Embed Size (px)
Citation preview

Cellular lipids and immunity: characterisation of
glycosphingolipids binding the antigen presenting molecule CD1
Karen MuindiOGBI seminar30th May 2007
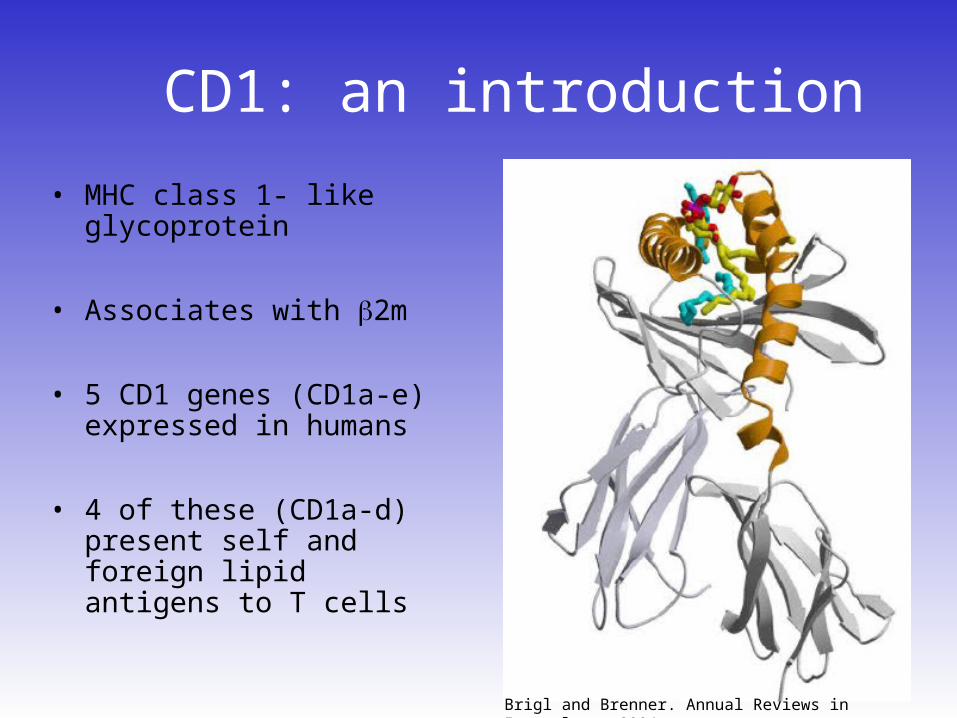
CD1: an introduction
• MHC class 1- like glycoprotein
• Associates with 2m
• 5 CD1 genes (CD1a-e) expressed in humans
• 4 of these (CD1a-d) present self and foreign lipid antigens to T cells
Brigl and Brenner. Annual Reviews in Immunology, 2004

CD1a intracellular trafficking
Adapted from Hava et al. Curr Opin Immunol. 2005

CD1b,c and d intracellular trafficking
Hava et al. Curr Opin Immunol. 2005

CD1 antigens

• CD1 presentation of self GSLs implicated in etiology of several autoimmune diseases
• MS patients have elevated numbers of T cells responding to a variety of GSLs including sulfatide and GM1
• sera from Guillain-Barré patients has elevated levels of Abs reactive to GM1, GA1 and GD1b
• The most potent activators of CD1d-restricted T cells (-galactosylceramide and iGb3) are GSLs
Endogenous glycosphingolipids

• Wt mCD1d or mCD1d-tail deleted (TD) expressing cells assayed for their ability to activate CD1d restricted NKT cells
• CD1d-TD presentation greatly reduced.
• Suggests that lysosomal processing is required for activation of NKT cells
CD1 lysosomal trafficking and T cell activation
Chiu et al. JEM. 1999, 103-10.

