
Insect Biochem., Vol. 12. No. 6, pp. 663 667. 1982 0020-1790:82'060663-05503.00/0 Printed in Great Britain. Pergamon Press Ltd
2 0 - H Y D R O X Y E C D Y S O N E A N D THE O C C U R R E N C E GIANT H n R N A IN EARLY T H I R D INSTAR
CALLIPHORA VICINA LARVAE
OF
ELI SHAAYA* and L. LEVI~NBOOK'J" Laboratory of Physical Biology, National Institute of Arthritis. Diabetes,
and Digestive and Kidney Diseases, National Institutes of Health, Bethesda. MD 20205, U.S.A.
(Received 24 February 1982)
Abstract The pattern of labelled uridine incorporation into epidermal RNA species of starved, early third instar Calliphora vicina larvae, shortly prior to pupariation, closely resembles that previously described for the corresponding fully grown larvae. The effects of exogenously administered 20- hydroxyecdysone on facultative feeding phase small larvae implicate the naturally occurring small burst of this hormone as being the signal for the cessation of feeding, giant HnRNA synthesis and the larwd commitment to pupariation.
Key Word Index: Starvation, giant HnRNA, pupariation, 20-hydroxyecdysone, Calliphora ~'icina. larva
I N T R O D U C T I O N
ABOUT TWO DAYS before pupariat ion, the epidermal cells of Calliphora vicina larvae commence to synthe- size a new type of RNA, termed giant hetero-disperse nuclear RNA or H n R N A (SHAAYA, 1976a, 1976b). This RNA species is of non-r ibosomal origin and con- sists of rapidly turning-over large molecules of aver- age tool. wt ~15 x 10 ~' (or 75S) having a lifespan of about 30min, H n R N A appears to be a prerequisite for subsequent pupariat ion, and its synthesis is regu- lated (SHAAYA, 1979) by the earlier product ion of a small "'peak" of ecdysteroids (SHAAYA and KARLSON, 1965: KOOLMAN, 1978). Al though the function of HnRNA in insect development is not yet unders tood precisely, there is evidence from other organisms that messenger RNA is transcribed from HnRNA (reviewed by SCHERRER et al., 1979). Further, active 75S RNA genes were recently characterized in the Balbiani rings of Chironomus tentans chromosomes (ANDERSSON et al., 1980), and RYLANDER et al. (1980) demonstra ted that such RNA from two of the Bal- biani rings code for high tool. wt secretory protein.
In a previous paper (SHAAYA and LFVENBOOK, 19821 we showed that young third instar C. vicina larvae in the facultative feeding phase (i.e. larvae which con- t inue development and puparia te when starved), re- spond to exogenous 20-hydroxyecdysone by curtail- ment, and even cessation, of feeding, and by shorten- ing the time to pupariat ion. In the present work we now examine whether such undersized, starved larvae also produce H n R N A before pupariat ion, and if so, how this is influenced by 20-hydroxyecdysone.
MATERIALS A N D M E T H O D S
Methods for rearing C. ticina larvae and maintenance of experimental animals were described previously (SHAAYA
* Present address: The Volcani Center, Institute of Tech- nology, Bet-Dagan, Israel
"t To whom reprint requests should be addressed.
and LI-VENBOOK, 19821. Third stage larvae weighing 22 24 rag, or 24 28 rag, in the facultative feeding phase, were utilized in this study (¢11~ SHAAYA and LEVENBOOK. 1982). To obviate the effects of endogenous ecdysteroids, larvae were loosely ligatured with a fine silk thread applied at the third anterior segment i,e. behind the ring gland, and the needle of a 10 i~1 Hamilton syringe inserted through the ligature from the anterior end. The ligature was then tight- ened after withdrawal of the needle l\311owing injection of either 2/~1 saline to stabilize subsequent incorporation of isotope {SHAAYA and CLEVER, 1972), or 2 #1 20-hydroxyec- dysone dissolved in saline. Only the posterior portion of the larva was used for the ensuing experiments. 20- Hydroxyecdysone (Sigma Chemical Co., St. Louis, Mis- souri. U.S.A.) was dissolved and assayed as described earlier (SHAAYA and Li'_vlynOOK, 1982).
