40
I-
zwo20
WL 0
HOME CAGE FEARFIRST RESPONSEDUAL FED RAISED
MEAN DAY OF AGE
Fig. 5. Differential responsiveness in feartests.
affectional attachment. It could merelyreflect the fact that the cloth mother isa more comfortable sleeping platformor a more adequate source of warmthfor the infant. However, three of thefour infants nursed by the cloth motherand one of the four nursed by the wiremother left a gauze-covered heating padthat was on the floor of their cages dur-ing the first 14 days of life to spend upto 18 hours a day on the cloth mother.This suggests that differential heatingor warmth is not a critical variablewithin the controlled temperature rangeof the laboratory.
Other tests demonstrate that the clothmother is more than a convenient nest;indeed, they show that a bond developsbetween infant and cloth-mother surro-gate that is almost unbelievably similarto the bond established between humanmother and child. One highly definitivetest measured the selective maternal re-sponsiveness of the monkey infants underconditions of distress or fear.
Various fear-producing stimuli, suchas the moving toy bear illustrated in Fig.4, were presented to the infants in theirhome cages. The data on differentialresponses under both feeding conditionsare given in Fig. 5. It is apparent thatthe cloth mother was highly preferred tothe wire mother, and it is a fact thatthese differences were unrelated to feed-ing conditions-that is, nursing on thecloth or on the wire mother. Above andbeyond these objective data are obser-vations on the form of the infants' re-sponses in this situation. In spite of theirabject terror, the infant monkeys, afterreaching the cloth mother and rubbingtheir bodies about hers, rapidly come tolose their fear of the frightening stimuli.Indeed, within a minute or two most ofthe babies were visually exploring thevery thing which so shortly before hadseemed an object of evil. The bravest ofthe babies would actually leave themother and approach the fearful mon-sters, under, of course, the protectivegaze of their mothers.
These data are highly similar, interms of differential responsiveness, tothe time scores previously mentioned andindicate the overwhelming importanceof contact comfort. The results are sostriking as to suggest that the primaryfunction of nursing may be that of in-suring frequent and intimate contactbetween mother and infant, thus facili-tating the localization of the source ofcontact comfort. This interpretationfinds some support in the test discussedabove. In both situations the infants
423
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
nursed by the cloth mother developedconsistent responsiveness to the softmother earlier in testing than did theinfants nursed by the wire mother, andduring this transient period the lattergroup was slightly more responsive to thewire mother than the former group.However, these early differences shortlydisappeared.
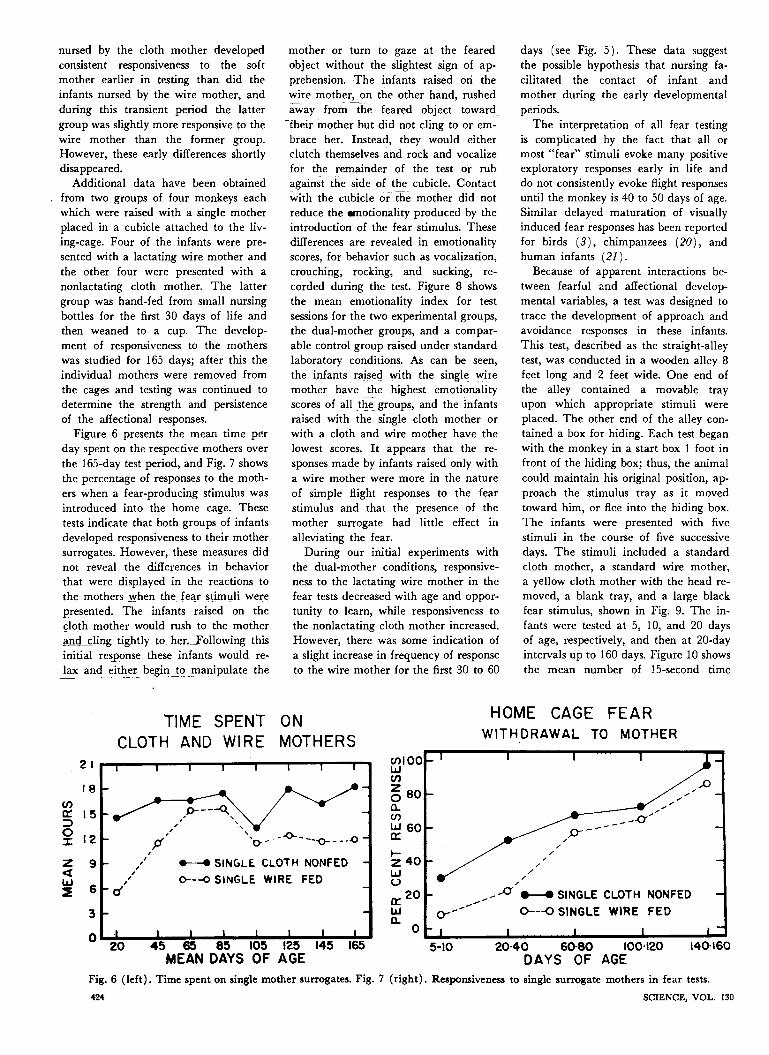
Additional data have been obtainedfrom two groups of four monkeys eachwhich were raised with a single motherplaced in a cubicle attached to the liv-ing-cage. Four of the infants were pre-sented with a lactating wire mother andthe other four were presented with anonlactating cloth mother. The lattergroup was hand-fed from small nursingbottles for the first 30 days of life andthen weaned to a cup. The develop-ment of responsiveness to the motherswas studied for 165 days; after this theindividual mothers were removed fromthe cages and testing was continued todetermine the strength and persistenceof the affectional responses.
Figure 6 presents the mean time perday spent on the respective mothers overthe 165-day test period, and Fig. 7 showsthe percentage of responses to the moth-ers when a fear-producing stimulus wasintroduced into the home cage. Thesetests indicate that both groups of infantsdeveloped responsiveness to their mothersurrogates. However, these measures didnot reveal the differences in behaviorthat were displayed in the reactions tothe mothers when the fear stimuli were
_presented. The infants raised on thecloth mother would rush to the motherand cling tightly to her.Following thisinitial response these infants would re-lax and either begin to manipulate the
mother or turn to gaze at the fearedobject without the slightest sign of ap-prehension. The infants raised on thewire mother, on the other hand, rushedaway from the feared object towardtheir mother but did not cling to or em-brace her. Instead, they would eitherclutch themselves and rock and vocalizefor the remainder of the test or rubagainst the side of the cubicle. Contactwith the cubicle or the mother did notreduce the ¬ionality produced by theintroduction of the fear stimulus. Thesedifferences are revealed in emotionalityscores, for behavior such as vocalization,crouching, rocking, and sucking, re-corded during the test. Figure 8 showsthe mean emotionality index for testsessions for the two experimental groups,the dual-mother groups, and a compar-able control group raised under standardlaboratory conditions. As can be seen,the infants raised with the single wiremother have the highest emotionalityscores of all the groups, and the infantsraised with the single cloth mother orwith a cloth and wire mother have thelowest scores. It appears that the re-sponses made by infants raised only witha wire mother were more in the natureof simple flight responses to the fearstimulus and that the presence of themother surrogate had little effect inalleviating the fear.
