Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Jan. 1989, p. 62-700019-9567/89/010062-09$02.00/0Copyright © 1989, American Society for Microbiology
Outbreaks of Pneumocystis carinii Pneumonia in Coloniesof Immunodeficient Mice
PETER D. WALZER,12.3* C. KURTIS KIM,"3 MICHAEL J. LINKE,"2'3 CHERYL L. POGUE,"2MICHAEL J. HUERKAMP,4 CLARENCE E. CHRISP,4 ANTHONY V. LERRO,5
SALLY K. WIXSON,6 ELAINE HALL,7 AND LEONARD D. SHULTZ7
Cincinnati VA Medical Center,' and Division of Infectious Diseases, Department of Internal Medicine,2 and DepartmentofPathology and Laboratory Medicine,3 University of Cincinnati College of Medicine, Cincinnati, Ohio 45220;
Unit for Laboratory Medicine, University of Michigan Medical School, Ann Arbor, Michigan 481094;Institute for Cancer Research, Fox Chase Cancer Center, Philadelphia, Pennsylvania 191115;
The Department of Laboratory Animal Science, Smith Kline & French Laboratories,Philadelphia, Pennsylvania 191016; and The Jackson Laboratory, Bar Harbor, Maine 046097
Received 1 July 1988/Accepted 19 September 1988
Outbreaks of Pneumocystis carinii pneumonia occurred in colonies of nulnu and scidlscid mice at fourdifferent institutions. The disease, which was characterized by chronic wasting and respiratory insufficiency,was more severe in older mice and in animals housed in cages with special protective tops. Histopathologicfeatures included alveolar filling with the typical foamy honeycomb material and a mild, nonspecific hostinflammatory response. Immunofluorescence and immunoblotting studies suggested the P. carinni isolate was ofmouse rather than of rat or human origin, and the outbreaks could be related to each other by common vendoror source of breeding animals. Once P. carinii became established in a mouse colony, the organism tended topersist for long periods of time. The principal control measure was depopulation of the colony, although limitedexperience with the administration of trimethoprim-sulfamethoxazole was encouraging. Thus, outbreaks ofpneumocystosis are a serious problem among colonies of immunodeficient mice, with important implicationsfor the use of these animals in biomedical research. Data obtained by studying these outbreaks should enhanceunderstanding of the pathogenesis of P. carinii pneumonia and be helpful in formulating improved methods ofdetection and control.
Pneumocystis carinii is an important opportunistic pulmo-nary pathogen. The organism exists widely in nature as asaprophyte in the lungs of humans and a variety of animals;while isolates from these sources are morphologically iden-tical, antigenic studies suggest that species and strain differ-ences exist (9, 20, 34, 39). Among humans, P. carinii causespneumonia in premature, malnourished infants; childrenwith primary immunodeficiency diseases (particularly severecombined immunodeficiency disease); patients receivingcorticosteroids or other immunosuppressive drugs; and per-sons with the acquired immunodeficiency syndrome (AIDS)(26, 35).The specific host immune defects which predispose to the
development of pneumocystosis are poorly understood. An-imal challenge studies involving exogenous P. carinii havebeen severely hampered by the low virulence of the organ-ism (2, 3, 27, 36). Rats, mice, and other animals, whenadministered corticosteroids, spontaneously develop P. ca-rinii pneumonia with histologic features closely resemblingthe human form of the disease by a process which involvesreactivation of latent infection (3, 27-29, 37, 38, 46). Thismodel system has been widely used to study P. carinii buthas not permitted direct analysis of the relationship betweenthe organism and host immune function. Sporadic cases ofpneumocystosis have been reported in aniamls with sus-
pected or proven immunodeficiency disorders, and a fewoutbreaks of the disease have occurred in colonies of athy-mic (nude) mice (31, 32, 45). Such "experiments of nature"provide unusual opportunities to study the epidemiology andpathogenesis of P. carinji pneumonia.
* Corresponding author.
The Cincinnati VA Medical Center has had a long-terminterest in clinical and experimental P. carinii infection.Over the past several years we have had the opportunity toparticipate in the investigation of large outbreaks of pneu-mocystosis in colonies of immunodeficient mice at fourdifferent institutions. In the current study we present theclinical and pathological features of the disease, antigeniccharacteristics of the P. carinii isolates, host serologicresponses, control measures, and the potential impact ofthese outbreaks on the use of immunodeficient mice inbiomedical research.
MATERIALS AND METHODS
Immunodeficient mice. The outbreaks of pneumocystosisoccurred in colonies of immunodeficient mice at the follow-ing institutions: (i) Crl:CDI (ICR) nulnu mice obtained froma commercial vendor and housed at the University of Mich-igan, Ann Arbor; (ii) BALB/c nulnu and C B-17/Icr scidlscidmice bred and housed at the Institute for Cancer Research,Fox Chase Cancer Center, Philadelphia, Pa. (the scidlscidmice are lymphopenic, lacking both B and T cells, and serveas a model of severe combined immunodeficiency disease inhumans [1]); (iii) Crl:CDI (ICR) nulnu mice obtained fromthe same commercial vendor as those at the University ofMichigan and housed at Smith Kline & French Laboratories,Philadelphia, Pa.; and (iv) C B-17/IcrJ scidlscid mice bredand housed in the Research Animal Facility at The JacksonLaboratory, Bar Harbor, Maine. The original breeding pairsof these mice were obtained from the Fox Chase animalcolony.
Investigators at each institution assumed primary respon-sibility for evaluating their own outbreak of pneumocystosis.
62
Vol. 57, No. 1
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

OUTBREAKS OF P. CARINII 63
Systematic steps were taken to define the clinical features ofthe illness, etiology, and extent of involvement of the animalpopulation. Procedures included autopsies, histopathologicexamination, cultures, and serologic studies. After docu-mented cases of P. carinii pneumonia were found, one of us(P.D.W.) was contacted about further studies. The timing ofthis contact in relation to the stage of the outbreak had amajor impact on the number of animal specimens availablefor investigation. In general, attempts were made to obtainfixed lung specimens for histologic examination, frozenlungs for immunofluorescence staining and immunoblottinganalysis, and serum specimens for the determination ofantibody titers to P. carinii.
Histopathology. Histopathologic examination of the lungsof the mice was performed by the veterinary pathologist ateach institution. Specimens were also requested from theinstitutions so that the histologic features of pneumocystosisin the different outbreaks would be compared by a singlepathologist (C.K.K.). Samples included already stainedslides as well as frozen and fixed sections. Since freezingartifacts made detailed histologic examination difficult, thefrozen lung sections were usually not included in the dataanalysis. To prepare permanent specimens, lung tissueswere fixed in 4% buffered formaldehyde, embedded inparaffin, sectioned at 4 to 6 ,um, and stained with hematox-ylin-eosin and Grocott methenamine silver stains. Speci-mens from 31 mice (11 nude and 20 scid mice) were exam-ined in detail. The severity of pneumocystosis was gradedusing a previously described scoring system (19, 37, 38) withminor modification: light infection (<25% alveoli involved);moderate infection (25 to 50% alveoli involved); heavyinfection (>50% alveoli involved).
