Embed Size (px)
Citation preview

Reconstitution of Hormone-sensitive Adenylate Cyclase Activity with Resolved Components of the Enzyme*
(Received for publication, March 20, 1978)
Elliott M. Ross, Allyn C. Howlett,$ Kenneth M. Ferguson, and Alfred G. Gilmans
From the Department of Pharmacology, University of Virginia School of Medicine, Charlottesville, Vtrginia 22.908
Adenylate cyclase can be resolved into at least two proteins, a thermolabile, N-ethylmaleimide-sensitive component and a second protein (or proteins) that is more stable to either of these treatments. Neither com- ponent by itself catalyzes the formation of cyclic AMP using MgATP as substrate. However, mixture of the two reconstitutes MgATP-dependent fluoride- and guanyl-5’-yl imidodiphosphate (Gpp(NH)p)-stimulata- ble adenylate cyclase activity. The more stable com- ponent can be resolved from the first in various tissues or cultured cells by treatment of membranes or deter- gent extracts with heat or N-ethylmaleimide. The two proteins have also been resolved genetically in two clonal cell lines that are deficient in adenylate cyclase activity. An adenylate cyclase-deficient variant of the S49 lymphoma cell (AC-) contains only the thermola- bile activity, while the activity of the more stable pro- tein is found in a complementary hepatoma cell line (HC-1).
In addition, AC S49 cell plasma membranes contain MnATP-dependent adenylate cyclase activity. The pro- tein that catalyzes this reaction appears to be the same as that which can combine with the thermostable com- ponent to reconstitute Mg”+-dependent enzyme activity because both activities co-fractionate by gel exclusion chromatography and sucrose density gradient centrif- ugation, both activities have identical denaturation ki- netics at 30°C and both activities are stabilized at 30°C and labilized at 0°C by various nucleotides and divalent cations with similar specificity. It is thus hypothesized that the thermolabile factor is the catalytic subunit of the physiological adenylate cyclase and that the Mn”+- dependent activity is a nonphysiological expression of the catalytic protein.
The thermostable moiety of the enzyme, which is proposed to serve a regulatory function, appears to consist of two functional components, based upon dif- ferential thermal lability of its ability to reconstitute hormone-, NaF-, or Gpp(NH)p-stimulated adenylate cy- clase activity. These components have not, however, been physically separated.
The thermolabile and thermostable components can interact in detergent solution or in a suitable mem- brane. Mixing of the detergent-solubilized regulatory component with AC membranes that contain only the
* This work was supported by llnited States Public Health Service Grants NS10193 and AM17042 and American Cancer Societv Grant BC240. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “nd~~ertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of 1Jnited States Public Health Service Postdoctoral Fellowship NS05530.
9 Recipient of United States Public Health Service Research Ca- reer Development Award NSWO26.
catalytic protein and P-adrenergic receptors reconsti- tutes catecholamine-stimulatable adenylate cyclase ac- tivity; however, addition of the catalytic protein to membranes that contain receptor and the regulatory component yields MgATP-dependent enzymatic activ- ity that is unresponsive to hormones.
The multiple regulatory responses of hormone-sensitive adenylate cyclase activity have suggested that the enzyme system may have a complex composition, and a number of investigators have now begun to resolve its individual com- ponents. Hormone receptor proteins have been separated from the enzymatic portion of adenylate cyclase, and the hydrodynamic parameters of each have been reported (1, 2). Independent components that are required for the regulatory effects of guanine nucleotides are also under study. Pfeuffer and Helmreich have described a guanine nucleotide-binding protein that is probably involved in the control of adenylate cyclase activity (3,4), and Cassel and Selinger have presented evidence that a catecholamine-stimulated GTPase activity is involved in regulation of the enzyme by hormones, guanine nucleotides, and cholera toxin (5, 6).
We recently described the reconstitution of catecholamine- sensitive adenylate cyclase activity by the combination of a detergent extract of plasma membranes, which contains ade- nylate cyclase activity, with plasma membranes from a variant clone of S49 lymphoma cells which is deficient in adenylate cyclase activity but which has retained the P-adrenergic re- ceptor (AC)’ (7). We have also shown that the adenylate cyclase activity in the detergent extract can be resolved into at least two protein components, the catalytic protein and one or more components required for the display of basal or fluoride- or guanine nucleotide-stimulated enzymatic activity (8). One of these proteins is retained in AC cells. We sug- gested that this is in fact the catalytic protein of the adenylate cyclase system, and that the adenylate cyclase-deficient phe- notype of these cells reflects the loss of the other protein component(s) of the system.
In the initial communication cited above (7), several phe- nomena were noted but not explained, and many questions of molecular mechanisms were raised. Among these were the origin of increased NaF- and Gpp(NH)p-stimulated enzymatic activity when detergent-solubilized adenylate cyclase was added to membranes that lacked this catalytic activity, the
’ The abbreviations used are: Gpp(NH)p, guanyl-5’.yl imidodi- phosphate; Hepes, 4-(2.hydroxyethyl))l-piperazineethanesulfonate; AC, an S49 lymphoma variant clone phenotypically deficient in adenylate cyclase activity; UNC, an S49 lymphoma variant clone that has NaF- and Gpp(NH)p-stimulated adenylate cyclase and ,f&adre- nergic receptors but in which the effect of hormones on the enzyme has been lost; C and G/F, two resolved components of adenylate cyclase. The properties of these components are described in the text.
6401

6402 Reconstitution of Catecholamine-sensitive Adenylate Cyclase
role of the individual components of adenylate cyclase in the reconstitution of hormone-sensitive activity, and the mode of interaction of each component with the others and with the membrane. This report contains data that shed light on each of these areas. In addition, more extensive evidence is provided to show that the catalytic protein of adenylate cyclase is retained in AC cells, and it is suggested that its intrinsic activity is demonstrable in the presence of Mn2+ rather than Mg”+.
EXPERIMENTAL PROCEDURES
The phenotypes of all cell lines used in this study and the conditions used for their growth have been described previously (9-12). Plasma membranes were prepared from cultured cells according to the method of Ross et al. (ll), from turkey erythrocytes according to a modification of the procedure of Rosen and Rosen (13) suggested by Dr. Z. Selinger, Hebrew University, and from rat or rabbit liver according to the procedure of Neville (14) through Step 12. A cytosolic fraction from rat testis, containing a soluble adenylate cyclase activity, was prepared according to the method of Braun and Dods (15). Adenylate cyclase activity was solubilized from each membrane source using the nonionic detergent Lubrol 12A9 as previously de- scribed (7), except that GTP was occasionally omitted from the solubilization buffer as noted in the text. We refer to a 150,000 x g supernatant obtained by this procedure as a membrane extract, regardless of whether it contains assayable adenylate cyclase activity. Reconstitution of various enzyme activities was achieved by simple mixing of the components noted in the text. All reconstituted mixtures were held for at least 20 min at 0-4°C before assay.
Adenylate cyclase was assayed as described previously (ll), except that 10 mM MnC12 was sometimes substituted for 10 mM MgClz in the assay medium when noted in the text or legends. When soluble testis enzyme activity was assayed, 0.5 mM 1 -methyl-3-isobutylxanthine was used in addition to 0.1 mu R020-1724 (4-(3-butoxy+methoxyben- zyl)-2-imidazolidinone) to inhibit cyclic nucleotide phosphodiesterase activity. Assays were conducted at 30°C for 20 min when membrane- bound enzyme preparations were used and for 15 min with soluble preparations. Activity was linear with time for these periods. When activators were used their concentrations were: GTP or Gpp(NH)p, 0.1 mM; NaF, 10 mu; (-)-isoproterenol, 1 pM. The radioactive product of the enzyme was measured according to the method of Salomon et al. (16). Since reconstituted systems represent mixtures of different proteins, enzyme activities are generally expressed simply as pico- moles of cyclic AMP produced. min-’ ml-’ reaction volume, and the amounts of protein added to 0.1.ml reaction volumes are given in the legends. Ligand binding to the /3-adrenergic receptor was quantified with [‘““Ijiodohydroxybenzylpindolol as described previously (17). Guanylate cyclase was assayed for us by Dr. Kikuo Ichihara using the method of Mittal and Murad (18). Protein was measured according to the method of Lowry et al. (19).
Gel exclusion chromatography in the presence of detergent was performed on Ultrogel AcA34 essentially as described by Haga et al. (l), except that the column dimensions were 0.9 x 12 cm and the flow rate was 7.5 ml/h; the column was equilibrated and eluted with 0.1% Lubrol 12A9, 20 mu NaHepes, pH 8, 2 mM MgCla, 1 mM EDTA, 20 mM NaCl. Sucrose density gradient ultracentrifugation was performed exactly as described previously (1).
