Embed Size (px)
Citation preview

The Plant Cell, Vol. 1,265-273, March 1989 © 1989 American Society of Plant Physiologists
Production of Cell Wall-Degrading Enzymes by Aspergillus nidulans: A Model System for Fungal Pathogenesis of Plants
Ralph A. Dean" and William E. Timberlake a,b,1
a Department of Plant Pathology, University of Georgia, Athens, Georgia 30602 b Department of Genetics, University of Georgia, Athens, Georgia 30602
The cell wall-degrading enzymes polygalacturonase and pectate lyase have been suggested to be crucial for penetration and colonization of plant tissues by some fungal pathogens. We have found that Aspergillus nidulans (-Emericella nidulans), a saprophytic Ascomycete, produces levels of these enzymes equal to those produced by soft-rotting Erwinia species. Induction of polygalacturonase and pectate lyase in A. nidulans requires substrate and is completely repressed by glucose. Surprisingly, inoculation of excised plant tissues with A. nidulans conidia leads to formation of necrotic, water-soaked lesions within which the organism sporulates. Thus, A. nidulans has phytopathogenic potential. The release of glucose and other sugars from wounded tissues may repress pectolytic enzyme production and limit disease development. Therefore, we tested creA204, a mutation that relieves glucose repression of some A. nidulans carbon utilization enzymes, for its effect on production of pectolytic enzymes. creA204 failed to relieve catabolite repression of polygalacturonase or pectate lyase and had no effect on disease severity.
INTRODUCTION
The cell walls of living plants carry out diverse functions, one of which is the exclusion of pathogens. When plant cells die, their walls provide a source of nutrients for many organisms, including potentially pathogenic microorga- nisms. The complex chemical composition and physical structure of plant cell walls make them difficult to penetrate and degrade. Various polymers, including cellulose and protein, are embedded in a matrix of highly branched polysaccharides (McNeil et al., 1984; Selvendran and O'Neill, 1987). One of the simplest of these polysaccha- rides, pectin, is a major component of the middle lamella and acts as an intercellular cement (Talmadge et al., 1973). When presented with purified cell walls in defined culture medium, the first enzymes produced by many microorga- nisms are pectin-degrading enzymes (Jones et al., 1972; Mankarios and Friend, 1980). Numerous pectin-degrading enzymes have been identified, including endo- and exo- forms of lyases and hydrolases (Bateman and Basham, 1976). Depolymerization of pectin has been reported to be a prerequisite for further cell wall breakdown (Bauer et al., 1973, 1977; Keegstra et al., 1973),
Many plant pathogenic fungi produce pectin-degrading enzymes in culture (Cooper, 1983; Keon et al., 1987). In planta, these enzymes may cause tissue maceration and cell death. However, the actual role they play in plant pathogenesis remains to be determined; whether these
1 To whom correspondence should be addressed.
enzymes are disease determinants or simply modify the course of disease development is unknown. It should be noted that microbial pectin-degrading enzymes may also serve to activate host defense systems; enzymatically released pectin fragments have been proposed to be endogenous elicitors of plant defense responses (Bishop et al., 1981; Lee and West, 1981; Bruce and West, 1982; Davis et al., 1986; Robertsen, 1987).
The mechanisms controlling expression of cell wall- degrading enzymes in eukaryotes are not well understood. In culture, production of pectolytic enzymes by many fungi, including plant pathogens, requires substrate and is re- pressed by preferred Carbon sources such as glucose (Cooper, 1983). Catabolite repression has been proposed to explain the diphasic growth pattern exhibited by some hemibiotrophs and thereby the way in which these patho- gens cause disease. For example, few symptoms are visible during initial infection of bean by Colletotrichum lindemuthianum. Subsequent tissue collapse has been correlated with and proposed to result from the production of large amounts of pectolytic enzymes (Wijesundera et al., 1984). On the other hand, with necrotrophs (Botrytis allii and Rhizoctonia solani, for example) production of pectolytic enzymes begins early during and may even precede infection (Cooper, 1983; Marcus et al., 1986). It is thus possible that some phytopathogens circumvent catabolite repression.
Aspergillus nidulans is one of only a few fungi that have

266 The Plant Cell
a highly developed molecular genetic system. A. nidulans is closely related to many phytopathogens but is not a significant etiological agent itself. However, in this paper, we show that A. nidulans has a limited, but significant, phytopathogenic potential. We further demonstrate that it can efficiently utilize polygalacturonic acid as sole carbon source. Polygalacturonase and pectate lyase are produced sequentially during growth on polygalacturonic acid and their production is subject to catabolite repression. Inter- estingly, the catabolite derepression mutation creA204 (Hynes and Kelly, 1977) does not relieve repression of these enzymes. In the accompanying paper (Dean and Timberlake, 1989), we report isolation, characterization, and inactivation of the single copy A. nidulans pectate lyase gene (pelA).
