Embed Size (px)
Citation preview

Naunyn-Sehmiedeberg's Arch. Pharmacol. 289, 99--109 (1975) �9 by Springer-Verlag 1975
Adrenal Cortex Adenylate Cyclase Specific B ind ing Sites for 5 ' -Guany ly l - im idod iphospha t e
in P a r t i a l l y Pur i f ied P l a s m a Membranes f rom Bovine Adrena l Cor tex
H. Glossmann with the technical assistance of K. Klape t t ek
Pharmakologisehes Institut der Justus Liebig Universit~t Giel3en
Received February 6 / Accepted March 18, i975
Summary. Speei fie binding sites for 5'-guanylyl-imidodiphosphate [Gpp(NH)p] have been identified in a partially purified plasma membrane fraction from bovine adrenal cortex. The apparent affinity of Gpp(NH)p at 30~ was 12 txM -1 and the concentration of binding sites was 100 pmoles per mg of protein. Binding of Gpp (NH)p is inhibited by Mn 2+ > Mg 2+ >> Ca 2+ and enhanced by low concentrations of the chelators ethylenediamino-tetraacetic acid (EDTA) and ethylene glycolbis- (fl-aminoethylether)-N,N'-tetraacetie acid (EGTA). High concentrations of EDTA are inhibitory and at 2.5 mM EDTA binding of Gpp(NH)p is only 10% of that observed in the absence of the chelator. The bound labeled GTP analogue ex- changed only slowly with the unlabeled nucleotide after a steady state has been reached. EDTA also releases the bound labeled Gpp(NH)p from its binding sites. The slow dissociation of Gpp(NH)p can explain the persistent activation of adeny- late cyelase observed after pretreatment of bovine adrenal cortex plasma mem- branes with Gpp(NH)io and subsequent washing.
I t is suggested that at least parts of these binding sites are identical to the sites identified earlier as regulatory sites for angiotensin high-affinity receptors (Glossmann et al., 1974a) and for ACTH-stimulated cyclase (Glossmann and Gips, 1974).
Key words: Adenylate Cyclase -- GTP -- 5'-Guanylyl-imidodiphosphate -- Binding Sites.
5 ' -Guanylyl- imidodiphosphate is a GTP analogue which st imulates adenyla te cyclase (ATP-pyrophosphate- lyase , EC 4.6.1.1.) ac t iv i ty in part iculate preparat ions f rom m a n y eukaryot ic cells (Londos et al., 1974; Lefkowitz, 1974). We have recent ly described t h a t Gpp(~rfI)p, like GTP, can dissociate labeled angiotensin I I f rom its high-affinity binding sites on bovine adrenal cortex plasma membranes (Glossmann et al., 1974a). The modula t ion of angiotensin binding was highly specific for GTP and Gpp(NH)p. I T P and much less U T P were also active, bu t ATP, cyclic AMP and 5 'GMP were ineffective to lower the s teady-s ta te binding of
Send o//print requests to: H. Glossmann, Pharmakologisches Institut der gustus Liebig Universitgt, ]:)-6300 Giel3en, Schubertstral3e 1, Federal Republic of Germany.
7*

100 H. Glossmann
angiotensin. Regula tory nucleotide sites for bovine adrenal cortex ACTH-sensit ive adenyla te cyclase in the same membrane prepara t ion have a specificity similar to those sites modula t ing angiotensin binding (Glossmann and Gips, in press). The impor tance of these sites for adrenal cortex adenyla te cyelase is emphasized b y the findings tha t ACTIt (at 0.1 mM ATP) stimulates ra t adrenal adenyla te eyclase only in the pres- ence of Gpp(NH)p to a significant extent, and t h a t the apparen t disso- ciation constant for GTP for ACTH-s t imula ted bovine adrenal cortex adenyla te cyelase is in the same range as for the t rophie hormone A C T H (20 nM) (Glossmann and Gips, in press). To fur ther invest igate the pro- posed nucleotide sites in bovine adrenal cortex plasma membranes we have performed quant i ta t ive binding studies with the labeled GTP analogue. This nueleotide offers the advan tage of no t being hydrolysed to a significant ex tent by nucleotide phosphohydrolases.
Materials 1 and Methods
3H Gpp(IqH)p, specific activity 8.7 Ci/mmole, was from ICN, Irvine, Cal. (U.S.A.). All nucleotides, except GTP and ATP which were from Sigma Chemicals Co. (St. Louis, Missouri, U.S.A.), were obtained from Boehringer, Mannheim. Assays for adenylate cyelase were performed as described (Glossmann and Gips, 1974) and cyclic AMP was determined as reported by Salomon et al. (1974). Binding of aH Gpp(NH)p was measured with the Millipore | filtration technique described earlier in detail (Glossmann and I~eville, 1972). In brief, aliquots of partially purified plasma membranes (prepared as described; Glossmann and Gips, in press) from bovine adrenal cortex were incubated at 30 ~ in 50 mM Tris-HC1, pH 7.4 containing labeled Gpp(NH)p and the listed additions.
