Embed Size (px)
Citation preview

Vol. 256, No. 20, Issue of October 25, pp. 10293-10305. 1981 Printed m U.S. A.
Primary Structure of 3-Phosphoglycerate Kinase from Horse Muscle Il. AMINO ACID SEQUENCE OF CYANOGEN BROMIDE PEPTIDES CBl-CB4 AND CB6-CB14, SEQUENCE OF
METHIONINE-CONTAINING REGIONS, AND COMPLETE SEQUENCE OF THE ENZYME*
(Received for publication, June 11, 1981)
Martin Merrett$ From the Department of Protein Chemistry, Microbiology Research Division, Wellcome Research Laboratories, Langley Court, Beckenham, Kent BR3 3BS, England
The amino acid sequences of 13 of the 14 cyanogen bromide peptides of horse muscle 3-phosphoglycerate kinase have been determined. These peptides together constitute 75% of the structure of the enzyme.
Except for the smallest peptides, automated sequence analysis of the parent peptide was employed together with the automated or mgnual(5dimethylaminonaph- thalene-1-sulfonyl (dansyl)-Edman method) sequencing of relevant peptides obtained by proteolytic digestion. In the case of the five smallest peptides, all between 6 and 11 residues in length, sequences were derived from analyses of the intact peptides.
CB14a, a peptide produced in low yield during the preparation of the cyanogen bromide fragments, was helpful in the sequence analysis of CB14. This peptide appeared to have arisen as the result of a cyanogen bromide cleavage on the carboxyl side of a trypto- phanyl residue.
The sequences of these 13 cyanogen bromide pep- tides, together with that of CBl, reported previously (Hardy, G. W., Darbre, A., and Merrett, M. (1981) J. Biol. Chem. 256, 10264-10292), have been combined with data obtained from the sequence analysis of tryp- tic peptides derived from the methionine-containing regions of the enzyme to determine the complete se- quence. Isolation of these overlap peptides was facili- tated by the labeling of methionyl residues with iodo[2- %]acetic acid prior to tryptic digestion. Tryptophan- containing tryptic peptides were also isolated, and their sequence analysis enabled the positions of the four tryptophanyl residues to be established. In this way, the problem caused by tryptophan degradation during the preparation of the cyanogen bromide fragments was overcome.
The NH; terminus of CBl, the NH*-terminal cyanogen bromide peptide, was identified as N-acetyl serine by mass spectrometry.
The alignment of the cyanogen bromide fragments of horse muscle 3-phosphoglycerate kinase described here is in complete agreement with that based on X-ray analysis of the enzyme reported previously (Banks, R. D., Blake, C. C. F., Evans, P. R., Haser, R., Rice, D. W., Hardy, G. W., Merrett, M., and Phillips, A. W. (1979) Nature 279, 773-777). The complete structure is com- prised of 416 residues and has a molecular weight of 44,519.
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
t Present address, Department of Protein Biochemistry, Biochem- istry Division, Wellcome Research Laboratories, Langley Court, Beckenham, Kent BR3 3BS, England.
3-Phosphoglycerate kinase (ATP:3-phospho-D-glycerate l- phosphotransferase, EC 2.7.2.3) from horse muscle has been the subject of x-ray crystallographic studies (1,2). The present investigations of the primary structure of this important en- zyme have been undertaken in order to complement the information obtained from x-ray maps and thus provide a detailed atomic model.
In the previous paper (3), the isolation of the 14 cyanogen bromide fragments of horse muscle 3-phosphoglycerate kinase was described, together with the sequence analysis of the largest of these fragments, CBl. In this paper, the sequence analysis of the remaining 13 cyanogen bromide peptides is described.
The results of the sequence analyses of the 14 cyanogen bromide peptides have already been used in combination with a high resolution x-ray map of the enzyme to derive a complete structure that has enabled a possible hinge-bending mecha- nism, essential to the action of the enzyme, to be proposed (4). The accuracy of this alignment of the cyanogen bromide peptides is confiied in the present work by the chemical evidence obtained from the sequence analysis of tryptic pep- tides derived from the methionine-containing regions of the enzyme.
METHODS’. ’
RESULTS
Amino Acid Sequence of CNBr Peptides CBl-CB4 and CBG-CBl4-Details of the sequence analysis of peptides CBl-CB4 and CB6-CB14 are described in the miniprint sup- plement.
Alignment of the CNBr Fragments in the Complete Se- quence -An unambiguous alignment of the CNBr peptides of phosphoglycerate kinase has been ascertained in the following manner.
