Embed Size (px)
Citation preview

Plant Physiol. (1968) 43, 756-766
The Respiratory Chain Components of Higher Plant Mitochondrial
Claude Lance2 and Walter D. Bonner, Jr.Th.e Johnson Researoh Foundation, University of Pennsylvania, Philadelphia, Pennsylvania 19104
Received January 9, 1968.
Abstract. Tightly coupled mitochondria have been prepared from a variety of plant sources:
white potato (Solanum tuberosum), Jerusalem artichoke (Heliantus tuberosus), cauliflower
buds (Brassica oleracea), and mung bean hypocotyls (Phaseolus aureus). Mitochondria with no
appreciable coupling were also prepared from skunk cabbage spadices (Symplocarpus foetidus).
Room temperature difference spectra show that these mitochondria are very similar in the
qualitative and quantitative composition of their electron carriers. The different cytochromesare present in the amounts of 0.1 to 0.3 mamole per mg of mitochondrial protein. The molarratios of the different eleotron carriers are, on the average: 0.7:0.7:1.0:3 to 4:10 to 15 respec-
tively for cytochrome aa3, cytochromes b, cytochromes c, flavoproteins, and pyridine nucleotides.From low tem!perature difference spectra carried out under particular experimental oondi-
tions, it can be deduced that these mitochondria contain 3 b cytoehromes whose a bands are
located at 552, 557, and 561 mi,, and 2 c cytochromes, one of which, a cl-like cytochrome, isfirmly bound to the mitochondrial membrane. Cytochrome oxidase can be optically resolvedinto its 2 components a and a.
For all kinds of mitochondria, the rates of oxidation of succinate are similar as well as theturnover of cytochrome oxidase (50-70 sec'1), regardless of the metabolic activities of thetissues. The number of mitochondria per cell appears to be the controlling factor of theintensity of tissue respiration.
Higher plant tilssues are characterized by agrea'ter number of components in the mitochondrialrespiratory chain than i,s the case wi'th animaltissues (6, 7, 37, 44). Some components (cyto-chrome f, cytochrome b, ), are presen(t only inohloroplasts and play a role in photosynthesis (28)or are a'ssociated with the microsomal fraction ofthe cell (41). However, there s!til,l exists someconfusion in the literatuire about the number andthe identity of the cytoohromes that are generaIllyfound in higher plant mitochondria. Thtis, in 1951,Bha.gvat and Hill (5), extending the work ofKeilin (33), found that the resspiratory chain inthe mittochondria o,f a nuimber of plant materialswa's simi,lar to t'hat of animal tissues and consistedof cytochrome oxi'dase, cytochrome b, and cyto-chrome c. Later, Martin and Morton (41), andWiskich, Morton, and Martin (50) found that therespiratory clhain of plant mitochondria consistedof cytochromes b, cl, c, a, and a3. To this sequience,Lundegardh has added cytochrome b3 (38, 39, 40),a soluble cytochrome first described by HiIll andScarisbrick (29). Bendall and Hill (4) and Ben-dalil (2) ihave suggested that cyto'chrome b7, an
1 This work was supported by a grant from theNational Science Foundation.
2 Present address: Laboratoire de Biologie Vegetale4, Faculte des Sciences, Universite de Paris, France.
autooxidizable component, is part of the respiratorychain of plant tissue's that exhibit a cyanide insen-sitive respiration such as the spadices of the flowersin the Aroid family. Finally, with improvementsin the observation techniques, the broad asymmetricband, originalfly attributed to cytochrome b, hasbeen reso,lveid into a ntumber oif components and thecharacterization or isolation of different types of bcytochromes from plant mitochondria has been re-ported (1, 6, 7, 32).
This paper will present evidence that the cvto-chromes characteri,stic of higher plants consist of3 b colmponents, one of which i,s b7, but the otlher2 are neither b nor b3, 2 c components, one of whichis not cl, and cytochrome oxidase (cyto'chrome a
and cytochrome a3). The concentrations of thesecomponen,ts, as well as those of pyridine nuicleo-tides an'd flavoproteins, have been measuired in a
variety of plant tissues and the results of thesemeasturements are presented and discussed in th.ispap.er. Historically, it is interesting to note thatthe c component descrilbed by Hill and his associate(5) was not cytochrome c, but rather the 'second c
type cytochrome that wiill 'be described in this pa'per.Fuirthermore, the c1 component described by Wis-kich et al. (50) and by Luindegardh (38) was notcytochrome c1, but rather one of the b components.Pertinent evidence stupporting these historical com-
ments are provided in this paper. A preliminaryreport of thi,s work has appeared (35).
