Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Nov. 1974, p. 687-696Copyright 0 1974 American Society for Microbiology
Vol. 120, No. 2Printed in U.S.A.
Isoleucine and Valine Metabolism in Escherichia coli K-12:Detection and Measurement of ilv-Specific Messenger
Ribonucleic AcidR. A. VONDER HAAR' AND H. E. UMBARGER
Department of Biological Sciences, Purdue University, West Lafayette, Indiana 47907
Received for publication 24 June 1974
Ribonucleic acid-deoxyribonucleic acid (RNA-DNA) hybridization was em-ployed for the determination of messenger RNA transcribed from the ilv genecluster of Escherichia coli K-12. Strains with derepressed levels of the isoleucineand valine biosynthetic enzymes owing to linked or unlinked genetic lesions werefound to exhibit ilv messenger RNA levels from 1.5- to 4-fold higher than didtheir isogenic parents. When grown under conditions that specifically repressedthe synthesis of isoleucine- and valine-forming enzymes, most strains exhibiteddrastically reduced ilv messenger RNA levels. Hybridization performed with theseparated strands of ilv DNA showed that all the ilv genes are transcribed fromthe same strand, the "I strand" of X480CI857St68di1v DNA. Sucrose gradientanalyses of RNA extracted from cells starved for isoleucine, valine, or leucineresulted in the detection of at least two distinct types of ilv messenger RNA.
The five genes specifying the isoleucine- andvaline-forming enzymes in Escherichia coliK-12 are clustered on the chromosome (Fig. 1).(28). Three of the genes, ilvA, ilvD, and ilvE,which specify threonine deaminase (EC4.2.1.16, L-threonine hydro-lyase [deaminating]),dihydroxy acid dehydrase (EC 4.2.1.9, 2.3-dihy-droxy acid hydro-lyase), and transaminase B(EC 2.6.1.42, branched-chain amino-acid:2-oxo-glutarate aminotransferase), respectively, ap-pear to comprise a single operon under the mul-tivalent repression control of isoleucine, valine,and leucine (10). Acetohydroxy acid synthase(EC 4.1.3.18, acetolactate pyruvate-lyase [car-boxylatingD, the product of the ilvB gene, isindependently controlled and fully repressedby valine and leucine alone (9). The product ofthe ilvC gene, isomeroreductase (EC 1.1.1.86,3-alkyl-2i3-dihydroxy-acid:NADP+ oxidoreduc-tase [isomerizing]), is induced by either of itssubstrates, a-acetolactate and a-acetohydrox-ybutyrate (2). These control patterns were elu-cidated from studies of the responses of eachenzyme activity to various types of repressingand derepressing conditions. The control mech-anisms are assumed to function at the level oftranscription as outlined by Jacob and Monod(15), but since the final enzyme complement isthe result of transcription and translation, andpossibly post-translational maturation or ag-
I Present address: Department of Biology, University ofUtah, Salt Lake City, Utah 84112.
gregation, alternative control processes mayexist.The ribonucleic acid-deoxyribonucleic acid
(RNA-DNA) hybridization techniques of Gilles-pie and Spiegelman (11), whereby RNA ishybridized to DNA immobilized on nitrocel-lulose filters, provide a means for direct mea-surement of the transcriptional activity of agene or group of genes. The levels of messengerRNA (mRNA) (under conditions of repressionor induction) have been determined for a num-ber of bacterial operons (3, 7, 13, 14, 17, 29). Inmost cases, transcriptional control has beendemonstrated, but some examples of apparentpost-transcriptional control have been detected(17, 21). To estimate mRNA levels, a source ofDNA enriched for the genes in question isrequired. This has usually been provided bypreparing a specialized transducing phage of Xor 480 that carries the desired bacterial genesand that can be isolated in bulk quantities (4, 6,27). The source of ilv DNA used in the presentstudy was extracted from the phage X480CI857-St68dilv isolated from the double lysogen M1199(kindly provided by M. laccarino). (This phagecarries all five genes of the ilv cluster andtransduces ilvA, ilvD ilvC, and ilvE strains toIlv+. No ilvB strains are available for testing astransduction recipients, but lysogens containingthe phage have nearly double the wild-typelevel of acetohydroxy acid synthase, the ilvBgene product.)This paper describes some parameters of the
687
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

VONDER HAAR AND UMBARGER
Pyruwate-, -Acetolactate- Iqhydroxy- - c.-Keto- -Valnlsovolerate isovalerate
AHS IR DH TRO
,Ketobutyrote et \q-Olhydrmy - loct-Keto-p- Isoleucine\tiy'roxy\utyrats \ G -methylvalerate methylvalerate
|TD- \________
Throonine\\I /
-t-E P B
met
I
CO0 A D
liv
FIG. 1. Biosynthetic pathways for isoleucine and valine. The enzymes catalyzing the indicated steps areabbreviated as follows: TD, threonine deaminase; AHS, acetohydroxy acid synthase; IR, acetohydroxy acidisomeroreductase; DH, dihydroxy acid dehydrase; TRB, transaminase B. All of the enzymes except threoninedeaminase catalyze corresponding steps in the valine and isoleucine pathways. Threonine deaminase is specificfor the isoleucine pathway alone. The broken lines indicate the structural genes in the ilv gene cluster thatspecify the respective enzymes. Gene loci ilvO and ilvP are repression recognition sites controlling the expres-sion of genes ADE and gene B, respectively. The gene order shown in the reverse (that is, left to right is
counterclockwise) of the way it is usually represented on the E. coli chromosome map. Adapted fromRamakrishnan and Adelberg (28).
system used to measure ilv mRNA and some
properties of the mRNA, and lists the ilv mRNAlevels of a number of ilv prototrophic andregulatory mutant strains under repressing andnonrepressing growth conditions.
MATERIALS AND METHODSOrganisms. The bacterial strains used in this
study are listed in Table 1. All are derived from E. coliK-12.
