Embed Size (px)
Citation preview

Acquisiaions and Direction des acquisitions et BiMiographicServices Branch des setvices biMiographiques
NOTICE AVIS
Tiie quality of this microform is La qualite de cette microforme heavily dependent upon the depend grandement de la qualit6 quality of the original thesis de ta thhse soumise au submitted for microfilming. microfilmage. Nous avons tout Every effort has been made to fait pour assurer une qualit6 -
ensure the highest quality of sup6rieure de reproduction. reproduction possible.
If pages are missing, contact the S'il manque des pages, veuillez university which granted the communiquer avec I'universite degree. qui a confer6 le grade.
Some pages may have indistinct print especially if the original pages were typed with a poor typewriter ribbon or if the university sent us an inferior photocopy.
Reproduction in full or in part of this microform is governed by the Canadian Copyright Act, R.S.C. 1970, c. C-30, and subsequent amendments.
La qualit6 d'impression de certaines pages peut laisser a desirer, surtout si les pages originales ont ete dactyiographi6es a I'aide d'un ruban us6 ou si I'universite nous a fait parvenir une photocopie de qualit6 inferieure.
La reproduction, mBme partielle, de cette microforme est soumise a la Loi canadienne sur le droit d'auteur, SRC 1970, c. C-30, et ses amendements subsequents.

VISUAL COVER AND SITE SELECTDN BY MULE DEER
Ann H, Rahme
B. Sc. (W.), University of British Columbia, 1985
THESIS SUBMfITED IN PAR1IAL mJLmLLMENT OF
THE REQUlREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
in the Department
of
Biological Sciences
0 Ann H. Rahme 1991
SIMON FRASER UNIVERSITY
NOVEMBER 1991
All rights reserved. This work may not be reproduced in whole or in part, by photocopy
or other means, without permission of the author.

National Lrbrary 1+1 ,,,"a* BiWiheqw nationale du Canada
Acquisitions and Direction des acquisitians et Bibhgraphic Services Branch des sewices bibllographiques
3% Welsngton Sfreel 3%. rue Wethngt~n Omm. Onlam Oaawa (Ontaro) KIA ON4 KIA ON4
The author has granted an irrevocable non-exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or sell copies of his/her thesis by any means and in any form or format, making this thesis available to interested persons.
L'auteur a accorde une licence irrevocable et non exclusive permettant a la Bibliothbque nationale du Canada de reproduire, priiter, distribuer ou vendre des copies de sa these de quelque maniere et sous quelque forme que ce soit pour mettre des exemplaires de cette these 5 la disposition des person nes interessees.
The author retains ownership of L'auteur conserve la propriete du the copyright in his/her thesis. droit d'auteur qui protege sa Neither the thesis nor substantial th8se. Ni la t h k e ni des extraits extracts from it may be printed or substantiels de celfe-ci ne otherwise reproduced without doivent &re imprimes ou &/her permission. autrement reproduits sans son
autorisation.
ISBN 0-315-76231-5

Wame :
Degree :
ANH HgLEH RAHME
Master of science
T i t l e of Thesis:
VISUAL COVER I W D SITE SELECTIONS BY MULE DEER
Examining Committee:
Chairman: Dr. R. C. Brooke
Dr. A.S. hbept*
- ~hacMeton, Associate Professor, imal Sciences, UBC, Vancouver, B.C.
- . D~.'M-L. Winston, Professor, Dept . Biologic,al Sciepces , SF'U
- - Dr. MID. Pitt, Associate Professor, Plant Science, Faculty of Agric, Sciences UBC, Vancouver, B. C, Public Examiner

I hereby grant to Simon Fraser University the rlght to lend
my thesis, proJect or extended essay-(the Xtle of whlch i s shown be low)
to users of the S imoh Frarer Unlversl ty 11 br&y, and to make part la t or
single coples only for such users or In response to a request from t h e
l ibrary of any other university, or other educational Fnstltutlon, on
i t s own behalf or for one of Its users. I further agree that permission for multfpfe copying of thls work for scholarly purposes may be gran+ed
by me or the Dean of Graduate Studles. I t Is understood that.copy1ng
or publIcat1on of this uork for ffnanclal galn shall not be allowed
wlthout my w r l tten perm~ssfon,
Tltfs of Thssls/Project/Extended Essay
Visual Cqver and S i t e Selection b y Mule Deer
Alfihor: . - [signature)
A n n 3. R a h m e
(name 1
N o v . 1 3 , 1991
(date)

iii
M t y cover has 3 functional components: auditory, olfactory, and
visual cover. In this study, I examined visual cover, as weU as crown closure and
other characteristics of sites used by Rocky Mountain mule deer (Odocoileus
Iremionus hemionus). I d e s c r i i the characteristics of visual cover and
evaluated the definition of adequate visual over used in ungulate habitat
studies. Visual cover reqrriremerrts for any species have not been well-defined,
but wildlife biologists assume that visual cover is adequate when an average of
10% of an adult ungulate is visible at a distance of 61 m or less. I found that 61 m
was too long of a sight distance to be used to describe quality of visual cover
available in different habitats, Also, 61 m was significantly longer than the
minimum distance at which 10% of a deer model was visible at sites used by
does. Thermal cover has been assumed to be adequate when crown closure
equals or exceeds 70% and tree height is at least 16 m. The mean crown closure at
sites used by does was 26.8% + 3.0 SE and was significantly less than 70%. The
ecrological validity of these assumptions about visual and thermal cover is
questioned a d discussed.
I measured visual cover w i d a profile board, a profile pole, and a deer
model, evaluated these apparatuses, and used them to desm'be characteristics of
visual cover for different habitats. I propose procedures that best discriminate
differences in visual cover among habitats. These procedures were used to
describe visual cover characteristics of 7 hiibitats. Unspaced sapling forests
provided the densest visual cover while open habitats provided the sparsest
v h d cover. Shrub, spaced sapling, immature forest and mature forest habitats
were intermediate in density of visual cover. The important vegetation
compents of visual aver were: crown closure, percent over of conifer trees

i v
in the top height strata, percent cover of amifas in the shrub layer, total percent
cover of non-conifers in the shrub layer, percent cover of deciduous trees in the
middle height strata, diameter (dbh) of deciduous trees of the middle height
strata, and percent cover of deciduous trees in the bottom height strata. Previous
modeis of visual cover used only characteristics of the coniferous overstory and
ignored the contribution of deciduous and under st or)^ vegetation to visual cover.
My results suggest that these omissions should be rectified.
I compared habitat characteristics at sites used by does to those at sites
randomly located within doe home ranges. I predicted that does used sites with
denser visual cover after parturition (to maximize survival of their fawns and
themselves) than that available at randomly located sites within their home
ranges, or than that at sites used before parturition. I examined visual cover,
crown dosure, and food and water resources at these sites to determine the
relative importance of these resources in site selection. Although climatic
conditions at the time of location were recorded, I was unable to determine
whether does required thermal cover. Availability of forb and minimum
distance to roads of sites used by does did not influence site selection, and there
was no apparent trade-off between either visual cover or crown closure and
percent cover of forbs. Visual cover and distance to water of sites influenced site
selection, but site selection behaviour differed significantly among does.

ACKNOWLEDGEMENTS
Financial support for the thesis project was provided by an NSERC grant
to Dr. A. S. Harestad, B-C. Ministry of Environment (Dave Jones and Bill Harper),
B.C. Ministry of Forests (Brian Nyberg), and B.C. Ministry of Transportation and
Highways (indirectly through Keith Sipson's contract). Personal funding was
provided by my parents, teaching assistantships, S.F.U.'s work study program, a
graduate bursary, stdent. loans, B.C.'s Chdenge program, and Dr. Harestad's
NSERC grant. The opportunity to study mule deer in the Okanagan was
provided by Keith Simpson. Keith also provided unlimited logistic support.
Allison Haney, Maureen Connelly, and Odilia were capable field
assistants, not to mention all my Peachland vistors that I put to work. I thank
Barry and Moira Rondeau for renting their truck to me. f must also thank the
girls for permitting me to follow them although they evaded my predictions:
Gabby, Easy, Brenda, Gimp, Sweetie, Shrimp, Trigger, Rodeo, Cindy, Fart, Myr,
Last, Atf&a, Daze, Wait, and Blaster. Fred Hovey, Alton, and the Midas
manuals guided me through data analyses. Editorial comments were supplied by
Alton, Dr. Mark Winston, Dr. Dave Shackleton, Dr. Mike Pitt, and Rob
Houtman. Moral support was provided by Rob, friends, colleagues, email pals,
r oomtes , and family. Thankfully, Melissa also took 4 years to finish her M.A.
The transitory crew in Peachland was great fun to live and work with:
Keith Simpson, Keith Hebert, Colleen Hodgson, Mike Demarchi, Graeme the
Scot, Jodi and the kids, ZRS Gyug and Sabeena. The folks at Hatheurne Lake and
Tepee Lakes Fishing Resorts provided hot beverages on cold days and an amused
tolerance of us tramping around their resorts. I thank Douglas Lake Ranch and
Brenda Mines for rescuing me that snowy and slippery New Year's Eve.

TABLE OF CONTENTS
. . ........................................................................................................................ Approval .ii
*.. Abstract ....................................................................................................................... .m
Acknowledgements ................... ... ........................................................................... v . . ................................................................................................................. List of Tables v 1 1
... ............................................................................................................. List of Figures v i i i
...................................................................................... 1. General Introduction ..I
.......................................................................................................... Study Area .4
2 Measurement and Characteristics of Visual Cover ................................. ..6
.................................................................................... Methods and Materials .8
Results and Discus6ort ....... ,.. ................................................................... 15 ..................................................................................................... Conclusions .38
3. Influences of Visual Cover and Other Factors on Diurnal Site
Selection by Doe Mule Deer .......................................................................... 41
................................................................................... Methods and Materids 44
............................................................................................................. Results.. .47
...................................................................................................... Discussion 7 4
...................................................................................................... Conclusions 86
4. Conclusions and Management Recommendations ............................... -88
............................................................................................................ Literature Cited 91

UST OF TABLES
Characteristics of 7 habitat t y p s in Thompson Plateau study area 1988-89- For each habitat type, mean percent cover of 3 plant groups, mean percent cover of 3 strata of coniferous and deciduous trees, mean tree diameters (dbh), percent crown dosure and sample sizes are indicated ........................................................ 14
Means of 8 variables that describe deer model visibility in 7 types of habitats ................................................................................................. 30
Independent variables in my multiple linear regression model and their mefkknts when the dependent variables are the visibility scores of the fourth pole section at 5 m and 15 m sequentially ....................................................................................................... 3 6
Mean percent cover of forbs at sites used by does and random sites ...................................................................................................................... 52
ANOVA table of percent cover of forb at sites used before and after parturition ........-......-..t.......-w..-................................................... ............ 53
Mean distances to water (m) from sites used by does and random sites ...................................................................................................... 54
Mean percent visibility of the deer model at coyote height and at 15 m for sites used by does and random sites ....-.................................. 5 7
Mean minimum distance (m) at which only 10% of the deer model can be seen at coyote height for sites used by does and
.......................... random sites .............................. ........................................... 6 0
Three micro-climate parameters estimated for does on the Thompson Plateau ................-.-.. ,.., ............................................................. 67
Mean percent crown closure for sites used by does and random sites ................................................................................................................... 73
Mean distance to the nearest road (m) for sites used by does and random sites .................................................................................................... 7 5

LIST OF FIGURES
Figure p a g ~
Study area on the Thompson Plateau in south-central British Columbia. Radicxallared deer winter in the Okanagan Valley near Peachland but spend spring and summer on the
........................................................................................... Thompson Plateau .5
Relationship between visibility of the profile board and pole at the fourth section (0.75-1.00 m). Visibility scores at 3 sites (n = 48) were measured at my height (1.5 m), 4 distances (5 m, 15 m, minimum distance for 10% deer model visibaty at coyote height and at my height), and 4 directions (updope, downslope, right and left across slope) ........................................................................... 16
Section numbers of the 8 pole sections and proportions of the deer model potentially visr'ble at different 25-cm sections of the
........................ pole are provided. Both apparatuses are drawn to scale 18
Relationship between mean percent of the deer model visible at 15 m at my height (IS m) and that at coyote height (0.75 m) ................ 20
Mean percent visibility of the deer m d d at my height (1.5 m) and 2 distances (5 m and 15 m) and the mean minimum distance for 10% deer model visibility for each habitat ........................tan.. 22
Mean and stambrd error of minimum distances (m) for 10% visibility of the deer model measured at my height (15 m) for 7 types of habitats ..............~...~......~...................................................................... 24
Mean and standard error of differences between habitats in visibility scores of each pole section, pole halves (where B is the bottom 4 sections of the pole and T is the top 4 sections of the pole) and total pole at 15 m (where BT is d l 8 sections of the pole) .............. --.. ........................................................................ ...-.......... -25
Mean and standard error of differences between habitats in visibility scores of the fourth pole section at 15 m for 4 observer positions and for the mean of the visibility scores recorded at
............................................................. the 4 positions ......-,...,,.........,.... ,. 27

i x Mean visibility score of the fourth pole section (0.75-1.00 m) at 15 m for each habitat and time period ...U.............1...od.ododododod.odod.-od.od.od....od..ododod..od.od.od 29
Mean and standard error percent visibility of the deer model measured at 15 m and my height (1-5 m) for 5 successional stages of a sere ................................................................................................... 32
Mean visibility score of each pole section for 7 habitats ......................... 33
Relationship b e e n percent visibility of the deer model measured at 15 rn and coyote height (0.75 m) and percent cover of forb ............................................ ... ................................................................ -49
Relationship between percent mown closure and percent cover of fork. .............................................................................................................. -50
Differences in mean percent visibility of the deer model at 15 m and coyote height (0.75 m) for random sites and sites used by 14 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for random sites is greater than that for sites used by that doe ................................................................................................ ............. 58
Differences in mean minimum distance (m) at which only 10% of the deer model can be seen at coyate height (0.75 m) for random sites and sites used by 14 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for random sites is greater than that for sites used by that doe.. ............................................... 62
Differences in mean minimum distance (m) at which only 10% of the deer model can be seen at coyote height (0.75 m) for random sites and bedding sites used by 11 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for random
.............. sites is greater than that for bedding sites used by that doe..--.- 63
Diffkrences in mean minimum distance (m) at which only 10% ~f the deer m& can be seen at coyote height (0.75 m) for bedding and nun-bedding sites used by 11 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for non- bedding sites is greater than that for bedding sites used by that doe-.- ..-....-...-- ............. ..,..,, ........................................................... .................... 64

X 18 Mean visibility of each pole section at IS m and my height (1.5
m) for doe and random sites .................................................................... 66
19 Frequency distribution for values of wind speeds recorded in 1988 (n = 42) ...................................................................................................... .a
20 Frequency distribution for values of short-wave radiation recorded in 1989 (n = 31) ................................................................................. 69
21 Frequency distributions of air temperature (n = 81) and operative temperature estimates for 1988 (when short-wave radiation was assumed to be U H ) ~ / m 2 ) and for 1989 (when
........................................................ wind speed was assumed to be zero) -70

CHAPTER1
GENERAL INTRODUCTION
All animals require fad , water and potentially cover to sunrive. Of these
requirements, cover is the least understood and only a few studies have
examined cover as a factor in habitat selection. Cover is best defined in the
context of the functiolls it perf-. Animals use cover in several ways:
thermal cover moderates climatic conditions which reduces thennoregulatory
energy requirements, security cover (i.e. hiding cover) reduces the risk of
detection and of attack by predators, escape cover provides a means of escape
from predators once the animal is detected, insect cover provides refuge from
harassing insecis, and snow-interception cover alters the depth and density of the
snowpack. The different functional types of cover are not structurally exclusive
because physical attributes of habitats may perform the different functions (Taber
1961, Peek et aI. 1982). f was able to avoid confounding use of security cover with
use of snow-interception cover by limiting my field season to Iate spring and
summer, but it was nttcessary to address h use of thermal, insect and escape
cover*
Security cave has 3 functional components: auditory, olfactory, and
visual cover- Only the visual tmmponent of security cover has been examined to
date because only visual cover can be measured effectively. Structurally, security
cover consists of vegetation and non-vegetation physical barriers that obstruct
predators' saw~y peK*eptim of prey- The bamers can operate to affect detection
by any of the following interfering with scent-laden air currents (oIfactory
cover), absorbkg or deaecoing sounds made by prey (auditory cover), or
obstnrcting the view of prey by predators (visual cover). The functional
compnenb of security cover may not be structurally exclusive. It is unlikely

that visual cover is physically distinct from olfactory and auditory cover.
Physical barriers that obstruct one of a predator's senses could obstruct other
senses.
The visual cover requirements have not been identified or measured for
any species, but for elk (Cmus elaphus) and mule deer (Odocoileus hemionus)
visual cover is thought to be adequate when an average of 10% of an adult
efk/deer is visible at a distance of 61 m or less (Thomas et al. 1979). Thermal
cover is thought to be adequate when crown closure equals or exceeds 70% and
tree height is at least 16 m in a stand of conifers (Thomas et al. 1986). Although
both of these criteria are currently being used to manage forests for many species
of ungulates, neither has been tested for ecological validity. I examined visual
cover, crown closure, and other characteristics of habitats used by doe mule deer
and their fawns to investigate the characteristics and use of visual cover and to
evaluate the definition of adequate visual cover proposed by Thomas et al.
(1979).
I measured visual cover in several ways. In Chapter 2, I evaluate the
procedures used in measuring visual cover with a profile pole (and profile
board) and a deer model. I propose a set of procedures that best discriminate
among habitats by providing an effective way of detecting differences in visual
cover. I document the phenology of visual cover over late spring and summer,
describe visual cover characteristics for 7 habitat types, and identrfy the
mmponents of vegetation which contribute to visual cover.
I used a natural experiment to determine if doe mule deer use visual
cover (Chapter 3)- I predicted that does will use sites with denser visual cover
after parturition than that available at randomly located sites within their home
range or than that at sites usxi before parturition. Habitat characteristics at sites
used by does were compared to sites randomly located within doe home ranges. I

also compared sites used by does before parturition to those used after
parturition. I examined visual cover, crown closure, food and water resources at
these sites to determine which resources were important in site selection. The
micro-climate conditions at the time of location also were examined to assess the
need of thermal cover. In this way, I attempted to determine which resources
(food, water, visual cover and crown closure) were correlated with sites used by
does in summer.

