Embed Size (px)
Citation preview

CocMl Lopm Pr'Oeeuu edited by B. KjerfYe(EJ..seyjer Oeeanocraphy Series. 60)C 1m Elsevier Science Publisben B.V. All richu retelTed
Chapter 12
Coastal Lagoons as Fish Habitats
Alejandro Yafiez-Arancibia-, Ana Laura Lara Dominguez- andDaniel Paulyb
-Programa de Ecolog{a, Pesquerlas y Oceanograf£a. del Golfo de Mexico(EPOMEXJ, UniversidadAuttSnoma de Campeche, Apartado Postal 520,Campeche 24000, Campeche, Mexico;blnternational Center for Liuing Aquatic Resources ManagementaCLARM), M.C. P.O. Box 2631, 0718 Makati, Manila, The Philippines
363
'Many fish species use coastal lagoons as habitats during at least some part of theirlife cycles. We discuss these characteristics with emphasis on latitudinal differences. Higher biomass of coastal fishes will occur when lagoon-estuarine habitatspace is available and accessible. The concept of seasonal programming describeshow juveniles use that lagoon habitat space. We also consider the lagoon~stuarine
dependence of subtropical and tropical coastal fishes.
Introduction
Coastal lagoons are defined in geomorphological terms (Phleger, 1969;Lankford, 1977; Kjerfve, 1986; Yanez-Arancibia, 1987). In this, they differgeologically from estuaries, which are defined in terms of the hydrologicalconditions and gradients of salinity and temperature (density) (Cameronand Pritchard, 1963; Pritchard, 1967; Fairbridge, 1980; Yciiiez-Arancibia,1987; Kjerfve and Magill, 1989). Kjerfve (1989) proposed a new classification for "inland waters which are connected to the sea-, based on six classes(estuaries, coastal lagoons, fjords, bays, tidal rivers and straits), defined byvarious combinations of physical and geomorphological processes.
Kjerfve's assignment of adjacent classes to estuaries and coastal lagoonsis due to their sharing numerous characteristics. This is also the reason whyDay and Y8.fiez-Arancibia (1982) had earlier proposed the (ecological) concept of "lagoon-estuarine environments-, Le. shallow semi-enclosed waterbodies ofvariable volume, connected to the sea in a permanent or ephemeralmanner', with variable temperature and salinities, permanent muddy bottoms,high turbidity, irregular topographic characteristics, and biotic elements.

364
In this chapter we present the fish fauna occurring in various lagoonestuarine environments, with emphasis on those features of lagoons thatare actually 'used' by the fish populations, and on some of the mechanismsinvolved. therein.
We concentrate on fishes because they generally constitute the overwhelming bulk (up to 99%) of the lagoon--estuarine nekton, both in terms ofnumbers and biomass (McHugh, 1967). The few nonpiscine members of thelagoon--estuarine nekton are squids, portunid crabs, penaeid shrimps, reptiles, and mammals such as dolphins.
Much information is now available on the life cycles of fishes in relationto lagoon--estuarine environment (McCleave et aI., 1984; Yanez-Arancibia,1985; Dadswell et aI., 1987; Weinstein, 1988; Henderson and Margetts.1988). and various schemes have been proposed to link these cycles withenvironmental parameters and the observed high abundances of fishes inlagoons and estuaries (Day el aL, 1981; Pauly, 1982; Weinstein, 1982;Yanez-Arancibia, 1986).
Two important points have emerged from these studies:- the utilization of the lagoon-estuarine environment is an integral part of
the life cycle ofnumerous fishes, particularly so in the Neotropics and theIndo--Pacific (Longhurst and Pauly, 1987);
- the lagoon--estuarine environment is mainly utilized by juveniles andyoung adults.We shall here consider: (1) the fishes that 'use' coastal lagoons; and (2) the
features of the lagoon habitat(s) that are actually being used by these fishes.
Which Fishes Use Coastal Lagoons?
Although the exact species and dominance ranks change between sites,the dominant fishes of lagoon--estuarine habitats usually belong to only afew taxonomic groups. High latitude estuaries have simple ichthyofaunas,dominated in the northern hemisphere, by SaImonidae (salmon and trout),Osmeridae (smelt and capelin), Gasterosteidae (sticklebacks),Ammodytidae(sandlance), and Cottidae (sculpins) and their relatives (Dadswell et aI.,1987; Day et al., 1989). In the southern hemisphere the SaImonidae arereplaced by their ecological analogs, the Galaxioidae (McDowall and Eldon,1980; McDowall and Frankenberg, 1981). Important families for temperatesystems are theAnguilIidae (freshwater eels), Clupeidtu (herring-like fish),Engraulidae. (anchovies),Ariidae (marine catfish), Cyprirwdonti.dae (killifish),Gadidtu (cod), Gasterosteidae (stickleback), Serranidae (basses), Sciaenicku (sea trout, drums, croakers), Sparidae (sea-breams), and Pleuroneetidae(flounders) (Haedrich,1983; McCleave et al., 1984).