The blocked NH,-terminal residue of CBI and the absence of homoserine from CB14 enabled these two peptides to be located at the NHz-terminal and COOH-terminal ends of the molecule, respectively. The remaining cyanogen bromide pep-
’ Portions of this paper (including “Methods,” part of “Results,” Figs. 2-4 and 6-9, Tables I-XLI, and additional references) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 81M-1394, cite author, and include a check for $27.00 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
‘The abbreviations used are: CNBr, cyanogen bromide; DMAA, N-dimethylallylamine; Quadrol, N,N,N’,N’.-tetrakis(2-hvdroxvnro- pyl)ethylenediamine; PTH, phenylthiohydantoin; Cm, S-carboxy- methyl; TLC, thin layer chromatography; dansyl, J-dimethylamino- naphthalene-l-sulfonyl.
10293

Primary Structure of 3-Phosphoglycerate Kinase 1
AC-Ser
141 1 4 2 1 4 3 1 4 4 1 4 5 1 4 6 1 4 1 148 140 150 1 5 1 1 5 2 153 154 1 5 5 156 1 5 7 158 150 I f 0 1 F 1 1 6 2 1 6 3 1 6 4 1 6 5 1 6 6 161 l h q 1 6 9 110 111 1 1 2 1 1 7 1 1 4 1 1 5
A l a G1U Pro A l a LVS Ile G l u Thr Phe A m llla Ser l e u Ser Lya Leu Cly ASP .'a1 "yr <??.I AJl l Asp A l a Ihe nly Thr A l a HIP Arq A l a HIS Ser Fer vet "_ ._ 4
'""-3
2 4 6 2 4 1 2 4 R 2 4 9 2 5 0 1 5 1 ? 5 2 7 5 3 2 5 4 2 5 5 2 5 6 257 758 25q ?60 7 6 1 ? 6 2 2 6 3 2 6 4 2 6 5 2 6 6 2 6 1 2 6 8 2 6 9 ?70 2 7 1 7 1 2 2 1 3 2 7 4 1 7 5 2 7 6 2 7 7 2 7 8 ? 1 9 2 8 C
V a l Leu A1811 P m "et Clu Ile 01y Thr Ser Leu Phe Asp 01u CIu Clv A l a Lys Ile Val I.ys AS" ~ e u r e t 8er LYS Ala clu Lvs nsn r l y !'a1 Lys Ile Thr ___I 1- CBll I :
'""~""~~""""' c- M 9 ""-( " "7
asp ~ y s val so= H i s v a l ser Thr o l y ~ l y Gly Ala scr Leu R1u U u Leu Glu ClY LYE V a l b U pro GIY ASP A'n
3 8 6 187 388 3 8 9 1 9 0 1 9 1 392 391 391 395 3 9 6 397 398 399 4 0 0 4 0 1 ((12 403 4 0 4 4 0 5 406 407 4 0 8 4 0 9 411 '12 'I' '" 'I5 'I6
I
FIG. 1. The complete amino acid sequence of 3-phosphoglycerate kinase from horse muscle, showing the positions of the cyanogen bromide fragments, CBl-CBl4, and the methionine overlap peptides, MTl-MTl l , whose sequence analysis results are summarized. -+, residues identified by automated Edman degradation; -, residues identified by the manual dansyl-Edman method.

Primary Structure of 3-Phosphoglycerate Kinase 10295
tides were aligned using the whole or partial sequences of the carboxymethylmethionine-containing tryptic peptides ob- tained from a digest of the whole enzyme as described under “Methods” in the miniprint supplement. The peptides used for the alignment of the cyanogen bromide fragments are shown in the diagram of the final sequence (Fig. 1). Details of the properties and sequence analysis of these peptides are described under “Results” in the miniprint supplement.
