Embed Size (px)
Citation preview

This article was downloaded by: [University of Alberta]On: 04 October 2014, At: 03:14Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Journal of InterdisciplinaryCycle ResearchPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/nbrr19
Transplantation of circadianpacemaker in the housecricket, Acheta domesticus L.Bronislaw Cymborowski aa Department of Invertebrate Physiology , WarsawUniversity , 93 Zwirki i Wigury, Warszawa, PL‐02–089,PolandPublished online: 22 Sep 2008.
To cite this article: Bronislaw Cymborowski (1981) Transplantation of circadianpacemaker in the house cricket, Acheta domesticus L., Journal of InterdisciplinaryCycle Research, 12:2, 133-140
To link to this article: http://dx.doi.org/10.1080/09291018109359734
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,

sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

J. interdiscipl. Cycle Res., 1981, vol. 12, number 2, pp. 133-140.
Transplantation of Circadian Pacemaker in the House Cricket,Acheta domesticus L.
by
Bronislaw Cymborowski*
ABSTRACT
Destruction of neurosecretory cells in the pars intercerebralis of the brain causesthe loss of locomotor activity rhythms and creates hyperactivity. When thebrain is taken from a rhythmic cricket having both short-period activityrhythm (S-PAR) or long-period activity rhythm (L-PAR) and is implanted intothe abdomen of such an arrhythmic recipient, it produces a characteristiclocomotor activity rhythm of the host cricket.
The implanted brain has an inhibitory effect on the locomotor activity of ahyperactive and arrhythmic recipient.
INTRODUCTION
The most intensive studies in physiology of insects' circadian rhythms werecarried out in the search for the cells generating the circadian behaviour (forreview see Saunders, 1976). Early experiments have demonstrated that singleeucaryotic cells (Bruce and Pittendrigh, 1956; Hastings and Sweeney, 1958;Harker, 1960) and insect tissues (Cymborowski and D.utkowski, 1968, 1969,1970; Cymborowski et al., 1970; Dutkowski and Cymborowski, 1970;
* Department of Invertebrate Physiology Warsaw University, 93 Zwirki i Wigury, PL-02-089Warszawa, Poland.Paper presented at the 9th International Inderdisciplinary Cycle Research Symposium, Trier,Fed. Rep. of Germany, 6-11 July 1980.Abstracting keywords: Circadian rhythms, circadian pacemaker transplantation, Achetadomesticus, neurosecretory cells, pars intercerebralis, neurohormones.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

Cymborowski, 1970b, 1973a), can exhibit rhythmicity. This suggested that anumber of centers within an organism itself can provide cues for the circadianrhythmicity.
The problem arises now to what extent the rhythmicity observed in a givenfunction, like for example in insect locomotor activity, is exclusively under thecontrol of a particular group of cells which can be termed "biological clock". Anumber of experiments were performed on insects in this respect. It has becomeobvious that the central nervous and endocrine systems are the main source ofcircadian rhythmicity (Brady, 1969; Cymborowski, 1970a, Truman, 1972;Roberts, 1974; Sokolove, 1975; Ciemior et al., 1977). In the cockroach, forexample, the main pacemaker controlling the locomotor activity rhythmsseems to be located in the optic lobes (Roberts, 1974; Sokolove, 1975), but incrickets it appears to reside in median neurosecretory cells of the brain(Cymborowski, 1970a,b; Cymborowski, 1973b).
What is the nature of the coupling between the pacemaker and the motorsystem of insects? This question has become recently of major importance inthe search for the controlling mechanism of the insect locomotor activityrhythms. Harker (1955) was the first to report that this activity could behormonally controlled. On the basis of transplant experiments she suggestedthat the subesophageal ganglion was the source of locomotor activity rhythm.Then, parabi'osis experiments confirmed hormonal involvement in thelocomotor activity rhythms of crickets and cockroaches (Cymborowski andBrady, 1972). Handler and Konopka (1979) have recently demonstrated in theirelegant experiments a humoral component in the control of Drosophilaactivity rhythms. They found that a short-period mutant brain can produce ashort-period activity rhythm when implanted into the abdomen of agenetically arrhythmic host.
In this paper I will present further evidence that a brain isolated fromrhythmic cricket can produce its characteristic activity rhythm when implantedto arrhythmic host.
MATERIAL AND METHODS
Culture conditions and activity recordings
The studies were carried out on male house crickets (Acheta domesticus L.)about a week after adult ecdysis. The insects were cultured under alternatingconditions of light and darkness (LD 12:12) at 24°C. The dark period began at18.00. Food and water were available ad libitum.
Locomotor activity was recorded using two types of actographs. One of themwas a rocking-box actograph, which consisted in essence of an endless circular
134
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