RESULTS
We recently reported the reconstitution of catecholamine- sensitive adenylate cyclase activity by the combination of a detergent extract of plasma membranes from B82 cells, which contain adenylate cyclase activity but no ,&adrenergic recep- tors, with membranes from a phenotypically adenylate cy- clase-deficient S49 lymphoma cell variant which, however, contain ,&adrenergic receptors (7). We subsequently showed that the adenylate cyclase in the extracts used in these exper- iments could be resolved into at least two protein components, neither of which was catalytically active when assayed in isolation (8). One component, which is quite thermolabile and sensitive to N-ethylmaleimide, is retained in membranes from the AC- cells (8). This finding demanded a re-evaluation of the mechanism by which the reconstitution of a hormone response takes place.
The thermolabile component of adenylate cyclase can be resolved from the thermostable component(s) that is present in detergent extracts by incubation at 30°C or 37°C. Fig. 1, A and B, shows the loss of total adenylate cyclase activity that results from incubation of such extracts of I382 and wild type S49 cells, respectively, at 30°C. In the latter case activity disappears rapidly with a half-time of approximately 4 min. With the B82 extract, the kinetics of denaturation does not appear to be fist order. However, if two fist order processes are assumed to be involved, the faster would also have a tli2 of approximately 4 min. If such incubated extracts are com- bined with membranes from AC S49 cells, which contain no demonstrable adenylate cyclase activity under these assay conditions, the reconstituted mixtures retain nearly all of the enzymatic activity that was present in the absence of incuba- tion of the extract (Fig. 2, A B). This is true even after the time when all detectable adenylate cyclase activity in the donor extract has been lost. Furthermore, the activity in the reconstituted mixtures is stimulated by multiple types of regulators of the system: NaF, Gpp(NH)p, or isoproterenol + GTP. The slight loss of activity of the reconstituted mixture
5 10 15 20 25 30
Time (min)
I I I 1 0 2 4 6 8 10
Time (min)
FIG. 1. Inactivation of adenylate cyclase at 30°C in Lubrol 12A9 extracts of plasma membranes from B82 cells (A) or from wild type S49 cells (B). Following incubation at 30°C for the indicated times, aliquots of extract were assayed in the presence of Gpp(NH)p (0) or NaF (0). Protein contents in 100 ~1 of assay reaction volume were 12 pg (A) and 15 pg (B).

Recowtitution of Catecholamine-sensitive Adenylate Cyclase 6403
b 5 IO 15 20 30
lime (min)
h ‘.
5 IO I5 20 25 30
lime (min)
FIG. 2. Reconstitution of hormone-sensitive adenylate cyclase us- ing donor extracts inactivated for varying times at 30°C as in Fig. 1 and AC membranes. Reconstitution is shown with B82 donor extract (A) and with wild type 349 donor extract (B). &constitutions were performed as follows: A, 40 ~1 of B82 donor extract (96 ).~g of protein) was added to 360 ~1 of AC membranes in Buffer A (20 mM NaHepes, 2 mM MgCL, 1 mM EDTA, pH 8.0) (360 ).~g of protein). Aliquots (40 ~1) were assayed (total volume 100 ~1) in the presence of GTP (O), isoproterenol (ZrVE) plus GTP (O), NaF (a), or Gpp(NH)p (V). R, 25 ~1 of wild type donor extract (20 pg of protein) was added to 340 ~1 of AC membranes (290 pg of protein). Aliquots were assayed as in A. Dotted lines in each panel represent the rates of decay of adenylate cyclase activity in the donor extracts that are shown in Fig. 1. The level of activity shown by the dotted line represents the adenylate cyclase activity contributed to the assay by the donor extract in this experiment.
that occurs as a function of time of incubation of the donor presumably represents denaturation of the thermostable com- ponent(s) of the enzyme.
Results are shown for both B82 and S49 cell donor extracts
to demonstrate the generality of this result for two different cell lines and to indicate that similar results are obtained with extracts from cells that do not (B82) and do (S49) contain ,& adrenergic receptors. The source of the receptor for the re- sponse to hormone is thus the AC membrane. B82 cells were the source of adenylate cyclase in the initial reconstitution experiments (7). However, extracts of wild type S49 cells appear to be entirely equivalent to those derived from B82
cells, and S49 cell membranes have thus been used as a source of components of adenylate cyclase because of their greater purity and because of previous characterization of the enzyme from this source (1, 11).
While the results of the experiments just described are consistent with the hypothesis that multiple components are necessary for adenylate cyclase activity, additional means of resolution of these components were sought to strengthen this argument. Accordingly, advantage was taken of the greater sensitivity of the thermolabile component of adenylate cyclase to N-ethylmaleimide (8). A detergent extract of wild type S49 cell membranes was treated with various concentrations of N- ethylmaleimide prior to reconstitution with AC- membranes. N-Ethylmaleimide at 1 to 10 mM completely inactivated ade- nylate cyclase activity in the detergent extract (Fig. 3); how- ever, the reconstituted mixture was again active and re- sponded not only to NaF and Gpp(NH)p but also to isopro- terenol. A thermostable, N-ethylmaleimide-resistant compo- nent (or components) is thus not only sufficient to restore adenylate cyclase activity to detergent extracts of AC mem- branes (8) but will also restore hormonal responsiveness when reconstituted with receptor-replete membranes from AC- cells.
Identity of the Thermolabile Component--We suggested that the thermolabile, N-ethylmaleimide-sensitive component of adenylate cyclase is in fact the catalytic subunit of the complex (8) and have referred to it as C for the sake of brevity. However, this proposal was based largely upon the require- ment for C for basal and both Gpp(NH)p- and NaF-stimulated adenylate cyclase activities, and further information on this possibility is thus necessary. Table I shows that while AC membranes are inactive when MgATP is used as substrate, these membranes and detergent extracts thereof do catalyze the formation of cyclic AMP when MnATP is the substrate. While this Mn’+-dependent adenylate cyclase activity is al- ways observed, the specific activity is quite variable, ranging, in different membrane preparations, from 10 to 50 pmol/min/mg of protein. To be noted are the observations
DOnOr
J
[N-ethylmaleimide] (mM)
FIG. 3. Reconstitution of hormone-sensitive adenylate cyclase us- ing AC membranes and N-ethylmaleimide-inactivated donor ext,ract. A detergent extract of wild type plasma membranes (1.5 mg/ml) was incubated for 20 min at 0°C with various concentrations of N-ethyl- maleimide. The reaction was terminated by the addition of an equi- molar amount of dithiothreitol. Extracts (40 ~1) were then added to 330 ALI of AC membranes in Buffer A (330 pg of protein) or to 330 ~1 of Buffer A. After 20 min at 0°C the reconstituted mixtures (40 ~1) were assayed (total volume = 100 ~1) in the presence of GTP (not shown), GTP plus isoproterenol (ZNE) (O), NaF (a), or Gpp(NH)p (7). Activities in the presence of GTP were approximately 1 pmol/min/ml. Adenylate cyclase in the diluted donor extracts was assayed in the presence of NaF (A) or Gpp(NH)p (V) (- - -).
6404 Reconstitution of Catecholamine-sensitive Adenylate Cyclase
TABLE I
Diualent cation specificity for adenylate cyclase activity
Me” -de- Mn’+-de-
Enzyme source Activator pen&?Pt en- pendent en- zyme activ- zyme activ-
itv itv”
Wild type mem- branes Ir
Heated wild type membranes’, ’
AC membranes”
AC extract’
GTP’ GTP + INE” NaF GPPU’WP NaF GppU’Wp GTP GTP + INE NaF GPPWWP GTP’ NaF GPPWWP
pmol min
44 235 253 119
2 1 1 1 2
3 2 2
mg-l
22 32
255 51
4 0
15 14 17 12 35 34 40
” Mg”+-dependent enzyme activity is defined as that detected in the presence of 10 mu MgCl, in the assay, while Mn’+-dependent activity is assayed in the presence of 10 rnM MnCl?.
’ All membranes were pelleted and resuspended in 20 IIIM NaHepes, pH 8, before assay.
’ GTP has a slight stimulatory effect on enzymatic activity (t2- fold) with wild type membrane preparations and no effect with AC membranes or extracts.
” INE, isoproterenol. ” Membranes incubated at 37°C for 30 min prior to assay. ’ The AC extract contributed a final concentration of 20 FM Mg”’
to the assay.
that the wild type, membrane-bound enzyme is still subject to regulation by NaF, Gpp(NH)p, or isoproterenol when MnATP is the substrate, although responses to guanine nucleotides and, particularly, the ,8-adrenergic agonist are much reduced. However, activity in AC membranes is not influenced by these ligands. This is consistent either with the loss of regu- latory components of the enzyme in AC membranes or with the possibility that the MnATP-dependent catalysis of cyclic AMP synthesis by AC membranes is performed by a protein that is not a normal constituent of the hormone-sensitive adenylate cyclase system.