STUDY AREA
My study was conducted on the Thompson Plateau (Fig. 1) in south-
central British Columbia (500 N, 1200 W). The plateau, with its rolling hills,
varies in elevation between 900 m and 1900 m. The Montane Spruce, Ponderosa
Pine, Engelmann Spruce-Subalpine Fir Biogeoclimatic Zones occur in this
area (B.C. Ministry of Forests 1988). The Montane Spruce Zone is the most
common zone and Engelmann spruce (Picea engelmnnii) and subalpine fir
(Abies lasiocarpa) are its climax conifer species. Because of fires, forest
harvesting and other disturbances much of the forest land in the study area is
not at the climax stage. Currently, the forests are dominated by lodgepole pine
(Pinus contorfa) with Engelmann spruce, subalpine fir, and rarely Douglas fir
(Pseudotsuga menziesii) as minor species. The landscape is interspersed with
lakes and sedge meadows, clear-cuts, roads, hydro-lines, the Brenda Mines open
pit mine, and the right-of-way for the new Okanagan Connector Highway.
The study site was chosen on the basis of the existing facilities. Keystone
Bio-Research was contracted by the B.C. Ministry of Transportatior. and
Highways in cooperation with the B.C. Ministry of Environment to conduct an
inventory of ungulates along the Okanagan Connector Highway. Their study
began in November 1986 and continues into 1992. Keystone Bio-Research had
15-30 adult male and female mule deer fitted with radio-collars. Deer were
trapped with baited Clover traps (Clover 1954) on their winter ranges during 4
consecutive winters (1986-87,1987-88,198&89,1989-90). Besides a radio-colJ.ar,
each deer was fifted with a unique pair of coloued ear tags. The deer use tht?
Thompson Plateau for their spring and summer ranges. This area has an
extensive mad system which facilitates relocation of deer by radio-telemetry.

Fig
ure
1. S
tudy
are
a on
the
Tho
mps
on P
late
au in
sou
th-c
entr
al B
ritis
h C
olum
bia.
Rad
io-c
olla
red
deer
win
ter
in t
he O
kana
gan
Val
ley
near
P
each
land
but
spe
nd s
prin
g an
d su
mm
er o
n th
e T
hom
pson
Pla
teau
(S
imps
on 1
988)
.

CHAPTER2
MEASUREMENT AND CHARACTERISTICS OF VISUAL COVER
Security cover consists of vegetation and non-vegetation physical barriers
that obstrud predators' sensory perception of prey. These barriers can operate to
affect detection by any of the following: interfering with scent-laden air currents
(olfactory cover), absorbing and deflecting sounds made by prey (auditory cover),
and by obstructing the view of prey by predators (visual cover). Although a
number of authors have claimed to have measured security cover (i.e. hiding
cover), they have measured only visual cover, which is the easiest of the 3 to
measure. However, it is probable that structural characteristics which determine
visual cover are the same characteristics which determine olfactory and auditory
cover.
Security cover is typically defined in terms of its visual cover component.
The visual cover requirement has not been determined for any species, but a
common assumption is that visual cover is adequate when an average of 10% of
a standing adult elk is visible at a distance of 61 m or less (Thomas et al. 1979).
This definition is used as a standard criterion for assessing visual cover in forest
stands for ungulates like elk and mule deer, although it remains untested for
ecological validity.
Visual cover has k n measured in a number of ways. Nudds (1977) first
suggested that the density board developed by Wight (1939) (see Gysel and Lyon
1980), and used to measure foliage structure in habitats of birds (MacArthur and
MacArthur 1961, Recher 1%9) and small mammals (Rosenzweig and Winakur
2969, WCloskey and Fieldwick 1973, muld be adapted to quantify vegetation
structure in large mammal habitats. In his critique of the use of the density
board, Nudds (1977) suggested that it is inadequate for measuring vegetation

structure because it does not allow vegetation density to be apportioned to
different heights above ground. Without this capability, density boards cannot be
used to disaiminate among habitats (i.e. detect differences in habitat structure).
Nudds (1977) developed a profile board with which he could describe vegetation
density at different heights and thus quantify habitat structure. Several
researchers have used Nudds' profile board with a variety of procedures for
studies of wildlife habitats (Riley 1982, Canfield et al. 1986, Krasowski and Nudds
1986, Loft et al. 1987, Griffith and Youtie 1988, *heen and Lyon 1989, Yeo and
Peek 1989, MacHutchon and Harestad 1990).
Several researchers also have measured visual cover with models or
targets (Canfield et al. 1986, Smith and Long 1987, Griffith and Youtie 1988).
Smith and Long (1987) used a model that simulated the broadside profile of an
elk. The model was divided into 98 equal-sized squares of 2 alternating colors
(unspecified) and an observer standing 61 m away counted the number of visible
squares to estimate the percent of an elk that would be concealed. Griffith and
Youtie (1988) used models of adult mule deer in bedded and standing positions.
Concealment of deer silhouettes was estimated as the percentage of 0.1-m squares
that were 225% concealed by vegetation. Canfield et al. (1986) used the "Hillis"
method for measuring visual cover in which a target individual moved
randomly along a transect while an observer on the opposite slope recorded the
percentage of 20 observations, taken at 5-sec intervals, that the target was not
visible as a human torso. Usefulness of models and targets is limited because
they are species-s-c and do not standardize descriptions of vegetation
structure. Profile boards, however, are not species-specific, and have broader
utility because measures of vegetation structure and density can be standardized.
While Nudds (19772 made a thorough critique of methods used to
measure visual cover up to 1977, Griffith and Youtie (1988) evaluated the

8
influence of the width of a profile device (board or pole) on estimates of visual
cover, and the repeatability of estimating visual cover by different observers.
They also correlated data collected with profile devices to that collected with a
model of a deer silhouette. My objectives were to confirm the findings of
Griffith and Youtie (1988) in their comparisons of a profile pole with both the
profile board and a deer model. I evaluate procedures used in measuring visual
cover. I propose a set of procedures that best discriminate among habitats by
providing an effective way of characterizing visual cover of different habitat
types. I present phenological changes of visual cover for 4 habitat types from late
spring through summer. I describe visual cover characteristics of 7 habitat types.
I idenhfy attributes of vegetation which contribute to visual cover. I comment
on models that have been developed to predict visual cover in different forest
types and on the use of the standard definition of adequate visual cover in forest-
wildlife management.
METHODS AND MATERIALS
Fifteen different does were radietracked regularly during the study: 11 in
1988 and 12 in 1989. Eight does were radio-tracked during both years. In both
1% and 1989, I began my field work in June and ended it in August. Radio-
tracking was conducted during daylight hours between 0800 and 2000 PST.
Triangulation was initiated from roads to obtain a rough estimate of the doe's
location. Triangulation was then continued on foot until I was very close to the
doe. I approached the does kom a downwind direction and as silently as
possible. Exact locations of does (doe sites) were determined by seeing or hearing
the does or coxifinning their location with physical signs such as backs, feces,
beds or browsed vegetation. Site centres were located in the centre of bedding

9
sites or at the point of greatest site dist.hubance or where the doe was sighted.
Some of the methodology used in the 1988 season was adapted for the 1989
season for increased efficiency and appropriateness. In 1988, random siies were
located by the following method. After I hished radio-tracking the does in
August, I joined ali the hat iom for each doe on an aerial photograph and used
the minimum convex polygon method Wohr 1947) to determine their home
ranges. I then used a grid and randomly chose coordinate pairs to locate random
sites within each home range. I followed this procedure for all does for which I
had at least 3 locations. In 1989, I used a method that allowed me to visit
random sites throughout the season rather than at the end of the season. When
I was at a dm site, I chose a random direction by spinning a pencil. Then, using a
hip chain, I walked 200 m in that direction. AU these random sites should be
within doe home ranges, which are 7 krn2 in summer (Simpson and Gyug 1991).
I measured visual cover by estimating horizontal visibility of a deer
model in 1988 and 1989, and using a vegetation profile board in 1988 and a
vegetation profi!e pole in 1989. Visual cover is an important component of
security cover for the system that I investigated. Coyotes, for which vision is an
effective sense for prey location (Wells and Lehner 19781, are a common predator
of fawns in my study area (Simpson 1988). Whereas other researchers measured
percent concealment of a model or profile device (Mudds 1977, Griffith and
Youtie 1988), I measured percent visibility. These 2 measures are complements
of each other and results are equivalent. Griffith and Youtie (1988)
demonstrated that visibility measures are repeatable because there were no
difftemcs in visibility values recorded by different observers. Nevertheless, I
estimated all deer model and pole visibilities myself to eliminate inter-observer
bias
A fullsized (i.e. the same size as an average doe mule deer, 1 m at the

1 0
shoulder) d e r mode1 was made of 7.5-an tltick foam rubber. The silhouette was
the largest one possible for a doe that is standing broadside with head and ears
facing the observer. The deer model was spray-painted orange and was not
intended to mimic natural colouring. I drew a 20-cm square grid on the deer
model to facilitate my estimation of percent visibility of the whole deer model
(e-g. head and ears = 1.5 squares = 10%). Griffith and Youtie (1988) used a model
of a standing deer silhouette that was the same size but a different shape than
mine; the head was in profile and only upper thighs of legs were represented.
Concealment of their deer model was estimated as the percentage of 0.1-m
squares, painted alternately black and white, that were 225% concealed by
vegetation.
I constructed both a vegetation profile board and pole to measure
vegetation structure and density. The board was made of l a thick plywood
that was 10 cm wide and 2 m high. It was divided into 8 25-cm long sections
which were painted alternately white and fluorescent orange. Nudds (1977) and
Griffith and Youtie (1988) used similar designs for their vegetation profile
boards. Both Nudds (1977) and Griffith and Youtie (1988) used profile boards
that were 30.5 cm wide with alternating 50-an long sections of black and white.
The board used by Nudds (1977) was 0.5 m higher than the 2-m board used by
Griffith and Youtie (1988). In 1989, I made a 2-m vegetation profile pole of 2.5-
an diameter woo& doweling. Like my board, it was divided into 8 25- long
sections that were painted alternately white and orange. Griffith and Youtie
(1988) also used a 2.- diameter pole that was 2 m high, but it was divided into
IO-an long intervals, within 0.5-m long sections (percent concealment was
m a t e d for the 0.5-m long sections using IO-an long intervals), that were
painted alternately black and white. I wed the board in 1988, but I used the less
crumbersome pole for the 1989 field season. To ensure continuity of data, I

11
measured visibility of both the pole and board at 3 sites (4 directiom/site, and 4
distances/direction). Two d the sites were in open habitats, and the other was in
a forested habitat
The model of a deer was placed at site centres and its visibility was
recorded at 4 distance: 5 m, 15 m, the minimum distance at which only 10% of
the deer model can be seen when the observer is at coyote height (0.75 m), and
the minimum distance at which only 10% of the deer model can be seen at my
height (1.5 m). At each site, the deer model was viewed at these 4 distances, in 4
directions (upslope, downslope, and right and left aaoss the slope), and at 2
observer heights. Six of 8 visibility variables for the deer model were measured
as percent visibility: percent visible at 5 m and coyote height, percent visible at 5
m and my height, percent visible at 15 rn and coyote height, percent visible at 15
m and my height, percent visible at coyote height when only 10% is visible at my
height, and percent visible at my height when only 10% is visible at coyote
height. The remaining 2 visibility variables were measured as distances rather
than percent visible: the minimum distance at which only 10% of the deer
model is visible at coyote height, and the minimum distance at which only 10%
of the deer model is visible at my heightt
Pole 6.e. board (1988) and pole (1989)) visibility was measured at 4
distances and 4 directions like deer model visibility, but unlike deer model
visibility, pole visibility was estimated at my height only. The proportion of
each 25-rrm interval visible through the vegetation was recorded as a single digit
visibility score fram 1 to 5 which corresponded to a range in percent visibility
(i-e. 1 = 0 to 20%,2 = 21 to 4U%, and so forth). Pole visibility was measured at 3
distances in 1988 and at 4 distances in 1989. In both years, pole visibility was
estimated at 5 m, 15 m and the minimum distance for 10% visib'ity of the deer
model at my height- In 1989, I also estimated pole visibility at the minimum

1 2
distance for l G % visibility of the deer model at coyote height. For each distance
that pole visibility was measured, there were 11 pole visibility variables: 1 for
each of the 8 intervals on the pale, the sum of visibility scores for the bottom 4
intervals, the sum of the visibility scores for the top 4 intervals and the sum of
visibility scores for all intervals.
A variety of understory Characteristics were assessed at random sites and
sites used by does. Within a 5-m radius of the site centre, ocular estimates of the
percent covers of shrubs and forbs were determined and recorded by species in
1988. In 1989, the percent cover of shrubs was examined by height class (<5 cm,
>5 cm - ~ 0 . 5 m, >0.5 m - ~ 1 . 0 m, 21.0 m - 4 . 5 m, 21.5 m - <2.0 m) to assess - - understory structure. In 1989, only total percent cover of forb was estimated.
The percent covers of grasses, sedges, and mosses were recorded both years.
For each tree species and height strata, I measured average diameter and
average height and estimated percent cover within in the 5-m radius plot with
one exception. I did not record the diameters of deciduous trees in the lower
strata because these diameters were too small to influence visibility. Within an
overstory canopy, there were potentially 3 height strata: a dominant upper strata
composed of the tallest trees (A), a middle strata of subordinate trees (8) and a
lower strata of trees that are 2 m to 10 m tall (C). Four species of conifers were
recorded in the study area: lodgepole pine, subalpine fir, en gel ma^ spruce,
and Douglas fir. There were also 3 species of deciduous trees: thin-leaved
mountain alder (Ainus incana), trembling aspen (Populus tremuloides), and
willow (Salix spp.). Crown closure was estimated with the simple ocular
method (Bunnell and Vales 1990).
Sites were classified as being 1 of 7 habitat types, defined by successional
stage open, shrub, riparian, spaced sapling forest, unspaced sapling forest,
immature forest, or mature forest Sedge meadows, recent clear-cuts, roads, and

13
hydro-line right-of-ways were A considered open habitat, while shrub habitat
(e.g 10-year-old clear-cuts) was all early successional stages that were comprised
mainly of shrubs rather than trees. Riparian habitat was characteristically
dominated by shrubs and associated with water. Sapling habitat was all early
forest successional stages with trees that were on average 6 m tall and 4-9 an in
diameter at breast height (dbh). Immature forest was comprised mainly of
conifer trees whose diameters (dbh) were less than 26 an. Mature forest was
comprised of older trees with diameters greater than 20 cm (dbh). The immature
and mature forests in my study area were uneven-aged and comprised of several
species of trees. The special and complex attributes of border habitats were not
addressed in this study. I included only random sites for habitat descriptions
because inclusion of sites used by does would be biased toward attributes selected
by deer. Habitat characteristics that were measured at random sites were
averaged for each habitat type (Table I).
Sample sizes for habitat types were unequal because sites were determined
randomly or by radiolocating the does. The only exception to this method was
used in locating sites of the 2 sapling habitat types. Sapling habitats were very
rare in my study area and were never used by does nor found within doe home
ranges. These sapling habitats did not have the same species composition: the
spaced sapling habitat was primarily lodgepole pine and the unspaced sapling
habitat was primarily subalpine fir. Five sites were located randomly within
each of these stands.
If visibility changed during the field season because of vegetation growth,
visibilities measured at random sites assessed in August 1988 may not have been
mmparabie to those measured at doe sites located in May through August (but
assessed in August) 1988. To estimate this potential bias, phenology plots were
established to justifv measuring visibility at doe sites and random sites at the end

Tab
le 1
. C
hara
cter
istic
s of
7 ha
bita
t typ
es in
Tho
mps
on P
late
au s
tudy
are
a 19
88-8
9. F
or e
ach
habi
tat t
ype,
mea
n pe
rcen
t cov
er o
f 3 p
lant
gro
ups,
mea
n pe
rcen
t cov
er o
f 3
stra
ta o
f co
nife
rous
and
dec
iduo
us tr
ees,
mea
n tr
ee d
iam
eter
s (d
bh),
perc
ent c
row
n cl
osur
e an
d sa
mpl
e si
zes
are i
ndic
ated
. T
ree
heig
ht st
rata
: A =
a
dom
inan
t upp
er st
rata
of
the
talle
st tr
ees,
B =
a m
iddl
e st
rata
of
subo
rdin
ate
tree
s, an
d C
= a
low
er st
rata
th
at a
re 2
m to
10 m t
au.
Varia
ble
Qpe
n S
hrub
R
ipar
ian
Uns
pace
d Sp
aced
Im
mat
ure
Mat
ure
sapl
ing
sapl
ing
fore
st
fore
st
Sam
ple
size
17
5
10
5 5
15
28
Cro
wn
1 11
7
9 7
56
41
clos
ure
(%)
Forb
s (9
6)
18
20
20
22
14
17
12
Gra
sses
and
43
3
53
0 32
1
4
13
sedg
es (%
) N
on-c
onife
r 14
55
73
67
63
38
4
5
shw
bs (%
) Tr
ee s
trata
A
,B,C
A
,B,C
A
IBIC
A
,B,C
A
,B,C
A
,f?
,C
A,B
,C
Con
ifer
(%)
1,
1,
0 7
,1
,2
1
0,5
,0
10
,24
,3
36,5,2
35
,20
,11
28,20,5
dbh
(cm
) 1,
0,
0
3,
1,
2 11
, 5,
2
16, 1
0,
1 9,
2,
1
13,
8,3
2
1,1
3,2
D
ecid
uous
(%I
0, 0,
7 Q,
0,25
0,
0,36
0,
0, O
0,
0,1
4
0,
1,1
2
0,
O,6

of the 1988 season. I estimated visibilities of the profile pole at standard
distances, directions, and my height. In 1989, visibility at phenology plots was
measured through the field season to monitor temporal changes in visibility.
Three plots in each of 4 habitat types (open, riparian, i m t u r e forest and
mature forest) were visited 5 times during the field season. The sites were
monitored 2-3 weeks apart, beginning in late May and ending in early August.
The plots were located in areas within home ranges of d e r that I had radio-
tracked in 1988.
RESIJL'l'S AND DISCUSSION
Evaluation of Apparatuses
Board - Pole Comparison
1 tested for the equivalency of the profile board, which was used in 1988,
and the profile pole, which was used in 1W, by comparing the visibility scores of
each section of the board with those of the pole. For all 8 height sections for both
the board and the pole (0-2 rn), I calculated the mean of visibility scores from the
4 directions at 15 m for 3 sites. With paired median test and Bonferroni's
conrections, I found that the visibility Sores of dl 8 sections of the board were not
significantly different from that of the pole (n = 3, P > 0.05). Figure 2 shows
visliity scores far the fourth d o n (0-75-1.00 m above the ground) for the 3
sites in 4 directions, and at 4 distances from site centre. Like Griffith and Youtie
t1988), 1 condude that the width of a pro•’ile device is unimportant. Thus, I have
nut distinguished between dab collected with the board or the pole in the rest of
my analyses, and so refer to pale and board data simply as pole data.