In the tropics and subtropics, the important families are the Clupeidae,Engraulidae,Ariidae, Scuumidae,Poecillidae (guppy), Chanidae (milkfish),.
365
Synodontidae (lizardfish). Belon-idae (silver gars), Mugilida.e (mullets), Poly·nemidae (threadfins), Gobiidae (gobies), Cichlidae (tilapia and related fishes),Dasyatidcu (rays), Telraodontidae (puffers), Gerreidae (mojarras), £eiegnathidae (slipmouth, ponyfish), Pomadasyidae (grunts), and various familiesofflatfishes, such as theBothidae, So/eidae and Cyrwglossidae (Pauly. 1982;yanez·Arancibia, 1985, 1986; Lowe·McConnell, 1987).
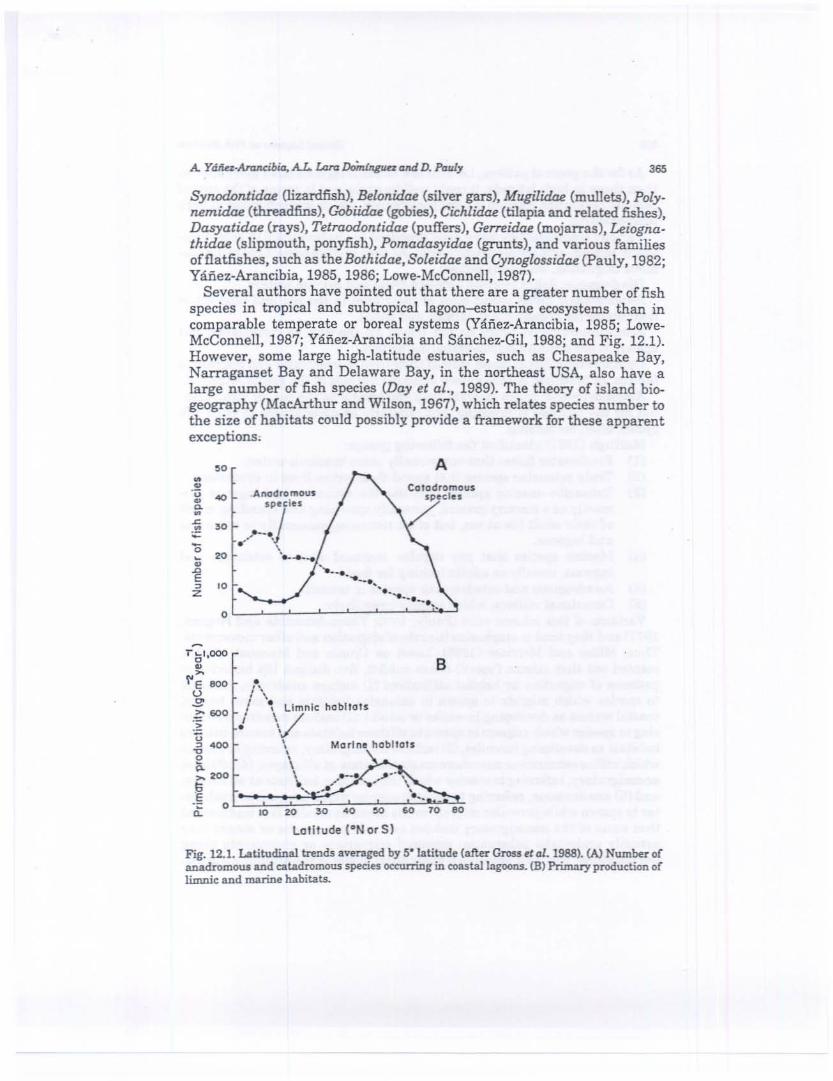
Several authors have pointed out that there are a greater number of fishspecies in tropical and subtropical lagoon--estuarine ecosystems than incomparable temperate or boreal systems (Yaftez-Arancibia, 1985; Lowe·McConnell, 1987; Yafiez·Arancibia and Sanchez-Gil, 1988; and Fig. 12.1).However, some large high-latitude estuaries, such as Chesapeake Bay,Narraganset Bay and Delaware Bay, in the northeast USA, also have alarge number of fish species Way et al., 1989). The theory of island biogeography (MacArthur and Wilson, 1967), which relates species number tothe size of habitats could possibly provide a framework for these apparentexceptions;
'0 A
'0- '"•~, '0z
COlodromousspec:les
B
Marine hoblTOTS
-r~l.000l'.