The use of trypsin to produce overlap peptides for the alignment of the CNBr fragments suffers from the disadvan- tage of giving only single-residue overlaps with fragments that have NHz-terminal basic residues. In the case of 3-phospho- glycerate kinase, there are three cyanogen bromide peptides of this type, CB2, CB3, and CB7; they did not, however, present a problem of alignment. The sequence analysis of MT2, in which cleavage at the arginine following the first Cm- methionine (position 20) had not occurred, demonstrated quite clearly the alignment of CB1 and CB2. Further evidence was provided from the sequence of MT1, since CB2 is the only cyanogen bromide peptide with an NHn-terminal arginyl res- idue. The sequence of MT2 showed Met28 to be followed by a lysyl residue, but the next peptide could not be aligned un- ambiguously from these data, since both CB3 and CB7 have NH2-terminal lysine. However, the peptides MT3 and MT4 provide clear evidence (minimum of three overlapping resi- dues) for the contiguity of the sequence from CB3 to CB6, and MT5 aligns CB6 with another NH2-terminal lysine pep- tide. In addition, the sequence from CB7 to the COOH ter- minus of the molecule is demonstrated by the overlap peptides MT6-MT11. Thus the true alignment of peptides must be as in Fig. 1, since the only alternative arrangement of the two lysine NHz-terminal cyanogen bromide peptides is unaccept- able, as it would place CB7 after CB2, omitting CB3-CB6 (161 residues) from the molecule. The single lysine overlaps do not exclude the possibility of an alternative sequence were there to exist additional lysine NH2-terminal cyanogen bromide peptides that were not isolated from the original digest. How- ever, the existence of such peptides seems most unlikely, since both major and minor components of the digest have been well resolved. In addition, unless any such putative peptide were derived from the COOH terminus (which is not possible,
since the COOH-terminal CNBr peptide, CB14, has been identified), then additional labeled Cm-methionine-containing peptides would have been expected in the tryptic digest. None was observed.
The only other CNBr peptides with identical NH2 termini are CB4 and CB12. No ambiguity arose here, since the overlap peptide MT3 contained the whole of the CB4 sequence over- lapping CB3, CB4, and CB5, tryptic cleavage at Arg,, being inhibited by the juxtaposed proline. In addition, a clear Ser- Lys overlap of the NHz-terminus of CB12 was obtained from MT9.
Identification of Tryptophanyl Residues-Amino acid analysis of 3-phosphoglycerate kinase indicated the presence of four tryptophanyl residues per enzyme molecule. However, the ultraviolet absorbance of the CNBr fragments suggested that only CB12, CB13, and CB14 contain tryptophan. Of these three, only CB14 gave a value of one tryptophan per molecule on amino acid analysis, the others giving values of 0.5 (CB13) and 0.2 (CB12). In addition, none of these CNBr fragments nor any of the peptides derived from them gave a positive reaction with Ehrlich’s reagent or yielded a tryptophan PTH- derivative from Edman degradation. Thus it appeared that some modikation of tryptophan had occurred in the CNBr fragments. The exact nature of this modification was not ascertained. However, a similar effect has been reported ( 5 ) in the case of muconolactone isomerase from Pseudomonas p u - tida; tryptophanyl residues of this protein were found to have been destroyed following treatment with CNBr.
Positive identification of the four tryptophanyl residues was achieved by the sequence analysis of relevant tryptic peptides. From the tryptic digest of 3-phosphoglycerate kinase, pre- pared for the isolation of methionine-containing peptides (see Fig. 5), three tryptophan-containing peptides were also iso- lated, having been identified by their reaction with Ehrlich’s reagent. One of these, MT12, also contained methionine. The sequence analysis of this peptide enabled the CNBr peptides CB12 and CB13 to be aligned and the first of the tryptophanyl residues in the sequence of the enzyme to be identified at position 310. The second tryptophan-containing tryptic pep- tide (TT1) was found in a pure state as an insoluble residue in the digest. It corresponded to a tryptic peptide expected
3-Phosphoglycerate kinase 1
Carboxymethylation at cysteine .1
C-carboxymethylation at methionine 3.
14
Tryptic digestion
1 - - Insoluble residue TTI 1 0.1 M NH,HCO~ solution
1 1 1 1
1 1 1 1
1 1
1 -1
2-Dimensional electrophoresis 2-Dimensional electrophoresis pH 6.5, 1.8
Chromo-Beads Chromo-Beads
2-Dimensional electrophoresis 2-Dimensional electrophoresis pH 6.5, 1.8 pH 6.5, 1.8
TLC TLC
MT2 MT1 MT6 MT5
MT9 MTl l
pH 6.5, 1.8
MT3 MT4 MT8 MT7 MTlO
FIG. 5. Summary of the isolation of the Cm-methionine-containing tryptic peptides.

10296 Primary Structure of 3-Phosphoglycerate Kinase
from a digest of CB13 but that could not be isolated. Trp335, T r p ~ , and the sequence between them were identified from automated sequence analysis of this peptide. The third tryp- tophan-containing tryptic peptide (TT2) was derived from the same region of the enzyme as CB14-T4 and enabled the putative identification of Trp at position 382 to be confirmed.
DISCUSSION
Results presented in this and the previous paper demon- strate that horse muscle 3-phosphoglycerate kinase consists of 416 amino acid residues corresponding to a molecular weight of 44,519.