Immunofluorescence. Immunofluorescence studies wereperformed on infected lungs in an attempt to determine thesource of the outbreaks (i.e., whether the P. carinii orga-nisms were of mouse, rat, or human origin). Lung specimenssent to us in the early stages of the study varied considerablyin the quality and quantity of material, storage conditions,method of preparation, and intensity of P. carinii infection.Later, a more uniform protocol was devised in which inves-tigators usually sacrificed the mice, removed and froze thelungs, and shipped them to us on dry ice; in a few instances,live mice were shipped directly to us to provide fresh lungs.The lung samples were studied by an indirect fluorescent-
antibody (IFA) technique as described previously (39, 40,42), with slight alteration. Specimens were prepared foranalysis either as frozen sections or as a preparation homog-enized in a Stomacher (Tekmar, Inc., Cincinnati, Ohio),added to the wells of a glass slide, air dried, and heat oracetone fixed. Lung specimens from immunosuppressednormal mice, rats, and humans with documented P. carinjipneumonia, prepared in a similar manner, served as positivecontrols. The presence of P. carinii was documented infrozen sections by methenamine silver stains and in lunghomogenates by the cresyl echt violet stain.
Reagents used in the IFA technique were prepared ac-cording to procedures outlined in earlier studies (25, 34, 41).Pooled serum specimens from different groups of normalmice with prolonged environmental exposure to mouse P.carinii served as the source of primary antibody to mouse-derived organisms. These specimens had good antibodytiters to mouse P. carinji but showed little or no reactivity(titer, <1:4) to rat or human P. carinii. Rabbits immunizedwith rat P. carinii obtained from infected lungs or grown intissue culture, and with human P. carinii from infectedlungs, served as the source of primary antibody for these
studies; the rabbit antisera were adsorbed with normal lungor uninfected tissue culture cells before use. Both rabbitantisera showed good reactivity against rat, human, andmouse P. carinii, suggesting cross-reacting determinants.Secondary antibodies included fluorescein-conjugatedF(ab')2 fragment goat anti-mouse immunoglobulin G (IgG)(heavy and light chain) (Cappel Labs, Malvern, Pa.), ad-sorbed with normal mouse lung, and fluorescein-conjuatedF(ab')2 fragment goat anti-rabbit IgG (heavy and light chain)(Cappel Labs).Frozen lung sections or lung homogenates were incubated
with pooled mouse serum, rabbit antiserum to rat P. carinii,or rabbit antiserum to human P. carinii in a moist chamberfor 30 minutes, washed with phosphate-buffered saline(PBS), incubated with the fluorescein-conjugated goat anti-mouse or anti-rabbit IgG for 30 min, washed with PBS,mounted with glycerol, and examined with a fluorescencemicroscope. P. carinii organisms stained brightly with atypical rim pattern of fluorescence. If P. carinii in a lungspecimen reacted by IFA with pooled mouse serum, theorganisms were considered to be of mouse origin. Reactivitywith either or both of the rabbit antisera did not necessarilyimply species specificity. Negative controls included thelack of fluorescence staining when lung specimens fromnormal or immunodeficient mice without P. carinii weresubstituted as target antigen or when PBS was used as theprimary antibody.The IFA technique was also used to measure serum
antibodies to P. carinii among the mice. Lung homogenatesof mouse, rat, or human P. carinii added to the wells of aglass slide were used as antigens. Serial dilutions of the testserum (beginning at 1:4) were added to the wells, the slidewas washed with PBS, fluorescein-conjugated goat anti-mouse IgG was added, and the slide was washed andprepared for examination as described above. The fluores-cence intensity was graded on a scale from 0 (negative) to 4+(maximum), and the highest serum dilution with a 1+intensity was considered to be the peak antibody titer. Nofluorescent staining occurred when PBS was substituted forthe mouse serum.
Immunoblotting. Immunoblotting studies were performedby procedures described in detail in earlier reports (34, 44).P. carinii-infected mouse lung homogenates served as anti-gen and were compared with rat and human P. carinii lunghomogenates prepared in a similar manner. Organisms weresolubilized for polyacrylamide gel electrophoresis using alysis buffer of 2% sodium dodecyl sulfate-0.06 M Tris (pH6.8)-1% glycerol-5% 2-mercaptoethanol, boiled for 3 min,cooled, and centrifuged at 9,000 x g for 20 min, and thesupernatants were stored at -20°C. The specimens wereelectrophoresed in discontinuous 0.1% sodium dodecyl sul-fate-12.5% polyacrylamide gels by the method of Laemmli(22).
Protein blotting was performed by the method of Towbinet al. (30) with slight modification. Separated proteins weretransferred from the gel to nitrocellulose paper in a Trans-Blot cell (Bio-Rad Laboratories, Richmond, Calif.) over-night at 30 V and then for 1 h at 60 V. The resulting proteinblots were either stained with ponceau red or blocked with3% gelatin and then incubated overnight at 4°C with the sameprimary antibody used in the IFA studies (i.e., pooled mouseserum or rabbit antiserum to rat or human P. carinii). Theblots were then washed, incubated with affinity-purified goatanti-mouse IgG or goat anti-rabbit IgG (Kirkegaard andPerry Laboratories, Gaithersburg, Md.), washed, and re-acted with Tris-buffered saline containing 0.6 mg of 4-chloro-
VOL. 57, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

64 WALZER ET AL.
1-naphthol per ml and 0.015% H202. Negative controlsincluded the lack of immunoreactivity when lungs without P.carinii were used as antigen or when Tris-buffered saline wassubstituted for the primary antibody.
RESULTS
Clinical features of the outbreaks. (i) University of Michi-gan. One outbreak occurred in a colony of 100 Crl:CDI (ICR)nulnu mice maintained under microbiologic barrier condi-tions at the University of Michigan. The mice were allobtained from the same commercial vendor and were group-housed on autoclaved corn cob bedding in 22 plastic shoeboxcages with filter tops. They were provided autoclaved com-mercial rodent chow and water ad libitum. The cages weredistributed among two positive-pressure, laminar flow, massair displacement racks within a room that was designatedsolely for housing nude mice. The animals were used exper-imentally to carry human squamous cell carcinoma cells orhuman breast cancer cells, but the majority of the animals inthe colony were not implanted with tumors.
P. carinii was first detected histopathologically in 12 of 16clinically healthy, 4- to 5-month-old outbred nude mice at theroutine termination of a cancer chemotherapy researchproject. Four months later, wasting and occasional sponta-neous deaths of 5- to 7-month-old nude mice belonging to theother investigators in the room began to occur. In addition toweight loss, clinical signs in some animals included cyanosisand dyspnea. Eight moribund nude mice from the totalpopulation representing all investigators were humanelyeuthanized and necropsied. Microscopic lesions were lim-ited to the lung and showed typical features of pneumocys-tosis.Anal tapes and fecal direct smears were negative for
parasites. Serology for Mycoplasma pulmonis and 11 majormurine viruses, including Sendai virus, mouse hepatitisvirus, pneumonia virus of mice, and polyomavirus, wasperformed (Charles River Biotechnical Services, Inc.).Three clinically ill nulnu mice aged 6 to 7 months wereseronegative on all 12 tests. Ten nul+ mice housed with thenude mice and which served as sentinel animals wereserologically negative for antibodies in the test panel.
