Embed Size (px)
Citation preview

Copyright � 2008 by the Genetics Society of AmericaDOI: 10.1534/genetics.107.085795
Genetic Analysis of the Role of Peroxisomes in the Utilization of Acetateand Fatty Acids in Aspergillus nidulans
Michael J. Hynes,1,2 Sandra L. Murray,1 Gillian S. Khew and Meryl A. Davis
Department of Genetics, University of Melbourne, Melbourne, Victoria 3010, Australia
Manuscript received December 11, 2007Accepted for publication January 17, 2008
ABSTRACT
Peroxisomes are organelles containing a diverse array of enzymes. In fungi they are important for carbonsource utilization, pathogenesis, development, and secondary metabolism. We have studied Aspergillusnidulans peroxin (pex) mutants isolated by virtue of their inability to grow on butyrate or by the inactivationof specific pex genes. While all pex mutants are able to form colonies, those unable to import PTS1 proteinsare partially defective in asexual and sexual development. The pex mutants are able to grow on acetate but areaffected in growth on fatty acids, indicating a requirement for the peroxisomal localization of b-oxidationenzymes. However, mislocalization of malate synthase does not prevent growth on either fatty acids oracetate, showing that the glyoxylate cycle does not require peroxisomal localization. Proliferation ofperoxisomes is dependent on fatty acids, but not on acetate, and on PexK (Pex11), expression of which isactivated by the FarA transcription factor. Proliferation was greatly reduced in a farAD strain. A mutationaffecting a mitochodrial ketoacyl-CoA thiolase and disruption of a mitochondrial hydroxy-acyl-CoA de-hydrogenase gene prevented growth on short-chain but not long-chain fatty acids. Together with previousresults, this is consistent with growth on even-numbered short-chain fatty acids requiring a mitochondrial aswell as a peroxisomal b-oxidation pathway. The mitochondrial pathway is not required for growth on valerateor for long-chain fatty acid utilization.
EUKARYOTES contain single membrane organellescalled microbodies containing specialized enzymes
involved in a wide range of metabolic activities. Com-monly these enzymes include oxidases generating hy-drogen peroxide. When this occurs, the microbodiescontain catalase and other antioxidative activities toremove reactive oxygen species and they are termed per-oxisomes. In some organisms and plant tissues, micro-bodies lack catalase but contain enzymes of the glyoxylatebypass and these microbodies are often called glyoxy-somes (for reviews see Titorenko and Rachubinski
2001; Platta and Erdmann 2007). In the filamentousfungus Neurospora crassa, microbodies contain glyoxy-late cycle enzymes but lack catalase as well as peroxi-somal oxidases but also contain enzymes for fatty acidb-oxidationwithacyl-CoAdehydrogenasesubstitutingforacyl-CoAoxidase(KionkaandKunau 1985; Thieringer
and Kunau 1991; Gainey et al. 1992; Schliebs et al.2006). It is unlikely that the two classes of microbodiesdiffer fundamentally and this is supported by the findingin silico of conserved orthologs of the full range ofproteins that constitute peroxisomes (peroxins) in N.crassa (Kiel et al. 2006). This is also likely to be the case forplants (e.g., Nishimura et al. 1986).
Peroxins are proteins required for peroxisome di-vision, for biogenesis from the endoplasmic reticulum,and for the import of proteins into the peroxisomalmatrix (reviewed by Platta and Erdmann 2007). Mu-tations in pex genes can result in the absence of perox-isomes, abnormal peroxisomal structures, mistargetingof matrix proteins, or an inability to respond to stimulithat cause increased numbers of peroxisomes. Twomajor classes of peroxisomal targeting signals occur inmatrix proteins. PTS1 sequences comprise three C-terminal amino acids (aa) usually of the form S/A R/K L/M although the context of the C-terminal sequencecan greatly affect targeting (Brocard and Hartig 2006)and some peroxisomal proteins have cryptic PTS1 se-quences (e.g., Klein et al. 2002). Other matrix proteinshave PTS2 sequences close to the N terminus with theconsensus R/K L/V/I X5 H/Q L/A/F/I (Petriv et al.2004). A large number of peroxins are involved in theimport of all matrix proteins while others are specificto each PTS class; e.g., Pex5 and Pex7 are the specificreceptors for PTS1 and PTS2 proteins, respectively(Lazarow 2006; Stanley and Wilmanns 2006).
Many microorganisms are able to use two carbon com-pounds and fatty acids as sole carbon sources. In fungi,there is increasing interest in the role of lipid catabolismin both animal and plant pathogenesis (e.g., Barelle
et al. 2006; Ramırez and Lorenz 2007; Schobel et al.2007; Wang et al. 2007) as well as in the involvement ofperoxisomes in secondary metabolism and develop-
1These authors contributed equally to this work.2Corresponding author: Department of Genetics, University of Mel-
bourne, Victoria 3010, Australia. E-mail: [email protected]
Genetics 178: 1355–1369 (March 2008)

ment (e.g., Berteaux-Lecellier et al. 1995; Kiel et al.2005; Maggio-Hall et al. 2005).
In Saccharomyces cerevisiae, all steps of b-oxidation arecarried out in peroxisomes, leading to the breakdown oflong-chain fatty acids such as oleate to acetyl-CoA, whichis then utilized via the glyoxylate cycle (Hiltunen et al.2003). PEX mutants are unable to use oleate but are stillable to use acetate or ethanol as carbon sources, in-dicating that the glyoxylate cycle does not depend onperoxisomal localization (Erdmann et al. 1989). Theenzymes isocitrate lyase (ICL) and malate synthase (MAS)are unique to the glyoxylate cycle and are required forgrowth on oleate. ICL is not peroxisomal in S. cerevisiaewhile MAS contains a PTS1 and, during growth on ole-ate, is in peroxisomes where the substrate acetyl-CoA isproduced by b-oxidation. However, mislocalization ofMAS to the cytoplasm does not prevent oleate utilizationand it is cytoplasmic during growth on acetate or eth-anol (Kunze et al. 2002, 2006). Genes specific to oleateutilization, including some PEX genes, are induced byfatty acids via the Oaf1-Pip2 heterodimer (Smith et al.2002, 2007; Gurvitz and Rottensteiner 2006) whileenzymes of the glyoxylate cycle are regulated separatelyby the gluconeogenic control mechanism involving theCat8 and Sip4 activators (reviewed in Schuller 2003).
The filamentous ascomycete, Aspergillus nidulans, isable to grow on both short-chain (C4-6) and long-chainfatty acids as well as on two carbon compounds assole sources of carbon and energy (Armitt et al. 1976;Hynes et al. 2006). Peroxisomes, which contain catalase,proliferate in response to oleate, and some enzymes ofb-oxidation are inducible by oleate (Valenciano et al.1996, 1998). Peroxisomal b-oxidation is indicated by thedeletion of foxA encoding a peroxisomal multifunc-tional enzyme affecting growth on long-chain fatty acids(Maggio-Hall and Keller 2004). The glyoxylate cycleenzymes ICL (encoded by acuD) and MAS (encoded byacuE) have been found to be present in peroxisomes(Gainey et al. 1992; Szewczyk et al. 2001). While AcuEhas a PTS1, AcuD lacks an obvious PTS (Sandeman et al.1991; Gainey et al. 1992). Here we show that, unex-pectedly, AcuD peroxisomal localization is dependenton the PTS2 receptor Pex7(G) and that deletion of aninternal sequence results in mislocalization.
By the isolation and complementation of mutantsunable to grow on butyrate we have discovered that thefarA, farB, and scfA genes specify transcription factorsrequired for fatty acid induction of genes of fatty acidutilization (Hynes et al. 2006). We report here the clon-ing by complementation of further genes required forgrowth on butyrate and identify some of these as pexgenes encoding peroxins. We have also inactivatedthe genes predicted to encode the orthologs of Pex5,the PTS1 receptor: Pex3 required for peroxisome bio-genesis and Pex11 involved in peroxisome prolifera-tion. With GFP-tagged proteins containing PTS1 orPTS2 sequences we have investigated peroxisomal pro-
tein localization in these mutants. The pex mutants havebeen found to affect growth on both short- and long-chain fatty acids. However, growth on acetate is notabolished in the mutants and we have also found thatmislocalization of AcuE (MAS) to the cytoplasm doesnot prevent growth on either fatty acids or acetate,showing that the glyoxylate cycle does not dependon peroxisomal localization. Proliferation of perox-isomes in response to fatty acids but not acetate hasbeen observed and this is dependent on the Pex11ortholog, PexK, and on the fatty acid regulator FarA.Overall, our results show that peroxisomes are dis-pensable for acetate metabolism but crucial for fattyacid utilization.
None of the mutants isolated had mutations in genesencoding peroxisomal enzymes of b-oxidation. How-ever, one mutant affecting growth on short- but notlong-chain fatty acids was predicted to result from a mu-tation affecting a mitochodrial ketoacyl-CoA thiolase.Maggio-Hall and Keller (2004) have demonstratedthe existence of a mitochondrial pathway for theb-oxidation of short-chain fatty acids. We have disrupteda gene for a predicted mitochondrial hydroxy-acyl-CoAdehydrogenase and found that this prevents growth onshort-chain fatty acids. The properties of these mutantsare consistent with growth on even-numbered short-chain fatty acids requiring a mitochondrial as well as aperoxisomal b-oxidation pathway. This mitochondrialpathway is not required for growth on the odd-numberedvalerate (C5) or for long-chain fatty acid utilization.
While the pex mutants form colonies on standardlaboratory media, all mutants affected in PTS1 proteinimport have reduced conidiation and are subfertile inhomozygous sexual crosses. These effects were shown tobe independent of the VelA pathway, which affects thebalance between asexual and sexual development (Kim
et al. 2002; Tsitsigiannis et al. 2004a). The possibilitythat these effects were due to an inability to formWoronin bodies containing the PTS1 HexA proteinwas eliminated by the isolation of a hexA deletion mu-tant that had only a minor conidiation defect and wascapable of normal sexual development. The pex mutantsdefective in PTS1 protein import were found to beauxotrophic for biotin due to an inability to synthesizethe intermediate pimelic acid. This collection of pexmutants now provides valuable tools for the study of therequirements for the correct compartmentalization ofenzymes and metabolite shuttling between the cyto-plasm and organelles during the growth and develop-ment of a filamentous fungus.
MATERIALS AND METHODS
A. nidulans strains, media, and transformation: Media andconditions for growth of A. nidulans were as described by Cove
(1966). Carbon and nitrogen sources were added as appro-priate to minimal salts. The pH of these was adjusted to
1356 M. J. Hynes et al.