The sequence analysis of the CNBr peptides has been used previously to derive a complete structure by combination with x-ray data (4). The complete sequence reported here is based on the alignment of CNBr fragments using sequence data from methionine overlap peptides. The two methods are in complete agreement.
Resolution of the complete structure has enabled the atomic interactions between the enzyme and bound nucleotides to be identified. In addition, it has enabled a probable 3-phospho- glycerate-binding site to be described as well as suggesting a mechanism by which the substrates occupying these sites may be brought into a suitable spatial arrangement to allow the reaction to proceed (4). Crystallographic studies have also demonstrated by means of x-ray difference maps using enzyme crystals grown in the presence of MnATP and MgADP that nucleotide binding occurs on a single site in the COOH-ter- minal domain (4). These agreed with NMR data obtained from studies of lanthanide-ADP and ATP complexes by Tan- swell et al. (6). However, it has not been possible to produce similar data on the 3-phosphoglycerate-binding site, as inter- action of the enzyme crystals with solutions of this substrate is accompanied by extensive conformational changes which obscure the binding site.
The 3-phosphoglycerate-binding site is expected to be a small cavity containing a number of positively charged side chains which would bind the negatively charged substrate. It is likely to be close to some residues that could play a catalytic role, since no charged or hydrophilic groups are seen near the y-phosphoryl group of ATP. Inspection of the model reveals only one possible site. This is situated on the NHZ-terminal domain opposite the nucleotide-binding site of the COOH- terminal domain. Around this small cavity in the NHz-termi- nal domain are located five arginyl, three histidyl, two gluta- myl, and one aspartyl residues. If this site does represent the 3-phosphoglycerate-binding site, then a relative movement of the two domains would be required in order to bring the 3- phosphoglycerate across a 1-nm gap to the ATP y-phosphate and at the same time exclude water from the active site. Relative motion of these two domains does appear possible if the NH2- and COOH-terminal domains were to hinge about the narrow neck region that joins them together. In this way, the two domains of the enzyme may move in a manner very similar to that of the motion of discrete subunits, although they are in fact both different regions of the same polypeptide chain.
Direct evidence for a major conformational change in yeast phosphoglycerate kinase upon binding of ATP and 3-phos- phoglycerate to form a ternary complex has been reported by Pickover et al. ( 7 ) . Low angle x-ray scattering in solution was used to measure the radius of gyration of the enzyme, both in the presence and the absence of substrates. The results sug- gested an 8-14’ rotation of one of the lobes upon formation of the ternary complex. Evidence for a conformational change in yeast phosphoglycerate kinase upon substrate binding has been provided by NMR studies (6) and from experiments with
affinity elution carried out by Scopes and Algar (8). In the latter case, it was shown that reduction in the interaction between the enzyme and carboxymethylcellulose in the pres- ence of substrates was caused a t least in part by a substrate- induced conformational change. The binding of 1,3-diphos- phoglycerate was seen to have the largest effect: this is con- sistent with the proposed hinge-bending model.
Apart from the similarity in shape of phosphoglycerate kinases from horse muscle and yeast (2, 9), x-ray diffraction studies on hexokinase (10, 11) and adenylate kinase (12) have shown that these enzymes also consist of two lobes separated by a deep cleft. The active site of phosphofructokinase has also been shown to consist of a cleft between two folding domains (13). Similarly, pyruvate kinase, although folded into three distinct domains, has been shown to bind phosphoenol- pyruvate in a cleft between two of these domains (14).
Anderson et al. (15), who used computer-generated space- fiiing models of hexokinase, adenylate kinase, and phospho- fructokinase to demonstrate the striking similarity of overall structure of these enzymes, have suggested that substrate- induced closure of a cleft between two lobes is a general feature of kinases. The mechanism of action and specificity of this group of enzymes may depend upon this effect. The exclusion of water from the active site would facilitate the nucleophilic attack of the phosphoryl acceptor on the ATP y-phosphate and at the same time prevent hydrolysis of ATP. Substrate specificity would be expected to preclude induction of the required conformational change were a water molecule to bind in place of the phosphoryl acceptor. Numerous reports have been made of conformational changes accompanying substrate binding in kinases other than phosphoglycerate kinase. These include hexokinase (16), adenylate kinase (17, 18), phosphofructokinase (13), and pyruvate kinase (19).