P. carinii pneumonia was eliminated from the animalfacility at the University of Michigan by quarantine anddepopulation of the entire nude mouse colony and repopu-lation with animals free of the organism.
(ii) Fox Chase Cancer Center. The Fox Chase CancerCenter outbreak in C B-17/Icr scidlscid mice was firstrecognized in mid 1984 as an increase in mortality within acolony of approximately 900 scidlscid mice. All of thesemice were housed in microisolator cages within the conven-tional rodent colony. Necropsies on about 100 animals wereperformed over the next 9 months (August, 1984, to April,1985). Of these, 66 animals were diagnosed as positive for P.carinii with methenamine silver stain. The age of affectedanimals ranged from 17 through 87 weeks, with the majoritybetween 35 and 40 weeks. Clinically, the animals presentedwith chronic debilitation, weight loss, and dyspnea whichprogressed to cyanosis. The mice were frequently ill forseveral months before they became moribund. The lungswere densely consolidated with a firm, rubber consistencyand did not "deflate" when the thoracic cavity was opened.The colony was euthanized and repopulated with animalsfrom a barrier colony which had always been free of P.carinii infection as judged by histologic examination. Thesemice have remained in good health.
The nulnu mouse colony was known to be infected with P.carinii for more than 10 years. In general, deaths wererestricted to older animals, at least 40 weeks old. Clinicalfeatures of pneumocystosis were indistinguishable fromthose in scid/scid mice. Lack of hair in the nulnu miceallowed earlier detection of infected animals, with weightloss usually being the first sign. As the disease progressed,the animals lost their normal pink color due to cyanosis. Thelungs of infected nulnu mice had the same morphologicfeatures as the lungs of infected scidiscid mice. P. carinjiwas eliminated from nulnu mouse stocks by caesarian rede-rivation and isolator maintenance.Among both groups of immunodeficient mice, pneu-
mocystosis appeared to develop at an earlier age and causemore severe clinical illness in mice housed in microisolatorcages or in crowded cages (i.e., four to five mice per cage)than in mice housed in conventional cages.
(iii) Smith Kline & French. The Smith Kline & Frenchoutbreak began in November, 1985, in one room of abarrier-operated facility containing Crl:CDI (ICR) nulnu andnul+ mice which had been obtained from the same vendor asthe mice at the University of Michigan. The nulnu mice weregroup-housed in polycarbonate shoebox cages with micro-isolator lids (Lab Products, Maywood, N.Y.) and autoclavedhardwood chip bedding, food, and water. All animal manip-ulations and cage changing was performed under a biologicalsafety cabinet by personnel wearing protective clothing(e.g., masks, gloves, etc.). Ultimately, about 75% of the 300nulnu mice housed in this room were involved in theoutbreak. A survey of nulnu mice housed in other roomsrevealed that P. carinii had spread to these locations as well.The clinical manifestations of pneumocystosis includedwasting, diarrhea, and cyanosis. The illness seemed to bemore severe in mice more than 4 months old than in youngeranimals and more severe in mice housed in microisolatorcages than in animals housed under conventional conditions.Lung histology and cultures, as well as serologic testing ofnulnu and sentinel nul+ mice, failed to reveal an etiologyother than P. carinii. The outbreak was controlled bydepopulation and restocking the colony with mice obtainedfrom a separate breeding facility maintained by the vendorknown to be free of P. carinii infection. These mice haveremained well.
(iv) The Jackson Laboratory. C B-17 scidiscid mice wereintroduced into a research colony at the Jackson Laboratoryby caesarian derivation from homozygotes kindly suppliedby Gayle and Mel Bosma at the Fox Chase Cancer Center in1984. The mice were housed at four to five animals per cagein pine shavings under conventional conditions in a researchanimal room. Lexon filter tops were used to minimizeairborne infections. The diet consisted of pasteurized WayneLab Blox and acidified water ad libitum. Although 60 scidlscid mice raised under the above conditions showed a meanlife span of 337 + 13.3 days over the first 18 months, therewas a gradual decrease in longevity associated with clinicalsigns of debilitation and chronic respiratory disease. Themean life span of scidiscid mice in this research colonydecreased to 133 + 2.98 days. Analysis of lung tissue by(Gomori methenamine silver stain and by a sensitive immu-noperoxidase stain of paraffin-embedded sections confirmedthe presence of P. carinii. The age of affected animals variedfrom 6 to 30 weeks.To protect other immunocompromised mice from P. cari-
nii infection and to minimize spread of this pathogen, thescidlscid mice were placed in a Maxi-Miser positive individ-ually ventilated (PIV) cage rack (Thoren Caging System,
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

OUTBREAKS OF P. CARINII 65
Hazelton, Pa.) which had a high-efficiency particulate air(HEPA)-filtered air supply and exhaust system and wasunder positive pressure relative to the outside. Rather thanimproving the health of the scidlscid mice, housing theseanimals in the PIV rack had a detrimental effect on life span.Within 6 months of placing scid/scid mice in the PIV rack,the mean life span had dropped further to only 91.2 + 2.64days. Histopathological examination of stained lungs con-firmed severe pneumonia caused by infection with P. carinii.
Since trimethoprim-sulfamethoxazole (TMP-SMZ) sus-pension had been previously shown to be effective in theprevention and treatment of P. carinii pneumonia in humansand in corticosteroid-suppressed rats (15-17), scidlscid micewere placed on a regimen of TMP-SMZ in the drinkingwater. To assess the efficacy of TMP-SMZ treatment in aconventional mouse room environment, breeding pairs ofscid/scid mice were removed from the PIV rack and placedin conventional mouse boxes. A dose of 500 of SMZ and 100mg of TMP per kg per day was supplied in the drinking wateras Sulfatrim suspension (Goldline Laboratories). The micewere supplied with TMP-SMZ suspension in drinking waterfor 3 days per week and were provided with water alone forthe remainder of the week in a manner similar to theintermittent chemoprophylactic regimen used in rats (18).Treatment with TMP-SMZ had a remarkable effect on the
health of the scid/scid mice, with improvement in theiroverall appearance and an increase in their life span. Thefirst scid/scid mice were started on the TMP-SMZ regimenshortly after birth and have been followed for 6 months; theoverall mortality has been <5%. Five of these animals werekilled at 3 to 4 months of age, and histopathological evalua-tion of methenamine silver-stained lung sections revealedwidely scattered P. carinii organisms with focal accumula-tions of macrophages. Moreover, the breeding performanceof homozygotes did not appear to be affected by the TMP-SMZ therapy. New generations of mice are currently beingderived by hysterectomy. On the other hand, six young scidlscid mice were first placed in the PIV rack for 6 weeks andthen begun on TMP-SMZ; analysis of the lungs of theseanimals killed at 6 to 7 months of age revealed severe P.carinii pneumonia.