Media. The minimal medium used was that de-scribed by Davis and Mingioli (8), except that citratewas omitted and the glucose concentration was in-creased to 0.5%. Supplements, when required, wereadded at the following concentrations: L-isoleucine,L-leucine, L-methionine, L-histidine, L-tryptophan,0.5 mM; L-valine, 0.8 mM; and thiamine, 0.1 mM.Tryptone broth contained (per liter): tryptone, 10 g;NaCl, 5 g; MgSO4, 1.22 g; and thiamine, 12 mg. Solidmedia were prepared by adding 1.5% agar (Difco) tothe media before autoclaving. Soft-agar overlays forphage titering consisted of tryptone broth with 0.7%agar.Growth of cells, preparation of cell extracts, and
enzymatic assays. Cells were grown, cell extractswere prepared, and enzymatic assays were performedas described previously (9, 32). Protein concentrationwas determined by the biuret method (18). Theoptical density of cultures was determined with a
Spectronic 88 spectrophotometer.Large scale production ofbacteriophage. Bacter-
ial strains lysogenic for heat-inducible bacteriophagewere grown, to the stationary phase, overnight inminimal medium containing the required supple-ments at 30 C in shaker flasks. The next moming,enough cells were added to 100 liters of tryptone brothin a New Brunswick Fermacell fermentor (model
F-250) to achieve an initial optical density at 660 nm(ODe,,o) of 0.1 to 0.15 (1 OD,,o unit = 7.3 x 10i cellsper ml). The cells were grown at 32 C to an OD,,o of0.4. At that time, the temperature of the medium was
increased to 42 C for 30 min, after which growth wascontinued at 37 C for 3 h. The induced cells werechilled in a New Brunswick Rapid Chiller (modelRC-200) and harvested by using a Sharples centrifuge(model AS-14). The cells were stored at -20 C in 20-gpackets.
Extraction of bacteriophage from induced cells.A 20-g amount of induced cells was suspended in 50 to70 ml of TMG buffer [0.01 M tris(hydroxyme-thyl)aminomethane (Tris)-hydrochloride, pH 7.4;0.01 M MgSO4; 10 ug of gelatin per ml]. The phagewere released by adding 3 to 5 ml of chloroform andincubating the mixture at 37 C for 15 min with oc-casional stirring. A 0.1-gg amount of deoxyribonucle-ase per ml was added, and the mixture was incubatedan additional 10 min at 37 C. The mixture wascentrifuged at 10,000 rpm for 10 min, and the result-ing supernatant fluid was transferred to a small flaskand kept on ice. Cesium chloride block gradients wereprepared by first adding 0.5 ml of CsCl (n = 1.3888) tothe bottom of polycarbonate or cellulose nitratecentrifuge tubes and adding successive 1 ml layers ofCsCl (n = 1.3788 and n = 1.3711). The supernatantfluid containing the phage was layered onto thegradients, and the tubes were centrifuged in a Ti-50rotor at 35,000 rpm for 45 min in a Spinco L-2ultracentrifuge. The resulting single phage band wascollected in one fraction and recentrifuged in a cesiumchloride equilibrium gradient (n = 1.3805) at 27,000rpm for 40 h. Phage prepared from strains MI199 andCSH44 were resolved into two bands upon equilib-rium centrifugation. Only the lower (defective) phagebands, X480CI857St68dilv (X080dilv) from strainMI199 and X080CI857St68dlac (A4,80dlac) from strain
E
r_b
688 J. BArEIOL.
I
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ilv-SPECIFIC MESSENGER RNA
TABLE 1. List of strains
Strain Relevant genotypea Relevant ilv phenotype Source or Reference
Cui Wild type H. E. UmbargerCU2 ilvE12 Tramsaminase B negative H. E. UmbargerCU4 gal Wild type J. BlattCU5 Hfr, met Wild type E. SimonCU16 metE200, rbs-215 Prototrophic, but with abnormally W. J. Pledger (26)
low threonine deaminase activitytrpE6185 trpE6185 Wild type R. SomervilleCU109 rnsC, trpE6185 ilvADE and ilvB derepressed J. Wasmuth (33)CUi11 rnsC, ilv0546, trpE6185, ilvADE and ilvB derepressed J. Wasmuth (33)
rbs-215CU113 ilv0468, metE200, rbs-215 ilvADE derepressed E. KlineCU115 ilv0266, metE200, rbs-215 ilvADE derepressed E. KlineCU339 ilv0468 ilvADE derepressed E. KlineCU340 ilv0469 ilvADE derepressed E. KlineCU341 ilv0264 ilvADE derepressed E. KlineCU342 ilv0266 ilvADE derepressed E. KlineCU1014 ilvC462, leu Isomeroreductase negative J. Wasmuth (33)CU5001 met, azl-2 ilvB derepressed S. Dwyer (9)CU5002 met, azi-1 ilvADE and ilvB derepressed S. Dwyer (9)PS164 thi (Xi434N7) M. LevinthalM1199 ilvDAC115, his, trp Heat-inducible dilv phage M. Iaccarino, probably
(XO80CI857St68)b transductant R2 in refer-(X480CI857St68dilv) ence (4)
CSH43 thi (X480CI857St68) J. Miller (22)CSH44 thi (AX80CI357St68) J. Miller (22)
(XA80CI857St68dlac)CU344 gal, ilvDAC115 Isoleucine and valine requirer E. Kline
a The genetic symbols are those employed for the recent E. coli linkage map (30): ilv, met, trp, leu, thi, andhis, genes for isoleucine and valine, methionine, tryptophan, leucine, thiamine, and histidine biosynthesis,respectively; gal, rbs, and lac, genes for galactose, ribose, and lactose utilization, respectively; rnsC, the locusgiving rise to the SuA phenotype; azi, locus for azaleucine-resistance.
b This is a heteroimmune lysogen with normal S function (cell lysis) and can, therefore, be used to titer thephages bearing the St68 (lysis defective) marker.
CSH44, were collected. Nondefective phage wereprepared from strain CSH43, singly lysogenic forX480CI857St68 (X080). All purified phage were col-lected in about 2 ml and diluted 1:1 with TMG bufferin the cold prior to extraction ofDNA. The titer of thenondefective phages in such preparations was about101 plaque-forming units per ml. The X480 bands inthe purification gradients from both the single anddouble lysogens were about the same size. The defec-tive phage bands were usually broader than the X480bands and, for X4080dilv, the transducing titers in thepurified preparations were about 1010 per ml.
Titering of XO8OCI57St68 and \X80CI857St68-dilv phage. The procedure of Adams (1), employingstrain PS164 (kindly supplied by Mark Levinthal) asindicator, was used to titer the number of plaque-forming phage. The procedure of Lennox (19) wasused to determine the titer of ilv transducing phage.