For RNA labeling a I hr long pulse of 5 ,t~Ci of [5-3H] - uridine (New England Nuclear, Boston, Massachusetts, 25 Ci/m-mole) in 2/tl saline solution was administered with a fine glass needle through the abdominal body wall (LEvENB~×)~:, 1979) at the desired time after the saline or hormone injection. Epidermal RNA was obtained from lar- val body walls, which were cleaned as much as possible of all tissues other than epidermis. RNA was extracted as described (SHAAVA, 1976a) and separated on 10 × 0.6cm gels of 2.2'!,, polyacrylamide according to St;AAYA (1976C). On such gels 18S and 28S rRNA, 38S pre-RNA and HnRNA could be displayed simuhaneously. A single '~epi- dermis" from the small larwle yielded about 30 ltg RNA.
RESULTS
Epidermal H n R N A J&mation in ,~tamed. early third instar larvae prior to pupariation
C. t'icina larvae weighing 20 nag or more are in the facultative feeding phase; if starved for 48 hr or longer they refuse to feed if replaced on food, and pupariate some 2-3 days later (SHAAYA and LfiVENBOOK, 1982). To assess the epidermal RNA pattern of such larvae irrevocably commit ted to pupariat ion, 24~28 mg lar- vae (weighing about one-third their fully grown weight of ~ 7 6 m g ) were starved for 0, 24, 48 and 72 hr, ligated, injected with saline and 3 hr later they
663
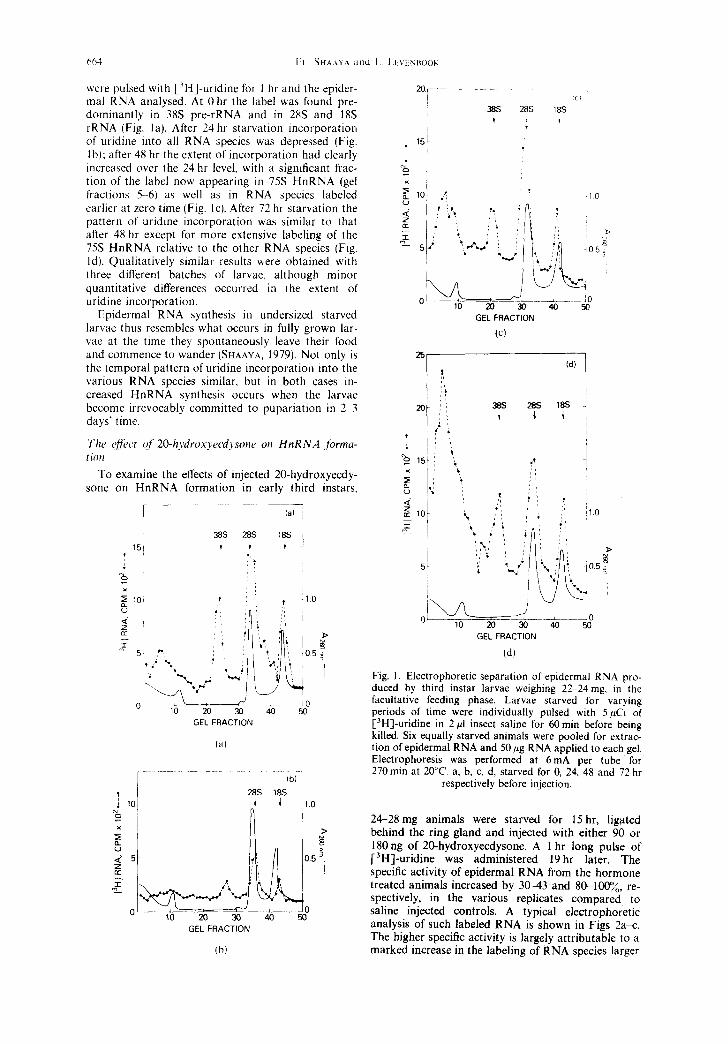
664 [~] SHA.kY,~. ztlld L. LI'~INIIOOK
were pulsed with [3H]-uridine tbr 1 hr and the epider- mal RNA analysed. At 0 hr the label was found pre- dominantly in 38S pre-rRNA and in 28S and 18S rRNA (Fig. lal. After 24 hr starvation incorporation of uridine into all RNA species was depressed (Fig. lb); after 48 hr the extent of incorporation had clearly increased over the 24 hr level, with a significant frac- tion of the label now appearing in 75S HnRNA (gel fractions 5 6) as well as in RNA species labeled earlier at zero time (Fig. lc). After 72 hr starvation the pattern of uridine incorporation was similar to that after 48 hr except for more extensive labeling of the 75S HnRNA relative to the other RNA species (Fig. ld). Qualitatively similar results were obtained with three different batches of larvae, although minor quantitative differences occurred in the extent of uridine incorporation.