756 www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from
Copyright © 1968 American Society of Plant Biologists. All rights reserved.

757LANCE AND BONNER-ELECTRON CARRIERS OF PLANT MITOCHONDRIA
Experimental Procedure
Mitochondria were prepared from a variety ofplant material's including tissues with a low meta-bolic activity sudh as tulbers of potato (Solanumtutberosum) or Jerusalem artichoke (Helianthustuberosus) or from actively respiring tiissues, suchas cauliflower bud's (Brassica oleracea) and fastgrowing mung bean hypocoityls (Phaseolus aureus).A cyanide insensitive tissue, the spadix of the skunkcabbage (Symplocarputs foetidus), was also includedfor comparison.
Standarrd techniques were used for the prepara-tion of the mitochondria (10). The tissues weredisintegrated using either a salad maker or a
Waring Blendor operated at low speed. The grind-ing medium 'used in a ratio of ,tissue to mediu1mof 1 to 2, consisted of 0.3 M mannitol, 1 mm EDTA,0.1 % 'bovine serum alibumin and 0.05 % cysteine.During the grinding and squeezing of the breithrough muslin, the pH was adjutsted in the vicinityof 7.2. The ho;mogenate was centrifuged at 1000 gfor 15 minutes to eliminate the cell debris, then at10,000 g for 15 minutes. The mitochondriad pelletwas sutspended and homogenized in a 'small volumeof washing medium identical to the grinding mediuimexcept that cysteine was omitted. Following 2 more
centr:ftigations-250 g for 10 minutes, and 6000 g
for 15 minutes-the mitochondria were collected and
stuspended in a small volume of washing mediumso that 'the concentration in mitoohondrial proteinwas around 100 mg per ml. A,lil operations were
carried out at a temperature of 2 to 4°.Protein estimations were made uisin(g a semi-
mi'cro Kjeldah'l technique (46) and multiplying by6.25 the amounts of nitrogen found after nesgleriza-tion of the digested samples.
Oxygen uptake was measured 'in a 3 ml 'stirredcel,l tusing a Clark oxygen electrode. The mediutmcontained 0.3 M mannitol, 10 mm KCI, 5 mM MgCloand 10 mm phosphate buffer. The ppH was adjusteda't 7.20. T'he oxygen concentration in ai;r saturatedlmedium at room temperauture (20-220) was takena's 240 FM (19).
Spectrophotometric measurements were per-
formed with the use of a split beam scanningspectroPhotometer as developed by Chance andcollaborators (13). The concentrations of thedifferent eleotron carriers were measured at room
temperature from reduced minus oxidized di,ffer-ence spectra after reduction of the mitoohondrialpreparation by succinate or by hydrosulfite. Thepairs of wavelengths selected for the measurement
of the individual component's, a's well as the ex-
tinction coefficients, were those given by Chanceand Wi'l'liams (20). However, necessary adjust-ments had to be made to take into account the factthat the absorption peaks of plant electron carriersare not always the same as those of simitlar carriersof animail mi;tochondria.
The identifi'cation of the various cytochromespresent in plant mitochondria was best performedat liquid nitrogen temperature (77°K). The oxi-dized and reduced samples were placed in a 3 mmlight path cuvette and cooled in a Dewar flaskfilled with liquid nitrogen during the scanning ofthe spectrum (6, 21). Low temperature brings ahigher resolution as wdlHl as an enhancement of theabsorption peak. By scanning the same sample,first at room temperature and then at low tempera-ture, it is possible to measure the magnitude of thelow temperature en'hancement and, by using thesame extinction coefficients as before (20), theconcentration of the various carriers can be ob-tained from the low temperature spectra. However,as the peaks of the pigments are overlapping eachother, and als the enhancemenit is not uniform overthe whole spectrum, the resulits obtained tunder theseconditions are only estimates and mutist be regardedwith caution (see Results). Although there havebeen some ten'tative determinations of extinctioncoefficients for cytochrome b (42), no definitivecoefficients are available for plant respiratory chaincomponents. Those values generally accepted forestimating the concentrations of the respiratorychain carriers fouind in animall mitochondria (20)have been u'sed in this study. The use of theseextinction coefficients limits the accuracy of ourresults, but in point of fact the error cannot bevery large because of the remarkable constancy ofthese coeffi-cients in the range of 16 to 23 mM'cm-for the a-bands of most known cytochromes.