Extraction ofDNA from phage. The procedure ofThomas and Abelson (31), employing phenol, wasused to extract DNA from X080 phage. DNA wasextracted from Xf80dilv or XA80dlac phage by os-motic shock. The phage were first dialyzed againsttwo changes of a solution containing 0.01 Methylenediaminetetraacetate, tetrasodium salt, and
0.01 M tris(hydroxymethyl)aminomethane-hydro-chloride (pH 7.4) at 4 C. The dialysis bag was thenplaced in 250 ml of the same buffer prewarmed to 37 Cand incubated for 15 min to lyse the phage. Theresulting DNA solution was then dialyzed against twochanges of 0.01 M ethylenediaminetetraacetate (pH8.2) followed by dialysis against two changes of 0.01M tris(hydroxymethyl)aminomethane-hydrochloride(pH 8.0). During the dialysis, the volume of the phagepreparations was increased approximately twofold.The yield of DNA produced by either method ( > 5001&g of DNAIml) was determined by measuring theOD... (1 OD unit = 50 sg of DNA/ml).Growth of cells and labeling and extraction of
RNA. Cells to be labeled with [3H]uridine were grownin minimal medium containing supplements to anOD... of 0.80. For labeling during exponential growth,the cells were labeled directly in the growth medium.A 25-ml (or 10-ml) sample of culture was transferredfrom the growth medium or the starvation medium toa small flask containing 0.25 ml (or 0.10 ml) of[3HJuridine (Schwarz/Mann, 0.5 mCi/ml; 28 Ci/mmol) and shaken at 37 C for 30 to 60 s. (Whereas thisamount of uridine would be expected to meet thepyrimidine needs of the amount of cells used for only
689VOL. 120, 1974
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
VONDER HAAR AND UMBARGER
about 30 s, the incorporation proceeded at a constantrate for at least 5 min. Thus, the uridine in themedium was not utilized well enough to quenchendogenous pyrimidine synthesis.) The cells werekilled and the RNA was extracted according to theprocedure of Okamoto, et al. (25). For labeling of anauxotroph during starvation, the cells were grown inminimal medium cQntaining excess isoleucine, valine,and leucine, harvested at room temperature, washedwith 0.05 M phosphate buffer (pH 7.5) at roomtemperature, recentrifuged, and suspended in theappropriate starvation medium, prewarmed to 37 C,lacking one of the branched-chain amino acids. Sam-ples were pulse-labeled for 60 s beginning 5 min afterthe onset of starvation. Since the strains employedexhibited a stringent control over RNA synthesis, thisprocedure for labelling mRNA during starvation isvery inefficient. However, it was sufficient for thequalitative detection of ilv mRNA.
Hybridization procedures. DNA was immobi-lized on nitrocellulose filters (type B-6, 25 mm,Schleicher and Schuell) as described by Nygaard andHall (24). Filter hybridizations were performed by themethod of Gillespie and Spiegelman (11). The prep-aration of the complementary I and r strands of phageDNA and hybridization of [3H]RNA to each were asdescribed by Miller (22).
Sucrose gradients. Sucrose gradients (5 to 20%) in0.02 M potassium acetate buffer (pH 5.1) containing0.1 mM ethylenediaminetetraacetate were formed innitrocellulose centrifuge tubes (1 by 3 inches [ca. 2.54by 5.62 cm]) using a Beckman gradient former. RNAsamples of 0.1 to 0.5 ml were layered onto thegradients. The tubes were centrifuged for 21.5 h at25,000 rpm in an SW25.1 rotor at 4 C. The gradientswere fractionated with an Isco model D fractionatorby pumping 60% sucrose into the bottom of each tubeand collecting from the top.
Liquid scintillation counting. [3H ]RNA was mea-sured by precipitation of a 50 !liter sample with cold5% trichloroacetic acid. The precipitates were col-lected on membrane filters (HA 0.45 um; MilliporeCorp.) and washed with cold 5% trichloroacetic acid.The filters were dried and immersed in toluene-Spec-trofluor (Amersham-Searle) scintillation fluid. Ni-trocellulose filters used for hybridizations were driedsimilarly and immersed in scintillation fluid. Radio-activity was determined with a Nuclear-Chicago Iso-cap 300 scintillation counter. The counting efficiencyfor 3H was about 50%.
Reagents. Thiamine, ribonuclease A, deoxyribonu-clease and L-leucine were obtained from Sigma Chem-ical Co. L-Isoleucine, L-valine, L-histidine, L-trypto-phan, and L-methionine were obtained from Calbio-chem. Poly(U,G) was obtained from Miles Labora-tory. Biochemical-grade CsCl was obtained fromGallard Schlessinger Chemical Co. Optical-gradeCsCl, sodium dodecyl sarcosine, and density gradientgrade sucrose were obtained from Schwarz/Mann.
RESULTSStandardization of the hybridization pro-
cedures. A number of DNA and RNA satura-tion studies were performed to insure the valid-
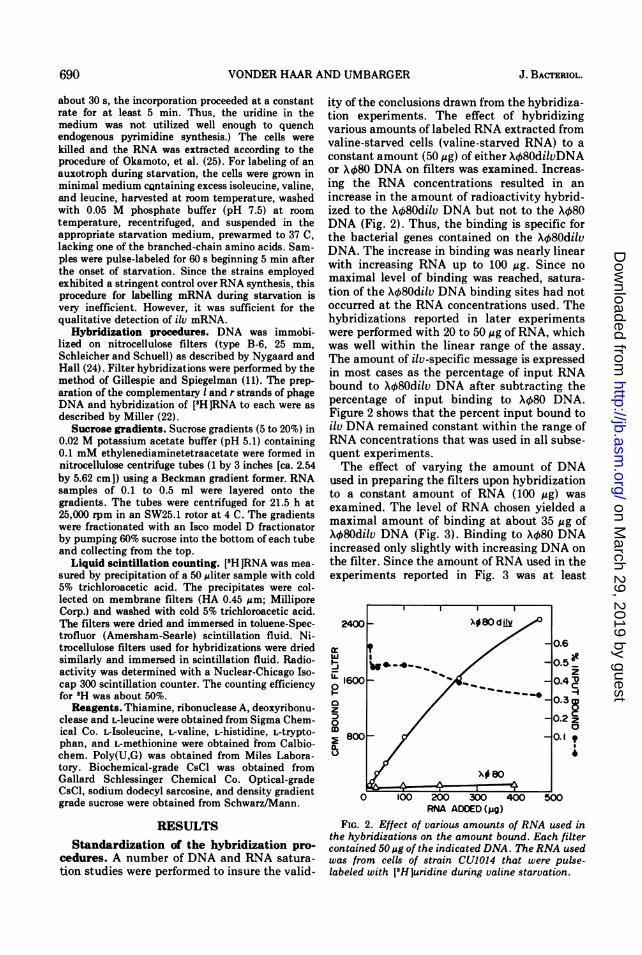