Observed similarities of secondary structure among kinases (11, 12, 20) appear to be related to nucleotide binding. The order of @-strands in the C-domain (CBADEF) in the horse muscle enzyme exactly parallels that of the NAD-binding domain of the dehydrogenases (21). In the N-domain (CDBAEF), there is one difference, and that relates to the location of strand D. However, the deep cleft between two lobes that appears to be a common feature among the kinases that have so far been studied is not related to any similarities of secondary structure. Also, the tertiary structures of these enzymes are quite diverse. Thus different patterns of folding of the polypeptide chain have given rise to a common feature, consisting of two lobes and a deep cleft, suggesting that this feature is an essential requirement for kinase action.
Acknowledgments-I wish to express my thanks to Drs. A. W. Phillips and D. Stone for their encouragement and helpful discussions, and to Dr. F. Cottee who carried out the mass spectrometry. I also thank M. Clackers, S. J. Paterson, and S. Ruston for their skilled technical assistance, and P. Bugby and D. J. Hambidge for their help in the preparation of the manuscript.
REFERENCES 1. Blake, C. C. F., Evans, P. R., and Scopes, R. K. (1972) Nut. New
2. Blake, C. C. F., and Evans, P. R. (1974) J. Mol. Biol. 84,585-601 3. Hardv. G. W.. Darbre. A,. and Merrett. M. (1981) J. Biol. Chem.
Biol. 235, 195-198
4.
5. 6.
7.
8.
256,’ 10284110292 Banks. R. D.. Blake. C. C. F.. Evans. P. R.. Haser. R.. Rice, D. W..
Hardy, G.’W., Merrett, M . , and Phillips, A. W. (1979) Nature 279, 773-777
Meagher, R. B. (1977) Biochim. Biophys. Acta 494, 33-50 Tanswell, P., Westhead, E. W., and Williams, R. J. P. (1976) Eur.
Pickover, C. A,, McKay, D. B., Engelman, D. M., and Steitz, T.
Scopes, R. K., and Algar, E. (1979) FEBS Lett. 106, 239-242
J. Biochem. 63,249-262
A. (1979) J. Biol. Chem. 254, 11323-11329

Primary Structure of 3-Phosphoglycerate Kinase 10297
9.
10.
11.
12.
13. 14.
15.
Wendell, P. L., Bryant, T. N., and Watson, H. C. (1972) Nut. New
Steitz, T. A., Fletterick, R. J., and Hwang, K. J. (1973) J. Mol.
Fletterick, R. J., Bates, D. J., and Hwang, K. J. (1975) Proc. Natl.
Schulz, G. E., Biedermann, K., Kabsch, W., and Schirmer, R. H.
Evans, P. R., and Hudson, P. J. (1979) Nature 279, 500-504 Stammers, D. K., and Muirhead, H. (1975) J. Mol. Biol. 96,213-
Anderson, C. M., Zucker, F. H., and Steitz, T. A. (1979) Science
Biol. 240, 134-136
Biol. 78,551-561
Acad. Sei. U. S. A. 72,38-42
(1973) J. Mol. Biol. 80,857-864
225
204,375-380
METHODS
16. Bennett, W. S., Jr., and Steitz, T. A. (1978) Proc. Natl. Acad. Sei.
17. Pai, E. F., Sachsenheimer, W., Schirmer, R. H., and Schulz, G. E.
18. Buttlaire, D. H., and Cohn, M. (1974) J. Biol. Chem. 249,5733-
19. Mildvan, A. S., and Cohn, M. (1966) J. Bid. Chem. 241, 1178-
20. Bryant, T. N., Watson, H. C., and Wendell, P. L. (1974) Nature
21. Buehner, M., Ford, G. C., Moras, D., Olsen, K. W., and Rossmann,
Additional references are found below.