After a given time the contents of the glass tube were rapidly diluted with 4 ml ice-cold 10 mM Tris-ItC1 buffer (pit 7.4) and filtered through Millipore (HAWP) filters. One rinse (4 ml) of the incubation vessel was employed to wash the filter. The dried filter was covered with I m] of 1~ (w/v) sodium dodeeyl sulfate. After 2 hrs standing, 10 ml of liquid scintillation mixture (Unisolve | was added. After vigorous shaking, radioactivity of the samples was determined. The 3H counting efficiency was 30% . Blank values (omitting bovine adrenal cortex plasma mem- branes) were determined in the same way. 0.5~ of the total radioactivity present during incubation was trapped by the filters in the absence of bovine adrenal cortex plasma membranes.
Results
Uiotalce and Dissociation o/ Gpp(NH)p
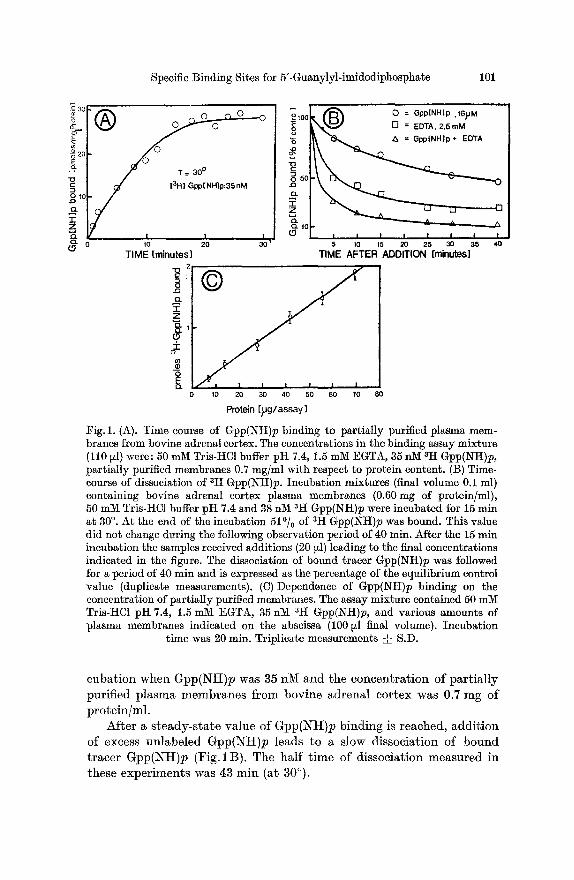
Labeled Gpp(NI t )p was rapidly bound by part ial ly purified plasma membranes f rom bovine adrenal cortex. An example is shown in Fig. 1A where equilibrium was reached a t 30 ~ after approximate ly 15 min in-
z The abbreviations used are: ACTHz_~, corticotropin-(1-- 24), tetraeosapeptide (Synaethen | Ciba-Geigy, Basel); Gpp(l~H)p, 5'-guanylyl-imidodiphosphate; Gpp(CH~)p; 5t-guanylyl-fl, y-methylenediphosphonate; cyclic AMP, adenosine 3 ~ : 5'- monophosphate.
3C o;
~2c
" c
z Q . CI.