If C is truly the inactive catalytic subunit of adenylate cyclase, then the Mn’+-dependent synthesis of cyclic AMP could be a nonphysiological assay of its inherent activity. Several types of experiments support the identity of the MnZ+- dependent activity and C. MnATP or Mn-App(NH)p (but not Mn’+ alone) stabilizes C activity at 3O”C, indicating that C has a nucleotide binding site. As shown in Fig. 4, the denatur- ation kinetic data of C, as assayed by reconstitution with the thermostable components from heated wild type S49 deter- gent extracts, and of the Mn’+-dependent adenylate cyclase activity are similar in the presence or absence of MnATP. Identical results are obtained if Mn-App(NH)p is used, but Mg”+ plus ATP or App(NH)p (0.5 111~) are ineffective. The dependence of this stabilization on the concentrations of nu- cleotide and Mn’+ has not been determined precisely, but 5 mM Mn’+ plus 0.5 mu ATP or App(NH)p produce a maximal stabilizing effect. The nucleotide specificity of this stabiliza- tion process is also similar when C (reconstituted activity)or Mn’+-dependent adenylate cyclase activity is measured. Either ADP or App(NH)p at 0.5 mM plus 5 to 10 IIIM MnC12 stabilize these activities, while adenosine, AMP, or cyclic AMP is ineffective at 0.5 or 2 mM. Guanylate cyclase activity in extracts of AC membranes, assayed with either Mn’+ or Mg”+, is not measurably decreased by heating for 10 min at 30°C.
In contrast to these observations, both C and Mn’+-depend- ent adenylate cyclase activity are markedly labilized by higher
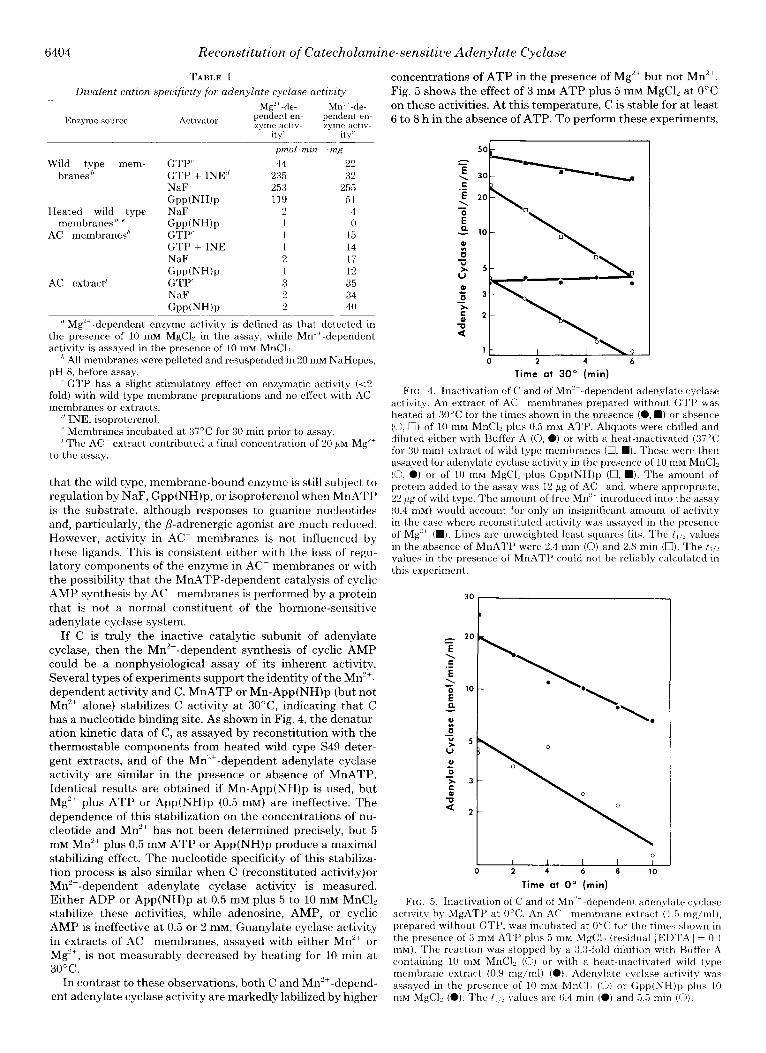
concentrations of ATP in the presence of Mg’+ but not Mn’+. Fig. 5 shows the effect of 3 IIIM ATP plus 5 mM MgC12 at 0°C on these activities. At this temperature, C is stable for at least 6 to 8 h in the absence of ATP. To perform these experiments,
0 2 4 6
Time at 30’ (min)
FIG. 4. Inactivation of C and of Mn’+-dependent adenylate cyciase activitv. An extract of AC membranes prepared without GTI’ was heated at 30°C for the times shown in the presence (0, n ) or absence (0, 0) of 10 mM MnCl, plus 0.5 mM ATP. Aliquots were chilled and diluted either with Buffer A (0, a) or with a heat-inactivated (37°C for 30 min) extract of wild type membranes (0, n ). These were then assayed for adenylate cyclase activity in the presence of 10 mM MnCli (0, 0) or of 10 mrvi M&l? plus Gpp(NH)p (0, n ). The amount of protein added to the assay was 12 Kg of AC and, where appropriate, 22 ;~g of wild type. The amount of free Mn” introduced into the assay (0.4 mM) would account for only an insignificant amount of activity in the case where reconstituted activity was assayed in the presence of Mg’+ (M). Lines are unweighted least squares fits. The t, d values in the absence of MnATP were 2.4 min (0) and 2.8 min (U). The t,,, values in the presence of MnATI’ could not be reliably calculated in this experiment.
30- t
1 0
I I I I I 0 2 4 6 8 IO
Time at 0” (min)
FIN;. 5. Inactivation of C and of Mn” dependent adenylate cyclase activity by MgATP at 0°C. An AC membrane extract (1.5 mg/ml), prepared without GTP, was incubated at 0°C for the times shown in the presence of 3 mM A’I’P plus 5 mM M&l, (residual [EIX’A] = 0.1 mM). The reaction was stopped by a 3.3.fold dilution with Buffer A containing 10 IllM MnCl? (0) or with a heat-inactivated wild type membrane extract (0.9 mg/ml) (0). Adenvlate cyclase activity was assqvved in the presence of 10 nlM MnCli (0) or Gpp(NH)p plus 10 mM MgCl, (0). The t , ; . ! values are 6.4 min (0) and 6.5 min (0).

Reconstitution of Catecholamine-sensitive Adenylate Cyclase
the inactivation reaction was stopped by dilution with buffer containing either MnCb (when M&dependent activity was to be measured) or a heated (37°C for 20 min) extract of wild type membranes (when C activity was to be measured by reconstitution), The nucleotide specificity of this inactivation process apparently differs from that for the Mn”+-dependent stabilization at 30°C in that ATP was the only nucleotide tested (at 3 111~) that caused this effect.
The identity of C and the Mn’+-dependent adenylate cy- clase activity is further supported by their identical patterns of fractionation by gel exclusion chromatography and sucrose density gradient ultracentrifugation. As shown in Figs. 6 and 7, both Mn’+-dependent adenylate cyclase activity and recon- stitutive C activity display virtually identical hydrodynamic properties, which are summarized in Table II. The small calculated differences in molecular weight and the amount of detergent bound result primarily from the different value for the partial specific volume of each activity. These differences, however, are not statistically significant. The variability in these values is probably in part due to the lability of the activities and the low specific activity of the Mn’+-dependent enzyme. The hydrodynamic properties of each activity are unchanged by the addition of MnC12 and ATP to the medium, and identical values are obtained if reconstitutive C activity is measured in the presence of NaF or Gpp(NH)p. Only purification will supply proof that the Mn”‘-dependent ade- nylate cyclase activity is that of the isolated catalytic subunit of the physiological Mg”+-dependent enzyme. However, we feel that these experiments argue strongly for that premise.
-c 0 C 6
‘ii 4
10 20 30 40 50
Fraction Number
Fro. 6. &I exclusion chromatography of C and Mn”-dependent adenylate cyclase. An extract of AC membranes was prepared and analyzed on IJltrogel AcA34 as described under “Experimental Pro- cedures.” Each column fraction was 175 al. Aliquots (60 ~1) were assayed for adenylate c,vclase activity either in the presence of 10 rnM MnCb (0) or, after reconstitution with a heat-inactivated wild type membrane extract, in the presence of IO mM MgCli and Gpp(NH)p (the example shown, 0) or NaF. Arrou~.s denote the elution volumes for blue dextran (Icft) and 2,4-dinitrophenylglycine (r&U). The upper panel is a calibration curve used for determination of the Stokes radius using the standards ,Kgalactosidase (6.84 nm), catalase (5.21 nm), lactate dehydrogenase (4.75 nm), and malate dehydrogenase (3.69 nm) (1).
L ,o v)
12 -
10 -
0 -
6
4 -
2 -
1
6405
25
Fraction Number
FIG. 7. Sucrose density gradient ultracentrifugation of C and Mn’+-dependent adenylate cyclase in an extract of AC plasma mem- branes. Centrifugation of a Lubrol 12A9 extract was carried out in medium prepared in Hz0 as discussed under “Experimental Proce- dures.” Fractions were assayed as described in legend to Fig. 6. The upper panel represents a standard curve used for obtaining the sedimentation coefficient of each activity with reference to the markers catalase (11.3 S), lactate dehydrogenase (7.3 S), malate dehydrogenase (4.32 S), and cytochrome c (1.71 S) (1). The position of Gpp(NH)p-treated wild type adenylate cyclase determined in these studies is shown for reference. The enzyme was exposed to Gpp(NH)p prior to solubilization. The value obtained is close to that determined previously (1). However, there was an error in calculation in the previous report and the s~,,.,~ value for the enzyme should have been 7.8 S rather than 7.5 S.
