Embed Size (px)
Citation preview

General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Association between pituitary adenylate cyclase-activating polypeptideand reproduction in the blue gourami
Gal Levy a,b,c, Karen Jackson a,b, Gad Degani a,b,*
a School of Science and Technology, Tel-Hai Academic College, Upper Galilee 12210, Israelb MIGAL-Galilee Technology Center, Kiryat Shmona 11016, Israelc Department of Neurobiology, George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv 69978, Israel
a r t i c l e i n f o a b s t r a c t
Article history:Received 24 April 2009Revised 30 July 2009Accepted 25 September 2009Available online xxxx
Keywords:PRPGHRHLPGHRHPACAPReproductionBlue gourami
0016-6480/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.ygcen.2009.09.015
* Corresponding author. Address: School of ScienAcademic College, Upper Galilee 12210, Israel. Fax: +
E-mail address: [email protected] (G. Degani).
Please cite this article in press as: Levy, G., et al.rami. Gen. Comp. Endocrinol. (2009), doi:10.10
In order to gain a better understanding of the roles of pituitary adenylate cyclase-activating polypeptide(PACAP) in reproduction and growth, the expression of the PACAP gene during the reproduction cycle andits potential role in regulating gonadotropin and growth hormone (GH) gene transcription in blue gou-rami were investigated. The cDNA sequences of the full-length blue gourami brain PACAP and that ofits related peptide (PRP) were acquired. PACAP cDNA had two variants, obtainable by alternative splicing:a long form encoding for both PRP and PACAP and a short form encoding only for PACAP. In females,mRNA levels of PACAP were very high only in individuals with oocytes in the maturation stage, as com-pared to levels in unpaired vitellogenic and non-vitellogenic fish. The PACAP mRNA levels in males werehigh only in nest builders, as opposed to in non-nest building males and juveniles. In pituitary culturecells from high vitellogenic females, PACAP38 (the 38 amino acid form) only brought about an increasein bFSH levels, without altering GH and bLH mRNA levels. On the other hand, in adult non-reproductivemale pituitary cells, PACAP38 decreased the GH mRNA level. Based on these results, we propose that inthe blue gourami, PACAP is involved in the final oocyte maturation stage in females, whereas in males, itis associated with sexual behavior. In addition, the effect of PACAP38 on pituitary hormone gene expres-sion is different in females and males, indicating that PACAP38 is potentially a hypophysiotropic regula-tor of reproduction, which mediates pituitary hormone expression.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
In teleosts, as in other vertebrates, growth and reproduction aretightly regulated, mainly via the hypothalamus–pituitary axis. Thehypothalamic decapeptide, gonadotropin-releasing hormone(GnRH), controls the release of the pituitary gonadotropins, folli-cle-stimulating hormone (FSH) and luteinizing hormone (LH),which in turn control gametogenesis (Yaron et al., 2003).
It is becoming increasingly evident that growth hormone (GH) hasa role in reproduction (Marchant et al., 1989; Trudeau et al., 1993;Ran et al., 2004). A complex regulatory system, involving multiplestimulatory and inhibitory factors, may control the GH axis. In addi-tion to its role in development and somatic growth, GH is involved inspermatogenesis regulation (Scott et al., 1984; Stacey et al., 1984;Hull and Harvey, 2001; Degani et al., 2003b). It has been reported thatin fish, GnRH augments the GH release from the pituitary in vivo(Melamed et al., 1996; Sohn et al., 2001; Klausen et al., 2002; Kiyaand Oka, 2003; Ran et al., 2004). Morphological features support
ll rights reserved.
ce and Technology, Tel-Hai972 4 6944980.
Association between pituitary a16/j.ygcen.2009.09.015
the existence of direct stimulatory actions of GnRH on GH release(Parhar and Iwata, 1994), and specific GnRH receptors have been de-tected in goldfish somatotrophs (Cook et al., 1991). However, theinfluence of GnRH on GH mRNA levels is still controversial. GnRH in-creases GH mRNA content in goldfish (Klausen et al., 2002) and sock-eye salmon (Taniyama et al., 2000), but direct exposure to GnRH3 intilapia did not alter the transcript levels (Melamed et al., 1996).
Pituitary adenylate cyclase-activating polypeptide (PACAP) andits related peptide (PRP) are members of the vasoactive intestinalpeptide (VIP)/secretin/glucagons family. Both peptides are encodedby a single gene (PRP–PACAP), which is further cleaved to releasethese two different peptides. In vertebrates, PACAP is a neuropeptidethat is associated with the release of pituitary GH (Montero et al.,2000), whereas PRP (also known as Growth hormone releasing hor-mone-like peptide GHRH-LP) is active in teleosts and not in mam-mals (Tam et al., 2007). Previously, a model of molecular evolutionof these peptides suggested that the mammalian growth hormonereleasing hormone (GHRH) evolved from the non-mammalianGHRH-like peptide. However, more recently, Lee et al. (2007) discov-ered a new GHRH peptide, which is homologous to mammalianGHRH, implying that the GHRH-like peptide is homologous to themammalian PACAP-related peptide (PRP). In teleosts, the full length
denylate cyclase-activating polypeptide and reproduction in the blue gou-

2 G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
of the PRP–PACAP cDNA has been cloned from the eel (Montero et al.,1998), salmon (Parker et al., 1993), catfish (McRory et al., 1995),grouper (Accession No. AY869693), carp (Accession No. EF592488),Atlantic cod (Accession No. DQ109988), trout (Accession No.AF343976) sturgeon, whitefish, grayling, flounder, halibut (Adamset al., 2002) and tilapia (Chi-Wei et al., 2007). In several teleosts,two transcripts of PRP–PACAP, resulting from exon skipping and apartial deletion of exon 4, have been reported (Parker et al., 1993;Parker et al., 1997; Small and Nonneman, 2001; Adams et al.,2002). In addition, in several teleosts, two genes encoding for PACAP(PACAPa and PACAPb) have been identified (Fradinger and Sher-wood, 2000; Cardoso et al., 2005). In the zebrafish, these two geneshave different cDNA sequences and an alternative splicing mecha-nism existing only in zfPACAP2 (Wang et al., 2003).
There are two peptides of PACAP, PACAP27 and PACAP38, derivedfrom the same precursor and consisting of 27- and 38-amino acids,respectively. The PACAP sequence was found to be more conservedamong taxa than PRP, indicating its significance as an importantmodulator in a variety of endocrine cells, as well as in various othercells situated in the hypothalamus, pituitary and peripheral organs(Arimura, 1992, 1998; Montero et al., 2000). The mRNA of PACAPreceptors, namely PACI and VPACI, have been identified in the pitui-tary of the goldfish and tilapia, hinting that they may have a role inpituitary hormonal regulation (Adams et al., 2002; Chi-Wei et al.,2007). In fish, PACAP has a more potent effect on GH secretion thandoes PRP (Goth et al., 1992; Wong et al., 2000). In addition, PACAP in-creased LH secretion and transcription in goldfish and tilapia, prob-ably by a paracrine mechanism (Wong et al., 2000; Chang et al.,2001; Yaron et al., 2001). In rats, PACAP stimulates a variety of ovar-ian functions, such as the synthesis of estradiol (E) and progesterone(P) in ovarian granulosa cells, through the cAMP signaling pathway(Kotani et al., 1998). However, PRP did not alter GtH release and tran-scription in the examined teleost (Montero et al., 2000).