Specific Binding Sites for 5'-Guanylyl-imidodiphosphate 10l
[3H] Gpp[NH]p:35 nM
! lO
TIME [minutes] 2
O
"I- 7
r~
o1001 ~ ~ 0 = Gpp[NHlp ,16/uM
| ] L~ I ! ! I I ! I I I
20 30 i 5 10 15 20 25 30 35 40 TIME AFTER ADDITION [minutes]
10 20 30 40 50 60 70 80
Protein [/ug/assay ]
Fig. 1. (A). Time course of Gpp(NH)p binding to partially purified plasma mem- branes from bovine adrenal cortex. The concentrations in the binding assay mixture (110 ~z]) were: 50 mlV[ Tris-HC1 buffer pI-I 7.4, 1.5 mM EGTA, 35 nM atI Gpp(NH)p, partially purified membranes 0.7 mg/ml with respect to protein content. (B) Time- course of dissociation of 3H Gpp(NH)p. Incubation mixtures (final volume 0.1 ml) containing bovine adrenal cortex plasma membranes (0.60 mg of protein/ml), 50 mM Tris-HC1 buffer pH 7.4 and 38 nM ~I-I Gpp(I~H)p were incubated for 15 rain at 30 ~ At the end of the incubation 51~ of 3H Gpp(Ntt)p was bound. This value did not change during the following observation period of 40 rain. After the 15 rain incubation the samples received additions (20 ill) leading to the final concentrations indicated in the figure. The dissociation of bound tracer Gpp(NH)p was followed for a period of 40 rain and is expressed as the percentage of the equilibrium control value (duplicate measurements). (C) Dependence of Gpp(NH)p binding on the concentration of partially purified membranes. The assay mixture contained 50 mlVf Tris-HC1 pH 7.4, 1.5 mM EGTA, 35 nM ~H Gpp(NI-I)p, and various amounts of plasma membranes indicated on the abscissa (100 ~l final volume). Incubation
time was 20 rain. Triplicafe measurements • S.D.
euba t ion when G p p ( N H ) p was 35 nM and the concen t ra t ion of pa r t i a l l y pur i f ied p l a sma m e m b r a n e s f rom bovine ad re na l cor tex was 0.7 rag of p ro te in /ml .
Af te r a s t e a d y - s t a t e va lue of G p p ( N t t ) p b ind ing is reached, a d d i t i o n of excess un labe led G p p ( N H ) p leads to a slow dissocia t ion of b o u n d t r ace r G p p ( N H ) p (Fig. lB ) . The ha l f t ime of d issocia t ion measu red in these expe r imen t s was 43 rain (at 30~

102 H. Glossmann
Addition of EDTA (final concentration 2.5 mM) leads to a more rapid dissociation of bound labeled Gpp(NH)p from its binding sites.
A combination of EDTA with unlabeled Gpp(NH)p (Fig. i B) releases 90o/0 of bound tracer Gpp(NH)p after 40 min.
As the concentration of partially purified plasma membranes is increased in the assay mixture bound tracer Gpp(NH)p increases at equilibrium. The linear relationship between bound Gpp(NH)p and pro- tein concentration shown in Fig, 1 C is more apparent than real. Higher concentrations of plasma membranes in the assay mixture bind up to 90o/o of tracer Gpp(NH)p and as expected from theory the "bound~free" ratio, but not bound Gpp(NH)p is proportional to the amount of plasma membrane present in the incubation mixture (not shown).
Binding Constants and Speei/icity o/the Gpp(NH)p Binding Sites The binding constants for the Gpp(NH)p binding sites were deter-
mined at equilibrium by lowering the specific activity of SH Gpp(NH)p. Seatehard and Lineweaver-Burk plots of equilibrium binding data were linear and the data could be described by the following equation:
N . K . ] b = I + K . /
where b is the concentration of bound ligand at free ligand concentration/. K, the association constant, and N, the concentration of binding sites,
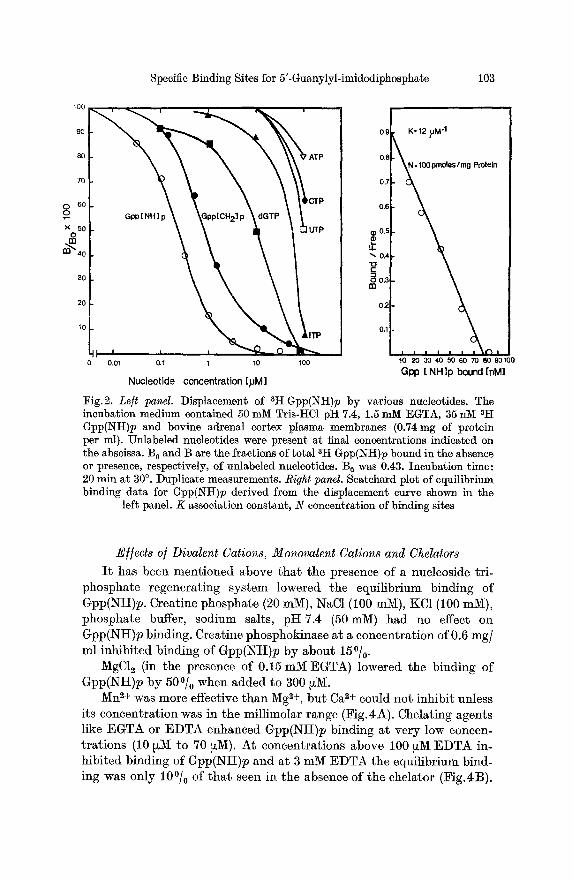
were determined for three different membrane preparations. In the presence of 1.5 mM EGTA K was 12, 11, 13 ~xM -1 and N 100, 89, 108 pmoles per mg of protein. An example is shown in Fig. 2. The specificity of the binding sites was determined by displacement studies using a fixed concentration of tracer Gpp(NH)p and various concentrations of unlabeled nueleotides from 10 nM to 100 ,~M (Fig. 2).
