Embed Size (px)
Citation preview

Copyright 2001 by the Genetics Society of America
The Formamidase Gene of Aspergillus nidulans: Regulation byNitrogen Metabolite Repression and Transcriptional
Interference by an Overlapping Upstream Gene
James A. Fraser, Meryl A. Davis and Michael J. Hynes
Department of Genetics, University of Melbourne, Victoria, 3010 Australia
Manuscript received June 19, 2000Accepted for publication September 18, 2000
ABSTRACTThe ability to utilize formamide as a sole nitrogen source has been found in numerous fungi. We have
cloned the fmdS gene encoding a formamidase from Aspergillus nidulans and found that it belongs to ahighly conserved family of proteins separate from the major amidase families. The expression of fmdS isprimarily regulated via AreA-mediated nitrogen metabolite repression and does not require the additionof exogenous inducer. Consistent with this, deletion analysis of the 59 region of fmdS has confirmed thepresence of multiple AreA-binding sites containing a characteristic core GATA sequence. Under carbonstarvation conditions the response to nitrogen starvation is eliminated, indicating that the lack of a carbonsource may result in inactivation of AreA. Sequence analysis and isolation of cDNAs show that a gene ofunknown function lies directly 59 of fmdS with its transcript overlapping the fmdS coding region. Disruptionof the 59 gene and analysis of the effects of overexpression of this gene on fmdS expression has shownthat expression of this upstream gene interferes with fmdS transcription, resulting in a strong dependenceon AreA activation for expression. Therefore the relative position of these two genes is essential for normalregulation of fmdS.
THE hydrolysis of amides via amidase activities has shown to be highly regulated in response to nitrogenbeen characterized in the filamentous fungus Asper- levels in the cell through nitrogen metabolite repres-
gillus nidulans (Hynes 1975a; Hynes and Pateman sion, which results in the preferential utilization of the1970). A wide variety of amides can be hydrolyzed via nitrogen sources ammonium and l-glutamine (for aa number of distinct enzyme activites (Hynes 1975a). review, see Marzluf 1997). In A. nidulans the positivelyThe acetamidase enzyme encoded by the amdS gene acting product of the areA gene is the major regulatoryallows growth on acetamide as a carbon and/or nitrogen factor involved in this control mechanism (Arst andsource by producing ammonium and acetate. Regula- Cove 1973; Kudla et al. 1990). Loss-of-function areAtion of amdS expression is complex, involving multiple mutants are unable to grow on most nitrogen sourcesinduction signals as well as control by carbon and nitro- other than ammonium, including formamide (Hynesgen metabolites (for a review, see Hynes and Davis 1972; Arst and Cove 1973; Hynes 1975b). In the1996). The general amidase encoded by gmdA contrib- absence of a preferred nitrogen source this GATA zinc-utes to growth on longer chain amides, producing am- finger-type DNA-binding protein recognizes and acti-monium and the corresponding carboxylic acid (Hynes vates expression through the consensus sequence1975a). A formamidase enzyme (formamide amidohy- HGATAR in the promoters of over 100 genes encodingdrolase, EC 2.5.1.49) mediates the highly specific hydro- permeases and enzymes required for the catabolism oflysis of formamide to produce formate and ammonium secondary nitrogen sources (Wilson and Arst 1998).(Hynes 1975a). Formamide therefore serves only as a Regulation by AreA gives rise to a hierarchy of genenitrogen source as formate is a single carbon molecule expression, with the expression of some genes beingand hence not a carbon source for Aspergillus spp. The more highly activated than others. Mutations within thefmdS gene on chromosome III was defined by isolation DNA-binding domain of AreA have been isolated thatof the formamide nonutilizing fmdS1 mutant lacking alter this hierarchy. The areA102 (L683V) mutationdetectable formamidase activity (Hynes and Pateman gives rise to a product that has a higher affinity for1970). TGATAR sites, allowing stronger growth on acetamide
In filamentous fungi the activities of many nitrogen and reduced growth on uric acid (Hynes 1972, 1975b;catabolic enzymes, including the amidases, have been Kudla et al. 1990; Ravagnani et al. 1997). Analysis of
the amdS promoter has revealed the presence of TGA-TAR sites (Corrick et al. 1987), while the promoters
Corresponding author: Michael J. Hynes, Department of Genetics, of the purine permeases contain primarily CGATARUniversity of Melbourne, Victoria, 3010 Australia.E-mail: [email protected] sites (Gorfinkiel et al. 1993; Diallinas et al. 1995).
Genetics 157: 119–131 ( January 2001)

120 J. A. Fraser, M. A. Davis and M. J. Hynes
TABLE 1Mirror-image mutants have been isolated (L683M) thatshow an opposite-yet-weaker phenotype with better A. nidulans strains used in this studygrowth on substrates dependent on transcription ofgenes whose promoters contain CGATAR sites (Ravag- Strain Genotypenani et al. 1997).
MH1 biA1The expression of areA itself is controlled by autoregu-MH50 yA1 areA102 riboB2 pyroA4
lation and by regulation at the mRNA stability level, with MH341 yA1 riboB2 su-adE20 adE20areA217the areA transcript half-life being significantly reduced MH3018 yA1 pabaA1 argB2under nitrogen-sufficient conditions through an ele- MH8375 yA1 riboB2 fmdS1 alX
MH9467 yA1 areA102 pyroA4 usgSD(pJAF4473)ment in the 39 untranslated region (UTR) of the tran-MH9469 yA1 areA102 pyroA4 usgSD(pJAF4472)script (Langdon et al. 1995; Platt et al. 1996a). The
activity of AreA is also affected by the NmrA protein Gene symbols are as described in Clutterbuck (1974).(Andrianopoulos et al. 1998). In Neurospora crassa, theNmrA homologue NMR1 has been shown to be requiredfor nitrogen metabolite repression through interaction ration and DNA transformation was performed using the
method of Andrianopoulos and Hynes (1988). Arabidopsiswith the zinc finger and 12 carboxyl-terminal residuesthaliana media was prepared as described in Cobbett et al.of the AreA homologue NIT2 (Dunn-Coleman et al.(1998) with nitrogen sources added at 5 mm. Growth tests used1981; Xiao et al. 1995). Mutations affecting nmr-1 or the Columbia ecotype. Schizosaccharomyces pombe Edinburgh
altering the interacting residues of NIT2 result in nitro- minimal media was prepared as described in Alfa et al. (1993)gen-derepressed phenotypes. The relevant regions of with nitrogen sources added at 100 mm.