The blue gourami (Trichogaster trichopterus) belongs to the subor-der Labyrinthici (characterized by the presence of an air-filledbreathing cavity [the labyrinth], located above the gills under theoperculum) and to the family Anabantidae. This fish serves as a use-ful model in studying the role of endocrine regulation on reproduc-tion, since it is multi-spawning and male dependent, withasynchronic ovary development. Thus, each stage of its gonadaldevelopment can be controlled and examined separately in the lab-oratory (Degani, 1993a,b; Jackson et al., 1999). The profiles of bFSH,bLH and GH secretion and gene expression, and the pattern of sexsteroid secretion during gonadal development in male and femaleblue gourami, have been previously reported (Degani and Boker,1992). The highest levels of blue gourami GH (bgGH) mRNA werefound in immature female pituitaries during high vitellogenesisand maturation. These results imply that GH may play a role in thegonadal cycle of the female blue gourami (Goldberg et al., 2004;Degani et al., 2006).
To date, the crosstalk between growth and reproduction is notfully understood. Molecular analysis of the central hypothalamicpeptides, regulating growth and reproduction, can provide an in-sight into their physiological role. Therefore, the aims of this studywere to: (1) clone and sequence the gene, PRP–PACAP, (2) study theexpression profiles of PACAP in the blue gourami during differentstates of reproduction, and (3) examine the role of PACAP in regu-lating pituitary hormone transcription.
2. Materials and methods
2.1. Fish and sampling procedure
Blue gourami fish (T. trichopterus), which were maintained andbred at MIGAL Laboratories in northern Israel, were used in this
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
study. Investigations were conducted in accordance with the Inter-national Guiding Principles for Biomedical Research Involving Ani-mals as promulgated by the Society for the Study of Reproduction.The fish were grown in containers (2 � 2 � 0.5 m) at a temperatureof 27 �C under a light regime of 12 h light:12 h darkness (Jacksonet al., 1994) and fed an artificial diet (45% protein, 7% fat) supple-mented by live food (Artemia salina). Brains were collected fromthree-month-old females and males (4�5 ± 0�6 g) at various stagesof gonadal development. The stages in females were: previtello-genesis (PV), low vitellogenesis (LV), high vitellogenesis (HV) andmaturation (MS) (Jackson et al., 1999). The somatic and gonadalweights were recorded for the gonadosomatic index (GSI) calcula-tions, and gonadal samples were processed for histological deter-mination of the reproductive stage. Males with an un-detectableGSI were considered to be juveniles.
2.2. Histological analysis
Gonadal samples were fixed in Bouin, and subsequently pro-cessed for light microscopy. Paraffin sections of 6 lm were stainedwith hematoxylin and eosin, as previously described (Jackson et al.,1994).
2.3. RNA extraction and cDNA synthesis
Total RNA was extracted from freshly excised whole brains ofmale (n = 17) or female (n = 24) gourami, using Trizol reagent(Invitrogen, CA) according to the manufacturer’s recommenda-tions. RNA samples were incubated with RNase-free deoxyribonu-clease-I (Roche Diagnostics, Germany) to eliminate contaminatinggenomic DNA. First strand cDNA was synthesized by the Reverse-IT1st Strand Synthesis Kit (ABgene, UK) from 0.5 to 2 lg total RNA for1 h at 57 �C, followed by 10 min at 75 �C.
2.4. cDNA cloning of PRP–PACAP
The 30 and 50 cDNA clones were amplified by employing the 30
and 50 rapid amplification of cDNA ends (RACE) protocol (RocheApplied Science, Germany). For 50 end cloning, the first cDNAstrand was synthesized with a specific primer (GHRHr183), whichwas designed based on the consensus sequence of PACAP cDNA se-quences of other teleosts. cDNA was purified (PCR Purification Kit,Bioneer Corp., Korea) and a tailing reaction was performed, usingthe dATP terminal deoxy transferase enzyme (Roche Diagnostics,Germany) and 3.5 mM MgCl2. Table 1A summarizes the primersused in the cloning of PRP–PACAP cDNA. Poly-A tailed cDNA wasamplified by the dT(17) anchor primer and GHRHr183 primer, fol-lowed by nested PCR, with the PCR anchor and GHRHr41. For the 30
end cloning, first strand cDNA was synthesized with the dT(17) an-chor primer and amplified with the PCR anchor and GHRH45 prim-ers, followed by nested PCR, with the PCR anchor and GHRH6primers. All PCR amplifications were carried out in a 50 ll reaction,with 2.5 U of Taq polymerase in a suitable reaction buffer, 1.5 mMMgCl2, 0.2 mM of each dNTP (Promega, USA), 2 mM specific prim-ers, 1 ll cDNA template and mineral oil. The PCR conditions for allPCR reactions were: an initial denaturation of 94 �C for 3 min, 50cycles of 94 �C for 1 min, 50 �C for 1 min and 72 �C for 1 min, fol-lowed by a final extension of 5 min at 72 �C. PCR products werepurified and cloned into pGEMT of the pGEMT Easy Vector System(Promega, USA) and propagated in Escherichia coli cells. The recom-binant plasmid was then extracted using the Qiaprep Mini-Prep Kit(Qiagen, Germany) and sent for nucleotide sequence determina-tion. To identify the two forms of the PRP–PACAP precursor cDNAs,a PCR primer set located on both sides of exon 4, GHRHr183 andGHRHf225, was used (Table 1A). The PCR products were electro-phoresed on a 2% agarose gel. PCR products were excised and
denylate cyclase-activating polypeptide and reproduction in the blue gou-

Table 1Nucleotide sequences of primers: (A) 30 and 50 RACE primers; (B) real-time PCR primers.
Primer name Synthesis direction Position Sequence
AGHRHr183 Reverse 818–837 50TACACAGGAGGGCAGTTTGG30
GHRHr41 Reverse 720–740 50CGCTGCCAGGTATTTCTTGAC30
GHRH45 Forward 710–728 50GCAAATGGCAGTCAAGAAA30
GHRH6 Forward 671–690 50AGACGGGATCTTCACAGACA30
GHRHf225 Forward 500–515 50CCCACCAGAAGAGACCAG30
dT(17) anchor Forward and reverse 50GACCACGCGTATCGATGTCGACdT(17)30
PCR anchor Forward and reverse 50GACCACGCGTATCGATGTCGAC30
BPrimer name Gene Synthesis Direction Sequence
GHRHf6 PACAP Forward 50AGACGGGATCTTCACAGACA30
GHRHr183 PACAP Reverse 50TACACAGGAGGGCAGTTTGG30
GLHexfor LH Forward 50CCTGACTGTCCTCCTGGTGT0 30
GLHexrev LH Reverse 50TTTGCTTTTGGTTTGCTGTG 30
GFSHexfor FSH Forward 50GTTGTCATGGCAGCAGTGTT 30
GFSHexrev FSH Reverse 50CCTCGTGGTAGCAATGTCCT 30
GGHExpfor GH Forward 50TTCACAACCGCTATGGACAA 30
GGHExprev GH Reverse 50TGACGCTGCTCTTCAATCTG 30
ExpG18Sfw 18S Forward 50CCGTCGTAGTTCCGACCATA 30
ExpG18Srev 18S Reverse 50CCCTTCCGTCAATTCCTTTA 30
G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx 3
ARTICLE IN PRESS
recovered by PCR purification kit (Bioneer Corp., Korea), and finallysequenced directly, using PRP–PACAP specific primers.
2.5. Sequence assembly and analysis
The PRP–PACAP sequencing data were analyzed with the Laser-gene 6 (DNASTAR, WI) software, which is comprised of several dif-ferent programs. DNA sequences were assembled with the SeqManprogram, and MegAlign was employed for sequence comparisonsby means of the ClustalW method.