6.5 where necessary. Long-chain fatty acids were dispersedin media by the addition of 0.5% tergitol (NP-40, Sigma,St. Louis) before melting or autoclaving. All strains werederived from the original Glasgow strain and contained thevelA1 mutation except where specifically mentioned and stan-dard genetic manipulations were as previously described(Clutterbuck 1974, 1994; Todd et al. 2007). Strains contain-ing the scdAD and echAD mutations were obtained from LoriMaggio-Hall. Preparation of protoplasts and transformationwere as described (Andrianopoulos and Hynes 1988).
Molecular techniques: Standard methods for DNA manip-ulations, nucleic acid blotting, and hybridization have beendescribed (Sambrook et al. 1989; Hynes et al. 2006). For allstrains generated by DNA manipulations followed by trans-formation of A. nidulans, the genome integration events werecharacterized by Southern blotting of restriction-enzyme-digested genomic DNA isolated from transformants.
Bioinformatic analysis: All A. nidulans DNA sequences werederived from the genome sequence at the Broad Institute(http://www.broad.mit.edu/annotation/genome/aspergillus_group/MultiHome.html). Predicted protein-targeting sequen-ces were analyzed by the use of the programs TargetP (http://www.cbs.dtu.dk/services/TargetP/) and Wolf Psort (http://wolfpsort.org/).
Isolation of mutants unable to grow on butyrate andmolecular cloning of genes: Mutants unable to utilize 20 mm
butyrate were isolated as previously described (Hynes et al.2006). For each mutant, it was shown by outcrossing that thephenotype resulted from a mutation in a single gene. PyrG�
double mutants were generated by crossing to a strain contain-ing the pyrG89 mutation and these strains were used forcloning the genes by functional complementation using thegenomic library in the autonomously replicating vectorpRG3AMA1 (Osherov and May 2000) as described (Hynes
et al. 2006). Sequences corresponding to the complementinggenes were recovered and analyzed either through rescue bytransforming into Escherichia coli or by direct PCR on genomicDNA from complementing transformants using primers flank-ing the insert DNA in the vector (Hynes et al. 2006). Sequenceswere compared with the A. nidulans DNA sequence (http://www.broad.mit.edu/annotation/genome/aspergillus_group/MultiHome.html) to identify the mutant genes. The clonedgenes are listed in Table 1 together with the corresponding or-thologs from S. cerevisiae (http://www.yeastgenome.org/). TheA. nidulans system of gene nomenclature has been followedwith letters corresponding to the numbers assigned to the S.cerevisiae genes.
The insert DNA in the complementing clones correspondedto the following coordinates in the genome sequence: pexA,contig 102—84,400–90,000; pexM, contig 23—52,763–58,765;antA, contig 5—338,689–341,912. For pexF, insert DNA from acomplementing clone was subcloned and the complementingsequence identified as corresponding to AN2925.3. In addi-tion, a strain containing a deletion of pexF was constructed byreplacing a BamHI–EcoRI fragment (corresponding to en-coded amino acid coordinates 343–1081) with the pyrG gene asa BamHI–EcoRI fragment (Borneman et al. 2001) and trans-formation of a pyrG89 strain selecting for PyrG1. The resultingstrain had an identical phenotype to the original pexF23 mu-tant and was found to be allelic in crosses between the strains.The pexG14 mutation was complemented by a subclone, cor-responding to contig 14 (47,157–49,109) derived from aninitial complementing plasmid, and contained AN0880.3as the only intact gene. DNA corresponding to the mutantgene was cloned from genomic DNA using the PCR primers59-ACCGTACGTATAAGTCATTCG-39 and 59-AAGAGAGGACCCAGTTGTAG-39 and sequenced, revealing the presence of aT insertion corresponding to codon 261 and resulting in an in-
frame stop codon after a further 18 codons, thereby eliminat-ing two conserved WD domains. Another gene was found to becomplemented by a plasmid containing a 6-kb insert, andsequencing from one end showed that this began at contig 68(18,464) and contained the gene AN10512.3 predicted tospecify a keto-acyl thiolase with a mitochondrial targetingsequence. This gene was designated mthA.
Gene deletions and disruptions: AN10215.3 is predicted toencode an ortholog of Pex5 (Table 1). The gene was clonedusing the PCR primers 59-GTTATATGGGATTGGCGTGG-39and 59-TCCGAATTCCGAGCTCTGC-39, generating a 3.86-kbfragment, which was cloned into pBluescript SK1 (Stratagene,La Jolla, CA). A deletion strain was made by replacing a SphI–SalI fragment (corresponding to amino acids 191–633 of thepredicted protein) with a SphI–XhoI fragment of the riboB gene(Oakley et al. 1987) and transforming the linear insert into ariboB2 strain selecting for RiboB1 transformants. The resultingphenotypes described in the results were complemented bycotransformation experiments using the original clone.AN1921.3 is predicted to encode an ortholog of Pex11 (Table1). The gene was cloned using the PCR primers 59-CATAACTAAGATGTGTCTCTGG-39 and 59-GTGCTGTGTCTGAAATATAGG-39, generating a 2-kb fragment, which was cloned intopGEMTEasy (Promega, Madison, WI). A deletion strain wasmade by replacing a HindIII–XhoI fragment (corresponding toamino acids 27–181 of the predicted protein) with a HindIII–XhoI fragment of the riboB gene and transforming the linearinsert into a riboB2 strain selecting for RiboB1 transformants.AN2281.3 is predicted to encode an ortholog of Pex3 (Table1). A 3.2-kb fragment containing this gene was amplified withthe primers 59-GAAGGGAAAGAATGAGAGA-39 and 59-ACTCAATTGCATCTAGGTCC-39 and inserted into pGEMTEasy.An internal 1-kb XbaI–EcoRI fragment lacking 59 and 39 se-quences of the gene was cloned into SpeI–EcoRI-cut pMT1612containing the bar selectable marker (Nayak et al. 2006). Theresulting plasmid was transformed into an nkuAD strain(Nayak et al. 2006) selecting transformants for glufosinateresistance. Transformants were screened for growth on buty-rate and a transformant showing inhibited growth wasisolated. Southern blot analysis showed that the predicteddisruption event resulting from homologous recombinationhad occurred, yielding two tandem truncated copies of thegene—one lacking 390 bp of 39 coding sequence correspond-ing to the last 129 amino acids and the other lacking 445 bp of59-UTR and the coding sequences corresponding to the first123 amino acids of the predicted protein. The resultingphenotypes described in the results were complementedby cotransformation experiments using the original clonedDNA. AN7008.3 was chosen for disruption because it waspredicted to encode a mitochondrial protein containing 3-hydroxyacyl-CoA dehydrogenase domains. An internal frag-ment of this gene was obtained using the PCR primers59-TATCCGCCCATTGCTAACTC-39 and 59-AGATGCGATCTTGCCCATAC-39 and cloned into a plasmid containing the A.fumigatus riboB gene as a heterologous marker (Nayak et al.2006). This plasmid was transformed into an nkuAD strainselecting for Ribo1 transformants. A transformant showinginhibited growth on short-chain fatty acids was selected forfurther analysis. The predicted homologous disruption eventgenerates truncated genes—one encoding a protein lacking thelast 103 amino acids and one lacking the 59 region and the se-quence corresponding to the first 216 amino acids. AN4695.3 ispredicted to encode the Woronin body protein HexA (see alsoaccession no. AAF67173). A 2.9-kb fragment encompassing thisgene was cloned using the PCR primers 59-CCATCCATGCCTACGATACC-39 and 59-CAGTTCGGGAGAGATTCGAG-39 andinserted into EcoRV-digested pBluescriptSK1. A deletion strainwas made by replacing an NsiI-partial SalI fragment (corre-
Peroxisomal Functions in A. nidulans 1357