~E 000
\;~ "'0~
~ ..oo
K~ >00
.~ ~-=~~.g:::~--:;;--;;:~~~ oL 10 20 30 "0 !lO 60 70 eo
Lotitude (ONorS)
Fig. 12.1. Latitudinal trends averaged by 5·'atitude (after Gross n aI. 1988). (A) Number ofanadromotU and eatadromous species occ:urring in COastallagOOllS. (.8) Primary production oflimnic: and marine habitats.

366 Coa.tai Lapotu eu FWi Habitats
As for the general pattern, i.e. that low latitude lagoons have more speciesthan those in high latitude, it could well be explained in terms of the cost ofosmotic regulation relative to total metabolism, which is much lower at hightemperatures.
Various classifications have been proposed to analyze the fish communities in Jagoon-estuarine environments. As they involve some form ofdiadromous migration, we briefly define:
Diadromous: fishes migrate between fresh and marine waters.Anadromous: fishes spend most of their lives in the sea and migrate to
freshwater for breeding (e.g. salmon and shad).Semi·aruuiromous: fishes do not travel all the way to freshwater, but
spawn in low-salinity lagoon--estuarine waters.Catadromous: fishes spend mostoftbeir lives in freshwater and migrate to
the sea for breeding. A well-known example are eels oftbe genus Anguilla.Amphidromous: fishes migrate from freshwater to the sea, or vice versa,
not for the purpose of breeding, but regularly at other stages in their lifecycles, often for feeding.
McHugh (1967) identified the following groups:(1) Freshwater fishes that occasionally enter brackish·water.(2) Truly estuarine species that spend their entire lives in estuaries.(3) Estuarine-marine species that use the estuaries and lagoons pri-
marily as a nursery ground, generally spawning and spending muchof their adult life at sea, but often returning seasonally to estuariesand lagoons.
(4) Marine species that pay regular seasonal visit to estuaries andlagoons, usually as adults looking for food.
(5) Anadromous and catadromous species in transit.(6) Occasional visitors, which appear. irregularly.Variants of this scheme exist (pauly, 1975; Yanez·Arancibia and Nugent,
1977) and they tend to emphasize the role ofmigration and other movements.Thus, Miller and Merriner (1985), based on Cronin and Mansueti (1971),pointed out that marine ('sport') fishes exhibit, five distinct life histories orpatterns of migration for habitat utilization: (1) inshore anadromy, referringto species which migrate to spawn in estuarine habitats and move towardcoastal waters as developing juveniles or adults; (2) inshore catadromy, referring to species which migrate to spawn in offshore habitats and toward inshorehabitats as developingjuveniles; (3) inshore nonmigratory, referring to specieswhich utilize estuarine or nearshore coastal habitats at all stages; (4) offshorenonmigratory, referring to species which utilize ocean habitats at all stages;and (5) anadromous, referring to oceanic species which migrate to freshwater to spawn while juveniles migrate to the ocean as smolts. It is understoodthat some of the nonmigratory inshore and offshore species or stages mayactually undertake substantial seasonal migrations or movements alongthe coastline. Fig. 12.2 presents a conceptual model of the stimuli involved.

Light. Temperature,Prey Distribution
Habitat:
physicalProcess:
FIshes'ActivIty:
FIshes'BehavIor:
Stlmull:
, ,r---Plankton----J;--Nearshore---+!-Channel or Mouth-l-Estuary or Lagoon.....
I. ' ...1i! ! ..' ,
Ekman Transport, Ii ..··..Counlefcurronts, ! Alongshore Currenls i Tidal Flux
'''1;'' I. i A/-----------: --: ., -....
j 6oontal~ 10 r--"'~.-.-,Drifl +_ L~90on-EsIUaIY-----...T~~~~~:~m ..... _ -~- ~~~~~:~
r ;oaVertical Movements _l_~ Circadian • __ .1_ ..... : Circa~dal H:lbilat Seloction
f.
I A,,;." rR,,,m. ! A';~."r' rl : Tidal Zeilgeber Habitat Slruclure: )0- Diel ! (Salinity, Temperature. (Covar. Depth.J Zeilgcber ! Turbidity, Pressure. Substrale, Proy.: : OUnclOry Cues. Turbulonce,! i Electric Fields) Conspecilics): : .'.. . .: : '.