Visibility score for profile pole
Figure 2 Relationship between visibility of the profile board and pole at the fourth section (0-75-1 -00 m). Visibility scores at 3 sites (n=48) were measured at my height (1.5 m), 4 distances (5 m, 15 m, minimum distance for 10% deer model visibility at coyote height and at my height), and 4 directions (upslope, downslope, right and left across slope). The number of occasions for each data point are given as values of "nH-

1 7
Deer Model - Pole Comparison
Following the procedure of Griffith and Youtie (1988), I assigned the mid-
values to the visibility scores of the pole data (i-e. 1 = lo%, 2 = 30%, 3 = SO%, 4 =
70%, and 5 = 90%). I then weighted these visibility scores by multiplying them by
the proportion of the deer model potentially visible at that section of the pole
(Fig. 3), and correlated the sum of these weighted visibility scores for the pole
with the deer model visibility (n = 2102, df = 2100, r = 0.91, P < 0.05). Data for this
analysis were collected at 164 sites in 4 directions, at my height, and at 3 or 4
distances for the deer model and pole (n = 2102). Before using correlation
analyses, I examined the residuals from a regression between deer model
visibility and the transformed visibility of the pole for normality and found that
the residuals of deer model visibility have a nonnal distribution.
I obtained higher correlation coefficients (r = 0.91 overall and r = 0.87-0.96
for my 7 habitat types) than did Griffith and Youtie (1988) (r ranged between 0.62
and 0.85 for 6 habitat types). I used a finer scale, (25-cm intervals) than did
Griffith and Youtie (1988) (5(Fcm intervals). In discussing the results of their
bedded deer model and pole comparisons, Griffith and Youtie (1988) suggested
that a finer scale may improve the correlation. Although I did not use a bedded
deer model, I conclude that decreasing length of profile pole sections increased
the amount of variation i? deer model concealment that can be explained by
pole concealnrent. Canfield et al. (1986) tried to correlate the visual cover
measured with the "Hillis" method to that measured with the profile board and
found they were only moderately correlated.
Pole data can be transformed into deer model-like data without a loss of
information, and the visibility of a standing deer can be indexed by either device.
With a profile pole, it is possible to record vegetation structure and density
where this is not possible with the deer model. Also in most situations, the pole

Figure 3. Section numbers of the 8 pole sections and proportions of the deer model potentially visible at different 25-an sections of the pole are provided. Both apparatuses are drawn to scale.

is easier to use than is the deer model.
Evaluation of Procedures
Observer Height
I used the deer model to compare visibility estimated at the 2 observer
heights because all pole visibility data were recorded at my height only and not at
coyote height. I used Spearman's rank correlations to compare the percent
visibility of the deer model recorded at the 2 observer heights (coyote height, 0.75
m; my height, 1.5 m) and 2 distances (5 m and 15 m). I also compared the
minimum distance at which there was 10% visibility of the deer model at coyote
height with that at my height. Visibility at coyote height was highly correlated to
the visibility at my height: 5 m, n = 164, rs = 0.92, P < 0.01; 15 m, n = 164, rs = 0.96,
P < 0.01 (Fig. 4). The minimum distance at which 10% visibility was observed at
coyote height was also strongly correlated with the minimum distance at which
10% visibility was observed at my height (n = 164, rs = 0.95, P < 0.01).
I compared visibility of the deer model at the 2 observer heights to
determine which observer height dowed for discrimination among habitats.
The mean visibility of the deer model at 15 m for each habitat was compared to
that for a l l other habitats; there were 7 habitats, and 21 unique pairs of habitat
comparisons. When the 21 differences resulting from the 21 habitat comparisons
were averaged for both coyote height (0.75 m) and my height (1.5 m), the mean of
the differences was only slightly greater for my height (24.7 m + 4.6 SE) than that
for coyote height (23.6 m + 4.1 SE) and was not significant (Mann-Whitney U test,
n = 42, df = I, U = 214.5, P > 0.05).
Only one other researcher (Riley 1982) measured visual cover at coyote
height; however, he did not compare visibility at coyote height with visibility at
other observer heights. Visib'liity of the deer model at coyote height (0.75 m) and

I I I I I I I I
0 1
20 40 60 80 100 Percent visibility at coyote height
Figure 4. Relationship between mean percent of the deer model visible at 15 m at my height (1.5 m) and that at coyote height (0.75).

2 1
that at my height (1 -5 m) were highly correlated, thus one can be transformed to
the other. Also, the comparison between 2 observer heights demonstrated the
same magnitude of differences among habitats. I recommend that other
researchers measure visibility at their own height because estimating visibility is
easier when standing.
Distance of Observer from Site Centre
At my height, visibility of the deer model in each habitat type decreased
with distance from the deer model (Fig. 5). The rate of decrease was greater for
shrub through mature forest successional stages than that for open habitat. I
compared visibility of the pole at 5 m and 15 m to determine which distance
allowed for discrimination among habitats. The mean visibility score of the
fourth pole section (0.75-1.00 m) at 5 m and 15 m of each habitat was compared to
that of all other habitats. The mean of the differences in visibility scores was
slightly greater for 15 m (1.18 + 0.19 SE) than for 5 m (0.88 + 0.12 SE), but was not
significantly different (Mann-Whitney U test, n = 42, df = 1, U = 187.0, P > 0.05).
Nudcis (1977) found differences in profile board visibility among habitats
to be greater at 15 m than at 5,10,20,25, or 30 m, and recommended that
visibility of a profile board be estimated at a standard distance to ensure
comparability of visibility measures. He also suggested that the standard distance
may be species and geographically specific. Yeo and Peek (1989) used 10 m as a
standard distance for their study of Sitka black-tailed deer (0. h. sifkensis) in
Alaska while Riley (1982) used 6 m for his study of mule deer fawns in Montana,
but none of these researchers discussed why they chose these distances. I
conclude that 15 m is an appropriate distance to use as a standard in my study
area for mule deer.
It has been assumed that visual cover is adequate when 10% of a standing

+ open
+ shrub
+ riparian
qc_ unspaced sapling
4 spaced sapling
-6-- immature forest
+ mature forest
o ] I I I I I I I I I I I I
0 I
10 20 30 40 50 60 70 Distance from deer model (m)
Figure 5. Mean percent visibility of the deer model at my height (1.5 m) and 2 distances (5 m and 15 m) and the mean minimum distance for 10% deer model visibility for each habitat.

23
elk is visible at 61 m or less (Thomas et al. 1979). I measured the minimum
distance for 10% deer model visibility for each habitat at randomly located sites
within doe home ranges and at my height. I found that the mean of this distance
was much less than 61 m for all habitats except open habitats (Fig. 6). A t-test
performed on the entire dataset reveals that the minimum distance for 10% deer
model visibility was sigxuficantly different from 61 m (n = 164, df = I, t = -15.0, P <
0.05). When the data for each habitat were analyzed individually with t-tests and
Bonferroni's sequential correction, open habitat was the only habitat that was not
different than 61 m (n = 33, df = 1, t = 0.5, P > 0.05). Thomas et al.3 (1979)
definition is useful only in determining whether visual cover is present or not.
It does not allow for detecting differences in visual cover quality among habitats
with visual cover. While 61 m is too long to discriminate among habitats, a
distance of 15 m is more suitable in describing quality of visual cover.
Pole Section
I compared visibility of all 8 sections of the pole to determine which
section allowed for discrimination among habitats. The mean visibility of each
pole section at 15 m for each habitat was compared to that of all other habitats.
These same comparisons were done for the 2 pole halves (the sum of the bottom
4 sections and the sum of the top 4 sections) and total pole (the sum of all 8
sections). The mean of the differences between the pairs was divided by 4 for the
2 pole halves and divided by 8 for the total pole to standardize the magnitudes of
visibility scores. The means of the differences were greatest for the fourth (0.75-
1-00 m) and fifth (1.00-1.25 m) sections 7). Although a multisample Kruskal-
Wallis test revealed that differences between habitats were not sigruficantly
different for any of the pole sections (n = 231, df = 10, H = 7.7, P > 0.05), I used the
fourth pole section as the visib'ity variable in further analyses, because more of

opn shb rip usp ssp imm mat Habitat type
Figure 6. Mean and standard error of minimum distances (m) for 10% visibility of the deer model measured at my height (1.5 m) for 7 types of habitats: opn = open, shb = shrub, rip = riparian, usp = unspaced sapling, ssp = spaced sapling, imm = immature forest, mat = mature forest.

Figure 7. Mean and standard error of differences between habitats in visibility scores of each pole section, pole halves (where B is the bottom 4 sections of the pole and T is the top 4 sections of the pole) and total pole at 15 m (where BT is a11 8 sections of the pole.
2.0
1.8- cn
1.6- 6 0 -
1.4- 2. -- - .- n 1.2- cn .- > 1.0- C .- a 0.8- Q) - U r 0.6- Q, L
0.4- ii -
0.2 - - 0.0 1 1 1 1 I 1 I 1 1 1 1
1 2 3 4 5 6 7 8 B T B T Pole section

26
a standing deer could be seen at 0.75-1-00 m than at 1.00-1.25 m (Fig. 3.); therefore
the fourth section was the most biologically meaningful section for estimating
visibilities in habitats. However, lower sections of the pole should be used when
evaluating habitat use by fawns (Riley 1982) and perhaps for bedded adults.
Observer Position
I compared visibility of the pole at 4 different observer positions to
determine which observer position allowed for discrimination among habitats.
Visibility scores were recorded at 4 different positions relative to the centre of the
site where the pole was placed: upslope, downslope, right and left across slope.
The mean visibility of the fourth pole section at 15 m of each habitat in 4
observer positions was compared to that of all other habitats. The mean of the
differences was greatest for the upslope position (Fig. 8), but was not significantly
different from those for the oiher positions (I-way multisample Kruskal-Wallis
test, n = 105, df = 4, H = 2.7, P > 0.05).
Other researchers used the 4 cardinal directions or randomly chosen
directions instead of slope directions (Nudds 1977, Griffith and Youtie 1988) and
made no comment on the influence of observer position. Canfield et al. (1986)
and Yeo and Peek (1989) measured visibility from upslope and downslope
positions. Canfield et al. (1986) hund that visibility of a target increased as the
elevation of a viewer outside the target's stand increased, however, they did not
comment on the influence of elevation of the observer on visibility within a
stand. Position did not influence estimates of visual cover because my study area
was gently sloped. In steeply sloped areas, position could affect estimates of
visual cover. To ensure comparability of my results with those from other study
areas I chose to use the mean of visibility scores recorded at the 4 observer
positions for each site for the remainder of my malyses.

Figure 8. Mean and standard error of differences between habitats in visibility scores of the fourth pole section at 15 m for 4 observer positions and for the mean of the visibility scores recorded at the 4 positions.
- 0.2 - 0.0 I I I I 1
UP down right left mean Obsewer position

Characteristics of Visud Cover
Phenology of Visual Cover
In 1988 and 1989, I used different methods for locating random sites within
doe home ranges, and I measured the visibility of doe and random sites either
during (1 989) or at the end of the fiefd season (1988). Thus, 1988 and 1989 datasets
may not be directly comparable. To assess whether they were comparable, I
monitored 12 phenolcgy plots through the 1989 field season to record temporal
effects on visibility at sites. A series of I-way median tests for each habitat, with
date as the only effect and corrected with Bonferroni's sequential correction,
showed that visibility did not change significantly during the field season for any
of the 4 habitats (n = 15, df = 4, P > 0.05) (Fig. 9).
I concluded that visibility did not change from the end of May until early
August in 1989 for 4 habitat types, and assuming that no significant temporal
changes in visibility occurred for the same period in 1988, I pooled visibility data
from 1988 and 1989 . Loft et al. (1987) found that there was no loss of visual cover
in the first half of suxnrner in plots (ungrzed) that he monitored from 20 June to
28 September. Nudds (1977) measured interseasonal variation in visual cover
and found that visual cover decreased from summer to fall as deciduous trees
and shrubs lost their leaves. He did not d e changes in visibility during late
spring and summer.
Visual Cover in Various Habitat Types
Visibility of the deer modeI in 7 different habitats was described by 8
visibility variables (Table 2). Figure 5 also demonstrates deer model visibility in
each habitat type. Only 2 varhbks of deer d e 1 visibility did not show
signifcant differences between habitats with a 1-way multisample Kruskaf-
Wallis tesk percent visrile at coyote height when only 10% of the deer d e 1 is

+ riparian
+ immature forest
+ mature b e s t
Date
Figure 9. Mean visibility score of the fourth pole section (0.75-1.00 m) at 15 m for each habitat and time period.

Table 2. Means of 8 variables that describe deer model visibility in 7 types of 3 0 habitats. Krusal-Wallis test, n = 42, df = 6, *I? < 0.05. Variable labels: c = coyote height (0.75 m), a = my height (1.5 m), 5 = 5 m, 15 = 15 m, dl0 = minimum distance for 10% visibility, cad10 = percent visible at coyote height and adlO, and acdlO = percent visible at my height and cdlO (see page 9).
Habitat c5' a5" c15* al5' cd1O' adlo* cad10 acd10
Open 94.7% 98-0% 69.2% 77.4% 55.5rn 65.7m 5.3% 27.7%
Shrub 67.4% 73.2% 22.8% 23.4% 15.0m 18.2rn 4.8% 26.3%
Unspaced 46.6% 52.8% 7.2% 9.8% 10.6 rn 11.6 m 8.8% 14.4% sapling
'paced 68.4% 67.4% 12.8% 18.0% 13.8 m 15.4 rn 7.2% 18.2% sapling
immature 84.5% 86.8% 36.7% 39.1% 23.0 rn 23.6 rn 12.8% 14.5% forest
Mature 76.5% 79.6% 27.6% 31.8% 19.6m 2 1 . 8 ~ 1 8.5% 19.7% forest

visible at my height (n = 42, df = 6, H = 8.4, P > 0.051, and percent visible at my
height when only 10% of the deer model is visible at coyote height (n = 42, df = 6,
H = 10.0, P > 0.05). For the other 6 visibility variables, there were significant
differences among habitats (n = 85, df = 6, P c 0.05).
Visibility in different successional stages were examined to determine how
visibility changes during the process of succession (Fig. 10). Visual cover was
sparsest in the earliest successional stage (open habitats). Visual cover was
densest in the shrub and sapling (unspaced) stages and was intermediate in
density in immature and mature stages- Yeo and Peek (1989) measured visual
cover and found that visual cover was greatest in sapling habitats and least in
mixed conifer old growth habitats. They did not include open habitats in their
investigation. Six of the habitats they examined are similar to 4 of my 7 habitat
types: shrub, sapling, immature, and mature or old growth forest. The mean
percent visibilities of their pole at 10 m for these habitats falls between the means
of percent visibility of my deer model at 5 m and 15 m. Becker et al. (1990)
discussed visual cover values of different successional stages for coastal
biogeoclimatic zones. Herb stages (i.e. open habitat) had very poor visual cover
values, while shrub-seedling stages (i.e. shrub habitat), sapling-pole (i.e. sapling),
and young/mature (i.e. immature and mature forests) had poor to excellent
visual cover value depending on density, height, and bole size of conifers and
evergreen shrubs. The values of visual cover of different successional stages that
I obtained in the interior of B.C. appear to be comparable to results obtained by
Yeo and Peek (1989) in coastal Alaska and Becker ei al. (1990) in coastal B.C.
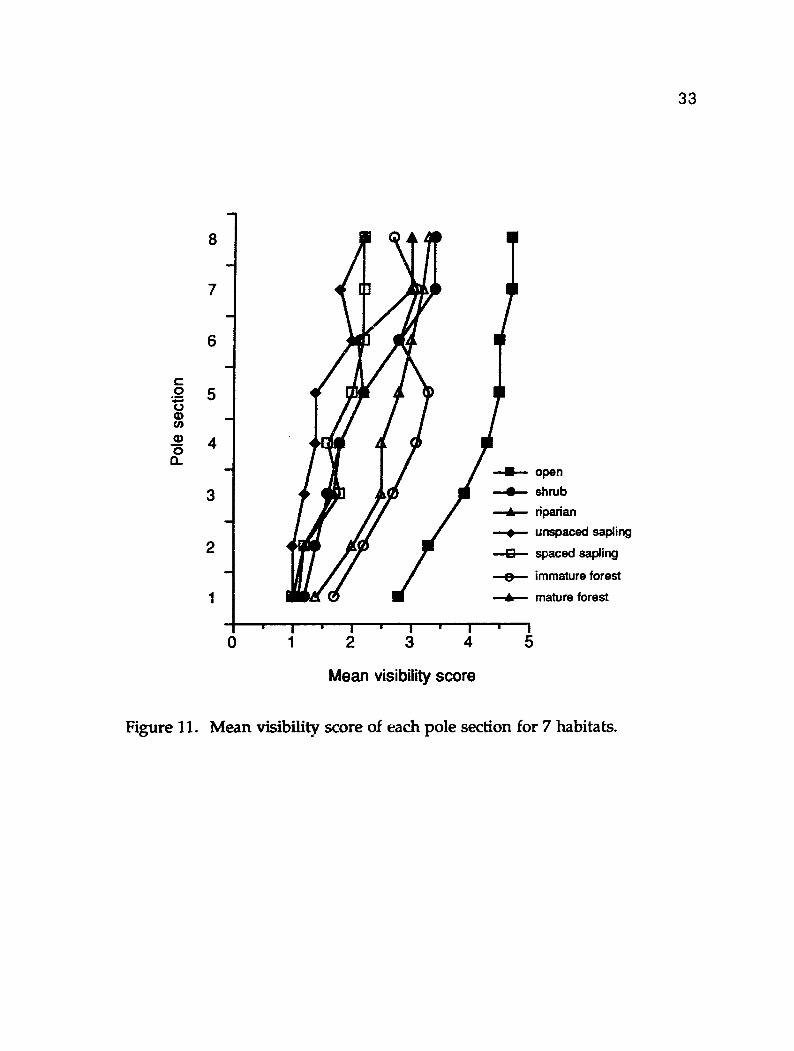
Visibility profiles for all habitats in my study area were similar in shape
=g. 11). A 1-way multisample Kruskal-Wallis test for the visibility of each pole
section showed that visibility of all pole d o n s were significantly different
among habitats (n = 85, df = 6, H = 36.7, P < 0.05). Visibility was least at the lowest

I I I I 1 OPn shb USP imm mat
Successional stages
Figure 10. Mean and standard error percent visibility of the deer model measured at 15 m and my height (1.5 m) for 5 successional stages of a sere. opn = open, shb = shrub, usp = unspaced sapling, imm = immature forest, mat = mature forest.
I
I I I I I I I I I I i 0 1 2 3 4 5
Mean visibility score
immature forest
mature forest
Figure 11. Mean visibility score of each pole section for 7 habitats.