During our initial experiments withthe dual-mother conditions, responsive-ness to the lactating wire mother in thefear tests decreased with age and oppor-tunity to learn, while responsiveness tothe nonlactating cloth mother increased.However, there was some indication ofa slight increase in frequency of responseto the wire mother for the first 30 to 60
days (see Fig. 5). These data suggestthe possible hypothesis that nursing fa-cilitated the contact of infant andmother during the early developmentalperiods.The interpretation of all fear testing
is complicated by the fact that all ormost "fear" stimuli evoke many positiveexploratory responses early in life anddo not consistently evoke flight responsesuntil the monkey is 40 to 50 days of age.Similar delayed maturation of visuallyinduced fear responses has been reportedfor birds (3), chimpanzees (20), andhuman infants (21).
Because of apparent interactions be-tween fearful and affectional develop-mental variables, a test was designed totrace the development of approach andavoidance responses in these infants.This test, described as the straight-alleytest, was conducted in a wooden alley 8feet long and 2 feet wide. One end ofthe alley contained a movable trayupon which appropriate stimuli wereplaced. The other end of the alley con-tained a box for hiding. Each test beganwith the monkey in a start box 1 foot infront of the hiding box; thus, the animalcould maintain his original position, ap-proach the stimulus tray as it movedtoward him, or flee into the hiding box.The infants were presented with fivestimuli in the course of five successivedays. The stimuli included a standardcloth mother, a standard wire mother,a yellow cloth mother with the head re-moved, a blank tray, and a large blackfear stimulus, shown in Fig. 9. The in-fants were tested at 5, 10, and 20 daysof age, respectively, and then at 20-dayintervals up to 160 days. Figure 10 showsthe mean number of 15-second time
TIME SPENT ONCLOTH AND WIRE MOTHERS
11 . -1 9.
~~~~~'D ---00 -~
-- SINGLE CLOTH NONFED -
O-0 SINGLE WIRE FED
LI A A A A A A
20 45 65 85 lOS 125 145 165MEAN DAYS OF AGE
(/100WU)O 800~n-W 60It
z 40
c 20
a- 0b
HOME CAGE FEARWITHDRAWAL TO MOTHER
5-10 20-40 60-80 100 120DAYS OF AGE
140160
Fig. 6 (left). Time spent on single mother surrogates. Fig. 7 (right). Responsiveness to single surrogate mothers in fear tests.
424 SCIENCE, VOL. 130
U)cr
0X
z
W
2 1
18
1 5
12
9
6
3
0
.0'
p-~~~~~~~~~
_~~~~~~~~~~,M
- @--*-*SINGLE CLOTH NONFED -o ~~CO-- SINGLE WIRE FED_
1,
A, ,,
a l,A
0-1
/' \
l1
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

HOME CAGE FEARTOTAL EMOTION SCORE
INCREASE FROM NORM-MECHANICAL STIMULI
DUAL SINGLE SINGLE CONTROLSFED CLOTH WIRE
NON-FED FED
Fig. 8. Change in emotionality index infear tests.
periods spent in contact with the appro-priate mother during the 90-second testsfor the two single-mother groups, andthe responses to the cloth mother by fourinfants from the dual-mother group.
During the first 80 days of testing, allthe groups showed an increase in re-sponse to the respective mother surro-gates. The infants fed on the single wiremother, however, reached peak respon-siveness at this age and then showed aconsistent decline, followed by an actualavoidance of the wire mother. Duringtest sessions 140 to 160, only one contactwas made with the wire mother, andthree of the four infants ran into thehiding box almost immediately and re-mained there for the entire test session.On the other hand, all of the infantsraised with a cloth mother, whether ornot they were nursed by her, showed aprogressive increase in time spent in con-tact with their cloth mothers until ap-proaches and contacts during the testsessions approached maximum scores..The development of the response of
flight from the wire mother by the groupfed on the single wire mother is, ofcourse, completely contrary to a derived-drive theory of affectional development.A comparison of this group with thegroup raised with a cloth mother givessome support to the hypothesis that feed-ing or nursing facilitates the early devel-opment of responses to the mother butthat without the factor of contact com-fort, these positive responses are notmaintained.The differential responsiveness to the
cloth mother of infants raised withboth mothers, the reduced emotionalityof both the groups raised with clothmothers in the home-cage fear tests,and the development of approach re-sponses in the straight-alley test indi-
21 AUGUST 1959
cate that the cloth mother providesa haven of safety and security for thefrightened infants The affectional re-sponse patterns found in the infant mon-key are unlike tropistic or even complexreflex responses; they resemble insteadthe diverse and pervasive patterns of re-sponse to his mother exhibited by thehuman child in the complexity of situa-tions involving child-mother relation-ships.The role of the mother as a source of
safety and security has been demon-strated experimentally for human infantsby Arsenian (22). She placed children11 to 30 months of age in a strange roomcontaining toys and other play objects.Half of the children were accompaniedinto the room by a mother or a substi-tute mother (a familiar nursery attend-ant), while the other half entered thesituation alone. The children in the firstgroup (mother present) were much lessemotional and participated much morefully in the play activity than thosein the second group (mother absent).With repeated testing, the security score,a composite score of emotionality andplay behavior, improved for the chil-dren who entered alone, but it still fellfar below that for the children who wereaccompanied by their mothers. In sub-sequent tests, the children from themother-present group were placed in thetest room alone, and there was a drasticdrop in the security scores. Contrariwise,the introduction of the mother raisedthe security scores of children in theother group.We have performed a similar series of
open-field experiments, comparing mon-keys raised on mother surrogates withcontrol monkeys raised in a wire cagecontaining a cheesecloth blanket fromdays 1 to 14 and no cloth blanket sub-sequently. The infants were introducedinto the strange environment of the openfield, which was a room measuring 6 by6 by 6 feet, containing multiple stimuliknown to elicit curiosity-manipulatoryresponses in baby monkeys. The infantsraised with single mother surrogates wereplaced in this situation twice a week for8 weeks, no mother surrogate being pres-ent during one of the weekly sessionsand the appropriate mother surrogate(the kind which the experimental infanthad always known) being present dur-ing the other sessions. Four infants raisedwith dual mother surrogates and fourcontrol infants were subjected to similarexperimental sequences, the cloth motherbeing present on half of the occasions.The remaining four "duial-mother" in-
Fig. 9. Response to the fear stimulus inthe straight-alley test.