Epidermal RNA synthesis in undersized starved larvae thus resembles what occurs in fully grown lar- vae at the time they spontaneously leave their food and commence to wander (SHAAYA, 1979)• Not only is the temporal pattern of uridine incorporation into the various RNA species similar, but in both cases in- creased HnRNA synthesis occurs when the larvae become irrevocably committed to pupariation in 2 3 days' time.
The ~,ffi, ct of 20-hydroxyecdysone on HnRNA forma- tion
To examine the effects of injected 20-hydroxyecdy- sone on HnRNA formation in early third instars,
38S 28S 18S 15r i i
t
% ×
10~ ,{ , !1.0
u I "~,T, @!
10 20 30 40 50 GEL FRACTION
(a)
I 10
×
¢.9 <" 5 Z iv-
(b) 28S 18S
,t,
10
1,0 !
0.5 < 20 30 40 50
GEL FRACTION
(b)
15
% x 10 0
20 m . . . . . . . . . . . . I tc~ i
38S 28S 18S , ,
!10
i i i ,: s j/ ~' "%" 5
° ~ 0 2b 3o .o ~0 GEL FRACTION
(c)
i (d) l
I I 38S 28S 18S
15 .I
10 ~ f" : ' "' E ~ ; i : f"
V" i; io. (
10 20 30 40 50 GEL FRACTION
(d)
Fig. I. Electrophoretic separation or epidermal RNA pro- duced by third instar larvae weighing 22-24 nag, in the [acultative feeding phase. Larvae starved for varying periods of time were individually pulsed with 5/~Ci of [3H]-uridine in 2 #I insect saline for 60 rain before being killed. Six equally starved animals were pooled for extrac- tion of epidermal RNA and 50 #6 RNA applied to each gel. Electrophoresis was performed at 6mA per tube for 270 rain at 20°C. a, b, c, d, starved for 0, 24, 48 and 72 hr
respectively before injection.
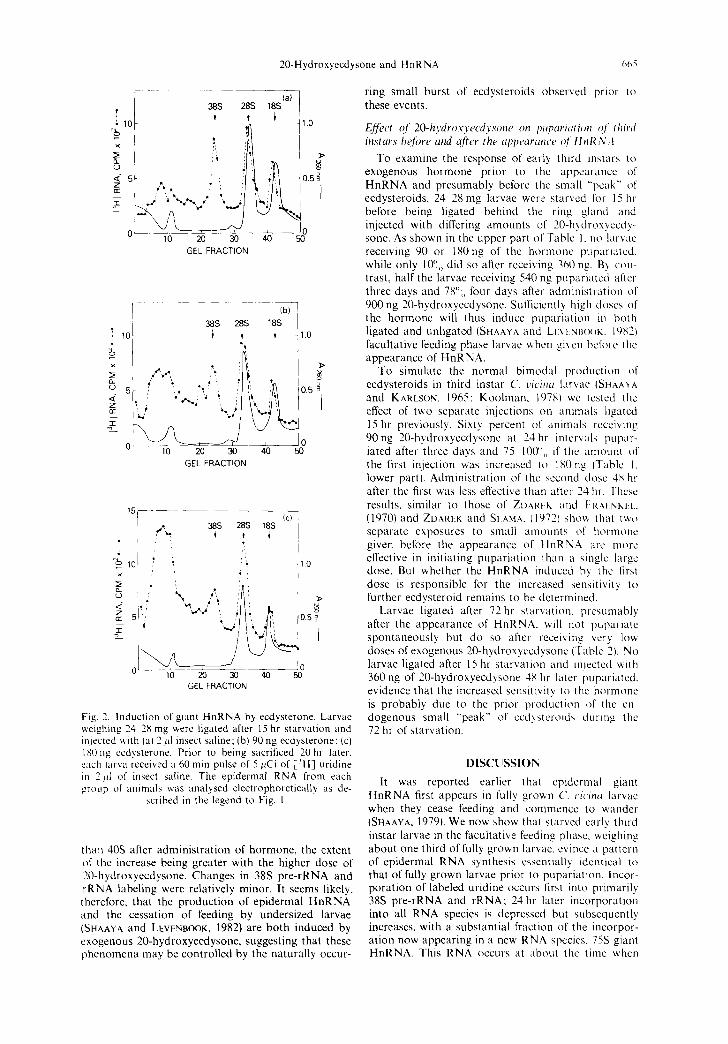
24-28 mg animals were starved for 15 hr, ligated behind the ring gland and injected with either 90 or 180ng of 20-hydroxyecdysone. A 1 hr long pulse of [3Hl-uridine was administered 19hr later. The specific activity of epidermal RNA from the hormone treated animals increased by 30-43 and 80--10(Oo, re- spectively, in the various replicates compared to saline injected controls. A typical etectrophoretic analysis of such labeled RNA is shown in Figs 2a-c. The higher specific activity is largely attributable to a marked increase in the labeling of RNA species larger
20-Hydroxyecdysone and HnRNA {m5
I (a) 1 ~, 38S 28S 18S
i ~° x '