2.6. Primary culture of dispersed pituitary cells
Pooled pituitaries were minced with a surgical blade under adissecting microscope with fine forceps and immediately trans-ferred to ice cold M-199 medium containing 10 mM HEPES, 0.3%BSA, 0.003 U/ml penicillin, 0.003 mg/ml streptomycin and0.0013 U/ml nystatin. After incubation at 25 �C for 1 h, cells weredissociated by trypsin and several aspirations through a needle,followed by centrifugation at 3000 rpm for 5 min and suspensionin M-199 medium, containing 10% FCS and 10 mM HEPES,0.003 U/ml penicillin, 0.003 mg/ml streptomycin and 0.0013 U/mlnystatin. The cells were immediately plated in 24-well plates at adensity of 5 � 105 cells/well for 72 h at 28 �C, 5% CO2 and 90%humidity. The medium was changed to M-199 medium, containing0.1% BSA, 10 mM HEPES and 0.003 U/ml penicillin, 0.003 mg/mlstreptomycin and 0.0013 U/ml nystatin (stimulating medium). Fol-lowing 15 min of incubation at 28 �C, the cells were stimulatedwith human PACAP38 (Sigma), diluted in stimulating medium.After incubation, the medium was removed, and the cells wereharvested by scraping for RNA extraction.
2.7. Real-time PCR
In order to compare the PACAP mRNA levels in the brain of thegourami individuals, or mRNA levels of the GH, bLH, and bFSH inthe primary culture of dispersed pituitary cells, the relative abun-dance of mRNA was normalized with the mRNA of the endogenousreference gene, 18S subunit of rRNA (18S rRNA), by the comparativethreshold cycle (CT) method, according to Pfaffl (2001). The relativeamount of each gene was calculated by the formula 2�DCT, whereDCT corresponds to the difference between the CT measured for eachtarget gene, and that determined for 18S rRNA. To validate this meth-
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
od, serial dilutions were prepared from a brain cDNA sample, and theefficiencies of gene amplifications were compared by plotting DCTversus log (template), according to the method of Muller et al.(2002). Linear regressions of the plots showed R2 values of 0.99.Gene-specific primers for the real-time PCR (Table 1B) were de-signed using Primer3 Software. To each of the above PCR mixturesSyber Green Master Mix (ABgene, UK) was added, and amplificationwas carried out in a RotorGene 3000 Sequence Detection System(Corbett Research, Australia) under the following conditions: for PA-CAP-initial denaturation at 95 �C for 15 min, followed by 40 cycles of95 �C for 10 s, 60 �C for 20 s, 72 �C for 20 s and 83 �C for 15 s; for bLH-initial denaturation at 95 �C for 15 min, followed by 40 cycles of95 �C for 10 s, annealing at 60 �C for 15 s and a final extension at72 �C for 20 s; for bFSH-initial denaturation at 95 �C for 15 min, fol-lowed by 40 cycles of 95 �C for 10 s, annealing at 57 �C for 20 s and afinal extension at 72 �C for 20 s; for GH-initial denaturation at 95 �Cfor 15 min, followed by 40 cycles of 95 �C for 10 s, 64 �C for 20 s, 72 �Cfor 20 s and 83 �C for 15 s and for 18S rRNA-initial denaturation at95 �C for 15 min, followed by 40 cycles of 95 �C for 15 s, 64 �C for20 s and a final extension at 72 �C for 20 s. Amplifications of each tar-get gene and reference gene (18S rRNA) cDNAs were performedsimultaneously in separate tubes in duplicates, and the results wereanalyzed with the Q-Gene software (BioTechniques Software Libraryat www.BioTechniques.com). Dissociation-curve analysis was runafter each real-time experiment to ensure that there was only oneproduct. To control for false positives, a non-template negative con-trol was run for each primer pair.
2.8. Statistical analysis
Data are presented as the means ± SEM. The significance of thedifferences between group means of hormone or mRNA levels wasdetermined by one-way analysis of variance (ANOVA) followed byan a posteriori Bonferroni post hoc test using SPSS 17.0 software.Differences were considered statistically significant at P < 0.05.
3. Results
3.1. Cloning and sequence analysis of PRP–PACAP
Schematic structures of the PRP–PACAP cDNA are presented inFig. 1A, and the full-length cDNA sequence of the PRP–PACAP
denylate cyclase-activating polypeptide and reproduction in the blue gou-

SP Cryptic peptide PRP PACAP
SP PACAP
Truncated PRP - PACAP cDNA
Full-length PRP -PACAP cDNA
Cryptic peptide
A
B
Fig. 1. Schematic structures and sequences of PRP–PACAP. (A) The schematic structures of the pre-pro PRP–PACAP cDNA of the blue gourami. (B) The cDNA sequence of PRP–PACAP, which has a signal peptide marked by a dashed black line, the cryptic peptide marked by dashed double lines, the PRP sequence indicated by a single black line, thecleavage site positions marked by boxes and the PACAP sequence, indicated by double black lines. The grey box designates the region deleted in the PRP–PACAP short form.The triangle denotes the 30 polyadenylation signal ATTAAA. UTR-untranslated region; PRP–PACAP-related polypeptide; PACAP-pituitary adenylate cyclase-activatingpolypeptide. (C) RT-PCR products resulting from primers, which were designed according to PRP–PACAP (GHRHr183 and GHRHf225) and which are located on both sides ofexon 4. The cDNA template was prepared from brain RNA of females or males. The size marker is in the lane farthest to the left.
4 G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
Please cite this article in press as: Levy, G., et al. Association between pituitary adenylate cyclase-activating polypeptide and reproduction in the blue gou-rami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015

G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx 5
ARTICLE IN PRESS
precursor, compiled from the data obtained from the 50 and 30
RACE, and its deduced amino acid (aa) sequence, are shown inFig. 1B. Sequence analysis of overlapping 50 and 30 RACE products,isolated from the brain of gourami, revealed that blue gouramiPRP–PACAP cDNA is 985 bp long, with an open reading frame of525 bp, encoding for a 174 aa precursor peptide. The 50 and 30 UTRswere found to be 275 bp and 184 bp in size, respectively, and theputative polyadenylation signal was discovered to be located14 bp upstream of the poly(A) tail. The blue gourami PRP–PACAPprecursor is comprised of a 20 aa long signal peptide, a 62 aa longcryptic peptide and a 45 aa long PRP segment, followed by a 2 aaprocessing site (KR) and a 38 aa long PACAP segment. The PACAP se-quence was found to be similar to the PACAP38 form, containingthe GRR sequence at a position, which could yield the PACAP27peptide.
The deduced nucleotide and aa sequences of this gene in theblue gourami were compared with homologous sequences from anumber of other fish species (Fig. 2). The peptide cleavage sites,which are located at positions 660, 747 and 780 and which pro-duce the PRP, PACAP27 and PACAP38 isoforms, were found to befully conserved. In addition, the gourami have a shorter PRP–PACAPform, 114 bp shorter than the longer form, corresponding to theexcision of the 3aa of the cryptic peptide and the first 35 aa ofPRP (Fig. 1A–C).
A sequence alignment of the PRP aa sequence revealed thatonly the first 32 aa at the N-terminus end are highly conservedin closely related fish. There is an 80.0–93.3% sequence identitybetween the blue gourami PRP sequence and that of other tele-osts, whereas there is only a 31.6–56.6% sequence identity be-tween other tetrapods and fish (Fig. 2A). On the other hand,the aa sequence of PACAP is highly conserved in mammalsthrough fish. The sequence identity between blue gourami PA-
Fig. 2. Sequence comparison of amino acids making up the PRP (A) and PACAP (B) peptidanalysis of PRP and PACAP mature peptide sequences. PRP and PACAP of blue gourami weare represented by dots, whereas differing residues are shown in the single letter code
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
CAP and other teleosts is over 90%, and between gourami andhuman PACAP is 92.1% (Fig. 2B).