Molecular techniques: Standard methods were as describedboth AreA and NmrA are highly conserved in A. nidu-by Sambrook et al. (1989). Restriction enzymes and Munglans, suggesting both organisms share this regulatoryBean Exonuclease (Promega, Madison, WI) were used withmechanism (Platt et al. 1996b; Andrianopoulos et al. the supplied buffers. DNA fragments were purified from aga-
1998). Deletion of the C-terminal amino acids of AreA rose gels using the BresaClean DNA purification kit (Genethus partially relieves nitrogen metabolite repression Works) following the manufacturer’s specifications. DNA de-
phosphorylation was performed by addition of 1 unit of Arcticwhile almost complete relief of repression is obtainedshrimp alkaline phosphatase (U.S. Biochemicals, Cleveland)when the stability-mediating element in the 39 UTR ofto 50 ng DNA in the recommended buffer.the areA transcript is also mutated (Platt et al. 1996a). PCR protocols: Sequences of primers referred to in the text
In addition, the product of the tamA gene is believed are as follows:to act as a coactivator of AreA function and to contribute
BTUB2: AGT TGT TAC CAG CAC CGG ACto the activation of a subset of nitrogen metabolite re-BTUB3: GCT CCG GTG TTT ACA ATG G
pression regulated genes and has also been shown to FMD1: GCC GAG CCT GGA GAT GTCinteract with the carboxyl terminus of AreA (Davis et FMD2: TCG CCG ATT CCA CTC AGC
FMDlac1: CCA AAC TCG AGG ACC CCC CAC CCA GCal. 1996; Small et al. 1999). The formamidase of A.FMDlac2: GCG GAG CTC GAG GGT TTT GCnidulans was previously shown to be strongly regulatedFMDlac3: GCA GCG CTC GAG CAA CCCby AreA-dependent nitrogen metabolite repressionFMDlac4: GCG ACC TCG AGG TAA CCG
(Hynes 1972, 1975b). No evidence for induction by FMDlac5: GCA ATG CTC GAG ACG GAA GGexogenous formamide was found and furthermore the fmdsGSP1: CAG ATA GCT TTT GCA GG
fmdsGSP4: TGC CGT AGC TAG GGA TAT CTresponse to nitrogen limitation was greatly reducedfmdSrt1: GGC ATC CAG ATA TCC CTA GCwhen carbon was also limiting (Hynes 1970, 1972).USG1: TGA AGG TAC CCT GAG AGT GAG CCUsing published sequences for formamidases fromUSG2: TTA TCA GAT CTC GCA CGG CTA CG
Methylophilus methylotrophus and Mycobacterium smegmatis USGrt1: GTG GTT GGC ACC GTG CGA GCwe have identified an A. nidulans expressed sequence tag USGrt2: GGG AGT CCA TGA ATA TCT CGT C(EST) sequence with high similarity. This has enabled us
59 rapid amplification of cDNA ends (RACE) was performedto characterize the structure and regulation of the fmdS using the 59 RACE System v2 kit (GIBCO BRL, Gaithersburg,gene of A. nidulans. MD) with the nested gene specific primers fmdsGSP1 and
fmdsGSP4, using total RNA from A. nidulans strain MH1 col-lected after transfer to nitrogen starvation, carbon-sufficientconditions. Semiquantitative RT-PCR was performed as de-MATERIALS AND METHODSscribed in Ha et al. (1999) using the Superscript One-step RT-PCR system (GIBCO BRL). A total of 22 PCR cycles were usedA. nidulans strains, growth media, and transformation:
Strains used in this study are shown in Table 1. Growth media for the benA control (BTUB2 and BTUB3), 26 for usgS (USGrt1and USGrt2), and 28 or 30 for fmdS (fmdSrt1 and FMD2)and conditions were as described by Cove (1966). Nitrogen
sources were used at a final concentration of 10 mm and with all reactions stopped while in their linear phase.Plasmid construction: The 7-kb BamHI and 3.3-kb ClaI fmdS-carbon sources at 1% w/v unless stated otherwise. Mycelia for
assays were grown at 378 for 16 hr, washed with carbon-free hybridizing fragments from cosmid L19H02 were cloned intopBluescript SK1 to create pJAF4136 and pJAF4510, respec-media, and transferred to the assayed growth condition for 4
hr. Genetic analysis was carried out using techniques as de- tively. A minimal fmdS subclone was created by subcloning the1.7-kb ScaI fragment from pJAF4136 into pBluescript SK1 cutscribed by Clutterbuck (1974). A. nidulans protoplast prepa-

121Regulation of the fmdS Gene of A. nidulans
with EcoRV to create pJAF4155. usgS inactivation plasmids were used to probe colony lifts of a chromosome III specificcreated by inserting a 2.4-kb BglII/SmaI riboB fragment from cosmid library (Brody et al. 1991). Two hybridizingpPL1 (Oakley et al. 1987b) into pJAF4510 digested completely
cosmids were identified, L19H02 and L20G07. Both cos-with BglII and partially with HincII. The HincII site at 21221mids were shown to complement the A. nidulans fmdS1was used to create pJAF4472 and the HincII site at 21023 to
create pJAF4473. Nucleotide positions are with reference to mutant (MH8375) in transformants directly selected onthe fmdS ATG at 11. usgS cDNA clones were isolated from a formamide as the sole nitrogen source. Neither of theselZAP library constructed by Dr. Rodolfo Aramayo at Texas cosmids lay within the region of the ordered minimalA&M University, which used RNA from a mixed vegetative
library indicated to contain the fmdS region of the chro-and 24-hr asexual development culture of A. nidulans strainmosome (Prade et al. 1997), nor were these cosmidsFGSC A26. Plaque lifts were probed with the 3.3-kb ClaI frag-
ment from pJAF4510. Clones were excised as described in adjacent to each other.Short et al. (1988). A 7-kb BamHI fragment capable of complementing
All fmdS::lacZ constructs were created through fusion of the fmdS1 mutation was subcloned from cosmidfmdS at the BglII site at 1472 in frame to the BamHI site in
L19H02. Sequencing 3.5 kb of the end of the clone tothe lacZ reporter construct pJS3524. pJAF4241 was constructedwhich the PCR fragment hybridized in Southern blotsusing the XhoI site at 2599 bp, inserting into pJS3524 cut with
XhoI and BamHI. pJAF4504 was created by inserting the 2.5- revealed that the fmdS gene contained four introns thatkb XhoI fragment, including the rest of usgS, into the XhoI were confirmed by cDNA sequence. Complementationsite of pJAF4241. pJAF4242 and pJAF4243 were created by of the fmdS1 mutation with the minimal 1.7-kb ScaIdigesting pJS3524 with KpnI, blunting with mung bean endo-
subclone (pJAF4155) confirmed the function of thisnuclease, cutting with BglII, and inserting fmdS fragments start-sequence.ing at the EcoRV site at 2123 and the ScaI site at 262, respec-
tively. The remaining fmdS fusion constructs were created by Two major startpoints of fmdS transcription withinintroducing a KpnI site via PCR within the promoter that was the fmdS promoter region were identified in multipleused (in conjunction with the BglII site) to clone the se- independent clones generated by 59 RACE. Thequenced PCR products into pJS3524. All PCR reactions used
startpoints of transcription (at 221 and 212 relative toprimer FMD2 in conjunction with a construct-specific primer.the predicted ATG) corresponded to the existence ofConstructs included pJAF4503 (primer FMDlac1), pJAF4502
(FMDlac2), pJAF4501 (FMDlac3), pJAF4500 (FMDlac4), and two possible TATA boxes (both with the sequence TTA-pJAF4499 (FMDlac5). A similar approach was taken to create TAC) lying at 259 and 248. An additional startpointthe usgS::lacZ construct pJAF4622, using primer USG1 to intro- was detected within the first exon at nucleotide 17,duce a KpnI site at 21330 and USG2 to create a BglII site
which did not correspond to any recognized TATA se-at 1144 relative to the usgS start codon, with this fragmentquence.sequenced and subcloned into pJS3524.
The usgS overexpression plasmids were made using a three- fmdS belongs to a highly conserved gene family: Thestep cloning procedure. The usgS overexpression plasmid predicted 45-kD 411-amino-acid FmdS does not sharepJAF4869 was generated by inserting a 400-bp EcoRI/BamHI significant similarity with AmdS (Corrick et al. 1987)alcA promoter fragment from pAL3 (Fillinger et al. 1995)
but has 56% identity and 77% similarity to the formami-into pJAF4510 digested with EcoRI/BglII. A 1.4-kb EcoRI/BamHIdase of M. methylotrophus, with conservation across thepyrG-containing fragment from pARB4342 was then inserted
into EcoRI/BamHI-digested pJAF4869 to give pJAF4870. The entire protein. Similarity to the M. smegmatis enzyme isgene replacement plasmid pJAF4871 was created by inserting lower (66%), with no homology at the carboxyl terminusthe 3.8-kb BamHI/ClaI fragment from pJAF4870 into the usgS (Figure 1). Database searches using the predicted FmdSsubclone pJAF4510 digested fully with BglII and partially with
protein sequence identified previously uncharacterizedClaI.highly conserved formamidase-like sequences in the ge-lacZ reporter gene assays: Reporter gene studies used strains
carrying a single copy of the relevant construct in single copy nomes of prokaryotic, eukaryotic, and archaeal speciesat the argB locus as described by Punt et al. (1990). b-Galactosi- (Figure 1). Full sequences have been isolated as part ofdase assays were carried out by the method of Davis et al. the genome sequencing projects of Bordetella pertussis, B.(1988).
bronchiseptica, Aeropyrum pernix, S. pombe, Candida albicansNucleotide sequence accession number: The sequence forand two (in tandem) from A. thaliana. Tests on definedthe fmdS and usgS genes has been deposited in GenBank under
accession no. AF274009. media showed that S. pombe could use formamide asa sole nitrogen source and that formamide enhancedgrowth of A. thaliana relative to growth in the absence
RESULTSof added nitrogen source, indicating that these for-mamidase sequences are likely to be functional. Incom-Cloning of fmdS: A BLAST search performed on the
University of Oklahoma A. nidulans EST database plete sequences were also identified as ESTs from theplant species Oryza sativa (accession nos. C72046,(http://www.genome.ou.edu/asper.html) revealed a se-
quence (EST c8h05a1.r1) with a high level of similarity D49000, D49028, and D46854), Zea mays (AI714639 andAI739746), Medicago truncatula (AW256497), Lotus ja-to the formamidases of the bacteria M. methylotrophus
and M. smegmatis (Mahenthiralingam et al. 1993; ponicus (AW720090, AW720252, and AW719264), Sor-ghum bicolor (AW564921), and Lycopersicon esculentumWyborn et al. 1996). Primers FMD1 and FMD2 based
upon the EST sequence were used to amplify a 317-bp (AI782257). All of these proteins are distinct from thetwo major amidase families (the nitrilase and amidaseproduct from A. nidulans genomic DNA, which was then

122 J. A. Fraser, M. A. Davis and M. J. Hynes
signature group) and more closely resemble ureases elements responsible for this response, an fmdS::lacZpromoter deletion series was constructed and integrated(Novo et al. 1995).in single copy at argB as described by Punt et al. (1990).Regulation of fmdS by nitrogen metabolite repression:Each strain was assayed under growth conditions of ni-Previous studies of the regulation of formamidase activ-trogen and carbon starvation or sufficiency (Figure 2).ity showed a lack of induction by formamide yet strongThe longest fmdS::lacZ fusion (pJAF4241, with 599 bpregulation by nitrogen metabolite repression (Hynesof fmdS promoter) displayed a 10-fold increase in expres-1970, 1972). In an attempt to localize the promotersion during nitrogen starvation. Four potential AreArecognition sequences (HGATAR) were identified withinthe fmdS promoter, and the deletion series was used todetermine the relative contribution of each site. Thesites at 270 (TGATAG), 2145 (CGATAA) and 2160(AGATAA) relative to the predicted fmdS start codonall contribute to fmdS::lacZ expression under nitrogen-limiting conditions, with the site at 2145 only playinga minor role. The site at 2315 (TGATAA) appeared tohave little effect on the regulation of fmdS, with thedeletion of this region actually corresponding to a slightincrease in the basal levels of b-galactosidase activity,perhaps through the removal of other regulatory se-quences (Figure 2).