corridor 30 cm in circumference. The corridor was pivoted across one diameterand operated two mercury-pot switches. Six such actographs were used. Theywere either coupled to a six-channel event pen recorder or insect's movementswere registered on a smoked revolving drum (Cymborowski, 1972).
The second type of actograph consisted of a photocell box which also had acorridor 30 cm in circumference, constructed in such a way that a passing insectcaused a change in light intensity setting a pen recorder or counter in motion.The stream of light consisted of a wave longer than 70 nm. Preliminaryexperiments have shown that crickets did not react to this wavelength.
Cautery of neurosecretory cells of the brain
In order to obtaine arrhythmic crickets neurosecretory cells of the parsintercerebralis of the brain were destructed by means of a radio frequencyoscillator (RF), after a previous exposition of this part of brain by cutting a holein the upper surface of the head capsule. The brain was easily located andneurosecretory cells destroyed. After cautery the opening was sealed withmelted wax, and the recording of the locomotor activity of these insects beganimmediately.
Control experiments consisted of all the stages of the experiment excludingthe destruction of the neurosecretory cells.
Brain transplantations
Before brain removal the locomotor activity of the cricket was recorded underconditions of DD. Only insects with clear free-running activity rhythms servedas brain donors. The brain was excised under the ringer solution after Yeagerand Hager (1934), and implanted into the abdomen of arrhythmic cricketswhose neurosecretory cells had been removed and locomotor activity wasrecorded. The wound was sealed with melted wax and the recording of thelocomotor activity started immediately. In control experiments the brains withcauterized neurosecretory cells were implanted into arrhythmic insects.
RESULTS
Brain-donor crickets
Under the conditions of alternating LD 12:12 crickets exhibit very distinctrhythms of locomotor activity (Fig. 1). About two-third of the total activityoccurs during the dark period. The majority of the registered insects begantheir intensive locomotor activity immediately after the darkness falls. If an
135
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4
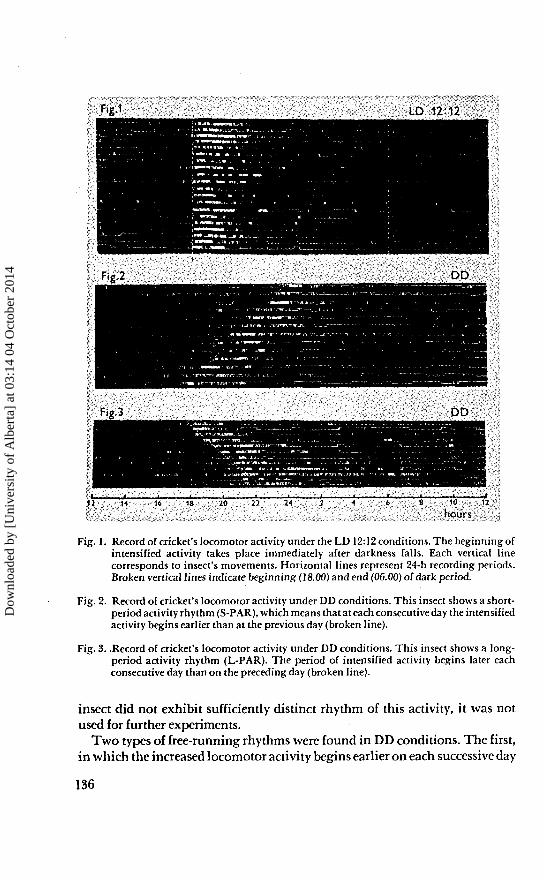
Fig. 1. Record of cricket's locomotor activity under the LD 12:12 conditions. The beginning ofintensified activity takes place immediately after darkness falls. Each vertical linecorresponds to insect's movements. Horizontal lines represent 24-h recording periods.Broken vertical lines indicate beginning (18.00) and end (06.00) of dark period.
Fig. 2. Record of cricket's locomotor activity under DD conditions. This insect shows a short-period activity rhythm (S-PAR), which means that at each consecutive day the intensifiedactivity begins earlier than at the previous day (broken line).
Fig. 3. .Record of cricket's locomotor activity under DD conditions. This insect shows a long-period activity rhythm (L-PAR). The period of intensified activity begins later eachconsecutive day than on the preceding day (broken line).
insect did not exhibit sufficiently distinct rhythm of this activity, it was notused for further experiments.
Two types of free-running rhythms were found in DD conditions. The first,in which the increased locomotor activity begins earlier on each successive day
136
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