3.2. Variations in the PACAP mRNA levels in females during maturationand sexual behavior
The relative mRNA levels of the PACAP were determined inbrains excised from females maintained alone (without males) inthe previtellogenic (PV), low vitellogenic (LV) and high vitellogenic(HV) stages, as well as from mature females (MS), which werethose paired with males. LV females were identified as those witha low percentage (<50%) of oocytes at the advanced vitellogenicstage, whereas HV females were detected by a high percentage(>50%) of oocytes at the advanced vitellogenic stage. Results ofthe histological staining are summarized in Fig. 3. The mRNA levelsof PACAP, which were determined by real-time PCR and normalizedwith the 18S rRNA levels, are summarized in Fig. 4. PACAP mRNAlevels were higher in the paired reproductively active females withoocytes in the maturation stage (MS), than in unpaired females(PV, LH and HV) (3.4-, 9.7- and 4-fold, respectively) (P < 0.05, byANOVA and Bonferroni post hoc test).
3.3. Variations in the PACAP mRNA levels in males during maturationand sexual behavior
Fig. 5 presents the different stages of gonadal development inthe male blue gourami. The variation in the relative mRNA levelsof PACAP was determined in the brain of juvenile and mature males(Fig. 6). PACAP mRNA levels were 4.77-fold higher in mature repro-ductive males (nest builders, paired with females), as compared toin mature non-reproductive (non-nest builders kept separatelyfrom females) and juvenile males (P < 0.05 ANOVA and Bonferronipost hoc test). No significant difference was found between the PA-
es. Sequence alignment was conducted using DNAStar WI Megalign ClustalW in there aligned with corresponding sequences of other vertebrates. Identical amino acids
for amino acids. PACAP is highly conserved in contrast to PRP peptides.
denylate cyclase-activating polypeptide and reproduction in the blue gou-

Fig. 3. Histological sections showing different stages of gonadal development in the blue gourami female. (A) and (B) are non-mature oocytes in the previtellogenic stage(PV). (A) Perinuclear oocyte with an intracellular spherical structure. (B) Oocyte at cortical alveolar stage. Note the irregular outgrowth of the nuclear periphery. (C) Oocyte atearly vitellogenic stage. Note the increase in oocyte size, lipid droplets (L) and the beginning of yolk vesicle accumulation (white arrow). D) Oocyte in the advancedvitellogenic stage. Note the high concentration of yolk vesicles supplanting the periphery of the oocyte (black arrow) and the lipid vesicles (L). (E) Oocyte at maturation stage(MS). Note that the lipid vesicles have been joined into one vesicle (L). The oocyte nucleus is denoted by (n). Sections were stained with hematoxylin and eosin. Bar = 100 lm.
6 G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
CAP mRNA levels of the mature non-reproductive fish and the juve-niles (P > 0.05 ANOVA and Bonferroni post hoc test).
3.4. Effect of PACAP38 on GH, bLH and bFSH gene expression indispersed pituitary cells of gourami females and males
To assess the direct effect of PACAP38 on GH, bLH and bFSH geneexpression, pituitary cells obtained from females and males weretreated with 10 nM human PACAP38 for different time periods,and mRNA levels of pituitary hormones and 18S rRNA were deter-mined by real-time PCR. PACAP38 treatment of the pituitary cellculture, obtained from advanced high vitellogenic females, signifi-cantly increased bFSH subunit mRNA levels after 48 h (2.6-fold). Nosignificant changes in bLH and GH mRNA levels were observed fol-lowing PACAP38 treatment (P > 0.05 ANOVA and Bonferroni post
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
hoc test, Fig. 7). On the other hand, in a pituitary cell culture de-rived from adult non-reproductive males, treatment with PACAP38brought about a significant decrease in GH mRNA levels after 24and 48 h (8.3- and 5.6-fold, respectively) (P < 0.05 ANOVA and Bon-ferroni post hoc test, Fig. 8).
4. Discussion
In this study, blue gourami (T. trichopterus) brain PRP–PACAPcDNA was cloned in order to determine the PACAP expression pro-file during the reproduction cycle in both males and females. Suchdata can contribute to a better understanding of the centralinvolvement of this peptide in the blue gourami, a model repre-senting reproduction control in the suborder Labyrinthici.
denylate cyclase-activating polypeptide and reproduction in the blue gou-

Fig. 4. Relative mRNA levels of PACAP in brains from blue gourami females atdifferent stages of oogenesis. The stages are previtellogenesis (PV), low vitellogen-esis (LV), high vitellogenesis (HV) and maturation (MS). Total brain RNA wasreverse transcribed and used for quantitative real-time PCR. The relative amount ofthe PACAP mRNA was normalized to that of 18S rRNA by the CT cycle method, where2�DCT reflects the relative amount of PRP–PACAP transcripts. Each histogramrepresents the average of at least five independent measurements (means ± SEM;n = 5–7). A different letter above the histogram denotes its significant differencefrom the mRNA levels of the other histograms (P < 0.05 ANOVA and Bonferroni posthoc test).
Fig. 6. Relative mRNA levels of PACAP in brains from blue gourami males atdifferent stages of reproduction: juvenile, mature non-reproductive and maturereproductive. Total RNA from brains was reverse transcribed for quantitative real-time PCR. The relative amount of PACAP mRNA was normalized to that of 18S rRNAby the CT cycle method, where 2�DCT reflects the relative amount of the specificgene precursor transcripts. Each histogram represents the average of at least fiveindependent measurements (means ± SEM; n = 5–6). A different letter above thehistogram denotes its significant difference from the mRNA levels of the otherhistograms (P < 0.05 ANOVA and Bonferroni post hoc test).
G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx 7
ARTICLE IN PRESS
The PRP–PACAP sequence encodes for the PRP as well as for thetwo PACAP peptide isoforms: PACAP27 and PACAP38. A compari-son of PRP sequences revealed that the PRP length and sequence
Fig. 5. Histological sections showing different stages of gonadal development in the bluethe middle of the lobule. (B) Mature non-reproductive male. A high concentration of SZ ithe decrease in the number of SZ in the center of the lobule. Cellular spermatogenesis sspermatocyte, ST-spermatids. Sections were stained with hematoxylin and eosin. Bar = 5
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
varies among other species. The PRP sequence is only moderatelyconserved; it shares over 80% of the sequence with other teleostPRPs and only 31.6% with mammalian PRP. In contrast, thePACAP38 sequence is tightly conserved, showing an identity of over86%, in fish through mammals. Similarly to reports on salmon (Par-ker et al., 1997), catfish (McRory et al., 1995), whitefish (Adams
gourami male. (A) Juvenile male. Note the low concentration of spermatozoa (SZ) ins observed in the middle of the lobule. (C) Mature fish during sexual behavior. Notetages are shown in A and B at the periphery of the lobule: SG-spermatogonia, SC-0 lm.