ity of the conclusions drawn from the hybridiza-tion experiments. The effect of hybridizingvarious amounts of labeled RNA extracted fromvaline-starved cells (valine-starved RNA) to aconstant amount (50 ug) of either X080dilvDNAor X480 DNA on filters was examined. Increas-ing the RNA concentrations resulted in anincrease in the amount of radioactivity hybrid-ized to the X480dilv DNA but not to the X480DNA (Fig. 2). Thus, the binding is specific forthe bacterial genes contained on the Xk80dilvDNA. The increase in binding was nearly linearwith increasing RNA up to 100 Mg. Since nomaximal level of binding was reached, satura-tion of the Xk80dilv DNA binding sites had notoccurred at the RNA concentrations used. Thehybridizations reported in later experimentswere performed with 20 to 50 Mg of RNA, whichwas well within the linear range of the assay.The amount of ilv-specific message is expressedin most cases as the percentage of input RNAbound to X80dilv DNA after subtracting thepercentage of input binding to X080 DNA.Figure 2 shows that the percent input bound toilv DNA remained constant within the range ofRNA concentrations that was used in all subse-quent experiments.The effect of varying the amount of DNA
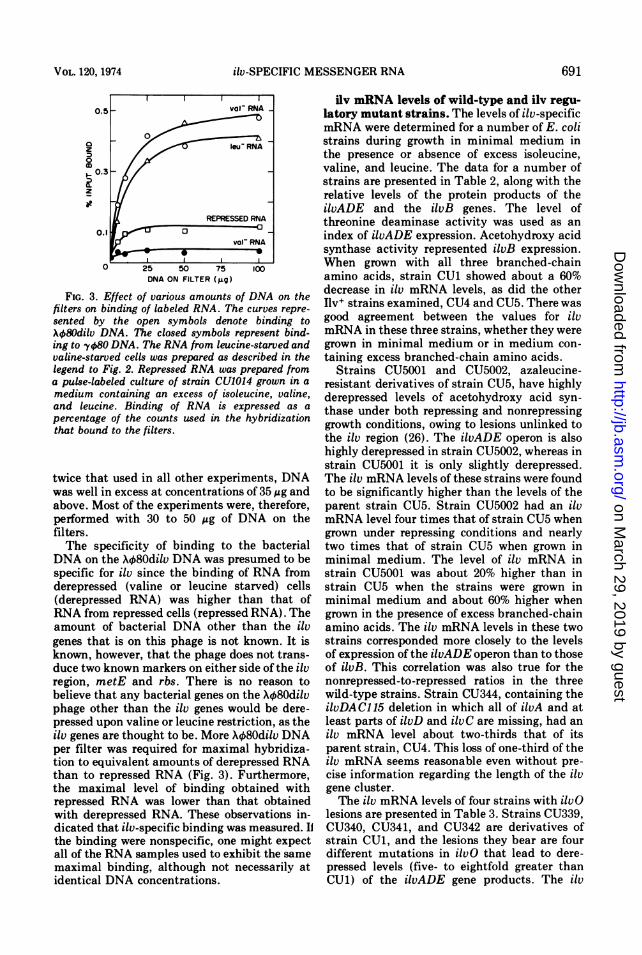
used in preparing the filters upon hybridizationto a constant amount of RNA (100 Mg) wasexamined. The level of RNA chosen yielded amaximal amount of binding at about 35 MAg ofXq080dilv DNA (Fig. 3). Binding to Xk80 DNAincreased only slightly with increasing DNA onthe filter. Since the amount of RNA used in theexperiments reported in Fig. 3 was at least
2400 - X080dilv
-0.6
tj -..0.5
U.1-0 0-40o ~~~~~~~~~~0.30 -~~~~~~~~~~0.2Z800_0.C
RNA ADDED (gig)FIG. 2. Effect of various amounts of RNA used in
the hybridizations on the amount bound. Each filtercontained 50Mug of the indicated DNA. The RNA usedwas from cells of strain CU1014 that were pulse-labeled with [3H]uridine during valine starvation.
690 J. BACTERIOL.
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
ilv-SPECIFIC MESSENGER RNA
og 0.3 IClelu RNAz
0
0.3-
REPRESSED RNA
0.1 OW ~~~~~~vol-RNA
0 25 50 75 100DNA ON FILTER (p9g)
FIG. 3. Effect of various amounts of DNA on thefilters on binding of labeled RNA. The curves repre-sented by the open symbols denote binding toX080dilv DNA. The closed symbols represent bind-ing to y080 DNA. The RNA from leucine-starved andvaline-starved cells was prepared as described in thelegend to Fig. 2. Repressed RNA was prepared froma pulse-labeled culture of strain CU1014 grown in a
medium containing an excess of isoleucine, valine,and leucine. Binding of RNA is expressed as a
percentage of the counts used in the hybridizationthat bound to the filters.
twice that used in all other experiments, DNAwas well in excess at concentrations of 35 ,ug andabove. Most of the experiments were, therefore,performed with 30 to 50 gg of DNA on thefilters.The specificity of binding to the bacterial
DNA on the VX80dilv DNA was presumed to bespecific for ilv since the binding of RNA fromderepressed (valine or leucine starved) cells(derepressed RNA) was higher than that ofRNA from repressed cells (repressed RNA). Theamount of bacterial DNA other than the ilvgenes that is on this phage is not known. It isknown, however, that the phage does not trans-duce two known markers on either side of the ilvregion, metE and rbs. There is no reason tobelieve that any bacterial genes on the X480dilvphage other than the ilv genes would be dere-pressed upon valine or leucine restriction, as theilv genes are thought to be. More X480dilv DNAper filter was required for maximal hybridiza-tion to equivalent amounts of derepressed RNAthan to repressed RNA (Fig. 3). Furthermore,the maximal level of binding obtained withrepressed RNA was lower than that obtainedwith derepressed RNA. These observations in-dicated that ilv-specific binding was measured. IIthe binding were nonspecific, one might expectall of the RNA samples used to exhibit the samemaximal binding, although not necessarily atidentical DNA concentrations.
ilv mRNA levels of wild-type and ilv regu-latory mutant strains. The levels of ilv-specificmRNA were determined for a number of E. colistrains during growth in minimal medium inthe presence or absence of excess isoleucine,valine, and leucine. The data for a number ofstrains are presented in Table 2, along with therelative levels of the protein products of theilvADE and the ilvB genes. The level ofthreonine deaminase activity was used as anindex of ilvADE expression. Acetohydroxy acidsynthase activity represented ilvB expression.When grown with all three branched-chainamino acids, strain CUi showed about a 60%decrease in ilv mRNA levels, as did the otherIlv+ strains examined, CU4 and CU5. There wasgood agreement between the values for ilvmRNA in these three strains, whether they weregrown in minimal medium or in medium con-taining excess branched-chain amino acids.