Histopathology. The histologic features of the differentoutbreaks of pneumocystosis were very similar, although theextent of disease varied among individual mice. Of the 11nude mice graded by a single examiner, 3 had light, 5 hadmoderate, and 3 had severe P. carinii pneumonia; of 20 scidmice analyzed, 5 had light, 4 had moderate, and 11 hadsevere disease.
Representative histologic features of pneumocystosis arepresented in Figure 1. In light infection, the underlying lungarchitecture was not altered except for slight interstitialthickening and mild chronic inflammatory cell infiltrates.Methenamine silver stain depicted P. carinii cyst formsalong the alveolar walls. As the disease progressed, thealveolar lumens contained the characteristic foamy honey-comb material (Fig. 1A to C) which revealed various num-bers of organisms (Fig. 1D). In some areas, the alveolicontained prominent collections of macrophages but few P.carinii organisms (Fig. 1E and F). Severe pneumocystosiswas characterized by extensive alveolar filling with thefoamy material and masses of P. carinii, interstitial thicken-ing with edema and mild fibrosis, and foci of atelectasis withcompensatory hyperinflation; however, host inflammatorycell infiltrates were still rather inconspicuous.
Immunofluorescence. Initial IFA studies were performedon nulnu mice obtained from the University of Michigan and
on scidlscid mice obtained from Fox Chase. Since we werecontacted late in the course of these outbreaks, we were onlyable to obtain four mice (two nude, two scid) with sufficientlung tissue and P. carinii infection available for testing. Allspecimens reacted with pooled mouse serum, suggesting amouse strain of organism. However, discrepancies (e.g.,variation in the intensity of the immunofluorescent staining)occurred between lung homogenates and frozen sectionswhen tested with rabbit antisera to rat and human P. carinii,thus making further data interpretation difficult.The outbreak at Smith Kline & French provided a larger
number of mice to study as well as the opportunity todevelop a more uniform protocol. Lung homogenates werechosen as the standard antigen preparation because theimmunofluorescent staining was consistently easier to inter-pret than were the frozen sections. Specimens from all ninenulnu mice examined reacted with pooled mouse serum,rabbit antisera to rat P. carinii, and rabbit antisera to humanP. carinii by IFA. In contrast, no lung homogenates from sixnul+ mice reacted with any source of antibody.The results of serologic testing in 22 mice by IFA are
presented in Fig. 2. Serum antibodies to mouse P. cariniiwere detected in 10 of 12 nul+ mice housed in the same roomas the nulnu mice; 6 of these animals had a reciprocal titer of>8. Serum antibodies to rat P. carinii were detected in onlyfour of the nul+ mice, and the titers were lower than thoseto mouse P. carinii. No antibodies to human P. carinfi weredetected. Only 1 of 10 nulnu mice had detectable serumantibodies to any source of P. carinii, and this was a low titerof antibodies to mouse-derived organisms.When we heard about the problem of pneumocystosis in
the nude mouse colony at Fox Chase, we obtained lungspecimens from five nulnu mice. The lung homogenates fromfour of these animals had P. carinii by cresyl echt violetstain; all of these specimens reacted with pooled mouseserum as well as with rabbit antisera to rat and human P.carinii by IFA.Serum antibodies to P. carinii were measured among nul
nu and nul+ mice; because of the small amounts of serumavailable, specimens from one to three animals were pooledfor analysis. All three serum specimens from the nul+ micehad reciprocal antibody titers of 128 (Fig. 2). Only one ofthese mice had antibodies to rat P. carinfii, and none hadantibodies to human organisms. No serum antibodies to anysource of P. carinii were found in two specimens from nulnumice.
Eight scid/scid mice were obtained from the ResearchAnimal Facility at The Jackson Laboratory for analysis. Theanimals had been housed for 3 to 5 months and appearedclinically ill. The lung homogenates from all eight scid/scidmice reacted with pooled mouse serum and rabbit antisera torat and human P. carinii by IFA.
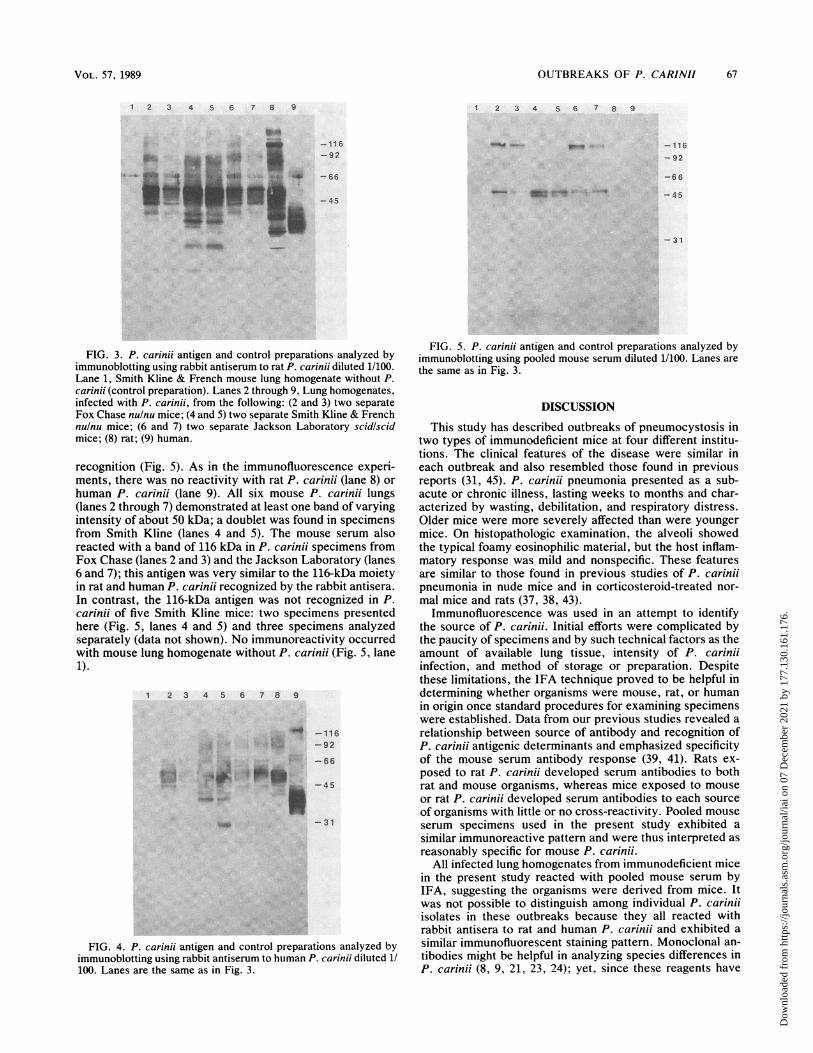
Immunoblotting. In the immunoblotting studies, lung ho-mogenates from mice in the different outbreaks were probedwith different sources of antibody and compared with theantigenic profile of rat and human P. carinii. Data obtainedusing rabbit antiserum to rat P. carinii are presented in Fig.3. As in our earlier reports (34), multiple bands were foundwith rat P. carinii, the most prominent of which weremoieties of about 45, 50, and 116 kilodaltons (kDa) (Fig. 3,lane 8). The principal antigen recognized in human P. carinuiwas a broad-based band of about 40 kDa (lane 9). The majorantigens recognized in mouse P. carinii were bands of about45 and 50 kDa, as seen in specimens from Fox Chase (lanes2 and 3), Smith Kline (lanes 4 and 5), and Jackson Labora-tory (lanes 6 and 7); these moieties were similar to the 45-
VOL. 57, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.