RESULTS
Growth of A. nidulans on Plant Tissues
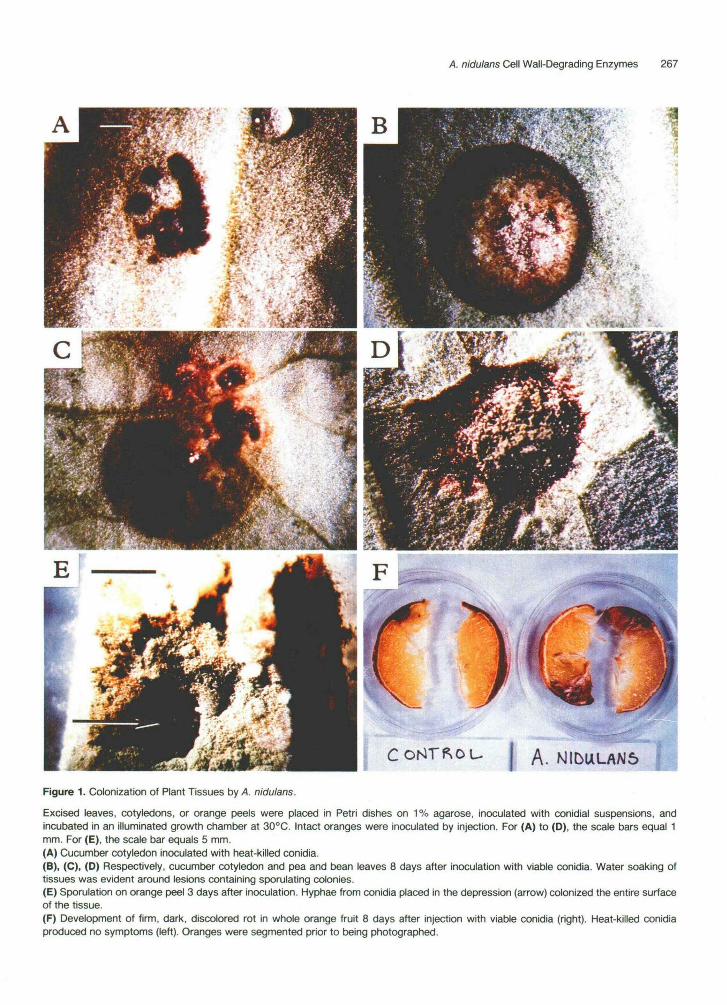
We investigated the ability of A. nidulans to cause plant disease with an infection assay utilizing detached plant tissues. Tissues were placed on 1% agarose, a medium that does not support growth of the fungus, and inoculated with drops of a conidial suspension. Results from prelimi- nary experiments showed that no detectable infection occurred unless the tissues were wounded at the site of inoculation. Inoculation of wounded sites with heat-killed conidia caused no further tissue damage (Figure 1A). However, when cucumber cotyledons or pea or bean leaves were inoculated with viable conidia, water-soaking and host cell death occurred within a few days. By 5 days, sporulation was evident in the lesions (Figure 1, B to D). Inoculation of the inner surface of orange fruit rinds re- sulted in rapid fungal growth and extensive sporulation. The tissues were completely colonized within 3 days of inoculation (Figure 1E). When whole orange fruits were inoculated by injecting conidia, the fungus grew into the flesh and by 8 days caused a firm, dark, discolored rot (Figure 1 F).
Utilization of Polygalacturonic Acid as Carbon Source
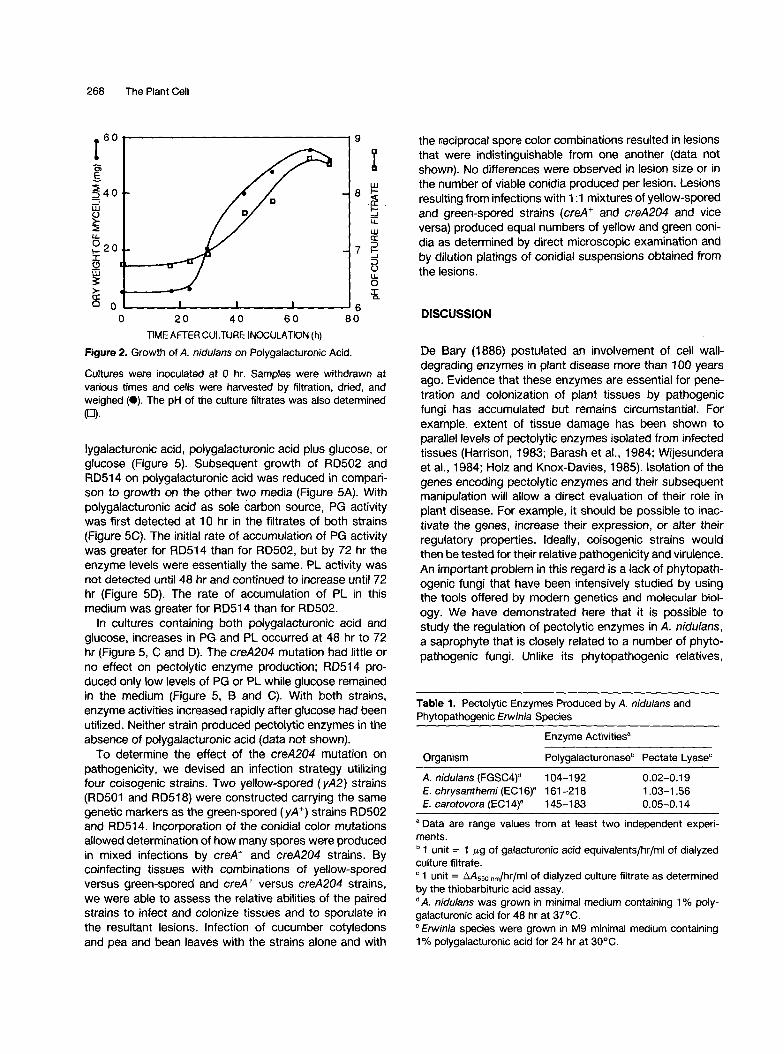
Formation of water-soaked lesions on leaves and cotyle- dons and growth on orange fruit rinds indicated that A. nidulans was capable of utilizing pectin as a nutrient source. To test this possibility, we grew the organism in minimal medium containing polygalacturonic acid as sole carbon source. After a lag of ~24 hr, mycelial dry weight increased in a typical sigmoidal fashion, reaching a maxi- mum by -65 hr (Figure 2). At this time culture filtrates contained two major classes of pectolytic enzymes, poly- galacturonase (PG) and pectate lyase (PL). Levels of PG activity produced by A. nidulans were similar to those produced by the soft-rotting Erwinia species, E. carotovora
and E. chrysanthemi (Table 1). The levels of PL produced by A. nidulans and E. carotovora were also similar, but were ~10-fold lower than the level produced by E. chrysanthemi.