Resultsand Discussion
Quantitative Determination of Electron Carriersat Rooml Temperature. Figure 1 shows a series ofroom tempera,ture difference spectra obtained withJerusa,lem artichoke mitoohondria. The tipper spec-trum shows a clear identification of the cytochromesafter reduotion by dithionite. The a!bsorption peakof cytochrome oxidase (cytochrome a + cyto-ohrome a3) is located at 602 mju, a few mjm closerto the blue region of the spectrum than cytoc-hromeoxidase from animal mitochondria whose a-bandis generally found at 605 mu (52). T.he a-bandof cytochrome c is at 550 m,L, and cytoohrome bappears as a shou,lder centered around 560 mu.The broad absorption peak aroun'd 530 m,u repre-sents the combined ,8 band's of both cytochrome cand cytochrome b. The Soret region is dominatedby the absorption peak of cytochrome b at 430 mu,whereas cytochrome oxidase appears as a shoulderaround 442 mju. If the reduction of the mitochon-driad carriers is brought about by anaerobiosis inthe presence of succinate, the picture (middlespectrum) i's quite similar except that the extent ofreduction of the b component is smaller than in thepresence of dithionite, while cytochrome c andcytochrome oxidase are reduced to the same pro-
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
portions. The cytochrome b shoutilderm,u is ilower than in the presence of diin the Soret region the contribution ofb appears as a shoutlder on the short
Je
430
428 442
i 11
1~
A.D=-0.01If
-rusalem Artichoke MitochondicRoom Temperature
550/560
+
AO,D=0 005
T ,' /
E
Succ. /i
.-- T- I T.400 440 480 520
Wavelength (mj)560 600
FIG. 1. Difference spectra of Jerusalmitochondria at room temperature (230).dithionite reduced minus aerobic (0 bubbltion. Middle trace: succinate (16 mM) ri
aerobic. Lower trace: succinate (16 mm
mycin A (0.0015 ,ug/ml) aerobic minus ae
path: 10 mm (quartz cuvettes). Mitochotconcentration: 10 mg per ml approxim;chondria suspended in the medium used factivity assay.
around 560 side o-f the cytochrome oxidase peak at 442 mju.thioni,te and The difference spectrtum between an antimycin Acytochrome inhibi,ted succinate aerobic preparation and an
wavelength oxygenated one gives the difference spectrum ofthe b component alone (lower spectrum). Thisspectrum presents a broad peak centered aroun(d560 my and a ;shar,p absorption peak in the Soretregion at 428 m,i. The flavoprotein contribution
602 to the difference spectrum, which is not shown in
s204-Ox figutre 1, is :represented by a trough between 510and 465 m,u. As for the pyridine nuoleotides, theircontribution is represented by an increase in ab-
Succ Anaerobc-Ox sorption in the ultraviolet between 374 and 340 mn,(9). The mitochondrial pyridine nucleotides con-
sist mainly o,f NAD + NADH, very little amounts'Antimycin/O2-Ox of NADP + NADPH are present (26). Very
similar spectra were obtained from al,l mito,chondriauised in thiis study, including the skunk cabbage,whose oxidation processes are insensitive to cyanide(6,7,51).
Iem artichoke The concentrations of the various electron car-
Upper trace: riers, obtained 'by using the method of Chance anding) prepara- Wililiams (20) for 5 different kinds of mitochon-educed minus dria, are given in table I. Expressed in term's ofrobic. Optical
ai mmoles per mg of mitoc'hondrial protein, theidria.l protein concentration's of the different carriers appear toately. Mito-
i
be very similar in each type of mitochondria. Theor respiratory concentration of the different cytochromes is of
the order o,f 0.1 to 0.3 mp,m,ole/mg protein, a range
Table I. Respiratory Chaint Comtponents of Plant MitochlondriaThe concentrations of the different electron carriers are determined from room temperature difference spectra
between a dithionite reduced preparation and a similar one maintained oxidized by 02 bubbling (fig 1, upper trace).The concentrations are measured using the wavelengths (sliightly modified) and absorption coefficients of Chanceand Williams (20). All figures are mean values of 8 to 10 different experiments for each type of mitochondria,except for the skunk cabbage (4 expts). Cvtochrome aa : cytochrome oxida,se, FP: flavoproteins, PN: pyridinenucl otiides.