The areA217 loss-of-function allele abolished the re-sponse to nitrogen starvation for all constructs showingthat the response was AreA dependent. Expression offmdS::lacZ was unaffected by the tamAD mutation (resultsnot shown) in agreement with previous studies (Arstand Sheerins 1996).
Effects of carbon starvation on fmdS expression: Theabsence of a carbon source led to loss of response tonitrogen starvation of fmdS::lacZ expression (Figure 2).Earlier studies on formamidase levels showed a reducedresponse to nitrogen starvation in the absence of anadded carbon source (Hynes 1972). This was observedwith all fusion constructs, indicating that the effect wasnot due to a specific site in the fmdS promoter. ReducedfmdS::lacZ expression was observed only upon completecarbon starvation, with the poorer carbon sources, lac-tose, fructose, or limiting glucose (0.1% w/v), givingexpression equivalent to 1% glucose (results not shown).In A. nidulans carbon catabolite repression (CCR) ismediated by the zinc-finger repressor protein CreA(Dowzer and Kelly 1991). Analysis of the fmdS pro-moter identified possible tandem CreA recognition sites
Figure 1.—Alignment of 10 predicted formamidase-typeenzymes. Identical residues are indicated by dark shading;similar residues are indicated by light shading. The alignmentwas generated using Pileup (GCG software package, Dever-eaux et al. 1984) and viewed with Boxshade (Bioinformaticsgroup, ISREC). At1, A. thaliana formamidase 1 (accession no.CAB38294); At2, A. thaliana formamidase 2 (CAB38295); An,A. nidulans ; Ap, A. pernix (BAA79495); Bb, B. bronchiseptica ; Bp,B. pertussis ; Ca, C. albicans ; Mm, M. methylotrophus (Q50228); Ms,M. smegmatis (Q07238); Sp, S. pombe (CAB60014). Preliminarysequence data for C. albicans, B. pertussis, and B. bronchisepticawas obtained from The Institute for Genomic Research websiteat http://www.tigr.org.

123Regulation of the fmdS Gene of A. nidulans
Figure 2.—Deletion analysisof fmdS::lacZ regulation. Re-porter gene constructs withvarying fmdS promoter lengthswere targeted in single copyat the argB locus (Punt etal. 1990). Constructs were de-signed to sequentially removepotential regulatory elements(represented by the indicatedsymbols) with the expressionof each construct assayed inareA1 and areA217 genetic back-grounds under different growthconditions. Mycelium was grownin 100 ml of 1% glucose and10 mm ammonium tartrate me-dium at 378 for 16 hr, washedwith carbon-free/nitrogen-freemedia, and then transferred tothe indicated growth media for4 hr. When present after trans-
fer, glucose was at 1% w/v. NH4 indicates 10 mm ammonium tartrate; 2N is nitrogen free. Values are given in units per minuteper milligram of protein and represent the results of at least three separate experiments with standard errors shown.
at 2409 and 2399 (CCCCACCCAGCCCCGCGGA) glucose), nitrogen-free media, a short phase of no de-tectable change in expression was followed by b-galac-(Cubero and Scazzocchio 1994; Panozzo et al. 1998).
Equivalent sites with almost identical flanking sequence tosidase levels increasing over a 3-hr period. Transferof mycelia to carbon-free media at 2.5 hr led to a rapid(CTCCAGCCCAACTCCGCGGA) are also seen in the
niaD promoter (Johnstone et al. 1990). Hynes (1973) loss of the response, comparable to that caused by theprotein synthesis inhibitor cycloheximide. This effectshowed that nitrate reductase levels are also low under
carbon starvation conditions. Introduction of thecreA204 loss-of-function allele led to a reduction offmdS::lacZ expression although the response to AreAwas still apparent (Table 2). Removal of the tandemCreA sites had no detectable effect on expression of thereporter construct (Figure 2).
A time course analysis of the effect of carbon starva-tion on fmdS::lacZ expression was performed (Figure 3).Following transfer of mycelia to carbon-sufficient (1%
TABLE 2
Effect of various regulatory mutations on pJAF4241 fmds::lacZexpression: b-Galactosidase activity
Glucose Carbon freeMutantgenotype NH4 2N NH4 2N
WT 9 6 1 111 6 7 12 6 0 14 6 1areA217 9 6 1 10 6 1 9 6 0 16 6 2
Figure 3.—Time-course analysis of fmdS::lacZ expression. AcreA204 2 6 0 31 6 4 6 6 0 13 6 1strain carrying fmdS::lacZ fusion plasmid pJAF4241 was assayedareA217 3 6 0 5 6 1 4 6 1 10 6 2using conditions as described in Figure 2, with mycelia col-creA204lected at half-hour intervals and assayed for b-galactosidaseareA102 13 6 1 113 6 11 14 6 1 16 6 2activity. Mycelia were transferred to 1% glucose/nitrogen-freehapCD 2 6 0 75 6 7 7 6 1 6 6 1growth media (e), carbon-free/nitrogen-free media (h), andcarbon-free/nitrogen-free plus cycloheximide (n). MyceliaA strain containing the fmdS::lacZ fusion pJAF4241 with 599
bp of fmdS promoter integrated at the argB locus was crossed were washed with carbon-free/nitrogen-free media prior totransfer. Values are given in units per minute per milligraminto the relevant mutant genetic background. Assay conditions
were as described in Figure 2. Wild-type (WT) and areA217 of protein and represent the results of at least three separateexperiments with standard errors shown.data are reproduced from Figure 2.

124 J. A. Fraser, M. A. Davis and M. J. Hynes
was also observable irrespective of the time of transfer(data not shown). The nonmetabolizable glucose ana-logue 2-deoxyglucose (2-DOG) is sufficient at 0.5% w/vto cause CCR of amdS expression mediated by CreA(M. J. Hynes, unpublished results). Transfer of myceliato carbon-free/nitrogen-free media containing 0.5%2-DOG for 4 hr did not simulate carbon sufficiency andallow fmdS::lacZ expression (data not shown). The lackof effect of the creA204 mutation on the response tonitrogen starvation combined with the inability of2-DOG to substitute for glucose implies that the loss ofAreA control of fmdS expression is in response to asignal independent of carbon catabolite repression.
Regulation of fmdS by AnCF: The 59 region of fmdShas a CCAAT sequence (2112) that was shown to bindthe A. nidulans CCAAT-binding factor (AnCF; Papagi-annopoulos et al. 1996; Steidl et al. 1999) by electro-mobility shift assay (data not shown). The same sitehas been shown to be required in an AnCF-dependentmanner for the formation of a nucleosome-free regionin the fmdS promoter (Narendja et al. 1999). A straindeleted for the hapC gene encoding one of the subunitsof AnCF (Papagiannopoulos et al. 1996) displayed re-duced growth on formamide as the sole nitrogen source.A hapCD background resulted in slightly reduced ex-pression of the fmdS::lacZ fusion (Table 2). However,deletion of the CCAAT sequence had no noticeableeffect on fmdS::lacZ expression (compare pJAF4242 withpJAF4499 in Figure 2). This could have been due tothe deletion fortuitously bringing other regulatory se-quences into the proximity of the fmdS promoter ormore probably this is due to loss of usgS in this construct(see discussion).
The 59 end of fmdS contains an overlapping gene:Previous studies of formamidase activity indicated a re-duction of fmdS expression in areA102 strains, corre-
Figure 4.—Identification of usgS, an upstream gene, withinthe promoter of fmdS. (A) Schematic representation and re-striction map of the fmdS region. Exons are shown as shadedboxes (usgS dark, fmdS light). The presence of a divergent ESTis indicated. Directions of transcription (arrows) are shown. B,BamHI; Bg, BglII; C, ClaI; H, HincII; S, ScaI; and X, XhoI. (B)usgS deletion constructs. Constructs allowed disruption of usgSthrough deletion of the first two exons (pJAF4472) and firstthree exons (pJAF4473) of usgS while still maintaining anuninterrupted fmdS promoter region. (C) Nucleotide and con-ceptual protein sequence of the area surrounding the usgS/fmdS intergenic region, showing UsgS residues 238–362 andFmdS residues 1–65. Putative fmdS TATA boxes are underlinedand AreA recognition sites boxed. A CCAAT site shown tobe required for the formation of a nucleosome-free region(Narendja et al. 1999) is doubly underlined. usgS transcriptpolyadenylation sites are indicated with down arrows and fmdStranscriptional startpoints as determined by RACE with solidcircles. Nucleotides are numbered with reference to the 11 atthe start of the fmdS coding region. Lowercase letters indicateintron sequences.