fants were given repetitive tests to ob-tain information on the development ofresponsiveness to each of the dual moth-ers in this situation. A cloth blanket wasalways available as one of the stimulithroughout the sessions. It should be em-phasized that the blanket could readilycompete with the cloth mother as a con-tact stimulus, for it was standard labora-tory procedure to wrap the infants insoft cloth whenever they were removedfrom their cages for testing, weighing,and other required laboratory activities.As soon as they were placed in the
test room, the infants raised with clothmothers rushed to their mother surro-gate when she was present and clutchedher tenaciously, a response so strong thatit can only be adequately depicted bymotion pictures. Then, as had been ob-served in the fear tests in the home cage,they rapidly relaxed, showed no sign ofapprehension, and began to demonstrateunequivocal positive responses of ma-nipulating and climbing on the mother.After several sessions, the infants beganto use the mother surrogate as a base of
TIME IN CONTACT WITH MOTHER
DAYS OF AGE
Fig. 10. Responsiveness to mother surro-gates in the 6traight-alley tests.
425
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Fig. 11. Subsequent response to cloth mother and stimulus in the open-field test.
operations, leaving her to explore andhandle a stimulus and then returning toher before going to a new plaything.Some of the infants even brought thestimuli to the mother, as shown in Fig.11. The behavior of these infantschanged radically in the absence of themother. Emotional indices such as vo-calization, crouching, rocking, and suck-ing increased sharply. Typical responsepatterns were either freezing in a
crouched position, as illustrated in Fig.12, or running aroynd the room on thehind feet, clutching themselves withtheir arms. Though no quantitative evi-dence is available, contact and manipu-lation of objects was frantic and ofshort duration, as opposed to the playfultype of manipulation observed when themother was present.
In the presence of the mother, thebehavior of the infants raised with sin-
Fig. 12. Response in the open-field test in the absence of the mother surrogate.
42.6
gle wire mothers was both quantitativelyand qualitatively different from that ofthe infants raised with cloth mothers.Not only did these infants spend littleor no time contacting their mother sur-rogates but the presence of the motherdid not reduce their emotionality. Thesedifferences are evident in the mean num-ber of time periods spent in contact withthe respective mothers, as shown in Fig.13, and the composite emotional indexfor the two stimulus conditions depictedin Fig. 14. Although the infants raisedwith dual mothers spent considerablymore time in contact with the clothmother than did the infants raised withsingle cloth mothers, their emotional re-actions to the presence and absence ofthe mother were highly similar, the com-posite emotional index being reducedby almost half when the mother was inthe test situation. The infants raised withwire mothers were highly emotionalunder both conditions and actuallyshowed a slight, though nonsignificant,increase in emotionality when the motherwas present. Although some of the in-fants reared by a wire mother did con-tact her, their behavior was similar tothat observed in the home-cage feartests. They did not clutch and cling totheir mother as did the infants withcloth mothers; instead, they sat on herlap and clutched themselves, or heldtheir heads and bodies in their arms andengaged in convulsive jerking and rock-ing movements similar to the autistic be-havior of deprived and institutionalizedhuman children. The lack of exploratoryand manipulatory behavior on the partof the infants reared with wire mothers,both in the presence and absence of thewire mother, was similar to that observedin the mother-absent condition for theinfants raised with the cloth mothers,and such contact with objects as wasmade was of short duration and of anerratic and frantic nature. None of theinfants raised with single wire mothersdisplayed the persistent and aggressiveplay behavior that was typical of manyof the infants that were raised with clothmothers.The four control infants, raised with-
out a mother surrogate, had approxi-mately the same emotionality scoreswhen the mother was absent that theother infants had in the same condition,but the control subjects' emotionalityscores were significantly higher in thepresence of the mother surrogate than inher absence. This result is not surpris-ing, since recent evidence indicates thatthe cloth mother with the highly orna-mental face is an effective fear stimulus
SCIENCE, VOL. 130
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
MEAN TIME PERIODS INCONTACT WITH MOTHER
DUAL SINGLE SINGLEFED CLOTH WIRE
NONFED FED
CONTROL
OPEN FIELDCOMPOSITE EMOTIONAL INDEX
w 500-4
w31-2zLu
DUAL FED SINGLE SINGLECLOTH WIRE
NONFED FED
Fig. 13 (left). Responsiveness to mother surrogates in the open-field test. Fig. 14 (right). Emotionality index in testing with and with-out the mother surrogates.