denylate cyclase-activating polypeptide and reproduction in the blue gou-
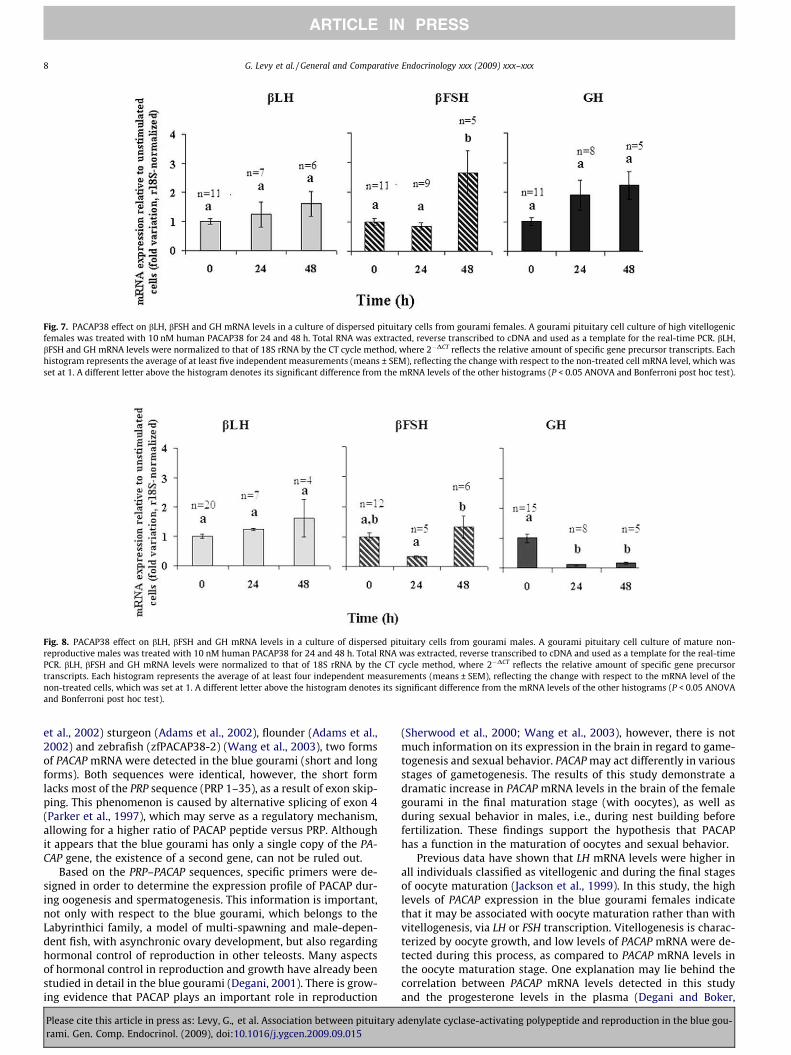
Fig. 7. PACAP38 effect on bLH, bFSH and GH mRNA levels in a culture of dispersed pituitary cells from gourami females. A gourami pituitary cell culture of high vitellogenicfemales was treated with 10 nM human PACAP38 for 24 and 48 h. Total RNA was extracted, reverse transcribed to cDNA and used as a template for the real-time PCR. bLH,bFSH and GH mRNA levels were normalized to that of 18S rRNA by the CT cycle method, where 2�DCT reflects the relative amount of specific gene precursor transcripts. Eachhistogram represents the average of at least five independent measurements (means ± SEM), reflecting the change with respect to the non-treated cell mRNA level, which wasset at 1. A different letter above the histogram denotes its significant difference from the mRNA levels of the other histograms (P < 0.05 ANOVA and Bonferroni post hoc test).
Fig. 8. PACAP38 effect on bLH, bFSH and GH mRNA levels in a culture of dispersed pituitary cells from gourami males. A gourami pituitary cell culture of mature non-reproductive males was treated with 10 nM human PACAP38 for 24 and 48 h. Total RNA was extracted, reverse transcribed to cDNA and used as a template for the real-timePCR. bLH, bFSH and GH mRNA levels were normalized to that of 18S rRNA by the CT cycle method, where 2�DCT reflects the relative amount of specific gene precursortranscripts. Each histogram represents the average of at least four independent measurements (means ± SEM), reflecting the change with respect to the mRNA level of thenon-treated cells, which was set at 1. A different letter above the histogram denotes its significant difference from the mRNA levels of the other histograms (P < 0.05 ANOVAand Bonferroni post hoc test).
8 G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
et al., 2002) sturgeon (Adams et al., 2002), flounder (Adams et al.,2002) and zebrafish (zfPACAP38-2) (Wang et al., 2003), two formsof PACAP mRNA were detected in the blue gourami (short and longforms). Both sequences were identical, however, the short formlacks most of the PRP sequence (PRP 1–35), as a result of exon skip-ping. This phenomenon is caused by alternative splicing of exon 4(Parker et al., 1997), which may serve as a regulatory mechanism,allowing for a higher ratio of PACAP peptide versus PRP. Althoughit appears that the blue gourami has only a single copy of the PA-CAP gene, the existence of a second gene, can not be ruled out.
Based on the PRP–PACAP sequences, specific primers were de-signed in order to determine the expression profile of PACAP dur-ing oogenesis and spermatogenesis. This information is important,not only with respect to the blue gourami, which belongs to theLabyrinthici family, a model of multi-spawning and male-depen-dent fish, with asynchronic ovary development, but also regardinghormonal control of reproduction in other teleosts. Many aspectsof hormonal control in reproduction and growth have already beenstudied in detail in the blue gourami (Degani, 2001). There is grow-ing evidence that PACAP plays an important role in reproduction
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
(Sherwood et al., 2000; Wang et al., 2003), however, there is notmuch information on its expression in the brain in regard to game-togenesis and sexual behavior. PACAP may act differently in variousstages of gametogenesis. The results of this study demonstrate adramatic increase in PACAP mRNA levels in the brain of the femalegourami in the final maturation stage (with oocytes), as well asduring sexual behavior in males, i.e., during nest building beforefertilization. These findings support the hypothesis that PACAPhas a function in the maturation of oocytes and sexual behavior.
Previous data have shown that LH mRNA levels were higher inall individuals classified as vitellogenic and during the final stagesof oocyte maturation (Jackson et al., 1999). In this study, the highlevels of PACAP expression in the blue gourami females indicatethat it may be associated with oocyte maturation rather than withvitellogenesis, via LH or FSH transcription. Vitellogenesis is charac-terized by oocyte growth, and low levels of PACAP mRNA were de-tected during this process, as compared to PACAP mRNA levels inthe oocyte maturation stage. One explanation may lie behind thecorrelation between PACAP mRNA levels detected in this studyand the progesterone levels in the plasma (Degani and Boker,
denylate cyclase-activating polypeptide and reproduction in the blue gou-

G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx 9
ARTICLE IN PRESS
1992). In female rats, PACAP was found to be involved in steroido-genesis (Kotani et al., 1998) and steroid-induced reproductivebehavior (Ha et al., 2000; Apostolakis et al., 2004).
Thus, PACAP may be involved in final oocyte maturation bymultiple mechanisms along the hypothalamic-pituitary axis: (i)alteration of LH or FSH transcription levels in the pituitary, (ii)upregulation of progesterone secretion in the ovary, and (iii) par-ticipation in the positive feedback of progesterone on GnRH syn-thesis and release from the hypothalamus.
In males, spermatogenesis is achieved by synchronous cell divi-sion and differentiation. Moreover, germ cell proliferation correlateswith the GSI (Nagahama, 1994). Our results demonstrate a correla-tion between PACAP mRNA levels and the expression levels of LHin the pituitary and the plasma (Degani et al., 1997, 2003a). Testos-terone levels also increase in mature fish, and in the rat, PACAP in-duced testosterone secretion from the testis (Rossato et al., 1997).In addition, 11 keto-testosterone induced nest building glue in malethree-spined sticklebacks (Jakobsson et al., 1999). According to all ofthe above data, PACAP may be involved in regulating sexual behav-ior, inducing gonadotropin release and steroidogenesis.
These conclusions are based only on changes in the mRNA lev-els, which may not always correlate with the protein expression.This limitation stems from the difficulty in measuring the bluegourami PACAP protein level in the brain. However, our data indi-cate the activity level of the PACAP neuronal system in the bluegourami brain (Canosa et al., 2008).
