Embed Size (px)
Citation preview

Kirill B. Gromadski and Marina V. Rodnina
Biochemistry 4000 Dora Capatos
Kinetic Determinants of High-Fidelity Discrimination on
the Ribosome

tRNA Selection
Ribosome selects aminoacyl transfer RNA (aa-tRNA) with anticodon matching to the mRNA codon in the A site from the bulk of nonmatching aa-tRNAs
30S subunit
50S subunit

Mismatches
• Cognate tRNA: matches codon in the decoding site
• Near-cognate tRNA: one mismatched base pair
• Frequency of mismatch is 10-3 to 10-4

tRNA Discrimination on the Ribosome
• Rejection of incorrect tRNAs occurs in 2 stages:
1. Initial selection of ternary complexes EF-Tu-GTP-aa-tRNA
2. Proofreading of aa-tRNA

What is Initial Selection?
• Steps of codon recognition and GTPase activation• Codon recognition occurs when the first codon-anticodon
base pair is stabilized by binding of the rRNA A1493 base pair’s minor groove in the decoding centre
• These interactions enable the ribosome to monitor whether an incoming tRNA is cognate to the codon in the A site.
• A non Watson-Crick base pair could not bind these ribosomal bases in the same way.
• An incorrect codon-anticodon provides insufficient free energy to bind the tRNA to the ribosome and it dissociates from it, still in its ternary complex with EF-Tu and GTP
• Occurs prior to GTP hydrolysis and must be fast

GTPase Activation & Hydrolysis• GTPase activation of EF-Tu• Release of inorganic phosphate induces conformational
transition of EF-Tu from GTP to GDP form• EF-Tu in GDP form loses affinity for aa-tRNA and
dissociates from the ribosome
Mg2+ ion

Accommodation
• After GTP hydrolysis, EF-Tu loses its affinity for aa-tRNA and the aminoacyl end of aatRNA is free to move into the peptidyl transferase centre on the 50S subunit
• tRNA accommodation occurs in the A site• Occurs when EF-Tu hydrolyzes its bound GTP
to GDP + Pi and is released from the ribosome permitting the aa-tRNA to fully bind to the A site

Proofreading
• Proofreading step is independent of the initial selection step
• Proofreading includes the conformational changes that occur after GTP hydrolysis and before peptide bond formation
• Rejection will occur if a mismatch is detected, and the aa-tRNA will dissociate from the ribosome
• Otherwise, peptide bond formation will occur.

The Decoding Problem
Crystal structure of 30S subunit with anticodon stem-loop fragmentsOf tRNA bound to codon triplets in the decoding site show that the codon-anticodon complex forms interactions with rRNA in the decoding site.
Free energy of Watson Crick base pairing alone cannot account for the high efficiency of tRNA selection!

Objective
What are the respective contributions of initial selection and proofreading to tRNA
selection that account for the low error rate of the ribosome?

I. Overall Selectivity
• Measure selectivity of the ribosome at high & low fidelity conditions:– Conditions at which overall fidelity of selection was
high due to high efficiency of both initial selection and proofreading
– Overall selectivity measured by competition between Leu-tRNAleu specific for the CUC codon
– Measure proofreading by

Results: Selectivity of the Ribosome
Since initial selection and proofreading steps are independent:
Probability of Overall Selection = Prob (Initial Selection) x Prob (proofreading)
At high fidelity: 1/450 = (1/30 x 1/15)

Results: Error Rates?
• Contribution of initial selection is calculated from overall selectivity to be about 30. Proofreading was calculated to be about 15.
• Overall selectivity is product of initial selection and proofreading and
• is approximately 450 at high fidelity conditions.
• Incorporation of 1 incorrect per 450 amino acids• This indicates an efficiency of initial selection of 30.

Kinetic Mechanism of EF-Tu-Dependent aa-tRNA Binding

II. Individual Steps of Selection
• Elemental rate constants of the steps contributing to initial selection of ternary complex EF-Tu-Phe-tRNAPhe
(anticodon 3’-AAG-5’) were determined on mRNA programmed (initiated) ribosomes with cognate (UUU) or near-cognate (CUC) codons in the A site.