3 4
sections of the pole and incheased with increasing height above the ground. The
pole was most visible at al l levels in the open habitat, less visible in immature
and mature forests and least visible in the remaining 4 habitats. Nudds (1977)
found that, structurally, vegetation was more similar betweex-, habitats in the first
meter above ground level than that in the second meter. In Nudds' study area,
the distribution of vegetation became more uniform at all heights as sera1 age
increased. The rank order of the habitat visibilities for the deer model was the
same as that for the profile pole (Fig. 10 and Fig. 11). The magnitude of
visibilities among habitats appear different because of the differences in shape of
the two apparatuses.
Nudds (1977) compared structural differences within habitat tykes and
concluded that forest habitats were not uniformly structured. I performed a 1-
way multisample Kruskal-Wallis test on deer model and pole visibility at 15 m
using site number as the effect and found no intra-habitat differences in visibility
for any of the 7 habitats (P > 0.05). I conclude that within each of my 7 habitats,
structure and density of their vegetation were uniform.
Components of Visual Cover
I performed a multiple linear regression to determine which physical and
vegetation characteristics contributed to visual cover. The frequency
distributions of the residuals of the regressions were normal, thus I did not
violale the assumption of normality for this parametric test. Because of
autocorrelation between the independent variables, I could not perform
regression analysis on data for each habitat separately. It is not surprising that,
within habitats, habitat characteristics were closely related.
The visibility of the fourth section of the pole (0.75-1.00 m) at 5 m and 15
rn were the 2 dependent variables used sequentially. There were 21 independent

variables used (Table 3). Because total percent cover of non-conifer shrubs was
directly related to the percent cover of non-conifer shrubs at each of 5 heights, I
ran 2 separate analyses, one with the total percent cover and the second with the
percent cover at the 5 heights. None of the percent cover variables for the 5
heights of non-conifer shrubs were significant in the regression, but total percent
cover of shrubs was significant (Tables 3). Thus the regression models presented,
used total percent cover sf shrubs rather than percent cover of non-conifer
shrubs at 5 heights.
Although all the habitat variables were measured for a 5-m-radius plot,
the regression of pole visibility at 15 m fitted the data better ( ~ 2 = 0.52) than that
at 5 m ( ~ 2 = 0.42). This may be further evidence that 15 m is a more appropriate
standard distance for measuring visual cover than 5 m. There were 4 significant
independent variables that the regression models had in common: aown
closure, total percent cover of non-conifers in the shrub layer, percent cover of
deciduous trees in the middle height strata, and percent cover of deciduous trees
in the bottom height strata. There were 3 other variables that were significant
only in the 15 m regression model: percent cover of conifer trees in the top
height strata, percent cover of conifers in the shrub layer, and the diameter of
deciduous trees in the middle height strata. There was 1 variable, percent cover
of conifer trees in the bottom height strata, that was sigruficant in the 5 m
regression model only. Of all the significant variables only 2 had positive
coefficients: aown closure and diameter of deciduous trees in the middle height
strata. In a multiple regression, the sign ~f a coefficient does not necessarily
indicate the nature of the relationship between the dependent and independent
variables.
It is not surprising that percent cover of moss or herbs were not sigmficant
variables because neither plant type occurred frequeny 0.75-1.00 m above the

Table 3. Independent variables in my multiple linear regression model and 3 6 their coefficients when the dependent variables are the visibility scores of the fourth pole section at 5 m and 15 m sequentially. (5 m: n = 155, P c 0.05, ~2 = 0.42; 15 m: n = 155, P < 0.05, ~2 = 0.52) ' P c 0.05.
Independent Variables Coefficient Coefficient
at 5 m at 15 m
Constant +4.90e +4.42'
Slope +0.003 -0.01 0
Crown closure +0.01' +0.014*
Percent cover of moss -0.001 -0.003
Percent cover of herbs -0.001 -0.001
Percent cover of conifer trees in the top height strata -0.01 -0.01 9*
Diameter of conifer trees of the top height strata +0.003 -0.01 4
Percent cover of conifer trees in the middle height strata -0.01 -0.01 3
Diameter of conifer trees of the middle height strata -0.01 -0.01 8
Percent cover of conifer trees in the bottom height strata -0.03' -0.020
Diameter of conifer trees of the bottom height strata +0.05 -0.01 2
Percent cover of conifer in the shrub layer -0.01 -0.030'
Total percent cover of non-conifer in the shrub layer -0.005' -0.006'
Percent cover of deciduous trees in the middle height strata -0.1 4' -0.1 36"
Diameter of deciduous trees of the middle height strata +0.06 +0.092*
Percent cover of deciduous trees in the bottom height strata -0.02' -0.030'

37
ground. As expected, slope did not influence visual cover because the terrain in
my study area was gently sloped. Canfield ei al. (1986) examined the visual cover
of stands when viewed from an opposing slope to the stand. They found that the
viewing angle explained 52% of the variation in visual cover but slope, tree
height, tree and shrub densities and distribution also affected the relationship
between viewing angle and visual cover.
My analyses suggest that shrub densities of coniferous and non-coniferous
plants were important components of visual cover. Although some researchers
(Taber 1961, Black et al. 1976, Loft et al. 1987, Becker et al. 1990) acknowledged that
understory vegetation provides both thennal and visual cover, researchers who
modeled visual cover have not. Two visual cover models for ungulates
proposed by Smith and Long (1987) and McTague and Patton (1989) were based
on the boles and live crowns of conifers and did not include shrubs. Modeling
the characteristics of shrubs is problematic, but their role in visual cover cannot
be ignored. Lyon (1987) used a model that included boles but did not include
shrubs, but later realized that tall shrubs can provide visual cover. At the shrub
densities he recorded in some treatments of lodgepole pine stands, visual cover
increased from less than 10% when shrubs were not considered to over 90%
when they were.
The importance of deciduous trees as a component of visual cover has not
been examined by other researchers because they attempted to simulate visibility
only in coniferous stands. My study examined visual cover for a spectrum of
successional stages, not all of which were pure coniferous forests. In my study
area, diameter of conifers was not an important predictor of visual cover, but
percent cover of conifers in the top strata was. Previous models of visual cover
(Smith and Long 1987, Lyon 1987, McTague and Patton 1989) tended to use
variables such as average diameters, crown closure, density, basal area and stand

3 8 density index, because they are common forestry measures; however, these may
have no direct relationship to visual cover. Smith and Long's (1987) model of
visual cover used only coniferous tree diameters (dbh), density and spatial
distribution of trees. When they tested their model in the field, they found that
observed visual cover did not agree with their simulated results. Smith and
Long (1987) attributed &is discrepancy to irregular spacing of trees in field
situations, but I suggest that this discrepancy may be attributed to the exclusion of
understory and deciduous components from their model.
My regression analyses provide important insights into the structure of
visual cover. Habitat characteristics that have not traditionally k n included in
models of visual cover are important components of visual cover: e.g. percent
cover of coniferous and non-coniferous shrubs, percent cover of deciduous trees
in the middle and lower height strata and the diameter of deciduous trees of the
middle height strata. My regression analyses also show that crown closure and
percent cover of conifers in the top height strata which are commonly used in
some form in models, are sigruficant factors affecting visual cover.
CONCLUSIONS
In evaluating the use of 3 apparatuses to estimate visual cover (profile
board, profile pole and deer model), I found that the board and the pole are
equivalent, and that data collected with the pole can be transformed into percent
visibility of a deer model. The pole is convenient to use and provides data on
vegetation structure and deflsify. In comparison, the deer model is less
convenient to lnse and does not provide adequate information on vegetation
structure. Therefore, the profile-pole is the most versatile and convenient
apparatus for measuring visual cover. My results confirmed those reported by

Griffith and Youtie (1988).
I evaluated procedures of measuring visual cover and proposed a set of
procedures that increase our ability to detect differences in visual cover values
among habitats. Visual cover can be measured at the observer's standing height
because there appears to be no advantage to h e l i n g at coyote height. It is best to
use a standard distance between observer and apparatus for habitat or site
comparisons, which probably will be species and geographically speafic. For
mule deer in my study area, it was more appropriate to measure visual cover
quality at 15 m than at 61 m. The fourth (0.75-1.00 m) and fifth section (1.00-1.25
m) were the best pole sections at which to measure visibility for conditions
encountered in my study area for adult mule deer. I suggest that lower sections
of the pole be used when evaluating habitat use by fawns and perhaps bedded
adults. In my study area, there was no significant advantage for the observer to
stand upslope, downslope or across-slope from the apparatus when estimating
visual cover.
Temporal changes in visibility in 4 habitats was not significant during late
spring and summer. Horizontal visibility for each habitat can be measured at
any time during late spring and summer to assess a representative value for
visual cover of the various biogeodirnatic zones present in my study area.
Visibility was greatest in open habitats and least in unspaced sapling stands.
Regression analysis revealed habitat characteristics that predict pole visibility and
thus visual cover. The most important habitat characteristics to measure appear
to be: crown closure, percent cover of conifer trees in the top height strata,
percent cover of coniferous shrubs, total percent cover of non-coniferous shrubs,
percent cover of deciduous trees in the middle and bottom height strata, and
diameter of deciduous trees of the middle height strata. Because there were
gentle slopes in my study area, vegetation characteristics rather than physical

characteristics such as slope were important predictors of visual cover.
Existing visual cover models that translated cover guidelines into forestry
prescriptions have 2 weaknesses. First, they do not include deciduous tree and
shrub densities. Second, they a l l use untested "standard" criteria (e.g. visual
cover is adequate when 10% of a standing elk is visible at 61 m or less) to
evaluate the value of visual cover in various forest types. Only Smith and Long
(1987) questioned this assumption. Wore evaluating visual cover, I recommend
that investigators conduct preliminary research to ascertain a standard distance
that is appropriate for their species and study area by examining visibility-
distance functions. Also, models that simulate visual cover should include
characteristics of shrubs and deciduous trees.

4 1
C W R THREE
INFLUENCES OF VISUAL COVER AND OTHER FACTORS ON DIURNAL SITE
SELECTION BY MULE DEER
Security cover is a structural resource that reduces the probability of
detection or attack by a predator (Elton 1939, MacHutchon 1988). Because
structural characteristics of habitats vary aaoss a forested landscape, security
cover also varies and could therefore affect use of forest lands by Rocky
Mountain mule deer. Although security cover appears to be an important
component of mule deer habitat, its role in habitat selection has not been clearly
demonstrated (Peek et al. 1982, Thomas et al. 1986). Several studies have assessed
visual cover (as the most easily measured structural component of security
cover) in various habitat types fox ungulates (Nudds 1977, Riley 1982, Canfield et
al. 1986, Loft et al. 1987, Smith and Long 1987, Griffith and Youtie 1988, Servheen
and Lyon 1989, Yeo and Peek 1989). None of these, however, determined the
importance of visual cover resources relative to other important resources in
habitat selection.
The selection of habitats is thought to occur at 3 levels (Johnson 1980).
First, the geographical range of a species is selected by individuals. Then, each
individual selects a mosaic of habitats for their home range (second level).
Lastly, sites for basic activities are chosen within home ranges (third level).
Individuals use habitats in which they are most likely to survive and reproduce
(Krebs and Davies 1984). Use of a particular site is believed to be a function of the
costs arrd benefits available to the animal at the site (Peek et al. 1982). For
example, a cost of using a particular site may De increased predation risk due to a
lack of visual cover. A benefit of using a particular site may be high availability
of a valuable resource such as food.

4 2
The availability and distribution of basic resources such as food, water and
cover could influence selection of sites by does. I compared the availability of
food, water and visual cover at sites used by does to that at randomly located sites
within doe home ranges to establish the relative importance of these resources
in site selection. There was also the possiwty that sites are used by d m to avoid
human disturbance, so minimum distance to roads of s i te is examined. Atso, I
investigated whether visual cover functioned for thermal or security cover. To
assess the n e d for t h e d cover, I examined micro-climate conditions (air
temperature, wind speeds, and light intensity) at each site used by does. Crown
closure was estimated as an index of thermal cover at random sites and sites used
by does (Black et al. 1976, Leckenby et a1 1982, Thomas et aL 1986, Lyon 1987,
Smith and Long 1987). Structural characteristics of visual cover for sites used by
does and random sites were described in terms of visibilities of the deer model
and profile pole. The minimum distance for 10% visibility of a deer model was
measured at sites used by does to further evaluate the validity of the common
definition of adequate visual cover (Thomas et al. 1979).
In m i n g wh&er deer were influenced by visual cover in site
selection, I proposed a null and an alternate hypothesis.
Hg Visual cover at sites used by does was not different than that at randomly
located sites within doe home ranges.
In this case, deer would have used sites on the basis of hbitat
characteristics other than visual cover or at random.
HI V i d cover at sites used by does was different than thzr at randomly
bcated sites wiihin doe b m e ranges.
More +ally, does should have used sites with less visual cover when

4 3
the costs of predation were low or the bedits available by using the site were
high, Does should have used sites with less visual mver in spring when their
immediate energy rqukments were high and if there is a trade-off between
f d resources and visual mver resources. During spring when their fat reserves
were depleted (Mautz 1978, Simpson 1988), deer should have risked foraging on
vernal vegetation which was first available in open habitats to avoid continued
weight loss. Access to high quality forage early in spring a d d have effettlvely
shorened the duration of low energy and nutrient intake and increased survival
(Stridand 1975). The l ~ s f s of predation would have been relatively low in
spring because adult does and yearlings would not have been as vulnerable to
predation as young-of-theyear in summer. Visual cover at sites used by does
before parturition should have been lower than that at randomly sites within
doe home ranges or than that at sites used after parturitioa Simpson (1988)
o b e ~ e d that some does used open habitats that provided high quality food
before moving to their summer ranges where they gave birth.
In another scenario, does should have used sites with more visual cover
than that which was available at randomly located sites when the costs of
predation were high. After parturition, if a doe was ammpanied by her
newborn fawn into an opening, she would be risking her fawn's life and her
own by using a site with W e visual cover. Fawns are more vulnerable to
predatioln than add& (Trainer 19751, therefore the costs of using an open habitat
d d be greater for a doe with a fawn than those for a solitary doe. I predicted
that sites used by a doe with a fawn would have greater visual cover than sites
used by a solitary doe Daa with fawns should have used edges (ecotones) and
small openings* whereas solitary does w d d have ventured further from visual
cover into openings to forage on high Quality food and to bed near forage sources.
Edwards (1983) found that moose (.Alas nlces) cows with calves "show a different

4 4
distribution and diet from solitary adults and yearling moose". During spring
just before calving, moose cows on Isle Royale moved from an area with high
quality food to a predator-free area with low quality food. Visual cover was not
investigated by Edwards (19831, but her data indicate that behaviour of females
with young was influenced by predation risk.
METHODS AND MATERIALS
In both 1988 and 1989, I began my field work in June and ended it in
August. Parturition occurred in June and July for the majority of does. By the
time the fawns were 6 weeks old, they were relatively mobile and tended to
follow their dams rather than hide. All radio-tracked females produced fawns.
Eleven females were radio-tracked regularly in 1988; 10 were adults and 1 was a
yearling. Twelve adult does were radbtracked in 1989. A total of 15 different
individuals were tracked during the study.
Radietracking was conducted dwing daylight between 0800 and 2000 PST.
Triangulation was initiated from roads to obtain a rough estimate of the doe's
location, Triangulation was then continued on foot until I was very close to the
doe. I approached the does from a downwind direction and as silently as
@ble to avoid detection. The exact location of the doe was determined by
seeing or hearing it or eonfinning its location with physical sign. I looked for
fawns and signs of feeding, bedding, running and defecating. Besides finding a
hiding fawn, small tracks, small pellets, and small beds were assumed to confirm
the presence of a fawn I a s s a d from the doe's behaviour whether the doe had
a fawn nearby (White et al. 1972).
I compared Ue behaviour of does before and after parturition. The
treatments for this analysis were does without fawns (i.e. before parturition) and

4 5
does with fawns (i.e. after parturition), and the control treatment was
represented by randomly located sites within doe home ranges. Thus, site types
were classified in 3 tiers. Sites were either where does were found, with or
without fawns, or where sites were randomly located within doe home ranges.
Sites used by does were classified as either being used before or after parturition.
Also, doe sites were classified as to what kind of activity the doe was engaged in
at the site: bedding, feeding or unknown. One random site was determined for
every doe site with only 6 exceptions. Methods for choosing random sites
within home ranges of does were described in Chapter 2.
Visual cover was an important component of security cover for the
system that I investigated. Coyotes, for which vision is an effective sense for
prey location (Wells and Lehner 1978), were a common predator of fawns in my
study area (Simpson 1988). I measured visual cover by estimating the horizontal
visibility of a deer model (1988,1989) and of a profile board in 1988, and a profile
pole in 1989. Although visibility of the profile pole (or board) was positively
correlated to visibility of the deer model ( s e Chapter 2), I focussed on the
visibility of the deer model at coyote height to assess the use of visual cover by
deer and to assess the validity of the common definition of adequate visual
cover (Thomas et al. 1979) because a direct measure is better for experimental
purposes than a correlated measure.
Descriptions of the apparatuses (e.g. the deer model, the profile board, and
the profile pole) used to measure visual cover and of the procedures used in
measuring visual cover were given in Chapter 2. The profile board was used in
1988, but I used the less ambersome pole in the 1989 field season. I have not
distinguished between data collected with the board and the pole in my analyses
because the width of a visual cover device was unimportant (Griffith and Youtie
1988, Chapter 2) and so referred to pole and board data simply as pole data.