125Regulation of the fmdS Gene of A. nidulans
sponding with a reduction in growth on formamide asa sole nitrogen source (Hynes 1972). However, expres-sion of the fmdS::lacZ construct was equivalent to wildtype in an areA102 mutant background (Table 2). Toinvestigate this discrepancy further analysis of the fmdSpromoter was undertaken. Sequence analysis andsearches of the A. nidulans EST database revealed theexistence of a gene within the promoter of fmdS tran-scribed in the same direction as the fmdS transcript. Tosequence the entire gene an additional 1.3 kb of fmdSupstream sequence was obtained through cloning of a3.3-kb ClaI fragment (pJAF4510) from cosmid L19H02.Isolation of three independent cDNA clones confirmedthat the upstream gene (usgS) contained four intronsand encoded a predicted 362-amino acid highly hy-drophobic 41-kD product (Figure 4A). The usgS/fmdSintergenic region (from the stop codon of usgS to thestart codon of fmdS) is only 242 bp and the usgS tran-
Figure 5.—Effects of deletion of the usgS gene on for-script overlaps that of fmdS by up to 150 bp, terminatingmamide utilization. Colonies were grown for 2 days at 378 onwithin either the first exon or first intron of fmdS (Figure 1% glucose minimal medium. Nitrogen sources and concen-
4C). Database searches with usgS revealed a single homo- trations are indicated. WT is wild-type strain MH97. The usgSDlogue of unidentified function as an EST from the fun- strain is MH9510, generated with the usgS knockout plasmid
pJAF4473. areA102 usgSD [usgS] designates strain MH9467gus Botrytus cinerea (accession nos. CNS01D74 and(areA102 usgSD) transformed with multiple ectopic copies ofCNS019CV). Application of a hidden Markov topologyusgS subclone pJAF4510 to give strain MH9487.prediction algorithm (Tusnady and Simon 1998) indi-
cated that usgS is likely to encode a six-transmembranehelix protein with a long extracellular C-terminal tail.
grew slightly better than wild type (MH1). ThereforeCharacterization of a usgS deletion strain: Two disrup-loss of usgS results in increased formamide utilizationtion constructs of usgS were created by replacing eitherin both areA102 and areA1 backgrounds.the first two (pJAF4472) or first three exons (pJAF4473)
The plasmid pJAF4510 bearing usgS was cotrans-of usgS in plasmid pJAF4510 with a riboB1 fragment fromformed with the pyroA1 plasmid pI4 (May et al. 1989)plasmid pPL1 (Oakley et al. 1987b; Figure 4B). Theinto the areA102 usgSD strain. All cotransformants wereriboB1-containing fragment was inserted in reverse ori-phenotypically identical to the usgSD strains. Southernentation to ensure that no novel usgS transcripts wereblot analysis indicated that the usgS1 plasmid had inte-generated. For each construct a linear ClaI fragmentgrated ectopically into the genome and the genomiccontaining the usgS::riboB insert was used to transformusgS was still disrupted, showing that the effects of thean areA102; pyroA4; riboB2 strain (MH50), selecting forusgSD could not be restored in trans (MH9487, Figureriboflavin prototrophy. No obvious morphological phe-5). To confirm the lack of a trans-acting regulatory rolenotype could be seen in any RiboB1 transformants.for the usgS product on fmdS expression, the usgSD alleleSouthern blot analysis of 30 transformants for each con-was introduced into an fmdS::lacZ background and nostruct revealed one disruptant for each containing thechange in fmdS::lacZ expression was observed (resultsdesired integration event at usgS (data not shown).not shown). These results are compatible with usgS hav-Plate tests were used to determine if loss of usgSing a cis-acting effect on fmdS expression.function gave rise to an altered growth phenotype on
Analysis of transcription of fmdS and usgS by semi-different carbon or nitrogen sources. Apart from for-quantitaive RT-PCR showed that the fmdS product in-mamide utilization, no other phenotypes were detected.creased with nitrogen starvation and that levels wereGrowth on either 10 mm or 50 mm formamide waslower with carbon starvation (in agreement with thenoticeably stronger, particularly when comparing thedata presented in Figure 2). The usgS transcript wasusgSD areA102 double mutant (MH4473) to the parentalpresent in high levels on ammonium and showed onlyareA102 strain (MH50), which showed poor growth anda slight response to nitrogen starvation and no changereduced mycelial density on formamide as the sole nitro-in response to carbon starvation (Figure 6). The loss-gen source (Figure 5). Phenotypes of disruptants gener-of-function areA217 allele resulted in levels of fmdS prod-ated with either of the deletion constructs were identi-uct not responding to nitrogen starvation. In agreementcal, indicating that the new phenotype was not due towith usgS having a cis-acting effect, the fmdS productthe particular positioning of the riboB1 fragment. Thelevel was increased under both nitrogen limitation andusgSD mutation was introduced by genetic crosses into
an areA1 background. usgSD areA1 (MH9510) colonies sufficiency in the usgSD mutant. This response was most

126 J. A. Fraser, M. A. Davis and M. J. Hynes
weakly regulated by AreA. No major effects of carbonstarvation were evident.
Inclusion of the upstream gene in the usgS-fmdS::lacZconstruct (pJAF4504) led to a reduction in b-galactosi-dase levels under all growth conditions assayed com-pared to those observed for the construct lacking theentire usgS gene (pJAF4241, Figure 7). Under nitrogen-sufficient conditions expression of fmdS::lacZ was easilyobserved in the absence of usgS, whereas inclusion ofthe entire usgS gene reduced fmdS::lacZ expression toalmost undetectable levels. The levels of usgS-fmdS::lacZ
Figure 6.—Semiquantitative RT-PCR analysis of fmdS and considered in conjunction with the phenotype of theusgS transcription. All reactions were stopped while in their usgSD strain on formamide suggested that transcriptionlinear phase as described in Ha et al. (1999). Twenty-two PCR of the upstream gene was interfering with expressioncycles were used for the benA control (using the primers
of fmdS. Despite little change in usgS::lacZ expression,BTUB2 and BTUB3), 26 cycles for usgS (USGrt1 and USGrt2),expression of usgS-fmdS::lacZ was highly dependent onand 28 cycles for fmdS (fmdSrt1 and FMD2) except for the
areA217 mutant where 30 cycles were used to enhance the an AreA-mediated response to nitrogen starvation (Fig-signal. Wild-type RNA was isolated from strain MH1, areA217 ure 7). Inclusion of usgS therefore changed fmdS fromRNA from strain MH341, and usgSD RNA from MH9510. Tran- having leaky expression to being tightly regulated byscription of the constitutively expressed b-tubulin gene benA
nitrogen metabolite repression.(May et al. 1987) was used as a loading control. Growth condi-Alteration of usgS regulation affects fmdS expression:tions were as described in Figure 2.
The areA102 mutation was previously found to result inz50% of wild-type formamidase activity (Hynes 1972,
noticeable in repressing (glucose and ammonium 1975b). This was not found for the assays of fmdS::lacZgrown) conditions. fusions lacking usgS (Table 2). The areA102 mutant
As none of the fmdS fusion constructs studied thus far background resulted in increased usgS::lacZ expressionhad included the upstream gene, additional constructs under conditions of nitrogen limitation (Figure 7), con-were generated: a usgS::lacZ fusion (pJAF4622) and a sistent with the usgS promoter containing three poten-usgS-fmdS::lacZ fusion (pJAF4504) that contained 3.1 kb tial TGATAR AreA recognition sites (at 2305, 2676,of fmdS promoter, incorporating the upstream gene and and 2821 relative to the usgS predicted start codon).its promoter (Figure 7). The expression of usgS::lacZ When usgS was present in the fmdS promoter (as in(pJAF4622) was not strongly regulated in agreement pJAF4504) the presence of the areA102 allele resultedwith the RT-PCR data (Figure 7). b-Galactosidase levels in an z50% reduction in fmdS expression (Figure 7),increased less than twofold under nitrogen starvation consistent with the effects of areA102 on formamideconditions, and this increase was eliminated by the utilization being due to increased usgS transcription that
interferes with fmdS expression.areA217 mutation, indicating that usgS expression is
Figure 7.—Effects of thepresence of usgS on fmdS::lacZexpression. b-Galactosidase as-says of the three lacZ constructsunder different growth condi-tions in the presence of differ-ent areA alleles were performedas described in Figure 2. Con-struct nucleotide coordinatesare given relative to the ATGof fmdS. Wild-type data are re-produced from Figure 2 andTable 2. The lacZ fusions werecrossed into the relevant mu-tant genetic background. Val-ues are given in units per mi-nute per milligram of proteinand represent the results of atleast three separate experimentswith standard errors shown.