sponding to amino acids 22–221 of the predicted protein) witha partial XhoI-partial PstI fragment of the bar gene frompMT1612 and transforming the linear insert into a nkuADstrain selecting for glufosinate-resistant transformants.
Construction of strains expressing GFP-tagged proteins: Astrain expressing a GFP-AcuE fusion protein from the gpdpromoter has been described (Szewczyk et al. 2001). Thisstrain was crossed with relevant strains to obtain strains ex-pressing the fusion protein in various mutant backgrounds. Astrain expressing a GFP-AcuD fusion protein from the alcA pro-moter (Maggio-Hall and Keller 2004; K. Weeradechapon
and G. Turner, personal communication) was also used forcrosses with various mutant strains. Sources of gfp codingsequences for the following constructs were the plasmidspALX125, -213, or -215 with gfp flanked by the gpd promoterand the trpC terminator. AN1050.3 is predicted to encode aketo-acyl-CoA thiolase with an N-terminal PTS2 (RLNSILSHL)sequence. A 1.5-kb PCR fragment containing 100 bp of the 59sequence of this gene together with the entire coding regionwas generated with the primers 59-CAACTCCTCATGTCTTCTCG-39 and 59-ACAACAAAGCGCAATGCATAAACC-39. Thesecond primer removed the stop codon inserting an NsiI site,allowing a C-terminal in-frame fusion with GFP by cloning intoEcoRV-cut pALX215. A ClaI fragment containing this fusionsequence was cloned into ClaI-digested pMT1612 containingthe bar selectable marker. This plasmid was transformed intoan nkuAD strain selecting for glufosinate resistance. A trans-formant was identified as arising by a single homologousintegration event at AN1050.3, resulting in tandem copies of atruncated gene and the fusion gene driven by the AN1050.3promoter and encoding 417 amino acids fused to GFP. Thisstrain was used for crosses into pex mutant backgrounds. An N-terminal GFP fusion with AcuE (AN6653.3; Sandeman et al.1991) was made by generating a PCR fragment with a primercontaining an EcoRI site (59-GGAATTCCGACCGCCCAGCTTAAGGATGTG-39) and the standard M13 reverse primer us-ing as template a PGEMTEasy clone of acuE. This was cloned asan EcoRI–SpeI fragment into pALX213, resulting in a fusiongene driven by the gpd promoter producing GFP fused toamino acids 4–540 including the C-terminal SKL and contain-ing 671 bp 39-UTR. A mutation of the C-terminal S codon(TCA) was mutated to a stop codon (TAA) using the primers59-GTACCCGGAGACGAAATCTC-39 and 59-TGCTTAAAAGCTCTAATTTC-39 for inverse PCR and recircularizing using astemplate an acuE clone in pGEMTEasy containing the entiregene together with 1533 bp of 59-UTR. An NcoI–SpeI fragmentcontaining the mutation was used to replace the correspond-ing fragment in the gfp-acuE clone described above, resultingin a predicted GFP-AcuE fusion protein lacking the C-terminalSKL sequence. For complementation and GFP localizationstudies, the resulting constructs were cotransformed into ariboB2 stain containing an acuED (Nayak et al. 2006) selectingfor RiboB1. The entire ORF of AcuD (AN5634.3) was am-plified with the primers 59-GCTGAAAGCTTCCATCATGTCTT-39 and 59-CTGCAGTTGAACTGATCCTCTGTC-39 withthe second primer mutating the stop codon. This fragmentwas cloned into pALX215 at the EcoRV site, creating a codingregion for the entire ORF of AcuD with GFP fused at the Cterminus driven by the gpd promoter with the trpC terminator.This plasmid was used as a template for inverse PCR to deletesequences corresponding to AcuD amino acids 148–158 and315–338 using the primer pairs 59-GGGGTTGACTACCTTCGTCC-39/59-GGGAGTCATGCGCTCCTCGC-39 and 59-GGGTCCAACCTCGAGGCCCG-39/59-GGGATCAACGACAGCGTCGT-39, respectively. NotI–XhoI fragments from each of thethree plasmids were inserted into a plasmid containing the A.fumigatus pyroA4 selectable marker (Nayak et al. 2006) as wellas an internal fragment of the wA gene. The resulting plasmids
were transformed into a pyroA4 nkuAD strain selecting forPyro1 and screening of transformants for white conidialcolonies arising from homologous integration of the plasmidat the wA locus. The pexK gene (AN1921.3) was amplified withthe primers 59-CATAACTAAGATGTGTCTCTGG-39 and 59-GTAAGCTGTCTTTCGCCA-39 to generate a product with a 728-bp59-UTR and with the stop codon changed to TAC. An in-framefusion was made by cloning this fragment into a plasmidcontaining gfp and the trpC terminator. The insert was am-plified using the first primer and the standard vector primerM13 forward and cloned into a plasmid containing the A.fumigatus pyroA4 gene cut with EcoICRI. This was transformedinto an nkuAD pyroA4 strain selecting for PyroA1 and atransformant identified in which integration had occurred atthe trpC locus. This strain expressed PexK-GFP from the pexKpromoter and was crossed into different backgrounds.
Microscopy: Mycelium for microscopy was grown on cover-slips in liquid media at 25�. Alternatively, conidia were grownon solid media (1% agar)-coated microscope slides. Microscopyand capture of images were as described (Szewczyk et al. 2001).
RESULTS
Isolation of mutants affected in peroxisomal func-tions: Mutants specifically showing reduced ability togrow on media containing butyrate as the sole carbonsource but able to utilize acetate and glutamate wereisolated as described previously (Hynes et al. 2006). Thephenotype of mutants on butyrate medium ranged fromcomplete inhibition to poorer utilization. The genescorresponding to some of these mutants were clonedby complementation using a genomic library in thepRG3AMA1 autonomously replicating vector as describedin materials and methods. Sequencing of comple-menting clones and comparison with the A. nidulansgenome sequence showed that these genes correspondedto those encoding characterized peroxisomal functions,including the peroxins Pex1, -6, -7, and -13, designatedPexA, -F, -G, and -M to follow the A. nidulans genenomenclature (Table 1). In addition, a gene encodingan ATP carrier (AntA) was identified. Notably, with oneexception (see below), no mutants affected in fatty acidmetabolic enzymes were identified.
Mutations affecting additional peroxisome functionswere generated by cloning genes by PCR using primersbased on the A. nidulans genome sequence and gener-ating deletions or disruptions (materials and methods).A deletion of the pexF gene resulted in a phenotypeidentical to that of the pexF23 mutation and was shownto be allelic to pexF23. The genes predicted to encodePex5 (designated PexE) and Pex11 (PexK) were deletedand the gene predicted to encode Pex3 (PexC) wasdisrupted by insertion of the bar gene encoding glufo-sinate resistance (Table 1). The predicted defects inperoxisomal functions based on characterized S. cerevi-siae genes are summarized in Table 1.
Effects of mutations on peroxisomal protein targeting:The effects of pex mutations on the localization of theglyoxylate bypass enzymes, ICL (AcuD) and MAS (AcuE),were investigated by the use of GFP-tagged proteins
1358 M. J. Hynes et al.

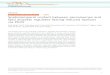
(Figure 1A). As shown previously (Szewczyk et al. 2001;Maggio-Hall and Keller 2004), GFP-AcuD and GFP-AcuE localized to punctate dots. This pattern was lost inpexF, pexM, and pexC mutant backgrounds, consistentwith the predicted roles of these genes in PTS1 andPTS2 protein import and peroxisomal biogenesis. AcuEcontains a predicted PTS1 (SKL), and mislocalizationwas observed in the pexE mutant background (PTS1receptor) but not in the pexG mutant (PTS2 receptor).Surprisingly, the reverse was observed for AcuD, whichhas no characterized peroxisomal targeting sequence,
indicating that this protein might have a cryptic PTS2sequence recognized by PexG (see below). The functionof PexG in the targeting of PTS2 proteins was confirmedby the finding that a GFP fusion with the product of thegene AN1050.3, predicted to encode a ketoacyl-CoAthiolase with a standard N-terminal PTS2 sequence, wasmislocalized in the pexG but not the pexE mutant (Figure1B).
The ICL of fungi show an unusually diverse range ofperoxisomal targeting signals (reviewed in Kunze et al.2006). AcuD has no obvious peroxisomal targeting
Figure 1.—Effects of pexmutations on the peroxi-somal localization of GFP-tagged proteins. Strainsexpressing the indicated GFPfusion proteins in differentpex mutant backgroundswere grown for 15–20 hron microscope slides coatedwith solid media. (A) AcuD-GFP stains were grown with0.25% ethanol except forthe pexED strain, which wasgrown on 10 mm threonineGFP-AcuE with 1% glucoseand (B) AN1050-GFP with0.25% Tween80 as carbonsources with ammonium asthe nitrogen source. (C) De-letion of amino acids 315–338 of AcuD results in lossof peroxisomal localization.Mycelium of strains contain-ing plasmids expressing thegfp fusion genes integratedat the wA locus were grownin liquid glucose minimalmedia on coverslips for mi-croscopy. DIC, differentialinterference contrast; FITC,standard fluorescein isothio-cyanate filter set. Bar, 20 mm.
TABLE 1
Summary of mutants affecting peroxisomal functions
Gene AlleleGenome
locusa
S. cerevisiaeorthologb
% identity(similarity)c
Length(aa)a Proposed function
pexA pexA9 AN5991.3 PEX1 39 (54) 1178 PTS1 and PTS2 protein importpexF pexF23 AN2925.3 PEX6 34 (50) 1477 PTS1 and PTS2 protein importpexM pexM15 AN1511.3 PEX13 35 (53) 437 PTS1 and PTS2 protein importpexC pexCTbar AN2281.3 PEX3 23 (39) 531 Peroxisome biogenesispexE pexED AN10215.3 PEX5 33 (48) 656 PTS1 protein import receptorpexG pexG14 AN0880.3 PEX7 41 (56) 356 PTS2 protein import receptorpexK pexKD AN1921.3 PEX11 27 (50) 236 Peroxisome proliferationantA antA15 AN0257.3 ANT1 37 (55) 337 ATP carrier
a Predicted gene and length of protein based on gene annotations (http://www.broad.mit.edu/annotation/genome/aspergillus_group/MultiHome.html).
b Sequences from the Saccharomyces Genome Database (http://www.yeastgenome.org/).c BlastP (Altschul et al. 1990) of S. cerevisiae protein against predicted A. nidulans sequence.
Peroxisomal Functions in A. nidulans 1359