Conc in electron carriersMaterial Cyt. aa3 Cyt. b Cyt. c FP PN
nIA.noles per ntig proteinJerusalem artichoke 0.10 0.10 0.15 0.38 0.92Mtung bean 0.11 0.12 0.17 0.58 4.10White potato 0.17 0.20 0.27 0.89 3.50Cauliflower 0.12 0.15 0.21 0.69 2.04Skunk cabbage 0.11 0.23 0.36 2.02 ...
Reference X used (m,,) 630 575 5440 510 374Measuring A used (mrn) 602 562 550 460 340E (mM-cm-1) 16 20 19 11 6
Table II. Stoichionzetry of the Respiratory Chain Components in Plant MitochondriaComputed from the vailues given in ta,ble I. The concentration of cytochrome c is taken as unity.
Material Relative concCyt. aa3 Cyt. b Cyt. c FP PN
Jerusalem artichoke 0.69 0.67 1.00 2.90 7.5Mung -bean 0.65 0.67 1.00 3.64 17.8White potato 0.65 0.76 1.00 3.30 14.1Cauliflower 0.60 0.75 1.00 3.40 1.1.5Skunk cabbage 0.32 0.65 1.00 5.80 ...
I I
1758
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.
LANCE AND BONNER-ELECTRON CARRIERS OF PLANT MITOCHONDRIA
of values quite similar to the concentrations gener-ally found in animal mitochondria (15, 17, 22).The filavoprotein's show more variability in therange of their concentrations as do the pyridinenuoleotides. Skunk cabbage mitochondria areslightly different from the other mitochondria, inthat the cytochrome c and flavoprotein contents arerather high compared to the cytochrome oxidasecontent.
Wthen expressed on a relaltive basils (table II),takinig the cytochrome c concentration as the unity,the resullts show that for most of the mitochondriathere is roughly 0.7 of each cytochrome oxidase(cytochrome aa3) and cytochrome b for 1 cyto-chrome c, 3 to 4 flavoprotein's and 10 to 15 pyridinenucleotides. The stoichiometry of the differentelectron carriers found in plant mi,toch-ondria issimillar to that reported for animal tissues, thoughthe concentration of the pyridine nuoleotides seemsto be slightly lower (15, 16, 18, 20). The flavo-protein content of skunik cajbbage mi,tochondria is
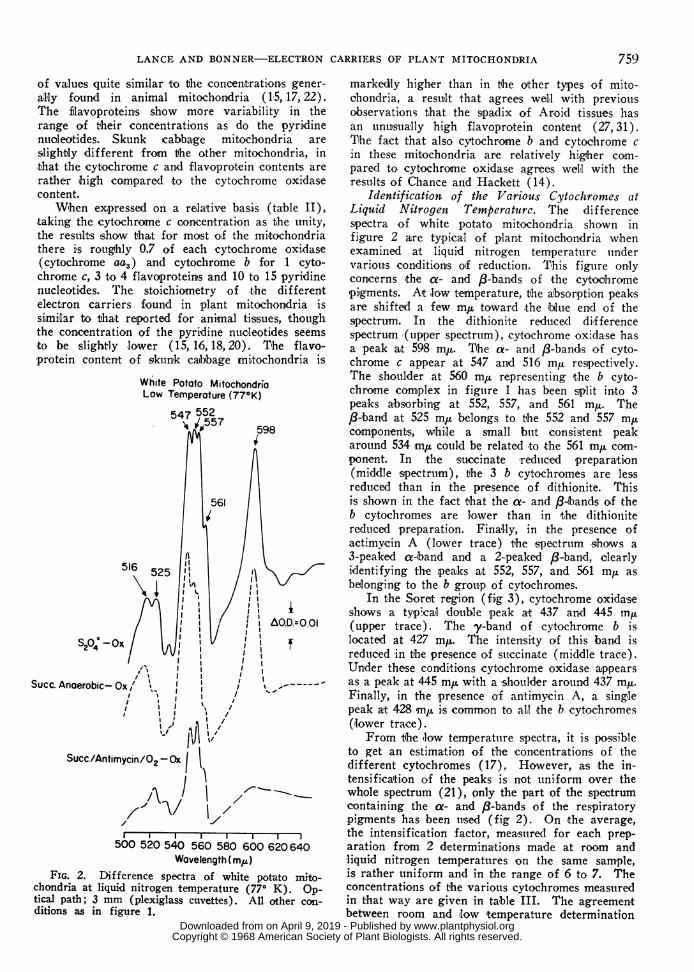
White Potato MitochondriaLow Temperature (77°K)
516
s2o Ox
/,% III
Succ. Anaerobic- Ox / % II
. II I
I 'II I I
;
Succ./Antinnycin/02-Ox
/ 7~~~I-
I I I I I I -Il500 520 540 560 580 600 620 640
Wavelength ( m.)FIG. 2. Difference spectra of white potato mito-
chondria at liquid nitrogen temperature (770 K). Op-tical path; 3 mm (plexiglass cuvettes). All other con-ditions as in figure 1.