Essentially, based on the examination of PRP–PACAP mRNA lev-els, it cannot be determined whether PRP takes part in reproduc-tion regulation, since PRP is independently expressed by analternative splicing mechanism, and PACAP is synthesized fromboth long and short forms. Therefore, further research is clearlywarranted to study the expression pattern of PRP during blue gou-rami oogenesis and sexual behavior. Furthermore, the existence ofa second PACAP gene that might affect pituitary hormones shouldbe considered.
In mammals and teleosts, accumulating data show that PACAPmay exert hypophysiotropic roles on gonadotropins and growthhormones, suggesting a role in reproduction regulation. Previousinvestigations demonstrated that PACAP directly stimulates bothtranscription and release of GH (Sze et al., 2007), LH and FSH frompituitary cells in teleosts (Melamed et al., 1995; Montero et al.,2000; Chang et al., 2001; Gur et al., 2002). In the zebrafish, PACAP2affects ovarian function (Wang et al., 2003). In mammalian pitui-tary cells, PACAP differentially regulates gonadotropins; continu-ous PACAP treatment suppressed bFSH mRNA levels and did notalter bLH mRNA levels (Tsujii and Winters, 1995). In addition, thepresence of immunoreactive PACAP receptor sites in the pituitaryof the goldfish supports the role of PACAP as a regulator of pituitaryhormones (Chan et al., 1998).
In this study, the action of PACAP38 on pituitary hormone tran-scription was investigated using pituitary dispersed cells of bothmales and females. Our data demonstrate that in the pituitary cul-ture obtained from high vitellogenic females, PACAP38 increasedmRNA levels of bFSH and did not alter the mRNA levels of bLHand GH. Our results are not in agreement with the findings in carpand goldfish pituitary cell cultures, in which PACAP increased GHmRNA levels after 48 h (Sze et al., 2007). In teleosts, it was sug-gested that PACAP is a potent GH secretagogue. Nevertheless, itseffect on GH transcription levels in females remains unclear. Theseresults imply that in high vitellogenic females, PACAP is involved inreproduction regulation via upregulation of bFSH genetranscription.
Interestingly, PACAP38 decreased the basal mRNA levels of GHin pituitary cultures obtained from mature non-reproductivemales. This is not in agreement with a recent study in zebrafish(Lin and Ge, 2009). Furthermore, it has been reported that PACAP
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
can induce GH secretion in teleosts (Parker et al., 1997; Wonget al., 2000; Rousseau et al., 2001; Xiao et al., 2002.) and can en-hance GnRH-induced-GH release in goldfish (Chang et al., 2001).Based on our data, we suggest that PACAP may negatively regulateGH mRNA transcription, only in mature non-reproductive gouramimales, since they have reached maximal growth, possibly underthe local effect of other peptides, e.g., somatostatin. In addition,our data demonstrate that PACAP did not alter bLH and bFSHmRNA levels. In other vertebrates, the effects of PACAP38 on bFSHand bLH are controversial. In the tilapia, PACAP38 increased bothbFSH and bLH mRNA levels in mature and immature males (Yaronet al., 2003); in the zebrafish PACAP decreased both bFSH and bLHmRNA levels (Lin and Ge, 2009) and in rats, PACAP down-regulatedonly bFSH mRNA through stimulation of follistatin and did not al-ter bLH mRNA levels (Inagaki et al., 1994). The variation in the PA-CAP effect on bFSH, bLH and GH gene expression among theseorganisms is related to the species, gender, sexual maturity, as wellas the experimental approach.
It should be noted that the culture system used in this studycontained all pituitary cell types, which may regulate GH, bLHand bFSH gene transcription by local mechanisms, such as para-crine regulation.
In conclusion, in this report, two PRP–PACAP cDNA forms werecloned from the gourami brain in order to facilitate research onthe differential expression patterns of PACAP during reproduction.The PRP and PACAP gene sequences have distinct phylogeneticcharacteristics, distinguishing them from that of other teleosts.The PACAP sequence was found to be highly conserved, comparedto the sequence of PRP. The seasonal fluctuations of PACAP tran-scripts have also demonstrated that this gene has a differentialreproduction control in males and females. In females, the dramat-ically high expression levels of PACAP during the maturation stagemay indicate that this hormone is involved in mating, as well as inoocyte maturation control and upregulation of bFSH gene expres-sion at the pituitary level. In contrast, in males, PACAP regulatesnest building and downregulates GH gene expression.
Yet, it is not excluded that other factors, such as pituitary hor-mones, steroids and gonadal peptides, may mediate these effects.More research is necessary in order to characterize the role(s) ofthese hormones in reproduction control.
Acknowledgments
We gratefully acknowledge to the support of Dr. Yoav Gothilf.We thank all of the members of our lab that contributed to thisstudy especially to Dr. Doron Goldberg for molecular biology ad-vice and Dr. Svetlana Yom-Don for the technical assistance.
References
Adams, B.A., Lescheid, D.W., Vickers, E.D., Crim, L.W., Sherwood, N.M., 2002.Pituitary adenylate cyclase-activating polypeptide and growth hormone-releasing hormone-like peptide in sturgeon, whitefish, grayling, flounder andhalibut: cDNA sequence, exon skipping and evolution. Regulatory Peptides 109,27–37.
Apostolakis, E.M., Lanz, R., O’Malley, B.W., 2004. Pituitary adenylate cyclase-activating peptide: a pivotal modulator of steroid-induced reproductivebehavior in female rodents. Molecular Endocrinology 18, 173–183.
Arimura, A., 1992. Pituitary adenylate cyclase activating polypeptide (PACAP):discovery and current status of research. Regulatory Peptides 37, 287–303.
Arimura, A., 1998. Perspectives on pituitary adenylate cyclase activatingpolypeptide (PACAP) in the neuroendocrine, endocrine, and nervous systems.Japanese Journal of Physiology 48, 301–331.
Canosa, L.F., Stacey, N., Peter, R.E., 2008. Changes in brain mRNA levels ofgonadotropin-releasing hormone, pituitary adenylate cyclase activatingpolypeptide, and somatostatin during ovulatory luteinizing hormone andgrowth hormone surges in goldfish. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology 295, R1815–R1821.
Cardoso, J.C., Clark, M.S., Viera, F.A., Bridge, P.D., Gilles, A., Power, D.M., 2005. Thesecretin G-protein-coupled receptor family: teleost receptors. Journal ofMolecular Endocrinology 34, 753–765.
denylate cyclase-activating polypeptide and reproduction in the blue gou-

10 G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx
ARTICLE IN PRESS
Chan, K.W., Yu, K.L., Rivier, J., Chow, B.K., 1998. Identification and characterization ofa receptor from goldfish specific for a teleost growth hormone-releasinghormone-like peptide. Neuroendocrinology 68, 44–56.
Chang, J.P., Wirachowsky, N.R., Kwong, P., Johnson, J.D., 2001. PACAP stimulation ofgonadotropin-II secretion in goldfish pituitary cells: mechanisms of action andinteraction with gonadotropin releasing hormone signalling. Journal ofNeuroendocrinology 13, 540–550.
Chi-Wei, L., Chang, S.L., Weng, C.F., 2007. Pituitary adenylate cyclase-activatingpolypeptide (PACAP) regulates the expression of PACAP in cultured tilapiaastrocytes. Experimental Biology and Medicine (Maywood) 232, 262–276.
Cook, H., Berkenbosch, J.W., Fernhout, M.J., Yu, K.L., Peter, R.E., Chang, J.P., Rivier, J.E.,1991. Demonstration of gonadotropin releasing-hormone receptors ongonadotrophs and somatotrophs of the goldfish: an electron microscopestudy. Regulatory Peptides 36, 369–378.