The question also arises as to the relationship between C, the Mn’+-dependent enzymatic activity of S49 AC cells, and a testicular adenylate cyclase that is soluble, not subject to regulation by NaF or Gpp(NH)p, and dependent on Mn2+ for activity (15). The data of Table III indicate that the ther- mostable component(s) of S49 cell adenylate cyclase does not appear to interact with the testicular enzyme as it does with C.” Furthermore the Stokes radius of the testicular enzyme under the conditions utilized in this study was found to be 3.4 nm, in agreement with the data of Neer (20). There is thus no apparent relationship between C and the soluble testicular enzyme.
Treatment of C with N-ethylmaleimide does allow differ- entiation between C (reconstitutive) activity and the Mn’+- dependent enzyme. Activity measured by reconstitution with the thermostable component(s) of a heat-inactivated, wild type extract is considerably more sensitive to the sulthydryl reagent than is the Mn’+-dependent enzymatic activity (Fig. 8). This observation will be discussed below.
Role of Thermostable Component(s) of Hormone-sensitive Adenylate Cyclase-Our previous analysis of the rate of ther- mal inactivation of the more heat-stable component(s) of
‘Extracts of HC-1 cell plasma membranes, which contain the thermostable component but not C (see below), also fail to influence the activity of soluble testicular adenylate cyclase.

6406 Reconstitution of Cntecholamine-sensitive Adenylate Cyclase
TABLE II
Molecular purameters for Mn” -clependent adenylntr cyclase and C in AC extract
Data were obtained as described under “Experimental Procedures” and in the legends to Figs. ii and 7. C was assayed by combination with a heated wild type extract in the presence of MgATP and either NaF or Gpp(NH)p (one activator only in each experiment). Since identical behavior was observed with either activator in the assay, these data have been combined for the purpose of calculation.
Parameter”
Stokes radius, a (nm) Partial specific volume, U
(ml/g) h Sedimentation coefficient
S2u.u (SY Molecular weight, M, Frictional ratio, f/f;, Lubroll2A9 bound
mg/mg protein mol/mol protein
Molecular weight of protein
7.0 I+_ 0.11 (6) 7.1 i 0.10 (7) 0.73 t 0.025 (6 0.76 i_ 0.013 (8
x 6) x 6)
6.6 I+ 0.35 (6 x 6.5 +I 0.23 (8 x
6) 6) 20 x IO’ 22 x IO’
1.8 1.8
0 0
20 x 10” (18-20 x 10‘7
0.13 40 19 x 10: (17-22
x 104)
IL The values given are the mean -+ S.D. for the number of deter- minations shown in parentheses.
’ Calculations are identical with those of Haga et al. (I) using data derived from six sucrose density gradient centrifugations in I>,0 and eight or six in Hz0 as denoted.
’ The calculations were made using the value of 0.735 ml/g as the partial specific volume observed for a typical soluble protein. The partial specific volume of Lubrol assumed for these determinations was 0.958 ml/g and its average molecular weight was assumed to be 635.
TARL~ III
Failure of C/F to allow MgATP utilization by testicular denylate cyclase
Activator
Mg”-de- pendent en- zyme activ-
itv
Mn’ ’ -de- pendent en- zyme activ-
it,,
pm01 min ’ mg-’
Testis” None <I 18 NaF <I 17
GPP(NH)P <I 17 Heated wild type ex- None <I <1
tract” NaF <I <l Gpp(NH)p <l <I
Testis plus heated None <l 15 wild type extract NaF <l 15
Gpp(NH)p <l 15
” Soluble fraction prepared according to Ref. 15. Each assay tube (100 ~1) contained 95 pg of protein.
Ir Lubrol 12A9 extract incubated for 20 min at 37°C. Each assay tube contained 44 pg of protein.
’ Mixture of 95 pg of testis protein plus 44 pg of S49 ceil protein. Specific activities are expressed per mg of testis protein.
adenylate cyclase remains the only means by which two functional entities have been distinguished (8). I f detergent extracts in which C has been inactivated (37°C incubation) are further incubated at 5O”C, enzymatic activity revealed by subsequent reconstitution with C (AC extracts) declines with
a t1/2 of 2 min if Gpp(NH)p-stimulated catalysis is measured or with a tllz of 5 to 8 min if NaF is the activator. It was suggested that the more stable component, referred to as F, can combine with C to yield basal and NaF-stimulated enzyme activity with MgATP as substrate. The second component, referred to as G, is hypothetically required for activation of the enzyme by Gpp(NH)p but is not essential for Mg”-de- pendent enzymatic activity (8). It must be emphasized that G and F are utilized only as designations of functional entities; their physical resolution has not been achieved and we make
0 4/ I I 1
0 Cl.03 0.1 0.3 1.0
[N-Ethylmaleimida] (mM)
FIG. 8. Inactivation of Mn”-dependent adenylate cyclase and C by N-ethylmaleimide. Aliquots of an AC plasma membrane extract (1.7 mg/ml; prepared without GTP) were incubated for 20 rnin at 0°C in the presence of N-ethylmaleimide at the concentrations shown. The reaction was terminated by the addition of equimolar dithiothre- itol. A portion of each aliquot was diluted with 2.3 volumes of 20 rnM NaHepes, pH 8, and a 40.~1 volume was assayed for adenylate cyclase activity in the presence of 10 mM MnCl, (0). A second portion was mixed with 1.0 volume of Buffer A and 1.3 volumes of a heated (37”C, 25 min) wild type extract (2.4 mg/ml). A 40.~1 sample of this recon- stituted mixture was assayed in the presence of 10 mM MgCl, and Gpp(NH)p (U). Original aliquots of extract to which dithiothreitol was added before N-ethylmaleimide displayed the same activities as did the aliquot that was exposed to neither reagent.
no claim that multiple proteins are involved. (We will there- fore usually refer to the activity as G/F.) Furthermore, while G/F appears to have a guanine nucleotide binding site, based on the ability of Gpp(NH)p and GTP to stabilize G/F activity during thermal denaturation, the designation F is not meant to imply that fluoride interacts directly with this protein.”
The experiment shown in Fig. 9 was designed to assess the necessity for G activity for regulation of reconstituted ade- nylate cyclase by hormone. A detergent extract of wild type membranes was incubated at 37°C to inactivate C and was then heated for various times at 50°C prior to reconstitution with AC- membranes. The ability of the extract to restore hormone- or Gpp(NH)p-stimulated activity to the membranes decays with the same t1,2, which is significantly shorter than that for the reconstitution of NaF-stimulated catalysis. The experiment suggests that G is required as an essential coupling factor between the receptor and C.
Alternative Sources and Preparations of G/F--Data pre-
sented above demonstrate that G/F can be resolved from C in solubilized preparations of adenylate cyclase from two cul- tured cell lines, either by heat or by treatment with N-ethyl- maleimide. If G/F is in fact a ubiquitous component of ade- nylate cyclase, it should also be assayable in other tissues. Furthermore, since the absence of either G/F or C should result in a cell without apparent adenylate cyclase activity, it seemed possible that other AC variants of the S49 cell might be deficient in C rather than G/F.