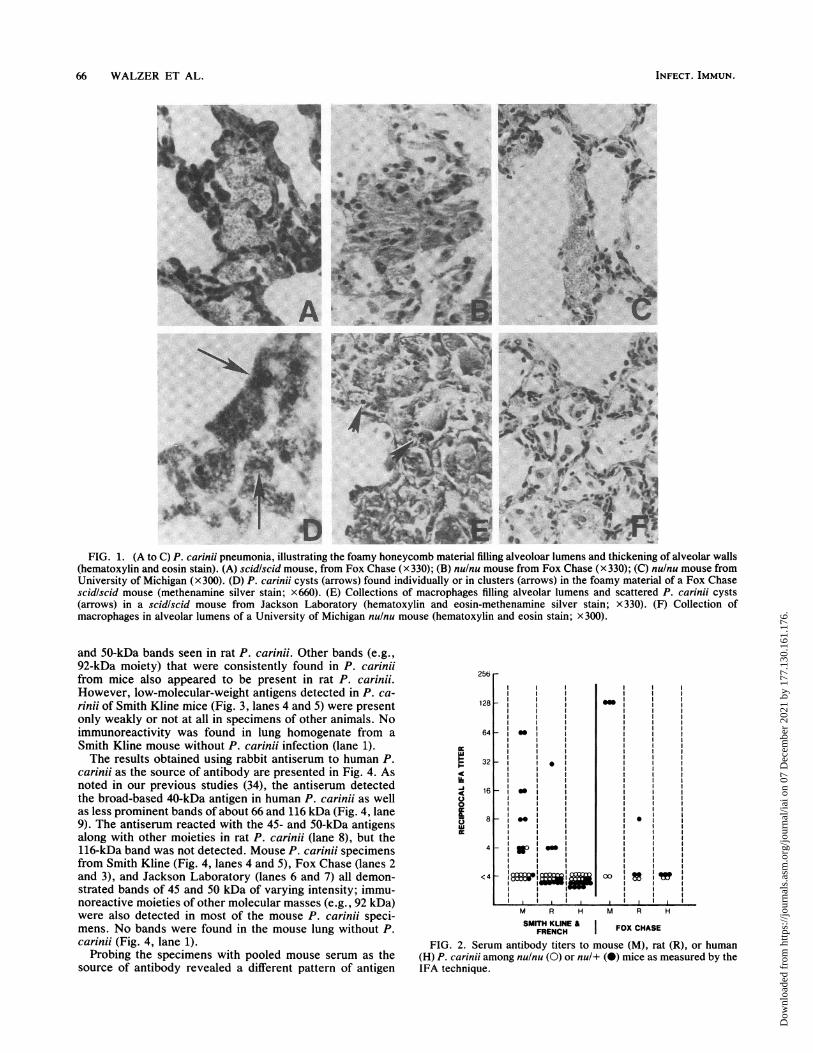
66 WALZER ET AL.
FIG. 1. (A to C) P. carinii pneumonia, illustrating the foamy honeycomb material filling alveoloar lumens and thickening of alveolar walls(hematoxylin and eosin stain). (A) scid/scid mouse, from Fox Chase (x 330); (B) nulnu mouse from Fox Chase (x 330); (C) nulnu mouse fromUniversity of Michigan (x300). (D) P. carinii cysts (arrows) found individually or in clusters (arrows) in the foamy material of a Fox Chasescidiscid mouse (methenamine silver stain; x660). (E) Collections of macrophages filling alveolar lumens and scattered P. carinii cysts(arrows) in a scid/scid mouse from Jackson Laboratory (hematoxylin and eosin-methenamine silver stain; x 330). (F) Collection ofmacrophages in alveolar lumens of a University of Michigan nulnu mouse (hematoxylin and eosin stain; X300).
and 50-kDa bands seen in rat P. carinii. Other bands (e.g.,92-kDa moiety) that were consistently found in P. carinfiifrom mice also appeared to be present in rat P. carinii.However, low-molecular-weight antigens detected in P. ca-rinii of Smith Kline mice (Fig. 3, lanes 4 and 5) were presentonly weakly or not at all in specimens of other animals. Noimmunoreactivity was found in lung homogenate from aSmith Kline mouse without P. carinii infection (lane 1).The results obtained using rabbit antiserum to human P.
carinii as the source of antibody are presented in Fig. 4. Asnoted in our previous studies (34), the antiserum detectedthe broad-based 40-kDa antigen in human P. carinfii as wellas less prominent bands of about 66 and 116 kDa (Fig. 4, lane9). The antiserum reacted with the 45- and 50-kDa antigensalong with other moieties in rat P. carinfii (lane 8), but the116-kDa band was not detected. Mouse P. carinii specimensfrom Smith Kline (Fig. 4, lanes 4 and 5), Fox Chase (lanes 2and 3), and Jackson Laboratory (lanes 6 and 7) all demon-strated bands of 45 and 50 kDa of varying intensity; immu-noreactive moieties of other molecular masses (e.g., 92 kDa)were also detected in most of the mouse P. carinii speci-mens. No bands were found in the mouse lung without P.carinii (Fig. 4, lane 1).
Probing the specimens with pooled mouse serum as thesource of antibody revealed a different pattern of antigen
256 r
128 F
64
I--
IcC,'U
32 F
16 _
8
<c4 _
0O
0
II
m
.
M R H M R H
SMITH KLINE & I FOX CHASEFRENCH
FIG. 2. Serum antibody titers to mouse (M), rat (R), or human(H) P. carinii among nulnu (0) or nul+ (-) mice as measured by theIFA technique.
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.
OUTBREAKS OF P. CARINII 67
1 2 3 4 5 6 7 8 9 2 3 4 5 6 7 8 9
-116-92
_-...r4,
FIG. 3. P. carinii antigen and control preparations analyzed byimmunoblotting using rabbit antiserum to rat P. carinii diluted 1/100.Lane 1, Smith Kline & French mouse lung homogenate without P.carinii (control preparation). Lanes 2 through 9, Lung homogenates,infected with P. carinii, from the following: (2 and 3) two separateFox Chase nulnu mice; (4 and 5) two separate Smith Kline & Frenchnulnu mice; (6 and 7) two separate Jackson Laboratory scidlscidmice; (8) rat; (9) human.
recognition (Fig. 5). As in the immunofluorescence experi-ments, there was no reactivity with rat P. carinii (lane 8) or
human P. carinii (lane 9). All six mouse P. carinii lungs(lanes 2 through 7) demonstrated at least one band of varyingintensity of about 50 kDa; a doublet was found in specimensfrom Smith Kline (lanes 4 and 5). The mouse serum alsoreacted with a band of 116 kDa in P. carinii specimens fromFox Chase (lanes 2 and 3) and the Jackson Laboratory (lanes6 and 7); this antigen was very similar to the 116-kDa moietyin rat and human P. carinii recognized by the rabbit antisera.In contrast, the 116-kDa antigen was not recognized in P.carinii of five Smith Kline mice: two specimens presentedhere (Fig. 5, lanes 4 and 5) and three specimens analyzedseparately (data not shown). No immunoreactivity occurredwith mouse lung homogenate without P. carinii (Fig. 5, lane1).