Strains CU5001 and CU5002, azaleucine-resistant derivatives of strain CU5, have highlyderepressed levels of acetohydroxy acid syn-thase under both repressing and nonrepressinggrowth conditions, owing to lesions unlinked tothe ilv region (26). The ilvADE operon is alsohighly derepressed in strain CU5002, whereas instrain CU5001 it is only slightly derepressed.The ilv mRNA levels of these strains were foundto be significantly higher than the levels of theparent strain CU5. Strain CU5002 had an ilvmRNA level four times that of strain CU5 whengrown under repressing conditions and nearlytwo times that of strain CU5 when grown inminimal medium. The level of ilv mRNA instrain CU5001 was about 20% higher than instrain CU5 when the strains were grown inminimal medium and about 60% higher whengrown in the presence of excess branched-chainamino acids. The ilv mRNA levels in these twostrains corresponded more closely to the levelsof expression of the ilvADE operon than to thoseof i1vB. This correlation was also true for thenonrepressed-to-repressed ratios in the threewild-type strains. Strain CU344, containing theilvDAC115 deletion in which all of ilvA and atleast parts of ilvD and ilvC are missing, had anilv mRNA level about two-thirds that of itsparent strain, CU4. This loss of one-third of theilv mRNA seems reasonable even without pre-cise information regarding the length of the ilvgene cluster.The ilv mRNA levels of four strains with ilvO
lesions are presented in Table 3. Strains CU339,CU340, CU341, and CU342 are derivatives ofstrain CU1, and the lesions they bear are fourdifferent mutations in ilvO that lead to dere-pressed levels (five- to eightfold greater thanGUi) of the ilvADE gene products. The ilv
VOL. 120, 1974 691
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
VONDER HAAR AND UMBARGER
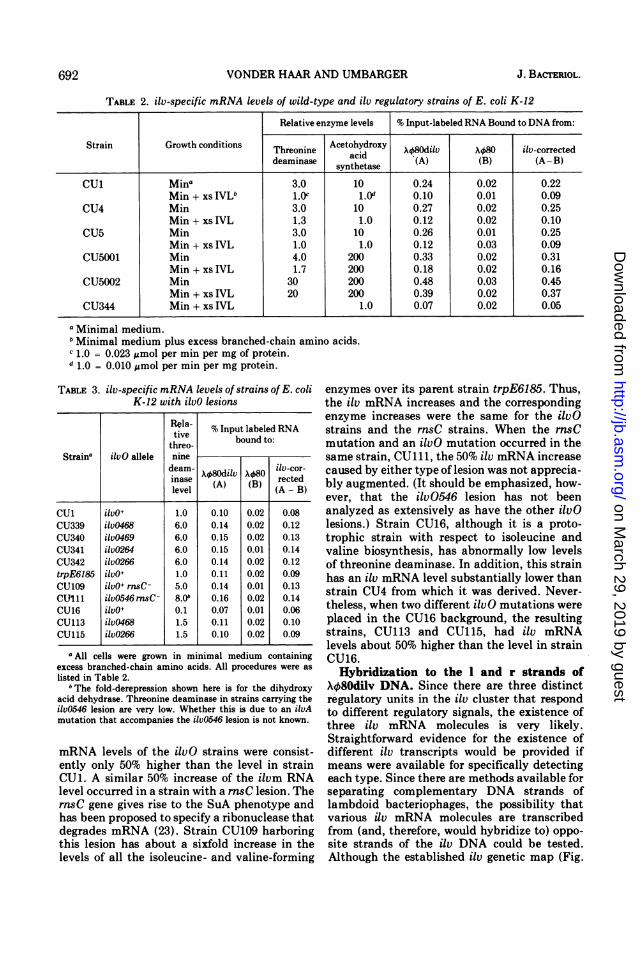
TABLE 2. ilv-specific mRNA levels of wild-type and ilv regulatory strains of E. coli K-12
Relative enzyme levels % Input-labeled RNA Bound to DNA from:
Strain Growth conditions Threonine Acetohydroxy X480di1v X48 ilv-correcteddeaminase synthetase (A) (B) (A- B)
Cui Mina 3.0 10 0.24 0.02 0.22Min + xs IVLU L.oc 1.0d 0.10 0.01 0.09
CU4 Min 3.0 10 0.27 0.02 0.25Min + xs IVL 1.3 1.0 0.12 0.02 0.10
CU5 Min 3.0 10 0.26 0.01 0.25Min + xs IVL 1.0 1.0 0.12 0.03 0.09
CU5001 Min 4.0 200 0.33 0.02 0.31Min + xs IVL 1.7 200 0.18 0.02 0.16
CU5002 Min 30 200 0.48 0.03 0.45Min + xs IVL 20 200 0.39 0.02 0.37
CU344 Min + xs IVL 1.0 0.07 0.02 0.05
a Minimal medium.'Minimal medium plus excess branched-chain amino acids.C 1.0 = 0.023 Amol per min per mg of protein.d 1.0 - 0.010 Mmol per min per mg protein.
TABLE 3. ilv-specific mRNA levels of strains of E. coliK-12 with ilvO lesions
Rela- % Input labeled RNA
tireo- bound to:Straina ilvO allele nine
deam- A080dilv A080 ilv-cor-inase (A) (B) (Ae-tB)level(A-B
Cui ilvO+ 1.0 0.10 0.02 0.08CU339 ilv0468 6.0 0.14 0.02 0.12CU340 ilv0469 6.0 0.15 0.02 0.13CU341 ilv0264 6.0 0.15 0.01 0.14CU342 ilv0266 6.0 0.14 0.02 0.12trpE6185 ilvO+ 1.0 0.11 0.02 0.09CU109 ilvO+ rnsC- 5.0 0.14 0.01 0.13CUI11 ilvO546 msC- 8.01 0.16 0.02 0.14CU16 ilvO+ 0.1 0.07 0.01 0.06CU113 ilv0468 1.5 0.11 0.02 0.10CU115 ilv0266 1.5 0.10 0.02 0.09
a All cells were grown in minimal medium containingexcess branched-chain amino acids. All procedures were aslisted in Table 2.