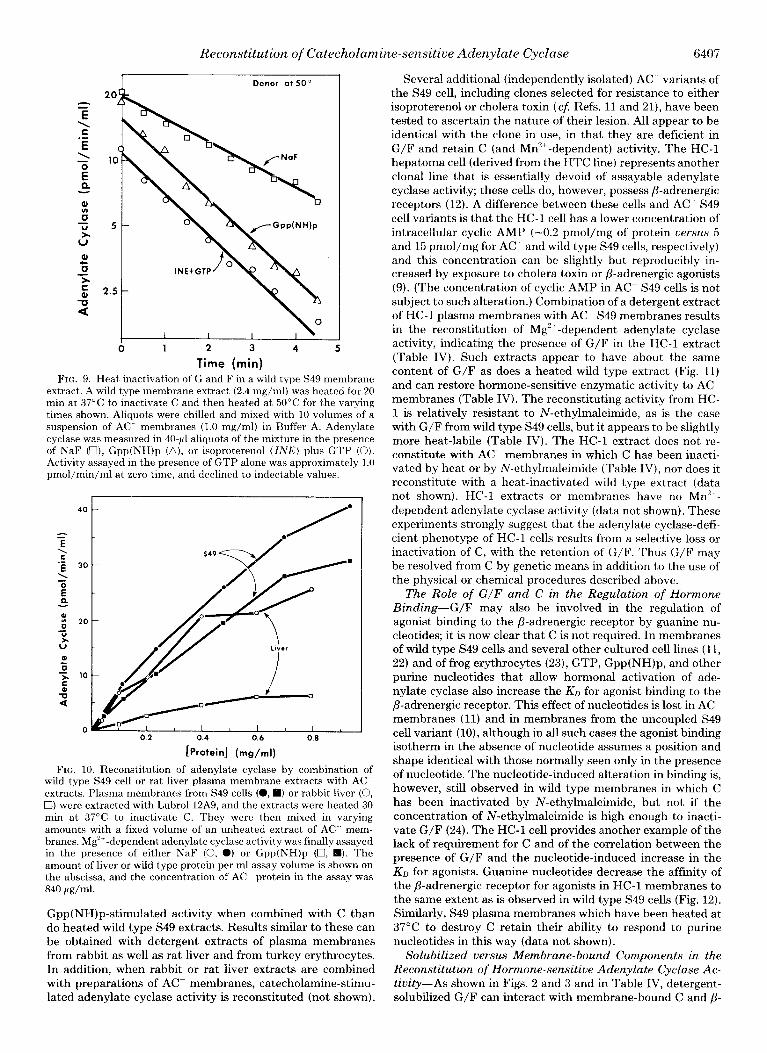
When a Lubrol 12A9 extract of rat liver plasma membranes is heated at 37°C for 30 min, it loses all assayable adenylate cyclase activity. As shown in Fig. 10, this extract can combine with an extract of AC membranes to yield both Gpp(NH)p- and NaF-stimulated adenylate cyclase activity, indicating the presence of G/F. Titration of the heated liver preparation and of a heat-inactivated extract of wild type S49 membranes suggests that they are about equally capable of restoring fluoride-stimulated Mg2+-dependent activity to the AC ex- tract. Heated !iver extracts reproducibly produce a lower
’ An effect of NaF on a component of the heated Lubrol extract has been detected. The result is to facilitate the binding of G/F to .4C membranes; this reaction also requires Mg” and a nucleotide (ADP or ATP). A. C. Howlett. I’. C. Sternweis, B. A. Macik, P. M. Van Arsdale, and A. G. Gilman, manuscript in preparation.
Reconstitution of Catecholamine-sensitive Adenylate Cyclase 6407
Donor at 50”
0 1 2 3 4 5
Time (min)
Several additional (independently isolated) AC-- variants of the S49 cell, including clones selected for resistance to either isoproterenol or cholera toxin (CL Refs. 11 and 21), have been tested to ascertain the nature of their lesion. All appear to be identical with the clone in use, in that they are deficient in G/F and retain C (and Mn”+-dependent) activity. The HC-1 hepatoma cell (derived from the HTC line) represents another clonal line that is essentially devoid of assayable adenylate cyclase activity; these cells do, however, possess P-adrenergic receptors (12). A difference between these cells and AC- S49 cell variants is that the HC-1 cell has a lower concentration of intracellular cyclic AMP (-0.2 pmol/mg of protein versus 5 and 15 pmol/mg for AC- and wild type S49 cells, respectively) and this concentration can be slightly but reproducibly in- creased by exposure to cholera toxin or /3-adrenergic agonists (9). (The concentration of cyclic AMP in AC S49 cells is not subject to such alteration.) Combination of a detergent extract of HC-1 plasma membranes with AC- S49 membranes results in the reconstitution of M$+-dependent adenylate cyclase activity, indicating the presence of G/F in the HC-1 extract (Table IV). Such extracts appear to have about the same content of G/F as does a heated wild type extract (Fig. 11) and can restore hormone-sensitive enzymatic activity to AC- membranes (Table IV). The reconstituting activity from HC- 1 is relatively resistant to N-ethylmaleimide, as is the case with G/F from wild type S49 cells, but it appears to be slightly more heat-labile (Table IV). The HC-1 extract does not re- constitute with AC- membranes in which C has been inacti- vated by heat or by N-ethylmaleimide (Table IV), nor does it reconstitute with a heat-inactivated wild type extract (data not shown). HC-1 extracts or membranes have no Mn’+-
dependent adenylate cyclase activity (data not shown). These experiments strongly suggest that the adenylate cyclase-defi- cient phenotype of HC-1 cells results from a selective loss or inactivation of C, with the retention of G/F. Thus G/F may be resolved from C by genetic means in addition to the use of the physical or chemical procedures described above.
FIG. 9. Heat inactivation of G and F in a wild type S49 membrane extract. A wild type membrane extract (2.4 mg/ml) was heated for 20 min at 37°C to inactivate C and then heated at 50°C for the varying times shown. Aliquots were chilled and mixed with 10 volumes of a suspension of AC membranes (1.0 mg/ml) in Buffer A. Adenylate cyclase was measured in 40.~1 aliquots of the mixture in the presence of NaF (U), Gpp(NH)p (a), or isoproterenol (INK) plus GTP (0). Activity assayed in the presence of GTP alone was approximately 1.0 pmol/min/ml at zero time, and declined to indectable values.
0.2 0.4 0.6 0.8
[Protein] (mg/ml)
FIG. 10. Reconstitution of adenylate cyclase by combination of wild type S49 cell or rat liver plasma membrane extracts with AC extracts. Plasma membranes from S49 cells (a, 0) or rabbit liver (0, 0) were extracted with Lubrol 12A9, and the extracts were heated 30 min at 37°C to inactivate C. They were then mixed in varying amounts with a fixed volume of an unheated extract of AC mem- branes. Mg’+-dependent adenylate cyclase activity was finally assayed in the presence of either NaF (0, 0) or Gpp(NH)p (U, q ). The amount of liver or wild type protein per ml assay volume is shown on the abscissa, and the concentration of AC- protein in the assay was 840 pg/ml.
Gpp(NH)p-stimulated activity when combined with C than do heated wild type S49 extracts. Results similar to these can be obtained with detergent extracts of plasma membranes from rabbit as well as rat liver and from turkey erythrocytes. In addition, when rabbit or rat liver extracts are combined with preparations of AC- membranes, catecholamine-stimu- lated adenylate cyclase activity is reconstituted (not shown).
The Role of G/F and C in the Regulation of Hormone Binding-G/F may also be involved in the regulation of agonist binding to the /3-adrenergic receptor by guanine nu- cleotides; it is now clear that C is not required. In membranes of wild type S49 cells and several other cultured cell lines (11, 22) and of frog erythrocytes (23), GTP, Gpp(NH)p, and other purine nucleotides that allow hormonal activation of ade- nylate cyclase also increase the Kn for agonist binding to the /3-adrenergic receptor. This effect of nucleotides is lost in AC membranes (11) and in membranes from the uncoupled S49 cell variant (lo), although in all such cases the agonist binding isotherm in the absence of nucleotide assumes a position and shape identical with those normally seen only in the presence of nucleotide. The nucleotide-induced alteration in binding is, however, still observed in wild type membranes in which C has been inactivated by N-ethylmaleimide, but not if the concentration of N-ethylmaleimide is high enough to inacti- vate G/F (24). The HC-1 cell provides another example of the lack of requirement for C and of the correlation between the presence of G/F and the nucleotide-induced increase in the KI, for agonists. Guanine nucleotides decrease the affinity of the P-adrenergic receptor for agonists in HC-1 membranes to the same extent as is observed in wild type S49 cells (Fig. 12). Similarly, S49 plasma membranes which have been heated at 37°C to destroy C retain their ability to respond to purine nucleotides in this way (data not shown).
Solubilized versus Membrane-bound Components in the Reconstitution of Hormone-sen‘sitive Adenylate Cyclase Ac- tivity-As shown in Figs. 2 and 3 and in Table IV, detergent- solubilized G/F can interact with membrane-bound C and /?-

6408 Reconstitution of Catecholamine-sensitive Adenylate Cyclase
TABLE IV
Reconstitution ofAC S49 cell membranes with HC-I extracts
AC membranes (1 .O mg/ml) were mixed with 0.1 volume of HC-1 eimide (MalNEt) treatment was for 20 min at 0°C and was stopped extracts (0.64 mg/ml) and the mixtures were held at 0°C for 20 min before assay in the presence of Mg’+. The treatments of the individual
by addition of equimolar dithiothreitol 10 min before mixing of membranes and extracts. Activity is expressed as picomoles of CAMP
extracts or membranes were performed before mixing. N-Ethylmal- produced/min/ml reaction volume.
Donor extract: HC-1 HC-1 HC-1 HC-1 HC-1 HC-1
37"; 15 min 50°C; 15 min HC-1 1mM 10 mhl HC-1 MalNEt MalNEt
Acceptor membrane: NOIE AC AC AC AC AC AC AC- 37"C;30 min 1 mu MalNEt
Activator GTP GTP + isoproterenol NaF Gpp(NH)p
ND” 2 1 0 0 2 1 1 ND” gh 3 0 0 6 3 1
0 16 4 0 0 10 3 2 0 26 7 1 1 19 6 3
n ND, not determined. ’ Activity of this sample in the presence of GTP plus isoproterenol plus 1 pM (-)-propranolol was 2 pmol/min/ml.