I---------AppetitiveBehavior---------.......~
wFig. 12.2. Conceptual model of the slimuli and behaviors involved in fish movements towarns estuaries and coastnllagoons (after Doehlert ~
and Mundy,198B),

368 Coastal LosooTl! as Fish Habitau
Recently, Zijistra (988) based on Louis and Lasserre (1982), presented aclassification consisting of six types, pertaining to different regions rangingfrom temperate to tropical climates and which may therefore have a worldwide application: (1) anadromous and catadromous species; (2) residentspecies spending their entire lives in the coastal areas; (3) marine species,spawning in the coastal region and using it as nursery ground; (4) marinespecies, which use the coastal area as a nursery area, but which spawn andspend most of their adult life in the offshore region; (5) marine species,which pay regular seasonal visits to the coastal zone, usually as adult; and(6) adventitious visitors, which appear irregularly and have no apparentestuarine or coastal requirements.
Real fishes, however, will not adhere strictly to any of these classifications. Rather, a number of populations will belong to several types, inde·pendent of how well-conceived the types are.
More interesting are classifications that are quantitative, even if thisimplies that they become site-specific in the process. Deegan and Thompson(1985) provided such classification for the Mississippi delta. Similarly,Yaiiez-Arancibia et al. (l988b) proposed a classification scheme for Terminos Lagoon, Southern GulfofMexico, which provides salinity ranges for thevarious groupings, i.e.:
freshwater spawners, occurring in waters < 10%0.brackish water groups,limited to 10-34%0.marine spawners, occurring in waters ~ 35%0.
Classifications of this type can be constructed, more or less rigorously, forany desired level of resolution, using clu~tering-typetechniques (i.e. usingstation species lists vs. observed environmental parameters; Fig. 12.3)andlor the results of physiological experiments, e.g. on salinity tolerance.
Fish production in lagoon-estuarine systems is generally high (Day et al.,1989), and this generally leads to high fishery catches. One reason for thishigh production is that many of the fish species of importance in lagoonestuarine environments are either: (1) r-selected sensu (Pianka, 1978), i.e.relatively small, fast-growing animals with high productionJbiomass (PIB)ratio; or (2) K-selected juveniles, i.e. fast-growing, high production stagefishes whose adult form, however, typically occurs outside the lagoon. Thehigh PIB ratios of the constituent species are not the only explanation forthe generally high production of lagoon-estuarine communities. The highbiomasses themselves, i.e., the high carrying capacities of these ecosystemsin terms of food availability to fishes also playa crucial role.

370 Coo.stal LcgOOM Q$ Fish Habitats
Which Features of Lagoon-Estuarine Habitats Do Fishes ActuallyUse?
The two most important processes for living organisms are:(1) energy acquisition, i.e. feeding, as needed to maintain their struc
ture, and to grow;(2) reproduction: only those organisms are still around whose ancestors
bred and who endowed them with ability to do the same.Except for very few species, fishes do not use lagoons for reproduction
proper, i.e. for late maturation, spawning/fertilization. It is tempting torelate this to the ephemeral nature and the strong environmental variability of these systems. Rather, it is as feeding grounds that lagoon-estuarineenvironments are important to fishes.