for monkeys that have not been raised
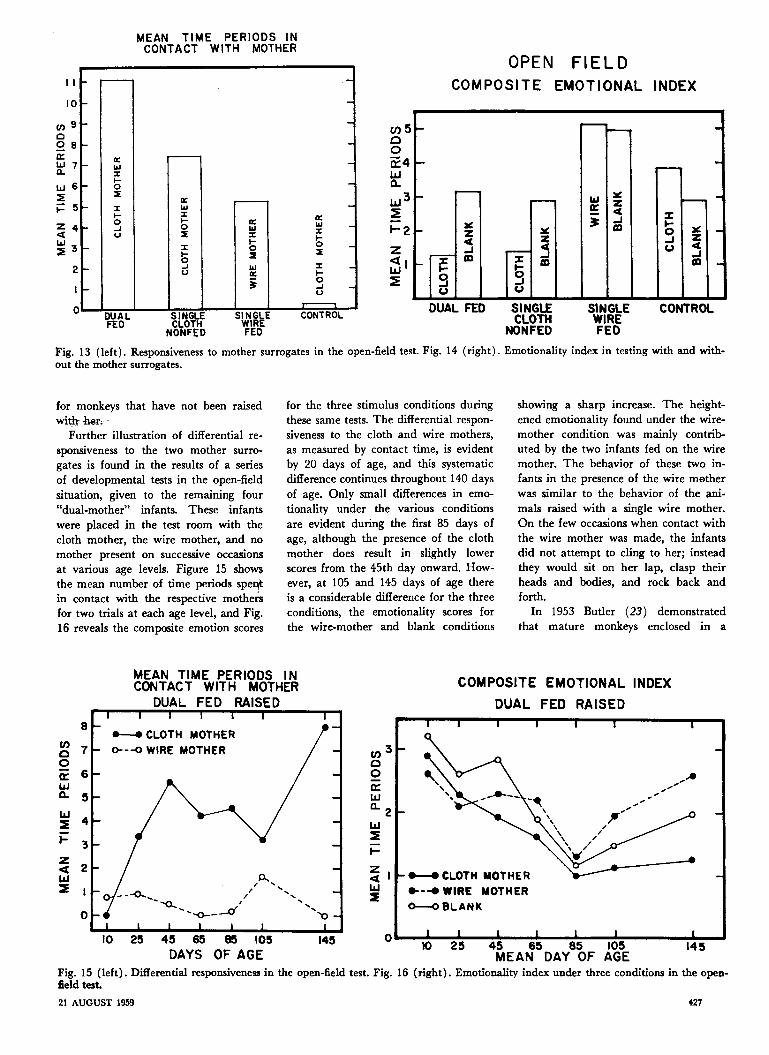
Further illustration of differential re-sponsiveness to the two mother surro-gates is found in the results of a seriesof developmental tests in the open-fieldsituation, given to the remaining four"dual-mother" infants. These infantswere placed in the test room with thecloth mother, the wire mother, and nomother present on successive occasionsat various age levels. Figure 15 showsthe mean number of time periods spertin contact with the respective mothersfor two trials at each age level, and Fig.16 reveals the composite emotion scores
for the three stimulus conditions duringthese same tests. The differential respon-siveness to the cloth and wire mothers,as measured by contact time, is evidentby 20 days of age, and this systematicdifference continues throughout 140 daysof age. Only small differences in emo-tionality under the various conditionsare evident during the first 85 days ofage, although the presence of the clothmother does result in slightly lowerscores from the 45th day onward. How-ever, at 105 and 145 days of age thereis a considerable difference for the threeconditions, the emotionality scores forthe wire-mother and blank conditions
showing a sharp increase. The height-ened emotionality found under the wire-mother condition was mainly contrib-uted by the two infants fed on the wiremother. The behavior of these two in-fants in the presence of the wire motherwas similar to the behavior of the ani-mals raised with a single wire mother.On the few occasions when contact withthe wire mother was made, the infantsdid not attempt to cling to her; insteadthey would sit on her lap, clasp theirheads and bodies, and rock back andforth.
In 1953 Butler (23) demonstratedthat mature monkeys enclosed in a
MEAN TIME PERIODS INCONTACT WITH MOTHER
DUAL FED RAISEDCOMPOSITE EMOTIONAL INDEX
DUAL FED RAISED
u) 3
W-
ZL
z41
Wu
10 25 45 65 85 105 145
DAYS OF AGE ~~~~~~1025 45 65 85 105 145DAYS OF AGE o lo 25 4MEAN DAY OF AGEFig. 15 (left). Differential responsiveness in the open-field test. Fig. 16 (right). Emotionality index under three conditions in the open-
field test.
21 AUGUST 1959 427
I0
) 900 8
W 7
W, 60~
:5
Z 4
W32
0
X Wa3.- Xz(o I- cca
_i ~ 0 W XU X
z 0k_04i WXU cc -
0Fi _4U-
1 A ra .-0 - - -^
z
E4K 4x0~ ~ ~~:x_1- 1 0094 J
4J
CONTROL
U,)0
0
CL
z
WU
8
7
6
5
4
3
2
0
A I I
*-@ CLOTH MOTHER- 0-0 WIRE MOTHER
/~~~~~~~
£ I--A l a A A
PI-,
'o -
A/
- Ag1 rI I I I`
- CLOTH MOTHER*--- WIRE MOTHER0---- BLANK
a .la a a .a
1
a a
1
a
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

dimly lighted box would open and re-open a door for hours on end with noother motivation than that of lookingoutside the box. He also demonstratedthat rhesus monkeys showed selectivityin rate and frequency of door-openingin response to stimuli of different de-grees of attractiveness (24). We haveutilized this characteristic of responseselectivity on the part of the monkey tomeasure the strength of affectional re-sponsiveness of the babies raised withmother surrogates in an infant versionof the Butler box. The test sequence in-volves four repetitions of a test batteryin which the four stimuli of clothmother, wire mother, infant monkey,and empty box are presented for a 30-minute period on successive days. Thefirst four subjects raised wth the dualmother surrogates and the eight infantsraised with single mother surrogateswere given a test sequence at 40 to 50days of age, depending upon the avail-ability of the apparatus. The data ob-tained from the three experimentalgroups and a comparable control groupare presented in Fig. 17. Both groups ofinfants raised with cloth mothers showedapproximately equal responsiveness tothe cloth mother and to another infantmonkey, and no greater responsivenessto the wire mother than to an emptybox. Again, the results are independentof the kind of mother that lactated,cloth or wire. The infants raised withonly a wire mother and those in thecontrol group were more highly respon-sive to the monkey than to either of themother surrogates. Furthermore, theformer group showed a higher frequencyof response to the empty box than tothe wire mother.
In summary, the experimental analy-sis of the development of the infantmonkey's attachment to an inanimatemother surrogate demonstrates the over-whelming importance of the variable ofsoft body contact that characterized thecloth mother, and this held true for theappearance, development, and mainte-nance of the infant-surrogate-mother tie.The results also indicate that, withoutthe factor of contact comfort, only aweak attachment, if any, is formed.Finally, probably the most surprisingfinding is that nursing or feeding playedeither no role or a subordinate role inthe development of affection as meas-ured by contact time, responsiveness tofear, responsiveness to strangeness, andmotivation to seek and see. No evidencewas found indicating that nursing medi-ated the development of any of theseresponses, although there is evidence in-
428
dicating that feeding probably facilitatedthe early appearance and increased theearly strength of affectional responsive-ness. Certainly feeding, in contrast tocontact comfort, is neither a necessarynor a sufficient condition for affectionaldevelopment.
300_
CL200
WI 100
XW X g W
uZs21L. .iO0 c
DUAL FED SINGLE SINGLE CONTROLSWIRE FED CLOTH FED
Fig. 17. Differential responses to visualexploration.
onU,U,
-J
z4tW&
VISUAL EXPLORATION RETENTIONDUAL FED
ovL I
- CLOTH MOTHER0-O WIRE MOTHER--- BLANK
200 _
100 _ K >Zz--za-
II
ORIGINAL 0-1 2-3 4-5 6-9 . 145RETENTION MONTH
Fig. 18. Retention of differential visual-exploration responses.
LOVE MACHINEMEAN LEVER PRESSESa f 1- A 1
350
3001-
CoW250COWX 200
W 150W
Z 1004tW
50c
o
ORIGINAL 09 30.39 6069 099DAYS OF RETENTION
Fig. 19. Retention of differential visual-exploration responses by single-surrogateinfants.
