Embed Size (px)
Citation preview

Proc. Nati Acad. Sci. USAVol. 78, No. 12, pp. 7417-7421, December 1981Biochemistry
Salts promote activation of fat cell adenylate cyclase by GTP:Special role for sodium ion
(cations/anions/5'-guanylyl imidodiphosphate/nucleotide regulatory protein)
MICHAEL S. KATZ*, JOHN S. PARTILLAt, MARCO A. PI&EYROt, CHRISTINE R. SCHNEYERt, ANDROBERT I. GREGERMANt**Division of Endocrinology and Metabolism, Department of Medicine, University of Texas Health Science Center, and Medical Service, Audie L. Murphy VeteransAdministration Hospital, San Antonio, Texas 78284; and fGerontology Research Center, National Institute on Aging, National Institutes of Health at Baltimore CityHospitals, Baltimore, Maryland 21224; and MDepartment of Medicine, Baltimore City Hospitals, and Department of Medicine, Johns Hopkins University School ofMedicine, Baltimore, Maryland 21205
Communicated by Frederic C. Bartter, August 27, 1981
ABSTRACT The effects of GTP on adenylate cyclase [ATPpyrophosphate-lyase (cyclizing), EC 4.6.1.1] ofhuman and rat fatcell membranes ("ghosts" and purified membranes) were exam-ined in the absence and presence of added inorganic salts. Withhuman ghosts GTP alone (0.1 mM) inhibited enzyme activity by40% at 30°C and had no significant effect at 3rC. At both tem-peratures Na' salts of Cl1, N3-, and S042- stimulated activity(up to 4-fold basal activity for 200mM NaN3), with maximal effectsat salt concentrations of 100-200 mM. Over the same concentra-tion range these salts also allowed temperature-dependent stim-ulation by GTP. GTP increased the maximal activity produced bysalt alone by about 2-fold at 30°C and about 4-fold at 37C. Na+(added as Cl-) was much more effective than other alkali metalcations in promoting activation by GTP. Na+ salts allowed acti-vation of the human enzyme by the GTP analog 5'-guanylyl imi-dodiphosphate and also promoted stimulation of rat fat cell aden-ylate cyclase by both nucleotides. In time course studies ofhumanand rat fat cell ghosts, GTP appeared to sustain an initial high rateof salt-stimulated activity, which in the absence of nucleotide sub-sequently fell to a lower rate, suggesting that salts might activateadenylate cyclase by promoting the stimulatory effect of endog-enous membrane-bound GTP. However, with purified human fatcell membranes and a GTP-free system, salts were still stimulatoryand promoted activation by added GTP. These results differ fromthose of previous reports in other systems in which Na+ has pro-moted only inhibitory effects in GTP regulation of adenylatecyclase.Guanine nucleotides regulate the activity of mammalian aden-ylate cyclase [ATP pyrophosphate-lyase (cyclizing), EC 4.6.1.1].Activation of the enzyme by a number of agents, notably hor-mones and cholera toxin, has been suggested to occur by oneor more mechanisms that facilitate an activating effect of GTPat the nucleotide regulatory subunit of the enzyme (1-4). Flu-oride anion, a powerful activator ofenzyme activity, also appearsto affect adenylate cyclase at the regulatory subunit, but it isas yet uncertain whether activation by fluoride requires inter-action ofguanine nucleotides with the regulatory subunit (5, 6).Recently inorganic salts ofnon-fluoride anions have been shownto stimulate adenylate cyclase activity in several tissues (7-13).Although studies of liver, kidney, and pancreas preparations(9-11) have shown that inorganic salts enhance activation ofadenylate cyclase by the synthetic GTP analog 5'-guanylyl im-idodiphosphate (GMP-P[NH]P), the mechanism ofenzyme ac-tivation by salts, and specifically the interaction ofions with thenaturally occurring stimulatory nucleotide GTP, remains spec-ulative (14).