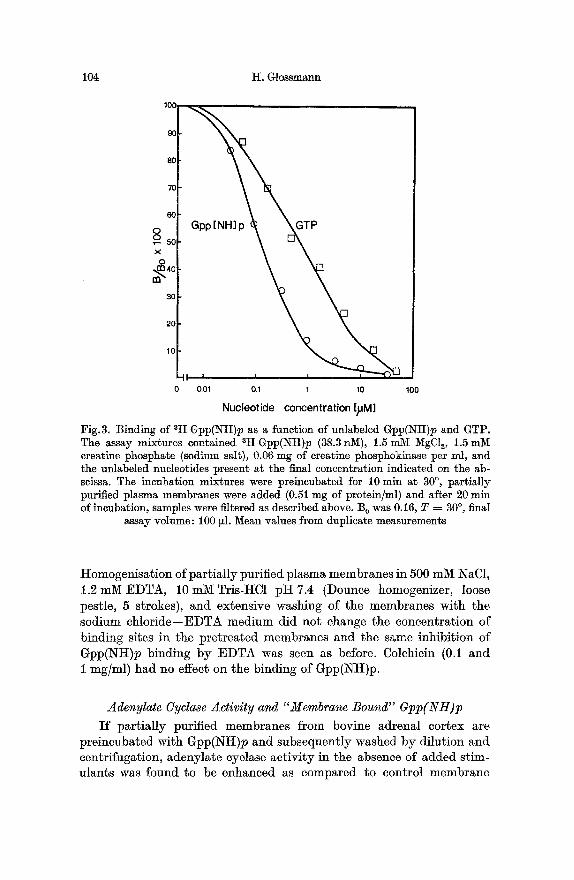
Cyclic AMP, 5'-CMP, 5'-AMP and dTTP did not inhibit binding of Gpp(NH)p even at concentrations of 100 ~M (not shown). Other nueleo- tides had apparen~ affinities to the Gpp(NH)p binding sites as shown in Fig.2. In some experiments a nucleoside triphosphate regenerating system consisting of creatine phosphate, creatine phosphokinase (ATP: ereatine N-phosphotransferase EC 2.7.3.2.) and MgCl~ was included in the binding assay mixture. Under these conditions GTP gained apparent affinity to the Gpp(NH)p binding sites as shown by its ability to inhibit Gpp(Ntt)p binding (Fig. 3).
I t was found that the presence of the regenerating system lowered the equilibrium binding of Gpp(NH)p% The divalent cation Mg 2+ was responsible for this effect, as will be shown below.
The following binding constants were found in the presence of the regenerating system: K = 18,8,9 ~lV[ -x, N = 44,43,48 pmoles per mg of protein (three different membrane preparations).
Specific Binding Sites for 5'-Guanylyl-imidodiphosphate 103
100
90
80
70
X O 50
II~ 40
3O
20
10
Gpp[NH]p
o.ol o.~ 1 ~o loo
Nucleotide concentration [pMl
0,~
O.E
0.'/
O,E
~. 0.4
0.2
0.1
K= 12 ruM-1
t e in
= = m m = i i ~ l t~ l
'iO 20 30 40 50 60 70 80 90100
Gpp [ NHlp bound [nMl
Fig.2. Left panel. Displacement of SH Gpp(NH)p by various nueleotides. The incubation medium contained 50 mM Tris-ttC1 pH 7.4, 1.5 mM EGTA, 35 nlY[ sH Gpp(I~H)p and bovine adrenal cortex plasma membranes (0.74 mg of protein per ml). Unlabeled nueleotides were present at final concentrations indicated on the abscissa. B 0 and B are the fractions of total SH Gpp(NH)p bound in the absence or presence, respectively, of unlabeled nucleotides. B 0 was 0.43. Incubation time: 20 min at 30 ~ Duplicate measurements. Right panel. Scatchard plot of equilibrium binding data for Gpp(Ntt)p derived from the displacement curve shown in the
left panel. K association constant, N concentration of binding sites
E]/ects of Divalent Cations, Monovalent Cations and Chelators I t has been mentioned above that the presence of a nucleoside tri-
phosphate regenerating system lowered the equilibrium binding of Gpp(NH)p. Creatine phosphate (20 raM), NaC1 (100 raM), KCI (100 mM), phosphate buffer, sodium salts, pH 7.4 (50mM) had no effect on Gpp(NH)p binding. Creatine phosphokinase at a concentration of 0.6 rag/ ml inhibited binding of Gpp(NH)p by about 15~
MgC12 (in the presence of 0.15 mM EGTA) lowered the binding of Gpp(57H)p by 500/o when added to 300 ~zM.