Retention of Affectional Responses
One of the outstanding characteristicsof the infant's attachment to its motheris the persistence of the relationship overa period of years, even though the fre-quency of contact between infant andmother is reduced with increasing age.In order to test the persistence of theresponsiveness of our "mother-surrogate"infants, the first four infant monkeysraised with dual mothers and all of themonkeys raised with single mothers wereseparated from their surrogates at 165to 170 days of age. They were tested foraffectional retention during the follow-ing 9 days, then at 30-day intervals dur-ing the following year. The results areof necessity incomplete, inasmuch asthe entire mother-surrogate program wasinitiated less than 2 years ago, butenough evidence is available to indicatethat the attachment formed to the clothmother during the first 6 months of lifeis enduring and not easily forgotten.
Affectional retention as measured bythe modified Butler box for the first 15months of testing for four of the infantsraised with two mothers is given in Fig.18. Although there is considerable varia-bility in the total response frequencyfrom session to session, there is a con-sistent difference in the number of re-sponses to the cloth mother as contrastedwith responses to either the wire motheror the empty box, and there is no con-sistent difference between responses tothe wire mother and to the empty box.The effects of contact comfort versusfeeding are dramatically demonstratedin this test by the monkeys raised witheither single cloth or wire mothers. Fig-ure 19 shows the frequency of responseto the appropriate mother surrogate andto the blank box during 'the presepara-tion period and the first 90 days of re-tention testing. Removal of the motherresulted in a doubling of the frequencyof response to the cloth mother andmore than tripled the difference be-tween the responses to the cloth motherand those to the empty box for the in-fants that had lived with a single non-lactating cloth mother surrogate. Theinfants raised with a single lactating wiremother, on the other hand, not onlyfailed to show any consistent preferencefor the wire mother but also showed ahighly significant reduction in generallevel of responding. Although incom-plete, the data from further retentiontesting indicate that the difference be-tween these two groups persists for atleast 5 months.
Affectional retention was also tested
SCIENCE, VOL. 130
I_
SINGI- CLOTH NONFEDW-_FLOTH MOTHER-0-4 BLANK
- SINGLE WIRE E.D -00 WIRE MOTHER
,O- --0 PLANKa9 9 5 5
AL
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
FREE FIELD RETENTIONMEAN TIME PERIODS INCONTACT WITH MOTHER
12
10to9
U) 9In°a
cl 7
aW 6
5
t._
Z4
3
2
0
l l IA IORIGINAL 0 30-39 6069 909
DAYS OF RETENTION
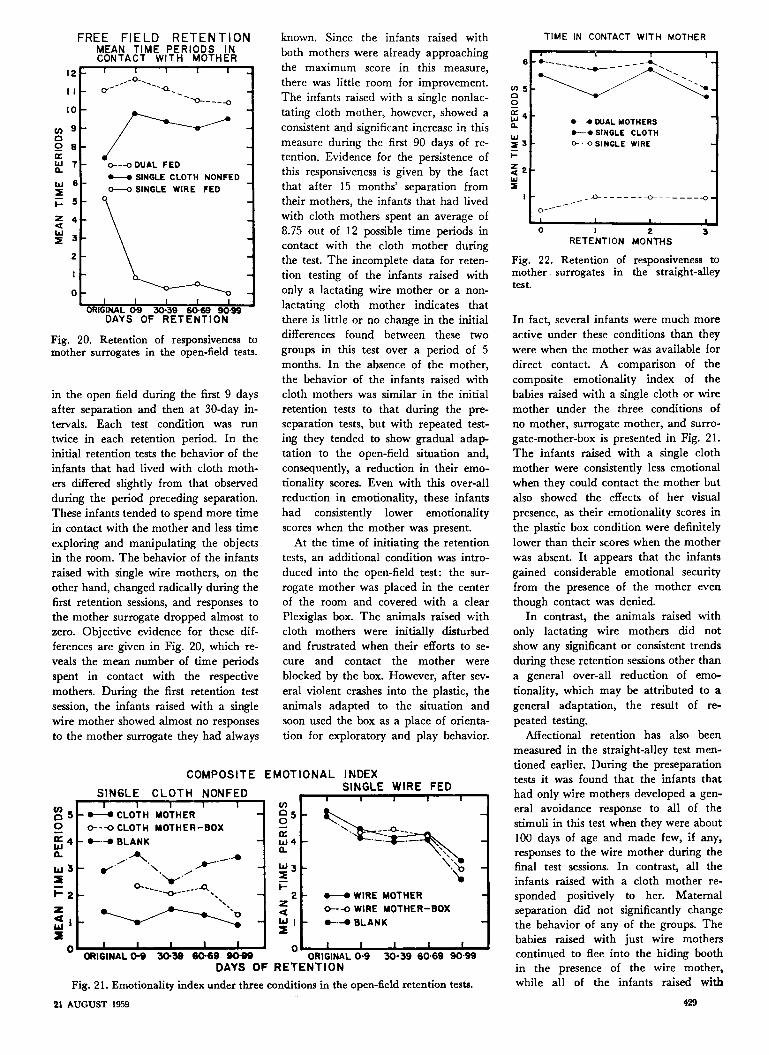
Fig. 20. Retention of responsiveness tomother surrogates in the open-field tests.
in the open field during the first 9 daysafter separation and then at 30-day in-tervals. Each test condition was run
twice in each retention period. In theinitial retention tests the behavior of theinfants that had lived with cloth moth-ers differed slightly from that observedduring the period preceding separation.These infants tended to spend more timein contact with the mother and less timeexploring and manipulating the objectsin the room. The behavior of the infantsraised with single wire mothers, on theother hand, changed radically during thefirst retention sessions, and responses tothe mother surrogate dropped almost tozero. Objective evidence for these dif-ferences are given in Fig. 20, which re-
veals the mean number of time periodsspent in contact with the respectivemothers. During the first retention testsession, the infants raised with a singlewire mother showed almost no responses
to the mother surrogate they had always
Ul)0 50
IdCC4
h3a
2
z
29
COMPOSITESINGLE CLOTH NONFED
- *---*CLOTH MOTHERo-o CLOTH MOTHER-BOX
- *---4 BLANK
0-=-- -~
a a A IA I
known. Since the infants raised withboth mothers were already approachingthe maximum score in this measure,
there was little room for improvement.The infants raised with a single nonlac-tating cloth mother, however, showed a
consistent and significant increase in thismeasure during the first 90 days of re-
tention. Evidence for the persistence ofthis responsiveness is given by the factthat after 15 months' separation fromtheir mothers, the infants that had livedwith cloth mothers spent an average of8.75 out of 12 possible time periods incontact with the cloth mother duringthe test. The incomplete data for reten-
tion testing of the infants raised withonly a lactating wire mother or a non-
lactating cloth mother indicates thatthere is little or no change in the initialdifferences found between these two
groups in this test over a period of 5months. In the absence of the mother,the behavior of the infants raised withcloth mothers was similar in the initialretention tests to that during the pre-
separation tests, but with repeated test-
ing they tended to show gradual adap-tation to the open-field situation and,consequently, a reduction in their emo-
tionality scores. Even with this over-allreduction in emotionality, these infantshad consistently lower emotionalityscores when the mother was present.