signals as also found for ICL in other filamentousascomycetes. ICL sequences from bacteria to plantsare highly conserved and Gainey et al. (1992) identifieda region relatively conserved in filamentous ascomy-cetes but absent from bacteria, plants, and S. cerevisiae.By a similar analysis, we identified two regions in AcuDfor analysis. In-frame deletions corresponding to thesesequences were made in a gfp-acuD fusion construct andtransformed into A. nidulans with integration at the wAlocus (materials and methods). The deletion corre-sponding to aa 315–338 resulted in mislocalization(Figure 1C), indicating that the 315–338 region maycontain a cryptic sequence acting as a PTS2 sequencerecognized by the pexG receptor. The gfp-acuD strainswere crossed to an acuD loss-of-function mutant, and itwas found that the wild-type fusion gene complementedfor growth on acetate while both deletion genes showedlack of complementation, suggesting that the deletionsresulted in loss of enzyme activity. We cannot excludethe possibility that AcuD is localized via piggybackingwith an unknown protein with a standard PTS2sequence.
Proliferation of peroxisomes is dependent on PexK:In S. cerevisiae and other fungi, including A. nidulans,peroxisome numbers increase in response to fatty acids(Valenciano et al. 1996). Pex11 has been shown to be aperoxisomal membrane protein that recruits dynamin-like proteins necessary for peroxisome division andproliferation in many species (Erdmann and Blobel
1995; Li et al. 2002; Li and Gould 2003; Kiel et al. 2005;Thoms and Erdmann 2005; Yan et al. 2005; Lingard
and Trelease 2006; Orth et al. 2007). When a strainexpressing GFP-AcuE from the constitutive gpd pro-moter was grown on glucose and transferred to mediacontaining different carbon sources, increased num-bers of peroxisomes were observed in the presence ofoleate or butyrate in comparison with glucose or acetatemedia (Figure 2A). This was not observed in the pexKD
background where there was no response to fatty acidmedia with only a few larger intensely fluorescentperoxisomes observed.
A sequence encoding PexK labeled at the C terminuswith GFP was inserted into the A. nidulans genome at thetrpC locus (materials and methods) and it was foundthat this fusion protein was able to almost fully comple-ment the pexKD for growth on fatty acid media. A pexKgfppexK1 strain was grown for 16 hr in glucose medium andtransferred to glucose-, butyrate-, or oleate-containingmedia for 4 hr for microscopy. During growth on glu-cose, labeling of the peroxisomal membrane by GFPwould be dependent on basal expression of the fusiongene. Fatty acid induction would be expected to in-crease expression of both tagged and untagged PexK,potentially resulting in peroxisomal proliferation. Thiswas observed with both butyrate and oleate resulting inincreased numbers of peroxisomes present in distinctclusters spaced along the hyphae (Figure 2B). Consistent
with PexC being required for de novo production of per-oxisomes from the endoplasmic reticulum (Hoepfner
et al. 2005; Kragt et al. 2005), the pexCTbar disruptionresulted in mislocalization of GFP-AcuE (Figure 1A)and also in loss of the formation of peroxisomes labeledwith PexK-GFP (Figure 2C). The pexF23 mutation re-sulted in a few small punctate fluorescent structures,presumably peroxisomal membrane ghosts, as well asincreased levels of mislocalized PexK-GFP (Figure 2C).
Deletion of farA, encoding a transcription factor re-quired for fatty acid induction of enzymes involved infatty acid utilization and of expression of some pex genes(Hynes et al. 2006), greatly reduced the expansion ofthe number of peroxisomes in response to butyrate andoleate as visualized by both GFP-AcuE and PexK-GFPlabeling (Figure 2, A and B). This indicated that ex-pansion of the numbers of peroxisomes in response tofatty acids results from FarA-dependent increased levelsof the proteins required for peroxisomal biogenesis andproliferation.
Growth of mutants on carbon sources: All mutantswere able to utilize glucose, a strong carbon source, aswell as the poorer carbon sources, proline and lactose,almost as well as wild type (Figure 3A). The only observ-able effect was a delay in growth on all media observableat 1 day of incubation, resulting in somewhat smallercolony diameters. Therefore, peroxisomal functions arenot essential for growth. The mutants were also able toutilize acetate with the exception of pexED (discussed inmore detail below). However, growth of all pex mutantswas clearly affected to some extent. Growth on ethanol,which is metabolized via acetyl-CoA, was similar to thaton acetate except for the somewhat greater effects ofthe pex mutants. Growth on the short-chain fatty acidsbutyrate (C4), valerate (C5), and hexanoate (C6) wasgreatly inhibited in all mutants defective in the perox-isomal targeting of PTS1 proteins and this was observedin the presence of lactose (Figure 3A). Utilization ofthese fatty acids was clearly reduced but not inhibited inthe pexG mutant defective in PTS2 protein targeting aswell as in the pexKD and the antA15 mutants.
As observed previously (Maggio-Hall and Keller
2004), A. nidulans is unable to grow on fatty acids ofchain length 7–10 due to strong inhibition but is able toutilize longer-chain fatty acids. However, on long-chainfatty acids conidiation was greatly reduced as illustratedfor oleate compared with the poor carbon source lac-tose (Figure 3B). Growth inhibition by long-chain fattyacids was observed for all mutants affected in PTS1 pro-tein localization while growth was reduced but not in-hibited in the pexF, pexK, and antA mutants (Figure 3C).These results showed that peroxisomes are involved inboth short- and long-chain fatty acid utilization.
Developmental defects in peroxisomal mutants: De-spite strong growth on standard glucose containingcomplete and minimal media, pex mutants defective inPTS1 protein localization conidiated poorly as shown by
1360 M. J. Hynes et al.

a brown colony appearance. As for wild type, this waspartly remediated by the presence of 1 m sorbitol (Fig-ure 4A) and 0.6 m KCl (not shown). The effects of thepexG mutation were much less extreme. Conidial countssupported these observations (Figure 4B). Mutants con-taining the pexKD were not detectably affected inconidiation (not shown).
These effects on conidiation were observed in thevelA1 mutant background that is standard for mostlaboratory work. In a velA1 background, conidiation isgreatly reduced (Mooney and Yager 1990; Kim et al.2002; Tsitsigiannis et al. 2004a). We therefore testedthe effects of the pexF and pexC mutations on conidia-tion in velA1-containing strains and found virtually no
Figure 2.—Peroxisomalbiogenesis and prolifera-tion. (A) Proliferation ofGFP-AcuE-labeled peroxi-somes in reponse to fattyacids is dependent on PexKand FarA. (B) FarA-depen-dent proliferation of perox-isomes in response to fattyacids visualized by peroxi-somal membrane labelingwith PexK-GFP. Enlarge-ments (arrowheads) showincreased numbers of per-oxisomes resulting fromfatty acid induction. In glu-cose media only 1–2 perox-isomes are observed whilebutyrate and oleate resultin 5–10 and .10 peroxi-somes in each cluster, re-spectively. In farAD, singleperoxisomes are observedexcept for oleate whereclusters of up to 5 peroxi-somes are infrequently ob-served. (C) Peroxisomebiogenesis visualized byPexK-GFP is dependent onPexC and is aberrant inthe pexF23 mutant. Myce-lium was grown on cover-slips in 1% glucoseminimal liquid media for18 hr and then transferredto the same media or tominimal media containing10 mm acetate, 10 mm buty-rate, or 0.5% Tween80 (as asource of oleate). All mediacontained ammoniumchloride as the nitrogensource.
Peroxisomal Functions in A. nidulans 1361

conidiation (Figure 4C). Again, this was relieved by thepresence of sorbitol. No effects of velA1 on growthinhibition by fatty acids were observed (not shown). Thepex mutations affecting PTS1-localized proteins, but not
the pexG or pexK mutations, were also observed to affectsexual reproduction with homozygous but not hetero-zygous crosses being subfertile with very few smallfruiting bodies (cleistothecia) being produced, indicat-ing a recessive defect. A velA1 background results ingreatly increased cleistothecial production (Kim et al.2002) and in this background pexF- and pexC-containingstrains produced increased numbers of small cleisto-thecia (Figure 4D).
Woronin bodies that are used to plug septal poresas well as broken hyphal tips, preventing cytoplasmicleakage ( Jedd and Chua 2000; Tenney et al. 2000; Tey
et al. 2005), contain the Hex1 PTS1 protein, andWoronin bodies have been shown to be derived fromperoxisomes in N. crassa (Yuan et al. 2003; Managadze
et al. 2007). Therefore, the gene AN4695.3 (designatedhexA), predicted to encode the Woronin body protein,was deleted (materials and methods). Growth andconidiation was only slightly affected by the hexAD
(Figure 4E) and homozygous crosses were fully fertile(Figure 4F). The pexF23 hexAD double mutant was notmore affected than the pexF23 single mutant in con-idiation (Figure 4E). Sensitivity to sorbose has beenobserved in other fungi lacking the Woronin bodyprotein and, although A. nidulans is highly sensitive tosorbose, some sensitivity of both pexF23 and hexAD
mutants was observable (Figure 4E).Signaling by the oxylipin Psi factors derived from
fatty acids has been shown to affect the balance be-tween asexual and sexual development (for review seeTsitsigiannis and Keller 2007). Oleate was found toenhance cleistothecial production in both wild-type andhexAD strains but not at all in a pexF23 mutant. Thismight indicate that peroxisomal fatty acid metabolism isrequired for oxylipin production but it cannot beexcluded that this is the result of the inhibitory effectsof oleate in pexF23. The pex mutants tested were alsofound to have reduced rates of conidial germination(Table 2). It is likely that this explains the initial slowergrowth and somewhat smaller colony size even onglucose media.
It was found that all pex mutants affected in PTS1protein targeting, but not the pexG mutant, were auxo-trophic for biotin. The pex mutants all responded to theaddition of the intermediate pimelic acid while a biA1mutant, predicted to lack biotin synthase, the final stepin biotin synthesis, did not respond to pimelic acid. Ithas been shown that pimelic acid is synthesized fromoleate and other long-chain fatty acids in some fungalspecies (Ohsugi et al. 1988).
The effects of mislocalization of glyoxylate cycleenzymes: The unique enzymes of the glyoxylate path-way, ICL and MAS, are peroxisomal in plants and insome fungi (for review see Kunze et al. 2006), as shownhere for A. nidulans. However, in S. cerevisiae, ICL isalways cytoplasmic and MAS, which contains a PTS1, isonly peroxisomal during growth on oleate, but this
Figure 3.—Growth of mutants on carbon sources. The fol-lowing carbon sources were added to minimal media (1%agar) with 10 mm ammonium chloride as the nitrogen sourceat the following concentrations: glucose (1%), proline and ac-etate (50 mm), ethanol and lactose (0.5%), butyrate and val-erate (10 mm), hexanoate (5 mm), and C12-C22 chain-lengthfatty acids (2.5 mm). Acids were adjusted to pH 6.5 with so-dium hydroxide where necessary and tergitol (NP-40; Sigma)was added before melting media to disperse longer-chain fattyacids. Growth was for 2–3 days at 37�. (A) Growth of mutantson short-chain fatty acids. (B) An enlargement of colonies of awild-type strain on oleate and lactose to show inhibition ofconidiation. (C) Growth of mutants on long-chain fatty acids.
1362 M. J. Hynes et al.

localization is not essential for oleate utilization (Kunze
et al. 2002).We investigated the effects of mislocalization of MAS
on growth on acetate and fatty acids by mutating theacuE gene to eliminate the encoded C-terminal PTS1(materials and methods). Cotransformation of anacuED riboB2 strain with plasmids encoding wild-type orthe PTS1 mutant AcuE, together with a riboB1-containingplasmid selecting for RiboB1, yielded �50% transform-ants capable of growth on acetate or oleate as carbonsources, indicating complementation of the acuED. Com-plementation was similarly found for constructs express-ing GFP-AcuE fusions with or without the PTS1. The
PTS1 mutation did not affect growth on acetate and onlyslightly reduced growth on butyrate and long-chain fattyacids (Figure 5A). Growth on valerate was not affected byacuE mutations, consistent with one cycle of b-oxidationresulting in the formation of propionyl-CoA, which ismetabolized via the methyl-citrate pathway, therebybypassing the glyoxylate cycle (Brock et al. 2000; Brock
2005). The PTS1 mutation was found to result in mis-localization of GFP-AcuE to the cytoplasm (Figure 5B).These results indicated that, like S. cerevisiae, peroxi-somal localization of MAS is not essential for a functionalglyoxylate cycle during growth on either acetate oroleate (Kunze et al. 2006). However, in A. nidulans ICL
Figure 4.—Develop-mental phenotypes of pexmutants. (A) Colony mor-phology of mutants. Re-duced conidiation is shownby the brown appearanceof colonies growing on com-plete and glucose minimalmedia. This is partially re-stored by the presence ofsorbitol (1 m) in completemedium.(B)Conidialcountsof mutants grown on com-plete media or completemediawith1m sorbitol.Con-idia were harvested from1.7-cm-diameter circular re-gions and counted with ahemocytometer. Error barsrepresent the standard er-rors of three replicates. (C)Colony morphology of pexmutants showing a furtherreduction of conidiation invelA1 backgrounds. (D)Mag-nified view of surface of col-onies growing on glucoseminimal medium showingthe production of smallcleistothecia in pexF andpexC mutants in a velA1 back-ground. The cleistotheciaareobservedasblackspherescoated by Hulle cells. (E)Colony morphology of mu-tants containing a deletionof the hexA gene. Conidia-tion is only slightly affectedin comparison with thepexF23 mutant. Both wild-type (green) and yA1 (yel-low) conidial colonies areshown. Increased sensitivityto sorbose (0.1% in the pres-ence of 50 mm proline as car-bon source) is shown by the
hexAD and pexF23 mutants. (F) Cleistothecia (white arrowheads) produced by wild-type and hexAD but not pexF23 strains in homo-zygous crosses viewed with an inverted microscope. (G) Induction of the production of cleistothecia by oleate in wild-type and hexADbutnot inpexF23 strains.Discs(1cm)saturatedwith1moleatewereplacedinthemiddleofglucoseminimalplatesspreadwithconidia.Plates were taped to exclude air after 2 days and incubated for another 5 days and viewed with an inverted microscope. Cleistotheciacoated with Hulle cells are seen at the periphery of the filters.
Peroxisomal Functions in A. nidulans 1363