Individual Steps of Selection
• Monitor GTP hydrolysis & peptide bond formation by quench flow using isotopes [γ-32GTP or aa-tRNA charged with 3H- or 14C-labelled amino acids
• All other rate constants measured by fluorescence experiments carried out by stopped-flow technique (measure conformational changes)
• Fluorophores are wybutine (binds to tRNA) and proflavin

Experimental Setup
• Measure binding or dissociation:
1. Syringe: ribosomes in excess
2. Syringe: Ternary complex
3. tRNA-labelled (fluorescence or radioactive isotope)
4. Use high fidelity buffer conditions (low Mg 2+ concentrations)
5. Do stopped flow or quench flow experiments

Rapid Kinetics
• Apparent rate constants• Do not follow Michaelis Menten Kinetics; must
use mathematical curve fitting to obtain kapparent
• Pre-steady state conditions• Use stopped flow or quench flow device • Single turnover conditions: [TC] << [ribosome] to
ensure that only one round of selection occurs

Initial Binding
•R + TC Complex
•k1 is 2nd Order•K-1 is 1st Order•K1 = 140 +/-20 uM-1 s-1 (slope)
•KM = (k2 + k-1) / k1•KM ~ [ribosome] at ½ Vmax
•Exponential curve Fitting
Kapp Increases linearly with [Ribosome]

Codon Recognition
• Kapp determined from fluorescence increased with ribosome concentration in a hyperbolic shape
• Kapp increased faster for cognate vs. near-cognate tRNAs
•K2 = 190 ± 20 s-1
Near-cognate
Cognate

Chase Experiments
• To a fluorescently labelled Phe-tRNA in complex with GTP and GTPase deficient EF-Tu(H84A), initiate dissociation by adding an excess of nonfluorescent ternary complex and monitor fluorescence decrease over time
• Use GTPase deficient EF-Tu to determine if GTP hydrolysis has an effect on fluorescence

Dissociation of Codon-RecognitionComplex
1 = k-2 = 0.23 ± 0.05 s-1 (Cognate) k-2 ~ 02 = k-2 = 80 ± 15 s-1 (Near-cognate)3 = Control: no dissociation occurs upon addition of buffer instead of non-fluorescent Ternary complex
•Initial binding of ternary complex reversible when there is no match betweencodon and anticodon
•Cognate dissociates very slowly compared to near-cognate
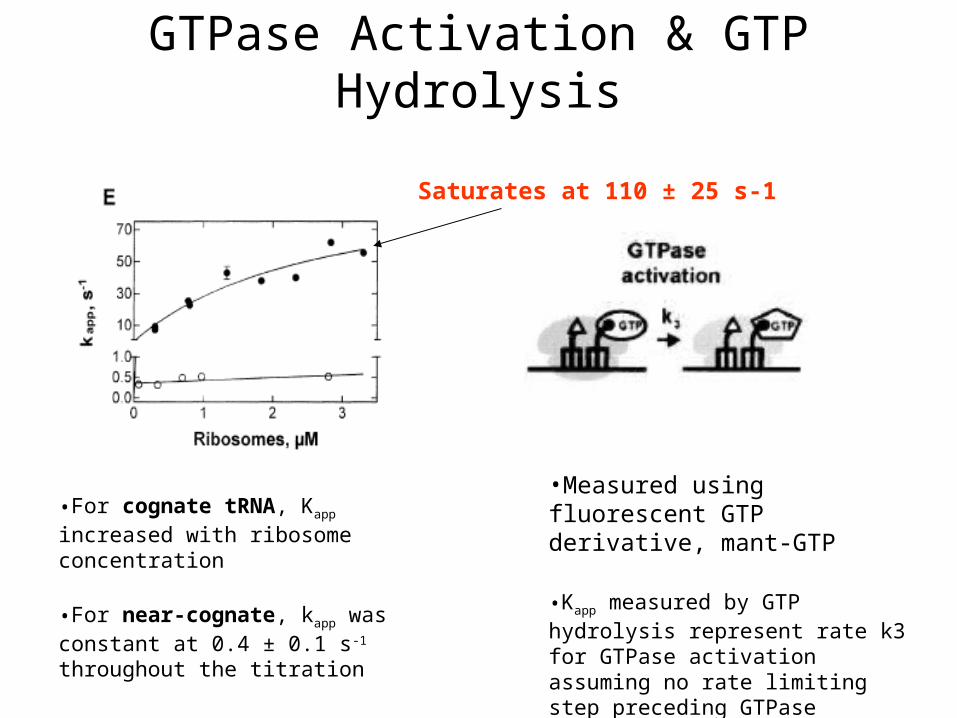
GTPase Activation & GTP Hydrolysis
•Measured using fluorescent GTP derivative, mant-GTP
•Kapp measured by GTP hydrolysis represent rate k3 for GTPase activation assuming no rate limiting step preceding GTPase activation
•For cognate tRNA, Kapp increased with ribosome concentration
•For near-cognate, kapp was constant at 0.4 ± 0.1 s-1 throughout the titration
Saturates at 110 ± 25 s-1