U. S. A. 76,4848-4852
(1977) J. Mol. Biol. 114,37-45
5740
1193
247, 14-17
M. G. (1973) Proc. Natl. Acad. Sei. U. S. A. 70, 3052-3054

10298 Primary Structure of 3-Phosphoglycerate Kinase
1 2 3 1 5 6 7 8 9 10 11 1 2 1 3 11 1 5 1 6 1 7 1 8 1 9 2 0 CH CO S e r L e u S e r Ann Lye Leu Thr Leu Asp Lys Leu ASD Val Lys Gly Lys Arq V a l Val HSe
l 3 T I _ _ _ ( &yz V-l'L-, =;' e.2 +<4= kT4bi
I- T h l "I 5 _Th2 ,- + - h g Thf -I e 5 e
2 1 2 2 2 3 24 25 26 27 28 Arq Val Asp Phe Asn V a l P r o HSe
"""" ""4-
CB3 - 29 30 31 32 3 3 34 35 36 37 38 39 4 0 4 1 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 5 8
Lys Ann Asn Gln Ile T h r Ann Ann Gln Arc 11s L y s Ala Ala Val Pro Ser 110 Lya Phe CyPLeu A s p Asn Gly Ala L y s S e r Val Val
T3 ____I T4 "--------I+ T5 - ~ ~ ~ ~ ~ ~ j j j j j 4 j ~ j ~ j j j ~ ~ ~ ~ ~ j j
' + " + - + + T + P ~ + 3 + 4 2 2 4 ' 777777777777777777
+"-+"%-?*+ 59 60
i a u HSe - 7
=T- CB4 -
61 62 63 64 65 66 67 68 69 70 71
++-+++--b+"-+ S e r His Leu Gly Arq P r o A s p V a l G l y P r o HSe
CB6 1 7 6 1 7 7 1 7 8 1 7 9 180 181 1 8 2 1 8 3 1 8 4 1 8 5 1 8 6 1 8 7 188 1 8 9
"+++--+"+" Val Gly Val Asn Leu Pro Gln Lya Ala Gly Gly Phe Leu HSe
-
i - ",TL-
- CB7 190 1 9 1 1 9 2 1 9 3 1 9 4 1 9 5 1 9 6 1 9 7 1 9 8 1 9 9 2 W 2 0 1 2 0 2 2 0 3 2 0 4 2 0 5 2 0 6 2 0 7 2 0 8 2 0 9 2 1 0 2 1 1 2 1 2 2 1 3 2 1 4 2 1 5 2 1 6 2 1 7 2 1 8 2 1 9 LYs LYa Clu Leu Asn Tyr Phe Ala Lys Ala Lcu G1U 8.1 P r o G l u Arq Pro Phe Leu Ala Ile Leu Gly Gly Ala LYn V a l A l a ASP LY. ~ ~ ~ j j j ~ j ~ 4 4 ~ ~ j j ~ ~ ~ ~ ~ ~ j ~ ~ j ~ ~ j 4 ~
L T1 ____(I- T2 P 7 T3 I - T4 _.I 7 9 -T ""
220 221 222 223 224 225 226 110 Gln Lau 11s Asn Ann HSe -+*

Primary Structure of 3-Phosphoglycerate Kinase
- CB8 227 228 229 230 231 232 233 Lsu Asp Lye V a l Asn C l u HSe """7 """
CB9 234 235 236 237 238 239 240 241 242 243 244 245 246 247 240 249 250 110 I h Gly G ly G ly HSe Ala Phe Thr Phe Lsu Lys Val Leu Asn Asn HS.3
- +"++-+++-++
- C B l l 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 Glu Ile Gly The Ser M U Phe ASP Glu Glu Gly Ala Lys I le V a l Ly. Aan l a u HSe +*4+4+*+-++"+"-+
I 7
T1 l = T 2 - ( F y T ! - - - ; I