is peroxisomal with the consequence that the twoenzymes would be predicted to be in separate compart-ments in acuE PTS1 mutants. It is likely that localizationof ICL is inefficient with some enzyme being present inthe cytoplasm; this was supported by the observation ofsome cytoplasmic fluorescence in GFP-AcuD strains.
Surprisingly, GFP-AcuD was mislocalized in a pexG butnot a pexE background (Figure 1), indicating a require-ment for the PTS2 receptor peroxin. However, growthon acetate or ethanol utilization was only slightly af-fected in the pexG14 mutant (Figure 5C), showing thatperoxisomal localization of ICL is not required. How-ever, pexED strains, where AcuE and other PTS1 proteinsare cytoplasmic, were inhibited on acetate and ethanolmedia (Figure 5C). Loss of targeting of all peroxisomalproteins in pexED pexG14 double mutants resulted inimproved growth on these media (Figure 5C) and thiswas also observed in pexED double mutants with pexF23,pexA9, pexM15, and pexCTbar (not shown). This indicatedthat, in the presence of acetate, ICL activity in the peroxi-some in the absence of MAS and other PTS1 proteinsmight result in the toxic accumulation of glyoxylate.
Mitochondrial pathways for fatty acid utilization: Amitochochondrial pathway for fatty acid b-oxidation hasbeen discovered in A. nidulans by Maggio-Hall andKeller (2004). Deletion of the echA gene, encoding amitochondrial enoyl-CoA hydratase, results in inhibitedgrowth on butyrate, hexanoate as well as oleate and eru-cic acid. In contrast, deletion of scdA, predicted to en-code a mitochondrial acyl-CoA dehydrogenase, does notresult in inhibited growth on fatty acids but in an in-ability to utilize hexanoate and butyrate (Maggio-Hall
et al. 2007). We have confirmed these results (Figure6A). One of the mutants that we isolated was comple-mented by a clone containing a gene, AN10512.3, pre-dicted to encode a mitochondrial ketoacyl-CoA thiolase(materials and methods). The phenotype of this mu-tant (designated mthA25) was identical to that of thescdAD: inability to grow on hexanoate and butyrate andno inhibition by long-chain fatty acids (Figure 6A). Theseresults supported the existence of a mitochondrial path-way for b-oxidation of even-numbered short-chain fattyacids (Figure 6C). Accumulation of enoyl-CoA in anechA mutant results in growth inhibition, while the scdAand mthA mutations result in loss of a functional path-
way without the accumulation of toxic metabolites.Consistent with this, an scdAD echAD double mutant isnot inhibited by fatty acids (Maggio-Hall et al. 2007)and we have isolated a putative scdA mutation as asuppressor of the echAD (our unpublished data).
The gene AN7008.3 is predicted to encode a mito-chondrial hydroxy-acylCoA dehydrogenase. We clonedthis gene (designated hadA) and generated a disruptionmutant by insertion of the riboB gene (materials and
methods). This mutation resulted in a phenotype sim-ilar to the echAD: inhibition by butyrate and the long-chain fatty acids elaidic, oleate, and erucic (Figure 6B).It was found that scdAD hadATriboB double mutantswere not sensitive to butyrate and oleate. These resultsshowed that the predicted hydroxyacyl-CoA dehydroge-nase functions in this pathway and that accumulation ofthe hydroxyacyl-CoA substrate results in inhibition(Figure 6C). The mutants affected in the mitochodrialpathway do not prevent growth on the odd-numberedvalerate (C5) nor is inhibition observed with tridecanoic(C13) or heptadecanoic (C17) (Figure 6, A and B).
DISCUSSION
By the isolation of mutants unable to use the short-chain fatty acid butyrate as a carbon source, we haveidentified mutants affected in peroxisomal functioning.These include pexA and pexF mutants predicted to bedefective in the recycling of PTS receptors to the cy-toplasm due to a loss of the AAA ATPase complex as wellas a pexM mutant defective in an essential protein of themembrane complex necessary for docking of receptor–cargo protein complexes (for review see Platta andErdmann 2007). An additional mutant due to a loss-of-function mutation in pexG, predicted to encode thereceptor for PTS2 proteins, results in an inability totarget a PTS2-containing protein but is unaffected inPTS1 protein localization. Deletion of the pexE generesults in loss of PTS1 but not PTS2 protein import,showing that the gene product is the specific PTS1receptor. These results confirm the proposal that thePTS receptors function independently in filamentousfungi unlike in mammals and plants (see Kiel et al.2006). The pexC gene (AN2281.3) is predicted toencode a protein orthologous to Pex3, which together
TABLE 2
Conidial germination in pex mutants
Strain
Time (hr) Wild type pexF2323 pexCTbar pexED pexG14 pexKD
4 53 (15) 23 (4) 10 (2) 30 (11) 8 (5) 13 (5)6 100 (0) 90 (7) 70 (12) 86 (9) 80 (7) 93 (4)
Conidia were diluted and germinated in glucose minimal medium and viewed with an in-verted microscope. Conidia visibly germinating were counted and are expressed as a percentageof the total. Values are the average of three replicates with standard errors shown in parentheses.
1364 M. J. Hynes et al.

with Pex16 and Pex19, have been found to be essentialfor peroxisomal membrane formation in S. cerevisiaeand mammals (Hettema et al. 2000; Hoepfner et al.2005; Kragt et al. 2005; Fujiki et al. 2006). Disruption ofpexC resulted in mislocalization of both PTS1 and PTS2proteins as well as the inability to form peroxisomalstructures as visualized by PexK-GFP.
Developmental phenotypes of mutants: Completeloss of import of matrix proteins, or indeed an inabilityto form peroxisomes as in the pexC mutant, does notresult in extreme phenotypes under laboratory condi-tions. Mutants are able to form colonies both on richcomplete media and on minimal media where the only
essential function missing is the ability to synthesizepimelic acid in the biotin biosynthetic pathway.
Incomplete developmental defects were observed inthe mutants unable to localize PTS1 proteins. Conidia-tion was significantly reduced and homozygous crossesproduced very few small cleistothecia. In velA1 back-grounds, where the balance between sexual and asexualdevelopment is altered (Kim et al. 2002), conidiation wasfurther reduced and production of cleistothecia in-creased, indicating that these effects were independentof the VelA-signaling pathway. In view of the major rolefor lipid metabolism in the generation of oxylipin-signaling molecules regulating development as wellas secondary metabolism (Tsitsigiannis et al. 2004b,2005; Tsitsigiannis and Keller 2007), it is surprisingthat a complete lack of peroxisomes does not have amore drastic effect on A. nidulans. The observation of analmost complete absence of conidiation in wild-typestrains growing on long-chain fatty acids as sole carbonsources might reflect gross alterations in oxylipin factorsignaling. Only a minor conidiation defect was observedin a mutant lacking the HexA Woronin body protein. Inthe heterothallic fungus Podospora anserina, loss of Pex2results in an inability to switch from mitotic to meioticdevelopment (Berteaux-Lecellier et al. 1995) andpex5D mutants have nuclear, mitochondrial, and sexualabnormalities (Bonnet et al. 2006). Interestingly, apex7D is epistatic to the effects of the pex5D for all phe-notypes, raising the possibility that mislocalization ofonly PTS1 proteins is more detrimental than mislocal-ization of all peroxisomal proteins (Bonnet et al. 2006).The developmental phenotype of the pexED describedhere is not more extreme than that of other pex mutantsor the pexED pexG14 double mutant. Reactive oxygenspecies have been found to be important in sexual de-velopment in A. nidulans (Scherer et al. 2002; Lara-Ortiz et al. 2003). It is possible that some developmentalabnormalities in the pex mutants result from impair-ment of normal reactive oxygen metabolism.
Fungal sexual and asexual spores have high lipid con-tents (e.g., Goodrich-Tanrikuluet al. 1998; Tsitsigiannis
et al. 2004b; Schobel et al. 2007). Messenger RNAs forglyoxylate cycle and gluconeogenic genes are found inA. nidulans conidia (Osherov and May 2000, 2001). Al-though rates of conidial germination in pex mutants arereduced, both conidia and ascospores of the pex mutantscan germinate, showing that peroxisomal fatty acid metab-olism is not essential for signaling the initiation of ger-mination and that lipids do not provide the only initialcarbon source for growth. It is likely that the other majorsource of carbon in the spore is trehalose (Fillinger et al.2001).
Regulation of peroxisome proliferation: The num-ber of peroxisomes greatly increases in response to bothshort- and long-chain fatty acids and this is dependenton PexK. GFP-PexK labels peroxisomes in the absenceof proliferation, indicating that it is a normal compo-
Figure 5.—Effects of mislocalization of peroxisomal en-zymes on growth on acetate and fatty acids. (A) Mislocaliza-tion of MAS encoded by acuE does not prevent growth onacetate or fatty acids. The phenotypes of representative co-transformants expressing GFP-AcuE with (SKL) and without(stop) PTS1 are compared with wild type and an acuE mutant.(B) Mislocalization of GFP-AcuE resulting from loss of PTS1.Mycelium for microscopy was grown in liquid glucose mini-mal media on coverslips. Similar results were observed for my-celia grown in acetate media and for five other transformantsfor each construct. (C) Growth inhibition on acetate and eth-anol media in a pexED mutant is partially restored in a doublemutant with the pexG14 mutation.
Peroxisomal Functions in A. nidulans 1365