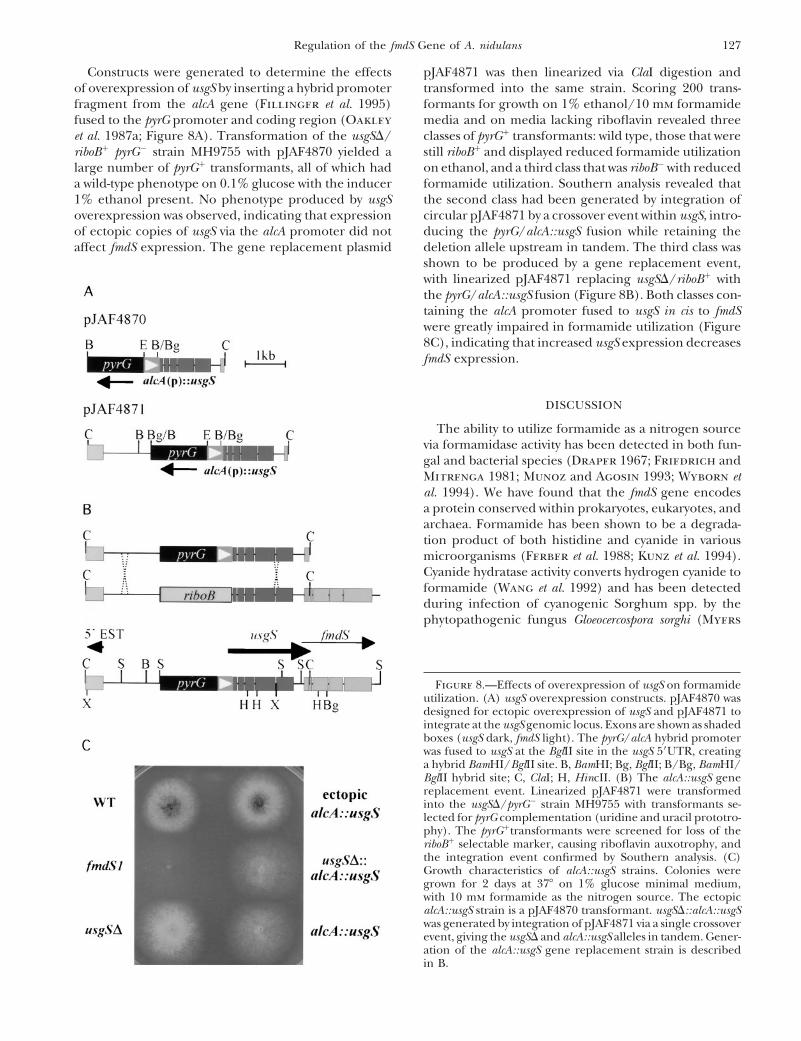
127Regulation of the fmdS Gene of A. nidulans
Constructs were generated to determine the effects pJAF4871 was then linearized via ClaI digestion andtransformed into the same strain. Scoring 200 trans-of overexpression of usgS by inserting a hybrid promoter
fragment from the alcA gene (Fillinger et al. 1995) formants for growth on 1% ethanol/10 mm formamidemedia and on media lacking riboflavin revealed threefused to the pyrG promoter and coding region (Oakley
et al. 1987a; Figure 8A). Transformation of the usgSD/ classes of pyrG1 transformants: wild type, those that werestill riboB1 and displayed reduced formamide utilizationriboB1 pyrG2 strain MH9755 with pJAF4870 yielded a
large number of pyrG1 transformants, all of which had on ethanol, and a third class that was riboB2 with reducedformamide utilization. Southern analysis revealed thata wild-type phenotype on 0.1% glucose with the inducer
1% ethanol present. No phenotype produced by usgS the second class had been generated by integration ofcircular pJAF4871 by a crossover event within usgS, intro-overexpression was observed, indicating that expression
of ectopic copies of usgS via the alcA promoter did not ducing the pyrG/alcA::usgS fusion while retaining thedeletion allele upstream in tandem. The third class wasaffect fmdS expression. The gene replacement plasmidshown to be produced by a gene replacement event,with linearized pJAF4871 replacing usgSD/riboB1 withthe pyrG/alcA::usgS fusion (Figure 8B). Both classes con-taining the alcA promoter fused to usgS in cis to fmdSwere greatly impaired in formamide utilization (Figure8C), indicating that increased usgS expression decreasesfmdS expression.
DISCUSSION
The ability to utilize formamide as a nitrogen sourcevia formamidase activity has been detected in both fun-gal and bacterial species (Draper 1967; Friedrich andMitrenga 1981; Munoz and Agosin 1993; Wyborn etal. 1994). We have found that the fmdS gene encodesa protein conserved within prokaryotes, eukaryotes, andarchaea. Formamide has been shown to be a degrada-tion product of both histidine and cyanide in variousmicroorganisms (Ferber et al. 1988; Kunz et al. 1994).Cyanide hydratase activity converts hydrogen cyanide toformamide (Wang et al. 1992) and has been detectedduring infection of cyanogenic Sorghum spp. by thephytopathogenic fungus Gloeocercospora sorghi (Myers
Figure 8.—Effects of overexpression of usgS on formamideutilization. (A) usgS overexpression constructs. pJAF4870 wasdesigned for ectopic overexpression of usgS and pJAF4871 tointegrate at the usgS genomic locus. Exons are shown as shadedboxes (usgS dark, fmdS light). The pyrG/alcA hybrid promoterwas fused to usgS at the BglII site in the usgS 59UTR, creatinga hybrid BamHI/BglII site. B, BamHI; Bg, BglII; B/Bg, BamHI/BglII hybrid site; C, ClaI; H, HincII. (B) The alcA::usgS genereplacement event. Linearized pJAF4871 were transformedinto the usgSD/pyrG2 strain MH9755 with transformants se-lected for pyrG complementation (uridine and uracil prototro-phy). The pyrG1transformants were screened for loss of theriboB1 selectable marker, causing riboflavin auxotrophy, andthe integration event confirmed by Southern analysis. (C)Growth characteristics of alcA::usgS strains. Colonies weregrown for 2 days at 378 on 1% glucose minimal medium,with 10 mm formamide as the nitrogen source. The ectopicalcA::usgS strain is a pJAF4870 transformant. usgSD::alcA::usgSwas generated by integration of pJAF4871 via a single crossoverevent, giving the usgSD and alcA::usgS alleles in tandem. Gener-ation of the alcA::usgS gene replacement strain is describedin B.

128 J. A. Fraser, M. A. Davis and M. J. Hynes
and Fry 1978a; Fry and Myers 1981; Wang et al. 1999). These results suggest that AreA is rapidly inactivatedin response to carbon starvation. Genes dependent onNo formamidase activity is detected in this fungus but
the formamide produced is not detectable as it is rapidly AreA, such as fmdS and other genes solely involved innitrogen source utilization, will not respond to nitrogenmetabolized by other organisms (Myers and Fry
1978b). Cyanide originating from cyanide-producing starvation in the absence of a carbon source. Genessuch as amdS, which are also involved in carbon sourcebacteria or cyanogenic glucosides in plants is likely to
be a source of formamide for utilization by saprophytes. utilization, will respond to carbon starvation by activa-tion by one or more other regulatory pathways. TheIn the phytopathogenic fungi Leptosphaeria maculans and
Fusarium solani, both cyanide and formamide can serve mechanism for signaling AreA inactivation due to car-bon starvation is yet to be determined. Due to its simplic-as a nitrogen source (Barclay et al. 1998; Sexton and
Howlett 2000). Our finding that formamidase homo- ity fmdS regulation provides an excellent system for thefurther study of this phenomenon. It is likely that inlogues occur in many plant species may indicate a detox-
ification role for these enzymes in plants. nature fungi may often face severe nutrient deprivationand derepress appropriate enzymes for the scavengingAs had been shown previously, fmdS expression was
strongly regulated in response to the nitrogen state of of trace sources of carbon and nitrogen.Analysis of the fmdS promoter revealed the presencethe cell (Hynes 1972, 1975b). Deletion analysis of the
fmdS promoter revealed multiple GATA sequences of an overlapping gene (usgS) whose transcript termi-nated within fmdS. The deletion of usgS resulted in in-through which this effect was modulated by AreA, as
found for other genes required for alternative nitrogen creased growth on formamide and cis/trans tests showedthat the effect of usgS was apparent only when in cissource utilization (Davis et al. 1993; Punt et al. 1995;
Gonzalez et al. 1997; Hutchings et al. 1999). Induction with fmdS. When considered in conjunction with thedata obtained when usgS expression was increased (inof fmdS expression by exogenous formamide does not
occur and it is unlikely that formamide is generated by an areA102 background and when overexpressed fromthe pyrG/alcA promoter), the results strongly suggestthe cell as an endogenous inducer (Hynes 1970, 1975a).