4 6
Where other researchers measured percent concealment of a model or profile
apparatus (Nudds 1977, Griffith and Youtie 19881, I measured percent visibility.
These 2 measures were complementary and results are equivalent (see Chapter
2). Griffith and Youtie (1988) demonstrated that visibility measures are
repeatable because there were no differences in visibility values recorded by
different observers. However, to eliminate inter-observer bias, 1 estimated all
deer model and pole visibilities myself.
A variety of habitat characteristics were assessed at sites used by deer and at
random sites. Within a 5-m radius of the site centre, total percent cover of forbs
was estimated in 1989, while a percent cover by each forb species was estimated
in 1988. Several other site characteristics were assessed that could affect use sf
visual cover: minimum distance to water and roads, air temperature,
occurrence of precipitation, incident light intensity (1989 only), wind speed (1988
only), and crown closure. Air temperature was measured in the shade just
above ground level with a thermometer. Incident light intensity was recorded at
shoulder height with a light meter at the site centre and in a nearby clearing.
Wind speed at shoulder height was measured with an anemometer at the site
centre. Crown closure was estimated with the simple ocular method (Bunnell
and Vales 1990). The same procedure for site description was followed at both
random and doe sites except micro-climate variables were measured only at doe
sites.
Understory characteristics also were measured in August for random sites
in 1988, whereas understory characteristics were measured throughout the
season at doe sites in 1988 and 1989 and at random sites in 1989. There was a
potential seasonal bias in comparing understory characteristics at doe sites with
those at random sites. I estimated percent cover of forb at phenology plots and
tested for seasonal &ects on percent cover of forbs with a multisample Kruskal-

4 7
Wallis test on the phenology dataset. Like Loft et al. (1987), I found that percent
cover of forb peaked in mid-summer and then declined for all four habitats, but
these changes in availability were insignificant whether I used date in a 1-way
analysis (n = 56, df = 4, H = 8.79, P 7 0.05) or date and habitat in a 2-way analysis (n
= 56, df = 19, H = 26.0, P > 0.05). Because there was no seasonal effect on percent
cover of forbs, the date on which data were collected does not confound forb
availability analyses. Although all of my 1988 random sites were assessed at the
end of the field season, I conclude that these sites can be compared without bias
to my doe site data and 1989 random site data which were collected throughout
the field season.
RESULTS
Sample Sizes
I located 81 sites used by does anci their fawns: 46 in 1988 and 35 in 1989.
During the study, 3-9 sites were identified for each of the 15 does that were
tracked. Among the 81 sites used by does, 56 were bedding sites and 25 were non-
bedding sites; 23 were used by does before parturition and 58 were used after
parturition. Of the 56 bedding sites, 12 were used before parturition and 44 were
used after parturition. f measured visibility at all but 2 of 81 doe sites. For 6 of 81
does sites, I did not have corresponding random sites (n = 75). Sample sizes for
my analyses of visibility data were 4 times greater than that of other datasets
because at sites, visibility variables were measured: upslope, downslope, and
right and left aaoss the slope. The sample size for percent cover of forbs was less
than 81 because I neglected to record it at 3 sites used by does (n = 781, and only 73
random-doe site pairs were available for forb analyses because not all of these 78
sites had random site partners. Light intensity was measured only in the first

4 8
field season because it did not occur to me to measure light intensity until after
my first field season (n = 31). I did measured wind speed in 1988 only because
there was no wind on 70% of deer locations in 1988, and when present it was
gusty and difficult to measure accurately (n = 42). Neither of these climate
variables was recorded on every occasion during their respective field seasons
because the equipment was not always functioning or available.
Percent Cover of Forbs
Although I measured the percent cover of several potential forage types,
my forage analyses focus on forbs because forb make up -97% of the diets of
Rocky Mountain mule deer during summer (Wilkins 1957, Lovaas 1958, Kufeld
et al. 1973, Willms et al. 1980, Collins and Urness 1983). Forb are also one of the
most digestible (65% digestibility) and most nutritious (13% crude protein) forage
types during summer (Schwartz and Hobbs 1985), and so they are considered to
be the most important forage type to deer.
Forb - Cover Tradeoff
I tested for the existence of a food - cover tradeoff with regression analyses.
If there was a tradeoff, then I expected to find a significant positive linear
regression between visibility and percent cover of forbs and a significant negative
linear relationship between crown closure and percent cover of forbs. A simple
linear regression between the visibility of the deer model at 15 m and coyote
height and forb coverage at doe and random sites (Fig. 12) was insignificant (n =
152, df = 151, r2 = 0.016, P = 059). A simple linear regression between crown
dosure and forb coverage was also insignrfiwt (n = 152, df = 151, r2 = 0.1 69, P =
0.11) (Fig. 13). There was no evidence that a forb - visual cover tradeoff existed in
my study area in summer.

Percent of model visible at 15 m
Figure 12. Relationship between percent visibility of the deer model measured at 15 m and coyote height (0.75 m) and percent cover of forb.

Percent crown closure
Figure 13. Relationship between percent crown closure and percent cover of forb.

Percent Cover of Forbs at Sites
Although does did not have to choose between good visual cover and
good forb availability, they may have used sites on the basis of percent cover of
forbs. I used 2 types of analyses to examine percent cover of forbs at doe sites.
First, a I-tailed Wilcoxon matched-pair rank-sum test demonstrated that percent
cover of forbs at doe sites was not sigruficantly (n = 73, P > 0.05) different from
that at random sites (Table 4). Also, percent cover of f o r b at sites used by does
before parturition was not significantly different from that at sites used by does
after parturition (n = 20, P > 0.05). Second, I performed 2 separate analyses of
variance for activity type and reproductive status because there was only one doe
for which I had all 4 combinations of activity type and reproductive status sites.
Although analysis of variance (ANOVA) data should k normally distributed, I
obtained the same results when I used data which had been normal rank
transformed. Becavse ANOVA is robust with respect to normality, results from
an ANOVA on untransformed data should be valid. The only signhcant main
effect was deer identity and there were no sigruficant interactions between
variables (Table 5). Does did not appear to use sites on the basis of percent cover
of forbs before or after parturition, but individual deer did use sites differently.
Water Availability
To test whether does used sites on the basis of their proximity to water, I
compared the minimum distance to water sowces (that I was able to identify) for
doe sites and random sites (Table 6). No significant results were found witk a 2-
tailed Wilcoxon matched-pair rank-sum test for the doe-random comparison (n
= 75, P > 0.05) or the beforeafter parturition comparison (n = 20; P > 0.05). With 2
3-way ANOVAs using site type (doe or random), deer identity and either activity
type (bedding or not) or reproductive status (fawn or not), both runs

Table 4. Mean percent cover of forbs at sites used by does and random sites. 5 2 Site types: R = random, D = doe, Bf = before parturition, A = after parturition, NBd = non-bedding, and Bd = bedding.
Site Type n R a n g e (%) M e a n (%) S E (%)
R 75 0-70 15.9 1.8
D 78 0-67 15.3 2.0
DBf 22 0-51 15.5 2.8
DA 56 0-67 15.3 2.5
DNBd 23 0-66 18.3 4.3
DBd 55 0-67 14.1 2.1
DBf Bd 12 0-28 12.9 2.7
D A M 43 0-67 14.4 2.6

Table 5. ANOVA table of percent cover of forbs at sites used before and after 5 3 parturition. Site = Site type, Id = Doe identification, Repro .=
Reproductive status.
- -
Factors Sum of df Mean F-ratio P Test term squares square
Site 49.9 1 49.9 0.32 0.58 Site x Id
Id 7350.1 1 1 668.2 2.97 0.00 Residual
Site x Id 1694.8 11 1 54.1 0.68 0.75 Residual
Repro 17.5 1 17.5 0.06 0.81 Id x Repro
Site x Repro 399.0 1 399.0 1.44 0.26 Site x Id x Repro
Id x Repro 3244.1 1 1 294.9 1.31 0.24 Residual
Site x Id x Repro 3052.5 1 1 277.5 1.23 0.28 Residual
Residual 16673.0 74 225.3
Total 32533.0 121

Table 6. Mean minimum distances to water (m) from sites used by does and 5 4 random sites. Site types: R = random, D = doe, Bf = before parturition, A = after parturition, NBd = non-bedding, and Bd = bedding.
Site Type n Range (m) Mean (m) SE (m)
DNBd 25 0 - 500 105 30

demonstrated that deer identity was the only sigruficant main effect and both
activity type (n = 120, df = 32, P < 0.05) and reproductive status (n = 126, df = 35, P
c 0.05) interacted with deer identity. Variation among individuals prevents
simple interpretation of results: some deer used sites near water sources while
others did not.
Deer may have responded to the proximity of water for reasons other than
as sources of drinking water. Other resources associated with riparian habitat,
such as lush forage and willow thickets which provided dense visual cover may
promote use of surrounding habitats. Also, areas with running or standing
water may provide non-visual (olfactory) security cover for deer by masking
their scent from predators (Sweeney et al. 1971, Simpson 1988). I tested for a
preference for riparian habitat by comparing the percent of sites used by does that
were in riparian habitats to the percent of random sites that were in riparian
habitats. Although a greater proportion of sites used by does are in riparian
habitats (24%) than that for random s i te (13%), a maximum likelihood test of
independence showed that site choice was independent of habitat type (n = 154,
df = 4, L = 5.69, P > 0.05). This result indicated site selection is independent of
habitat type (as I had classified them). Simpson (1988) suggested that does
switched from using forested or open habitats to using riparian areas during and
after parturition. I found this to be true for only 3 of 15 does and the opposite is
m e for 4 of 15 while I doe used riparian areas both before and after parturition (7
of 15 does were not located both before and alter parturition). It appears that
preference for riparian habitats varied among individuals.
Visual Cover
Visibility of the Deer Model at Sites used by Does
I tested for a preference for sites with low visibility by does with a I-tailed

Wilcoxon matched-pair rank-sum test with Bonferroni's sequential correction
for all 8 visibility variables. The test showed that only the minimum distance at
which 10% of the deer model could be seen at coyote height was significantly
different between sites used by does and random sites (n = 380, P < 0.05). The
visibility of the deer model at coyote height at 15 m, and the minimum distances
at which 10% of the deer model could be seen at coyote height and my height
were significantly lower at sites used after parturition than that at random sites
(n = 212, P < 0.05). No variables were significantly different between sites u s 4 by
does before parturition and random sites.
I chose 2 visibility variables among the 8 on which to conduct further
statistical tests. Because coyotes are a common predator of deer, I decided to use
the variables which were measured at coyote height. I used the visibility at 15 m
and coyote height because 15 m is a more biologically relevant distance than 5 m
for deer in terms of predator-prey interactions di.e. 5 m is too short). Also, 15 m is
better than 5 m for distinguishing among habitats (see Chapter 2). I also was
interested in the minimum distance at which only 10% of the deer model can be
seen at coyote height in order to compare it to 61 m which is used in the
definition of adequate visual cover proposed by Thomas et at. (1979).
Visibility of the Deer Model at 15 rn and Coyote Height
I tested the visibility of the deer model at 15 m and ooyote height (Table 7)
with 2 3way ANOVAs. I used 3 independent factors: site type (random or doe),
d e e ~ identity, and either activity type (bedding or not) or reproductive s t a m
before or after parturition). For both ANOVAs, deer identity was the only
significant variable and the only significant interaction was between site type and
deer identity (activity type run, n = 476, df = 10, P < 0.05; rqmiutive status ntn,
n = 500, df = 11, P < 0.05). In Figure 14, I plotted the interaction between site type

Table 7. Mean percent visibility of the deer model at coyote height and at 15 5 7 m for sites used by does and random sites. Site types: R = random, D = doe, Bf = before parturition, A = after parturition, NBd = non- bedding, and Bd = bedding.
Site Type n Range (%) Mean (%) SE (%)
R 75 0-1 00 36.6 3.2
D 79 0-1 00 30.9 2.9
DBf 22 0-1 00 38.9 6.2
DA 57 0-1 00 27.9 3.2
DNBd 25 0-1 00 28.6 5.0
Df3d 54 0-1 00 32.0 3.6
O B W 11 0-1 00 45.1 8.6
DAM 43 0-1 00 28.7 3.8

- 3 0 - 0 , 0 2 4 6 8 10 12 14 16
Rank
Figure 14. Differences in mean percent visibility of the deer model at 15 m and coyote height (0.75 m) for random sites and sites used by 15 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for random sites (R) is greater than that for sites used by that doe (D).

and deer identity. For 8 of 15 deer, mean visibility at random sites was greater
than that at the sites used by doe, but in 7 of 15 deer the mean visibility at
random sitc:; was less than that at sites used by does . When I added a categorical
variable that was rested in deer identity and either activity type or reproductive
status, the nested variable was the only sigruficant variable and the only
significant interaction was between it and site type for both runs (activity type, df
= 39, P < 0.05; reproductive status, df = 37, P < 0.05). When I plotted the
interaction terms, there were no particular deer acting as outliers. These results
were consistent for 3 forms of the data: untransformed, arcsin(square root)
transformed, and ranked-normal transformed. There was not a consistent trend
in site selection behaviow exhibited by does regarding visual cover available at
the sites they used.
Distance at Which Only 10% of the Deer Model Can be Seen at Coyote Height
Mean minimum distances at which only 10% of the deer model can be
seen at coyote height are described for 8 site types in Table 8. A t-test performed
on the entire dataset demonstrated that the minimum distance for 10% deer
model visibility was significantIy Merent than 61 m (n = 154, df = 1, t = -16.2, P <
0.05). When datasets for each site type were analyzed individually with t-tests
and Bonferroni's sequential correction, the minimum distance for 10% deer
model visibility for every site type was also significantly different from 61 m (df =
1, P a 0.05).
Minimum distance at which only 10% of the deer model can be seen at
coyote height was tested with 2 3-way anafyxs of variance. The 3 effects were:
site type (random or doe), deer identity, and either activity type (bedding or not)
or reproductive status Wore or after parturition). For both ANOVAs, deer
identity was the only significant variable. Only 11 of 15 does could be used in the

Table 8. Mean minimum distance (m) at which only 10% of the deer model 6 0 can be seen at coyote height for sites used by does and random sites. Site types: R = random, D = doe, Bf = before parturition, A = after parturition, N M = non-bedding, and Bd = bedding.
Site Type n Range (m) Mean (m) SE (in)
R 75 5-1 55 27.4 2.7
D 79 6-274 23.7 3.4
DBf 22 8 - 42 22.6 2.3
DA 57 6-274 24.4 4.7
DNBd 25 8-274 30.0 10.2
D M 54 6 - 66 20.8 1.6
DBfBd 11 8 - 42 25.4 3.2
DAM 43 6 - 66 19.7 1.8 - il

8 1
ANOVA for activity type because I identified both bedding and non-bedding sites
for only I1 of 15 does. Two sipficant interactions occurred between site type
and deer identity (n = 476, df = 43, P < 0.05) and between site type, deer identity
and activity type (n = 500, df = 47, P < 0.05)- Figure 15 demonstrates the
interaction between site type and deer identification while Figures 16 and 17
demonstrate the 3-way interaction between activity type, site type and deer
identification. These interactions were difficult to describe because there is no
regular pattern. For 8 of 15 does, the minimum distance was longer for random
sites than that fur sites used by does. Results were similar for bedding sites: for 7
of 11 does the minimum distance for 10% visibility is longer for random sites
than that for bedding sites used by does. When comparing non-bedding sites
with bedding sites, for 6 of I1 does, the minimum distance was longer for non-
bedding sites than that for bedding sites.
When I added a categorical variable that was nested in deer identity and
either activity type or reproductive status, the nested variable was the only
significant variable and the only signrficant interaction was between it and site
type for both runs (activity type, n = 476, df = 43, P < 0.05; reproductive status, n =
500, df = 47, P < 0.05). When I plotted the interaction tenns, there were no
particular deer acting as outliers. These results were not completely consistent
for 2 forms of the data: untransfonned and ranked-nod transformed.
It seems that individual does use sites differently with regard to visual
cover. The does that used sites with lower visibility at 15 m were the same does
that used sites with shorter minimum distances for 10% visibility than those
measured for random sites. One would srpect this result because these visibility
varhbfes are correlated (n = 154, df = 152, rs = 0.92, P < 0.01). k e is some
indication that does did not use sites r m d d y , but there was not a clear trend in
the relationship between the visibility at sites used by does and that at random

Q, U t E? a C s
Figure 15.
Rank
Differences in mean minimum distance (m) at which only 10% of the deer model can be seen at coyote height (0.75 m) for random sites and sites used by 15 does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for random sites (R) is greater than that for sites used by that doe (D).