markedly higher than in the other types of mito-ohondria, a result that agrees well with previousobservations that the spadix of Aroid tissues hasan unusually high flavoprotein content (27, 31).The fact that also cytochrome b and cytochrome cin theise mitochondria are relatively higher com-pared to cytochrome oxidase agrees well with therestu.lts of Chance and Hackett (14).
Identification of the Various Cytochrontes atLiquid Nitrogen Temperature. The differencespectra of white potato mitochondria shown infigure 2 are typical of plant mitochon'dria whenexamined at liquid nitrogen temperature uindervarious conditions oif reduction. This figure onlyconcerns the az- and 8f-bands of the cytochromepigments. Alt low temperature, the absorption peak'sare shifted a few m,u toward the lbluie end of thespectru.m. In the dithionite reduced differencespectrum (upper spectrum), cytochrome oxidase hasa peak at 598 m,,. The a- and 8-band's of cyto-chrome c appear at 547 an'd 516 mp respectively.The shoulder at 560 mJL representing the b cyto-chrome complex in figture 1 has been split into 3peaks absorbing at 552, 557, and 561 m,u. The,8-band at 525 m,l belongs to the 552 an'd 557 mucomponents, While a small but consistent peakarotund 534 mp could be related to the 561 mtu com-ponent. In the 'succinate reduced preparation(middle spectrum), the 3 b cytochromes are lessreduced than in the presence of dithionite. Thisis shown in the fact that the a- and 8l-bands of theb cytochromes are lower than in the dithioiiitereduced preparation. Finally, in the presence ofactimvcin A (lower trace) the ispectrtum slhows a3-peaked a-band and a 2-peaked f8-band, cilearlyidentifying the peaks at 552, 557, and 561 my asbelonging to the b group of cytochromes.
In the Soret region (fig 3), cytochrome oxidaseshows a typi,cal double peak at 437 and 445 tnu(upper trace). The y-band of cytochrome b islocated at 427 m,. The intensity of this band isreduced in the presence of stiiccinate (middle trace).Under these conditions cytochrome oxidase appearsas a peak at 445 mp with a shoulder around 437 my.Finally, in the presence of antimycin A, a sing-lepeak at 428 mnu i,s common to aLl the b cytochromes(lower trace).
From the 'low temperatture spectra, it is possibleto get an estimation of the concentrations of thedifferent cytochromes (17). However, as the in-tensificaltion of the peaks is not uniform over thewhole spectrum (21), only the part of the spectrumcontaining the oa- and f8-bands of the respiratorypigments has been ulsed (fig 2). On the average,the intensification factor, meastured for each prep-aration from 2 determinations made at room andliquid nitrogen temperatures on the same sample,is rather tuniform and in the range of 6 to 7. Theconcentrations of the various cytochromes measuredin that way are given in table III. The agreementbetween room and llow itemperature determination
759
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.