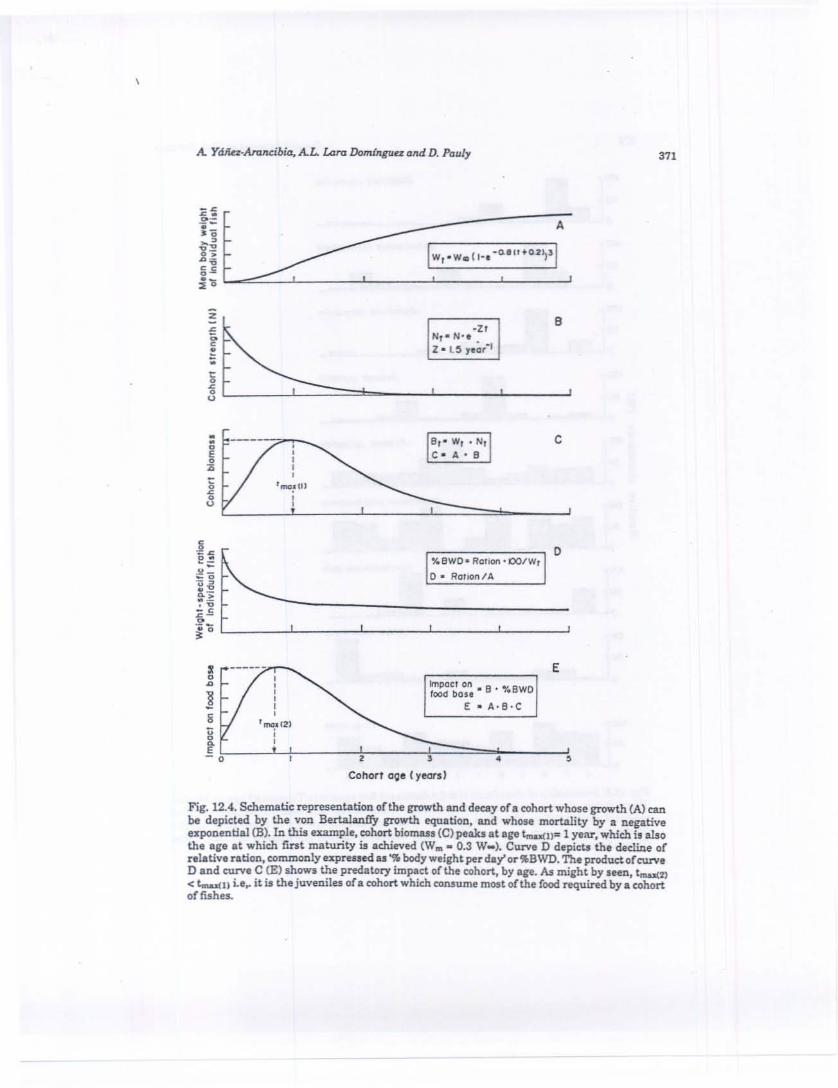
The growth/natural mortality schedules of unexploited fish cohorts generally imply that biomasses are maximized near the mean age at firstmaturity ofthe individual fish, an adaptation obviously related to maximizing the output of reproductive products. However, small fishes always eatmore per unit body weight than large fishes (Pauly, 1986). This results inthe maximum of the absolute food consumption curve, of any stable fishpopulation being shifted to the left, toward the juveniles (Fig. 12.4).
Thus, it is the juveniles of fish populations, and not the adults, whichexert the greatest pressure on the food supply. If a coastal population (B')of adult fishes is to maintain itself, it must place its juveniles in a habitatwith a carrying capacity »B'. This point, apparently not made in theextensive literature on lagoon and estuaries, provides an objective criterionfor referring to these water bodies as critical habitats.
However, for any coastal fish population, randomly spawning on the shelfadjacent to lagoon-estuary systems and letting the juveniles find their wayinto these systems, just does not happen. Rather, seasonal variations ofabiotic parameters and of food supplies and competitors must be accommo~dated, and this is what leads to the phenomenon labeled here as seasonalprogramming, which refers to the temporal and spatial sequence of lagoonhabitat uses by juvenile fishes (Yaiiez-Arancibia et al., 1988a).
Seasonal programming of food resource utilization by various species inTerminos Lagoon is illustrated in Fig. 12.5. Various species use the lagoonat different times, thus reducing their competitive interactions. At the levelof a given species, seasonal programming implies that its representatives(mainly juveniles, see above) feed, in the course of their ontogeny, on asuccession of different food types, often gathered from different subareaswithin a given lagoon (Fig. 12.6). This sequential utilization oHoOO type andfeeding location may be highly repetitive, hence predictable, between seasons.
In areas where lagoon systems have maintained themselves over longperiods, evolutionary mechanisms may emerge which stabilize and refinesuch seasonal programming, making the fish population in question'gradu~
A Ydlln·Atundbio., AI.. Lmu Dominguu and D. Paul, 371
!Hz
•o<
~•
·z,NT- N·.z· L~ )'.0('
B
§L --'-=::::::::=*= -'- ...Je.. ---'
----7-.._•,,,
'1IIG~1I1
c
o% ewo - RlInan' CIO/WI
O· RlIlilIn/A
!..~
~:_ 0'.§.~.~-<c.2'_.0 '---__-'-__-'-__-'-__-'-_--'
EImpacr ont<lQd bon· B' %BWO
E • A·B·C
,,,,,•
'''''',,12)u ,
~ !:..---.J''-t------t-----=!f=::::::==-;------:- 0 2 3 .. :)
Cohor' ot;le I years}
•o~
Fig, 12.4. SchematierepresentatioD oflhe growth and decayofa cohort whose growth (A) canbe depicted by the von Bertalanfl'y growth equation, and whose mortality by a negativeexponential {B}. In this ezample, cohort biomus eC) peaks at age t...u(n= 1 yelU', which is alsothe are at which. flnlt maturity is achieved (W... 0.3 W..). Curve D depicts the decline ofrelative ration. commonly expressed as'" body wei,ht per day' or'J'BWD. The product. ofeurveD and curve C (E) shows the predatory impact ofthe cohort, by age. M might by seen, tlDU(%)
< t-..(n i.e.. it is thejuveniles oia cohort which con.sume most ofthe food nquired by a cohortofflShe!.