1 2 3 4 5 6 7 8 9
_ -116~:.- -92:.._ DA
.:...*
!*#c !.Io"jP-45
-31
FIG. 4. P. carinii antigen and control preparations analyzed byimmunoblotting using rabbit antiserum to human P. carinii diluted 1/100. Lanes are the same as in Fig. 3.
FIG. 5. P. carinii antigen and control preparations analyzed byimmunoblotting using pooled mouse serum diluted 1/100. Lanes are
the same as in Fig. 3.
DISCUSSION
This study has described outbreaks of pneumocystosis intwo types of immunodeficient mice at four different institu-tions. The clinical features of the disease were similar ineach outbreak and also resembled those found in previousreports (31, 45). P. carinii pneumonia presented as a sub-acute or chronic illness, lasting weeks to months and char-acterized by wasting, debilitation, and respiratory distress.Older mice were more severely affected than were youngermice. On histopathologic examination, the alveoli showedthe typical foamy eosinophilic material, but the host inflam-matory response was mild and nonspecific. These featuresare similar to those found in previous studies of P. cariniipneumonia in nude mice and in corticosteroid-treated nor-
mal mice and rats (37, 38, 43).Immunofluorescence was used in an attempt to identify
the source of P. carinii. Initial efforts were complicated bythe paucity of specimens and by such technical factors as theamount of available lung tissue, intensity of P. cariniiinfection, and method of storage or preparation. Despitethese limitations, the IFA technique proved to be helpful indetermining whether organisms were mouse, rat, or humanin origin once standard procedures for examining specimenswere established. Data from our previous studies revealed a
relationship between source of antibody and recognition ofP. carinii antigenic determinants and emphasized specificityof the mouse serum antibody response (39, 41). Rats ex-
posed to rat P. carinii developed serum antibodies to bothrat and mouse organisms, whereas mice exposed to mouse
or rat P. carinii developed serum antibodies to each source
of organisms with little or no cross-reactivity. Pooled mouse
serum specimens used in the present study exhibited a
similar immunoreactive pattern and were thus interpreted as
reasonably specific for mouse P. carinii.All infected lung homogenates from immunodeficient mice
in the present study reacted with pooled mouse serum byIFA, suggesting the organisms were derived from mice. Itwas not possible to distinguish among individual P. cariniiisolates in these outbreaks because they all reacted withrabbit antisera to rat and human P. carinii and exhibited a
similar immunofluorescent staining pattern. Monoclonal an-
tibodies might be helpful in analyzing species differences inP. carinii (8, 9, 21, 23, 24); yet, since these reagents have
- 66
-45
- 116
-92
-6 6
-45
- 31
VOL. 57, 1989
-UU)
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

68 WALZER ET AL.
mainly been prepared using rat- or human-derived organismsas antigen and are not yet widely available, their applicabil-ity to studies of mouse P. carinii is unclear.Serum antibody results indicated that the principal sero-
logic response of nul+ mice at Smith Kline and Fox Chasewas directed towards mouse P. carinii, thus lending furthersupport to the hypothesis that mouse-derived organismswere the cause of illness in these outbreaks. As in ourprevious studies (41), nude mice were unable to mount ameaningful antibody response to any source of P. carinii.
In recent years, immunoblotting has been used to comparethe antigenic characteristics of rat and human P. carinii (8-10, 20, 23, 34). The present study has applied this techniqueto mouse P. carinii, and overall, the data are consistent withthe results obtained with immunofluorescence. Rabbit anti-sera to rat and human P. carinii reacted with bands of 45kDa, 50 kDa, and other molecular masses in infected-lunghomogenates of three different groups of mice, suggestingshared or cross-reacting determinants. The antigenic char-acteristics of mouse P. carinii appear to be more closelyrelated to those of rat than to those of human P. carinii. Thespecificity of the pooled mouse serum for mouse P. cariniiwas again demonstrated by its lack of reactivity with rat- orhuman-derived organisms. This serum did react with a116-kDa band and other moieties in infected mouse lungs.The 116-kDa antigen in mouse P. carinii is very similar to the116-kDa bands in rat and human P. carinii. It is of interestthat, when tested with different reagents in this study, theseantigens only reacted with the homologous source of anti-body. Whether the differences in immunoreactivity amongthe Smith Kline & French nulnu mice represent true speciesor strain differences in P. carinii, or perhaps are merely dueto technical factors, is unclear; however, all lung specimenswere prepared and analyzed in a similar manner.
Introduction of P. carinii into a colony of immunodeficientmice may be related to the manner in which these animalsare obtained. Nude mice at the University of Michigan andSmith Kline & French were purchased from the samecommercial vendor. This breeder had been implicated as thesource of a previously reported outbreak of pneumocystosisin nulnu mice (45), and conversations among veterinariansrevealed that several additional institutions were experienc-ing problems with P. carinii among nude mice obtained fromthis company. Investigations by the vendor indicated that P.carinii was present among mice in certain breeding facilitiesbut not in others. This information is important not only forresearchers but also because it can enable a vendor to adjustbreeding activities to ensure an adequate supply of mice freeof P. carinii infection.Although the scidlscid mice at The Jackson Laboratory
were originally developed from breeding pairs of animalsobtained from Fox Chase, the source of the pneumocystisoutbreaks at these two institutions could not be determined.Once P. carinii became established at The Jackson Labora-tory, the organism apparently spread to other mice as thecolony expanded. The fact that P. carinii may be present inresearch colonies of scidlscid or nulnu mice has importantimplications for investigators who wish to develop coloniesof these immunodeficient animals at their own institutions.The spread of pneumocystosis among the animal colonies
described in the present study emphasizes the communica-bility of the organism. Previous reports have implicated theairborne route as the mode of transmission (11, 13, 14, 43),but questions about the infective form of P. carinii (cyst ortrophozoite), environmental sources of the organism, andvertical transmission remain largely unanswered. Data from
the present study suggest that cages with special filters orlids protect mice from environmental exposure to P. carinii.However, immunodeficient mice already infected with P.carinii, housed in these cages, develop a more severe form ofpneumocystosis than do animals housed in conventionalopen cages. It is unclear whether this difference in diseasereflects a higher P. carinii burden within the inspired air,animal crowding, or other factors (e.g., temperature, humid-ity, ammonia, carbon dioxide levels) within the milieu ofthese microisolator cages.The outbreaks described in this report illustrate the patho-
genicity of P. carinii for immunodeficient mice under naturalconditions. While these animals might serve as experimentalmodels for P. carinii, efforts to develop a reliable system forgeneral use have been rather disappointing. Our originalstudies suggested that nude mice could be infected with ratand human P. carinii by different methods of inoculation butdeveloped little overt clinical illness (43). In later studiesperformed at another institution with a different strain ofnude mice and with nude rats, we were unable to achieve theexperimental transmission of P. carinii with any degree ofconsistency (36). Similar negative results have been reportedto us in unpublished studies by other investigators. On theother hand, Japanese workers have conducted a variety ofP.carinii transmission experiments among nude mice and nuderats and have used these animals to analyze host immuneresponses to P. carinii and to perpetuate the infection intheir colonies (4-7). Yet P. carinii replication in the animalswas modest, and corticosteroids had to be used in somecases to facilitate development of infection. scidlscid mice,which are even more immunologically impaired than are nulnu mice, might be explored as an alternative animal model.