+t+ + 5 • : 'i 0.5
r r ' i i I
+ 4 2 "'+" Z - . > 0 ~ ~ a J O 10 20 30 40 50
GEL FRACTION
10
,5 x
5 I 4 Z
38S +
,,' ,w V, ,~ i, + + ,'
10 20 30
(b) 28S 18S
+ 1.0 i't
, ~ los ~ ~,.i' 1+}
40 50 GEL FRACTION
15[
J ~1o i'
I
[--,,,a_ 0 L ~ - - 10
38S 28S +
..J L
20 3~
(c) l 18S~, 1
10 : I
;', ', 05 ~[
~ ] 0 4O 5O
GEL FRACTION
Fig. 2. Induction of giant HnRNA by ecdysterone. Larvae weighing 24 28 mg were ligated after 15 hr starvation and injected with (a) 2 id insect saline: (b) 90 ng ecdysterone: (c) 180 ng ecdysterone. Prior to being sacrificed 20 hr later, each larva received a 60 min pulse of 5/~Ci of [3HI uridine in 21d of insect saline. The epidermal RNA from each group of animals was analysed electrophoretically as de-
scribed in the legend to Fig. I.
than 40S after administration of hormone, the extent of the increase being greater with the higher dose of 20-hydroxyecdysone. Changes in 38S pre-rRNA and rRNA labeling were relatively minor. It seems likely, therefore, that the production of epidermal HnRNA and the cessation of feeding by undersized larvae (SHAAYA and LEVENBOOK,, 1982) are both induced by exogenous 20-hydroxyecdysone, suggesting that these phenomena may be controlled by the naturally occur-
ring small burst of ecdysteroids observed prior to these events.
Effect of 20-hydroxyecdysone on pupariation ql third instars b¢~fi?re and qlter the appearam'e o1 HnRN A
To examine the response of earl~ third instars to exogenous hormone prior to the appearance of HnRNA and presumably before the small "+peak" of ecdysteroids, 24 28 mg larvae were starved for 15 hr before being ligated behind the ring ghmd and injected with differing amounts of 20-hydroxyecdy- sone. As shown in the upper part of Table 1. no larvae receiving 90 or 180 ng of the hormone pupariated, while only 100,, did so after receiving 360 ng. By con- trast, half the larvae receiving 540 ng pupariated after three days and 78% four days after admmistration of 900 ng 20-hydroxyecdysone. Sufficiently high doses of the hormone will thus induce pupariatiot~ in both ligated and unligated (SHAAYA and kl','t xu{uJK. 19821 facultative feeding phase larvae when given before the appearance of HnRNA.