372
AnchrH1 mitch/III
Bairrlirlla cllrysouNI
=
_5:L__
000
'"0
':f Cd 11 .;-;H~ Gl1
"~f ~•Ii·
cucin<>slomus~
Ckh/tnomt1 lIrr'phlht1/mus
h' i3iEl. !
•~ooo0~< ",~ .' A .0 0
.•" 000
E• '"<r0
"""
0
:r B$hto
~ ~ MAW J J A , 0 N 0
Fig. 12.5. Seasonalityofabundance ofeight dominant fish 5pecies inTermiDOfI Lagoon, MWoo.These fishes &how a clear seasonal sequence of abundance and habitat utilization calledseasonal programming. Note that the sum of the relative abundance of these eight speciesOower graph) suggests an even utiliution of Tumin05 Lagoon throughout the year (aft.erYAiiu-Aranc:ibia d oJ., 1988a).

A Ydllu-Arandbia, AI.. L4ra Domingu.a and~. Pau.ly
OnlOQenic chonQu
373
LOQOOn/esluory
SeosonolchonQet
<. '- IDiurnol_ ..,:, chonQu.~ '-
"So.
-----"" Y,," ",de.'-----Y consume.
-'- seC.,nd o,de'-., conSllmo,
-----Irr. Tn;.d ."de,----,. c.,nsllme,
Fig. 12.6. Conceptual diagram illustrating the trophodynarnic eetllogy of a hypotheticallagoon-estuarine fish species. The elements of time and time of the day, season and fish age(size), and space are shown. The spatial elements include transfers between ecosystem, e.g.,from marine areas via lagoons/estuarine to riverine areas, as well as between habitats ofthesame system, e.g. from oyster reefs to seagrass beds within the lagoon/estuarine system (afterYlii\.ez·Arancibia et 01.,1986).
ally more dependent on the lagoon system for the maintenance of highbiomass.
In other areas, where the lagoon systems do not persist or are notregularly open to juvenile immigrations, e.g. off West Africa, use of thelagoons are a matter of random movements along the coasts, and of aninshore movements, i.e. errances. This implies (1) a lower conversion ofprimary and secondary production into fish flesh, and hence (2) lowerbiomasses of coastal fishes,
The difficulty inherent in practical separation of random alongshore!inshore movements, i.e. errances, from evolutionarily fine-tuned, aimedmovements towards and within lagoons, and the difficulties involved inpreoise field eltimatioo ofbIomass and conversion efficiencies, make rigorous testing of these hypotheses difficult. This is the main reason for thedebate on the degree of dependence of tropical coastal fishes 00 coastallagoons.
References
Boehlert, G.W. and Mundy, B.C. 1988 Roles of behavioral and physical factors in larvaland juvenile fish recruitment in estuarine nursery areas. In Larval fish and shellfishtraMport throu6h tnld. (Weinstein, M.P., ed.). American Fisheries Society Sympo.slum S. Bethesda, Maryland, 51-67.

374
Cameron, W.M. and Pritchard, D.W. 1963 Estuaries. In The Sea 2. Wiley Intersciencc,London, 306-324.
Cronin, L.E. and Mansueti, A.J. 1971 The biology oftbe estuary. In Symposium on thebiological significance of estuaries (Douglas, P.A. and Stroud. R.H.A., eds). SportFishing Institute, Washington, DC, 14-39.
Dadswell. M.J., KJauda, R.J., Moffitt, C.M., Saunders, R.L., Rulifson. RA and Cooper,J.E. 1987 Common slrotegUs of anadromous and catadromous (/,SMS. AmericanFisheries Society Symposium 1, Bethesda, Maryland.
Day, J.H., BIaber, S.J.M. and Wallace, J.H. 1981 Estuarine fishes. In Estuarine ecofugywith particu.lar ~fennce to SoutJum Africa (Day, J.H., ed.), A.A. Balkema, Rotterdam, 197-221.
Day, Jr., J.W. and Yadez-Arancibia, A. 1982 Coastal lagoons and estuaries: ecosystemapproach. Cieneia Intcram 22, 12-26.
Day, Jr., J.W., Hall, C., Kemp, M.W. and YAfiez-Aranc:ibia. A. 1989 Nekton, the freeswimming consumers. In Estuarine ecology. Wiley-Interscience, John Wiley andSons, New York, 377-437.