In support of this the promoter deletion series shows that expression of usgS results in transcriptional interfer-ence of the fmdS promoter. The fmdS::lacZ and RT-PCRdependence only on the GATA sites. This indepen-
dence from pathway-specific induction has been ob- analysis clearly indicate that the usgS effect is at thetranscriptional level.served for other nitrogen catabolic enzymes, e.g., histi-
dase (Polkinghorne and Hynes 1982) and the general As discussed by Eggermont and Proudfoot (1993)a genuine interference effect should be seen only in cisamidase (Hynes 1975b). It is likely that, upon nitrogen
starvation, the expression of many genes producing en- and not in trans, which is clearly shown in the usgS/fmdS system and indicates that the intergenic regionzymes responsible for nitrogen scavenging is activated,
irrespective of the presence of the relevant substrates. between usgS and fmdS lacks interference blocking ele-ments such as transcriptional pause or polyadenylationReduced formamidase expression in response to car-
bon starvation (Hynes 1972) was further confirmed by sites. In fact, the usgS poly(A) sites within fmdS showlittle similarity to the characterized diffuse fungal poly-this work. This has also been observed for nitrate reduc-
tase, NADP-GDH, and histidase (Hynes 1973, 1974; adenylation sites rich in A and U bases (for a review,see Proudfoot 1991). Multiple polyadenylation sitesPolkinghorne and Hynes 1982). Expression of the
fmdS::lacZ fusion follows the same trend as formamidase for the usgS transcript exist and by definition must beinefficient, because the fmdS transcript itself must passlevels and, together with the RT-PCR data, is consistent
with an effect at the transcriptional level. The time- through these sites. In contrast the fmdS gene appearsto contain an efficient polyadenylation signal, althoughcourse analysis of fmdS::lacZ expression suggests that
carbon starvation results in loss of activation by AreA. the definition of a fungal poly(A) site and the mecha-nisms involved are not as well understood as in mamma-This is supported by the loss of dependence on AreA
for expression under carbon starvation conditions and lian systems (Levitt et al. 1989; Proudfoot 1991).One model that can be proposed is that there is athe finding that the element responsible for this regula-
tion could not be localized to any specific sites in the direct effect of transcription through a downstream pro-moter on transcription initiation of the downstreamfmdS promoter deletion series. It is highly unlikely that
this is a general effect of carbon starvation on gene gene. Positive and negative supercoils generated by RNApolymerase could prevent access to the downstream pro-expression. Carbon starvation leads to increased levels
of acetamidase activity even in areA loss-of-function mu- moter by RNA polymerase (Wu et al. 1988). This phe-nomenon, which has been called promoter occlusiontants and relieves ammonium repression (Hynes 1972,
1975b). This has been supported by more recent data (Adhya and Gottesman 1982), has been characterizedin the compact genomes of phage and bacteria (e.g.,using an amdS::lacZ fusion that shows that the response
to carbon starvation is independent of AreA but depen- Adhya and Gottesman 1982). There are also rare eu-karyotic examples. In Saccharomyces cerevisiae it has beendent on an element in the 59 region of amdS (M. J.
Hynes, M. A. Davis and J. A. Sharp, unpublished data). shown that expression from the promoter of the actin

129Regulation of the fmdS Gene of A. nidulans
gene completely occludes a cryptic promoter in the first tion effects on fmdS expression as demonstrated here.Position effects are often seen with ectopic integrationintron (Irniger et al. 1992). The Drosophila melanogaster
Adh larval promoter is also occluded in this case by of genes and with directed gene rearrangements thatresult in alterations of gene regulation (e.g., Miller ettranscription from the upstream adult promoter (Cor-
bin and Maniatis 1989). In these situations transcrip- al. 1987). In general, in lower eukaryotes genes canbe relatively close together. In S. cerevisiae the averagetional interference is absolute with the downstream pro-
moter being completely inactive. distance betweeen tandemly oriented open readingframes is only 517 bp (Dujon 1996), indicating thatA second but not mutually exclusive model is that
expression of an upstream gene affects the chromatin transcription overlap and interference may occur insome cases. Gene clusters occur commonly in fungi,structure of the downstream regulatory region and pro-
moter such that access by transcription factors and/or an example of which is in A. nidulans sterigmatocystinbiosynthesis where the 25 genes involved are clusteredRNA polymerase is altered. Studies in S. cerevisiae have
generally indicated that transcript overlap of convergent within only 50 kb (Brown et al. 1996). The relativepositions of genes within these clusters may affect theirgenes has little effect on transcriptional levels (e.g.,
Peterson and Myers 1993), although some exceptions expression via transcriptional interference.have been observed (Puig et al. 1999). Recent studies J.A.F. was a recipient of an Australian Postgraduate Award. Thisof the convergent POT1 and YIL161w open reading work was supported by a grant from the Australian Research Council.frames (ORFs; with 84–115-bp transcript overlap)showed that deletion of the YIL161w promoter leads toa twofold increase in POT1 expression and alteration LITERATURE CITEDof the intergenic chromatin structure (Puig et al. 1999).
Adhya, S., and M. Gottesman, 1982 Promoter occlusion: transcrip-If this imposed alteration in nucleosome structure oc- tion through a promoter may inhibit its activity. Cell 29: 939–944.curred instead over the promoter of a tandemly arrayed Alfa, C., P. Fantes, J. Hyams, M. McLeod and E. Warbrick (Editors),
1993 Experiments with Fission Yeast: A Laboratory Course Manual.gene, the effects could be expected to be more signifi-Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.cant through the alteration of transcription factor ac- Andrianopoulos, A., and M. J. Hynes, 1988 Cloning and analysis
cessibilty. In the unr/N-ras system in Mus musculus, the of the positively acting regulatory gene amdR from Aspergillusnidulans. Mol. Cell. Biol. 8: 3532–3541.unr transcript, although shown to interfere with expres-
Andrianopoulos, A., S. Kourambas, J. A. Sharp, M. A. Davis andsion of the downstream N-ras promoter, actually termi- M. J. Hynes, 1998 Characterization of the Aspergillus nidulansnates 150 bp before the N-ras transcription initiation nmrA gene involved in nitrogen metabolite repression. J. Bacte-
riol. 180: 1973–1977.sites (Boussadia et al. 1997). The effect observed inArst, H. N., Jr., and D. J. Cove, 1973 Nitrogen metabolite repressionthis instance is proposed to occur through interference in Aspergillus nidulans. Mol. Gen. Genet. 126: 111–141.
with upstream promoter elements, and this could be Arst, H. N., Jr., and A. Sheerins, 1996 Nitrogen metabolite repres-sion in Aspergillus nidulans: A farewell to tamA? Curr. Genet. 6:via the enforcement of an altered chromatin structure.245–257.The fmdS promoter has been shown to have an or- Barclay, M., V. A. Tett and C. J. Knowles, 1998 Metabolism and
dered nucleosome structure centered around the enzymology of cyanide/metallocyanide biodegradation by Fu-sarium solani under neutral and acidic conditions. Enzyme Micro-CCAAT sequence, which may be required for overcom-biol. Technol. 23: 321–330.ing such an effect (Narendja et al. 1999). Reporter gene Boussadia, O., F. Amiot, S. Cases, G. Triqueneaux, H. Jacquemin-
assays in this study revealed that the CCAAT-binding Sablon et al., 1997 Transcription of unr (upstream of N-ras)down-modulates N-ras expression in vivo. FEBS Lett. 420: 20–24.complex AnCF has a relatively minor role when usgS is
Brody, H., J. Griffith, A. J. Cuticchia, J. Arnold and W. E. Tim-absent. In a genomic context the role of AnCF may be berlake, 1991 Chromosome-specific recombinant DNA librar-much greater and be required, in conjunction with ies from the fungus Aspergillus nidulans. Nucleic Acids Res. 19:
3105–3109.AreA, to open up the fmdS promoter to overcome theBrown, D. W., J. H. Yu, H. S. Kelkar, M. Fernandes, T. C. Nesbittnegative effects of the upstream gene. Expression of et al., 1996 Twenty-five coregulated transcripts define a sterig-
usgS altering nucleosome positioning would enforce a matocystin gene cluster in Aspergillus nidulans. Proc. Natl. Acad.Sci. USA 93: 1418–1422.greater dependence on both AnCF and AreA binding,
Clutterbuck, J. A., 1974 Aspergillus nidulans Genetics. Plenum Pub-resulting in tightly controlled fmdS expression. It has lishing Corp., New York.been recently shown that the effects of readthrough Cobbett, C. S., M. J. May, R. Howden and B. Rolls, 1998 The
glutathione-deficient, cadmium-sensitive mutant, cad2-1, of Arabi-transcripts from the S. cerevisiae GAL10 gene into thedopsis thaliana is deficient in gamma-glutamylcysteine synthetase.adjacent GAL7 promoter result from displacement ofPlant J. 16: 73–78.
the Gal4p transcriptional activator (Greger et al. 2000). Corbin, V., and T. Maniatis, 1989 Role of transcriptional interfer-ence in the Drosophila melanogaster Adh promoter switch. NatureThe effects of transcriptional interference result in337: 279–282.expression of the usgS and fmdS genes being functionally
Corrick, C. M., A. P. Twomey and M. J. Hynes, 1987 The nucleotidelinked. Alterations in expression of usgS result in altered sequence of the amdS gene of Aspergillus nidulans and the molecu-
lar characterization of 59 mutations. Gene 53: 63–71.fmdS expression. In an evolutionary sense selectionCove, D. J., 1966 The induction and repression of nitrate reductasemight maintain the tandem gene arrangement in order
in the fungus Aspergillus nidulans. Biochim. Biophys. Acta 113:to retain appropriate controlled expression of fmdS. Al- 51–56.