Production of PG and PL
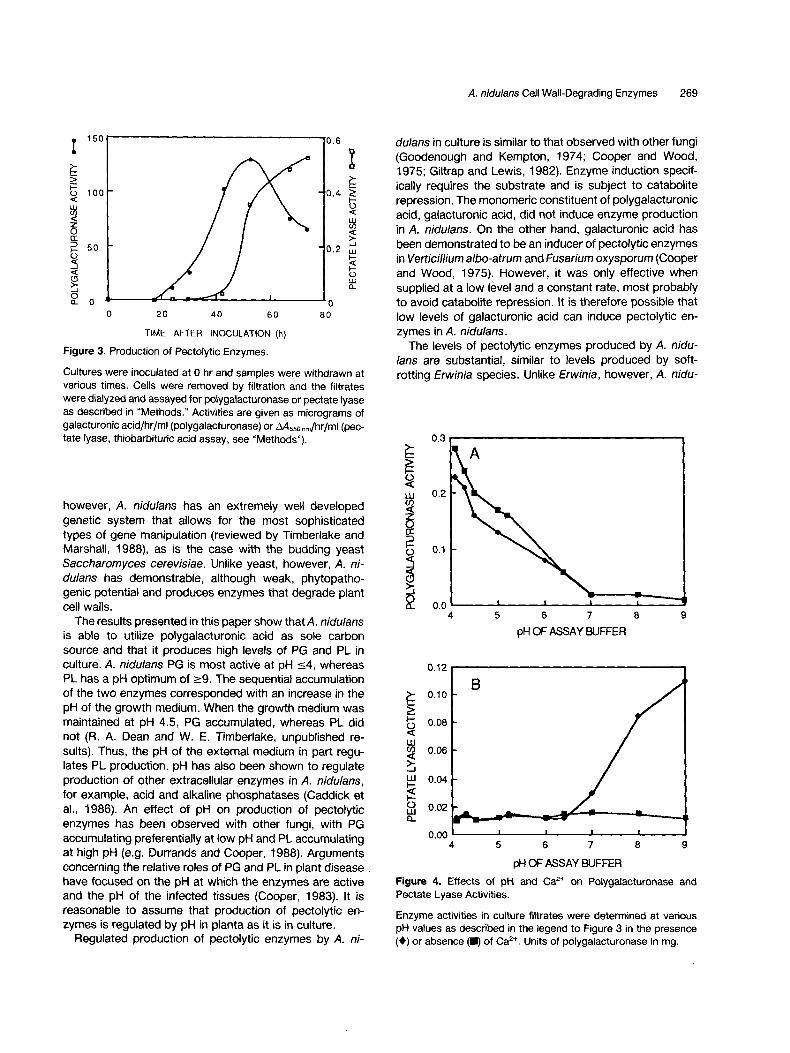
The time course of production of extracellular PG and PL by A. nidulans grown in minimal medium containing poly- galacturonic acid as sole carbon source is shown in Figure 3. PG activity was first detected at 24 hr after inoculation and its appearance coincided with the onset of growth (see Figure 1). Activity in the filtrate continued to increase until it reached a maximum at ~50 hr and then began to decrease. PL activity was first detected at 42 hr and continued to increase until 72 hr. The pH of the culture filtrate increased from 6.6 to 8.6 during the course of the experiment (Figure 1). The optimum pH for PG was _<4, whereas that for PL was ->9 (Figure 4). Thus, the times of accumulation of the two enzymes were consistent with their pH optima. PL activity was dependent on Ca 2÷, whereas PG activity was not (Figure 4).
Catabolite Repression of Pectolytic Enzyme Production
A. nidulans was grown on a variety of carbon sources to investigate the induction and repression of pectolytic en- zymes. Table 2 shows that pectolytic enzyme activities were only detected in filtrates from cultures grown with polygalacturonic acid. When glucose was added with po- lygalacturonic acid, no pectolytic enzyme activities were detected. Thus, pectolytic enzyme production is subject to catabolite repression. Polygalacturonic acid appeared to be a specific inducer of pectolytic enzymes because the enzymes were not produced when A. nidulans was grown on other suboptimal carbon sources, including galacturonic acid.
Effect of the creA204 Mutation on Pectolytic Enzyme Production and Growth on Plants
One possible explanation for the limited pathogenic ability of A. nidulans is that enzymes for cell wall breakdown are subject to carbon catabolite repression in planta. In an attempt to test this hypothesis, we examined the ability of the creA204 mutation to relieve glucose repression of pectolytic enzymes. This mutation partially relieves catab- olite repression of a number of enzymes involved in ac- quisition of suboptimal carbon sources such as ethanol and certain amino acids (Hynes and Kelly, 1977; Kelly and Hynes, 1977). Coisogenic strains RD502 (creA +) and RD514 (creA204) were germinated in medium containing glucose and transferred to medium containing either po-
A. nidulans Cell Wall-Degrading Enzymes 267
MI5ULAN6
Figure 1. Colonization of Plant Tissues by A. nidulans.
Excised leaves, cotyledons, or orange peels were placed in Petri dishes on 1% agarose, inoculated with conidial suspensions, andincubated in an illuminated growth chamber at 30°C. Intact oranges were inoculated by injection. For (A) to (D), the scale bars equal 1mm. For (E), the scale bar equals 5 mm.(A) Cucumber cotyledon inoculated with heat-killed conidia.(B), (C), (D) Respectively, cucumber cotyledon and pea and bean leaves 8 days after inoculation with viable conidia. Water soaking oftissues was evident around lesions containing sporulating colonies.(E) Sporulation on orange peel 3 days after inoculation. Hyphae from conidia placed in the depression (arrow) colonized the entire surfaceof the tissue.(F) Development of firm, dark, discolored rot in whole orange fruit 8 days after injection with viable conidia (right). Heat-killed conidiaproduced no symptoms (left). Oranges were segmented prior to being photographed.
268 The Plant Cell
60 9 I
W _ 8
2 2 0 7 z
0
0 >-
Q 0 I I I 6 0 20 40 60 80
TIME AFTER CULTURE INOCULATION (h)
Figure 2. Growth of A. nidu/ans on Polygalacturonic Acid.
Cultures were inoculated at 0 hr. Samples were withdrawn at various times and cells were harvested by filtration, dried, and weighed (0). The pH of the culture filtrates was also determined (~).