2 construct system to resolve CD1 pre- and post- lysosomal GSL load
TEV protease
TEV protease
Secretorypathway only
Secretory and endocytic pathways
Adapted from Brigl and Brenner. Annual Reviews in Immunology, 2004
B2m Extracellular Domainlinker Hinge CH3CH2
IgG2a Fc
B2m Extracellular Domainlinker TM CYTEV
hCD1b-Fc
hCD1b-TEV

hCD1b-Fc is expressed and maintains native conformation
0 250 500 750 1000 1250 15000.00
0.25
0.50
0.75
1.00
1.25
hCD1b-Fc (ng/ml)
OD
@ 4
05n
m0 250 500 750 1000 1250 1500
0.00
0.25
0.50
0.75
1.00
1.25
hCD1b-Fc (ng/ml)
OD
@ 4
05n
m
markers hCD1b-Fc-
175
85
62
47.5
32.5
25
16.5
6.5
hCD1b-Fc (72.7)
KDamarkers hCD1b-Fc-
175
85
62
47.5
32.5
25
16.5
6.5
hCD1b-Fc (72.7)
KDa

TEV protease cleaves hCD1b-TEV: cleaved protein maintains
native conformation
100 101 102 103 104FL1-H
0
20
40
60
80
100
% o
f M
ax
100 101 102 103 104FL1-H
100 101 102 103 104100 101 102 103 104FL1-H
0
20
40
60
80
100
% o
f M
ax
0
20
40
60
80
100
0
20
40
60
80
100
% o
f M
ax
100 101 102 103 104FL1-H
100 101 102 103 104100 101 102 103 104FL1-H
0
20
40
60
80
100
% o
f M
ax
0
20
40
60
80
100
0
20
40
60
80
100
% o
f M
ax
100 101 102 103 104100 101 102 103 104FL1-H
100 101 102 103 104100 101 102 103 104FL1-H
0
20
40
60
80
100
0
20
40
60
80
100
% o
f M
ax
0
20
40
60
80
100
0
20
40
60
80
100
% o
f M
ax
mock wt hCD1b hCD1b-TEV (3:2-2)0
100
200
300
400
500
600
700untreated+ TEV protease
HeLa transfectants
hC
D1b
(n
g/m
l)