0 0.2 0.4
[Protein] (mg/ml)
0.6
FIG. Il. Reconstitution of Mg”-dependent adenylate cyclase from extracts of AC and HC-I membranes. Plasma membrane extracts of HC-1 cells and S49 AC cells were mixed in varying proportions and assayed for adenylate cyclase in the presence of 10 mM MgCl, (0, W). For comparison, a heated wild type S49 extract was mixed with the same amount of ACT extract (0, 0). The amount of HC-I or of heated wild type protein per ml of assay volume is shown on the abscissa. The concentration of AC protein in the assay was I20 Fg/ml. Activators were Gpp(NH)p (0,O) or NaF (0, W).
adrenergic receptors to reconstitute hormone-sensitive, Mg”‘- dependent enzyme activity. However, not all interactions within the system are so amenable to manipulation, as typified by our persistent inability to reconstitute catecholamine-re- sponsive adenylate cyclase activity using preparations of de- tergent-solubilized P-adrenergic receptor and membrane-
bound enzyme.* It was therefore of interest to determine whether C, once
solubilized, could interact with membrane-bound G/F and receptor to yield a hormone-responsive enzyme or if soluble C and G/F could interact with receptor bound to a membrane that was deficient in both of these activities. The experiment
shown in Fig. 13 was undertaken to attempt to answer the second of these questions. An untreated wild type membrane extract was added to suspensions of AC membranes that had been heated at 37°C for various times in order to inactivate endogenous C. (C is more stable in membranes than in deter- gent solution: tl12 s 5 min at 37°C.) Fluoride- and Gpp(NH)p- stimulated enzyme activity are retained in the reconstituted mixture, even after extensive heating of the recipient mem- brane. At long times, this activity is equal to the activity added as the wild type extract (compare 30-min time points
’ T. Haga and A. G. Gilman, unpublished observations.
0 b, I I I I I 0 10-P 10-1 10-7 10-b 10-S
[Isoproterenol] (M)
FIG. 12. Effect of GTP on the binding of isoproterenol to HC-1 hepatoma cell membranes. Specific binding of isoproterenol to p- adrenergic receptors in HC-1 membranes was assayed by its ability to compete for binding sites with [““IJiodohydroxybenzylpindolol ([““I]IHYP) according to the method of Maguire et al. (17) in the presence (0) or absence (0) of 50 pM GTP.
with solid bars at right). This experiment demonstrates the increment in Gpp(NH)p- and NaF-stimulated activity that is observed when an active wild type extract is added to AC- membranes, as compared with its simple dilution into buffer (compare zero time values with solid bars at right of figure). This effect was noted, but not explained, previously (7). We are now able to hypothesize that this increment is due to the increased amount of C that is added in the AC- membranes. Thus, heating the AC- acceptor membranes in this experi- ment abolishes this incremental activity (closed triangles), and the increment decays with tllz that is typical of mem- brane-bound C. It is striking, furthermore, that the total isoproterenol-stimulated activity in this experiment decreases with this same tIj2, suggesting that only the C already in the acceptor membrane and not that added in detergent solution can interact with G/F and the P-adrenergic receptor in such a way as to produce hormone-stimulated activity. This exper- iment can also be performed by using N-ethylmaleimide, rather than heat, to inactivate C in the AC acceptor mem-
branes (Fig. 14). Hormone-stimulated activity and the incre- ments in fluoride- and Gpp(NH)p-stimulated activities are lost with the same dependence on N-ethylmaleimide concen- tration, again arguing that C added in detergent solution is not reincorporated into the membrane or is in an orientation such that it cannot interact functionally with receptor, or G/F, or both.

Reconstitution of Catecholamine-sensitive Adenylate Cyclase 6409
0.5 ! .
L 1 I I 1 .
0 5 10 15 20 25 30
Time (min)
FIG. 13. Reconstitution of hormone-sensitive adenylate cyclase by combination of wild type membrane extract with AC membranes incubated for various times at 37°C. AC membranes suspended in Buffer A at a concentration of 0.8 mg/ml were incubated at 37°C for the times shown on the abscissa. Following this incubation, the membranes (320 ~1) were mixed with untreated wild type extract (40 ~1,2.3 mg/ml). After 20 min at O”C, aliquots (40 ~1) of the reconstituted mixtures were assayed (total volume = 100 ~1) in the presence of GTP (O), isoproterenol (INE) plus GTP (II), NaF (A), or Gpp(NH)p (V). An aliquot (40 ~1) of the untreated wild type extract was also diluted with 320 ~1 of Buffer A and was assayed in the presence of NaF or Gpp(NH)p (solid bars to right). Solid triangles depict the difference in activity between each reconstituted mixture and the activity at- tributable to the donor extract alone (open triangle minus solid bar).
The above data show that detergent-solubilized C does not reconstitute hormone-stimulated adenylate cyclase activity when added along with solubilized G/F to a membrane con- taining hormone receptors. As shown in Table V, the addition of detergent-solubilized C to HC-1 membranes, which already contain G/F and the /3-adrenergic receptor, does result in the reconstitution of some NaF- and Gpp(NH)p-stimulated en- zymatic activity. However, the specific activity is considerably less than when membrane-bound C and soluble G/F are used, and the combination of soluble C and membrane-bound G/F does not result in the restoration of hormone-stimulated cat- alytic activity. Similar results have been obtained using heated or N-ethylmaleimide-treated wild type membranes as sources of membrane-bound G/F and P-adrenergic receptor (data not shown). While detergent-solubilized C (from S49 AC) can thus interact with membrane-bound G/F to yield Mg”-de- pendent activity, it apparently is unable to do so in a manner that allows regulation by the hormone receptor complex.
DISCUSSION
In a previous publication (8) we presented data that suggest that adenylate cyclase is composed of at least two protein components in addition to hormone receptors. One, a ther- molabile protein, is retained in a variant S49 lymphoma clone that is phenotypically deficient in Mg’+-dependent adenylate cyclase both in vivo and in vitro. A more heat-stable compo- nent (or components) of the system was identified in wild type S49 cells by showing that a heat-inactivated detergent extract of wild type membranes could combine with an AC- detergent
[N-ethylmaleimidcj (uM)
FIG. 14. Reconstitution of hormone-sensitive adenylate cyclase by combination of wild type membrane extract with N-ethylmaleimide- treated AC membranes. AC membranes suspended in Buffer A (0.83 mg/ml) were incubated at 0°C for 20 min with the indicated concentrations of N-ethylmaleimide. The reaction was terminated by the addition of an equimolar amount of dithiothreitol. The mem- branes (360 ~1) were then mixed with untreated wild type extract (40 ~1; 1.1 mg/ml). After 20 min at O”C, aliquots (40 ~1) of the reconstituted mixtures were assayed (total volume = 100 ~1) in the presence of GTP (0), isoproterenol (INE) plus GTP (C!), NaF (a), or Gpp(NH)p (V). An aliquot (40 ~1) of the untreated wild type extract was also diluted with 360 ~1 of Buffer A and assayed in the same way in the presence of NaF or Gpp(NH)p (solid bars to right).
TABLE V
Reconstitution of catecholamine responses using HC-1 and AC membranes and extracts
Extract (0.04 ml) (1.7 mg/ml HC-1, 1.5 mg/ml AC) was mixed with 0.4-ml membranes (1 mg/ml), and 0.04.ml aliquots of the mixture were assayed for Mg’+-dependent adenylate cyclase activity in the presence of the activators listed above.
Adenvlate cvclase activitv
Extract Mem- brane Basal
Isopro- tereno1
plus GTP
NaF GPPWWP
pndmm- ml-’
None” HC-1 to.5 <0.5 to.5 0.5 AC HC-1 0.6 0.5 2.5 4.6 Heated AC?’ HC-1 co.5 to.5 0.6 1.1 HC-1 AC 1.6 10 16.7 20.4
n Solubilization buffer (0.04 ml) (7) added to membranes. * Extract heated 30°C for 15 min before reconstitution.
extract to reconstitute Mgzf-dependent enzymatic activity. We argued that the thermolabile component that is found in AC membranes is identical with that which is lost upon heating the wild type extract. This contention is consistent with the identical denaturation kinetics of soluble S49 cell adenylate cyclase (Fig. 1B) and the thermolabile component (Fig. 3 in Ref. 8). In addition, the inactivation of the solubilized thermolabile component by N-ethyhnaleimide has the same concentration dependence as does inactivation of soluble wild type adenylate cyclase activity (compare Fig. 3 with Table I in Ref. 8). The alternative explanation, that something in the AC- extract is reactivating a damaged adenylate cyclase in the heated or N-ethylmaleimide-treated wild type extract, seems to us very unlikely because of the widely differing

6410 Reconstitution of Catecholamine-sensitive Adenylate Cyclase
treatments used to inactivate the wild type membranes. This explanation is also not supported by the presence of the thermostable component and absence of the activity of the labile protein in untreated membranes of the HC-I hepatoma cell.