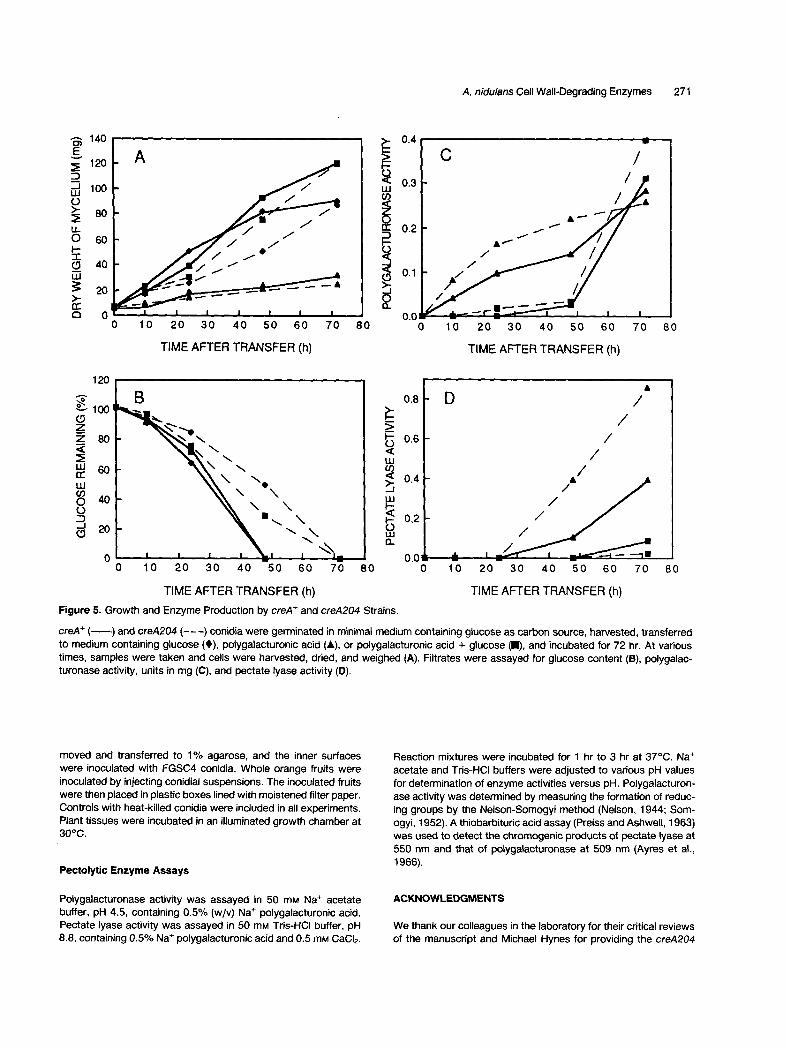
lygalacturonic acid, polygalacturonic acid plus glucose, or glucose (Figure 5). Subsequent growth of RD502 and RD514 on polygalacturonic acid was reduced in compari- son to growth on the other two media (Figure 5A). With polygalacturonic acid as sole Carbon source, PG activity was first detected at 10 hr in the filtrates of both strains (Figure 5C). The initial rate of accumulation of PG activity was greater for RD514 than for RD502, but by 72 hr the enzyme levels were essentially the same. PL activity was not detected until 48 hr and continued to increase until 72 hr (Figure 5D). The rate of accumulation of PL in this medium was greater for RD514 than for RD502.
In cultures containing both polygalacturonic acid and glucose, increases in PG and PL occurred at 48 hr to 72 hr (Figure 5, C and D). The creA204 mutation had little or no effect on pectolytic enzyme production; RD514 pro- duced only low levels of PG or PL while glucose remained in the medium (Figure 5, B and C). With both strains, enzyme activities increased rapidly after glucose had been utilized. Neither strain produced pectolytic enzymes in the absence of polygalacturonic acid (data not shown).
To determine the effect of the creA204 mutation on pathogenicity, we devised an infection strategy utilizing four coisogenic strains. Two yellow-spored (yA2) strains (RD501 and RD518) were constructed carrying the same genetic markers as the green-spored (yA ÷) strains RD502 and RD514. Incorporation of the conidial color mutations allowed determination of how many spores were produced in mixed infections by creA ÷ and creA204 strains. By coinfecting tissues with combinations of yellow-spored versus green-spored and creA ÷ versus creA204 strains, we were able to assess the relative abilities of the paired strains to infect and colonize tissues and to sporulate in the resultant lesions. Infection of cucumber cotyledons and pea and bean leaves with the strains alone and with
the reciprocal spore color combinations resulted in lesions that were indistinguishable from one another (data not shown). No differences were observed in lesion size or in the number of viable conidia produced per lesion. Lesions resulting from infections with 1:1 mixtures of yellow-spored and green-spored strains (creA ÷ and creA204 and vice versa) produced equal numbers of yellow and green coni- dia as determined by direct microscopic examination and by dilution platings of conidial suspensions obtained from the lesions.
DISCUSSION
De Bary (1886) postulated an involvement of cell wall- degrading enzymes in plant disease more than 100 years ago. Evidence that these enzymes are essential for pene- tration and colonization of plant tissues by pathogenic fungi has accumulated but remains circumstantial. For example, extent of tissue damage has been shown to parallel levels of pectolytJc enzymes isolated from infected tissues (Harrison, 1983; Barash et al., 1984; Wijesundera et al., 1984; Holz and Knox-Davies, 1985). Isolation of the genes encoding pectolytic enzymes and their subsequent manipulation will allow a direct evaluation of their role in plant disease. For example, it should be possible to inac- tivate the genes, increase their expression, or alter their regulatory properties. Ideally, coisogenic strains would then be tested for their relative pathogenicity and virulence. An important problem in this regard is a lack of phytopath- ogenic fungi that have been intensively studied by using the tools offered by modern genetics and molecular biol- ogy. We have demonstrated here that it is possible to study the regulation of pectolytic enzymes in A. nidulans, a saprophyte that is closely related to a number of phyto- pathogenic fungi. Unlike its phytopathogenic relatives,
Table 1. Pectolytic Enzymes Produced by A. nidulans and Phytopathogenic Erwinia Species
Enzyme Activities a
O r g a n i s m Polygalacturonase b Pectate Lyase c
A. nidulans (FGSC4) d 104-192 0.02-0.19 E. chrysanthemi (EC16) e 161-218 1.03-1.56 E. carotovora (EC14) e 145-183 0.05-0.14
a Data are range values from at least two independent experi- ments. b 1 unit = 1 ~g of galacturonic acid equivalents/hr/ml of dialyzed culture filtrate. c 1 unit = AAs~0 om/hr/ml of dialyzed culture filtrate as determined by the thiobarbitudc acid assay. A. nidulans was grown in minimal medium containing 1% poly-
galacturonic acid for 48 hr at 37°C. e Erwinia species were grown in M9 minimal medium containing 1% polygalacturonic acid for 24 hr at 30°C.