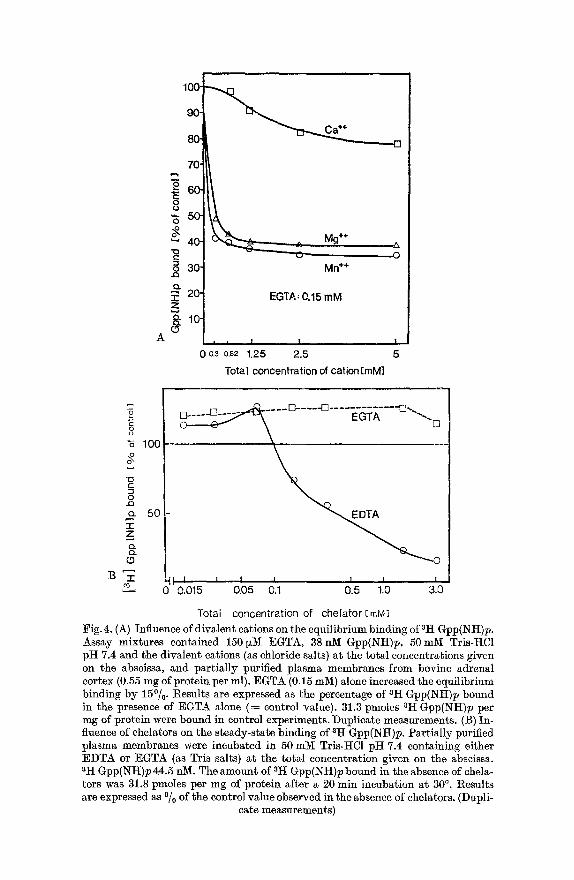
Mn 2+ was more effective than Mg 2+, but Ca ~+ could not inhibit unless its concentration was in the millimolar range (Fig.4A). Chelating agents like EGTA or EDTA enhanced Gpp(NH)p binding at very low concen- trations (10 ~M to 70 ~M). At concentrations above 100 ~M EDTA in- hibited binding of Gpp(NIt)p and at 3 mM EDTA the equilibrium bind- ing was only 10~ of that seen in the absence of the chelator (Fig.4B).
104 H. Glossmann lo0 90
8O
7O
SO
o Gpp[NH]p o 5o x
o
\
20
10
o.m o.~ ~ ~o ~0o
Nucleotide concentration DJM]
Fig.3. Binding of 3H Gpp(NH)p as a function of unlabeled Gpp(NH)p and GTP. The assay mixtures contained 8H Gpp(Ntt)p (38.3 r~), 1.5 mM MgC12, 1.5 mM ereatine phosphate (sodium saIt), 0.06 mg of ereatine phosphokinase per ml, and the unlabeled nucleotides present at the final concentration indicated on the ab- scissa. The incubation mixtures were preincubated for 10 rain at 30 ~ partially purified plasma membranes were added (0.51 mg of protein/ml) and after 20 rain of incubation, samples were filtered as described above. B 0 was 0.16, T = 30 ~ final
assay volume: 100 tB. Mean values from duplicate measurements
Itomogenisation of partially purified plasma membranes in 500 mM NaCI, 1.2 mM EDTA, 10 mM Tris-IIC1 pH 7.4 (Dounce homogenizer, loose pestle, 5 strokes), and extensive washing of the membranes with the sodium chloride--EDTA medium did not change the concentration of binding sites in the pretreated membranes and the same inhibition of Gpp(NH)p binding by EDTA was seen as before. Colehiein (0.1 and 1 mg/ml) had no effect on the binding of Gpp(NtI)p.
Adenylate Cyclase Activity and "Membrane Bound" Gpp(lgH)p I f partially purified membranes from bovine adrenal cortex are
preincubated with Gpp(NH)p and subsequently washed by dilution and centrifugation, adenylate cyclase activity in the absence of added stim- ulants was found r be enhanced as compared to control membrane
A
100-
90-
80-
70- e60.~oo =
-6 50-
40- Mg ++ i .a
" o r -
3 t MR** 20 EGTA: 0.15 mM
I I ! !