It is not certain from these data whether the heat-stable and heat-labile components in fact form an active complex or whether the adenylate cyclase catalyst requires the tonic activity of a second protein to maintain it in an active state. While it is clear that one component must have the necessary catalytic capability, it is not known whether the other com- ponent is a true regulatory “subunit” of the enzyme or whether it might exert its role by, for example, catalyzing the covalent modification of the catalytic protein. The mechanism by which the catalyst is activated is of course profoundly impor- tant for understanding the regulation of adenylate cyclase activity, but our current ignorance of this mechanism does not call into question the essential role of the regulatory compo- nent.
We initially suggested that the thermolabile component of adenylate cyclase, which is found in membranes of the AC variant, was in fact the catalytic moiety of the enzyme and that the heat-stable factors fulfilled the activating regulatory function discussed above (8). We have referred to the heat- labile component, therefore, as C. This report provides evi- dence to support this contention. First, the data in Table I demonstrate that plasma membranes of AC cells or detergent extracts therefrom contain a Mnzf-dependent adenylate cy- clase activity. Membranes or extracts that are devoid of C activity but that contain the thermostable component(s) of the enzyme (heated wild type S49 or untreated HC-1) do not contain the Mn”+-dependent activity. In addition, the co-frac- tionation of Mn”+-dependent catalytic activity and C during gel exclusion chromatography and sucrose density centrifu- gation appears to provide the best evidence for their identity, since these procedures measure the size, shape, and partial specific volume of a given protein. Finally, the finding that these activities have identical thermal denaturation kinetics complements the fractionation experiments. Both activities are quite labile at 30°C (Fig. 4), and both are similarly stabi- lized at this temperature by Mn2’ and 0.5 mu ATP or App(NH)p. At higher ATP concentration, in the presence of MgZf but not Mn”‘, both activities are labilized at 0°C (Fig. 5). It seems unlikely that such distinctive similarity of behav- ior under differing circumstances is merely fortuitous.
The hydrodynamic parameters determined for C may be compared to those published previously for Gpp(NH)p- treated adenylate cyclase from wild type S49 cells (1). While there is a clear difference in sedimentation coefficient (6.5 S for C versus 7.8 S for wild type, Gpp(NH)p-treated enzyme), it is not large. The fact that the Gpp(NH)p-activated enzyme appears to be larger than C by approximately 40,000 daltons is provocative, in view of Pfeuffer’s characterization of a Gpp(NH)p-binding protein of this size that appears related to adenylate cyclase (3) (see below). However, G/F appears to be considerably larger than this (M, - 1 to 2 x lo”), and the relationship between G/F and C and the Gpp(NH)p-treated enzyme may not be straightforward. Experimentation on these questions is clearly warranted.
At this time we are unsure of the significance of the effects of nucleotides and divalent cations on C. In addition to the catalytic site (which utilizes a metal’ nucleotide complex), adenylate cyclase has a divalent cation binding site (25), and it is certainly plausible to assume that this site, in addition to possible regulatory nucleotide binding sites, exist on the cat- alytic protein. The disparate effects of cation-nucleotides on the stability of C could result from actions at the same or
different sites. The stabilizing effect is observed with Mn’+ and is pronounced in the presence of moderate concentrations of ATP, App(NH)p, or ADP. Furthermore, GTP and Gpp(NH)p plus Mg’+ stabilize C, although this phenomenon has not been studied in detail. In contrast, only ATP at concentrations greater than 2 mu, and only in the presence of Mg”+, has been observed to labilize these activities at 0°C. While it is possible that these effects could all result from binding to the catalytic site of the enzyme, more detailed data are needed to determine the mechanism of these effects on enzyme stability.
C activity in extracts of AC membranes, as assayed by reconstitution of Mg’+-dependent adenylate cyclase with the heat-stable component(s), is more sensitive to N-ethylmaleim- ide than is the Mn”‘-dependent enzymatic activity. While C is inhibited 50% at about 30 to 50 pM N-ethylmaleimide, Mn”+- dependent activity is inhibited to this extent only at 0.5 to 1 mu (Fig. 8). While this disparity might be taken as evidence for the nonidentity of these activities, we think it more likely that there is a particularly reactive sulfhydryl group on C that is required for interaction with the thermostable component but which is not required for simple catalytic function.
Two other sources of MI?+-dependent adenylate cyclase activity must be considered. One is the soluble testicular adenylate cyclase discussed above. This appears not to be related to the Mn’+-dependent activity characterized here. The other has been described by Mittal and Murad (18), who reported that soluble guanylate cyclase can utilize MnATP as an alternative substrate. This activity is stimulated slightly by NaF and markedly by NaN.?, NO, or NaZFe(CN)r,NO; these responses are variable with the tissue of origin. While there is guanylate cyclase activity in AC membranes, it is low (about 10 to 20% of Mn”+-dependent adenylate cyclase) and is not denatured at 30°C under conditions identical with those of Fig. 4 (data not shown). We have attempted, without success, to reconstitute Mg’+-dependent adenylate cyclase activity using a heat-inactivated wild type extract and a fraction of rat liver guanylate cyclase purified by DEAE-cellulose chroma- tography (18).
We know less about the thermostable regulatory compo- nent(s) of adenylate cyclase than we do about C. Thermal inactivation analysis at 50°C distinguishes two relevant activ- ities, G and F, in heated extracts of wild type S49 (8) or B82 (not shown) plasma membranes. Because hormone- and Gpp(NH)p-stimulated activities decline at the same relatively rapid rate (Fig. 9) and NaF-stimulated activity declines at a slower rate, it is suggested that G activity is required for the guanine nucleotide-mediated effect of hormones on the en- zyme and that F activity is essential for the observation of any Mg2+-dependent catalytic activity. Furthermore, stabili- zation of G activity by Gpp(NH)p (8) or G/F activity by GTP (not shown) indicates the presence of a guanine nucleotide binding site on G/F. Verification of these speculations will require purification of G/F and determination of whether one or more polypeptide chains is involved. To date, we have been unable to separate G and F activities in a convincing fashion. They co-fractionate according to the following procedures: gel exclusion chromatography in the presence of 0.75% sodium cholate or 0.1%~ Lubrol 12A9, sucrose density gradient centrif- ugation, hydroxyapatite chromatography in 0.1% Lubrol12A9, ammonium sulfate precipitation, or DEAE-cellulose chroma- tography in 5 M urea plus 0.1% Lubrol 12A9 or 1% sodium cholate.
Circumstantial evidence suggests that G/F might also be involved with the purine nucleotide-mediated changes in af- finity of hormone binding; much firmer evidence indicates that C is not. Maguire et al. (22) showed that those purine

Reconstitution of Catecholamine-sensitive Adenylate Cyclase 6411
TABLE VI
Correlation of G/F and C with nucleotide-induced alterations in hormone binding to the ,l-adrenergic receptor
G/F and C are defined in the manuscript. The effect of nucleotides (particularly guanine nucleotides) on the binding of agonists, but not antagonists, is to decrease affinity and to increase the apparent Hill coefficient. When the effect of nucleotides is lost (designated -), the agonist binding isotherm in the absence of nucleotide assumes a position and shape indistinguishable from that normally found in the presence of nucleotide (IO, 11, 22, 24).
Nucleotide ef- Membrane G/F C feet on hor- Ref.
mane binding
Wild type S49 + + + (11) AC S49 - + - (11) Wild type S49 - 10 mM - - - (24)
MalNEt” HC-1 + + ,I -
Wild type S49 - 1 mM + - + (24) MalNEt
Wild type S49 - 37’C for 45 + - + h
min UNC + + - (10) Wild type S49 - ffiipin + + - (24
n MalNEt, N-ethylmaleimide. ’ Described in this report.
nucleotides that permit activation of adenylate cyclase by /?- adrenergic agonists also cause a decrease in their affinity for
the receptor and an increase in the apparent Hill coefficient of the binding isotherm (11, 22). Antagonist binding is not affected. Receptors for glucagon and prostaglandin E2 appear to be similarly affected by these purine nucleotides (26-28). At the current level of resolution of components, the presence of G/F seems to parallel the ability of these nucleotides to alter hormone binding, and its absence coincides with the lack of such alteration. Agonist binding and its regulation by guanine nucleotides are not changed by loss of C. These relationships are summarized in Table VI. The reconstitution of nucleotide regulation of agonist binding to AC membranes by the addition of solubilized G/F has so far been unsuccessful, presumably because of interference by added detergent. It should be noted that in several tissues, where coupling of hormone binding to the activation of adenylate cyclase is poor, no measurable effects of nucleotide on hormone binding are observed (cfi Ref. 29).
It has been suggested that there may be two regulatory sites for guanine nucleotides, one whose occupation by Gpp(NH)p leads to activation of adenylate cyclase and an- other whose occupation is manifest particularly by alterations in the binding of hormones. The evidence for this is indirect and is based particularly on different concentration require- ments for the two effects, different structural specificity (30), and the slow reversibility of the effect of Gpp(NH)p on the enzyme despite its readily reversible effect on binding (11). I f in fact there are two sites, G/F would appear to encompass that required for activation of adenylate cyclase by Gpp(NH)p. The hypothetical second site might also be asso- ciated with G/F or with the function whose absence or alter- ation is responsible for the phenotype of the uncoupled (UNC) S49 cell variant. These possibilities may be synonymous, however, since UNC and AC- S49 cells appear to have over- lapping defects (7, 31); it is as yet unknown if the UNC phenotype results from the loss or modification of a specific unidentified factor or from an alteration of G/F. This question is now under investigation, since UNC membranes can be recoupled with detergent extracts that contain G/F, but not C, activity.”