Figure 16.
Rank
Differences in mean minimum dktance (m) at which only 10% of the deer model can be seen at coyote height (0.75 m) for random sites and bedding sites used by I1 does where deer identification is ranked in order of difference. A positive value for a difference indicates Uat the mean visibility for random sites (R) is greater than that for I d d i n g sites used by that doe (B).

Rank
Figure 17. Differences in mean minimum distance (m) at which only 10% of the deer model can be seen at coyote height (0.75 m) for bedding and non-bedding site used by I I does where deer identification is ranked in order of difference. A positive value for a difference indicates that the mean visibility for non-bedding sites (NB) is greater than that for bedding sites used by that doe (B).

sites. Activity type appears to have influenced the choice of sites regarding the
minimum distance for 10% deer model visibility at coyote height, but not for the
visibility of the deer model at 15 m and coyote height. Reproductive status did
not play a significant role in the selection of site visibility.
Visibility of the Profile Pole at Sites Used by Does
Two Sway multi-sample median tests adjusted with Bonferroni's
sequential correction were performed on 11 pole variables which were measured
at 15 m: visibilities of the 8 pole sections, sum of visibilities of all 8 pole sections,
sum of bottom 4 sections of the pole and sum of the top 4 sections of the pole.
Again, deer identity, site type, and either activity type or reproductive status were
used as sample strata. There were sig"ificant differences for 9 of the 11 variables,
according to both Sway median tests (activity type, n = 377, df = 43, P < 0.05;
reproductive status, n = 405, df = 47, P < 0.05); the exceptions were the visibilities
of the bktorn 2 sections of the pole (P > 0.05). Despite the si-cant differences
at 6 s f 8 pole sections, profiles of pole visibility for 6 site types look very similar
(Fig. 18). Significant differences could be attributed to variation in doe behaviour
rather than site type, activity type or reproductive status. Visibilties of the pole
and deer model were highly correlated (Chapter 2) so analysis of pole visibility
would duplicate analysis of deer model visibility.
Miao-Chate Con& tions
I measured air temperature, wind speed and incident light intensity to
assess the need for thermal cover by does. Air temperature, wind speed, and
light intensity at the time md place of doe locations are described in Table 9.
Figures 19,20 and 21 are frequency distn'butions of wind speed, short-wave
radiation, and air and operative temperature. Estimates of short-wave radiation

Random
Doe Before
Doe After
I . I t 1 , f . I t
0 1 2 3 4 5 1
Mean visibility score
Figure18. Meanvisibilityof eachpolesection at 15m and my height (1.5 m) for doe and random sites. I = 0-20%, 2 = 21-40%,3 = 41-60%, 4 = 61-8096,s = 81-100%.

Table 9. Three miaeclimate parameters estimated for does on the Thompson Plateau.
Factors n Range Mean SE
Air Temperature (C) 81 1.0-28.0 14.8 0.6
Wind Speed (rn/sec) 42 0.0-8.0 0.9 0.3
Light Intensity (lux) 31 15,000- 50,000 12,000 251,000

Wind speed (m/sec)
Figure 19. Frequency distribution for values of wind speeds recorded in 1988 (n = 42).

100 150 200 250 300 350 400 450 500 550
Short wave radiation (W/m *)
Figure 20. Frequency distribution for values of short-wave radiation recorded in 1989 (n = 31).

Air temperature
Operative temperature
Temperature (C)
Figure 21. Frequncy distributions of air temperature (n = 81) and operative temperature estimates for 1988 (when short-wave radiation was assumed to be 200 W/rn2) and for 1989 (when wind speed was assumed to be zero.

7 1
were dfulated from recorded light intensities using equations in Appendix 1
from Parker and Giklingharn (1986) assuming that a l l measurements were taken
at 1200 PST. Tkte equations used gave estimates of incident short-wave radiation
for standing, captive mule deer. Demarchi (unpubl. data) who measured global
radiation (which includes both long-wave and short-wave radiation) in an open
habitat in my study area reported 100-1000 ~ / m 2 from 1200-1400 PST. His
results confirmed that my values for short-wave radiation were reasonable.
I calculated estimates of operative temperature from air temperature,
wind speeds, and short-wave radiation, md compared them iu lower (+5.0 C)
and upper (+23.5 C ) aitid operative temperatures for adult mule deer in
summer pefage (Parker and Rdbbins 19&Q, Parker and Gillingham 1%).
Operative temperature describes the effective temperature experienced by a doe
by incorporating the effects of air temperature, wind speed and indent radiation
(Parker and GIUingham 1986). Because neither wind speed nor Iight intensity
were measured during both field seasons, the operative temperatures are only
approximate estimates. T'he operative temperatures for the 1989 dataset were
calculated for a zero wind speed which was the most ~OIIUIZD~ condition in 1988
(fig- 191 and in 1990 (Dernarchi, pers. anrm.), and the operative temperatures for
the 1988 dataset were estimated with 200 ~ / m 2 which was the mode of the 2989
short-wave radiation distribution W I ~ . 20).
In both 1388 and 1 W 8 there were d y 11 occasions of 81 when the
operative temperature fell outside the thermoneutral zone. On all of these
od=catSiofls operative temperatures were higher than the upper critical operative
teml~peraiture, and does were bedded on 10 of these 11 OCCaSiOm. Tfiere were
limitatio~ to the accnrav of opgative temperature estimates because they were
dculated h r standing adults. Values for aperative temperahues would have

Un ni least 8646 of the occasions, does used sites where operative
temperature was within the thennoneutral zone. However, it is not known if
the sites used were used for their moderate micro-climate conditions because I
neglected to measure m i a h t e conditions at random sites. It is possible that
the sites used experienced similar miaeclimates to that which was available
randomly, and no selection for sites with thermal cover occurred. Examination
of mown dosure at random and doe sites provided further insight into the
potential use of thermal cover.
Although my sample size was small with only 9 occurrences of rain, there
was no indication that rain affkted habitat selection during summer. I used a 2-
way multi-sarnple median test with rain occurrence and habitat type (n = 81, df =
4, M = 7.6, P P 0.05), d I-tailed 2-sample Mann-Whitney U test of deer model
visibility at 15 m and coyote height with rain occurrence (n = 79, U = 302.5, P >
0105). In both tests, the results were insignificant.
Crown Closure
The quality of thermal cover is commonly indexed by percent crown
damre (Black et al. 1976,l,edcenby et al. 1982, Thomas et d. 1986, Lyon 1987,
Smith and Long 19873. The average crown dosllres of sites used by does and
mdon sites are ckscdxd in Table 10- In comparing a m closufs at sites used
by does with that at random sites with 2 Sfactor ANOVAs, the only significant
main &kt was deer identifimrion (n = 75, Qf = 2, P < 0.007) for bottt runs. There
were no sigdkmt inferactions for the ANUVA wing site type, deer
identification and repradwtiwe status (RI = 75, df = 2, P > 0.05). Ali 3 fa-s
in- with each other for the ANOVA using site type8 deer identification
andactkvitytype(n=75,df=2,P<0.007). Fc~3of11daes,a0~md~~1rewas
greater at bedding s&s than at Ilon-bedding sites, while fbr 5 of 11 does, the

Table 10. Mean percent crown dosure for sites used by does and random 73 sites. Site types: R = random, D = doe, Bf = before parturition, A = after parturition, NBd = non-bedding, and Bd = bedding.
Type of Site n Range (%) Mean (%) SE (%)
DBf 23 0 - 70 23.0 4.8

7 4
aown dosure at bedding sites was less than that at non-bedding sites. For the
remaining 3 does, the crown closure at bedding sites was approximately equal to
that for non-bedding sites. Adequate thermal cover has been defined by different
researchers as different minimum crown closure values (6096, Black et al. 1976;
70%, Thomas et al. 1986, Lycm lW, Smith and Long 1987; 752, Black et ai. 1976,
Leckenby et d. 1982). Crown closure at random sites and at sites used by does
was siWmtly different from &I%, 7046, and 75% (random, t-test., n = 150, t =-
128, P < 0.05; doe, t-test, n = 150, t =-12.8, P < 0.05).
Human Disturbance
There was a large amount of variation in distance to the nearest road as
indicated in Table 11. The only significant main effect for 2 3-way ANOVAs was
doe identification (n = 75, df = 2, P > 0.051, and there were no significant
interactions. The factors used were doe identification, site type and either
activity type or reproductive status. A I h g h does behaved differently from
each other when using sites with respect to distance to the nearest road, there
was no indication that site selection by active or inactive does was influenced
before or after parhuitim by distmce of sites to the nearest mad.
Site choice d d have been influenced by the availability and quality of a
number of essential reso- food, waterI a m dosureI visual cover, and
mer. bxitr:'; 4 si ts to areas of htrman activity couid have
idlueQcced w k t i m of sites by does. Although I expeaed that both food and
visual cover rescrur.-es to be major inawnces on site selectionI tire results of my
analyses shamed o t k w k In fact, none of the basic resources mentioned above

Table 11. Mean distance to the nearest road (m) for sites used by does and 75 random sites. Site types: R = random, D = doe, Bf = before parturition, A = after parturition, NB; = non-bedding, and Bd = Mding.
Site Type n Range (m) Mean (m) SE (m)
R 75 0-2000 271 41
D 81 5-1800 31 6 40
DBf 23 5-1 500 277 73
DA 58 1 0-1 800 332 49
DNBd 25 5-1 500 271 74
DBd 56 10-1 800 336 48
OBfBd 12 30-900 319 76
DABd 44 10-1 800 342 58

76
appeared to strongiy influence diurnal site selection in late spring and summer.
The results contained considerable variation that was largely due to the different
behaviour of individuals.
Forage Resources
I could not test my prediction that deer used sites with less visuai cover
when percent cover d forbs associated with these areas was greater than that
d a t e d with areas with more visual cover because there was no apparent
trade-off between percent cover of forb and visual cover (or crown closlue).
Although open habitats may have had a higher average percent cover of forb
h did furest habitats8 &me was great variation in percent cover of forb within
each habitat type, and no signifbnt relationship betweem visual cover and food
resouces. Other researchers found that forage was more abundant in open
habitats than forest Wtats (Wallmo et al. lm, Collins and Umess 1983). In
Utah, C d h s and Urnes (1983) found a high percent cover of forage in aspen
forests and in open habitas such as dear-cuts. In Washingtun8 forage was most
abundant in stands less than 20-years-dd (Hanley 1984). Atthough I expected to
find a significant relationship between both horizontal visibility and crown
cbmre and percent cover of forbs8 it appears that these site characteristics were
not accurate predicbors of foabs It appears that it was not necessary for does to
between forage resources and aver r s a m e s when they selected sites.
Furthennope, there is no evidence that does were influenced by percent
cover of forbs when dwashg sites within their home ranges. Does were not
using sites with higher percent awer of forbs than that at random sites. There
a r e a t k a s t 3 r r x p l a n a ~ i o r t h i s ~ First,doesmaynothavehad todect
sites with high percent am- of f& because forb were not a iimited resource
a d were readily availaMe in sammer. WhiEe many re-cparckers correlated forb

7 7
availability with habitat seiecticm by deer in summer (Wallmo et d. 1972, Collins
and Umess 1983, W e y 1984, Harestad 1985, b f t et al. 1986, Ordway and
Krausman 1986, l3uduttha et al. 1989, Griffith and Peek 1989), d y Clary and
Larson (1971) found that the distribution of mule deer was not correlated to
forage abundance.
Second, other measures of quality and availability of food may have been
more appropriate. Forb quality, plant phenology and the distniution of
particular forage species may have influenced site selection, but I did not
examine these factors. There may be a more appropriate way of measuring forb
availability although the method I used had been used by other researchers
WEUms et ai. 1980, Harestad 1985). Forb availability also can be measured by
dipping and drying forbs within sample quadrants (Wallmo et al. 1972, Collins
and UIJL~SS 1983) or by cmmhg total number of plants per species or per forage
type in sample areas to calculate density (Grover and Thompson 1986, Urdway
and Krausman 1986)
Third, it may be that percent aver of f o r b is not an important factor in
choos'tng bedding sites which 709& of the sites that I idenW were. Percent
cover of fastlw at bedding sites were diffaent than that at non-bedding sites used
by deer, but rsot significantly di f ferent It is not known if does feed at or near
their Idding sitesI but it w d be ewqpthdy efficient if food resources were
h M d at or near their bedding sites. Several resear- described structural
chmaddst i~ of beddirrg siw lrsed by juvenile ungulates (Walher 1968, Fichter
1974, qrrah 1974, AutenrEsth 9nd Fichter I97?5, Auhmrieth 1976, Bmdey 1977,
Barrett 1981, Tucks and Ga;mer 1983, GaiUard and Ddonae 1989, and Alldredge
et d.1990, but none of these shtdies described the forage resou~as ai bedding
sites. E o a d a n d ~ c o ( r r e r ~ h a v e n o t ~ d ~ f o r b e d d i n g s i t e s
of adult unrgulstes;.

Water Resources
As indicated by significant interactions between individual deer and both
activity type and reproductive status, there was some evidence that site selection
by does was influend by the distribution of water. Behaviour varied betweex?
individuals; some individuals used sites close to water white others did not.
Local distributions of mule deer in deserts of the southwest US. (and possibly
Idaho, Griffith 1983) are determined by the distribution of water (Clark 1953,
Swank 1958, Vdood et al- 1970, Truett 1972, Ordway and Kritusman 1986). This is
a geographic phenomenon a d not the case in Montana (Mackie 1970) and
unlikdy to be true in my study area. The mean distance to water from the
locations where doe mule deer were found in Arizona (1.14 km + 0.03 SE)
(Ordway and Krausman 1986) is much greater than that for the Thompson
PIateau (0.13 km + 0.02 SE)- The same is true for the mean distances to water
from randomly bciited points, (Arizona, 1.28 km + 0.05 SE; Thompson Plateau,
0.14 km + 0.02 SE), which indicates that water souras are more common and
dispersed in my study area than in Arizona's desert. My study area was located
on a plateau -which has many small depressim that collect water and form lakes
and aeeks of various shes.
Some does may have been attracted to areas near water for reasons other
than access to chinking water. Deer do not rreciessarily need access to drinking
water because su&t fa- a n provide enough water to meet their
meOlbalic TeqzLireLztents d deer on mwt ranges ( w d h 1981). A&huugh does
did not s b w a prefeemce for ripariaa habitafs when choosing sites within their
bane rangeI Simpson a d Gyug (1991) who studied the same radio-c:*ed does
ckmmstrated drat dses used summer home ranges with a higher frequency of
~ k a b i t a t s t h a n t h a t w h i c h w a s a ~ e i n t k s t u d y a r e k Greateruseof
a r e e ~ n e a a w a t e r o ~ ~ b a s i n s h a s b e e n o b s e r v e d e i s e w ~ , ~ i ~ n a t u r e

79
of h e relationship betwem habitat choice and proximity to water is unclear
(Patton and Judd 1970, Black et al. 1976). Riparian habitats have several
characteristics that ma). appeal to does: dense visual cover, abundant forbs, and
w i n g or standing water (which provide olfactory cover by masking scents).
Other researchers found that does preferred habitats near water which had an
abundance of succulent forage in summer (Kauffman and Krueger 1!284, Gillen et
al. 1985, Loft et al. 1986, Carson and Peek 1987)- Collins and Umess (1983)
coffefated availability of high quaiity p h t s with high water tables of wet
meadows. Leckenby et a1 (1982) suggested that riparian areas are especially
important during fawn-rearing because all the basic resouras (Le. thermal cover,
security cover, sudent foragef and water) are concentrated within them.
Several researchers have mted the use of water by deer for escape cover
(Barkaiow and Kekr 1950, lhmmut and Taber 1956, Pimlott et al. 1%9, Sweeney
et id- 19n). Does king chased by dogs used a variety of escape patterns, one of
whichwasrmningthn,ughwata. h38of4Qchaseswhezethedeerusedthis
strategy, the dogs lost d&s &ail (Sweeniqr et al. 1971). In summaryf it is unlikely
that access to drinking water was important fix does in my study areaf but it may
have been important for does to have access to riparian habitats which offer a
variety of ~esources~ "Rmse doa that did not wse riparian habitats probably had
Im quality home ranges that did not indude riparian habitats.
v w covtr Individual deer differed in their use of visual aova- Sane deer used sites
with more visual mver than was present at random sites, while other deer used
sites with less visual ama than at random si-. Activity type influenced site
selection by does through an inters- with doe identity, but reproductive
status did not influence dte s e k c t i ~ ~ . This r d t was incunsistent with my

80
alternate hypothesis that does would use sites with dense visual cover when the
consequences of predation were high. I can offer severid possible explanations
for this inconsistency.
First, if the vulnerability of does to predation was low both before and after
parturition, and if the selection of sites by does was not tightly linked to fawn
survival then I expect no difference in the visual cover at the sites used before
and with that at sites used after parturition. There is some evidence that fawn
survival is correlated with use of visual cover at bedding sites (Kjos and
Montgomery 1969, Robinette et al. 1973, Bromley 1978, Dood 19713, Sheehy, 1978,
Smith and Mount 1979, Barrett 1981, Riley 1982, Bowyer and Bleich 1984), but
bedding sites are chosen by fawns not their dams &insdale and Tomich 1953,
Eant 1974). However, a dam may influence choice of bedding sites by its fawns in
potentially 2 ways. First, dams choose a general bedding area for its fawns before
they choose bedding sites &indate and TOlltjch 1953, Lmt 1974). Second, a fawn
may be influenced by its dam's site choices through habitat imprinting.
Pronghorn fawns are found to use bedding sites with simiIar vegetation cover to
that at their birth sites which were chosen by their dams mchter 15)74). The same
~ v i o u r m a ) . o c c u r i n m u l e k *
Second, it is possible that some deer use visual VET while others do not;
deer may use m e r k~ a facultative way. Use of cover may be a behaviour that is
individd or SiaUationspeclfic Same individuals may prefer to use open sites
that aIlav them to be visually vigilant and detect predators quickly. In this
situation, the doe has ample time to decide when to flee and seek visual cover,
whenas other individuals prefer being inconspicuous at sites with visual cover
whae detection by predakcm (and of predators) is more difficult visually. This
maybetruewhether~av~isbeingusedprimarilyforswrurityornot, Ye0
and Peek (I=) suggested that dense vita& cover is not neaessarily better &an

8 1
sparse visual cover. They identified a e b I e tradeoff for prey that use visual
cover: when visual mver is dense then prey may not be detected by predators but
prey may not be able to deteet predators and an ambush may result This trade-
off could explain differences in doe behaviour.
In a study of alarm and flight responses of whitetailed deer in Florida,
Mary (1987) found that deer in dense vegetation fled even when a predator was
at a considerable distance. He hyp&w&d that deer exhibited this behaviour
because of the danger of losing sight of the predator. He further suggested that
deer sensed greater vulnerability to predators in dense vegetation, presmbly
because of the greater danger of surprise attack. Deer in forests seemed to
compensate for this increased danger by fleeing even when a predator was seen
at distances greater than 100 m. Consequently, solitary deer in the forest seemed
mare alert than those in bie pasfam Deer in open habitats could mnsenre
energy by mt attempting @ escape while the predator was at a s& distance.
AUdredge et aL (1991) found that w e n t pronghorns and fawns used denser
s h b cover than what was available at random sites, but the Mest, mat dense
coyer in their study area was avoided-
Third, it a h is possibk that visual over is so abundant in summer,
espedally for an animal that evdved on epm phim Gekt 19741, that most sites
cam meet the deer's cover mpimments- Perhaps selectian fix areas with visual
mis l x r c lu~ at the hame range level r a w than the site heL The random sites
that I used were randw\ within home ranges and therefore not truly randomly
1oc;ated. Sinrpn and Gyug (1991) evaluated summer hone range &ectictn by
mule deer does by comparing use and availability of various habitat types. They
found that does preferred riparian habitats and 2 immature hest types: spruce
alpine fir and mixed decidwus evergreen. Riparian habitats offer g o d visual
cover while immature folrests provide Illoderate visuaI cover <e Chapter 2).