(short-period activity rhythm - S-PAR) than on the preceding day (Fig. 2). This"acceleration" takes place at different rates with different individuals, being onthe average 25 min. About 80% of the examined insects have this type of rhythm;a period shorter than 24 h. The second type is formed by insects with a rhythmlonger (long-period activity rhythm - L-PAR) than 24 h (Fig. 3). In this group ofinsects the beginning of increased activity takes place with a certain constantdelay on each successive day, 35 min on average.
These two groups of insects were used as a brain donors: S-PAR and L-PAR.
Brain recipient crickets
Crickets which underwent cautery of their median neurosecretory cells wereused as brain recipients. All the insects in this group were arrhythmic andhyperactive (Fig. 4). They usually died within 1-2 weeks after the brain surgerywithout noticeable changes in their locomotor activities.
"ÖD*a n m u n u. n t r n n t i r i *»*•««« m i w » r • inn« : * » ••n4 n J F « * wrr.« rmiwu. •• r i n f ~JI 1 1 « '*•- * > r « m
w j w r i w r t i i E i . f i / ï J H i i ï i H » M M i i -i , 1 * r i - 4 Ï * « 1 1 1 i j i - i . a r j u ' i i i i i n n M I « Iw w w i n n 11411 »ii 1 »!- . ,« :*-- « H i.sM»«.f.. in-.i".-)«..iiT»:-r..<i -»• im •rt^.i •*••> > 1 n - . j . 1..;.« iiiI T « * I '•••'«' '-* il 1 rul- 'T J 1 I t i l i l t 1 . J « i 1 i. i 1 1 1 1 ' . a i ' n i ï» !•••» 11 -4 " J ' . j r i;s 1 I I ;«•
i n u : I ' » . - , j t , i i t U i i Jiii*.:.»! .. L U . vi .11 i M J i r s r . u . r . - i . j 1 . 1 s \r r - _ , . r . i : : . r . - : iI t t l I..) i ( 1 . 4 - i i i .«r : i -4. r ( i . - | . j - r :i 1 1 11 i.-j..-.: 1 . 1 . " .r 1 'I I 1. . . I ..- J - •. •' r r . r . | IM r n r u . 1 . i . n i i ; v » i ; ; j r a ^ r ' s - . l J i ) T 4 r « I » I - S ; ( ; J » » • 1 •...• 1 ; i 1 . ' i . 1 ( " " » . M r . i u
DD
-. . -lia: t'H Ii-'4IIII i! I.I ••! I ! '-.-. '• . .. , ' ..- ..: ".: . I .r:J. . r n u M i i i - u n . ,..m i ..i i n i i -n .« ,i «i : ,-a ...i Ï . B I i - i iu r ! . - i
i l l i « i i.iiuj-if ;.i m i . * r -rii »I- r J - , I . i -.m.t u wits »<ti irt it
M C .1 * Ljf" i-f i ' 1 . m i l r ; i if r-,.|--
Fig. 4. Record of locomotor activity of cricket after cautery of the median neurosecretory cells.Insect is aperiodic and hyperactive.
Fig. 5. Record of locomotor activity of an arrhythmic cricket after implantation of a brain takenfrom a short-period activity rhythm (S-PAR) donor. A clear short-period activity rhythmsimilar to that seen in the brain donor insect is observed for 3 days (cf Fig. 2).
Fig. 6. Record of locomotor activity of an arrhythmic cricket after implantation of a brain takenfrom a L-PAR donor. A clear long-period activity rhythm similar to that observed in thebrain donor insect can be seen for 3 days (cf Fig. 3).
137
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