Lysosomal trafficking of hCD1b-TEV
wt hCD1b I I L Y W R N P T S I G S I V L A
hCD1b-TEV (2b4) I I L Y W A S G S E N L Y F Q G G S G S R N P T S I G S I V L A
hCD1b-TEV (151-150) I I L Y W G S E N L Y F Q G R N P T S I G S I V L A
hCD1b-TEV (05-150) I I L Y W E N L Y F Q G R N P T S I G S I V L A
hCD1b-tev (3:2-2) I I L Y W E N L Y F Q G S I V L A
hCD1b-a-tev-a (3a:2-2) I I L Y W A E N L Y F Q G A S I V L A
B2m Extracellular Domain linker TM CYTEV
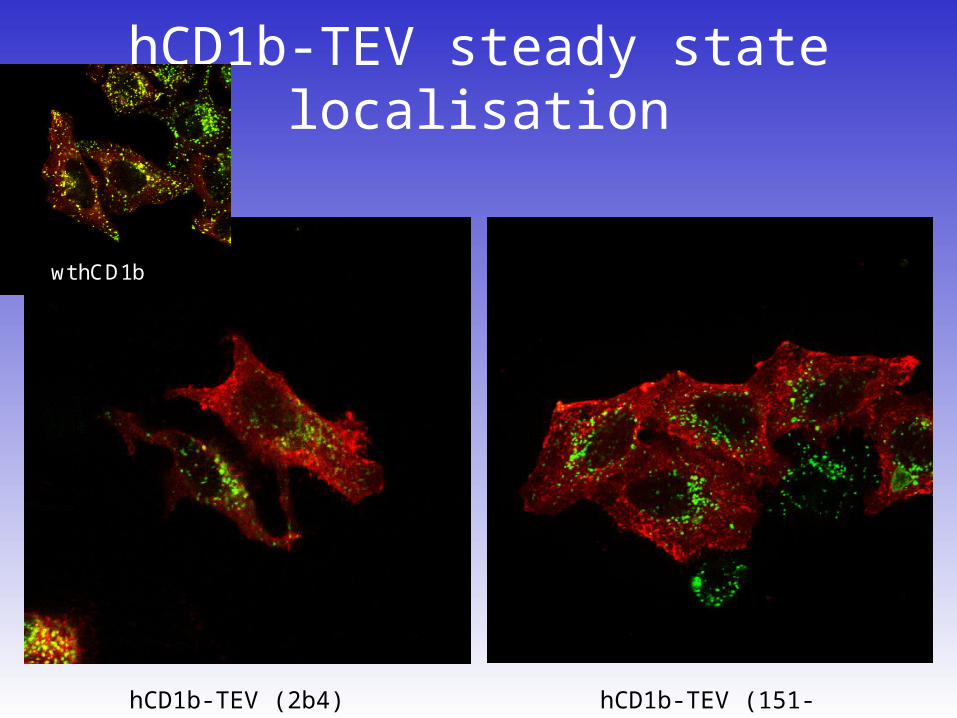
hCD1b-TEV steady state localisation
hCD1b-TEV (151-150)hCD1b-TEV (2b4)
wt hCD1b
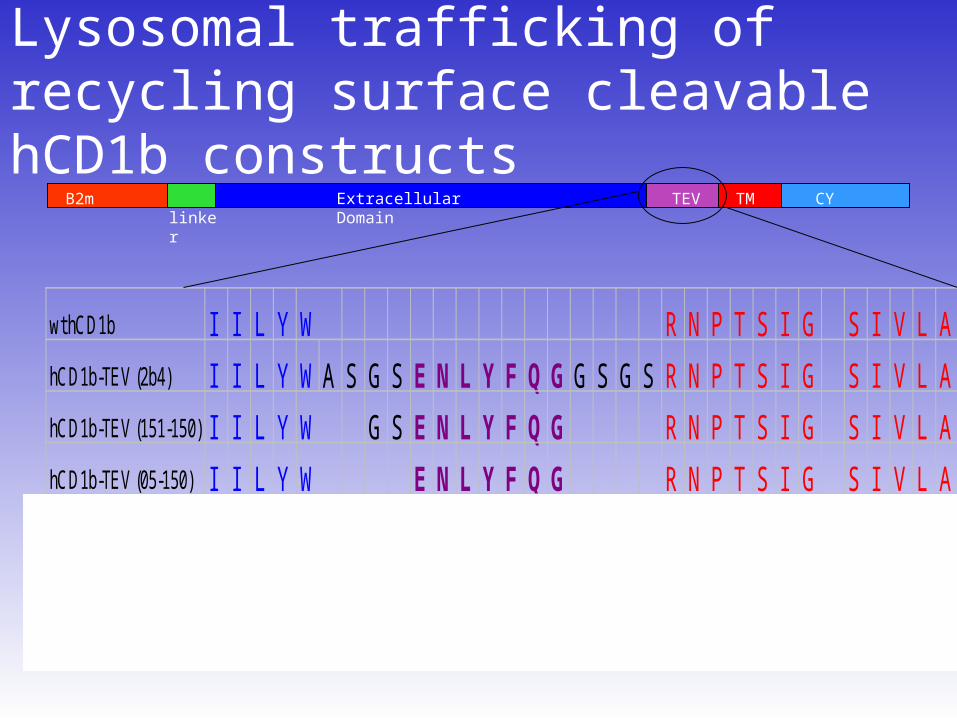
Lysosomal trafficking of recycling surface cleavable hCD1b constructs
wt hCD1b I I L Y W R N P T S I G S I V L A
hCD1b-TEV (2b4) I I L Y W A S G S E N L Y F Q G G S G S R N P T S I G S I V L A
hCD1b-TEV (151-150) I I L Y W G S E N L Y F Q G R N P T S I G S I V L A
hCD1b-TEV (05-150) I I L Y W E N L Y F Q G R N P T S I G S I V L A
hCD1b-tev (3:2-2) I I L Y W E N L Y F Q G S I V L A
hCD1b-a-tev-a (3a:2-2) I I L Y W A E N L Y F Q G A S I V L A
B2m Extracellular Domain linker TM CYTEV