nent of the peroxisomal membrane. Strains containingthe pexKD grow more poorly than wild type on both long-and short-chain fatty acids but no inhibition by fattyacids is observed and no detectable growth or develop-mental defects are observed in pexKD strains. Over-expression of Pex11 in Penicillium chrysogenum has beenfound to result in peroxisome proliferation (Kiel et al.2005). While in S. cerevisiae there is one Pex11 orthologrequired for peroxisome proliferation (Erdmann andBlobel 1995), multiple Pex11 proteins are found inhumans and plants (Li et al. 2002; Lingard andTrelease 2006). It has been suggested that there aretwo additional paralogs of Pex11 in A. nidulans as well asin A. fumigatus and P. chrysogenum (Kiel et al. 2006). It isnot known if these additional proteins play a role in theresponse to fatty acids or in the response to other stimuli.
The expression of pexK is induced in response to fattyacids but not acetate, and this is dependent on thetrancription factors ScfA and FarB for short-chain fattyacid induction and on FarA for both long- and short-chain induction (Hynes et al. 2006). Both FarA andFarB have been found to bind in vitro to DNA containingthe core CCGAGG sequence found in the 59 upstreamregion of pexK and other genes inducible by fatty acids.The greatly reduced peroxisome proliferation observedin farAD is consistent with proliferation dependent onfatty acid induction of PexK expression. The 59 sequencesof many A. nidulans pex genes contain one or morecopies of the CCGAGG-binding sequence. These genesinclude pexE and pexG encoding the specific PTS re-ceptors well as pexC and AN5113.3 (the predicted Pex16ortholog) but not AN4899.3 (Pex19), all of which arerequired for peroxisome biogenesis from the endo-plasmic reticulum (Hettema et al. 2000; Fujiki et al.2006). Genes for enzymes of b-oxidation (both mito-chondrial and peroxisomal), the methyl-citrate pathwayof propionyl-CoA metabolism, lipases, and cutinasesalso have 59-CCGAGG sequences (Hynes et al. 2006).
Therefore, during growth in the presence of fatty acidsit is predicted that there is a major reorganization ofmetabolism with expansion of the number of perox-isomes containing enzymes relevant to fatty acid catab-olism resulting from gene induction.
The role of peroxisomes in acetate utilization: In S.cerevisiae, the unique enzymes of the glyoxylate cycle,MAS and ICL, are cytoplasmic during growth on acetateor ethanol and pex mutants are able to grow on thesecarbon sources (Kunze et al. 2006). In A. nidulans, ICLand MAS are present in peroxisomes independently ofthe carbon source. However, we have shown that mu-tants with loss of peroxisomal targeting of matrix pro-teins, loss of peroxisome biogenesis, or peroxisomeproliferation are able to grow on acetate. Mislocaliza-tion of MAS resulting from loss of the PTS1 of AcuEdoes not abolish growth on acetate. AcuD targetingrequires the PTS2 receptor PexG and yet the pexG14mutant can grow on acetate, indicating that ICL doesnot have to be peroxisomal. However, pexG-dependentICL localization results in the pexE mutant having ICL inperoxisomes in the absence of MAS as well as otherPTS1-containing proteins. This results in inhibitedgrowth on acetate and ethanol, which is restored by lossof targeting of ICL in double mutants with other pexmutations. This might be explained if peroxisomal ICLactivity results in the production of glyoxylate, which, inthe absence of the PTS1-dependent MAS, cannot beused for malate production and is unable to escape theperoxisome, resulting in toxicity. This scenario dependson the import into the peroxisome of isocitrate gener-ated from citrate in the cytoplasm by aconitase.
Peroxisomes and fatty acid utilization: Complete lossof protein targeting to peroxisomes or a lack of peroxi-somes results in growth inhibition by fatty acids as doesthe mislocalization of PTS1-containing proteins in thepexED mutant. The pexG14 mutant, where only PTS2proteins are mislocalized, also resulted in reduced
Figure 6.—Effects of mu-tations in the mitochondrialshort-chain fatty acid b-oxi-dation pathway. (A) Growthon even- but not odd-num-bered chain fatty acids is in-hibited in the echAD mutant.Growth on butyrate andhexanoate but not valerateor long-chain fatty acids isgreatly reduced in the scdADand mthA25 mutants. (B)ThehadAdisruptionmutanthas properties similar toechAD. Growth was for 2–3days at 37�. (C) Proposedmitochondrial b-oxidationpathway. Accumulation ofintermediates in echA andhadA mutants leads togrowth inhibition.
1366 M. J. Hynes et al.

growth, but not inhibition, on fatty acid medium withthe greatest effects observed with short-chain comparedwith long-chain fatty acids. The lack of a strong pheno-type of pexG14 on long-chain fatty acids might reflect theinvolvement of multiple ketoacyl-CoA thiolases witheither PTS1 or PTS2 signals. Similar effects of pex mu-tants in P. anserina have been reported with inhibitedgrowth on oleate in pex2 and pex5 mutants and reducedoleate utilization in a pex7 mutant (Bonnet et al. 2006).Magnaporthe grisea and Colletotrichum lagenarium pex6deletions show complete loss of growth on long-chainfatty acid media (Kimura et al. 2001; Ramos-Pamplona
and Naqvi 2006). One possible mechanism for fattyacid inhibition is that cytoplasmic b-oxidation enzymesresult in the generation of hydrogen peroxide, whichcannot satisfactorily be detoxified by mislocalized per-oxisomal catalase (Kawasaki and Aguirre 2001) orglutathione peroxidase (AN5440.3) as suggested for P.anserina (Ruprich-Robert et al. 2002). We have pre-viously reported that downregulation of the expressionof genes involved in fatty acid breakdown leads to someresistance to oleate inhibition of a pexFD strain by theisolation of mutations in the farA regulatory gene aspartial suppressors of this phenotype (Hynes et al.2006). An additional possibility is that pex mutants aresensitive to fatty acids because of pertubations in mem-brane composition as observed for S. cerevisiae mutants(Lockshon et al. 2007). However, this is not an obviousconsequence of exposure to short-chain fatty acids and,in the S. cerevisiae studies, sensitivity was observed in apex7D, which was not observed here.
Further evidence for peroxisomal metabolism of fattyacids is provided by the requirement for PexK, which isnecessary for peroxisomal proliferation. However, theeffects of pexKD are not complete with growth being lessaffected on long- than on short-chain fatty acids, in-dicating that proliferation is not absolutely essential. Inaddition, mutation of the antA gene predicted to en-code a peroxisomal ATP carrier necessary for b-oxida-tion (Palmieri et al. 2001; van Roermund et al. 2001)results in greatly reduced growth on all fatty acids.
In our screen for butyrate-nonutilizing mutants wehave failed to isolate any affected in peroxisomal en-zymes and previous attempts to isolate mutants inA. nidulans have not yielded any lacking long-chainb-oxidation enzymes although an uncharacterised pexmutant was isolated (Kawasaki et al. 1995; De Lucas
et al. 1997). A deletion mutant of foxA encoding a per-oxisomal multifunctional enzyme is unable to grow onerucic acid (C22) but is only partially defective for growthon oleate and is unaffected for growth on short-chainfatty acids (Maggio-Hall and Keller 2004). Further-more, analysis of the genome sequences of A. nidulansand other filamentous fungi reveals the presence ofmany possible genes coding for b-oxidation enzymeswith potential PTS sequences, and deletion or disrup-tion of some of these has failed to result in complete loss
of growth on any one fatty acid (K. Reiser, M. A. Davis
and M. J. Hynes, unpublished results). It is likely thatfilamentous fungi have evolved multiple pathways allow-ing growth on a diversity of fatty acid substrates.
The normal situation for fatty acid utilization in A.nidulans is the production of acetyl-CoA by b-oxidationand its metabolism via the glyoxylate cycle within per-oxisomes, resulting in the production of malate andsuccinate. It is not clear how these metabolites exit theperoxisome and in all organisms there are many unan-swered questions relating to metabolite trafficking be-tween the cytoplasm, peroxisomes, and mitochondria(for discussion see Kunze et al. 2006; Visser et al. 2007).
Mitochondrial b-oxidation: Our data suggest that acomplete peroxisomal b-oxidation pathway exists as forS. cerevisiae. However, as shown by Maggio-Hall andKeller (2004), an additional mitochondrial pathwayexists. Mutations of either echA or hadA genes lead toinhibited growth on short-chain fatty acids as well ason even-numbered long-chain fatty acids due to accu-mulation of toxic intermediates. With even-numberedlong-chain fatty acids, butyryl-CoA produced in thepenultimate cycle of peroxisomal b-oxidation can enterthe mitochondria for b-oxidation, accounting for thetoxicity observed. The lack of butyrate and hexanoateutilization (but not inhibition) in the scdA and mthAmutants shows that the mitochondrial pathway is essen-tial for growth on these fatty acids. However, the scdAand mthA mutants do not affect growth on long-chainfatty acids, indicating that this mitochondrial pathway isnot required for their utilization. A possible reason foran essential mitochondrial short-chain b-oxidationpathway is that only one or two cycles of the peroxisomalpathway are insufficient for efficient carbon flux and sothe direct generation of acetyl-CoA within the mito-chondrion is required for energy production. The mito-chondrial pathway is also required for the utilization ofthe amino acids valine and isoleucine as carbon sources(Maggio-Hall et al. 2007).
Mutants affected in the mitochondrial pathway areable to grow on the odd-numbered valerate (C5) and ontridecanoic (C13) and heptadecanoic (C17). This isconsistent with valerate not being a substrate for thedefined mitochondrial pathway but undergoing oneround of b-oxidation in the peroxisome to produceacetyl-CoA and propionyl-CoA. The final cycle of b-oxidation of odd-numbered long-chain fatty acids wouldalso result in acetyl-CoA and propionyl-CoA in the per-oxisome with propionyl-CoA shuttling to the mitochon-dria where it can be metabolized via the methyl-citratepathway (Brock et al. 2000; Brock 2005). The possibil-ity that there is a distinct mitochondrial b-oxidationpathway for valerate remains.
Initial characterization of the pexG mutant was carried out by AnnaDuncan. Kimberly McMahon and Clarissa Moreira assisted with thedisruption of the hadA gene as part of an undergraduate project. Theresearch and technical assistance of Sophie Delimitrou, Rosemary
Peroxisomal Functions in A. nidulans 1367