GTPase Activation & GTP Hydrolysis
= absence of ribosomesKapp = 62 +/- 3 s-1 (UUU codon)Kapp = 0.35 +/- 0.02 s-1 (CUC)

Proofreading & Peptide Bond Formation
Kapp = 6.6 +/- 0.4 s-1 (Cognate)Kapp = 0.19 +/- 0.04 s-1 (Near cognate)
Proofreading = fraction of dipeptides that undergo peptidyl transfer = k5 /(k5 + k7)

Kinetic Determinants of Initial Selection
k1, k-1, k2, were for the same for cognate and near-cognate ternarycomplexes, thus the only rate constant that contributes to the different affinity is k-2. So k-2 near cognate /k-2cognate = 80/0.23 ≈ 350.Free energy difference: ∆∆Go = -RTlnk = -RTln(350) = 3.4 kcal/molGTPase activation of EF-Tu is rate limiting for GTP hydrolysis

Kinetic Determinants of Initial Selection
•GTP hydrolysis by EF-Tu regulates initial selection•K3cognate/k3near-cognate = 650 => 650-fold GTP hydrolysis of cognate compared to near cognate •K1 and K2 do not reach equilibruim (would be too slow otherwise)

Cognate vs. Near Cognate Binding
Efficiency of initial selection = Kcat/KmFor cognate tRNA, Kcat = K2

Summary
• Both initial selection prior to and proofreading after GTP hydrolysis are required for efficient tRNA discrimination in vitro.
• Fidelity of initial selection:Finitial selection = 60 ± 20 is close to 30 • Rate constants of GTPase activation and tRNA
accommodation in the A site are much faster for the correct than the incorrect substrates
• k1, k-1, k2, were for the same for cognate and near-cognate ternary complexes

Discussion
• Thermodynamic vs. Kinetic Discrimination?
• tRNA selection at the initial selection step is kinetically controlled and is due to much faster (650-fold) GTP hydrolysis of cognate vs. near-cognate substrate
• Thermodynamic stability differences between cognate and near-cognate tRNAs: RTln350 is the ratio of rate constants: k-2near cognate /k-2cognate and 650 for GTP hydrolysis gives RTln(650) = 2.7 kcal/mol.

Discussion
• An incorrect codon-anticodon provides insufficient free energy to bind the tRNA to the ribosome and it therefore dissociates from it, still in its ternary complex with EF-Tu and GTP bound
• Free energy of base-pairing alone is insufficient to discriminate between cognate (correct) and near-cognate (incorrect) tRNAs
• May differ by as little as a single mismatch in the codon-anticodon duplex

Discussion
• GTPase activation of EF-Tu requires precise alignment of catalytic groups in active sites
• Changes of ribosome structure caused by the correct substrate may not occur or may be different with an incorrect substrate
• Reflect finding that rate constants of GTPase activation and tRNA accomodation in A site are much faster for correct vs. incorrect substrates

Discussion
• A-site binding is a non-equilibrium process that is driven by the rapid irreversible forward reactions of GTP hydrolysis and peptide bond formation
• Discrimination is based on the large differences in the forward reaction rates of GTPase activation and accomodation

Discussion
• Induced Fit Model • Ribosome may be capable of preferential
stabilization of complexes with the correct substrate in both ground state and transition state
• Incorrect substrates may be poorly or not at all stabilized
• Suggests ribosome increases selection potential by checking structure of intermediates by an induced fit mechanism.

Future Questions
• Further structural studies -Solve structure of the codon-anticodon complex in the decoding centre at high resolution
• Investigate induced fit discrimination mechanism of the ribosome
• Structure of conformational changes in proofreading
• Structural determinants that sense cognate base pairing

References
• Gromadski, K.B., Rodnina, M.V. 2004. Mol. Cell 13: 191-200.• Rodnina, M.V., Gromadski, K.B., Kothe, U., Wieden, H. FEBS Lett.
579: 938-942.• Rodnina, M.V., Wintermeyer, W. 2001. TIBS 26 (2): 124-130.• Voet, D., Voet J. 2004. Biochemistry. Wiley, New York.
![Ribosome Stoichiometry: From Form to Function · Ribosome abundance: A major model, also termed the ribosome concentration hypothesis [3], that explains how ribosomes could exert](https://img.pdfslide.us/doc/110x75/60de31e56d30fc4fb30719b8/ribosome-stoichiometry-from-form-to-function-ribosome-abundance-a-major-model.jpg)