A. nidulans Cell Wall-Degrading Enzymes 269
1 5 0
>__
O 1 0 0 < u..I (,3
Z o r r ~, s o
5
2 0 4 0
T I M E
I
6 0
A F T E R I N O C U L A T I O N (h)
0 . 6
0.3
0 . 4 >--_, l - iD < ILl u3
0 . 2 ,,, F--
0 t ~
o 8 0
Figure 3. Production of Pectolytic Enzymes.
Cultures were inoculated at 0 hr and samples were withdrawn at various times. Cells were removed by filtration and the filtrates were dialyzed and assayed for polygalacturonase or pectate lyase as described in "Methods." Activities are given as micrograms of galacturonic acid/hr/ml (polygalacturonase) or ,~Ass0 nm/hr/ml (pec- tate lyase, thiobarbituric acid assay, see "Methods").
however, A. nidulans has an extremely well developed genetic system that allows for the most sophisticated types of gene manipulation (reviewed by Timberlake and Marshall, 1988), as is the case with the budding yeast Saccharomyces cerevisiae. Unlike yeast, however, A. ni- dulans has demonstrable, although weak, phytopatho- genic potential and produces enzymes that degrade plant cell walls.
The results presented in this paper show that A. nidulans is able to utilize polygalacturonic acid as sole carbon source and that it produces high levels of PG and PL in culture. A. nidulans PG is most active at pH <_4, whereas PL has a pH optimum of _>9. The sequential accumulation of the two enzymes corresponded with an increase in the pH of the growth medium. When the growth medium was maintained at pH 4.5, PG accumulated, whereas PL did not (R. A. Dean and W. E. Timberlake, unpublished re- sults). Thus, the pH of the external medium in part regu- lates PL production, pH has also been shown to regulate production of other extracellular enzymes in A. nidulans, for example, acid and alkaline phosphatases (Caddick et al., 1986). An effect of pH on production of pectolytic enzymes has been observed with other fungi, with PG accumulating preferentially at low pH and PL accumulating at high pH (e.g. Durrands and Cooper, 1988). Arguments concerning the relative roles of PG and PL in plant disease have focused on the pH at which the enzymes are active and the pH of the infected tissues (Cooper, 1983). It is reasonable to assume that production of pectolytic en- zymes is regulated by pH in planta as it is in culture.
Regulated production of pectolytic enzymes by A. ni-
dulans in culture is similar to that observed with other fungi (Goodenough and Kempton, 1974; Cooper and Wood, 1975; Giltrap and Lewis, 1982). Enzyme induction specif- ically requires the substrate and is subject to catabolite repression. The monomeric constituent of polygalacturonic acid, galacturonic acid, did not induce enzyme production in A. nidulans. On the other hand, galacturonic acid has been demonstrated to be an inducer of pectolytic enzymes in Verticillium albo-atrum and Fusariurn oxysporum (Cooper and Wood, 1975). However, it was only effective when supplied at a low level and a constant rate, most probably to avoid catabolite repression. It is therefore possible that low levels of galacturonic acid can induce pectolytic en- zymes in A. nidulans.
The levels of pectolytic enzymes produced by A. nidu- lans are substantial, similar to levels produced by soft- rotting Erwinia species. Unlike Erwinia, however, A. nidu-
pH OF ASSAY BUFFER
LU 0.2
0.1
l 0.0
A
5 6 7 8 9
0.12
I I I I
0.10
.~ 0.08
LU 0.06
uJ 0.04
0.02 t3_
0.00 4 5 6 7 8 9
pHOFASSAYBUFFER
Figure 4. Effects of pH and Ca 2+ on Polygalacturonase and Pectate Lyase Activities.
Enzyme activities in culture filtrates were determined at various pH values as described in the legend to Figure 3 in the presence (0) or absence (I) of Ca 2÷. Units of polygalacturonase in mg.

270 The Plant Ceil
Table 2. Growth and Pectolytic Enzymes Produced by A. nidulans on Various Carbon Sources
Carbon Source a Growth ~ Pectolytic Enzymes c
Polygalacturonic acid ++ + Polygalacturonic acid + glucose +++ - Glucose +++ - Lactose +++ - Maltose +++ - Sucrose +++ - Ethanol ++ - Acetate + + - Galacturonic acid ++ -
a A. nidulans was grown in liquid culture on minimal medium containing 1% carbon source for 50 hr. b +++ = good, ++ = moderate. c+ = polygalacturonase and pectate lyase activities detected in dialyzed culture filtrate, - = neither activity detected.
lans is capable of causing only limited plant disease, even when inoculated onto excised and wounded plant tissues. There are many possible explanations for this weak patho- genic ability. One parameter affecting aggressiveness may be the sensitivity of pectolytic enzyme production by A. nidulans to catabolite repression. Catabolite repression has been suggested to underlie disease resistance in some plant-pathogen interactions (Holz and Knox-Davies, 1986a, 1986b). It has also been shown that treatments resulting in increased soluble carbohydrates in plant tis- sues decrease enzyme production by pathogens and re- duce symptom severity (Horton and Keen, 1966; Patil and Dimond, 1968; Weinhold and Bowman, 1974). Genes af- fecting carbon catabolite repression have been described in A. nidulans (Bailey and Arst, 1975; Hynes and Kelly, 1977). We determined that a mutant allele of one such gene (creA204) had little effect on production of pectolytic enzymes. Mutations at the creA locus have been shown to derepress many enzymes involved in carbon acquisition. However, exceptions, for example quinate dehydrogenase and/~-galactosidase, are known (Bailey and Arst, 1975). To alleviate carbon catabolite repression of pectolytic en- zymes, we are taking advantage of the A. nidulans molec- ular genetic system. We have cloned the unique gene encoding pectate lyase (pelA) as described in the accom- panying paper (Dean and Timberlake, 1989). It is possible to place this and other genes under the control of various regulatory elements and promoters and then to examine the effects of altered levels of expression and regulatory patterns on phenotype (Adams et al., 1988). Evaluation of the importance of PL and PG for pathogenicity and viru- lence will involve forcing their expression at high levels independent of carbon catabolite repression, pH, and other factors.