82
While these use and availability comparisons suggest some preference for use of
visual cover at the home range level, this type of analyses have limited value
because of inherent biases against common habitats and biases for rare habitats
( J o h n 1980).
Fourth, visuai a v e r may have a stronger influence on site selection than
what is indicated by m y results because of a lack of power in my analyses. Small
sample sizes (i.e. less than 10 sites per doe) increase the probability that a null
hypothesis is wrongly accepted. Also, it may be that visual cover as I measured i t
is not equivalent to how deer perceive or use security cover. Auditory or
olfactory cover may be structurally different from visual cuver and more
important.
While several researchers have measured visual cover in ungulate
habitats (Nudds 1977, Riley 1982, Canfield et al. 1986, b f t et al. 1987, Smith and
b n g 1987, Griffith and Youtie 1988, Servheen and Lyon 1989, Yeo and Peek 1989,
Simpson and Gyug f992), no other study has compared visual cover at sites used
by deer to that at random sites located within home ranges. It has been observed
that habitat use of an area by deer increases when visual cover is available (Short
et aL 1977, Loft and M d c e 1984, toft et al. 1987). Bladc et al. (1976) suggested that
d t y cover may be required even in absence of predation risk if hLU use of a
habitat is to occur, implying psychological need. Dasmann and Taber (1956)
observed that adult h e s used dense brush during parturition and during the
first weeks of the fawn's life, while bucks fed in the open at this time. Yeo and
Pa& (1989) measwed visual cover at random sites within core areas and in non-
care areas within home ranges of Hack-tailed deer. Their d t s indicate that
lrisual cover was anfy in one habitat type when t h y ampared core
areas with mnum areas Visual mer was 16% greater within axe areas than
w i t h i n I u m ~ a r e a s o f ~ h r u b d ~ t a t

83
Aithough several researchers related LW of visual cover by fawns to fawn
survival by inferring that use of visual cover reduces predation risk and
vulnerability (Kjos and Montgomery 1969, Robinette et al. 1973, Brornley 1978,
Dood 1978, Sheehy, 1978, Smith and LeCount 1979, Barrett 1981, Riley 1982,
Bowyer and Bleich 1984), no one has attempted to correlate the use of visual
cover by adults to adult survival. Consequently, I am not convinced that does
are influenced by visual cover in their site selection within their home ranges.
Perhaps, visual cover is vital to fawns at the site selection level and important to
does at the home range selection ievel. If this is true, habitat managers should
focus on the understory component of visual cover which is important for fawn
bedding sites rather than conifer overstory. S h b s are a significant component
of visual cover for adult ungulates as well (see Chapte~ 2).
Universal use of the definition of adequate visual cover proposed by
Th- et al. (1979) may be inadequate and inappropriate. This definition is
inadequate because it gives no idonnation regarding quality of visual cover. It
only indicates whether visual cover is present or not Ah, 1 find that it is
inappropriate in my study area because the mean minimum distance for 10%
visibility of a standing dea IIbOdel at sites used by does was 24 m which is
sigdhntly less 61 m. Simpson and Gyug (1991) also found that the minimum
distancefbr 10% visibilittybfastandingperson) wasIessthan30mat78% of all
sites used by does. Yeo d Peek O W ) suggest that W% standing visual cover at
1ommaybeoptimaL LedEenbyetaL(1982)usedasimilarcriteriatoThomaset
aL (1979 that was Npposedly mare ampropriabe for their study area. in the Great
Basin Theyusedasightdis~of45mratherthan61 mandappliedittothe
view04abeddeddgerntherthanastandingW. Isuggestthatvisualwver
requitemen6 are species and geographicalIy specific Adapte visual cover as
-by ThomasetaL ~1979)maybevalidforeIkh theBlueMountainsof

8 4
Oregon and Washington, but not for deer in areas of denser vegetation like parts
of Alaska and Brtish Columbia.
Micro-Climate Conditions and Crown Closure
Although there is no conclusive evidence that does used visual cover, I
have examined the possibility that visual cover was used as thermal cover rather
than security cover. Peek et al. (1982) proposed that thermal cover is preferred
but not required by mule deer in summer when heat is extreme. kkenby et al.
(1982) noticed that forest stads that did not provide thermal or security mver
were rarely used during the day. Several investigators observed that habitat
selection can be influenced by micro-chate conditions (Dasmam 1954, Darling
1964, Loveless 1964, Staines 1976, Boyd 1981, Collins and Umess 1983, Brindley et
aL 1989)- Miao-climate conditions at most sites used by does were moderate in
terms of air temperature, wind, short-wave radiation, and rainfall which may
have resulted from moderating effects of thermal cover or may have indicative
of the climate of the general area.
There appears to be less than adequate thermal cover (as indexed by crown
closure> at both random sites and doe sites. There are 3 possible explanations for
tfre moderate conditions at sites used by does when crown dosure was low. First,
the climate in the general area was moderate and selection of sites with dense
a m ~~ was ll~~~ecessary. Second, sparse crown closures were sufficient
to provide t h d coyer. There is some dis<rrepancy as to which minimum
value of crow clamre provides adequate thermal cover in summer. Several
authors have defined adequate thermal cover as coniferous forests with an
average cravn ctusure exceding 70% and t n s at least 12 m Kgh (Wmas et al.
1986, Lyon 1987, Smith and Lcmg IP87) while other re~ea~chers used a greater
75% crown dosure dterhn for trees or shrubs taller than IS m (Black et ale

8 5
1976, Ledamby et at 1982). Black et aL (1976) also stated that for forest stands at
the sapling stage or older, a crown dasure of 60% provided adequate thermal
cover in summer. Third, understory vegetation provided ~~1 cover which
moderated the micrQ-climates at sites. Taber (1%1) acknowledged that
understory vegetation provides thermal cover. Dense aown closures do not
seem especially necessary to maintain high populations of mule deer because
topography and lower vegetation can act as thermal cover (Peek et al. 1982). I am
unable to determine whether thanaI cover was required by does in my study
area during summer, but I would suggest that the definition c' thermal cover
should not be limited to a standard aown closure value. The influence of
understory vegetation on rniao-climates should be widely acknowledged.
Jnsect Cover
It has been noticed by several researchers that harassment by insects can
influence habitat selection by ungulates (Darling 1964, Collins and Urness 1982,
Jakimchuk et al. 1987, B ~ d l e y et al. 1989). When harassed by insects, reindeer
and red deer seek open, efevated, windy places (Darling 19641, while feral goats
retreat to shady areas (Brindey et al. 1989). I did not measure levels of insect
harassment? but of sites used by does only 21% were in open habitats. Although
it is p s i b l e that insect harassment influend site selection by does, it is
unlikeiy that it was a major influence.
Disturbance
The distance from sites to nearest roads did not influence site selection.
Because the density of roads in my study area was relatively high, does may not
have had the opportunity to avoid roads. Deer may use visual cover to avoid
disturbane rather than avoiding habitats adjacent to roads. Habitat selection

86
strategies of elk appear to he based primarily on the availability of fwd and
hwnan bisturkce Peek et aL 1982). V d cover seems to be a requirement for
dk in the presence of human disturbance (Peek et al. 1982). Visual cover affects
responses of elk to hunting and road activities Perry and Overly 1976, Basile and
Lonner 1979, Irwin and Peek lm, Lyon 1979). Three researchers reported use of
visual cover by d- on north dopes during hunting season (Wilkins 1957,
lavaas 1958, and Madtie 1970), although they did not indicate whether deer used
visual cover outside of hunting season.
CONCLUSIONS
Density of visual cover and minimum distance to water influenced site
selection by doe muie deer, but perent cover of forb did not Does were not
confronted with a choice between food and visual cover (or aown closure)
because there was no apparent trade-off between these resources. The influence
of visual cover and water availability on site selection was ambiguous because of
variation in the behaviour between individuals. Although does did not show a
preference for riparian habitats at the site selection level of habitat choice,
Sipson and G p g (1W1) showed that does d riparian habitats more
frequently than what would be predicted from the availability of riparian habitats
when choosing their summer h e ranges. I have offered a number of
explanations for variation in doe behaviour with respect to visual cover. 1 was
unable to determine if either visual cover or crown closure were wed as thermal
cover- It is possible that does required insect cover, but I did not measure levels
of insect harassment Does did not appear to be influenced by disturbance from
roads in choosing sites.
I question universal use of the definition of adequate visual cover

87
proposed by Thomas et aL (19'7%. I find that 61 m is too long to be appropriate as
a standard sight dis-ce value for my study area. Perhaps it is inappropriate to
describe security mver which is functionally and structurally complex with a
single sight distance vatue. I suggest that more attention should be focussed on
the value of undmtory vegetation as visual cover for 2 reasons. First, because it
is more likely that fawns require visual cover than do adults and because fawns
primarily use understory structure for visual cover, understory vegetation
should be managed for its value as visual cover. Second, shrubs are structurally
an important component of visual cover for adults (see Chapter 2).

(zmwmR4
CONCLUSIONS AhTD MA3TAGEMENT RECOMMENDATIONS
In evaluating the use of 3 apparatuses to estimate visual cover (profile
bcrard, profile pole and deer model), I conclude that the profile pole is the most
versatile and convenient apparatus for measuring visual cover. The following
set of procedures for detecting differences in visual cover among habitats. Visual
cover can be measured at the observer's natural height because there is no
advantage to kneeling at coyote height For mule deer in my study area, the
optimal distance for measuring visibility is 15 m. The faurth (0.75-1.00 m) and
fifth section (1.00-1.25 m) are the best pole sections to measure visibility at for
conditions encountered in my study area for adult mule deer. I recommend that
lower sections of the pole be d when evaluating habitat use by fawns. There
is no significant advantage for the observer to stand upslope, downslope or
aaoss-slope from the apparatus when estimating visual cover.
Unspad sapling forests provided the densest visual cover while open
habitats provided the sparsest visual cover. Shrub, spaced sap;ng, immature
forest and mature forest habitats were intermediate in density of visual cover.
The important vegetation components of visual cover were: crown closure,
percent cover of conifer trees in the top height strata, percent cover of conifers in
the shrub layer, total percent cover of non-conifers in the shrub layer, percent
cover of dedduous trees in the middle height strata, diameter (dbh) of deciduous
trees of the middle height strata, and percent cover of decid~~)us trees in the
bottam height strata Mod& of visual cover use only characteristics of the
mnifmus overstory and ignore the contribution of deciduous and understory
vegetation to visual cover. This amision should be rectified.
The importance of Security mer to wildlife has long been acknowledged

89
1933, Egler 1m, 'I'bmas et d. lm, Leckenby et al. 1982. DeByle 1985). I
cannot claim that does required visual cover, although the distribution of visual
cover did influence site selection by does differently. Likewise, Wallmo and
Schoen (1980) found that while the structure of coniferous forests influenced
habitat selection, there was no evidence indicating that deer required forests (as
thermal and visual cover) for survival. Peek et al. (1982) suggested that thermal
cover is preferred but not required by mule deer in summer when heat is
extreme and when forage becomes desiccated. However, visual cover may be
required in the presence of human activity (Peek et al. 1982).
Perhaps, the distribution of visual cover influenced does more in their
selection of summer home ranges rather than in their se1don of sites within
summer home ranges. 5mm.I segregation and differential habitat use by
ungulates in summer is proposed to be a result of antipredator strategies by
maternal females (Skigas and Flinders 1980, Edwards 1983, Jakimchuk et al.
1987, Festa-Bianchet 1988). In these 3 studies, security cover was not visual cover
provided by vegetation but predator-free habitats in isolated areas. The
importance of visual cover in site selection is probably greater for young (that are
"hider" types like mule deer (Geist 1981)) than for adult ungulates. Several
researchers have correlated survival of young with the distribution and structure
of understory vegetation (Kjus and Montgomery 1%9, Robinettc et al. 1973,
Bromley 1978, Dood 1978, Sheehy, 1978, Smith and M o u n t 1979, Barrett 1981,
Riley 1982, Bowyer and BMch 1984). Because understory vegetation influences
survival of young and because it is an important component of visual cover for
adult deer, understory vegetation should be included in management plans and
for models of visual cover.
The universal application of the definition of adequate visual cover
pmposed by Thomas et aL (1979) seems inappropriate. Sixty-one meters is too

90 - . - long for dscmnmathn among habitats, a distance of 15 m is more appropriate
for this function for mule deer in my study area The mean minimum distance
for 10% vki'biility at sites used by dws was significantly less than 61 rn. It may be
neceSSafy to empirically redefine adequate visual cover for different species and
ecosystems like Leckenby et al. (1982) did for their mule deer study in the Great
Basin. However, security cover is functionally and structurally complex and it is
unlikeIy that a single sight distance value can describe security or visual aver
adequateIy.

Aildredge, A. W., R D. Deblinger, and J. Peterson. 1991. Birth and fawn bed site selection by pronghorns in a sagebrushsteppe community. J. Wildl.
Manage. 55222-227,
Autenrieth, R E. 1976. A study of birth sites selected by pronghorn does and the M sites of fawns. Pmc Bienn. Pronghorn Antdope Workshop 2127-134.
, and E. Fichter. 1m. On the behavior and socialization of pronghorn fawns. Wildl. Monogr. 42 llpp.
Barblow, F. S., Jr., and W. E. H e r . 1950. Escape behavior of the white-tailed deer. J. Wildl. Manage 1k246-247.
Barrett, M- W. 1981. Environmental characteristics and functional significance of pronghorn fawn bedding sites in Alberta. J. Wild. Manage 4S:l2O-l3l.
Basile, J. V., and T. N. Lomer. 1979. Vehicle restrictions influence elk and hunter distribution. Mont. J. For. 77355-159.
Beckr, D. Q, F. L. Bu.mdl, D. W. Janz, J. B. Nyberg, and E. L. Richardson. 1990. Techniques for managing habitat. Pages 133-195 in J. B. Nyberg and D. W. Janz, tech- eds. Deer and elk habitats in coastal forests of southern British
Columbia B.C. Minist For. Res. Branch Spec. Rep. Ser. 5. 310pp.
Black, H., R ScherzInger, and J. W. Thomas. 1976. Relationships of Rocky Mountain Elk and Rocky Mountain mule deer habitat to timber
management in the Blue Mountins of Oregon and Washington. Fages 11-31 in J. M. Peek and S. R Hieb, eds. Proc Elk-logging-roads Symp., Vniv. Idaho For. Wildl Range Exp. Stn.
BodPntha, T. S, j. M. Peek, and J. L Lam. 1989. Mule deer habitat use related to succession in a bcfigrass uxnmunity. J. Wid. Manage. 53314319-

Bowyer, R T-, and V. C. BIe id~ 1 9 4 . Weds of cattle grazing on selected habitats of southern mule deer. Calif. Fish Game 7&240-247.
Boyd, 1. L. 1981. Population changes and distribution of a herd of feral goats on Rhum, hner Hebrides 1960-78. J. Zool., Lond. 193287-304.
Brindley, E. L., D. J. Bullodr, and F. Maisels. 1989. Effects of rain and fly harassment on the feeding behavim of free-ranging feral goats. Appl. Anim. Behav. Sci. 2431-41.
B. C. Ministry of Fofests. 1!%8. Biogedimatic zones of British Columbia 1988. Map B.C. Minist For., Victoria, B.C.
Bromley, P. T. 1977. Aspects of the behavioural ecology and sociobiology of the pronghorn (Anf i lmup ameriuzna ). Ph.D. Thesis, Univ. Calgary, Alta.
370.8~.
. 1978. Ultimate factors, behavis-w and fawn recruitment. Proc Bienn. Pronghorn Antdope Workshop &449-471.
Bunneil, F. L., and D. J. Vales- 1990. Comparison of methods for estimating forest overstory cover: Me~ences among techniques. Can. J. For. Res. U3:IOI-107.
Canfield, J. E., L J. Lyan, and J. M. Hillis. 1986. The influence of viewing angle on elk hiding cover in young timber stands. U.S.D.A. For. Serv. Kes. Pap. INT-371. 5pp.
Carson, R G., and J M. Peek 1987. Mule deer habitat selection patterns in northcentral Washington. J. Wddl. Manage. 51:46-51.
Clark, E. D. 1953- A study of the behavior and movements of the Tucson Mountain mule deer. MS. Thesis, Univ. Arizona, Tucson. lllpp.
Ctary, W. P-, and F. R Latson. l97l. Elk and deer use ars related to food