E 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 ser Lys A l a Glu Lya Asn Gly Val Lys 110 T h r h u P r o Val Anp Phe Val T h r A l a Asp Lys Phe Asp Glu H i 8 A l a Lys Thr G ly C ln ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ j ~ ~ ~ ~ ~ + ~ ~ ~ ~ ~ +""
- T1 - I T3 ""+ "-r + 300 301 302 303 304 305 306 307 308 309 310 311
T h r Val A l a Ser ~ l y Ile P r o Ala G l y T r p nsc 777
++e*-++-++++ I
3 + 4 4 yrn% 9 + + "z -to LYs321
- CB13 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 Gly Leu A s p C y P G l y T h r O l u S e r Ser Lys Lys Tyr Ala Glu Ala V a l Ala Arg A l a Lys Gln 110 V a l T r p Asn Gly Pro V a l Gly V a l ~ ~ ~ ~ j 4 ~ ~ 4 ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~
I T 1 7 'Wl-24- 77 T3 -ST' "" + + "i, * 3 " s1-7 342 343 344 345 346 347 348 349 350 351 352 353 354 355 Phe Glu TrP 0111 A l a . P h e A l a Arg Gly Thr Lys Ala Lnu HSe
c 77777777- ' S l
- CB14 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374 375 376 377 378 379 380 381 382 383 384 385 A s p Glu Val V a l L y s Ala T h r Ser U g G l y C y 8 Ile T h r I l e Ile Gly Gly Gly A s p T h r A l a T h r C y 8 C y 8 Ala L y s T r p A m Thr G lu ~ ~ ~ ~ ~ 4 ~ 7 ~ ~ ~ 4 ~ ~ ~ j ~ j ~ j ~ j ~ ~ ~ j +4+
F T 1 - p T 2 - 1 7
T3 : : + T%
" + m s
386 387 386 389 390 391 392 393 394 395 396 397 398 399 4 0 0 (01 402 403 404 405 406 407 408 409 410 4 1 1 412 413 414 415 416
+3"-++34 +++ A s p Lys Val Ser H i s V a l Sar Thr G ly G ly G ly Ala Sar m u Glu Leu Leu Glu G ly Lys Val Leu Pro Gly V a l Aap Ala Leu Ser A m V a l
~ ~ ; ~ ~ ~ ~ 7 ~ ~ ~ ~ ~ ~ ~ ~ j ~ ~ ~ ~ : ~ ~ ~ ~ ~ ~ 6 - ~ j ~ ~ '
~ ~ ~ ~ j ~ j ~ ~ j ~ ~ ~ 4 ~ ~ ~ ~ ~ ~ ~ j ~ ~ ~ I
= I 7 Sl 4 i s2 l

10300 Primary Structure of 3-Phosphoglycerate Kinase
0.2
Fraction Number (1.7ml) Fraction N u m k r
2
1 2 3 "
I - r
\,\L JL d
10 20 30 40 50 Fraction Number (3 ml)
5 .- 8 " 6 7 -
Fraction Number (6 m l )
20 40 64 80 100 120
T a b l e I
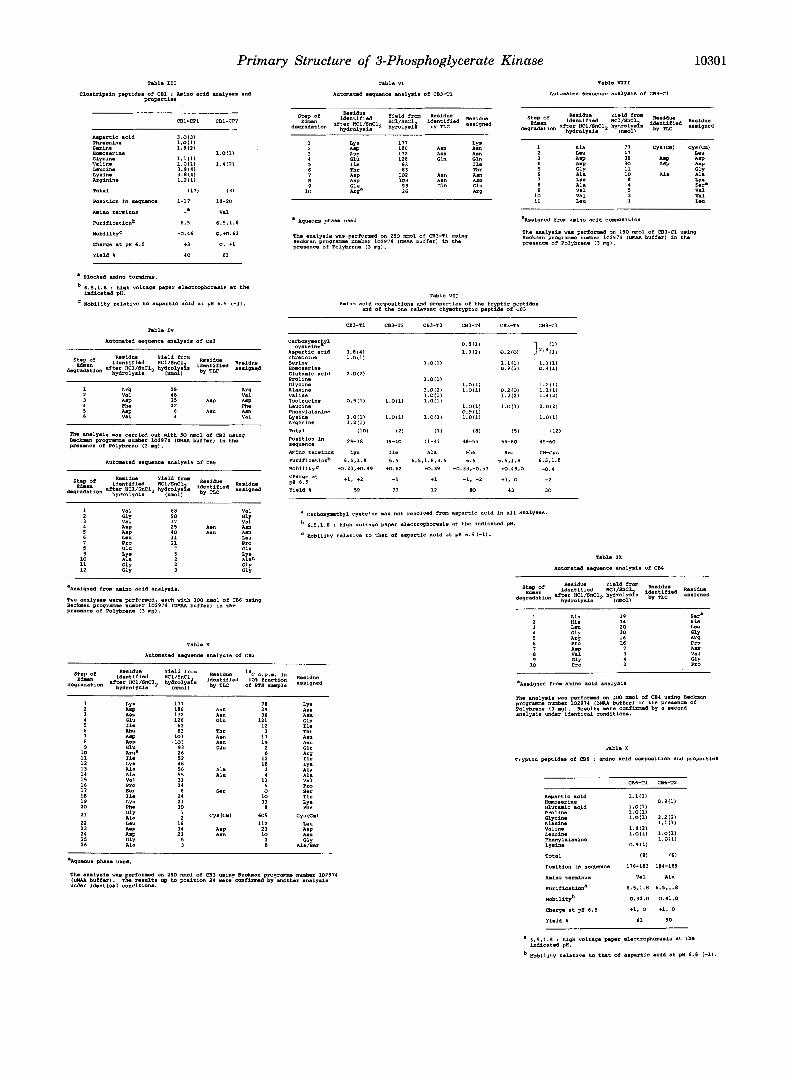
Tryptic peptides of CB1 : m i n o acid C o m p n i t i O n B a n d properties
C81-T1 CBl-TZ CB1-TI C81-TI CB1-TIa CB1-Tkb CB1-TSa CB1-T5aa C i l l
Primary Structure of 3-Phosphoglycerate Kinase 10301
177 186 132 I28
102
63
103
83
9 3 26
A," li." Cln
n.n ?.sn Cl" 8
9 10 11
3.8141 1.0111
25 48 35 12
0.9111
1.0111 1.2111
6 4
1 2 3 4 5 6 7 8 9
10 11 12
68 90
25 37
4 0
11 11
1 5 2 2 3
all a n a l y s e s .