'The fold-derepression shown here is for the dihydroxyacid dehydrase. Threonine deaminase in strains carrying theilv0546 lesion are very low. Whether this is due to an ilvAmutation that accompanies the ilv0546 lesion is not known.
mRNA levels of the ilvO strains were consist-ently only 50% higher than the level in strainCUi. A similar 50% increase of the ilvm RNAlevel occurred in a strain with a rnsC lesion. ThernsC gene gives rise to the SuA phenotype andhas been proposed to specify a ribonuclease thatdegrades mRNA (23). Strain CU109 harboringthis lesion has about a sixfold increase in thelevels of all the isoleucine- and valine-forming
enzymes over its parent strain trpE6185. Thus,the ilv mRNA increases and the correspondingenzyme increases were the same for the ilvOstrains and the rnsC strains. When the rnsCmutation and an ilvO mutation occurred in thesame strain, CU111, the 50% ilv mRNA increasecaused by either type of lesion was not apprecia-bly augmented. (It should be emphasized, how-ever, that the ilv0546 lesion has not beenanalyzed as extensively as have the other ilvOlesions.) Strain CU16, although it is a proto-trophic strain with respect to isoleucine andvaline biosynthesis, has abnormally low levelsof threonine deaminase. In addition, this strainhas an ilv mRNA level substantially lower thanstrain CU4 from which it was derived. Never-theless, when two different ilvO mutations wereplaced in the CU16 background, the resultingstrains, CU113 and CU115, had ilv mRNAlevels about 50% higher than the level in strainCU16.
Hybridization to the 1 and r strands ofXk80dilv DNA. Since there are three distinctregulatory units in the ilv cluster that respondto different regulatory signals, the existence ofthree ilv mRNA molecules is very likely.Straightforward evidence for the existence ofdifferent ilv transcripts would be provided ifmeans were available for specifically detectingeach type. Since there are methods available forseparating complementary DNA strands oflambdoid bacteriophages, the possibility thatvarious ilv mRNA molecules are transcribedfrom (and, therefore, would hybridize to) oppo-site strands of the ilv DNA could be tested.Although the established ilv genetic map (Fig.
J. BACTERIOL.692
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ilv-SPECIFIC MESSENGER RNA
1) shows that all three operons have their''control elements" to the same side of thestructural genes (thus being transcribed in thesame direction and from the same strand),recent genetic linkage data have cast some
doubt on this map order (16). The possibilitythat the separate I and r strands of X080dilvDNA could each hybridize with ilv mRNA wasexamined not only for the possible developmentof separate assays for specific ilv messages, butalso as an indirect check on the direction oftranscription of the ilv genes.
Separate I and r strands were prepared fromXA80dilv, X480, and Xk80dlac DNA, and hy-bridizations were performed. The RNA samplesused were extracted from cells starved for eitherisoleucine, leucine, or valine. Such cells wouldpresumably be derepressed for the expression ofilvADE alone, ilvB and ilvADE, or ilvB, ilUvC,and ilvADE, respectively (9). RNA from strainCUi grown in minimal medium, which shouldcontain substantial amounts of each type ofmessage, and RNA from-valine-inhibited cellsof strain CU1, which contains high levels ofilvADE message, were also used. RNA wasprepared as described in Materials andMethods. The amount of radioactivity bound toeach strand of X480 DNA or Xb80d1ac DNA wassubtracted from the amount of binding to thesame strand of X480dilv DNA to determine theilv-specific binding. All of the ilv-specificmRNA from every sample tested bound specifi-cally to the I-strand of Xq80dilv DNA (Table 4).Since each presumed type of ilv mRNA wasrepresented in at least two of these RNA sam-ples, it is likely that all the ilv-specific mRNA istranscribed from the same strand in vivo.Therefore, the direction of transcription of allthe ilv genes appears to be the same. Recentwork has shown that induction of the ilvC gene
in strain CUi by a-acetohydroxy butyrate re-sults in ilv-specific mRNA that hybridizespreferentially to the I strand of ilv DNA (A. L.Williams, R. A. Vonder Haar, and H. E. Um-barger, Abstr. Annu. Meet. Amer. Soc. Micro-biol., p. 171, 1974).
Sucrose gradient analysis of ilv-specificmRNA. If distinct species of ilv mRNA doindeed exist, it is possible that they are ofdifferent lengths and, therefore, separable bysucrose gradient centrifugation. The sucrose
gradients were prepared, centrifuged, and frac-tionated. Each fraction was analyzed for ilv-specific mRNA by filter hybridization toXb80dilv DNA (5 to 10 Asg ofDNA per filter) andto AX80 DNA. The RNA samples used wereprepared from cells starved for either isoleucine,valine, or leucine. Figures 4, 5, and 6 show theOD,,*, and ilv-specific mRNA profiles of thesucrose density gradient centrifugations of thesethree RNA samples. When isoleucine-starvedRNA was centrifuged (Fig. 4), a single band ofilv-specific mRNA was detected. This band,presumably the ilvADE mRNA, was at a posi-tion in the gradient corresponding to 18.8S.Centrifugation of the leucine-starved RNA (Fig.5) resulted in two bands of ilv-specific mRNA,one of which was 18.4S, and a more slowlysedimenting band which was about 12S. Themore rapidly sedimenting species was probablythe ilvADE message and the slower one wasprobably the ilvB message. The ilvC messagethat should be present in the valine-starvedRNA may have co-sedimented with the ilvBmessage. Presumptive support for such co-sedimentation can be obtained by comparingthe heights of the peaks corresponding to ilvBmRNA and ilvADE mRNA in Fig. 5 and 6.However, the difference between the two figuresis not striking. Indeed, the possibility that the
TABLE 4. Hybridizations to separated strands of X48Odilv DNA and Xb80dlac DNAa
Counts/min bound to:Presumed
['H]RNA sample iv-mRNA I strand I strand rilv sstrand r strand r-ilvPresent Of of speific Of of seiiX810dilv 8Odlac scific XA iv A80dac specific
CUi during valine inhibi-tion ................... ilvADE 347 44 323 44 36 8
CU1014, isoleucine starved ilvADE 680 168 512 100 108 (-8)CU1014, leucine starved .. ilvADE and ilvB 369 90 279 63 67 (-4)CU1014, valine starved ... ilvADE, ilvB, and
ilvC 614 91 523 65 76 (-9)CU1, minimal grown ...... ilvADE, ilvB, and
ilvC 1594 404 1190 243 349 (-86)
a Similar results were obtained when AX80 DNA was used in place of X080d1ac DNA.
VOL. 120, 1974 693
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

VONDER HAAR AND UMBARGER
75.75 ~~~~~~~~~~~~~.75~
.50/- .500
.25 I.25
° 10 20 30TOP BOTM
FIG. 4. Sucrose gradient centrifugation of RNAfrom isoleucine-starved cells of strain CU1014. Su-crose gradients (5 to 20%) were centrifuged for 21.5 hat 25,000 rpm.
I I
FIG. 5. Sucrose gradient centrifugation of RNAfrom leucine starved cells of strain CU1014. Centrifu-gation was as indicated in the legend to Fig. 4.
' -
1.00 - 2.0
.75 21.