0 0.3 0.62 1.25 2.5 Total concentration of cation[mM]
_6, O
-6 EF . . . . . . . . ~7"~--'k EGTA " ' - .
"s 100 . . . .
- 0 c
0 ..Q
50 "r z (D. (3.
B " r I I ~ I I I I ] co 0 0.015 0.05 0.1 0.5 1.0 3.0
Total concentration of chelator EmM] Fig.4. (A) Influence of divalent cations on the equilibrium binding of 3It Gpp(NH)p. Assay mixtures contained 150 ~tM EGTA, 38 nM Gpp(NH)p, 50 talk Tris-HC1 pH 7.4 and the divalent cations (as chloride salts) at the total concentrations given on the abscissa, and partially purified plasma membranes from bovine adrenal cortex (0.55 mg of protein per ml). EGTA (0.15 mM) alone increased the equilibrium binding by 15~ Results are expressed as the percentage of ZH Gpp(Iqtt)p bound in the presence of EGTA alone (= control value). 31.3 pmoles aH Gpp(NH)p per mg of protein were bound in control experiments. Duplicate measurements. (B) In- fluence of chelators on the steady-state binding of 3H Gpp(NH)p. Partially purified plasma membranes were incubated in 50 mY[ Tris-HC1 pH 7.4 containing either EDTA or EGTA (as Tris salts) at the total concentration given on the abscissa. sit Gpp(Ntt)p 44.5 n]VI. The amount of 3It Gpp(NH)p bound in the absence of chela- tors was 31.8 pmoles per mg of protein after a 20 rain incubation at 30 ~ Results are expressed as ~ of the control value observed in the absence of chelators. (Dupli-
cate measurements)

106 H. Glossmann
preparat ions incubated in the absence of Gpp(NH)p (Glossmann and Gips, in press). We were, however, unable to decide if this enhancement of adenyla te cyelase observed in Gpp(NH)p pre t rea ted membranes was due to "bound" Gpp(NH)p or the result of a change of the enzyme or its envi ronment induced by the nucleotide.
We have, however, ruled out t ha t the enhancement of adenyla te cyclase seen after preincubat ion was due to traces of t r apped Gpp(NH)p present in the free form and carried over to the subsequent incubation. Results obtained with 3H Gpp(NH)p presented in Table 1, now suggest t h a t specifically bound Gpp(NH)p is responsible for the act ivat ion of adenylate cyclase seen after preincubat ion of plasma membranes with the GTP analogue.
Table 1. E]ects o] preincubation with Gpp(21H)p on adenylate cyclase activity. Par- tially purified plasma membranes from bovine adrenal cortex were preincubated in the following medium: 0.1 mM ATP, 5 mM MgCl~, 1 ram cyclic AMP, 50 ml~ Tris-HC1 pH 7.4, 2 mM 1-methyl isobutyl xanthine, 2 mlg ereatine phosphate, 0.6 mg creatine phosphokinase per ml. Gpp(NH)p, containing aH Gpp(NH)p, was present during the preineubation period at the final concentration indicated in the table. The concentration of partially purified plasma membranes was 0.4 mg of protein per ml. After 10 rain of preineubation at 30 ~ the preincnbated plasma membranes were pipetted into 40 ml ice-cold 20 m ~ NaHCO a containing 1 mM dithiothreitol and spun at 2~ and 40000• for 40rain. The supernanant was decanted and the walls of the centrifuge tubes wiped free from adhering medium. The membrane pellet was rinsed two times with 5 ml of distilled water, and each time the walls of the tubes were blotted dry. The rinsed pellet was then taken up in 0.6 ml distilled water, homogenized and tested subsequently for adenylate cyclase activity in the absence or presence of ACTHI_2t (0.6 tzg/ml). The incubation medium for adenylate eyclase was identical to the preincubation medium except that 0.1 m ~ [a-a2P]ATP (110 epm/pmole) was used. Aliquots of the resuspended plasma mem- branes were tested for "bound" Gpp(l~H)p by liquid scintillation counting. An aliquot of plasma membranes not preineubated was also tested for adenylate cyclase activity. The following activities were determined (pmoles cyclic AMP/rag protein • min): no addition (basal activity), 90 :k 8; ACTH1-24 (0.6 9g/ml), 447 • 18; Gpp(NH)p (6 ~zNI), 387 4- 27; ACTHI_2t (0.6 ~g/ml) -k Gpp(NH)p (6 ~zM), 900 4- 72.