PH. A." I."
T h r *." I.= Gl"
605 CY.IQII 22 23 24 25 26
112 21 le
2 8

10302 Primary Structure of 3-Phosphoglycerate Kinase
1 2 3 k I 6 7 8 9
11 10
12 1 3 1. 15 16 1 7 18 19 20
22 23 2. 25
158 54 73 66 17 53 36 52 5 1 '9 30 23 16
I 10
7 9
Le" PhE
LY. *.X
8
1.0111 4.4141
1.0111
0.9111
2.1121 1.0111
1.0111
1.0111
I31
264-266
11.
W . 5 5
6.5
+I
75
. 6 . 5 6.5 6.5.1.8 6.5.1.8 6 . 5 6.5.1.8 6.5.1.8 6.5.1.8
4 4 3.1 1 2 2 3 2
W.45 10.60 0 0 , W . 5 -0.18 -0.26 0 0
tl +I 0 0 , tl -1.3 -1 0 0
10 30 30 74 2a 68 58 35
1131
251-263
Clu
6.5.1.8
-0.60
-3
55
131
267-269
6.5.1.8
I."
-0.60.0
-1, a
7,
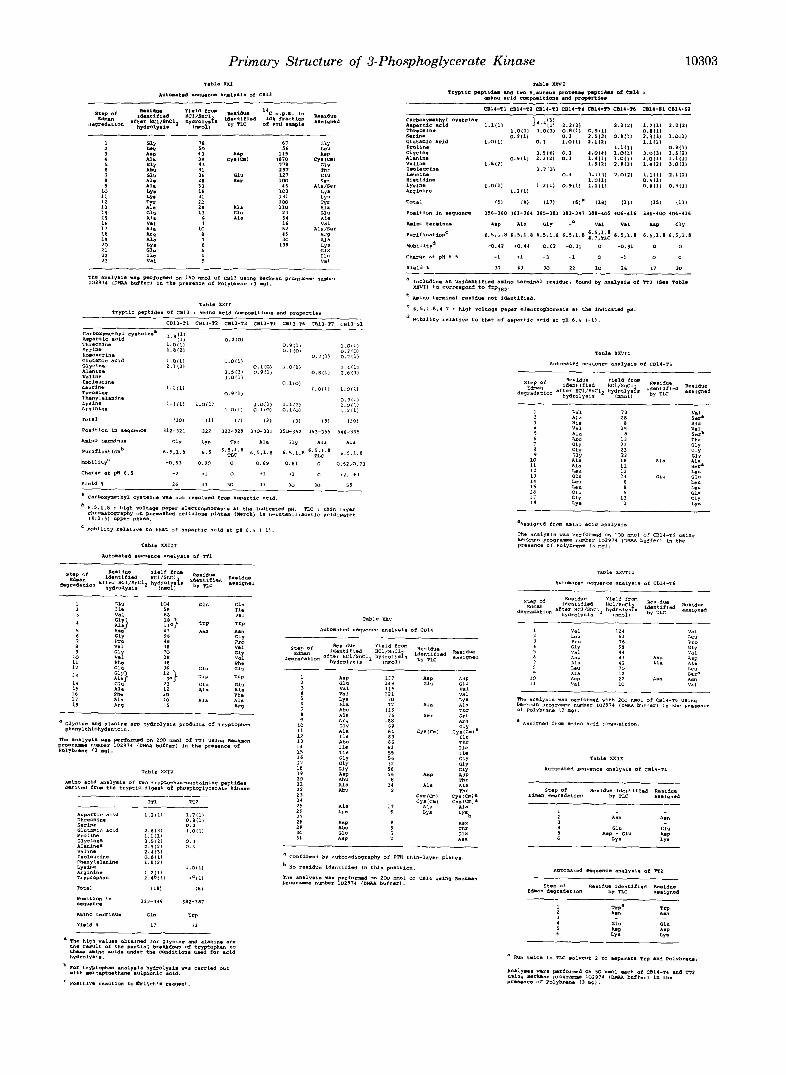
Primary Structure of 3-Phosphoglycerate Kinase 10303
h*P Ai.
lis"

10304 Primary Structure of 3-Phosphoglycerate Kinase
i a 2 0 21 2 2 2 1 2 4 2 5 26 17
Primary Structure of 3-Phosphoglycerate Kinase 10305
I." Cl"