"mssng ivC mRN ispr fteivD
025 .50
010 _20 30TOP FRACTION NUMBER sro
FIG. 6. Sucrose gradient centrifugation of RNAfrom valine-starved cells of strain CU1O14. Centrifu-gation was as indicated in the legend to Fig. 4.
"fmissing" ilvC mRNA is part of the ilvADEtranscript should also be considered.
DISCUSSIONThe results presented in this report demon-
strate the detection and estimation of ilv-specific mRNA made under a variety of physio-logical conditions. Several wild-type and ilv
regulatory strains were monitored for their ilvmRNA levels under both repressing and non-repressing growth conditions. Most strains stud-ied responded as expected to the repressionsignal generated by excess branched-chainamino acids, in that their ilv mRNA levels werelowered. In addition, regulatory mutantsshowed levels of ilv mRNA higher than in theirwild-type parents. (It may be particularly sig-nificant that the presumed "operator" ilvOmutants showed less increase in ilv mRNA thandid the azl strain CU5002.) These results indi-cated that repression of the ilv genes is exerted,at least in part, at the level of transcription. Thefact that ilv mRNA differences were usually notas striking as corresponding enzyme differencesis not without precedent. For example, trpmRNA has been found to be more efficientlytranslated immediately upon derepression thanit is during steady-state conditions of derepres-sion (5). It is possible that mRNA molecules canbe acted upon specifically to result in enzymeincreases greater than the corresponding mRNAincrease. It should be emphasized, however, thata significant portion of the mRNA measured as"ilv-specific" in this study could be mRNAhybridizing to other, unidentified E. coli genescarried by the X480diiv phage that would maskmuch of the control over "true ilv mRNA."
According to the model for regulation of theilv genes put forward by Hatfield and Burns(12), threonine deaminase, the first enzyme ofthe isoleucine pathway, is involved in ilv regula-tion. However, a clear-cut role for threoninedeaminase in control of transcription does notemerge from the limited amount ofmRNA datafrom strains with altered ilvA gene products.Strains CU344 and CU345 are strains with theentire ilvA gene and at least parts of the ilvDand ilvC genes deleted. Neither of these strainsappeared to have more ilv mRNA than would beexpected in strains that have a large portion ofthe ilv cluster missing. (The data for strainCU345 was not shown.) If ilvA specified sometype of transcriptional repression element, theilv mRNA levels in these strains might havebeen derepressed.The sucrose gradient analyses demonstrated
the existence of at least two distinct ilv mRNAtranscripts, one about 18.8S and one about12.2S. Since the molecular weights of all of theenzymes of the isoleucine-valine pathways in E.coli are not known, accurate calculations cannotbe made for the expected sizes of the mRNAfrom the three ilv operons. Estimates of 22S forthe ilvADE message, 14S for the ilvC message,and 14.3S for the ilvB message can be obtainedfrom the molecular weights of the same or
694 J. BACTERIOL.
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ilv-SPECIFIC MESSENGER RNA
similar enzymes from other organisms. The factthat the computed values are slightly higherthan the actual values may have resulted from anumber of factors. The secondary structure orthe amount of hydration of the RNA moleculesmay have caused a slower sedimentation rate.The value assigned as the actual value of theRNA in the band was measured at the center ofthe peak in the graphical representation. Per-haps only the leading edge of the band con-tained the completed messages and the rest ofthe band consisted of smaller trailing pieces ofmRNA that had not been completed or hadbeen enzymatically cleaved. If one used theleading edges of the peaks to determine theexperimental values, they are closer to theexpected values.Although the established genetic map of the
ilv region places the ilvADE operon adjacent toits presumed repression recognition site, theilvO gene, some recent genetic data haveplaced the ilvC gene between ilvA and ilvO (16).This placement of the ilvC gene between theilvADE operon and its presumed operator geneand the fact that all the ilv genes are tran-scribed in the same direction, present a problemin explaining the observation that the ilvC geneand the ilvADE operon are expressed independ-ently of each other. If transcriptional control ofboth ilvC and ilvADE is exerted at sites lying tothe right of ilvC, there must exist some mecha-nism for the differential control of RNA polym-erase activity as it proceeds along the operons.Another possibility that should be considered
is that ilvC may be transcribed onto the samemRNA molecule as iIvADE and that differen-tial control of expression of the ilvC gene andthe ilvADE operon is exerted at the level oftranslation. Translational control is found incertain bacteriophages with RNA genomes inwhich the secondary structure of the RNAmolecule determines the proteins that are made(20). The sucrose gradient analysis of RNA fromvaline-starved cells, which is the only sampleanalyzed on sucrose gradients that should haveappreciable amounts of ilvC message, showedthat the component presumed to be the ilvADEtranscript sedimented slightly faster than didthe corresponding component in the isoleucineor leucine-starved samples. However, it did notsediment nearly as fast as the,predicted mRNAmolecule corresponding to the ilvADE and ilvCgenes. The possibility that the ilvADE and ilvC(and even i1vB) genes are transcribed into asingle molecule which is rapidly degraded toilvADE and ilvC (and ilvB) messages is notexcluded. It is interesting, however, that strainswith an rnsC lesion, which lack an endonucle-
ase, have derepressed levels of the ilv enzymes,perhaps owing to increased stability and trans-lational efficiency of some polycistronic mes-sage. The nature of the ilvC gene message willbe ascertained with greater certainty when ananalysis is made of the RNA under conditions ofilvC induction and ilvADE and ilvB generepression. For the present, we can only con-clude that ilvADE and ilvB occur in separatetranscripts.
ACKNOWLEDGMENTSThis investigation was supported by Public Health Service
grant GM 12522 from the National Institute of GeneralMedical Sciences. R.A.V.H. was a Public Health Servicepredoctoral trainee, biochemistry training grant 5 T0l GM1195 from the National Institute of General Medical Science.
LITERATURE CITED1. Adams, M. H. 1959. Bacteriophages. Interscience, New
York.2. Arfin, S. M., B. J. Ratzkin, and H. E. Umbarger. 1969.
The metabolism of isoleucine and valine in Escherichiacoli. XVII. The role of induction in the derepression ofacetohydroxy acid isomeroreductase. Biochem. Bio-phys. Res. Commun. 37:902-908.
3. Attardi, G., S. Naono, J. Rouviere, F. Jacob, and F. Gros.1963. Production of messenger RNA and regulation ofprotein synthesis. Cold Spring Harbor Symp. Quant.Biol. 28:363-372.
4. Avitabile, A., M. S. Carlomagne-Cerillo, R. Favre, and F.Blasi. 1972. Isolation of transducing bacteriophages forthe histidine and isoleucine-valine operons in Esche-richia coli K-12. J. Bacteriol. 112:40-47.