Deegan, L.A. and Thompson, B. 1985 The ecology offish communities in the MississippiRiver deltaic plain. In Fish community ecology in ~stuari~s and. coastal lagoons:towards an ~cosystem integration (Yanez-Arancibia, A., ed.). UNAM Press, Mbico,DF,35-56.
Fairbridge, R W. 1980 The estuary: its definition and geodynamics cycle. In Chemistryand biochemistry of estuaries (Olausson, E. and CaOO, I., eds). Wiley-lnterscience.John Wiley and Sons, Inc., New York, 1-36.
Gross, M.R, Coleman, R..M. and McDowall, R..M. 1988 Aquatic productivity and theevolution of diadromous fish migration. &una 2.39, 1291-1293.
Haedric.b, RL. 1983 Estuarine fishes. In Ecosystem 0{ tM world. 26. Estuariu alUien.cloud seas (Ketchum, RH., ed.). Elsevier Scientific Publishing Company. Amsterdam, 182-207.
Henderson. P.A. and Margetts, A.R 1988 Fish in estuaries. Journal ofFish Biclogy 33,254 pp.
Kjerfve, B. 1986 Comparative oceanography of eoastallagoons. In Estuarin~ lJariability(Wolie, D., ed.). Academic Press, Inc., New York, 63-82.
K;ierfve, R 1989 Estuarine geomorphology and physical oceanography. In Estuarineecology (Day, Jr., J.W., Hall, CA.S., Kemp, M.W. and yanez.Arancibia, A., eds).Wiley·Interscience, John Wiley and Sons, New York, 47-78.
K;ierfve, R and Magill, K 1989 Geographic and hydrodynamic characteristics ofshallowcoastal lagoons. MarifU! Geology 88, 187-199.
Lankford, RR 1977 Coastal lagoons of Mexico: their origin and classification. InEstUlJrifU! PrrJCe3SeS (Wiley, M., ed.). Academic Press, Inc., New York, 182-215.
Longhurst, A.R and Pauly, 0.1987 Ecology oftropU:aJ. OCNJU. Academic Press Inc., SanDiego, California.
Louis, M. and La.sserre, G. 1982 Etude des peuplementa des poissons dans lea lagunesdes mangroves de Guadeloupe (Antilles franli8ises). In Coa.stal Wgoons (Lasserre, P.and Postma.. H., eds). Oceano'. Acta 5, 333--338.
Lowe-McConnell, R.H. 1987 Ecological studies in tropica1fish communitu$. CambridgeUniversity Press, Cambridge, England.
MacArthur, R.H. and Wilson, E.D. 1967 The theory of island biogeography. PrincetonUniversity Press, Princeton, New Jersey.

, A. YWln-Amncibia, A.I.. Lara Domlng~ and D. Pauly 375
McCleave, J.D., Arnold, G.P., Dodson, J.J. and Neill, W.N. 1984 Mechanisms ofmigranons infislus. Plenum Press, New York.
McDowall, RM. and Eldon, G.A. 1980 The ecology ofwhitebait migrations (Galaxiidae:Galaxias spp). fulunes Resource Bulletin 20. 172 pp.
McDowall, RM. and Frankenberg, R.S. 1981 The Galaxiid fishes ofAustralia. Rec. Austr.Mus. 33, 443-605.
McHugh. J .L. 1967 Estuarine nekton. InEstuanes (Lauff, G.H., ed,). Special Publication83. American Association for the Advancement ofScience, Washington, DC, 581-619.
Miller. J.M. and Merriner, J.V. 1985 Determinants of habitats dependency in marinesport fishes. In Proceedings of First World Angling Conference, France Cap·d'Agde,12-18 September 1984. IGFA, Fort Lauderdale, Florida, 119-130.
Pauly, D. 1975 On the ecology of a small West-Africa lagoon. Ber. dt. wiss. Kommn.Meeresforsch 24, 4&-62.
Pauly, D. 1982 The fishes and their ecology. In Small-scale fisheries ofSan Miguel Bay.Philippines: biology and stock assessment (Pauly, D. and Mines. A.N., eds.). ICLARMTechnical Reports 7, 124 p. International Center for Living Aquatic ResourcesManagement, Manila Philippines. 15-33.
Pauly, D. 1986 A simple method forestimatingthe food consumption offish populationsfrom growth data and food conversion experiments. Fisheries Bulletin 84, 827839.