Locomotor activity after the brain implantation
When a brain is taken from a rhythmic cricket, either S-PAR or L-PAR, andimplanted into the abdomen of an arrhythmic recipient in which theneurosecretory cells of pars intercerebralis had been destroyed, this results in acreation of locomotor activity rhythms of the recipient. There were 12experiments with brain implantation taken from S-PAR crickets in which atleast 2-3 cycles were observed (Table 1). Only the recipients with distinctlocomotor activity rhythms were taken into account (Fig. 5). As can be seenfrom this figure, a clear short-period free-running rhythm persists for 3 cyclesand then becomes less obvious. Striking similarities are observed in theperiodicity of rhythms of an unoperated brain donor and recipient afterreceipt of the brain (compare Fig. 2 and Fig. 5).
Table 1. Activity rhythms in arrhythmic hosts implanted with S-PAR of L-PAR brains.
_. No. of No. of Average period*n o r experiments rhythmic hosts in h ± S.D.
SPAR 12 7 23 ±0.8L-PAR 8 5 25 ±0.6
• Average period was calculated by periodogram analysis.
The implantation of a brain from L-PAR cricket into an arrhythmic hostalso restores the periodicity characteristic for the donor (Table 1, Fig. 6).Among 8 successful transplantations the period of locomotor activity rhythmwas on the average 25 ± 0.6 h. It is remarkably close tot that observed inunoperated brain donor insects (compare Fig. 3 and Fig. 6).
Both S-PAR and L-PAR brains have a similar inhibitory effect on thelocomotor activity of recipients. Immediately after the brain implantation thetotal activity level declines and stays so for 2-3 cycles of rhythmic behaviour andthan the host insect becomes again hyperactive and aperiodic. In controlexperiments none of the arrhythmic host showed evidence of rhythmicity(N = 11 ) after receiving brain with cauterized neurosecretory cells. Neither wasthere any change in the level of the activity, except for the short periodfollowing immediately the surgery when the activity increased.
In none of the post mortem examined crickets the neural connectionsbetween the implanted brain and the host tissues have been found.
138
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

DISCUSSION
It is obvious that a brain isolated either from a short-period free-runningactivity cricket or from a long-period activity one can create its characteristicactivity rhythm when implanted into an arrhythmic host. Since no neuralconnections between the brain and the host tissues were found, the conferredrhythmicity must occur due to a hormonal factor periodically released from thedonor brain into the host hemolymph. The implantation of a brain withremoved median neurosecretory cells does not produce a periodicity in arr-hythmic animals. It seems, therefore, that neurosecretory cells of the parsintercerebralis of the brain are the source of the observed rhythmicity.
The presented data strongly suggest that crickets' main circadian pacemakeris located in the pars intercerebralis of the brain as postulated earlier (Cymbo-rowski, 1970a,b; Cymborowski, 1973a,b). These findings are in agreementswith studies of Hardier and Konopka ( 1979) on Drosophila fly. In both cricketsand flies, the brain has an inhibitory effect on the total level of the locomotoractivity. In crickets the hormonal factor produced by the median neurosecre-tory cells of the brain affects the motor system of the recipient, either directly orindirectly through the host brain. The same seems to be true for the Drosophilasystem, becauseKonopka and Wells (1980) have found that a mutation whicheliminates circadian locomotor activity rhythm of this insect affects the loca-tion in median neurosecretory cells of the brain.
ACKNOWLEDGEMENT
This work was supported in part by scientific programme R-III-14 coordinatedby the Jagiellonian University, Cracow, Poland.
REFERENCES
BRADY, J. (1969): How are insect circadian rhythms controlled? Nature 223: 781-784.BRUCE, V. G. and PITTENDRIGH, C. S. (1956): Temperature independence in a unicellular
"clock". Proc. Nat. Acad. Sci. USA. 42: 676-682.CIEMIOR, K. E., SEHNAL, F. and CYMBOROWSKI, B. (1977): The role of the neuro-
endocrine system as a pacemaker of insect moulting cycles. J. interdiscipl. Cycle Res. 8:301-303.
CYMBOROWSKI, B. (1970a): Investigation on the neurohormonal factors controllingcircadian rhythm of locomotor activity of the house cricket (Acheta domesticus L.). I.The role of the brain and subesophageal ganglion. Zool. Pol. 20: 103-125.
CYMBOROWSKI, B. (1970b): Investigation on the neurohormonal factors controlling circadianrhythm of locomotor activity of the house cricket (Acheta domesticus L.). II. Dailyhistochemical changes in the neurosecretory cells of the pars intercerebralis andsubesophageal ganglion. Zool. Pol. 20: 127-151.
139
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4