Degani, G., 1993a. The effect of sexual behavior on oocyte development and steroidchanges in Trichogaster trichopterus. Copeia 4, 1091–1096.
Degani, G., 1993b. Reproduction control in multi-spawning asynchronicTrichogaster trichopterus (Pallas) as a model for the anabantidae family.Trends in Comparative Biochemicals and Physiology 1, 1269–1275.
Degani, G., 2001. Blue gourami (Trichogaster trichopterus) model for labyrinth fish.Laser Pages Publishing, Israel.
Degani, G., Boker, R., 1992. Sensitivity to maturation-inducing steroids andgonadotropin in the oocytes of blue gourami Trichogaster trichopterus(Anabantidae, Pallas, 1770). General and Comparative Endocrinology 85, 430–439.
Degani, G., Mananos, E.L., Jackson, K., Abraham, M., Zohar, Y., 1997. Changes inplasma and pituitary GtH-II levels in female blue gourami Trichogastertrichopterus during the end of vitellogenesis and final oocyte maturation.Journal of Experimental Zoology 279, 377–385.
Degani, G., Jackson, K., Goldberg, D., Sarfati, R., Avtalion, R.R., 2003a. BetaFSH,betaLH and growth hormone gene expression in blue gourami (Trichogastertrichopterus, Pallas 1770) during spermatogenesis and male sexual behavior.Zoological Science 20, 737–743.
Degani, G., Tzchori, I., Yom-Din, S., Goldberg, D., Jackson, K., 2003b. Growthdifferences and growth hormone expression in male and female European eels[Anguilla anguilla (L.)]. General and Comparative Endocrinology 134, 88–93.
Degani, G., Jackson, K., Yom-Din, S., Goldberg, D., 2006. CDNA cloning and mRNAexpression of growth hormone in Belontiidae (Anabantoidei suborder). IsraeliJournal of Aquaculture 58, 124–136.
Fradinger, E.A., Sherwood, N.M., 2000. Characterization of the gene encoding bothgrowth hormone-releasing hormone (GRF) and pituitary adenylate cyclase-activating polypeptide (PACAP) in the zebrafish. Molecular and CellularEndocrinology 165, 211–219.
Goldberg, D., Jackson, K., Yom-Din, S., Degani, G., 2004. Growth Hormone ofTrichogaster trichopterus: cDNA Cloning, Sequencing and Analysis of mRNAExpression during Oogenesis. Journal of Aquacultural Tropics 19, 215–229.
Goth, M.I., Lyons, C.E., Canny, B.J., Thorner, M.O., 1992. Pituitary adenylate cyclaseactivating polypeptide, growth hormone (GH)-releasing peptide and GH-releasing hormone stimulate GH release through distinct pituitary receptors.Endocrinology 130, 939–944.
Gur, G., Bonfil, D., Safarian, H., Naor, Z., Yaron, Z., 2002. Pituitary adenylate cyclaseactivating polypeptide and neuropeptide Y regulation of gonadotropin subunitgene expression in tilapia: role of PKC, PKA and ERK. Neuroendocrinology 75,164–174.
Ha, C.M., Kang, J.H., Choi, E.J., Kim, M.S., Park, J.W., Kim, Y., Choi, W.S., Chun, S.Y.,Kwon, H.B., Lee, B.J., 2000. Progesterone increases mRNA levels of pituitaryadenylate cyclase-activating polypeptide (PACAP) and type I PACAP receptor(PAC(1)) in the rat hypothalamus. Brain Research. Molecular Brain Research 78,59–68.
Hull, K.L., Harvey, S., 2001. Growth hormone: roles in female reproduction. Journalof Endocrinology 168, 1–23.
Inagaki, N., Yoshida, H., Mizuta, M., Mizuno, N., Fujii, Y., Gonoi, T., Miyazaki, J., Seino,S., 1994. Cloning and functional characterization of a third pituitary adenylatecyclase-activating polypeptide receptor subtype expressed in insulin-secretingcells. Proceedings of the National Academy of Sciences of the United States ofAmerica 91, 2679–2683.
Jackson, K., Abraham, M., Degani, G., 1994. Oocyte maturation triggered by thepresence of male in the blue gourami (Trichogaster trichopterus). Journal ofMorphology 220, 1–9.
Jackson, K., Goldberg, D., Ofir, M., Abraham, M., Degani, G., 1999. Blue gourami(Trichogaster trichopterus) gonadotropic beta subunits (I and II) cDNA sequencesand expression during oogenesis. Journal of Molecular Endocrinology 23, 177–187.
Jakobsson, S., Borg, B., Haux, C., Hyllner, S.J., 1999. An 11-ketotestosterone inducedkidney-secreted protein: the nest building glue from male three-spinedstickleback, Gasterosteus aculeatus. Fish Physiology and Biochemistry 20, 79–85.
Kiya, T., Oka, Y., 2003. Glutamate receptors in the terminal nerve gonadotropin-releasing hormone neurons of the dwarf gourami (teleost). NeuroscienceLetters 345, 113–116.
Klausen, C., Chang, J.P., Habibi, H.R., 2002. Time- and dose-related effects ofgonadotropin-releasing hormone on growth hormone and gonadotropinsubunit gene expression in the goldfish pituitary. Canadian Journal ofPhysiology and Pharmacology 80, 915–924.
Kotani, E., Usuki, S., Kubo, T., 1998. Effect of pituitary adenylate cyclase-activatingpolypeptide (PACAP) on progestin biosynthesis in cultured granulosa cells from
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
rat ovary and expression of mRNA encoding PACAP type IA receptor. Journal ofReproduction and Fertility 112, 107–114.
Lee, L.T., Siu, F.K., Tam, J.K., Lau, I.T., Wong, A.O., Lin, M.C., Vaudry, H., Chow, B.K.,2007. Discovery of growth hormone-releasing hormones and receptors innonmammalian vertebrates. Proceedings of the National Academy of Sciencesof the United States of America 104, 2133–2138.
Lin, S.W., Ge, W., 2009. Differential regulation of gonadotropins (FSH and LH) andgrowth hormone (GH) by neuroendocrine, endocrine, and paracrine factors inthe zebrafish – an in vitro approach. General and Comparative Endocrinology160, 183–193.
Marchant, T.A., Chang, J.P., Nahorniak, C.S., Peter, R.E., 1989. Evidence thatgonadotropin-releasing hormone also functions as a growth hormone-releasing factor in the goldfish. Endocrinology 124, 2509–2518.
McRory, J.E., Parker, D.B., Ngamvongchon, S., Sherwood, N.M., 1995. Sequence andexpression of cDNA for pituitary adenylate cyclase activating polypeptide(PACAP) and growth hormone-releasing hormone (GHRH)-like peptide incatfish. Molecular and Cellular Endocrinology 108, 169–177.
Melamed, P., Eliahu, N., Levavi-Sivan, B., Ofir, M., Farchi-Pisanty, O., Rentier-Delrue,F., Smal, J., Yaron, Z., Naor, Z., 1995. Hypothalamic and thyroidal regulation ofgrowth hormone in tilapia. General and Comparative Endocrinology 97, 13–30.
Melamed, P., Gur, G., Elizur, A., Rosenfeld, H., Sivan, B., Rentier-Delrue, F., Yaron, Z.,1996. Differential effects of gonadotropin-releasing hormone, dopamine andsomatostatin and their second messengers on the mRNA levels of gonadotropinII beta subunit and growth hormone in the teleost fish, tilapia.Neuroendocrinology 64, 320–328.
Montero, M., Yon, L., Rousseau, K., Arimura, A., Fournier, A., Dufour, S., Vaudry, H.,1998. Distribution, characterization, and growth hormone-releasing activity ofpituitary adenylate cyclase-activating polypeptide in the European eel, Anguillaanguilla. Endocrinology 139, 4300–4310.