At the time of initiating the retentiontests, an additional condition was intro-duced into the open-field test: the sur-
rogate mother was placed in the centerof the room and covered with a clearPlexiglas box. The animals raised withcloth mothers were initially disturbedand frustrated when their efforts to se-
cure and contact the mother were
blocked by the box. However, after sev-
eral violent crashes into the plastic, theanimals adapted to the situation andsoon used the box as a place of orienta-tion for exploratory and play behavior.
EMOTIONAL INDEXSINGLE WIRE FED
U)C/)
cwId
I'
&4
CL
ORIGINAL 0-9 339 69 90-99 ORIGINAL 0-9 30-39 60-69 9099DAYS OF RETENTION
Fig. 21. Emotionality index under three conditions in the open-field retention tests.
21 AUGUST 1959
TIME IN CONTACT WITH MOTHER
AA.6~
U)500.
0:4
3pz2
Wa
0 32RETENTION MONTHS
Fig. 22. Retention of responsiveness tomother surrogates in the straight-alleytest.
In fact, several infants were much more
active under these conditions than theywere when the mother was available fordirect contact. A comparison of thecomposite emotionality index of thebabies raised with a single cloth or wiremother under the three conditions ofno mother, surrogate mother, and surro-
gate-mother-box is presented in Fig. 21.The infants raised with a single clothmother were consistently less emotionalwhen they could contact the mother butalso showed the effects of her visualpresence, as their emotionality scores inthe plastic box condition were definitelylower than their scores when the motherwas absent. It appears that the infantsgained considerable emotional securityfrom the presence of the mother even
though contact was denied.In contrast, the animals raised with
only lactating wire mothers did notshow any significant or consistent trendsduring these retention sessions other thana general over-all reduction of emo-
tionality, which may be attributed to a
general adaptation, the result of re-
peated testing.Affectional retention has also been
measured in the straight-alley test men-
tioned earlier. During the preseparationtests it was found that the infants thathad only wire mothers developed a gen-
eral avoidance response to all of thestimuli in this test when they were about100 days of age and made few, if any,
responses to the wire mother during thefinal test sessions. In contrast, all theinfants raised with a cloth mother re-
sponded positively to her. Maternalseparation did not significantly changethe behavior of any of the groups. Thebabies raised with just wire motherscontinued to flee into the hiding boothin the presence of the wire mother,while all of the infants raised with
429
I I
* 0 - -_ 0 =
o-o DUAL FED*- SINGLE CLOTH NONFED0-0 SINGLE WIRE FED
__---- --- '
"0-
v - 4DUAL MOTHERS-- SINGLE CLOTH
0- SINGLE WIRE
0-_-
5 t4
3
2 - WIRE MOTHERO---0 WIRE MOTHER-BOX
I - --- BLANK
O I IA a _
5
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Fig. 23. Typical response to cloth mother in the modified open-field test.
cloth mothers continued to respondpositively to the cloth mother at ap-
proximately the same level as in thepreseparation tests. The mean numberof time periods spent in contact with theappropriate mother surrogates for thefirst 3 months of retention testing are
given in Fig. 22. There is little, if any,
waning of responsiveness to the clothmother during these 3 months. Thereappeared to be some loss of responsive-ness to the mother in this situation after5 to 6 months of separation, but the test
was discontinued at that time as the in-fants had outgrown the apparatus.The retention data from these mul-
tiple tests demonstrate clearly the im-portance of body contact for the futuremaintenance of affectional responses.
Whereas several of the measures in thepreseparation period suggested that theinfants raised with only a wire mothermight have developed a weak attach-ment to her, all responsiveness disap-peared in the first few days after themother was withdrawn from the living-cage. Infants that had had the oppor-
tunity of living with a cloth mothershowed the opposite effect and eitherbecame more responsive to the clothmother or continued to respond to herat the same level.
These data indicate that once an af-fectional bond is formed it is maintainedfor a very considerable length of timewith little reinforcement of the contact-
comfort variable. The limited data avail-able for infants that have been sepa-
rated from their mother surrogates fora year suggest that these affectional re-
430
sponses show resistance to extinctionsimilar to the resistance previously dem-onstrated for learned fears and learnedpain. Such data are in keeping withcommon observation of human behavior.
It is true, however, that the infantsraised with cloth mothers exhibit some
absolute decrease in responsiveness withtime in all of our major test situations.Such results would be obtained even ifthere were no true decrease in thestrength of the affectional bond, be-cause of familiarization and adaptationresulting from repeated testing. There-fore, at the end of 1 year of retentiontesting, new tests were introduced intothe experimental program.
Our first new test was a modificationof the open-field situation, in whichbasic principles of the home-cage feartest were incorporated. This particularchoice was made partly because the lat-ter test had to be discontinued when themother surrogates were removed fromthe home cages.