In our own studies of human and rat fat cell membranes("ghosts"), we have found that a number of non-fluoride inor-ganic salts stimulate fat cell adenylate cyclase activity. More-over, fat cell ghosts have proven to be a convenient system inwhich to study salt interaction with GTP. GTP added alone tofat cell preparations has generally been found to be inhibitory,although under some assay conditions the nucleotide has slightstimulatory effect (14-24). In the human system we showed thatGTP assumes a stimulatory role only in the presence ofhormone(25); similarly, in the rat stimulation by GTP has for the mostpart required hormone, special assay conditions, or both (14,16, 17, 20, 22, 24, 26). On the other hand, we now demonstratethat in both human and rat fat cell preparations GTP becomesmarkedly stimulatory in the presence of inorganic salts thatthemselves activate adenylate cyclase. In examining the effectsof various salts, it has become apparent that the sodium ion hasa relatively specific enhancing effect on GTP action. In thispaper we have examined the effects of salts and GTP primarilyin human fat cell ghosts, in which salt-induced GTP activationof adenylate cyclase is more marked than in ghosts preparedfrom rat fat cells. Similar results were obtained with purifiedhuman membranes.
METHODSFat cell ghosts were prepared from human subcutaneous fat andrat epididymal fat pads as described (25). Some studies (seeResults) used plasma membranes prepared from human fat cellsas described for the rat by Harwood et aL (16). Adenylate cyclaseactivity was assayed in a total volume of50 ,ul as described (25),and cyclic AMP product was isolated by the method of Salomonet al. (27). Total Na+ concentration under routine assay condi-tions was 4 mM, and total Cl- was 18 mM. As noted in the ear-lier studies (16, 25), membranes but not ghosts were preparedin the presence of 1 mM EDTA, whereas no EDTA was addedto the enzyme assay mixture.
RESULTSGTP added alone to human fat cell ghosts inhibited adenylatecyclase activity at 30°C and had no effect at 37°C. At 30°C basalenzyme activity of 0.48 ± 0.05 (SEM) nmol of cyclic-AMP permg ofprotein per 10 min was inhibited by GTP (0.1 mM) to 0.29± 0.04 (P < 0.001 by paired Student's t test; n = 28). At 37°Cbasal activity was 0.43 ± 0.06 and was not significantly affected
Abbreviation: GMP-P[NH]P, 5'-guanylyl imidodiphosphate.*To whom reprint requests should be addressed at the GerontologyResearch Center, NIA, NIH, Baltimore City Hospitals, Baltimore,MD 21224.
7417
The publication costs ofthis article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 78 (1981)
by added GTP (0.39 ± 0.05; n = 17). Individual ghost prepa-rations varied to the extent that they occasionally showed someinhibition by GTP at 370C or no inhibition at 30'C. Neverthe-less, these results provide statistical evidence that GTP inhi-bition ofenzyme activity is temperature dependent. Moreover,GTP alone was clearly not stimulatory at any assay condition orany concentration of added GTP tested.
In the presence ofadded inorganic salts, however, GTP stim-ulated adenylate cyclase at both 30'C and 370C. Fig. 1 showsthe effects on adenylate cyclase activity of increasing concen-trations of Na+ salts ofCl- and N3- in the absence and presenceof GTP. Similar effects were seen with Na' salts of S042- andHP042- (data not shown). The magnitude of GTP effect in thepresence of salts was partially dependent on the anion species.
Fig. 1 also shows that the magnitude of GTP activation ofadenylate cyclase in the presence of salts is clearly temperaturedependent. For all three Na' salts (Cl-, N3-, and S042-) testedat 30'C and 370C, activation by GTP was much greater at 370Cthan at 30'C even though stimulation by salts alone was essen-tially the same at both temperatures or even slightly less at 370Cthan at 30'C. At maximally effective concentrations ofeach salt,activation by GTP at 370C (expressed as increase of activityabove that elicited by salt alone) was more than twice that seenat 300C.
Our usual assay system undoubtedly contained small butpossibly significant amounts of GTP, in part adherent to the fatcell ghosts and in part as a contaminant of the ATP substrateordinarily used (28). It seemed conceivable that this endoge-nous GTP might have contributed to or otherwise altered thestimulatory effects of exogenous GTP in the presence of salts.Therefore, the effects of exogenous GTP and Na+ salts wererestudied in the presence of substantially reduced GTP con-taminants-i.e., in a "GTP-free" system using purified humanfat cell plasma membranes rather than ghosts and using ATPpresumably free of GTP. This ATP (Sigma 2383) was preparedby the phosphorylation ofadenosine. The results with NaCl andNaN3 were very similar to those obtained by using the routineassay system (Fig. 1), making it unlikely that endogenous GTPcontamination contributed substantially to the GTP effects seenin the presence of Na+ salts.