’ P. C. Sternweis and A. G. Gilman, manuscript in preparation.
G/F presumably relates to other components of adenylate cyclase that are currently under study in other laboratories.
Pfeuffer (3) has reported that affinity chromatography of detergent-solubilized avian erythrocyte membranes on GTP- substituted Sepharose decreases Gpp(NH)p-stimulated ade- nylate cyclase activity. A protein that is eluted from the column by GTP or Gpp(NH)p restores this activity and also stimulates depleted NaF-stimulated activity. The relevant polypeptide that binds to the affinity support has a molecular weight of 42,000 by dodecyl sulfate polyacrylamide gel elec- trophoresis, but it is not possible to compare this value to our preliminary data that indicate a rather larger size for native G/F. It is possible that Pfeuffer has isolated either G or G/F by his procedure, since roughly a quarter of G/F activity in a heated wild type extract will bind to an affinity column synthesized according to the method of Pfeuffer (3). This low efficiency and possible difference in size may be due to species differences. Since Pfeuffer found adenylate cyclase activity in the fractions that do not bind to the affinity column (3), he may not, according to our model, have effected a total sepa- ration of C from G/F.
It is also plausible that G/F may represent at least a portion of the hormone-stimulated GTPase described by Cassel and Selinger (5). Molecular characteristics of this protein are as yet unavailable, however, and the stimulation of its activity by hormone would presumably not be assayable in detergent solution. The assay of any GTPase activity associated with G/F will depend on its purification to avoid large background nucleoside triphosphatase activity (5).
The existence of multiple components of adenylate cyclase and the presence of one of these in AC plasma membranes
forces us to re-evaluate the mechanism by which solubilized adenylate cyclase and AC- membranes combine to reconsti- tute catecholamine-stimulated activity. The data of Figs. 1 to 3 and Table IV indicate that the addition of G/F to the AC- membranes (which contain C and the ,&adrenergic receptor) is sufficient, considering only the known components of the system, for the reconstitution of a hormone-responsive en- zyme. In fact, the data shown in Figs. 13 and 14 argue that when G, F, and C (i.e. an untreated B82 or wild type S49 extract) are added to AC membranes, the only source of hormone-stimulated activity arises from the interaction of the added G/F with the C already in the membrane. The activity of the C that is added in the extract is expressed only as NaF or Gpp(NH)p-stimulated activity. Similarly, an AC extract cannot reconstitute hormone-stimulated activity when added to a membrane that already contains G/F and the P-adrener- gic receptor, such as membranes from HC-1 cells (Table V) or heated or N-ethylmaleimide-treated membranes from wild type S49 cells. Since NaF- and Gpp(NH)p-stimulated activity can be observed, these findings suggest that C must be ori- ented in the membrane in a specific fashion in order to interact productively with other components of the system that are necessary for the response to hormone, and we have so far
been unsuccessful in attempts to reinsert C into an appropriate membrane in the correct orientation. Since C appears to bind detergent in solution, as evidenced by its partial specific volume (Table II), it is plausible that it has a significant hydrophobic surface that normally interacts with the hydro- carbon moieties of membrane phospholipids (32). Hence it may be necessary to disrupt the membrane partially in order to integrate C into the bilayer or to remove detergent that may block hydrophobic sites necessary for the binding of C to phospholipids in the membrane. A similar situation may also obtain for the P-adrenergic receptor. This protein has a sig- nificant hydrophobic surface (l), and we have not succeeded
in the reconstitution of a catecholamine-stimulated adenylate

6412 Reconstitution of Catecholamine-sensitive Adenylate Cyclase
cyclase using detergent-solubilized receptor.
In contrast, solubilized G/F, which may bind little or no
detergent in solution, interacts freely with membrane-bound
C and receptor to promote a response of the enzyme to
hormone. It is tempting to speculate that C and hormone
receptors are integral membrane proteins situated primarily
in the core of the bilayer or bound to it via a hydrophobic
extension, and that G/F is a peripheral protein on the cyto- plasmic surface of the plasma membrane. Such speculations
can be tested in increasingly defined reconstituted systems.
Acitrzowledgments-We thank Debra L. Jordan and Pamela M. Van Arsdale for excellent technical assistance, Hannah J. Anderson for maintenance of cell cultures, and Wendy J. Deaner for preparation of the manuscript. We are indebted to Doctors Ferid Murad and Kikuo Ichihara for performing guanylate cyclase assays and providing partially purified liver guanylate cyclase. Lubrol 12A9 was a gift of ICI, Ltd.
11.
12.
13.
14. 15.
16.
17.
18. 19.
20. 21.
1.
2.
3. 4.
5.
6.
7.
8.
9.
10.
REFERENCES 22.
Haga, T., Haga, K., and Gilman, A. G. (1977) J. Biol. Chem. 252, 23. 5776-5782
Limbird, L. E., and Lefkowitz, R. J. (1977) J. Biol. Chem. 252, 24. 799-802
Pfeuffer, T. (1977) J. Biol. Chem. 252, 7224-7234 25. Pfeuffer, T., and Helmreich, E. J. M. (1975) J. Biol. Chem. 250,
867-876 26. Cassel, D., and Selinger, Z. (1976) Biochim. Biophys. Acta 452,
538-551 27. Cassel, D., and Selinger, Z. (1977) Proc. N&Z. Acad. Sci. U. S. A.
Proc. Natl. Acad. Sci. U. S. A. 74,2016-2020 Ross, E. M., Maguire, M. E., Sturgill, T. W., Biltonen, R. L., and
Gilman, A. G. (1977) J. Biol. Chem. 252,5761-5775 Insel, P. A., Maguire, M. E., Gilman, A. G., Bourne, H. R., Cofflno,
P., and Melmon, K. L. (1976) Mol. Pharmacol. 12, 1062-1069 Rosen, 0. M., and Rosen, S. M. (1969) Arch. Biochem. Biophys.
131,449-456 Neville, D. M., Jr. (1968) Biochim. Biophys. Acta 154, 540-552 Braun, T., and Dods, R. F. (1975) Proc. Natl. Acad. Sci. U. S. A.
72,1097-1101 Salomon, Y., Londos, C., and RodbeII, M. (1974) Anal. Biochem.
58, 541-548 Maguire, M. E., Wiklund, R. A., Anderson, H. J., and Gilman, A.
G. (1976) J. Biol. Chem. 251, 1221-1231 Mittal, C. K., and Murad, F. (1977) J. Biol. Chem. 252,3136-3140 Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193, 265-275 Neer, E. J. (1978) Adu. Cyclic Nucleotide Res. 9, 69-83 Bourne, H. R., Coffino, P., and Tomkins, G. M. (1975) Science
187, 750-752 Maguire, M. E., VanArsdale, P. M., and Gilman, A. G. (1976) Mol.
Pharmacol. 12, 335-339 Lefkowitz, R. J., MuIlikin, D., and Caron, M. G. (1976) J. Biol.
Chem. 251,4686-4692 Hewlett, A. C., Van Arsdale, P. M., and Gilman, A. G. (1978) Mol.
Pharmacol. 14, 531-539 Johnson. R. A.. Garbers. D. L.. and PiIkis. S. J. (1976) J. Suara-
mol. Siruct. 4, 205-226 Rodbell, M., Krans, H. M. J., Pohl, S. L., and Birnbaumer, L.
(1971) J. Biol. Chem. 246, 1872-1876 Lin, M. C., Nicosia, S., Lad, P. M., and Rodbell, M. (1977) J. Biol.
Chem. 252, 2790-2792 74,3307-3311
Ross, E. M., and Gilman, A. G. (1977) Proc. Natl. Acad. Sci. U. S. A. 74, 3715-3719
Ross, E. M., and Gilman, A. G. (1977) J. Biol. Chem. 252, 6966-6969
Brunton, L. L., Maguire, M. E., Anderson, H. J., and Gilman, A. G. (1977) J. Biol. Chem. 252, 1293-1302
Haga, T., Ross, E. M., Anderson, H. J., and Gilman, A. G. (1977)
28. Moore, W. V., and Wolff, J. (1973) J. Biol. Chem. 248,5705-5711 29. Maguire, M. E., Ross, E. M., and Gilman, A. G. (1977) Adu. Cyclic
Nucleotide Res. 8, 1-83 30. Lad, P. M., Welton, A. F., and RodbeII, M. (1977) J. Biol. Chem.
252, 5942-5946 31. Schwarzmeier, J. D., and Gilman, A. G. (1977) J. Cyclic Nucleo-
tide Res. 3, 227-238 32. Clarke, S. (1975) J. Biol. Chem. 250, 5459-5469





