For the new experiment a Masonitefloor marked off in 6- by 12-inch rec-
tangles was placed in the open-fieldchamber. Both mother surrogates were
placed in the test room opposite a
plastic start-box. Three fear stimuli, se-
lected to produce differing degrees ofemotionality, were placed in the centerof, the room directly in front of the start-box in successive test sessions. Eighttrials were run under each stimulus con-
dition, and in half of the trials the mostdirect path to the cloth mother was
blocked by a large Plexiglas screen, il-lustrated in Fig. 23. Thus, in these trials
the infants were forced to approach andbypass the fear stimulus or the wiremother, or both, in order to reach thecloth mother. Following these 24 trialswith the mothers present, one trial ofeach condition with both mothers ab-sent was run, and this in turn was fol-lowed by two trials run under the mostemotion-provoking condition: with a me-chanical toy present and the direct pathto the mother blocked.We now have complete data for the
first four infants raised with both a clothand a wire mother. Even with this scantyinformation, the results are obvious. Aswould be predicted from our othermeasures, the emotionality scores forthe three stimuli were significantly dif-ferent and these same scores were in-creased greatly when the direct path tothe mother was blocked. A highly sig-nificant preference was shown for thecloth mother under both conditions (di-rect and blocked path), although thepresence of the block did increase thenumber of first responses to the wiremother from 3 to 10 percent. In allcases this was a transient response andthe infants subsequently ran on to thecloth mother and clung tightly to her.Objective evidence for this overwhelm-ing preference is indicated in Fig. 24,which shows the mean number of timeperiods spent in contact with the twomothers. After a number of trials, theinfants would go first to the cloth motherand then, and only then, would go outto explore, manipulate, and even attackand destroy the fear stimuli. It was asif they believed that their mother wouldprotect them, even at the cost of her life-little enough to ask in view of her con-dition.The removal of the mother surrogates
from the situation produced the pre-dictable effect of doubling the emotion-ality index. In the absence of the moth-ers, the infants would often run to thePlexiglas partition which formerly hadblocked their path to the mother, orthey would crouch in the corner behindthe block where the mother normallywould have been. The return of themothers in the final two trials of thetest in which the most emotion-evokingsituation was presented resulted in be-havior near the normal level, as meas-ured by the emotionality index and con-tacts with the cloth mother.Our second test of this series was de-
signed to replace the straight-alley testdescribed above and provide morequantifiable data on responsiveness tofear stimuli. The test was conducted inan alley 8 feet long and 2 feet wide.
SCIENCE, VOL. 130
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

At one end of the alley and directlybehind the monkeys' restraining cham-ber was a small stimulus chamber whichcontained a fear object. Each trial wasinitiated by raising an opaque slidingdoor which exposed the fear stimulus.Beginning at a point 18 inches from therestraining chamber, the alley was di-vided lengthwise by a partition; thisprovided the infant with the choice ofentering one of two alleys.The effects of all mother combina-
tions were measured; these combina-tions included no mothers, two clothmothers, two wire mothers, and a clothand a wire mother. All mother con-ditions were counterbalanced by twodistance conditions-distances of 24and 78 inches, respectively, from therestraining chamber. This made it pos-sible, for example, to provide the in-fant with the alternative of running tothe cloth mother which was in closeproximity to the fear stimulus or to thewire mother (or no mother) at a greaterdistance from the fear stimulus. Thus,it was possible to distinguish betweenrunning to the mother surrogate as anobject of security, and generalized flightin response to a fear stimulus.
Again, the data available at this timeare from the first four infants raisedwith cloth and wire mothers. Neverthe-less, the evidence is quite conclusive. Ahighly significant preference is shownfor the cloth mother as compared to thewire mother or to no mother, and thispreference appears to be independent ofthe proximity of the mother to the fearstimulus. In the condition in which twocloth mothers are present, one 24 inchesfrom the fear stimulus and the other 78inches from it, there was a preferencefor the nearest mother, but the differ-ences were not statistically significant.In two conditions in which no clothmother was present and the infant hadto choose between a wire mother andno mother, or between two empty cham-bers, the emotionality scores were almosttwice those under the cloth-mother-pres-ent condition.No differences were found in either
of these tests that were related to previ-ous conditions of feeding-that is, towhether the monkey had nursed on thecloth or on the wire mother.The results of these two new tests, in-
troduced after a full year's separationof mother surrogate and infant, are com-parable to the results obtained duringthe preseparation period and the earlyretention testing. Preferential responsesstill favored the cloth as compared tothe wire mother by as much as 85 to
21 AUGUST 1959
TIME IN CONTACT WITH MOTHERSDUAL FED
6
(0
Z2LW
CLOTH WIREMOTHER MOTHER
Fig. 24. Differential responsiveness in themodified open-field test.
90 percent, and the emotionality scoresshowed the typical 2: 1 differential ratiowith respect to mother-absent andmother-present conditions.The researches presented here on the
analysis of two affectional variablesthrough the use of objective and ob-servational techniques suggest a broadnew field for the study of emotionaldevelopment of infant animals. Theanalogous situations and results foundin observations and study of human in-fants and of subprimates demonstratethe apparent face validity of our tests.The reliability of our observational tech-niques is indicated, for example, by thecorrelation coefficients computed for thecomposite emotional index in the open-field test. Four product-moment correla-tion coefficients, computed from foursamples of 100 observations by five dif-ferent pairs of independent observersover a period of more than a year,ranged from .87 to .89.
Additional Variables
Although the overwhelming impor-tance of the contact variable has beenclearly demonstrated in these experi-ments, there is reason to believe thatother factors may contribute to the de-velopment of the affectional responsepattern. We are currently conducting aseries of new experiments to test someof these postulated variables.
For example, Bowlby (14) has sug-gested that one of the basic affectionalvariables in the primate order is notjust contact but clinging contact. Totest this hypothesis, four infant mon-keys are being raised with the standardcloth mother and a flat inclined plane,
tightly covered with the same type ofcloth. Thus, both objects contain thevariable of contact with the soft cloth,but the shape of the mother tends tomaximize the clinging variable, whilethe broad flat shape of the plane tendsto minimize it. The preliminary resultsfor differences in responsiveness to thecloth mother and responsiveness to theinclined plane under conditions thatproduce stress or fear or visual explor-ation suggest that clinging as well ascontact is an affectional variable of con-siderable importance.
Experiments now in progress on therole of rocking motion in the develop-ment of attachment indicate that thismay be a variable of measurable im-portance. One group of infants is beingraised on rocking and stationary moth-ers and a second group, on rocking andstationary inclined planes. Both groupsof infants show a small but consistentpreference for the rocking object, asmeasured in average hours spent on thetwo objects.
Preliminary results for these threegroups in the open-field test give addi-tional evidence concerning the variableof clinging comfort. These data revealedthat the infants raised with the stand-ard cloth mother were more responsiveto their mothers than the infants raisedwith inclined planes were to the planes.The discovery of three variables of
measurable importance to the formationand retention of affection is not surpris-ing, and it is reasonable to assume thatothers will be demonstrated. The dataso far obtained experimentally are inexcellent concordance with the affec-tional variables named by Bowlby (14).We are now planning a series of studiesto assess the effects of consistency andinconsistency with respect to the mothersurrogates in relation to the clinical con-cept of rejection. The effects of early,intermediate, and late maternal depri-vation and the generalization of the in-fant-surrogate attachment in social de-velopment are also being investigated.Indeed, the strength and stability ofthe monkeys' affectional responses to amother surrogate are such that it shouldbe practical to determine the neurolog-ical and biochemical variables that un-derlie love.
References and Notes
1.2.
3.4.
5.6.