Genovese, and Khanh Nguyen is gratefully acknowledged. This workwas supported by a grant from the Australian Research Council.
LITERATURE CITED
Altschul, S. F., W. Gish, W. Miller, E. W. Myers and D. J. Lipman,1990 Basic local alignment search tool. J. Mol. Biol. 215: 403–410.
Andrianopoulos, A., and M. J. Hynes, 1988 Cloning and analysisof the positively acting regulatory gene amdR from Aspergillus ni-dulans. Mol. Cell. Biol. 8: 3532–3541.
Armitt, S., W. McCullough and C. F. Roberts, 1976 Analysis ofacetate non-utilizing (acu) mutants in Aspergillus nidulans. J.Gen. Microbiol. 92: 263–282.
Barelle, C. J., C. L. Priest, D. M. MacCallum, N. A. R. Gow, F. C.Odds et al., 2006 Niche-specific regulation of central metabolicpathways in a fungal pathogen. Cell. Microbiol. 8: 961–971.
Berteaux-Lecellier, V., M. Picard, C. Thompson-Coffe, D. Zickler,A. Panvier-Adoutte et al., 1995 A nonmammalian homolog ofthe PAF7 gene (Zellweger syndrome) discovered as a gene involvedin caryogamy in the fungus Podospora anserina. Cell 81: 1043–1051.
Bonnet, C., E. Espagne, D. Zickler, S. Boisnard, A. Bourdais et al.,2006 The peroxisomal import proteins PEX2, PEX5 and PEX7are differently involved in Podospora anserina sexual cycle. Mol.Microbiol. 62: 157–169.
Borneman, A. R., M. J. Hynes and A. Andrianopoulos, 2001 AnSTE12 homolog from the asexual, dimorphic fungus Penicilliummarneffei complements the defect in sexual development of anAspergillus nidulans steA mutant. Genetics 157: 1003–1014.
Brocard, C., and A. Hartig, 2006 Peroxisome targeting signal 1: Is itreally a simple tripeptide? Biochim. Biophys. Acta 1763: 1565–1573.
Brock, M., 2005 Generation and phenotypic characterization of As-pergillus nidulans methylisocitrate lyase deletion mutants: methyl-isocitrate inhibits growth and conidiation. Appl. Environ.Microbiol. 71: 5465–5475.
Brock, M., R. Fischer, D. Linder and W. Buckel, 2000 Methyl-citrate synthase from Aspergillus nidulans: implications for propi-onate as an antifungal agent. Mol. Microbiol. 35: 961–973.
Clutterbuck, A. J., 1974 Aspergillus nidulans genetics, pp. 447–510 inHandbook of Genetics, Vol. 1, edited by R. C. King. Plenum, New York.
Clutterbuck, A. J., 1994 Linkage map and locus list, pp. 791–795in Aspergillus: 50 Years On, edited by S. D. Martinelli and J. R.Kinghorn. Elsevier, Amsterdam.
Cove, D. J., 1966 The induction and repression of nitrate reductasein the fungus Aspergillus nidulans. Biochim. Biophys. Acta 113: 51–56.
De Lucas, J. R., S. Valenciano, A. I. Dominguez, G. Turner and F.Laborda, 1997 Characterization of oleate-nonutilizing mu-tants of Aspergillus nidulans isolated by the 3-amino-1,2,4-triazolepositive selection method. Arch. Microbiol. 168: 504–512.
Erdmann, R., and G. Blobel, 1995 Giant peroxisomes in oleic acid-induced Saccharomyces cerevisiae lacking the peroxisomal mem-brane protein Pmp27p. J. Cell Biol. 128: 509–523.
Erdmann, R., M. Veenhuis, D. Mertens and W. H. Kunau,1989 Isolation of peroxisome-deficient mutants of Saccharomycescerevisiae. Proc. Natl. Acad. Sci. USA 86: 5419–5423.
Fillinger, S., M. K. Chaveroche, P. Van Dijck, R. de Vries, G.Ruijter et al., 2001 Trehalose is required for the acquisitionof tolerance to a variety of stresses in the filamentous fungus As-pergillus nidulans. Microbiology 147: 1851–1862.
Fujiki, Y., Y. Matsuzono, T. Matsuzaki and M. Fransen, 2006 Im-port of peroxisomal membrane proteins: the interplay of Pex3p-and Pex19p-mediated interactions. Biochim. Biophys. Acta 1763:1639–1646.
Gainey, L. D., I. F. Connerton, E. H. Lewis, G. Turner and D. J.Balance, 1992 Characterization of the glyoxysomal isocitratelyase genes of Aspergillus nidulans (acuD) and Neurospora crassa(acu-3). Curr. Genet. 21: 43–47.
Goodrich-Tanrikulu, M., K. Howe, A. Stafford and M. A. Nelson,1998 Changes in fatty acid composition of Neurospora crassa ac-company sexual development and ascospore germination. Micro-biology 144: 1713–1720.
Gurvitz, A., and H. Rottensteiner, 2006 The biochemistry of ole-ate induction: transcriptional upregulation and peroxisome pro-liferation. Biochim. Biophys. Acta 1763: 1392–1402.
Hettema, E. H., W. Girzalsky, M. Van denBerg, R. Erdmann and B.Distel, 2000 Saccharomyces cerevisiae Pex3p and Pex19p are re-quired for proper localization and stability of peroxisomal mem-brane proteins. EMBO J. 19: 223–233.
Hiltunen, J. K., A. M. Mursula, H. Rottensteiner, R. K. Wierenga,A. J. Kastaniotis et al., 2003 The biochemistry of peroxisomalbeta-oxidation in the yeast Saccharomyces cerevisiae. FEMS Micro-biol. Rev. 27: 35–64.
Hoepfner, D., D. Schildknegt, I. Braakman, P. Philippsen and H.F. Tabak, 2005 Contribution of the endoplasmic reticulum toperoxisome formation. Cell 122: 85–95.
Hynes, M. J., S. L. Murray, A. Duncan, G. S. Khew and M. A. Davis,2006 Regulatory genes controlling fatty acid catabolism andperoxisomal functions in the filamentous fungus, Aspergillus ni-dulans. Eukaryot. Cell 5: 794–805.
Jedd, G., and N. H. Chua, 2000 A new self-assembled peroxisomalvesicle required for efficient resealing of the plasma membrane.Nat. Cell Biol. 2: 226–231.
Kawasaki, L., and J. Aguirre, 2001 Multiple catalase genes are differ-entially regulated in Aspergillus nidulans. J. Bacteriol. 183: 1434–1440.
Kawasaki, L., A. Farres and J. Aguirre, 1995 Aspergillus nidulansmutants affected in acetate metabolism isolated as lipid nonutil-izers. Exp. Mycol. 19: 81–85.
Kiel, J. A., I. J. van der Klei, M. A. van den Berg, R. A. Bovenberg
and M. Veenhuis, 2005 Overproduction of a single protein, Pc-Pex11p, results in 2-fold enhanced penicillin production by Pen-icillium chrysogenum. Fungal Genet. Biol. 42: 154–164.
Kiel, J. A., M. Veenhuis and I. van der Klie, 2006 PEX genes infungal genomes: common, rare or redundant. Traffic 7: 1291–1303.
Kim, H-S., K-Y. Han, K-J. Kim, D-M. Han, K-Y. Jahng et al., 2002 TheveA gene activates sexual development in Aspergillus nidulans.Fungal Genet. Biol. 37: 72–80.
Kimura, A., Y. Takano, I. Furusawa and T. Okuno, 2001 Peroxi-somal metabolic function is required for appressorium-mediatedplant infection by Colletotrichum lagenarium. Plant Cell 13: 1945–1958.
Kionka, C., and W. H. Kunau, 1985 Inducible beta-oxidation path-way in Neurospora crassa. J. Bacteriol. 161: 153–157.
Klein, A. T., M. Van denBerg, G. Bottger, H. F. Tabak and B.Distel, 2002 Saccharomyces cerevisiae acyl-CoA oxidase followsa novel, non-PTS1, import pathway into peroxisomes that is de-pendent on Pex5p. J. Biol. Chem. 277: 25011–25019.
Kragt, A., T. Voorn-Brouwer, M. van den Berg and B. Distel,2005 Endoplasmic reticulum-directed Pex3p routes to peroxi-somes and restores peroxisome formation in a Saccharomyces cer-evisiae pex3D strain. J. Biol. Chem. 280: 34350–34357.
Kunze, M., F. Kragler, M. Binder, A. Hartig and A. Gurvitz,2002 Targeting of malate synthase 1 to the peroxisomes of Sac-charomyces cerevisiae cells depends on growth on oleic acid. Eur. J.Biochem. 269: 915–922.
Kunze, M., I. Pracharoenwattana, S. M. Smith and A. Hartig,2006 A central role for the peroxisomal membrane in glyoxy-late cycle function. Biochim. Biophys. Acta 1763: 1441–1452.
Lara-Ortiz, T., H. Riveros-Rosas and J. Aguirre, 2003 Reactiveoxygen species generated by microbial NADPH oxidase NoxAregulate sexual development in Aspergillus nidulans. Mol. Micro-biol. 50: 1241–1255.
Lazarow, P. B., 2006 The import receptor Pex7p and the PTS2 tar-geting sequence. Biochim. Biophys. Acta 1763: 1599–1604.
Li, X., and S. J. Gould, 2003 The dynamin-like GTPase DLP1 is es-sential for peroxisome division and is recruited to peroxisomes inpart by PEX11. J. Biol. Chem. 278: 17012–17020.
Li, X., E. Baumgart, G. X. Dong, J. C. Morrell, G. Jimenez-Sanchez
et al., 2002 PEX11alpha is required for peroxisome prolif-eration in response to 4-phenylbutyrate but is dispensable forperoxisome proliferator-activated receptor alpha-mediated per-oxisome proliferation. Mol. Cell. Biol. 22: 8226–8240.
Lingard, M. J., and R. N. Trelease, 2006 Five Arabidopsis peroxin11 homologs individually promote peroxisome elongation, du-plication or aggregation. J. Cell Sci. 11: 961–972.
Lockshon, D., L. E. Surface, E. O. Kerr, M. Kaeberlein and B. K.Kennedy, 2007 The sensitivity of yeast mutants to oleic acid im-plicates the peroxisome and other processes in membrane func-tion. Genetics 175: 77–91.
Maggio-Hall, L. A., and N. P. Keller, 2004 Mitochondrial b-oxida-tion in Aspergillus nidulans. Mol. Microbiol. 54: 1173–1185.
1368 M. J. Hynes et al.