METHODS
Strains, Media, and Growth Conditions
Aspergillus nidulans FGSC4 (Glasgow wild-type) and FGSC237 (yA2, pabaA 1; trpC801 ) were obtained from the Fungal Genetics Stock Center. Strains RD501 (yA2; creA+), RD502 (yA+; creA+), RD514 (yA÷; creA204) and RD518 (yA2; creA204) were meiotic progeny from a cross of FGSC237 and MH664 (biA1; niiA4; creA204; provided by M. Hynes). Bacterial strains of Erwinia carotovora (EC14) and E. chrysanthemi (EC16) were provided by D.P. Roberts and N.T. Keen, respectively. Fungal strains were maintained on appropriately supplemented minimal medium (Pon- tecorvo et al., 1953; Clutterbuck, 1974; K~fer, 1977; Timberlake and Hamer, 1986).
For enzyme induction studies on different carbon sources and time course experiments with FGSC4, minimal medium cultures containing 1% (w/v) carbon source were inoculated to a density of 1 x 105 conidia/ml and shaken at 37°C. Cultures were har- vested by filtration through Whatman No. 1 paper. Filtrates were dialyzed against deionized water overnight at 4°C prior to being assayed for pectolytic enzyme activities. For time course experi- ments, duplicate 20-ml samples were withdrawn at intervals over 72 hr and filtered through tared Whatman No. 1 filter papers. The filters were dried overnight at 55°C and the dry weights of mycelia were determined. The pH of the filtrates was measured prior to dialysis. For time course experiments with creA204 mutants, strains were inoculated at a density of 1 × 106 conidia/ml in minimal medium containing 1% glucose and shaken at 37°C for 14 hr. Cells were harvested by filtration through Mira-Cloth (Behr- ing Diagnostics), washed with minimal medium, divided equally into minimal medium containing either 1% glucose, 1% polygalac- turonic acid, or 1% glucose + 1% polygalacturonic acid and incubated as above. Polygalacturonic acid was washed with ethanol prior to use.
The amount of glucose remaining in the culture filtrates was determined by glucose oxidase coupled to the formation of col- ored quinoneimine dye by peroxidase (Sigma Diagnostic Kit No. 315). To compare levels of pectolytic enzymes produced by A. nidulans to other organisms, E. carotovora and E. chrysanthemi cells were grown in M9 minimal medium containing 1% polygalac- turonic acid at 30°C with shaking for 24 hr. Cells were pelleted at 10,000g for 10 min. Supernatants were dialyzed and assayed for pectolytic enzyme activities as described above.
Inoculation of Plant Tissues
Leaves from pea (Pisum sativum cv Alaska) and bean (Phaseolus vulgaris cv White half runner) and cotyledons from cucumber (Cucumis sativus cv Marketer) were detached from growth cham- ber-grown seedlings. The tissues were surface-sterilized with 10% bleach for 5 rain, rinsed in sterile distilled water, and placed on 1% agarose in Petd dishes. Tissues were inoculated with 10- #1 drops of a suspension containing 1 x 10 ~ conidia/ml and then wounded through the drops with a sterile needle. To determine the effect of the creA204 mutation, plant tissues were inoculated as above with either RD501, RD502, RD514, and RD518 sepa- rately or in all pairwise combinations. For the infection of orange, fruits were surface-sterilized with 70% ethanol. Rinds were re-
A. nidulans Cell Wall-Degrading Enzymes 271
m
10o 03b iZ
e / / "
~ / ~, .
0 -' - oo . . . . . . . , ' , ' 0 10 20 30 40 50 60 70 80 0 10 20 30 40 50 60 70 80
TIME AFTER TRANSFER (h) T IME AFTER T R A N S F E R (h)
120
100 C9 _z z 8O <
w 60 n" IJJ o9 O 4O
-J 20
B
~ \ \e\
\ . .x , , , ,%._
0 10 20 30 40 50 60 70
0.8 >
0.6 < uJ
~ 0.4
~ 0.2 o.
D
/ /
/
/
/
/JJ 0 0.0"- u
80 0 10 20 30 40 50 60 70 80
TIME AFTER TRANSFER (h) T IME AFTER TRANSFER (h)
Figure 5. Growth and Enzyme Production by creA ÷ and creA204 Strains.
creA + ( ) and creA204 ( - - - ) conidia were germinated in minimal medium containing glucose as carbon source, harvested, transferred to medium containing glucose (0), polygalacturonic acid (A), or polygalacturonic acid + glucose ( i ) , and incubated for 72 hr. At various times, samples were taken and cells were harvested, dried, and weighed (A). Filtrates were assayed for glucose content (B), polygalac- turonase activity, units in mg (C), and pectate lyase activity (D).
moved and transferred to 1% agarose, and the inner surfaces were inoculated with FGSC4 conidia. Whole orange fruits were inoculated by injecting conidial suspensions. The inoculated fruits were then placed in plastic boxes lined with moistened filter paper. Controls with heat-killed conidia were included in all experiments. Plant tissues were incubated in an illuminated growth chamber at 30°C.
Pectolytic Enzyme Assays
Polygalacturonase activity was assayed in 50 mM Na + acetate buffer, pH 4.5, containing 0.5% (w/v) Na ÷ polygalacturonic acid. Pectate lyase activity was assayed in 50 mM Tris-HCI buffer, pH 8.8, containing 0.5% Na + polygalacturonic acid and 0.5 mM CaCI2.