Fig. 2 shows the time courses ofthe effects ofGTP and NaCl.In this experiment GTP alone was mildly inhibitory at 37°C overthe entire 10-min incubation although less so than at 300C. The
2-
0--
0M,
E
CZ-
C-,
EC)
z 1.2 I. -
/'GTP+NaCI0- 1.0 /0cm / GTP+NaCIE O~~//0.8~ ~ ~
C-,_-a O/~~~~~~~
E 0.4 .
BASAL /0.2 *..-/*..BASAL
IGTP GTP
0 2 4 6 8 10 0 2 4 6 8 10
MINUTES
FIG. 2. Time course of GTP activation of human fat cell adenylatecyclase in the presence of NaCl. A fat cell ghost preparation was in-cubated at 300C (A) and 370C (B). Concentrations of GTP, 0.1 mM;NaCl, 100 mM. Each point represents a single determination.
time course of stimulation by NaCl alone was nonlinear at bothtemperatures; i.e., the initial high rate of activity decreasedwith time. The addition ofGTP to NaCl appeared to sustain theinitial high rate of activity produced by salt alone, resulting inlinear time courses to 10 min at both temperatures. The ap-parent magnitude ofGTP effect was therefore time dependent.In this respect "stimulation" or "activation" by GTP could beviewed as the result of fall-off of activity in the presence of saltalone with preservation by GTP of the initial rate of salt-stim-ulated activity. This interpretation ofthe results seems strongerat 300C; at 370C the curve with NaCl alone is probably also non-linear even during the first few minutes, but enhancement byGTP is apparent from the first point measured. When purifiedhuman membranes were used, stimulation at 37°C by NaClalone was linear for at least the first 8 min and clear further stim-ulation by GTP was similar to that with ghosts.We also examined the effect of salts on GMP-P[NH]P-mod-
m
C=U.02C-
0-
EJ
C-
C-,
Cl-
ECL
100 200 300 400 100 200 300 400
[SALT]ImM)FIG. 1. GTP activation of human fat cell adenylate cyclase in the
presence of NaCl or NaN3. Fat cell ghost preparations were incubatedat 3000 (-) and 3700 (----) in the absence (o, o) and presence (o,o) of 0.1 mM GTP. NaCl and NaN3 effects were determined in one ex-
periment at 3000 and 3700, using a single ghost preparation. Eachpoint is the mean of duplicate determinations.
NaCI7
6
5 _ -
+GMP-PINH)P..4
d3
2-
1 .
I
I
NaN3
/+GMP-FPNHIF
i
I
(1.~~~~~~~
0 200 400 80 0 200 400 600
[SALT] ImM)
FIG. 3. GMP-P[NH1P activation of human fat cell adenylate cy-clase in the presence of NaCl and NaN3. Fat cell ghosts were incubatedat 30°0 in the absence (o) and presence (o) of 0.1 mM GMP-P[NH1P.Each point is the mean of duplicate determinations. In a separate ex-periment stimulation by GMP-P[NH]P in the presence of NaCl wasfound to be maximal at 600 mM salt.
I
-j I1
7418 Biochemistry: Katz et al.
I
Proc. Nati Acad. Sci. USA 78 (1981) 7419
+GTP~~~~~~~~~~~+T
0 20 0 01S 0 6D s 0 0 0 O 9
0~~~~~~~~+G
LGTP
0 200 000 2004O a 005
[CtSALT] ImMI
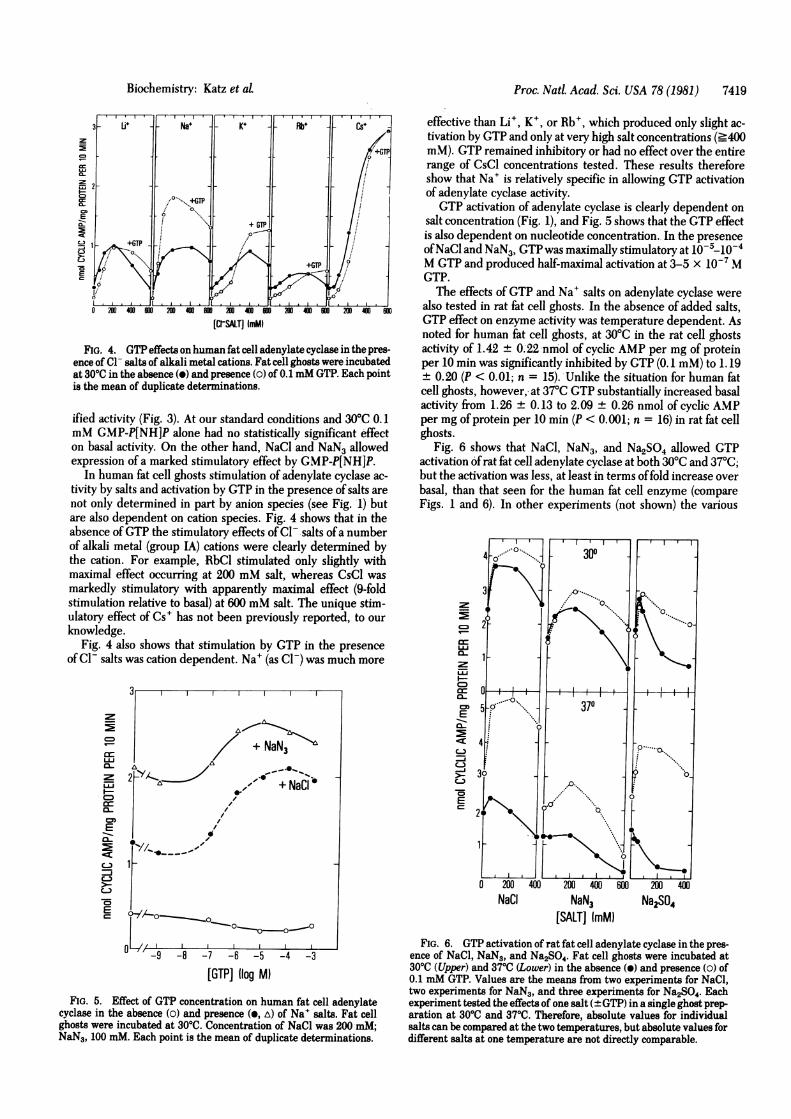