5. Baker, R., and C. Yanofsky. 1972. Tryptophan initiationfrequency and translational yield for the tryptophanoperon of Escherichia coli. J. Mol. Biol. 69:89-102.
6. Beckwith, J. R., and E. R. Signer. 1966. Transposition ofthe lac region of Escherichia coli. I. Inversion of the lacoperon and transduction of lac by 080. J. Mol. Biol.19:254-265.
7. Cunin, R., and N. Glansdorff. 1971. Messenger RNA fromarginine and phosphoenolcarboxylase genes in argR+and argR- strains of E. coli K-12. FEBS Lett.18:135-137.
8. Davis, B. D., and E. S. Mingioli. 1950. Mutants ofEscherichia coli requiring methionine or vitamin B,,. J.Bacteriol. 60:17-28.
9. Dwyer, S. B., and H. E. Umbarger. Isoleucine and valinemetabolism in Escherichia coli. XVI. Pattern of mul-tivalent repression in strain K-12. J. Bacteriol.95:1680-1684.
10. Freundlich, M., R. 0. Burns, and H. E. Umbarger. 1962.Control of isoleucine, valine and leucine biosynthesis. I.Multivalent repression. Proc. Nat. Acad. Sci. U.S.A.48:1804-1808.
11. Gillespie, D., and S. Spiegelman. 1965. A quantitativeassay for DNA-RNA hybrids with DNA immobilized ona membrane. J. Mol. Biol. 12:829-842.
12. Hatfield, G. W., and R. 0. Burns. 1970. Specific bindingof leucyl transfer RNA to an immature form of L-threo-nine deaminase: its implications in repression. Proc.Nat. Acad. Sci. U.S.A. 66:1027-1035.
13. Hill, C. W., and H. Echols. 1966. Properties of a mutantblocked in inducibility of messenger RNA for thegalactose operon of Escherichia coli. J. Mol. Biol.19:38-51.
14. Imamoto, F., and C. Yanofsky. 1967. Transcription of the
VOL 120, 1974 695
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

VONDER HAAR AND UMBARGER
tryptophan operon in polarity mutants of Escherichiacoli. 11. Evidence for normal production of tryp-mRNAmolecules and for premature termination of transcrip-tion. J. Mol. Biol. 28:25-35.
15. Jacob, F., and J. Monod. 1961. Genetic regulatory mech-anisms in the synthesis of proteins. J. Mol. Biol.3:318-356.
16. Kline, E. L., C. S. Brown, W. G. Coleman, Jr., and H. E.Umbarger. 1974. Regulation of isoleucine-valine bio-synthesis in an ilvDAC deletion strain of Escherichiacoli K-12. Biochem. Biophys. Res. Commun.57:1144-1151.
17. Lavalle, R., and G. DeHauwer. 1970. Tryptophan mes-senger translation in Escherichia coli. J. Mol. Biol.51:435-447.
18. Layne, E. 1957. Spectrophotometric and turbidimetricmethods of measuring proteins, p. 448-451. In S. P.Colowick and N. 0. Kaplan (ed.), Methods in enzymol-ogy, vol. 3. Academic Press Inc., New York.
19. Lennox, E. S. 1955. Transduction of linked geneticcharacters of the host by bacteriophage P1. Virology1:190-206.
20. Lodish, H. F., and N. D. Zinder. 1966. Mutants of thebacteriophage f2. VIII. Control mechanisms for phage-specific syntheses. J. Mol. Biol. 19:333-348.
21. McLellan, W. L., and H. J. Vogel. 1970. Translationalrepression in the arginine system of Escherichia coli.Proc. Nat. Acad. Sci. U.S.A. 67:1703-1709.
22. Miller, J. H. 1972. Experiments in molecular genetics.Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
23. Morse, D. E., and P. Primakoff. 1970. Relief of polarity inE. coli by "suA." Nature (London) 226:28-31.
24. Nygaard, A. P., and B. D. Hall. 1963. A method for thedetection of RNA-DNA complexes. Biochem. Biophys.Res. Commun. 12:98-104.
25. Okamoto, K., Y. Sugino, and M. Momura. 1962. Synthe-sis and turnover of phage messenger RNA in E. coliinfected with bacteriophage T4 in the presence ofchloromycetin. J. Mol. Biol. 5:527-534.
26. Pledger, W. J., and H. E. Umbarger. 1973. Isoleucineand valine metabolism in Escherichia coli. XXI. Muta-tions affecting derepression and valine resistance. J.Bacteriol. 114:183-194.
27. Press, R., N. Glansdorff, P. Miner, J. DeVries, R.Kadner, and W. K. Maas. 1971. Isolation of transduc-ing particles of N80 bacteriophage that carry differentregions of the Escherichia coil genome. Proc. Nat.Acad. Sci. U.S.A. 68:795-798.
28. Ramakrishnan, T., and E. A. Adelberg. 1965. Regulatorymechanisms in the biosynthesis of isoleucine andvaline. m. Map order of the structural genes andoperator genes. J. Bacteriol. 89:661-664.
29. Rogers, P., R. Krzycek, T. M. Kaden, and E. Afrman.1971. Effect of arginine and canavanine on argininemessenger RNA synthesis. Biochem. Biophys. Res.Commun. 44:1220-1226.
30. Taylor, A. L., and C. D. Trotter. 1972. Linkage map ofEscherichia coli strain K-12. Bacteriol. Rev.36:504-524.
31. Thomas, C. A., Jr., and J. Abelson. 1966. In G. L. Cantoniand D. R. Davis (ed.), Procedures in nucleic acidresearch, vol. 1, p. 553-556. Harper and Row, NewYork.
32. Vonder Haar, R. A., and H. E. Umbarger. 1972. Isoleu-cine and valine metabolism in Escherichia coli. XIX.Inhibition of isoleucine biosynthesis by glycyl-leucine.J. Bacteriol. 112:142-147.
33. Wasmuth, J. J., and H. E. Umbarger. 1973. Effect ofisoleucine, valine or leucine starvation on the potentialfor formation of the branched-chain amino acid biosyn-thetic enzymes. J. Bacteriol. 116:548-561.
696 J. BArRIOL.
on March 29, 2019 by guest
http://jb.asm.org/
Dow
nloaded from






![[Frenkel.] Vertex Algebras (Bourbaki org](https://img.pdfslide.us/doc/110x75/577d24401a28ab4e1e9bfeba/frenkel-vertex-algebras-bourbaki-org.jpg)






