Lysosomal trafficking of recycling surface cleavable hCD1b constructs
wt hCD1b I I L Y W R N P T S I G S I V L A
hCD1b-TEV (2b4) I I L Y W A S G S E N L Y F Q G G S G S R N P T S I G S I V L A
hCD1b-TEV (151-150) I I L Y W G S E N L Y F Q G R N P T S I G S I V L A
hCD1b-TEV (05-150) I I L Y W E N L Y F Q G R N P T S I G S I V L A
hCD1b-tev (3:2-2) I I L Y W E N L Y F Q G S I V L A
hCD1b-a-tev-a (3a:2-2) I I L Y W A E N L Y F Q G A S I V L A
B2m Extracellular Domain linker TM CYTEV

hCD1b-TEV (3:2-2) traffics to the lysosome …
wt CD1b hCD1b-TEV (3:2-2)
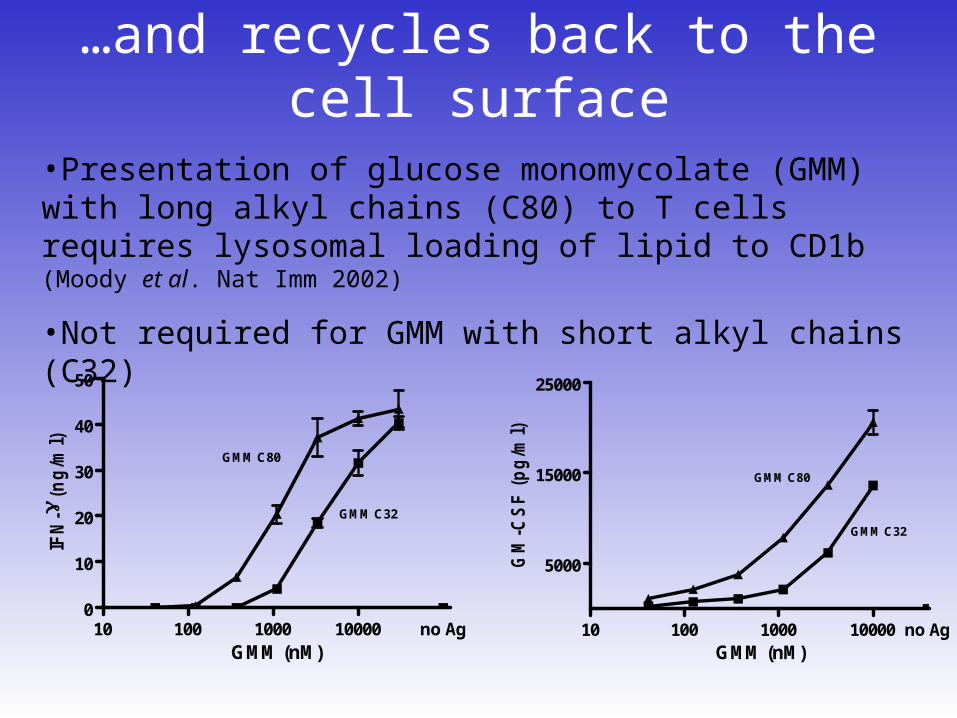
…and recycles back to the cell surface
10 100 1000 10000 no Ag0
10
20
30
40
50
GMM (nM)
IFN
- (
ng
/ml)
GMM C80
GMM C32
10 100 1000 10000 no Ag
5000
15000
25000
GMM (nM)
GM
-CS
F (
pg
/ml)
GMM C80
GMM C32
•Presentation of glucose monomycolate (GMM) with long alkyl chains (C80) to T cells requires lysosomal loading of lipid to CD1b (Moody et al. Nat Imm 2002)
•Not required for GMM with short alkyl chains (C32)