K. Lorenz, Auk 54, 245 (1937).R. A. Hinde, W. H. Thorpe, M. A. Vince,Behaviour 9, 214 (1956).E. Fabricius, Acta Zool. Fennica 68, 1 (1951).E. H. Hess, 1. Comp. and Physiol. Psychol.,in press.J. Jaynes, ibid., in press.H. Moltz and L. Rosenblum, ibid. 51, 658(1958).
431
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

7. C. R. Carpenter, Comp. Psychol. MonographNo. 10 (1934), p. 1.
8. A. Nolte, Z. Tierpsychol. 12, 77 (1955).9. S. Zuckerman, Functional Affinities of Man,
Monkeys and Apes (Harcourt Brace, London,1933).
10. W. Kohler, The Mentality of Apes (Humani-ties Press, New York, 1951).
11. R. M. Yerkes and M. I. Tomilin, 1. Comp.Psychol. 20, 321 (1935).
12. J. Dollard and N. E. Miller, Personality andPsychotherapy (McGraw-Hill, New York,1950), p. 133; P. H. Mussen and J. J. Con-ger, Child Development and Personality(Harper, New York, 1956), pp. 137, 138.
13. M. A. Ribble, The Rights of Infants (Co-
lumbia Univ. Press, New York, 1943); D. W.Winnicott, Brit. J. Med. Psychol. 21, 229(1948).
14. J. Bowlby, Intern. J. Psycho-Analysis 39,part 5 (1958).
15. Support for the research presented in thisarticle was provided through funds receivedfrom the graduate school of the University ofWisconsin; from grant M-772, National Insti-tutes of Health; and from a Ford Foundationgrant.
16. G. van Wagenen, in The Care and Breedingof Laboratory Animals, E. J. Farris, Ed.(Wiley, New York, 1950), p. 1.
17. J. P. Foley, Jr., J. Genet. Psychol. 45, 39(1934).
18. We no longer make the cloth mother out ofa block of wood. The cloth mother's body issimply that of the wire mother, covered by aterry-cloth sheath.
19. H. F. Harlow, Am. Psychologist 13, 673(1958); and R. R. Zimmermann, Proc.Am. Phil. Soc. 102, 501 (1958).
20. D. 0. Hebb, The Organization of Behavior(Wiley, New York, 1949), p. 241 ff.
21. A. T. Jersild and F. B. Holmes, Child De-velop. Monograph No. 20 (1935), p. 356.
22. J. M. Arsenian, J. Abnormal Social Psychol.38, 225 (1943).
23. R. A. Butler, 1. Comp. 9nd Physiol. Psychol.46, 95 (1953).
24. , J. Exptl. Psychol. 48, 19 (1954).
A number of cell lines derived fromnormal and malignant tissues have nowbeen serially propagated in culture.These may be grown adherent to a glasssurface and overlaid with a fluid me-dium, or they may be grown in suspen-sion; but in either case the opportunityis presented for the study of metabolismat the cellular level in a system whichoperationally resembles bacterial cul-tures in most essential respects. The cellsand the medium can be separately ana-lyzed, balance experiments can be set up,metabolic processes can be examinedqualitatively and quantitatively undercontrolled conditions, and the corre-sponding enzymatic activities can be ex-plored in cell-free extracts.A relatively limited number of metab-
olites have been shown to suffice for theapparently indefinite propagation of allthe human cell lines so far studied. Theminimal medium, in which every com-ponent is demonstrably essential, is listedin Table 1 and includes 29 components:13 amino acids, 8 vitamins, 6 ionic spe-cies, glucose, and serum protein (1, 2).The role of the serum protein is not en-tirely clear. Although a few cell lineshave been serially propagated in a pro-tein-free medium (3), serum protein isrequired by most mammalian cell cul-tures. In monolayer cultures, the proteinplays a role in the adhesion of the cellsto glass (4). However, its essentiality in
432
suspension cultures indicates that this isnot its only function, and it seems rea-sonable to assume that it acts in part asa carrier of as yet unidentified growthfactors which are bound to the protein,and which are slowly released into themedium.The present article, dealing with the
amino acid metabolism of these culturedcells, is a progress report rather than areview and in large part summarizesstudies from a single laboratory. Theenormous body of information availablewith respect to amino acid metabolismin bacterial cultures (5, 6) has served asa stimulus and prototype for the studieshere reported. The relevant findings inbacterial systems have not been referredto in detail, only because that importantexercise in comparative biochemistry isbeyond the scope of the present report.
Nutritionally Essential Amino Acids
The classic experiments of Rose andhis coworkers (7) have shown that eightamino acids (isoleucine, leucine, lysine,methionine, phenylalanine, threonine,tryptophan, and valine) suffice for nitro-gen balance in feeding experiments inman. In contrast, -every cell culture sofar examined, whether human or animal(8) in origin, and whether deriving fromnormal or malignant tissue (9), has re-
quired at least 13 amino acids for sur-vival and growth. Over and above theeight amino acids required for nitrogenbalance, these cell cultures require argi-nine, cyst(e)ine, glutamine, histidine,and tyrosine, and on the omission of anyone of these, the cells degenerate anddie. In their early stages these degenera-tive processes are reversible. This pre-sents an opportunity to explore the inti-mate structural derangement caused byspecific amino acid deficiences, and thereparative processes which occur on res-toration of these amino acids to themedium.None of the D-amino acids substitute
for the L-isomer (8) except for D-cystine,and this apparently acts by mobilizingcyst (e)ine residues bound to the serumprotein of the medium. (This is dis-cussed below.) Dipeptides were foundto be active, substituting for both com-ponent amino acids (10). Recent ex-periments indicate that, although dipep-tides are hydrolysed extracellularly byserum peptidases, and perhaps by cell-derived peptidases, some of the dipep-tide is transported into the cell and therehydrolyzed.A number of keto acids and other
amino acid congeners have been testedwith respect to their ability to substitutefor the corresponding amino acid (11).The results with several human and ani-mal cell lines are summarized in Table2. The cells contain a wide variety oftransaminases (12), and, as determinedin feeding experiments in rats (5), mostof these keto acids do in fact substitutefor the corresponding amino acid.Of particular interest is the fact that,
with all the human cell lines studied,citrulline substituted for arginine, whileornithine was inactive [Table 2; see also(13)]. Attempts to encourage citrullinebiosynthesis by progressive removal of
- The author is chief of the laboratory of cellbiology, National Institute of Allergy and In-fectious Diseases, National Institutes of Health,Bethesda, Md.
SCIENCE, VOL. 130
Amino Acid Metabolism inMammalian Cell Cultures
Harry Eagle
on
Apr
il 22
, 200
7 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from





