Control of pneumocystosis within colonies of immunode-ficient mice has been difficult. As evidenced by the persis-tence of P. carinii in Fox Chase nulnu mice for over 10years, once infection with the organism has been establishedit tends to become endemic. The principal control measurehas been to depopulate the colony and repopulate usinganimals with no known exposure to the organism. Althoughdrugs such as TMP-SMZ have been successful in the treat-ment and prevention of pneumocystosis in humans and rats(15-18), there has been reluctance to use these agents inimmunodeficient mice. These drugs are not microbicidal forP. carinii and hence are only effective in chemoprophylaxisas long as they are being given (12). Since TMP-SMZ andrelated drugs work by inhibiting synthesis of folic acid, therehas been concern about their effects on animal breeding,immune function, or use in tumor studies. The apparentsuccess of TMP-SMZ on the scidlscid mice in a convention-ally housed research colony at The Jackson Laboratory isencouraging. Since P. carinii organisms could still be foundin healthy animals, further studies will be necessary to findthe optimal dose regimen.
Outbreaks of pneumocystosis constitute a major problemto colonies of immunodeficient mice and may be becomingmore common. Successful strategies to meet this challengeshould include strictly controlled policies for breeding andmaintaining these colonies, careful surveillance for P. cari-nii, and early intervention at the first signs of infection. Theonly available method for detecting P. carinii in mousecolonies has been lung histology, which is cumbersome, timeconsuming, expensive, and relatively insensitive. Thepresent study has demonstrated the feasibility of usingserology as a sensitive test to monitor sentinel heterozygotemice for exposure to P. carinii in a manner similar to thatused for viruses and mycoplasma. The major impediment to
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

OUTBREAKS OF P. CARINII 69
the use of serology is a lack of a ready supply of antigen.Moreover, serological tests cannot be used to diagnosepneumocystosis in immunodeficient mice since some ofthese animals lack immunoglobulins or are poor antibodyresponders. Systems for detecting P. carinii antigens orgenetic material in tissues might be explored as alternativediagnostic techniques (33). Regardless of the method used,the heightened interest in P. carinii and widespread use ofimmunodeficient mice in biomedical research emphasize theneed to develop practical methods of detecting and control-ling infection with the organism in these animal colonies.
ACKNOWLEDGMENTS
This study was supported by the Medical Research Service,Veterans Administration, and by Public Health Service ContractNO1-AI-72646 and research grants AI-25897 and CA-20408 from theNational Institutes of Health.We thank David Stanforth, Howard Blatt, Gayle Bosma, Roy
Tietjen, and Angela Birch Smith for their valuable help.
LITERATURE CITED1. Bosma, G. C., R. P. Custer, and M. J. Bosma. 1983. A severe
combined immunodeficiency mutation in the mouse. Nature(London) 301:527-530.
2. Farrow, B. R. H., A. D. J. Watson, and W. J. Hartley. 1972.Pneumocystis pneumonia in the dog. J. Comp. Pathol. 82:447-453.
3. Frenkel, J. K., J. T. Good, and J. A. Schultz. 1966. LatentPneumocystis infection of rats, relapse, and chemotherapy.Lab. Invest. 15:1559-1577.
4. Furuta, T., and K. Ueda. 1987. Intra- and inter-species trans-mission and antigenic differences of Pneumocystis carinii de-rived from rat and mouse. Jpn. J. Exp. Med. 57:11-17.
5. Furuta, T., K. Ueda, and K. Fujiwara. 1984. ExperimentalPneumocystis carinii infection in nude rats. Jpn. J. Exp. Med.54:65-72.
6. Furuta, T., K. Ueda, K. Fujiwara, and K. Yamanouchi. 1985.Cellular and humoral immune responses of mice subclinicallyinfected with Pneumocystis carinii. Infect. Immun. 47:544-548.
7. Furuta, T., K. Ueda, S. Kyuwa, and K. Fujiwara. 1984. Effect ofT-cell transfer on Pneumocystis carinii in nude mice. Jpn. J.Exp. Med. 54:57-64.
8. Gigliotti, F., D. D. Stokes, A. B. Cheatham, D. S. Davis, andW. T. Hughes. 1986. Development of murine monoclonal anti-bodies to Pneumocystis carinii. J. Infect. Dis. 154:315-322.
9. Graves, D. C., S. J. McNabb, M. H. Ivey, and M. A. Worley.1986. Development and characterization of monoclonal antibod-ies to Pneumocystis carinii. Infect. Immun. 51:125-133.
10. Graves, D. C., S. J. McNabb, M. A. Whorley, J. D. Downs, andM. H. Ivey. 1986. Analysis of rat Pneumocystis carinii antigensrecognized using Western immunoblotting. Infect. Immun. 54:96-103.
11. Hendley, J. O., and T. H. Weller. 1971. Activation and trans-mission in rats of infection with Pneumocystis carinii. Proc.Soc. Exp. Biol. Med. 137:1401-1404.
12. Hughes, W. T. 1979. Limited effect of trimethoprim-sulfameth-oxazole prophylaxis on Pneumocystis carinii. Antimicrob.Agents Chemother. 16:333-335.
13. Hughes, W. T. 1982. Natural mode of acquisition for de novoinfection with Pneumocystis carinii. J. Infect. Dis. 145:842-848.
14. Hughes, W. T., D. L. Bartley, and B. M. Smith. 1983. A naturalsource of infection due to Pneumocystis carinii. J. Infect. Dis.147:595.
15. Hughes, W. T., S. Feldman, C. C. Chaudhary, M. J. Ossi, F.Cox, and S. K. Sanyal. 1978. Comparison of pentamidineisethionate and trimethoprim-sulfamethoxazole in treatment ofPneumocystis carinii pneumonia. J. Pediatr. 92:285-291.
16. Hughes, W. T., P. C. McNabb, and T. D. Makres. 1974. Efficacyof trimethoprim and sulfamethoxazole in the prevention andtreatment of Pneumocystis carinii pneumonitis. Antimicrob.Agents Chemother. 5:289-293.
17. Hughes, W. T., G. K. Rivera, M. J. Schell, D. Thornton, and L.Lott. 1987. Successful intermittent chemoprophylaxis for Pneu-mocystis carinii pneumonitis. N. Engl. J. Med. 316:1627-1632.