Cubero, B., and C. Scazzocchio, 1994 Two different, adjacent andteration of this gene arrangement would result in posi-

130 J. A. Fraser, M. A. Davis and M. J. Hynes
divergent zinc finger binding sites are necessary for CREA-medi- drogenase activities in Aspergillus nidulans. J. Gen. Microbiol. 81:165–170.ated carbon catabolite repression in the proline gene cluster of
Aspergillus nidulans. EMBO J. 13: 407–415. Hynes, M. J., 1975a Amide utilization in Aspergillus nidulans: evi-dence for a third amidase enzyme. J. Gen. Microbiol. 91: 99–109.Davis, M. A., C. S. Cobbett and M. J. Hynes, 1988 An amdS-lacZ
fusion for studying gene regulation in Aspergillus. Gene 63: 199– Hynes, M. J., 1975b Studies on the role of the areA gene in theregulation of nitrogen catabolism in Aspergillus nidulans. Aust. J.212.
Davis, M. A., J. M. Kelly and M. J. Hynes, 1993 Fungal catabolic Biol. Sci. 28: 301–313.Hynes, M. J., and M. A. Davis, 1996 Regulation of acetamide catabo-gene regulation: molecular genetic analysis of the amdS gene of
Aspergillus nidulans. Genetica 90: 133–145. lism, pp. 381–394 in The Mycota, edited by G. A. Marzluf andR. Brambl. Springer-Verlag, Berlin.Davis, M. A., A. J. Small, S. Kourambas and M. J. Hynes, 1996 The
tamA gene of Aspergillus nidulans contains a putative zinc cluster Hynes, M. J., and J. A. Pateman, 1970 The use of amides as nitrogensources by Aspergillus nidulans. J. Gen. Microbiol. 63: 317–324.motif which is not required for gene function. J. Bacteriol. 178:
3406–3409. Irniger, S., C. M. Egli, M. Kuenzler and G. H. Braus, 1992 Theyeast actin intron contains a cryptic promoter that can be switchedDevereaux, J., P. Haeberli and O. Smithies, 1984 A comprehen-
sive set of sequence analysis programs for the VAX. Nucleic Acids on by preventing transcriptional interference. Nucleic Acids Res.20: 4733–4739.Res. 12: 387–395.
Diallinas, G., L. Gorfinkiel, H. N. Arst, Jr., G. Cecchetto and Johnstone, I. L., P. C. McCabe, P. Greaves, S. J. Gurr, G. E. Coleet al., 1990 Isolation and characterisation of the crnA-niiA-niaDC. Scazzocchio, 1995 Genetic and molecular characterization
of a gene encoding a wide specificity purine permease of Aspergil- gene cluster for nitrate assimilation in Aspergillus nidulans. Gene90: 181–192.lus nidulans reveals a novel family of transporters conserved in
prokaryotes and eukaryotes. J. Biol. Chem. 270: 8610–8622. Kudla, B., M. X. Caddick, T. Langdon, N. M. Martinez-Rossi,C. F. Bennett et al., 1990 The regulatory gene areA mediatingDowzer, C. E., and J. M. Kelly, 1991 Analysis of the creA gene, a
regulator of carbon catabolite repression in Aspergillus nidulans. nitrogen metabolite repression in Aspergillus nidulans. Mutationsaffecting specificity of gene activation alter a loop residue of aMol. Cell. Biol. 11: 5701–5709.
Draper, P., 1967 The aliphatic acylamide amidohydrolase of Myco- putative zinc finger. EMBO J. 9: 1355–1364.Kunz, D. A., C. S. Wang and J. L. Chen, 1994 Alternative routes ofbacterium smegmatis: its inducible nature and relation to acyl-trans-
fer to hydroxylamine. J. Gen. Microbiol. 46: 111–123. enzymic cyanide metabolism in Pseudomonas fluorescens NCIMB11764. Microbiology 140: 1705–1712.Dujon, B., 1996 The yeast genome project: What did we learn?
Trends Genet. 12: 263–270. Langdon, T., A. Sheerins, A. Ravagnani, M. Gielkens, M. X. Cad-dick et al., 1995 Mutational analysis reveals dispensability of theDunn-Coleman, N. S., A. B. Tomsett and R. H. Garrett, 1981 The
regulation of nitrate assimilation in Neurospora crassa: biochemical N-terminal region of the Aspergillus transcription factor mediatingnitrogen metabolite repression. Mol. Microbiol. 17: 877–888. (er-analysis of the nmr-1 mutants. Mol. Gen. Genet. 182: 234–239.
Eggermont, J., and N. J. Proudfoot, 1993 Poly(A) signals and ratum: Mol. Microbiol. 20: 239).Levitt, N., D. Briggs, A. Gil and N. J. Proudfoot, 1989 Definitiontranscriptional pause sites combine to prevent interference be-
tween RNA polymerase II promoters. EMBO J. 12: 2539–2548. of an efficient synthetic poly(A) site. Genes Dev. 3: 1019–1025.Mahenthiralingam, E., P. Draper, E. O. Davis and M. J. Colston,Ferber, D. M., F. Khambaty and B. Ely, 1988 Utilization of histidine
by Caulobacter crescentus. J. Gen. Microbiol. 134: 2149–2154. 1993 Cloning and sequencing of the gene which encodes thehighly inducible acetamidase of Mycobacterium smegmatis. J. Gen.Fillinger, S., C. Panozzo, M. Mathieu and B. Felenbok, 1995 The
basal level of transcription of the alc genes in the ethanol regulon Microbiol. 139: 575–583.Marzluf, G. A., 1997 Genetic regulation of nitrogen metabolismin Aspergillus nidulans is controlled both by the specific transactiva-
tor AlcR and the general carbon catabolite repressor CreA. FEBS in the fungi. Microbiol. Mol. Biol. Rev. 61: 17–32.May, G. S., M. L. Tsang, H. Smith, S. Fidel and N. R. Morris, 1987Lett. 368: 547–550.
Friedrich, C. G., and G. Mitrenga, 1981 Utilization of aliphatic Aspergillus nidulans beta-tubulin genes are unusually divergent.Gene 55: 231–243.amides and formation of two different amidases by Alcaligenes
eutrophus. J. Gen. Microbiol. 125: 367–374. May, G. S., R. B. Waring, S. A. Osmani, N. R. Morris and S. H.Denison, 1989 Proceedings of the EMBO-Alko Workshop onFry, W. E., and D. F. Myers, 1981 Hydrogen cyanide metabolism
by fungal pathogens of cyanogenic plants, pp. 321–334 in Cyanide Molecular Biology of Filamentous Fungi. Foundation for Biotech-nical and Industrial Fermentation Research, Helsinki.in Biology, edited by B. Vennesland, E. E. Conn, C. J. Knowles,
J. Westley and F. Wissing. Academic Press, London. Miller, B. L., K. Y. Miller, K. A. Roberti and W. E. Timberlake,1987 Position-dependent and -independent mechanisms regu-Gonzalez, R., V. Gavrias, D. Gomez, C. Scazzocchio and B. Cub-
ero, 1997 The integration of nitrogen and carbon catabolite late cell-specific expression of the SpoC1 gene cluster of Aspergillusnidulans. Mol. Cell. Biol. 7: 427–434.repression in Aspergillus nidulans requires the GATA factor AreA
and an additional positive-acting element, ADA. EMBO J. 16: Munoz, G. A., and E. Agosin, 1993 Glutamine involvement in nitro-gen control of giberellic acid production in Gibberella fujikuroi.2937–2944.
Gorfinkiel, L., G. Diallinas and C. Scazzocchio, 1993 Sequence Appl. Environ. Microbiol. 59: 4317–4322.Myers, D. F., and W. E. Fry, 1978a The development of Gloecercosporaand regulation of the uapA gene encoding a uric acid-xanthine
permease in the fungus Aspergillus nidulans. J. Biol. Chem. 268: sorghi in Sorghum. Phytopathology 68: 1147–1155.Myers, D. F., and W. E. Fry, 1978b Enzymatic release and metabo-23376–23381.