Reaction mixtures were incubated for 1 hr to 3 hr at 37°C. Na + acetate and Tris-HCI buffers were adjusted to various pH values for determination of enzyme activities versus pH. Polygalacturon- ase activity was determined by measuring the formation of reduc- ing groups by the Nelson-Somogyi method (Nelson, 1944; Som- ogyi, 1952). A thiobarbituric acid assay (Preiss and Ashwell, 1963) was used to detect the chromogenic products of pectate lyase at 550 nm and that of polygalacturonase at 509 nm (Ayres et al., 1966).
ACKNOWLEDGMENTS
We thank our colleagues in the laboratory for their critical reviews of the manuscript and Michael Hynes for providing the creA204

272 The Plant Cell
strain. This research was supported by grants to W.E.T. from the U.S. Environmental Protection Agency (with O. Yoder and H. Van Etten), the U,S. Department of Agriculture/Central Research Grants Office, and National Institutes of Health.
Received January 12, 1989.
REFERENCES
Adams, T. H., Boylan, M. T., and Timberlake, W. E. (1988). brlA is necessary and sufficient to direct conidiophore development in Aspergi//us nidulans. Cell 54, 353-362.
Ayres, W. A., Papavizas, G. C., and Diem, A. F. (1966). Poly- galacturonate transeliminase and polygalacturonase production by Rhizoctonia so/ani. Phytopathology 56, 1006-1011.
Bailey, C., and Arst, H. N., Jr. (1975). Carbon catabolite repres- sion in Aspergi//us nidulans. Eur. J. Biochem. 51,573-577.
Barash, I., Zilberman, E., and Marcus, L. (1984). Purification of Geotrichum candidum endopolygalacturonase from culture and from host tissue by affinity chromatography on cross-linked polypectate, Physiol. Plant Pathol. 25, 161-169.
Bateman, D. F., and Basham, H. G. (1976), Degradation of plant cell walls and membranes by microbial enzymes, In Encyclo- pedia of Plant Physiology, Vol. 4, R. Heitefuss and P. H. Williams, eds (New York: Springer-Verlag), pp. 316-355.
Bauer, W. D., Talmadge, K. W., Keegstra, K., and Albersheim, P. (1973). The structure of plant cell walls II, The hemicellulose of the cell walls of suspension-cultured sycamore cells. Plant Physiol. 51,174-187.
Bauer, W. D., Bateman, D. F., and Whalen, C. H. (1977). Purifi- cation of an endo-/~-1,4-galactanase produced by Sc/erotinia sc/erotiorum: Effects on isolated plant cell walls and potato tissue. Phytopathology 67, 862-868.
Bishop, P. D., Makus, D. J., Pearce, G., and Ryan, C. A. (1981). Proteinase inhibitor-inducing factor activity in tomato leaves resides in oligosaccharides enzymatically released from the cell walls. Proc. Natl. Acad. Sci. USA 78, 3536-3540.
Bruce, R. J., and West, C. A. (1982). Elicitation of casbene synthetase activity in castor bean. The role of pectic fragments of the plant cell wall in elicitation by a fungal endopolygalactu- ronase. Plant Physiol. 69, 1181-1188.
Caddick, M. X., Brownlee, A. G., and Arst, H. N., Jr. (1986). Regulation of gene expression by pH of the growth medium in Aspergi//us nidu/ans. Mol. Gen. Genet. 203, 346-353.
Clutterbuck, A. J. (1974). Aspergi//us nic/u/ans. In Handbook of Genetics, Vol. 1, R. C. King, ed (New York: Plenum Publishing), pp. 447-510.
Cooper, R. M. (1983). The mechanisms and significance of en- zymatic breakdown of host cell walls by parasites. In Biochem- ical Plant Pathology, J. A. Callow, ed (New York: John Wiley and Sons), pp. 101-135.
Cooper, R. M., and Wood, R. K. S. (1975). Regulation of synthesis of cell-wall degrading enzymes by Vertici//ium a/bo-atrum and Fusarium oxysporum f. sp. lycopersici. Physiol. Plant Pathol, 5, 135-156.
Davis, K. R., Darvill, A. G., AIbersheim, P., and Dell, A. (1986). Host-pathogen interactions XXX. Characterization of elicitors of phytoalexin accumulation in soybean released from soybean cell walls by endopolygalacturonic acid lyase. Z. Naturtorsch. 41c, 39-48.
Dean, R. A., and Timberlake, W. E. (1989). Regulation of the Aspergillus nidulans pectate lyase gene (pelA). Plant Cell 1, 275-284.
De Bary, A. (1886). Ueber einige Sclerotinien und Sclerotien- Krankheiten. Botanische Zeitung 44, 376-480.
Durrands, P. K., and Cooper, R. M. (1988). Development and analysis of pectic screening media for the use in the detection of pectinase mutants. Appl, Microbiol. Biotechnol. 28, 463-467.
Giltrap, N. J., and Lewis, D. H. (1982). Catabolite repression of the synthesis of pectin-degrading enzymes by Suillus luteus (L. ex Fr.) S.F. Gray and Hebeloma oculatum Bruchet. New Phytol. 90, 485-493..
Goodenough, P. W., and Kempton, R. J. (1974). Regulation of the synthesis of an extracellular polygalacturonase by Pyreno- chaeta lycopersici when grown in media with limited carbon sources. Phytopathol. Z. 81, 78-84.
Harrison, J. G. (1983). Growth of lesions caused by Botrytis fabae on field bean leaves in relation to foliar bacteria, non-enzymatic phytotoxins, pectic enzymes and osmotica. Ann, Bot. (Lond.) 52, 823-838.
Holz, G., and Knox-Davies, P. S. (1985). Production of pectic enzymes by Fusarium oxysporum f. sp. cepae and its involve- ment in onion rot. Phytopathol. Z. 112, 69-80.
Holz, G., and Knox-Davies, P. S. (1986a). Possible involvement of apoplast sugar in endo-pectin-trans-eliminase synthesis and onion bulb rot caused by Fusarium oxysporum f. sp. cepae. Physiol, Mol. Plant Pathol. 28, 403-410.
Holz, G., and Knox-Davies, P. S. (1986b). Relation between endo-pectin-trans-eliminase and apoplast-symplast sugars in Fusarium bulb rot of onions. Physiol. Mol. Plant Pathol. 28, 411-421.