18. Hughes, W. T., and B. L. Smith. 1983. Intermittent chemo-prophylaxis for Pneumocystis carinii pneumonia. Antimicrob.Agents Chemother. 24:300-301.
19. Kim, C. K., J. M. Foy, M. T. Cushion, D. Stanforth, M. J.Linke, H. L. Hendrix, and P. D. Walzer. 1987. A comparison ofhistologic and quantitative techniques in the evaluation ofexperimental Pneumocystis carinii pneumonia. Antimicrob.Agents Chemother. 31:197-201.
20. Kovacs, J. A., J. L. Halpern, J. C. Swan, J. Moss, J. E. Parrillo,and H. Masur. 1988. Identification of antigens and antibodiesspecific for Pneumocystis carinii. J. Immunol. 140:2023-2031.
21. Kovacs, J. A., J. C. Swann, J. Shelhamer, V. Gill, F. Ognibene,J. E. Parillo, and H. Masur. 1986. Prospective evaluation of amonoclonal antibody in diagnosis of Pneumocystis carinii pneu-monia. Lancet ii:1-3.
22. Laemmli, U. K. 1980. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
23. Lee, C. H., C. D. Bolinger, M. S. Bartlett, R. B. Kohler, C. E.Wilde, and J. W. Smith. 1986. Production of monoclonal anti-body against Pneumocystis carinii by using a hybrid of ratspleen and mouse myeloma cells. J. Clin. Microbiol. 23:505-508.
24. Matsumoto, Y., T. Amayai, M. Yamada, J. Imanishi, and Y.Yoshida. 1987. Production of a monoclonal antibody with spec-ificity for the pellicle of Pneumocystis carinii by hybridoma.Parasitol. Res. 73:228-233.
25. Milder, J. E., P. D. Walzer, J. D. Coonrod, and M. E. Rutledge.1980. Comparison of histological and immunological techniquesfor detection of Pneumocystis carinii in rat bronchial lavagefluid. J. Clin. Microbiol. 11:409-417.
26. Mills, J. 1986. Pneumocystis carinii and Toxoplasma gondiiinfections in patients with AIDS. Rev. Infect. Dis. 8:1001-1011.
27. Minielly, J. A., S. D. Mills, and K. E. Holley. 1969. Pneumocys-tis carinii pneumonia. Can. Med. Assoc. J. 100:846-854.
28. Sheldon, W. H. 1959. Experimental pulmonary Pneumocystiscarinii infection in rabbits. J. Exp. Med. 110:147-160.
29. Stokes, D. C., F. Gigliotti, J. E. Rehg, R. L. Snellgrove, andW. T. Hughes. 1987. Experimental Pneumocystis carinii pneu-monia in the ferret. Br. J. Exp. Pathol. 68:267-276.
30. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
31. Ueda, K., Y. Goto, S. Yamazaki, and K. Fujiwara. 1977. Chronicfatal pneumocystosis in nude mice. Jpn. J. Exp. Med. 47:475-482.
32. Walzer, P. D. 1984. Experimental models for Pneumocystiscarinii infection, p. 7-75. In L. S. Young (ed.), Pneumocystiscarinii infections. Marcel Dekker, Inc., New York.
33. Walzer, P. D. 1988. Diagnosis of Pneumocystis carinii pneumo-nia. J. Infect. Dis. 157:629-632.
34. Walzer, P. D., and M. J. Linke. 1987. A comparison of theantigenic characteristics of rat and human Pneumocystis cariniiby immunoblotting. J. Immunol. 138:2257-2265.
35. Walzer, P. D., D. P. Perl, D. J. Krogstad, and P. G. Rawson.1974. Pneumocystis carinii pneumonia in the United States:epidemiologic, clinical and diagnostic features. Ann. Intern.Med. 80:83-93.
36. Walzer, P. D., and R. D. Powell. 1982. Pneumocystis cariniiinfection in nude and steroid treated normal mice, p. 123-132. InN. D. Reed (ed.), Proceedings of the Third International Work-shop on Nude Mice. Gustav Fisher, New York.
37. Walzer, P. D., R. D. Powell, and K. Yoneda. 1979. ExperimentalPneumocystis carinii pneumonia in different strains of cortiso-nized mice. Infect. Immun. 24:939-947.
38. Walzer, P. D., R. D. Powell, K. Yoneda, M. E. Rutledge, andJ. E. Milder. 1980. Growth characteristics and pathogenesis ofexperimental Pneumocystis carinii pneumonia. Infect. Immun.27:928-937.
VOL. 57, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.

INFECT. IMMUN.
39. Walzer, P. D., and M. E. Rutledge. 1980. Comparison of rat,mouse, and human Pneumocystis carinii by immunofluores-cence. J. Infect. Dis. 142:449.
40. Walzer, P. D., and M. E. Rutledge. 1981. Humoral immunity inexperimental Pneumocystis carinii pneumonia. I. Systemic andlocal antibody responses in rats. J. Lab. Clin. Med. 97:820-833.
41. Walzer, P. D., and M. E. Rutledge. 1982. Serum antibodyresponses to Pneumocystis carinii among different strains ofnormal and athymic (nude) mice. Infect. Immun. 35:620-626.
42. Walzer, P. D., M. E. Rutledge, and K. Yoneda. 1979. A new
method of separating Pneumocystis carinii from infected lungtissue. Exp. Parasitol. 47:356-368.
43. Walzer, P. D., V. Schnelle, D. Armstrong, and P. P. Rosen. 1977.
The nude mouse: a new experimental model for Pneumocystiscarinii infection. Science 197:177-179.
44. Walzer, P. D., D. Stanforth, M. J. Linke, and M. T. Cushion.1987. Pneumocystis carinii: immunoblotting and immunofluo-rescent analysis of serum antibodies during rat infection andrecovery. Exper. Parasitol. 63:319-328.
45. Weir, E., D. G. Brownstein, and S. W. Barthold. 1986. Sponta-neous wasting disease in nude mice associated with Pneumo-cystis carinii infection. Lab. Animal Sci. 36:140-144.
46. Yoshida, Y., M. Yamada, T. Shiota, T. Ikai, S. Takeuchi, and K.Ogino. 1981. Provocation experiment: Pneumocystis carinii inseveral kinds of animals. Zentralbl. Bakterrol. Parasitenkd.Infektionskr. Hyg. Abt. 1 Orig. Reihe A 250:206-212.
70 WALZER ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 07
Dec
embe
r 20
21 b
y 17
7.13
0.16
1.17
6.


![The homology domainofBruton C - PNASas atk, BPK,oremb)], whichis variously mutatedin chromo-some X-linked agammagobulnemia patients and X-linked immunodeficient(xid)mice,hasthepleckstrinhomology(PH)](https://img.pdfslide.us/doc/110x75/60dff19c14fc6d137802578b/the-homology-domainofbruton-c-pnas-as-atk-bpkoremb-whichis-variously-mutatedin.jpg)


