FIG. 4. GTP effects on human fat cell adenylate cyclase in the pres-ence of Cl- salts of alkali metal cations. Fat cell ghosts were incubatedat 3000 in the absence (W) and presence (o) of 0.1 mM GTP. Each pointis the mean of duplicate determinations.
ified activity (Fig. 3). At our standard conditions and 30°C 0.1mM GMP-P[NH]P alone had no statistically significant effecton basal activity. On the other hand, NaCl and NaN3 allowedexpression of a marked stimulatory effect by GMP-P[NH]P.
In human fat cell ghosts stimulation of adenylate cyclase ac-tivity by salts and activation by GTP in the presence of salts arenot only determined in part by anion species (see Fig. 1) butare also dependent on cation species. Fig. 4 shows that in theabsence ofGTP the stimulatory effects of Cl- salts ofa numberof alkali metal (group IA) cations were clearly determined bythe cation. For example, RbCl stimulated only slightly withmaximal effect occurring at 200 mM salt, whereas CsCl wasmarkedly stimulatory with apparently maximal effect (9-foldstimulation relative to basal) at 600 mM salt. The unique stim-ulatory effect of Cs' has not been previously reported, to ourknowledge.
Fig. 4 also shows that stimulation by GTP in the presenceofCl salts was cation dependent. Na+ (as Cl-) was much more
T- ~~~~~+NaN3LU~
2-
O /s'
E
E
0
-9 -8 -7 -6 -5 -4 -3
[GTP] (log M)
FIG. 5. Effect of GTP concentration on human fat cell adenylatecyclase in the absence (o) and presence (e, A) of Na+ salts. Fat cellghosts were incubated at 3000. Concentration of NaCl was 200 mM;NaN3, 100 mM. Each point is the mean of duplicate determinations.
effective than Li', K+, or RbV, which produced only slight ac-tivation by GTP and only at very high salt concentrations (-'400mM). GTP remained inhibitory or had no effect over the entirerange of CsCl concentrations tested. These results thereforeshow that Na' is relatively specific in allowing GTP activationof adenylate cyclase activity.GTP activation of adenylate cyclase is clearly dependent on
salt concentration (Fig. 1), and Fig. 5 shows that the GTP effectis also dependent on nucleotide concentration. In the presenceofNaCl and NaN3, GTP was maximally stimulatory at 10-L0-4M GTP and produced half-maximal activation at 3-5 X 10-7 MGTP.The effects of GTP and Na' salts on adenylate cyclase were
also tested in rat fat cell ghosts. In the absence of added salts,GTP effect on enzyme activity was temperature dependent. Asnoted for human fat cell ghosts, at 30TC in the rat cell ghostsactivity of 1.42 ± 0.22 nmol of cyclic AMP per mg of proteinper 10 min was significantly inhibited by GTP (0.1 mM) to 1.19+ 0.20 (P < 0.01; n = 15). Unlike the situation for human fatcell ghosts, however, at 37TC GTP substantially increased basalactivity from 1.26 ± 0.13 to 2.09 ± 0.26 nmol of cyclic AMPper mg of protein per 10 min (P < 0.001; n = 16) in rat fat cellghosts.