93 resources in Arizona ponderosa pine. U.S.D.A. For. Serv. Res. Note M- 202 4pp.
Clover, M- R 2954. Single gate deer trap. Calif. Fish Game 40:367-373.
Collins, W. B., and P. J. Urnen. 19&2 Mule deer and elk responses to horsefly attacks. Northwest Sci. 56:299-302
A and - . 1983. Feeding behavior and habitat selectim of mule deer and eik on northern Utah summer range. J. Wild. Manage. 47646-663.
Darling, F. F. 1964. A herd of red deer: a study in the animal behaviour. Fourth ed- Oxford University Press, London.
Dasmann, R F. 1954. Emlogy and social behavior of a population of the Columbian black-tailed deer in California. Ph.D. Thesis, Univ. California, Berkeley. 155pp.
, and R D. Taber. 19%. Behavior of Colunnbian black-tailed deer with reference to population ecology. J. Mammal. 32143-164.
DeByle, N. V. 1985. Wildlife. Pages 135-152 in N. V. DeByle and R. P. Winokur, eds. Aspen: -logy and management in the western United States.
U.S.D.A. Fbr. Sem. Gen. Tech. Rep. RM-119.
Demarchi, M. W. personal communication. Faculty d Forestry, Univ. British Columbia.
Dood, k R 1978- Summer movements, habitat use, and mortality of mule deer fawns in the Missouri River Breaks, Montana. Mont. Dep. Fish Game Fed. Aid Job F d Rep. Mont. Proj. W-120-R-8,9, Job No. 9 . 5 5 ~ ~ .
Egler, F- E. 1977- Tlw n a v e of vegetation, its management and mismanagement: an introduction to vegetation science. Frank E. Egier, Norfolk, Conn. 527pp-

Elton, C. 1939. On the nature of cover. J. WW. Manage. 3532-338.
Festa-Bianm, M. 1988. SeasonaI range dxtion in bighorn sheep conflicts between forage Quaifity, forage quantity and predator avoidance. Owologia
?!kts&o-586.
Fichter, E. 1974. On the bedding bebviou~ of p~cfnghorn fawns. Pages 352-355 in V. GeiSt and F- R Wafther, eds. The beha- of ungulates and its relation to management. Vot I. IUCN New Series Publ. 24. Morges, Switzerland.
Gaiilard, J. M., and D. Delonne- 1989. Seleebn des sites de repas par les W e s
chevreuils (Coprwhw urprwk 1. Acta Oecologica 20:421-418.
Geist, V. 1974. On the relationship of social dutim and d o g y in ungulates. Am. &I. 14905-220-
- 1981. Behavior adaptive strategies in mule deer. Pages 157-223 in 0. C. W h o , ed- Mule deer and black-tailed deer of North America. Univ. Nebraska Press, Lincoln. 605pp.
Gillen, R L, W. C. Krueger, and R F. Miller. 1985. Cattle use of riparian meadows in the Hue Mountains of northeastern Oregon. J. Range Maltage. 38:sm.
Griffith, B., and J- M. Peek 1989. Mule deer use of sera1 stage and habitat type in bittttrbntsh communi~e- J. WddL Manage. 53636-642.
, and B. A, Youtie 1 s . Two ckvices for estimating foliage density and deer hiding cover. WM. SIC. BuIl- 16-206-210.
Griffith, D. S. 1983. E t ~ ) k @ d ~ ~ C S of mule k. aaters of the Moon

9 5 National Monument, Idaho. Univ. Idaho Coop. Park Stud. Unit, Moscow.
w?'. Grover, K E. and M. J- TRompson- 1986. Factors influencing spring feeding site
selection by elk in the Elkhum Mountains, Montana. J. Wildl. Manage. 50.466-470.
Gysel, L W., and L. J. Lyon. 1980. Habitat analysis and evaluation. Pages 305-327 in S. C. S d ~ d t z , ed. management techniques. Fourth ed. The Widlife Sodety, Washington, D.C.
Hanley, T. A. 1984. Habitat patches and tfi& selection by wapiti and black-tailed deer in a coastal montane coniferous forest. J. Appl. E d . 21:423-436.
Harestad, A. S. 1984. Seasonal movements of black-taiIed deer. Pages 403-409 in W. R Meehan, T. R Merrell, Jr. and T. A. Hanley, eds. Proc. Symp. on fish and wildlife relationships in old-gmwth forest. Am. Inst. Fish. Res. Biol.
- 1985. Habitat w by black-tailed deer on northern Vancouver Island. J. Wildl. Manage. 49946-950.
Irwin, L L, and J. M. Peek 1979. Relationship between road closures and elk behavior in northern W o . Pages 199-204 in M. S. Boy- and L. D. Hayden-Wing, eds. North American eik: ecology, behavior and management. Univ. Wyoming, Warnie.
Jakimchuk, R D., S. H. F&gman, and L. G. Sopuck. 1987. Differential habitat use and sexual segregation in the Central Arctic caribou herd. Can. J. Zool. 65-SM-541.
Jdmson, D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 6 : M .
Kauffman, J. B., and W. C. Krueger. 1984. Livestock impacts on riparian easystems and &reamside management impiications ... a review. J. Range Minage 3E430-438.

K,&, C. G-, and G. G. M<mtpmery, 1%9. Daytime locations and bedsites of whitetailed deer fawns. Trans. Illinois State Acad. Sci. 621 17-119.
Krasowski, T. P., and T. D. Nudds. 1986. Mimhabitat structure of nest sites and nesting success of diving ducks. J. Wddl. Manage 9203-208.
Kr*, J- R, and N- B. Davies, eds. 1984- Second ed. Behavioural Ecology. Btaikwd Scientific, Oxford. 493pp.
Kufeld, R C., 0. C. W d h o 8 and C. Fetid- 1973. Foods of the Rocicy Mountain mule deer- US.D.A For. Serv. Res. Pap. RM-Ill. 31pp.
LaGay, IC E. 1987. The influence of habitat and group characteristics on the alarm and flight response of white-tailed deer. Anim. Behav. 3520-25.
Leckenby, D. A., D. P- Sheehy, C H. Nellis, R J. Scherzinger, L D. Luman, W. Elmore, J. C. Lemos, L Doughty, and C. E. Trainer. 1982 Mule deer. Wildlife habitats in managed rangelands - the Great Basin of southeast Oregon. U.S.D.A. For. Sem. Cen. Tech. Rep. PNW-139. 40pp.
Lent, P. C. 1974. Mother-infant relationships in ungulates. Pages 14-53 in V. Geist and F. R Walther, eds. The Maviour of ungulates and its relation to management Vol. 1. rUCN New Series Publ. 24. Morges, Switzerland.
-Id, A. 1933. Game managemt. Charles Scribner's Sons, New York, NY.
4 1 ~ ~ -
Linsdale, j. M=, and P. Q. T&ch 1953. A herd of mule deer: a record of
obseFvatioxs made on the Hastings Natural History Reservation. Univ. Calif. Press, l3erkeley. •̃ 67pp.
M, E R, and J. W- Mmke. 1984. DeeP use and habitat characteristics of transmission-he corridors in a Dou@as-fu forest. J. Wildl. Manage. 4&1311-1316.

I and J. 6. Kie 1%. fnteractkrn of cattle and deer an mountain rangeland. Cdif. AAgric 406-9-
, , . and R C- Bertraa 1987. Influence of cattle stocking rate on the structural profile of deer hiding cover. J. Wild. Manage. 51:655-661.
Lovaas, A. L 19%. Mule deer food habits and range use, Little Belt Mountains, Montana. J. WiU. Manage. 22275-283.
bveless, C. M. 1%4, Some relationships between wintering mule deer and the physical environment. Trans. North Am. Wildl. Nat. Resour. Conf. 29415-431,
Lyon, L J. 1979. Habitat &&v- for elk as influenced by roads and cover. J. For. 77:658-66U.
- 1987. Effects of thinning smallstem lodgepole pine stands on big game habitat. Pages 162-165 in R L. Garger, mpiler. Workshop Proc. Management of snall sfmn stands of ladgepole pine. U.S.D.A For. Serv. Gen. Tech. Rep. W-237.
MacArthur, R H, and J. W. MacArthu~~ 1961. On bird species diversity. Ecology 42594-598-
M'Qoskey, R T., and B. Fieldwick 1975. Ecological separation of sympatric rodents (Permyscus and Micro&). J. Mammal. 56:119-129.
MacHutchon, A G. 1988- Risk af predation and use of security cover and vigilance cover by foraging ~ I u m b i a n ground squirrels. U S Thesis, Simon Fraser UNV, Burnaby, B-C. 90pp.
, and A- S- Harestad- 1990. Vigilance behaviour and use of rocks by CoIumbian ground squirrels. Can. J, Zool. 6tk142&1432
McTague, J. P., and D. R Patton. 1989. Stamd density index and its application in

A&&, R J. 1970. Range ecology and relations of mule deer, elk, and cattle in the Missouri River Breaks, Montana. Wild. Monogr. 2 0 . 7 9 ~ ~ .
Mautz, W. W. 1978. Sledding on a bushy hillside: the fat cycle in deer. Wildl. Sac Bull. 6:8$-90.
Mob, C. 0. 1947. Table of equivalent populations of North American small mammals. Am, Midl. Nat. 37223-249-
Nudds, T. D. 19n. Quantifying the vegetative structure of wildlife cover. Wildl. Sac. Bull. 5113-117,
Ordway, L L., and P. R Kra- 1986. Habitat use by desert mule deer. J. WildI. Manage. W677-683.
Parkerf K. L, and M. P. G i g h a m . 1986. Estimating thermal environments for mule deer (Odclroiieus hemionus hemisnus ). B.C. Minist. For. Lands I&. B m c h WHR-24. 63pp.
. and C T. Wbifls. 1384. Thermoregulation in mule deer and elk. Can. J. Zeal. 621409-1422
Palton, D. R, and B- I- Jud& 1970. The roIe of wet meadows as wildlife habitat in the southwest J. Range Martage 23-272-275.
Peek, J. M, M D. ScottI L J- Ndsa, DD J. Piercef and L L Irwin. 1982. Role of amer in habitat miimagematt for big game in Northwestern United States. Trans, Nc&l Am, wild Conf- 47-372
P q , C, and R OvetEy- 1976. Impact of roads on big game distribution in Eportions of the Blue Mountains of Washington. Pages 62-68 in J. M. Peek and S. R Hieb, eds. Roc Elk-magging--roads Symp, Univ. Idaho For., WilaL, Range Exp. Stn,

Pimlott, D. H., J. A. Shannon, and G. B. Kolenosky. 1%9. The ecology of the
timber wolf in Algonquin Provincial Park Ontario Dep. Lands For. Res.
Rep. (Wildl.) 8 7 . 9 2 ~ ~ .
Pyrah, D. 1974. Antelope fawn bedding cover selection in central Montana.
Prm. Bienn. Pronghorn Antelope Workshop 6:113-121.
Recher, H. F. 1%9. Bird species diversity and habitat diversity in Australia and
North America. Am. Nat. 103:75-80.
Riley, S. J. 1982 Survival and behavior of radio-collared mule deer fawns during summers, 1978-1980, in the Missouri River breaks, Montana. M.S.
Thesis, Montana State Univ., Bozeman. S4pp.
Robinette, W. L., C. H. Baer, R E. Pillmore, and C. E. Knittle. 1973. Effects of nutritional change on captive mule deer. J. Wildl. Manage. 37:312-326.
Rawuweig, M. L., and J. Winakur. 1969. Population ecology of desert rodent communities: habitats and environemental complexity. Ecology 50:558- 572.
Schwartz, C. C., and N. T. Hobbs. 1985. Forage and range evaluation. Pages 25-51
in R J. Hudson and R G. White, eds. Bioenergetics of Wild Herbivores. CRC Press, Boca Raton, FL.
Servheen, G., and L. J. Lyon. 1989. Habitat use by woodland caribou in the Sellcirk Mountains. J. WMl. Manage. 53:230-237.
Sheehy, D. P. 1978. Characteristics of shrubland habitat associated with mule deer fawns at birth and during early life in southeastern Oregon. Oreg. Dep. Fish Wad. Info. Rep. Ser. Wildl. No. 78-1. 31pp.
Short, EL L, W. Evans, and E. L h k e r . 1977. The use of natural and modified pinyon pinejuniper woodlands by deer and elk. J. Wildl. Manage. 41543-
559.

100 Simpson, K. 1988. Okanagan Connector Highway ungulate telemetry and
inventory. Consultant's report to the Minist. Environ. Parks. 25pp.
, and L. W. G p g . 1991. Environmental assessment of the Okanagan Connector Freway. Part iI. Pages 1-60 in Keystone Biersearch. Movements, habitats and status of moose and mule deer populations influenced by the Okanagan Comector Highway. Consultant's report to B.C. Minist. Highways and Minist. Environ.
Smith, F. W., and J. N. Long. 1987. Elk hiding and thermal cover guidelines in the context of lodgepole pine stand density. Western J. Appl. For. 2:6-10.
Smith, R H., and A. LeCount. 1979. Some factors affecting survival of desert mule deer fawns. f. Wildl. Manage. 43657-665.
Stains, B. W. 1976. The use of natural shelter by red deer in relation to weather in northeast Scotland. J. Zool., Land. 180:l-8.
Steigers, W. D. and J. T. Flinders. 1980. Mortality and movements of mule deer fawns in Washington. J. Wildl. Manage. 44381-388.
Strickland, D. 1975. Mule deer in the Medicine Bow Mountains, southeastern Wyoming. Wyo. Game Fish Dep. Wild. Tech. Rep. 2. 103pp.
Swank, W. G. 1958. The mule deer in Arizona chaparral and an analysis of other important deer herds: a research and management study. Ariz. Game Fish 9)ep. Wildif Bull. 3.109pp.
Sweeney, J. R, R L. Marchinton, and f. M. Sweeney. 19n. Responses of radio- monitored whitetailed deer chased by hunting dogs. J. Wildl. Manage. 35707-7l6.
Taber, R D. 1961. The black-tailed deer, a review of ecology and management. Terre Vie 2221-245.
Thomas, J. W., H. Black, Jr., It. J. Scherzinger, and R J. Pedersen. f 979. Deer and

101 elk. Pages 104-127 in J. W. Thomas, ed. Wildlife habiiats in managed forests - the Blue Mountains of Oregon and Washington. U.S.D.A. For. %IT. Agric. Handb. 9%. 512pp.
. D. A. W e n b y , L. J. Erickson, S- R Thomas, D. L. hacson, and R. J.
Murray. 19%. Wildlife habitat by design: National forests in the Blue Mountains of Oregon and Washington. Trans North Am. Wildl. Nat. Ftes Conf. 51:203-214.
Trainer, C. E. 1975. Direct causes of mortality in mule deer fawns during summer m d winter periods on Stem Mountain, Oregon. West. Assoc. State Fish Game Comm. Proc. 55163-170.
Truett, J. C. 1972. Ecology of the desert mule deer (Odocoifeus hemionus crooki ) in southeastern Arizona Ph.D. Thesisf Univ. Arizona, Tucson. 64pp.
Tucker, R D., and G. W. Garner. 1983. Habitat selection and vegetational characteristics of antelope fawn bedsites in west Texas. J. Range Manage. 36A10-113.
Wallmo, 0. C. 1981. Mule and black-tailed deer distribution and habitats. Pages 1-25 in 0. C. Wallmo, ed. Mule and black-tailed deer of North America. Univ. Nebraska Press, Lincoln. 605pp.
. W. L. Regelin, and D. W. Reichglrt. 1972. Forage use by mule deer relative to logging in Colorado. J. Wildl. Manage. 36:1025-1033.
, and J. W. Schoert. 1980. Response of deer to secondary forest succession in southeast Alaska. For. S' 26:44&462.
WaIther, F. 1968. Verhalt en der Gazellen. Die Neue Brehm Bucherei, no. 373.
Wittenberg-Lutherstadt A. Ziemsen Verlag. 144pp.
Wells, U C., and P. N. Lehner. 1978. The relative importance of the distance senses in coyote predatory behavior. Anim. Behav. 26251-258.

102 White, M., F. F. Knowlton, and W. C. Glamer. 1972. Effect of dam-newborn
fawn behavior on captive and mortality. J. Wildl. Manage. 36:897-906.
Wight, H. M. 1939- Field and laboratory technic in wildlife management. Univ. Michigan Press, Ann Arbor. 107pp.
Wilkins, B. T. 1957. Range use food habits and agricultural relationships of the mule deer, Bridger Mountains, Montana. J. Wildl. Manage. 21:159-169.
Wiilms, W. A., A. McLean, R Tucker, and R Ritcey. 1980. Deer and cattle diets on summer range in British Columbia. J. Range Manage. 33:55-59.
Wood, J. E., T. S. Bickle, W. Evans, J. C. Germany, and V. W. Howard, Jr. 1970. The Fort Stanton mule der herd: some ecological and life history characteristics with special emphasis on the use of water. New Mexico State Univ. A@C Exp. Stn. BdI. 567. 32pp.
Yeo, J. J., and J. M. Peek. 1989. Habitat characteristics selected by Sitka black-tailed deer in logged forests of southeastern Alaska. unpubl. report to U.S.D.A. For. Serv., Ketchikan, AM. 48pp.















![[Training] Claves Marketing Digital de ITStrap MTY](https://img.pdfslide.us/doc/110x75/557c2be7d8b42aa77f8b4f36/training-claves-marketing-digital-de-itstrap-mty.jpg)