Each value is the mean q- 1 S.D. from triplicate determinations
Gpp(NH)p (gM) Gpp(NH)p bound (pmoles/mg)
Adenylate cyelasc activity a
without ACTH with ACTH
- - -- 1184- 6 581::k16 112 63.74-13 4074-22 8224-60 56 61 4- 9 345-4-15 6854-34 11.2 54 4-12 3264-39 6854-49 5.6 42.74- 6 3624-19 6854-22 0.56 36.14- 8 2894-17 6534-30
a pmoles cyclic AMP per mg of protein • 15 rain.

Specific Binding Sites for 5'-Gu~nylyl-imidodiphosph~te 107
Discussion
Binding sites for the GTP analogue Gpp(NH)p have been demonstra- ted in partially purified membranes from bovine adrenal cortex. The affinity constant for Gpp(NH)r was 12 ~zM -1 (at 30~ and is close to the values determined by indirect methods in experiments with angiotensin binding (6 ~zM -1, Glossmann et al., 1974a) and stimulation of adenylate eyclase in the same membrane preparation (2 [zM-1; Glossmann and Gips, 1974).
The Gpp(NH)p binding sites could be saturated with respect to the nucleotide. The concentration of binding sites was 100 pmoles per mg of protein. This number exceeds that of high-affinity angiotensin receptors in partially purified membranes from bovine adrenal cortex by two orders of magnitude (Glossmann et al., 1974b). Contrary to our expectations, equilibrium binding data with Gpp(NIt)p did not reveal a marked hetero- geneity of the binding sites. GTP and possibly Gpp(NH)p as well interact with a variety of mitoehondrial enzymes, eofae~ors in protein biosynthesis and other subcellular structures. Probably these binding sites are con- nected neither to adenylate cyclase nor to angiotensin receptors. If their affinity is close to that of the sites responsible for modulation of angio- tensin binding and adenylate cyelase activation, a discrimination based on equilibrium binding data alone would be extremely difficult. Two or more sets of binding sites can be diffentiated from binding curves only in favourable cases (Glossmann and Neville, 1972).
The specificity of the Gpp(NH)p binding sites was tested by dis- placement studies with other nueleotides. The apparent affinities of other nueleotides were close but not identical to those found for the stim- ulation of ACTIt-sensitive adenylate cyclase and angiotensin binding. GTP was as potent as Gpp(NH)p to inhibit high-affinity angiotensin binding (Glossmann et al., 1974a) whereas GTP enhanced ACTtI-stim- ulated bovine adrenal cortex adenylate eyclase with a lower apparent Km than Gpp(NH)2D did (Glossmann and Gips, in press). GTP, even in the presence of a nueleotide regenerating system, was less potent than Gpp(NtI)p itself to inhibit binding of tracer Gpp(NIt)/) (Fig.3). The effects of the components of the regenerating system on the equilibrium binding of Gpp(NH)p were mostly due to the divalent cation Mg ~+. Mg ~+ and Mn 2+ at very low concentrations could lower the steady state binding of Gpp(NH)p whereas Ca ~+ was ineffective unless present at millimolar concentrations. Analysis of equilibrium binding data demonstrated an apparent reduction of Gpp(NH)p binding sites by Mg 2+ in the presence of a nueleoside triphosphate regenerating system. The affinity of the re- mMning sites was unchanged.
The assumption that divalent cations participate in the binding of the GTP analogue was supported by the finding that the chelators EGTA

108 H. Glossmann
and EDTA at low concentrations increased the binding of Gpp(NH)p; probably by complexing traces of divalent cations contaminating the membrane preparation and the binding assay medium.
The chelator EDTA could inhibit the binding of Gpp(NH)10 when its concentration exceeded 100 [~M. Addition of EDTA also rapidly disso- ciates Gpp(NH)p from its binding sites. EGTA on the other hand in- hibits the binding of Gpp(NH)p only at much higher concentrations. The chelator EDTA has higher affinity for Ca 2+ than EGTA. EGTA has much lower affinity for Mg 2+ than for Ca 2+ (Wolf, 1973). The effects of the chelator EDTA observed at high concentrations (above 100 ~M) could indicate that a tightly membrane-bound divalent cation, perhaps Ca 2+, participates in Gpp(NH)p binding. It is noteworthy to mention here that the effects of Ca 2+ and especially EGTA on ACTH-sensitive adenylate cyclase in adrenal cortex are still obscure.
We have found e.g. that EGTA at very low concentrations (150 [~M) enhances Gpp(NH)p effects on adenylate cyclase in bovine adrenal cortex in the abscence of ACTIt and that Mn 2+ antagonizes the inhibitory effects of Ca 2+ and EGTA on ACTIt-stimulated adenylate cyclase (un- published observations).