To simulate the normal bimodal production of ecdysteroids in third instar C. vicim; larwte ( S H A A Y A
and KARLSON, 1965; Koolman, 1978) we tested thc effect of two separate injections on animals ligatcd 15 hr previously. Sixty percent of animals receiving 90ng 20-hydroxyecdysone at 24 hr interwds pupar- iated after three days and 75 100", if the amount of the first injection was increased to 180 ng (Table 1. lower part). Administration of the second dose 4g hr after the first was less effective than after 24 hr. fhese results, similar to those of Zt)A~I{K and FI',AINKIiL. (1970) and ZDARt~K and St.A~4A. (1972) shov, that two separate exposures to small amounts of hormone given before the appearance of HnRNA are tnore effective in initiating pupariation than a single large dose. But whether the HnRNA induced b~ the first dose is responsible for the increased sensitivity to further ecdysteroid remains to be determined.
Larvae ligated after 72 hr starvation, presumably after the appearance of HnRNA. will not pupariate spontaneously but do so after receiving very low doses of exogenous 20-hydroxyecdysone (Table 2). No larvae ligated after 15 hr starvation and rejected with 360 ng of 20-hydroxyecdysone 48 hr later pupariated. evidence that the increased sensitivity to the hormone is probably due to the prior production of the en- dogenous small +'peak" of ecdysteroids during the 72 hr of starvation.
DISCUSSION
It was reported earlier that epidermal giant HnRNA first appears in fully grown C. cicina larvae when they cease feeding and commence to wander ( S H A A Y A , 1979}. We now show that starved carly third instar larvae in the facuhative feeding phase, weighing about one third of fully grown larvae, evince a pattern of epidermal RNA synthesis essentially idetuical to that of fully grown larvae prior to pupariation. Incor- poration of labeled uridine occurs first into primarily 38S pre-rRNA and rRNA: 24 hr later incorporation into all RNA species is depressed but subsequently increases, with a substantial fraction of the incorpor- ation now appearing in a new RNA species, 75S giant HnRNA. This RNA occurs at about the time when

666 kLt SHAAYA and I_.. LEVE~OOK
Table 1. Effect of ecdysterone on induction of pupariation in larvae ligated prior t t~ the appearance of giant HnRNA
First injection Second injection ecdysterone ecdysterone hours alter the
(ng) (ng) first injection
"o Pupariation on indicated day after
last injection Day
I 2 3 4
90 0 180 0 360 - 0 0 10 10 540 0 32 52 900 - 0 30 72 78
0 0 48* 0 90 90 24 0 60 60 60 90 90 48 0 9 15 37
180 90 24 0 90 100 180 90 48 0 75 95 95 180 180 24 0 90 90 90
Third instar larvae weighing 24-28 mg were starved for 15 hr before being ligated and given different amounts of ecdysterone. The upper series was given a single injection, the other series received two injections at 24 or 48 hr intervals. Each test was performed on 12-18 larvae.
* Control larvae injected with saline.
Table 2. Effect of ecdysterone on induction of pupariation in larvae ligated after the appearance of giant HnRNA
Ecdysterone ?~o Pupariation on indicated injected (ng) day after injection
Day l 2 4
0 0 0 0 22 8 8 8 45 12 25 50 90 90 100 -
360* 0 0 0
Third instar larvae weighing 24-28 mg were ligated after 72hr starvation and injected with different amounts of ecdysterone.
*Controls ligated after 15hr starvation and given hormone 48hr later. Each test was performed on 12-18 animals.
starved larvae refuse to feed and, as shown earher (SHAAVA and LEVENBOOK, 1982) are committed to pupariate 2-3 days later. Thus, although there appears to be a close relationship between epidermal HnRNA synthesis and the larval commitment to pupariation, further work is necessary to establish the sequence of events and the precise role of HnRNA.