Fig. 6 shows that NaCl, NaN3, and Na2SO4 allowed GTPactivation ofrat fat cell adenylate cyclase at both 30°C and 37°C;but the activation was less, at least in terms offold increase overbasal, than that seen for the human fat cell enzyme (compareFigs. 1 and 6). In other experiments (not shown) the various
4 ci 300
3
LAJ~~~~~~~~~~~~O
C" 50 370E
0
0~~~~~
1 p~~~~c
0 200 400 200 400 600 200 400NaCI NaN3 Na2SO4
[SALT] (mM)
FIG. 6. GTP activation of rat fat cell adenylate cyclase in the pres-ence of NaCl, NaN3, and Na2SO4. Fat cell ghosts were incubated at3000 (Upper) and 3700 (Lower) in the absence (.) and presence (o) of0.1 mM GTP. Values are the means from two experiments for NaCl,two experiments for NaN3, and three experiments for Na2SO4. Eachexperiment tested the effects of one salt (±GTP) in a single ghost prep-aration at 30°C and 370C. Therefore, absolute values for individualsalts can be compared at the two temperatures, but absolute values fordifferent salts at one temperature are not directly comparable.
Biochemistry: Katz et aL

Proc. Natl. Acad. Sci. USA 78 (1981)
characteristics ofsalt-induced GTP activation ofrat fat cell aden-ylate cyclase were qualitatively similar to those described herefor human fat cell ghosts.
DISCUSSIONWe and others have discussed possible mechanisms ofadenylatecyclase stimulation by salts (10, 11, 14, 29, 30). Our presentresults exclude a number of mechanisms by which Na' saltsmight be stimulatory and allow activation by GTP. The relativespecificity of Na' (Fig. 4) makes it unlikely that salt effects onGTP action are mediated by a nonspecific alteration of the ionicstrength of the assay medium. Guanine nucleotides were in-hibitory in the presence of glycerol (100-500 mM) and stimu-latory concentrations ofF- (data not shown), indicating that Na+salt-induced activation of adenylate cyclase by GTP does notresult from a nonspecific solute effect and appears to be a dif-ferent process from that by which F- stimulates enzyme activ-ity. Ions are known to affect nucleotidase activity (31, 32) as wellas creatine kinase activity (33, 34). However, Na+ salt effect onactivation by GTP is probably not due to "GTP-sparing" at thenucleotide regulatory subunit in the presence of salts becausesalts also elicited stimulation by the GTP analog GMP-P[NH]P(Fig. 3), which is resistant to hydrolysis at the regulatory subunit(35). Furthermore, Na+ salts do not appear to act by sparingGTP in the incubation medium, because others have shownunder conditions similar to our own that ATP or an ATP-re-generating system prevents the bulk hydrolysis of GTP by fatcell membranes (35).
Recent studies of adenylate cyclase in purified plasma mem-branes from rat fat cells demonstrating effects ofGTP and aden-osine suggest possible mechanisms by which Na+ salts maystimulate enzyme activity and allow activation by GTP. Cooperet al. (24) found that increasing the concentration of GTP stim-ulated enzyme activity of rat fat cell membranes up to a peakactivity, and even higher concentration ofGTP caused a decline,or "inhibition," ofactivity from this peak value. It is conceivablethat our fat cell ghost preparations, which might contain moreendogenous GTP than does a plasma membrane preparation,may in fact contain high, inhibitory, concentrations of nucleo-tide. In this case salts might "stimulate" enzyme activity andallow "activation by GTP" by eliminating the inhibitory phaseof GTP action while maintaining its stimulatory phase. How-ever, unlike the purified rat membranes used by Cooper et aL,neither the human fat cell ghosts nor the purified human mem-branes used in the present study exhibited concentration-de-pendent biphasic stimulation and inhibition ofadenylate cyclaseby GTP. Whether the difference between the results with therat and human membranes represents some varying experi-mental condition or is a biological (species) difference cannotbe stated with certainty, although we favor the latter explanation.