Greger, I. H., A. Aranda and N. Proudfoot, 2000 Balancing tran- lism of hydrogen cyanide in Sorghum infected by Gloeocercosporasorghi. Phytopathology 68: 1717–1722.scriptional interference and initiation on the GAL7 promoter of
Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 97: 8415–8420. Narendja, F. M., M. A. Davis and M. J. Hynes, 1999 AnCF, theCCAAT binding complex of Aspergillus nidulans, is essential forHa, S. B., A. P. Smith, R. Howden, W. M. Dietrich, S. Bugg et
al., 1999 Phytochelatin synthase genes from Arabidopsis and the the formation of a DNase I-hypersensitive site in the 59 regionof the amdS gene. Mol. Cell. Biol. 19: 6523–6531.yeast Schizosaccharomyces pombe. Plant Cell 11: 1153–1164.
Hutchings, H., K. P. Stahmann, S. Roels, E. A. Espeso, W. E. Novo, C., R. Tata, A. Clemente and P. R. Brown, 1995 Pseudomonasaeruginosa aliphatic amidase is related to the nitrilase/cyanideTimberlake et al., 1999 The multiply-regulated gabA gene en-
coding the GABA permease of Aspergillus nidulans: a score of hydratase enzyme family and Cys166 is predicted to be the activesite nucleophile of the catalytic mechanism. FEBS Lett. 367: 275–exons. Mol. Microbiol. 32: 557–568.
Hynes, M. J., 1970 Induction and repression of amidase enzymes 279.Oakley, B. R., J. E. Rinehart, B. L. Mitchell, C. E. Oakley, C.in Aspergillus nidulans. J. Bacteriol. 103: 482–487.
Hynes, M. J., 1972 Mutants with altered glucose repression of ami- Carmona et al., 1987a Cloning, mapping and molecular analysisof the pyrG (orotidine-59-phosphate decarboxylase) gene of Asper-dase enzymes in Aspergillus nidulans. J. Bacteriol. 111: 717–722.
Hynes, M. J., 1973 The effect of lack of a carbon source on nitrate- gillus nidulans. Gene 61: 385–399.Oakley, C. E., C. F. Weil, P. L. Kretz and B. R. Oakley, 1987breductase activity in Aspergillus nidulans. J. Gen. Microbiol. 79:
155–157. Cloning of the riboB locus of Aspergillus nidulans. Gene 53: 293–298.Hynes, M. J., 1974 The effects of carbon source on glutamate dehy-

131Regulation of the fmdS Gene of A. nidulans
Panozzo, C., E. Cornillot and B. Felenbok, 1998 The CreA repres- Sexton, A. C., and B. J. Howlett, 2000 Characterisation of a cya-sor is the sole DNA-binding protein responsible for carbon catab- nide hydratase gene in the phytopathogenic fungus Leptosphaeriaolite repression of the alcA gene in Aspergillus nidulans via its maculans. Mol. Gen. Genet. 263: 463–470.binding to a couple of specific sites. J. Biol. Chem. 273: 6367– Short, J. M., J. M. Fernandez, J. A. Sorge and W. D. Huse, 19886372. Lambda ZAP: a bacteriophage lambda expression vector with in
Papagiannopoulos, P., A. Andrianopoulos, J. A. Sharp, M. A. Davis vivo excision properties. Nucleic Acids Res. 16: 7583–7600.and M. J. Hynes, 1996 The hapC gene of Aspergillus nidulans is Small, A. J., M. J. Hynes and M. A. Davis, 1999 The TamA proteininvolved in the expression of CCAAT-containing promoters. Mol. fused to a DNA-binding domain can recruit AreA, the majorGen. Genet. 251: 412–421. nitrogen regulatory protein, to activate gene expression in Asper-
Peterson, J. A., and A. M. Myers, 1993 Functional analysis of mRNA gillus nidulans. Genetics 153: 95–105.39 end formation signals in the convergent and overlapping tran- Steidl, S., P. Papagiannopoulos, O. Litzka, A. Andrianopoulos,scription units of the S. cerevisiae genes RHO1 and MRP2. Nucleic M. A. Davis et al., 1999 AnCF, the CCAAT binding complex ofAcids Res. 21: 5500–5508. Aspergillus nidulans, contains products of the hapB, hapC, and
Platt, A., T. Langdon, H. Arst, D. Kirk, D. Tollervey et al., 1996a hapE genes and is required for activation by the pathway-specificNitrogen metabolite signalling involves the C-terminus and the regulatory gene amdR. Mol. Cell. Biol. 19: 99–106.GATA domain of the Aspergillus transcription factor AREA and Tusnady, G. E., and I. Simon, 1998 Principles governing aminothe 39 untranslated region of its mRNA. EMBO J. 15: 2791–2801. acid composition of integral membrane proteins: application to
Platt, A., A. Ravagnani, H. Arst, Jr., D. Kirk, T. Langdon et al., topology prediction. J. Mol. Biol. 283: 489–506.1996b Mutational analysis of the C-terminal region of AREA, Wang, P., D. E. Matthews and H. D. VanEtten, 1992 Purificationthe transcription factor mediating nitrogen metabolite repression and characterization of cyanide hydratase from the phytopatho-in Aspergillus nidulans. Mol. Gen. Genet. 250: 106–114. genic fungus Gloeocercospora sorghi. Arch. Biochem. Biophys. 298:
Polkinghorne, M. A., and M. J. Hynes, 1982 L-histidine utilization 569–575.in Aspergillus nidulans. J. Bacteriol. 149: 931–940. Wang, P., R. W. Sandrock and H. D. VanEtten, 1999 DisruptionPrade, R. A., J. Griffith, K. Kochut, J. Arnold and W. E. Tim- of the cyanide hydratase gene in Gloeocercospora sorghi increasesberlake, 1997 In vitro reconstruction of the Aspergillus (5 Emeri-
its sensitivity to the phytoanticipin cyanide but does not affectcella) nidulans genome. Proc. Natl. Acad. Sci. USA 94: 14564–its pathogenicity on the cyanogenic plant sorghum. Fungal Genet.14569.Biol. 28: 126–134.Proudfoot, N., 1991 Poly(A) signals. Cell 64: 671–674.
Wilson, R. A., and H. N. Arst, Jr., 1998 Mutational analysis ofPuig, S., J. E. Perez-Ortin and E. Matallana, 1999 TranscriptionalAREA, a transcriptional activator mediating nitrogen metaboliteand structural study of a region of two convergent overlappingrepression in Aspergillus nidulans and a member of the “streetwise”yeast genes. Curr. Microbiol. 39: 369–373.GATA family of transcription factors. Microbiol. Mol. Biol. Rev.Punt, P. J., M. A. Dingemanse, A. Kuyvenhoven, R. D. Soede,62: 586–596.P. H. Pouwels et al., 1990 Functional elements in the promoter
Wu, H. Y., S. H. Shyy, J. C. Wang and L. F. Liu, 1988 Transcriptionregion of the Aspergillus nidulans gpdA gene encoding glyceralde-generates positively and negatively supercoiled domains in thehyde-3-phosphate dehydrogenase. Gene 93: 101–109.template. Cell 53: 433–440.Punt, P. J., J. Strauss, R. Smit, J. R. Kinghorn, C. A. van den
Wyborn, N. R., D. J. Scherr and C. W. Jones, 1994 Purification,Hondel et al., 1995 The intergenic region between the diver-properties and heterologous expression of formamidase fromgently transcribed niiA and niaD genes of Aspergillus nidulansMethylophilus methylotrophus. Microbiology 140: 191–195.contains multiple NirA binding sites which act bidirectionally.
Wyborn, N. R., J. Mills, S. G. Williams and C. W. Jones, 1996 Mo-Mol. Cell. Biol. 15: 5688–5699.Ravagnani, A., L. Gorfinkiel, T. Langdon, G. Diallinas, E. Adjadj lecular characterisation of formamidase from Methylophilus methyl-
et al., 1997 Subtle hydrophobic interactions between the seventh otrophus. Eur. J. Biochem. 240: 314–322.residue of the zinc finger loop and the first base of an HGATAR Xiao, X., Y. H. Fu and G. A. Marzluf, 1995 The negative-actingsequence determine promoter-specific recognition by the Asper- NMR regulatory protein of Neurospora crassa binds to and inhibitsgillus nidulans GATA factor AreA. EMBO J. 16: 3974–3986. the DNA-binding activity of the positive-acting nitrogen regula-
Sambrook, J., E. F. Fritsch and T. Maniatis, 1989 Molecular Clon- tory protein NIT2. Biochemistry 34: 8861–8868.ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY. Communicating editor: R. H. Davis





