hCD1b-bound lipid extraction and characterisation
hCD1b-Fc harvested from cell culture
supernatant by passing it over protein A
beads
hCD1b-TEV recovered from cell surface using TEV
protease and purified over -CD1b coated beads
lipids extracted from CD1b
glycan headgroups of GSLs were cleaved off using ceramide glycanase
glycans were labelled with 2-aminobenzamide (2-AB), or anthranilic acid (2-AA), and analysed by HPLC.
Species structure was confirmed by exoglycosidase digests
Cleavage of GM1 by ceramide glycanase
Adapted from Expert Reviews in Molecular Medicine © 2002 Cambridge University Press
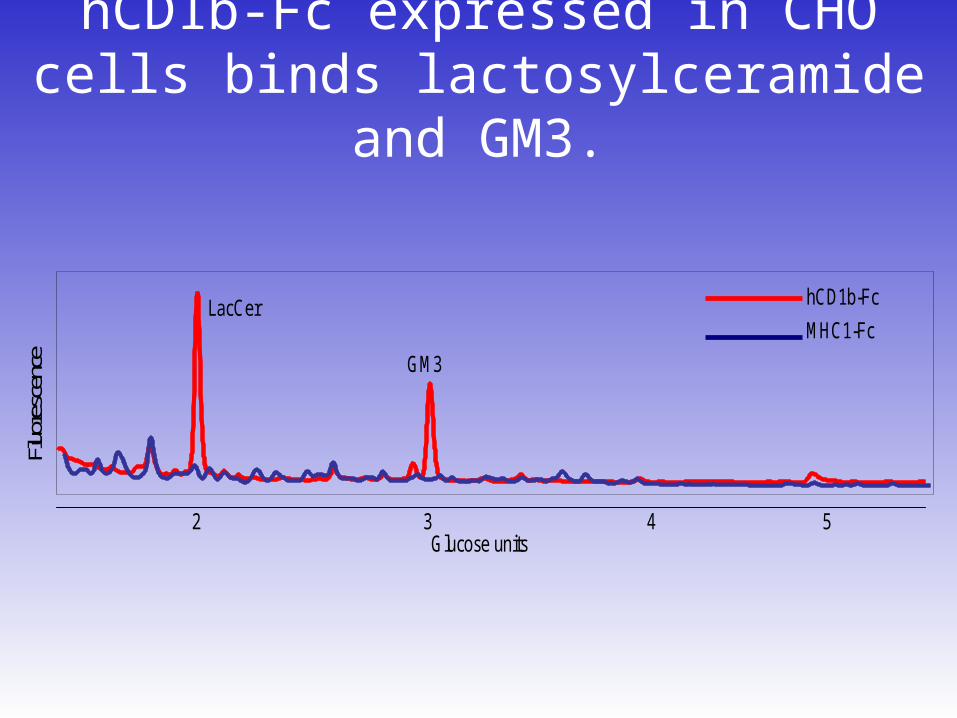
hCD1b-Fc expressed in CHO cells binds lactosylceramide and GM3.
Fluo
resc
ence
42 3 5Glucose units
LacCer
GM3
hCD1b-Fc
MHC1-Fc