Adenosine has been found to be a potentially important in-hibitor of adenylate cyclase activity. In rat fat cell membranesadenosine analogs seem to produce inhibition by two distinctprocesses, one antagonized competitively by methylxanthinesand dependent on the presence of "inhibitory" concentrationsof GTP (see above), the other unaffected by methylxanthinesand independent of GTP concentration (24, 36). In our systemNa+ salt effects do not appear to be explained by simple antag-onism of either inhibitory action of adenosine for two reasons:first, 10mM theophylline was always included in our incubationmixture and, second, Na+ salt-induced enzyme activation byGTP was dependent on GTP concentration (Fig. 5).The time courses of Na+ salt- and GTP-stimulated adenylate
cyclase activities may give a clue to the mechanism of Na+ ac-tion. Fig. 2 indicates that at 30'C added GTP appeared to pre-serve the initial rate of salt stimulation, which in the absence
of nucleotide subsequently fell to a lower rate approaching thatof basal activity. The results were similar at 37TC, although theinitial rate was greater in the presence of GTP. The nonlineartime course for Na' salt alone in Fig. 2 may imply time-de-pendent depletion of GTP. Preservation of the initial high rateof salt stimulation by added GTP is consistent with replacementofthe endogenous GTP, through which salt might exert its stim-ulatory effect and which is otherwise functionally lost over timeduring salt stimulation. Functional loss of GTP during its ac-tivation ofadenylate cyclase conceivably occurs even undercon-ditions in which GTP levels in the assay medium remain con-stant (i.e., the presence of regenerating system and ATP, seeabove). For example, as has been suggested previously (2, 4,35), the GTP bound, presumably to the regulatory subunit ofthe enzyme, might be rapidly degraded to GDP or another in-active metabolite that dissociates slowly from the binding site.In this case relatively large amounts of added GTP might berequired to maintain an equilibrium favoring active GTP inproximity to its presumed binding site. We have in fact foundthat GTP activates with half-maximal effect at 0.3-0.5 AM.
The time course observations are thus consistent with theidea that Na' stimulates adenylate cyclase by allowing expres-sion of the stimulatory effect of endogenous membrane-boundGTP. However, this notion may be simplistic. Enzyme activityin the presence of GTP alone exhibits a more nearly linear ratethan that of basal activity (Fig. 2), suggesting a "linearization"of the time course by GTP that is independent of monovalentcations. Previous studies ofadenylate cyclase ofrat fatcell mem-branes have concluded that nonlinear time courses representinterconversion between active and inactive forms of the en-zyme and that this interconversion is regulated in a complexfashion by multiple factors (hormones, nucleotides, and cations)(37).
Na' salts also stimulated enzyme activity under conditionsof reduced endogenous GTP-i.e., in purified human fat cellplasma membranes assayed with ATP free ofGTP. It is probablethat these manipulations of the usual assay conditions do notremove all endogenous GTP from the membrane-bound frac-tion. Even in this "GTP-free" assay system, therefore, Na' saltscould act as in our routine assay system to stimulate adenylatecyclase by promoting the stimulatory effect of membrane-bound GTP. On the other hand, enzyme stimulation by Na'was linear for a longer period in membranes than in ghosts (seeResults and Fig. 2). If membranes in fact contain less endoge-nous GTP than ghosts do, this result suggests that Na+ activatesadenylate cyclase by a mechanism other than promoting stim-ulation by membrane-bound GTP. Resolution of this issue mayrequire direct measurement of endogenous GTP in ghosts andpurified membranes.Our present studies raise the question ofhow Na+ salt effects
bear upon stimulation ofadenylate cyclase by hormones, whichare also thought to allow GTP to exert an activating effect at thenucleotide regulatory subunit (4). Our own work with adenylatecyclase of rat liver has suggested an interaction of salt and hor-mone action (29). Not only was it shown that lower concentra-tions of Na+ salt were required for enzyme stimulation in thepresence of glucagon than in its absence, but also that saltsclearly enhanced hormone action. On the other hand, while aninteraction of salt and hormone effects is clear, Na+ and hor-mone probably do not activate the fat cell enzyme by preciselythe same GTP-mediated mechanism, because the time coursesof Na+ and hormone effects are strikingly different. In humanfat cell ghosts, epinephrine preserves the initial high rate ofbasal activity, whereas true hormone stimulation over the initialbasal rate occurs only in the presence of GTP (19, 25). Theseeffects of hormone are clearly different from those of Na' in the
7420 Biochemistry: Katz et aL

Proc. NatL Acad. Sci. USA 78 (1981) 7421
absence and presence ofGTP as described in the present work(see Fig. 2 and above). In preliminary experiments we haveexamined the effects ofNa' plus GTP on epinephrine activationof adenylate cyclase of human fat. Low concentrations of Na'enhanced GTP activation of hormone action, but activity stim--ulated by high concentrations of both GTP and Na' decreasedwhen hormone was added. On the other hand, we have pre-viously reported that in rat fat epinephrine-stimulated activityin the presence ofGTP is unaffected by Na', at least under thelimited conditions tested (14).