CYMBOROWSKI, B. (1972): Construction of an apparatus for recording the locomotor activityof insects. Exp. Physiol. Biochem. 5: 229-255.
CYMBOROWSKI, B. (1973a): Les variations diurnes de l'activite des cellules neurosecretricesdu cerveau du grillon (Acheta domesticus L.). Acrida 2: 299-306.
CYMBOROWSKI, B. (1973b): Control of the circadian rhythm of locomotor activity in thehouse cricket. J. Insect Physiol. 19: 1423-1440.
CYMBOROWSKI, B. and DUTKOWSKI, A. (1968): The RNA synthesis in the neurosecretorycells of brain and in the subesophageal ganglion of house cricket (Acheta domesticusL.): Oithoptera, Gryllidae. Bull, Pol. Acad. Sci. Cl II. 16: 497-499.
CYMBOROWSKI, B. and DUTKOWSKI, A. (1969): Circadian changes in RNA synthesis in theneurosecretory cells of brain and subesophageal ganglion of the house cricket (Achetadomesticus L.). J. Insect Physiol. 15: 1187-1197.
CYMBOROWSKI, B. and DUTKOWSKI, A. (1970): Circadian changes in protein synthesis inthe neurosecretory cells of the central nervous system of Acheta domesticus L. J. InsectPhysiol. 16: 341-438.
CYMBOROWSKI, B., DUTKOWSKI, A. and SKANGIEL-KRAMSKA, J. (1970): Circadianchanges of acetylcholinesterase activity in the brain of house cricket (Acheta domesticusL.). Comp. Biochem. Physiol. 32: 367-370.
CYMBOROWSKI, B. and BRADY, J. (1972): Insect circadian rhythms transmitted by parabiosis- a re-examination. Nature new Biol. 236: 221-222.
DUTKOWSKI, A. and B. CYMBOROWSKI, (1970): The diurnal changes in RNA synthesis inthe epithelium cells of the egg vesicles of the house cricket. Bull. Pol. Acad. Sci. Cl II.18: 241-243.
HASTINGS, J. W. and SWEENEY, B. M. (1958): A percistent diurnal rhythm of luminescencein Gonyaulax polyedra. Biol. Bull. Woods Hole, 115: 440-458.
HARDLER, A. M. and KONOPKA, R. J. (1979): Transplantation of a circadian pacemaker inDrosophila. Nature 279: 236-238.
HARKER, J. E. (1955): Control of diurnal rhythms of activity in Periplaneta americana L.Nature, Lond. 175: 773.
HARKER, J. E. (1960): Endocrine and nervous factors in insect circadian rhythms. Cold SpringHarb. Symp. quant. Biol. 25: 279-287.
KONOPKA, R. J. and WELLS, S. (1980): Drosophila clock mutations affect the morphology ofa brain neurosecretory cells group. J. Neurobiol. (in press).
SAUNDERS, D. S. (1976): Insect Clocks, Pergamon Press.SOKOLOVE, P. G. (1975): Localization of the cockroach optic lobe circadian pacemaker with
microlesions. Brain Res. 87: 13-21.YEAGER, I. F. and HAGER, A. (1934): On the rates of contraction of the isolated heart and
Malpighian tube of the insect Periplaneta orientalis. Iowa State Coll. J. Sci. 8: 391-395.
140
Dow
nloa
ded
by [
Uni
vers
ity o
f A
lber
ta]
at 0
3:14
04
Oct
ober
201
4





