Maggio-Hall, L. A., R. A. Wilson and N. P. Keller, 2005 Fun-damental contribution of b-oxidation to polyketide mycotoxinproduction in planta. Mol. Plant Microbe Interact. 18: 783–793.
Maggio-Hall, L. A., P. Lyne, J. A. Wolff and N. P. Keller, 2007 Asingle acyl-CoA dehydrogenase is required for catabolism of iso-leucine, valine and short-chain fatty acids in Aspergillus nidulans.Fungal Genet. Biol. 45: 180–189.
Managadze, D., C. Wurtz, M. Sichting, G. Niehaus, M. Veenhuis
et al., 2007 The peroxin PEX14 of Neurospora crassa is essentialfor the biogenesis of both glyoxysomes and Woronin bodies. Traf-fic 8: 687–701.
Mooney, J. L., and L. N. Yager, 1990 Light is required for conidia-tion in Aspergillus nidulans. Genes Dev. 4: 1473–1482.
Nayak, T., E. Szewczyk, C. E. Oakley, A. Osmani, L. Ukil et al.,2006 A versatile and efficient gene-targeting system for Aspergil-lus nidulans. Genetics 172: 1557–1566.
Nishimura, M., J. Yamaguchi, H. Mori, T. Akazawa and S. Yokota,1986 Immunocytochemical analysis shows that glyoxysomes aredirectly transformed to leaf peroxisomes during greening ofpumpkin cotyledons. Plant Physiol. 80: 313–316.
Oakley, C. E., C. F. Weil, P. L. Kretz and B. R. Oakley, 1987 Clon-ing of the riboB locus of Aspergillus nidulans. Gene 53: 293–298.
Ohsugi, M., K. Miyauchi, K. Tachibana and S. Nakao, 1988 For-mation of a biotin precursor, pimelic acid, in yeasts from C18fatty acids. J. Nutr. Sci. Vitaminol. 34: 343–352.
Orth, T., S. Reumann, X. Zhang, J. Fan, D. Wenzel et al., 2007 ThePEROXIN11 protein family controls peroxisome proliferation inArabidopsis. Plant Cell 19: 333–350.
Osherov, N., and G. S. May, 2000 Conidial germination in Aspergil-lus nidulans requires RAS signaling and protein synthesis. Genet-ics 155: 647–656.
Osherov, N., and G. S. May, 2001 The molecular mechanisms ofconidial germination. FEMS Microbiol. Lett. 199: 153–160.
Palmieri, L., H. Rottensteiner, W. Girzalsky, P. Scarcia, F.Palmieri et al., 2001 Identification and functional reconstitu-tion of the yeast peroxisomal adenine nucleotide transporter.EMBO J. 20: 5049–5059.
Petriv, O. I., L. Tang, V. I. Titorenko and R. A. Rachubinski,2004 A new definition for the consensus sequence of the per-oxisome targeting signal type 2. J. Mol. Biol. 341: 119–134.
Platta, H. W., and R. Erdmann, 2007 Peroxisomal dynamics.Trends Cell Biol. 17: 474–484.
Ramırez, M. A., and M. C. Lorenz, 2007 Mutations in alternative car-bon utilization pathways in Candida albicans attenuate virulenceand confer pleiotropic phenotypes. Eukaryot. Cell 6: 280–290.
Ramos-Pamplona, M., and N. I. Naqvi, 2006 Host invasion duringrice-blast disease requires carnitine-dependent transport of per-oxisomal acetyl-CoA. Mol. Microbiol. 61: 61–75.
Ruprich-Robert, G., V. Berteaux-Lecellier, D. Zickler, A. Panvier-Adoutte and M. Picard, 2002 Identification of six loci inwhich mutations partially restore peroxisome biogenesis and/or alleviate the metabolic defect of pex2 mutants in Podospora.Genetics 161: 1089–1099.
Sambrook, J., E. F. Fritsch and T. Maniatis, 1989 Molecular Clon-ing: A Laboratory Manual, Ed. 2. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Sandeman, R. A., M. J. Hynes, J. R. S. Fincham and I. F. Connerton,1991 Molecular organisation of the malate synthase genes ofAspergillus nidulans and Neurospora crassa. Mol. Gen. Genet. 228:445–452.
Scherer, M., H. Wei, R. Liese and R. Fischer, 2002 Aspergillus nidu-lans catalase-peroxidase gene (cpeA) is transcriptionally inducedduring sexual development through the transcription factorStuA. Eukaryot. Cell 1: 725–735.
Schliebs, W., C. Wurtz, W. Kunau, M. Veenhuis and H.Rottensteiner, 2006 A eukaryote without catalase-containingmicrobodies: Neurospora crassa exhibits a unique cellular distribu-tion of its four catalases. Eukaryot. Cell 5: 1490–1502.
Schobel, F., O. Ibrahim-Granet, P. Ave, J. P. Latge, A. A. Brakhage
et al., 2007 Aspergillus fumigatus does not require fatty acid me-tabolism via isocitrate lyase for development of invasive aspergil-losis. Infect. Immun. 75: 1237–1244.
Schuller, H. J., 2003 Transcriptional control of nonfermentative me-tabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 43: 139–160.
Smith, J. J., M. Marelli, R. H. Christmas, F. J. Vizeacoumar, D. J.Dilworth et al., 2002 Transcriptome profiling to identifygenes involved in peroxisome assembly and function. J. Cell Biol.158: 259–271.
Smith, J. J., S. A. Ramsey, M. Marelli, B. Marzolf, D. Hwang et al.,2007 Transcriptional responses to fatty acid are coordinated bycombinatorial control. Mol. Syst. Biol. 3: 115.
Stanley, W. A., and M. Wilmanns, 2006 Dynamic architecture ofthe peroxisomal import receptor Pex5p. Biochim. Biophys. Acta1763: 1592–1598.
Szewczyk, E., A. Andrianopoulos, M. A. Davis and M. J. Hynes,2001 A single gene produces mitochondrial, cytoplasmic, andperoxisomal NADP-dependent isocitrate dehydrogenase in Asper-gillus nidulans. J. Biol. Chem. 276: 37722–37729.
Tenney, K., I. Hunt, J. Sweigard, J. I. Pounder, C. McClain et al.,2000 hex-1, a gene unique to filamentous fungi, encodes themajor protein of the Woronin body and functions as a plugfor septal pores. Fungal Genet. Biol. 31: 205–217.
Tey, W. K., A. J. North, J. L. Reyes, Y. F. Lu and G. Jedd, 2005 Po-larized gene expression determines Woronin body formation atthe leading edge of the fungal colony. Mol. Biol. Cell 16: 2651–2659.
Thieringer, R., and W. H. Kunau, 1991 b-Oxidation system of thefilamentous fungus Neurospora crassa. Structural characterizationof the trifunctional protein. J. Biol. Chem. 266: 13118–13123.
Thoms, S., and R. Erdmann, 2005 Dynamin-related proteins andPex11 proteins in peroxisome division and proliferation. FEBSJ. 272: 5169–5181.
Titorenko, V. I., and R. A. Rachubinski, 2001 The life cycle of theperoxisome. Nat. Rev. Mol. Cell Biol. 2: 357–368.
Todd, R. B., M. A. Davis and M. J. Hynes, 2007 Genetic manipula-tion of Aspergillus nidulans: meiotic progeny for genetic analysisand strain constuction. Nat. Protoc. 2: 811–821.
Tsitsigiannis, D. I., and N. P. Keller, 2007 Oxylipins as develop-mental and host-fungal communication signals. Trends Micro-biol. 15: 109–118.
Tsitsigiannis, D. I., T. M. Kowieski, R. Zarnowski and N. P. Keller,2004a Endogenous lipogenic regulators of spore balance in As-pergillus nidulans. Eukaryot. Cell 3: 1398–1411.
Tsitsigiannis, D. I., R. Zarnowski and N. P. Keller, 2004b Thelipid body protein, PpoA, coordinates sexual and asexual sporu-lation in Aspergillus nidulans. J. Biol. Chem. 279: 11344–11353.
Tsitsigiannis, D. I., T. M. Kowieski, R. Zarnowski and N. P. Keller,2005 Three putative oxylipin biosynthetic genes integrate sex-ual and asexual development in Aspergillus nidulans. Microbiol-ogy 151: 1809–1821.
Valenciano, S., J. R. Lucas, A. Pedregosa, I. F. Monistrol and F.Laborda, 1996 Induction of beta-oxidation enzymes and mi-crobody proliferation in Aspergillus nidulans. Arch. Microbiol.166: 336–341.
Valenciano, S., J. R. De Lucas, I. Van der Klei, M. Veenhuis and F.Laborda, 1998 Characterization of Aspergillus nidulans peroxi-somes by immunoelectron microscopy. Arch. Microbiol. 170:370–376.
van Roermund, C. W., R. Drissen, M. Van denBerg, L. Ijlst, E. H.Hettema et al., 2001 Identification of a peroxisomal ATP car-rier required for medium-chain fatty acid b-oxidation and nor-mal peroxisome proliferation in Saccharomyces cerevisiae. Mol.Cell. Biol. 21: 4321–4329.
Visser, W. F., C. W. T. van Roermund, L. Ijlst, H. R. Waterham andR. J. A. Wanders, 2007 Metabolite transport across the perox-isomal membrane. Biochem. J. 401: 365–375.
Wang, Z. Y., D. M. Soanes, M. J. Kershaw and N. J. Talbot,2007 Functional analysis of lipid metabolism in Magnaporthe gri-sea reveals a role for peroxisomal fatty acid b-oxidation duringappressorium-mediated plant infection. Mol. Plant Microbe In-teract. 20: 475–491.
Yan, M., N. Rayapuram and S. Subramani, 2005 The control of per-oxisome number and size during division and proliferation.Curr. Opin. Cell Biol. 17: 376–383.
Yuan, P., G. Jedd, D. Kumaran, S. Swaminathan, H. Shio et al.,2003 A HEX-1 crystal lattice required for Woronin body func-tion in Neurospora crassa. Nat. Struct. Biol. 10: 264–270.
Communicating editor: A. P. Mitchell
Peroxisomal Functions in A. nidulans 1369