Our findings indicate that 20-hydroxyecdysone in young third instar larvae increases the incorporation of labeled uridine into HnRNA as compared to other RNA species. To demonstrate specificity it is necess- ary to establish that our observations are not due to variations in precursor pool size, membrane transport or associated phenomena, or are merely the conse- quence of differences in turn-over rates between H n R N A and rRNA. The former has a very short half- life, whereas the latter is much more stable with a half-life in C. vicina of some 2.2 days (PROTZEL et al., 1976). Thus, assuming all RNA species are synthe- sized from the same uridine pool, a 1 hr long pulse of labeled uridine would be expected to label the various
RNA species as a function of their respective turn- over rates, resulting in the observed experimental findings. Fortunately, 38S RNA serves as an "internal standard" in this context; 38s RNA is a rapidly turn- ing-over precursor of ribosomal 28S and 18S rRNA (e.g. GREENBERG, 1969; RUBINSTEIN and CLEVER, 197l), and the labeled rRNA of Figs 1 and 2 is first processed through the 38S species. If the effect of 20-hydroxyecdysone (or of development) on HnRNA synthesis were non-specific, there should be a more or less constant relationship between the labeling of HnRNA and 38S RNA. In fact, 20-hydroxyecdysone scarcely affected the total radioactivity in 38S RNA, whereas the ratio of total counts (corrected for base- line) in HnRNA/38S RNA increased from about 0.5 in control larvae to approx. 2.4 in larvae receiving 90 ng of hormone, to approx. 5 for larvae receiving 180ng. In view of this ten-fold difference, we are led to conclude that 20-hydroxyecdysone specifically aug- ments epidermal HnRNA formation prior to puparia- tion.
Small larvae weighing at least 20 rag, ligated behind the ring gland, can be induced to pupariate by injec- tion of sufficient 20-hydroxyecdysone, the amount required depending upon the time when the ligature was applied. Larvae ligatured before the appearance of HnRNA required, in a single injection, about ten times more hormone than if the ligature was applied after HnRNA formation. But, the high dose could be reduced by some 80~ if two consecutive injections 24 hr apart were administered. It is noteworthy that 20-hydroxyecdysone administered after, in contrast to before, HnRNA formation reduces the time to pupar- iation from two days to one day after injection of the hormone. However, despite an apparent correlation between HnRNA synthesis and increased sensitivity to ecdysteroids inducing pupariation, it is not clear whether such changes in sensitivity are due to HnRNA formation, to other developmental differ- ences, or to both of these.
The probable explanation for increased tissue corn-

20-Hydroxyecdysone and HnRNA 667
petence to react to exogenous hormone involves the role of the small amount of ecdys tero id- - the small "peak"- - re leased prior to the wandering stage (SHAAYA and KARLSON, 1965; KOOLMAN, 1978). In last stage insect larvae the small "peak" appears to be the signal for several subsequent events such as the in- itiation of wandering (RIDDIFORD, 1980), the cessation of feeding (SHAAYA and LEVENBOOK, 1982), inhibi t ion of fat body protein synthesis (PAu et al., 1979), H n R N A formation (SHAAYA, 1979), and the change in epidermal commitment from larval to pupal cuticular proteins (RIDDIFORD, 1978). Small C. vicina larvae ligatured after the appearance of H n R N A have already produced the small "peak" and are poised to pupariate; it requires only a small dose of exogenous 20-hydroxyecdysone to make them do so. Conversely, larvae ligatured earlier, before the small "peak" or H n R N A synthesis, require either very large doses of hormones or two smaller ones 24 hr apart. The first of the two smaller doses probably simulates the effects of the small "peak".
Our experiments indicate that the sequence of physiological events prior to, and including puparia- tion, are the same in undersized, starved larvae as in fully grown, fed animals. In bo th cases giant H n R N A has an essential function since no larvae which failed to produce this RNA ever pupariated.
REFERENCES
ANDIzRSSON K., BJORKROTH B. and DANEHOLT B. (1980) The in situ structure of the active 75S RNA genes in Balbiani rings of Chironomus tentans. Expl Cell Res. 130, 313- 326.
GREEN13ERG J. R. (1969) Synthesis and properties of riboso- mal RNA in Drosophila melanoqaster. J. molec. Biol. 46, 85 98.