hCD1b-TEV expressed in CHO cells binds lactosylceramide and GM3.
LacCer GM3CHO
CHO
hCD1b-TEV
Fluore
scenc
e (Mv
)
0.0
125
220
250
450LacCer GM3
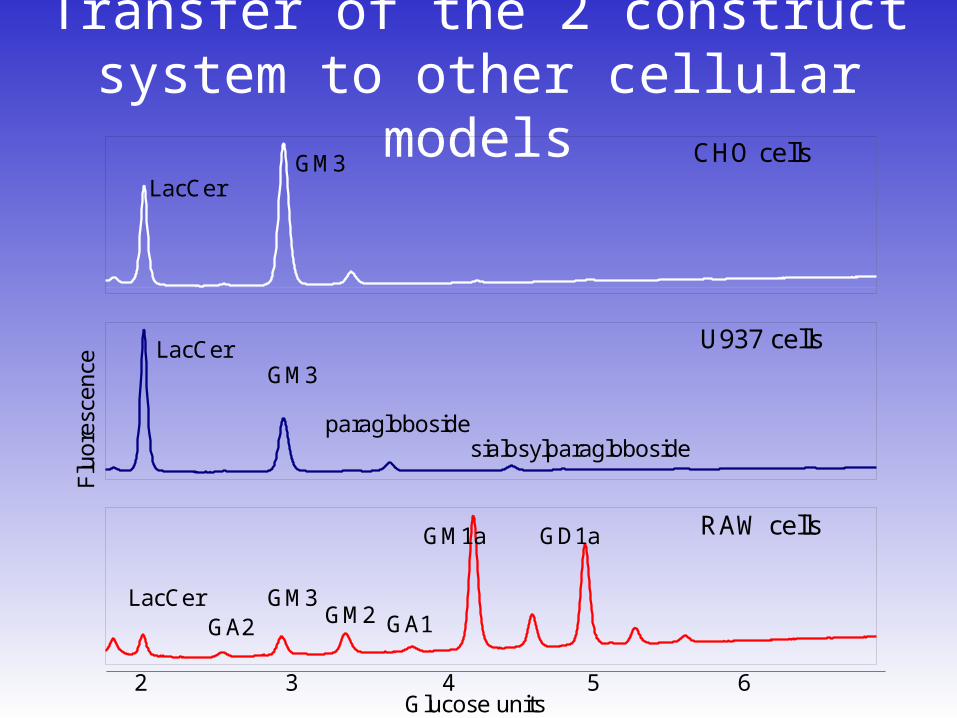
Transfer of the 2 construct system to other cellular models
Flu
ores
cenc
e
CHO cells
RAW cells
U937 cells
LacCer
LacCerGM3
GM3
GM3
GD1aGM1a
LacCer
2 3 4 5 6Glucose units
GA2 GM2 GA1
paraglobosidesialosylparagloboside

Lysosomal processing of GSLs bound to mCD1d expressed in RAW
cellsmCD1d-Fc
MHC1-Fc
mCD1d-TEV
control
GM1a GD1a
GD1a
Flu
ore
scen
ce
A
B
53 4 6Glucose units
GM1a
Neu5Ac2-3Gal1-3GalNAc 1-4Gal 1-4GlcCer 3
Neu5Ac 2
Gal1-3GalNAc 1-4Gal 1-4GlcCer 3
Neu5Ac 2
GD1a GM1a

Conclusions
• We have successfully designed a 2 construct system to study lysosomal processing of CD1 GSL antigens
• Studies of mCD1d in RAW cells suggest that trimming of GSL headgroups from the non reducing end occurs in the lysosome (GD1a to GM1a)

Future Studies
• Using the U937 system, determine whether exchange or processing of hCD1b-bound GSLs occurs in the lysosomes
• Determine which of pre- and post- lysosomal GSL antigens is more antigenic
• Determine whether the GSL tails are modified in the lysosome
• Determine whether activation of APCs results in a change in GSL profile and CD1 GSL antigen load.

Acknowledgements
HMS/BWH (Boston)• Manuela Cernadas• Gerald Watts• Duarte Barral• Michael Brenner
UCD (Ireland)• Louise Royle• Pauline Rudd
OGBI• David Neville• Terry Butters• Raymond Dwek
Work supported by; • NIH grants to Michael
Brenner• OGBI studentship • DPhil scholarship awards
from ORS and Clarendon Fund schemes





