Horton, J. C., and Keen, N. T. (1966). Sugar repression of endopolygalacturonase and cellulase synthesis during patho- genesis by Pyrenochaeta terrestris as a resistance mechanism in onion pink root. Phytopathology 56, 908-916.
Hynes, M. J., and Kelly, J. M. (1977). Pleiotropic mutants of Aspergillus nidulans altered in carbon metabolism. Mol. Gen. Genet. 150, 193-204.
Jones, T. M., Anderson, A. J., and Albersheim, P. (1972). Host- pathogen interactions IV. Studies on the polysaccharide de- grading enzymes secreted by Fusarium oxysporum f. sp. lyco- persicL Physiol. Plant Pathol. 2, 153-166.
Kafer, E. (1977). Meiotic and mitotic recombination in Aspergillus and its chromosomal aberrations, Adv. Genet. 19, 33-131.
Keegstra, K., Talmadge, K. W., Bauer, W. D., and Albersheim, P. (1973). The structure of plant cell walls II1. A model of the wall of suspension-cultured sycamore cells based on intercon- nections of the macromoleeular components. Plant Physiol, 51, 188-197.
Kelly, J. M., and Hynes, M. J. (1977), Increased and decreased sensitivity to carbon catabolite repression of enzymes of acetate metabolism in mutants ofAspergi//us nidu/ans. Mol. Gen. Genet. 156, 87-92.
Keon, J. P. R., Byrde, R. J. W., and Cooper, R. M. (1987). Some

A. nidulans Cell Wall-Degrading Enzymes 273
aspects of fungal enzymes that degrade plant cell walls. In Fungal Infection of Plants, G. F. Pegg and P. G. Ayres, eds (Cambridge: Cambridge University Press), pp. 133-157.
Lee, S. C., and West, C. A. (1981). Polygalacturonase from Rhizopus stolonifer, an elicitor of casbene synthetase in castor bean (Ricinus communis L.) seedlings. Plant Physiol. 67, 633- 639.
McNeil, M., Darvill, A. G., Fry, S. C., and Albersheim, P. (1984). Structure and function of the primary cell walls of plants. Annu. Rev. Biochem. 53, 625-663.
Mankarios, A. T., and Friend, J. (1980). Polysaccharide degrad- ing enzymes of Botrytis a//ii and Sc/erotium cepivorum. Enzyme production in culture and the effect of the enzymes on isolated onion cell walls. Physiol. Plant Pathol. 17, 93-104.
Marcus, L., Barash, I., Sneh, B., Koltin, Y., and Finkler, A. (1986). Purification and characterization of pectolytic enzymes produced by virulent and hypovirulent isolates of Rhizoctonia so/ani. Kuhn. Physiol. Mol. Plant Pathol. 29, 325-336.
Nelson, N. (1944). A photometric adaption of the Somogyi method for the determination of glucose. J. Biol. Chem. 153, 375-380.
Patil, S. S., end Dimond, A. E. (1968). Repression of polygalac- turonase synthesis in Fusarium oxysporum f. sp./ycopersici by sugars and its effect on symptom reduction in infected tomato plants. Phytopathology 58, 676-682.
Pontecorvo, G., Roper, J. A., Hemmons, L. M., MacDonald, K. D., and Bufton, A. W. J. (1953). The genetics of Aspergi//us nidu/ans. Adv. Genet. 5, 141-238.
Preiss, J., and Ashwell, G. (1963). Polygalacturonic acid metab- olism in bacteria I. Enzymatic formation of 4-deoxy-L-threo-5-
hexoseulose uronic acid. J, Biol. Chem. 238, 1571-1576. Robertsen, B. (1987). Endo-polygalacturonase from Cladospor-
ium cucumerinum elicits lignification in cucumber cotyledons. Physiol. Mol. Plant Pathol. 31,361-374.
Selvendran, R. R., and O'Neill, M. A. (1987). Isolation and analysis of cell walls from plant material. In Methods in Bio- chemical Analysis, Vol. 32, D. Glick, ed (New York: Johri Wiley and Sons), pp. 25-153.
Somogyi, M. (1952). Notes on sugar determination, J, Biol. Chem. 195, 19-23.
Talmadge, K. W., Keegstra, K., Bauer, W. D., and Albersheim, P. (1973). The structure of plant cell walls I. The macromolecular components of the walls of suspension-cultured sycamore cells with a detailed analysis of the pectic polysaccharides. Plant Physiol. 51, 158-173.
Timberlake, W. E., and Hamer, J. E. (1986). Regulation of gene activity during conidiophore development in Aspergillus nidu- lans. In Genetic Engineering, Vol. 8, J. E. Setlow and A. Hol- laender, eds (New York: Plenum Publishing), pp. 1-29.
Timberlake, W. E., and Marshall, M. A. (1988). Genetic regulation of development in Aspergillus nidulans. Trends Genet. 4, 162- 169.
Weinhold, A. R., and Bowman, T. (1974). Repression of virulence of Rhizoctonia solani by glucose and 3-O-methyl glucose. Phy- topathology 64, 985-990.
Wijesundera, R. L. C., Bailey, J. A., and Byrde, R. J. W. (1984). Production of pectin lyase by Collectotrichum lindemuthianum in culture and in infected bean (Phaseolus vulgaris) tissue. J. Gen. Microbiol. 130, 285-290.

DOI 10.1105/tpc.1.3.265 1989;1;265-273Plant Cell
R A Dean and W E Timberlakepathogenesis of plants.
Production of cell wall-degrading enzymes by Aspergillus nidulans: a model system for fungal
This information is current as of November 18, 2018
Permissions 8X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw153229
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists








![Index [] · 2011-03-23 · Index 937 Ascochyta rabiei 507 Aspergillus nidulans 755 Aspergillus ochraceus 135, 158 Aspergillus terreus 506, 755 Aspidosperma alkaloids 106 – fragmentation](https://img.pdfslide.us/doc/110x75/5e2b9234e715b60f857d1b3b/index-2011-03-23-index-937-ascochyta-rabiei-507-aspergillus-nidulans-755-aspergillus.jpg)




