KOOtMAN J. (1978) Ecdysone oxidase in insects. Hoppe- Seyler's Z. physiol. Chem. 359, 1315 1321.
LWNBOOK L. (1979) Hemolymph volume during growth of Calliphora ricina larvae. Ann. ent. Soc. Am. 72, 454-455.
PAU R., LEVENBOOK L. and BAUER A. C. (1979) Inhibitory effect of fl-ecdysone on protein synthesis by blowfly fat body in t, itro. Experientia 35, 1449 1451.
PROTZEL A., SRIDHARA S. and LEVENBOOK L. (1976) Ribo- somal replacement and degradation during metamor-
phosis of the blowfly Calliphora vicina. Insect Biochem. 6, 571-578.
RIDDIrORD L. M. (1978) Ecdysone-induced change in cellu- lar commitment of the epidermis of the tobacco horn- worm Manduca sexta, at the initiation of metamor- phosis. Gen. Comp. Endocrinol. 34, 438-446.
RIDDIFORD L. M. (1980) Interaction of ecdysteroids and juvenile hormone in the regulation of larval growth and metamorphosis of the tobacco hornworm. In Pro qress in Ecdysone Research (Ed. by HOFFMAN J. A.), pp. 409-429. Elsevier/North Holland, Amsterdam.
RUBINSTEIN L. and CLEVER U. (1971) Non-conservative processing of ribosomal RNA in an insect Chronomus tentans. Biochim. Biophys. Acta 246. 517 529.
RYDLANDER L., PIGON A. and EDSTROM J. E. t1980) Sequences translated by Balbiani ring 75S RNA in vitro are present in giant secretory protein from Chironomus tentans. Chromosoma 81, 101 113.
SCHERRER K., IMAIZUMI-SCHERRER M. T., REYNAtJD C. A. and THERWATH A. (1979) On pre-messcnger RNA and transcriptions. Molec. Biol. Rep. 5, 5 28.
SHAAYA E. (1976a) Synthesis and characterization of giant RNA in the epidermal cells of Calliphora erythrocephala. Insect Biochem. 6, 553 559.
SHAAVA E. (1976b) Giant RNA in insects I. Differential changes during Calliphora development and the role of ecdysone. Biochem. Biophys. Acta 447, 395 405.
SHAAVA E. (1976c) Separation of high molecular weight heterodisperse RNA and rRNA by polacrylamide gels. Analyt. Biochem. 75, 325 328.
SHAAYA E. (1979) Synthesis of giant HnRNA in the epider- mal cells of Calliphora and the role of the ring gland. Hoppe-Seyler's Z. physiol. Chem. 360, 445 449.
SHAAYA E. and CLEVER U. (19721 In rivo effects of 7-ama- natin on RNA synthesis in Calliphora erythrocephala. Biochim. Biophys. Acta 272, 373 381.
SHAAYA E. and KARLSON P. (1965) Der Ecdysontiter wah- rend der lnsektenentwicklung II. Die postembryonale Entwickhmg der Schmeissfliege Calliphora erythroee- phala Meig. J. Insect. Physiol. i l , 65 69.
SHAAVA E. and LFVENBOOK L. (1982) The effects of star- vation and ecdysterone on feeding and pupariation of early 3rd instar Calliphora vicinu larvae. J. Insect. Phy- siol. (in pressL
ZDAREK J. and FRAENKt~L G. (1970) Overt and covert effects of endogenous and exogenous ecdysone in pupar- ium formation of flies. Proc. hath. Acad. Sci. U.S.A. 67, 331 337.
ZDAREK J. and SCAMA K. (1972) Supernumerary larval instars in cyclorrhaphous Diptera. Biol. Bull. mar. biol. Lab. Woods Hole 142, 35(~357.
Recommended












![The intrinsically disordered region of GCE protein adopts a ......hydroxyecdysone (20E) and juvenile hormone (JH) [2]. While a high JH titer maintains cell divisions without differentiation](https://img.pdfslide.us/doc/110x75/60ce0d93ffc8442c4a5c8497/the-intrinsically-disordered-region-of-gce-protein-adopts-a-hydroxyecdysone.jpg)