Interpretation of interactions of salts, GTP, and hormoneshould be made in the context oftwo other sets ofobservations.In several systems both Na' and GTP are involved in the reg-ulation of hormone binding to receptors (38). Of more directrelevance to our results is evidence that hormone inhibition ofadenylate cyclase from multiple tissues requires Na' and GTP(39, 40). The above-mentioned inhibition of adenylate cyclaseby epinephrine (at high Na' and GTP concentrations) in humanbut not rat fat cell ghosts may reflect a Na+- and GTP-depen-dent interaction ofepinephrine with an inhibitory a-adrenergicmembrane receptor in the human tissue that appears to be ab-sent in the rat. Recent studies have in fact demonstrated an a-adrenergic receptor negatively coupled to the adenylate cyclaseof human fat cell membranes (41). Further work on the inter-action ofNa+, GTP, and hormone in human fat cell membraneswill have to take into account the opposing effects of a- and 3-adrenergic interactions with GTP and the mixed function ofepinephrine.
Thus, while others have reported on Na+ involvement inGTP-regulated inhibitory effects on adenylate cyclase (39, 40),we present here clear evidence of a role for Na+ in modulatingthe stimulatory function of GTP. Recent work with EDTA-treated plasma membranes of rat liver has also briefly shownthat monovalent cations reverse GTP inhibition of basal aden-ylate cyclase activity and promote activation by the nucleotide(39). Unlike our results with fat cell membrane preparations,in these liver membranes Li+ is a more potent stimulator ofenzyme activity (in the presence of GTP) than is Na+. In thefat cell system the modulation of nucleotide effect by Na+ ap-pears to differ from that produced by stimulatory hormone (seeabove) and, from a practical viewpoint, may provide a usefulprobe of the interaction between GTP and the nucleotide reg-ulatory subunit or between the latter and the catalytic unit.
Note Added in Proof. We are now aware of recent work which dem-onstrates that inorganic salts activate solubilized brain adenylate cyclaseand enhance the stimulatory effects ofGTPanalogs in that system. Theeffect of GTP was minimal in both the absence and presence of salts(42).
We thank the surgical staff of the Baltimore City Hospitals, and es-pecially Drs. Robert Castadot and Courtney J. Robinson, for their as-sistance in obtaining the human fat specimens used in these studies.Ms. Jerrie Coby provided skillful secretarial assistance in the prepa-ration of the manuscript.
1. Rodbell, M., Lin, M. C. & Salomon, Y. (1974)J. BioL Chem. 249,59-65.
2. Sevilla, N., Steer, M. L. & Levitski, A. (1976) Biochemistry 15,3493-3499.
3. Cassel, D. & Selinger, Z. (1977) Proc. Nati Acad. Sci. USA 74,3307-3311.
4. Cassel, D. & Selinger, Z. (1978) Proc. Nati Acad. Sci. USA 75,4155-4159.
5. Eckstein, F., Cassel, D., Levkovitz, H., Lowe, M. & Selinger,Z. (1979)J. Biot Chem. 254, 9829-9834.
6. Downs, R. W., Jr., Spiegel, A. M., Singer, M., Reen, S. & Aur-bach, G. D. (1980)1. BioL Chem. 255, 949-954.
7. Kalish, M. I., Pinieyro, M. A., Cooper, B. & Gregerman, R. I.(1974) Biochem. Biophys. Res. Commun. 61, 781-787.
8. Rahmanian, M. & Jarett, L. (1974) Biochem. Biophys. Res. Com-mun. 61, 1051-1056.
9. Johnson, R. A., Pilkis, S. J. & Hamet, P. (1975) J. BioL Chem.250, 6599-6607.
10. Roy, C., LeBars, N.-C. & Jard, R. (1977) Eur. J. Biochem. 78,325-332.
11. Svoboda, M. & Christophe, J. (1978) FEBS Lett. 86, 230-234.12. Schwabe, U., Puchstein, C., Hannemann, H. & Sochtig, E.