A number of observations with ACTH-sensitive adenylate cyclase in bovine adrenal cortex and rat adrenals led us to believe that Gpp(NtI)p dissociates only slowly from the sites responsible for the activation of the enzyme.
Preincubation of bovine adrenal cortex plasma membranes with Gpp(NH)p and subsequent extensive washing of the membranes to remove unbound nucleotide revealed persistent activation of the enzyme (Glossmann and Gips, in press). Based on kinetic data alone it was diL ficult to decide whether the effects of the nucleotide were due to "bound" nucleotide or due to change in the properties of the enzyme or its en- vironment induced by the nucleotide without necessarly being bound.
The experiments described above support the view that "bound" Gpp(NH)p is responsible for the activation of adcnylate cyclase seen after preineubation and washing of the partially purified plasma mem- brane preparation.
After completion of this study a report on the binding of Gpp(NH)p to turkey erythrocyte membranes by Spiegel and Aurbach (1974) has come to our attention. The specificity of the Gpp(NIt)p binding sites on turkey erythrocyte membranes is close to that found by us with partially purified membranes from bovine adrenal cortex. Spiegel and Aurbach (1974) also observed that EDTA inhibited Gpp(NIt)p binding. The con- centration of Gpp(NH)p binding sites in turkey erythrocytes membranes is similar to that found in bovine adrenal cortex plasma membranes. I t is perhaps justified to expect such binding sites ih other plasma membrane

Specific Binding Sites for 5'-Guanylyl-imidodiphosphate 109
preparat ions as well. Since the concent ra t ion of binding sites is more in the range expected for enzymes than for hormone receptors, some of the G p p ( N g ) p binding sites m a y well t u r n out as enzymes possibly contain- ing a t ight ly bound divalent cation.
Acknowledgements. The Deutsche Forschungsgemeinschaft supported this in- vestigation with a grunt.
References Glossmann, H., Baukal, A., Cart, K. J.: Angiotensin I I receptors in bovine adrenal
cortex: modification of angiotensin l I binding by guanyl nucleotides. J. biol. Chem. 249, 664-- 666 (1974 a)
Glossmann, H., Baukal, A., Cart, K. J.: Properties of angiotensin I I receptors in the bovine and rut adrenal cortex. J. biol. Chem. 249, 824--834 (1974b)
Glossmann, H., ~eville, D. M., Jr. : Phlorizin receptors in isolated kidney brush border membranes. J. biol. Chem. 247, 7779--7789 (1972)
Glossmann, H., Gips, It.: Adrenal cortex adenylate cyclase, comparison between the action of GTP and 5'-guanylylimidodiphosphate on the particulate enzyme from bovine adrenal cortex and rat adrenals. Natmyn-Schmiedeberg's Arch. Pharmacol. 286, 239--249 (1974)
Glossmann, tt., Gips, tI.: Bovine adrenal cortex adenylate eyclase: Properties of the particulate enzyme and effects of guanyl nucleotides. Naunyn-Schmiede- berg's Arch. Pharmacol. 2S9, 77--97 (1975)
Lefkowitz, R .J . : Stimulation of catecholamine-sensitive adenylate eyelase by 5'-guanylyl-imidodiphosphate. J. biol. Chem. 249, 6119--6124 (1974)
Londos, C., Salomon, Y., Lin, M. C., Harwood, I. P., Sehramm, M., Wolff, I., Rod- bell, M. : 5'guanylylimidodiphosphate, a potent activator of adenylate cyclase systems in eukaryotic cells. Proc. nat. Acad. Sci. (Wash.) 71, 3087--3090 (1974)
Salomon, Y., Londos, C., Rodbell, M. : A highly sensitive adenylate cyclase assay. Analyr Biochem. 58, 541--548 (1974)
Spiegel, A. M., Aurbach, G. D. : Binding of 5'-guanylylimidodiphosphate to turkey eryChrocyte membranes and effects on fl-adrenergic-aetivated adenylate cyclase. J. biol. Chem. 249, 7630--7636 (1974)
Wolf, H.U.: Divalent metal ion buffers with low ptI sensitivity. Experientia (Basel) 29, 241--249 (1973)









![Index [link.springer.com]978-3-642-38487...Index A ACA. See Adenylyl cyclase acaA. See Adenylate cyclase A ACC. See 1-aminocyclo-propane-1-carboxylic acid Acinetobacter baumannii,](https://img.pdfslide.us/doc/110x75/5b47af067f8b9af5078c45af/index-link-978-3-642-38487index-a-aca-see-adenylyl-cyclase-acaa-see-adenylate.jpg)



