Phleger, F.B. 1969 Some general features of coastal lagoons. In Coastal lagoons. asymposium (Ayala-Castanares. A. and Phleger, F.R, OOs). Symposium International,Coastal Lagoons UNAM-UNESCO, Mexico, DF, 5-26.
Pianka, E.R. 1978 Evolutionary ecology. Second Editon. Harper and Row Publishers.New York.
Pritchard, D.W. 1967 ObselVation on circulation in coastal plain estuaries. In Estuaries(Lauff, G.H., ed.). Special Publication 83. American Association for the AdvancementofScience, 37-44.
Weinstein, M.P. 1982 Commentary: a need for more experimental work in estuarinefisheries ecology. Northeast GulfScience 5, 59-64.
Weinstein, M.P. 1988 Laroal fish and shellfish transport through inlets. AmericanFisheries Society Symposium 3. Bethesda, Maryland.
Ydiiez-Arancibia, A. 1985 Fish community ecology in estuaries and coastal lagoons:towards an ecosystem integration. UNAM Press, Mexico, OF.
Yaiiez-Arancibia, A. 1986 Ecologia de comunidades de peces en sistemas costeros tropi.cales. In Ecologia de la zona costera: andlisis de sjete t6picos. AGT Editor. Mexico.OF. 127-154.
Yaiiez-Arancibia, A. 1987 Lagunas costeras y estuarios: cronologfa criterios y conceptospara una clasificaci6n ecol6gica de sistemas costeros. Publicacion ACIESP 54, 1-36.
YAiiez-Arancibia, A. and Day, Jr.• J.W. 1982 Ecological characterization of TerminosLagoon, a tropical Jagoon-estuarine system in the Southern Gulf of Mexico. InCoastal lagoons (Lasserre. P. and Postma, H., eds.). Oceanol. Acta 5 431-440.
Yanez-Arancibia, A. and Nugent, R. 1977 El papel ecol6gico de los peces en estuarios ylagunas costeras. An. Cento. Cienc. del Mar y Limnol. UnilJ. Nac. AUlon. M~:r.ico 4,107-114.
Ydil.ez-Arancibia, A. and Sanchez-Gil, P. 1988 Ecologia de los recursos demersaltsmarinos: fundamentos en costas tropicalts. AGT Editor. Mexico. DF.
yanez·Arancibia, A., Lara-Dominguez, A.L., Aguirre, A. and Diaz, S. 1986 Feeding

376 CoaswI Lagoons a. FisA HabitatS
ecology oftropieal estuarine fishes in relation to recruitment processes. In IDeI FADWorJuhop on rrcruitnwtl in tropical coastal demersal communitiu, worksJwp &portNo. 44-Suppumen1 (Yat\.e%-Arancibia.A and Pauly, D., eds). United Nations Educational, Scientific and Cultural Organization, Pam, 73-94..
Yiftez-Arancibia. A.. Lara.Dominguez. A.L.. Rojas Galaviz, J.L., SAochez-Gil. p. Day.J r", J.W. and Madden, C.J. 1988a Sea.sonafbiomass and diversity ofestuarine fishescoupled with tropical habitat heterogeneity (Southern Gulf of Mexico). Journal ofFish Biology 33, 191-200.
YA1l.ez·Arancibia, A., Lara.Dominguez, AL.. SAnchez·Gil, P., Rojas Galaviz. J.L., Alvarez. H.t Sober6n Chavez. G. and Day, Jr., J.W. 1988b Dynamics of coastal nektoniccommunities in the Southern GulfofMe.xico. In EcolO/l:J ofcoo.stal ecosysunu in tJuSoutJum. GulfofMhU:o: TJu Terminosl."ogoon region (ylU'iez-Arancibia, A and Day,Jr., J.W., eds). Instituto de Ciencias del Mar y Limnologia Universidad NacionalAut.6noma Mexico, Coastal Ecology Institute, Louisiana State University, Mexico,DF, 357-380.
Zi,)ll1tra, J.J. 1988 Fish migrations between coastal and offshore areas. In Coastal offs1wre~cosysum inluactions: Iutlln rlOUS on coa.Jto.I end utlUJ.riru. studus (Jansson, B.O.,ed.). Springer-Verlag, Berlin, 257-272.










![Environmental Deterioration of the Tecomate Coastal Lagoon ...In tropical regions, the coastal lagoons are surrounded by a wide man-grove fringe [3]; this vegetation plays an important](https://img.pdfslide.us/doc/110x75/5e9e0359470e0840c75e5100/environmental-deterioration-of-the-tecomate-coastal-lagoon-in-tropical-regions.jpg)


