(1979) Nature (London) 277, 143-145.13. Richards, J. M. & Swislocki, N. I. (1979) J. Biot Chem. 254,
6857-6860.14. Katz, M. S., Partilla, J. S., Pinieyro, M. A. & Gregerman, R. I.
(1980) Biochim. Biophys. Acta 613, 229-237.15. Cryer, P. E., Jarett, L. & Kipnis, D. M. (1969) Biochim. Biophys.
Acta 177, 586-590.16. Harwood, J. P., Low, H. & Rodbell, M. (1973) J. BioL Chem.
248, 6239-6245.17. Ebert, R. & Schwabe, U. (1974) Naunyn-Schmiedeberg's Arch.
Pharmacot 286, 297-313.18. Poupon, R. (1975) Biomedicine 23, 438-442.19. Cooper, B., Partilla, J. S. & Gregerman, R. I. (1976) Biochim.
Biophys. Acta 445, 246-258.20. Pairault, J. (1976) Eur. J. Biochem. 62, 323-334.21. Kather, H. & Geiger, M. (1977) Eur. J. Clin. Invest. 7, 363-371.22. Yamamura, H., Lad, P. M. & Rodbell, M. (1977)J. BioL Chem.
252, 7964-7966.23. Kather, H., Z6llig, K. & Simon, B. (1978) Horm. Metab. Res. 10,
46-49.24. Cooper, D. M. F., Schlegel, W., Lin, M. C. & Rodbell, M.
(1979)J. BioL Chem. 254, 8927-8931.25. Katz, M. S., Partilla, J. S., Pifneyro, M. A. & Gregerman, R. I.
(1981)J. Lipid Res. 22, 113-121.26. Rodbell, M. (1975)J. BioL Chem. 250, 5826-5834.27. Salomon, Y., Londos, C. & Rodbell, M. (1974) AnaL Biochem.
58, 541-548.28. Kimura, N. & Nagata, N. (1977) J. BioL Chem. 252, 3829-3835.29. Katz, M. S., Kelly, T. M., Pifneyro, M. A. & Gregerman, R. I.
(1980) Biochim. Biophys. Acta 632, 11-25.30. Wolff, J. & Cook, G. H. (1975) J. BioL Chem. 250, 6897-6903.31. Ebel, R. I. & Lardy, H. A. (1975)J. BioL Chem. 250, 191-196.32. Izutsu, K. T., Siegel, I. A. & Brisson, D. L. (1974) Biochim. Bio-
phys. Acta 373, 361-368.33. Chegwidden, W. R. & Watts, D. C. (1975) Biochim. Biophys.
Acta 410, 99-114.34. Morin, L. G. (1977) Clin. Chem. 23, 1569-1575.35. Salomon, Y. & Rodbell, M. (1975)J. BioL Chem. 250, 7245-7250.36. Londos, C., Cooper, D. M. F., Schlegel, W. & Rodbell, M.
(1978) Proc. Nati Acad. Sci. USA. 75, 5362-5366.37. Torres, H. N., Flawia, M. M., Medrano, J. A. & Cuatrecasas, P.
(1978) 1. Membr. BioL 43, 19-69.38. U'Prichard, D. C., Bylund, D. B. & Snyder, S. H. (1978)J. BioL
Chem. 253, 5090-5102.39. Jard, S., Cantau, B. & Jakobs, K. H. (1981) J. BioL Chem. 256,
2603-2606.40. Blume, A. J., Lichtshtein, D. & Boone, G. (1979) Proc. NatL
Acad. Sci. USA 76, 5626-5630.41. Bums, T. W., Langley, P. E., Terry, B. E., Byland, D. B., Hoff-
man, B. B., Tharp, M. D., Lefkowitz, R. J., Garcia-Sainz, A. &Fain, J. N. (1981)J. Clin. Invest. 67, 467-475.
42. Neer, E. J. & Salter, R. S. (1981)J. BioL Chem. 256, 5497-5503.
Biochemistry: Katz et aL





