Montero, M., Yon, L., Kikuyama, S., Dufour, S., Vaudry, H., 2000. Molecularevolution of the growth hormone-releasing hormone/pituitary adenylatecyclase-activating polypeptide gene family. Functional implication in theregulation of growth hormone secretion. Journal of Molecular Endocrinology25, 157–168.
Muller, P.Y., Janovjak, H., Miserez, A.R., Dobbie, Z., 2002. Processing of geneexpression data generated by quantitative real-time RT-PCR. BioTechniques 32,1372–1379.
Nagahama, Y., 1994. Endocrine regulation of gametogenesis in fish. InternationalJournal of Developmental Biology 38, 217–229.
Parhar, I.S., Iwata, M., 1994. Gonadotropin releasing hormone (GnRH) neuronsproject to growth hormone and somatolactin cells in the steelhead trout.Histochemistry 102, 195–203.
Parker, D.B., Coe, I.R., Dixon, G.H., Sherwood, N.M., 1993. Two salmon neuropeptidesencoded by one brain cDNA are structurally related to members of the glucagonsuperfamily. European Journal of Biochemistry 215, 439–448.
Parker, D.B., Power, M.E., Swanson, P., Rivier, J., Sherwood, N.M., 1997. Exonskipping in the gene encoding pituitary adenylate cyclase-activatingpolypeptide in salmon alters the expression of two hormones that stimulategrowth hormone release. Endocrinology 138, 414–423.
Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 29, e45.
Ran, X.Q., Li, W.S., Lin, H.R., 2004. Stimulatory effects of gonadotropin-releasinghormone and dopamine on growth hormone release and growth hormonemRNA expression in Epinephelus coioides. Sheng Li Xue Bao 56, 644–650.
Rossato, M., Nogara, A., Gottardello, F., Bordon, P., Foresta, C., 1997. Pituitaryadenylate cyclase activating polypeptide stimulates rat Leydig cellsteroidogenesis through a novel transduction pathway. Endocrinology 138,3228–3235.
Rousseau, K., Le Belle, N., Pichavant, K., Marchelidon, J., Chow, B.K., Boeuf, G., Dufour,S., 2001. Pituitary growth hormone secretion in the turbot, a phylogeneticallyrecent teleost, is regulated by a species-specific pattern of neuropeptides.Neuroendocrinology 74, 375–385.
Scott, A.P., MacKenzie, D.S., Stacey, N.E., 1984. Endocrine changes during naturalspawning in the white sucker, Catostomus commersoni. II. Steroid hormones.General and Comparative Endocrinology 56, 349–359.
Sherwood, N.M., Krueckl, S.L., McRory, J.E., 2000. The origin and function of thepituitary adenylate cyclase-activating polypeptide (PACAP)/glucagonsuperfamily. Endocrinological Reviews 21, 619–670.
Small, B.C., Nonneman, D., 2001. Sequence and expression of a cDNA encoding bothpituitary adenylate cyclase activating polypeptide and growth hormone-releasing hormone-like peptide in channel catfish (Ictalurus punctatus).General and Comparative Endocrinology 122, 354–363.
Sohn, Y.C., Kobayashi, M., Aida, K., 2001. Regulation of gonadotropin beta subunitgene expression by testosterone and gonadotropin-releasing hormones in thegoldfish, Carassius auratus. Comparative Biochemistry and Physiology. Part B,Biochemistry and Molecular Biology 129, 419–426.
Stacey, N.E., MacKenzie, D.S., Marchant, T.A., Kyle, A.L., Peter, R.E., 1984. Endocrinechanges during natural spawning in the white sucker, Catostomus commersoni. I.Gonadotropin, growth hormone, and thyroid hormones. General andComparative Endocrinology 56, 333–348.
Sze, K.H., Zhou, H., Yang, Y., He, M., Jiang, Y., Wong, A.O., 2007. Pituitary adenylatecyclase-activating polypeptide (PACAP) as a growth hormone (GH)-releasingfactor in grass carp: II. Solution structure of a brain-specific PACAP by nuclearmagnetic resonance spectroscopy and functional studies on GH release andgene expression. Endocrinology 148, 5042–5059.
denylate cyclase-activating polypeptide and reproduction in the blue gou-

G. Levy et al. / General and Comparative Endocrinology xxx (2009) xxx–xxx 11
ARTICLE IN PRESS
Tam, J.K., Lee, L.T., Chow, B.K., 2007. PACAP-related peptide (PRP) – molecularevolution and potential functions. Peptides 28, 1920–1929.
Taniyama, S., Kitahashi, T., Ando, H., Kaeriyama, M., Zohar, Y., Ueda, H., Urano, A.,2000. Effects of gonadotropin-releasing hormone analog on expression of genesencoding the growth hormone/prolactin/somatolactin family and a pituitary-specific transcription factor in the pituitaries of prespawning sockeye salmon.General and Comparative Endocrinology 118, 418–424.
Trudeau, V.L., Sloley, B.D., Wong, A.O., Peter, R.E., 1993. Interactions of gonadalsteroids with brain dopamine and gonadotropin-releasing hormone in thecontrol of gonadotropin-II secretion in the goldfish. General and ComparativeEndocrinology 89, 39–50.
Tsujii, T., Winters, S.J., 1995. Effects of pulsatile pituitary adenylate cyclaseactivating polypeptide (PACAP) on gonadotropin secretion and subunit mRNAlevels in perifused rat pituitary cells. Life Sciences 56, 1103–1111.
Wang, Y., Wong, A.O., Ge, W., 2003. Cloning, regulation of messenger ribonucleicacid expression, and function of a new isoform of pituitary adenylate
Please cite this article in press as: Levy, G., et al. Association between pituitary arami. Gen. Comp. Endocrinol. (2009), doi:10.1016/j.ygcen.2009.09.015
cyclase-activating polypeptide in the zebrafish ovary. Endocrinology 144,4799–4810.
Wong, A.O., Li, W.S., Lee, E.K., Leung, M.Y., Tse, L.Y., Chow, B.K., Lin, H.R., Chang, J.P.,2000. Pituitary adenylate cyclase activating polypeptide as a novelhypophysiotropic factor in fish. Biochemistry and Cell Biology 78, 329–343.
Xiao, D., Chu, M.M., Lee, E.K., Lin, H.R., Wong, A.O., 2002. Regulation of growthhormone release in common carp pituitary cells by pituitary adenylate cyclase-activating polypeptide: signal transduction involves cAMP- and calcium-dependent mechanisms. Neuroendocrinology 76, 325–338.
Yaron, Z., Gur, G., Melamed, P., Rosenfeld, H., Levavi-Sivan, B., Elizur, A., 2001.Regulation of gonadotropin subunit genes in tilapia. Comparative Biochemistryand Physiology. Part B, Biochemistry and Molecular Biology 129, 489–502.
Yaron, Z., Gur, G., Melamed, P., Rosenfeld, H., Elizur, A., Levavi-Sivan, B., 2003.Regulation of fish gonadotropins. International Review of Cytology 225, 131–185.
denylate cyclase-activating polypeptide and reproduction in the blue gou-






![Index [link.springer.com]978-3-642-38487...Index A ACA. See Adenylyl cyclase acaA. See Adenylate cyclase A ACC. See 1-aminocyclo-propane-1-carboxylic acid Acinetobacter baumannii,](https://img.pdfslide.us/doc/110x75/5b47af067f8b9af5078c45af/index-link-978-3-642-38487index-a-aca-see-adenylyl-cyclase-acaa-see-adenylate.jpg)






