760 PLANT PHYSIOLOGY
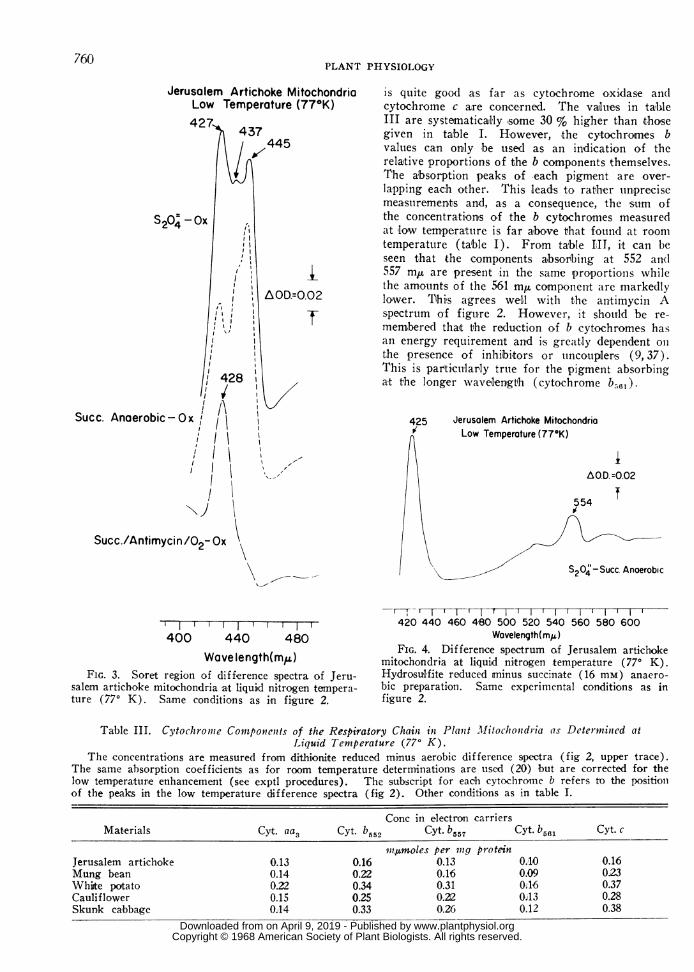
Jerusalem Artichoke MitochondriaLow Temperature (77°K)
437445
S2O4 Ox
I
AOD.=0.02
T
Succ. Anaerobic-Ox
is quite good as far as cytochrome oxidase andlcytochrome c are concerned. The vailu!es in tableIII are systematicallly some 30 % higher than t-hosegiven in table I. However, the cytochromes bvalues can on,ly be used as an indication of therela,tive proportion's of the b components themselves.The absorption peaks of each pigment are over-lapping each other. This leads to rather tunprecisemeasturemen,ts and, as a consequence, the sum ofthe concentrations of the b cytochromes mea-suredat 'low tempera!tuire is far above that found at roomtemperature (table I). From table HIT, it can beseen tjhat the components absorbing at 552 and557 m,u are present in the same proportionis whilethe amounts of the 561 m,u componen,t are markedlylower. TIhi,s agrees well w.ithl the antimycini Aspectrum of figure 2. However, it should be re-membered tha't the reduction of b cytochromes hasan energy requirement and i!s greatily dependent olnthe presence of inhibitors or tuncoupler's (9, 37).This is parti'cularly true for the pigment absorbingat the longer wavelenigth (cytochrome b5,,6,).
425 Jerusalem Artichoke MitochondriaLow Temperoture (770K)
AO.D. =0.02
Succ./Antimycin /02- Ox
S204'-Succ Anoerobic
400 440 480Wave length(mEs)
FIG. 3. Soret region of difference spectra of Jeru-salem artichoke mitochondria at liquid nitrogen tempera-ture (770 K). Same conditions as in figure 2.
'- I rI420 440 460 4-80 500 520 540 560 580 600
Wdvelength(mE)FIG. 4. Differen'ce spectrum of Jerusalem artichoke
mitochondria at liquid nitrogen temperature (770 K).Hydrosulfite reeduced minus succinate (16 mM) anaero-bic preparation. Same experimental conditions as infigure 2.
Table III. Cytochromle Contponenits of the Respiratory Chaint in Plan1t Mitochon'dr-ia as Determinbed atLiquid Temperature (770 K).
The concentrations are measured from dithionite reduced minus aerobic difference spectra (fig 2, upper trace).The same absorption coefficients as for room temperature determinations are used (20) but are corrected for thelow temperature enhanicement (see exptl procedures). The subscript for each cytochroime b refers to the positionof the peaks in the low temperature difference spectra (fig 2). Other conditions as in table I.
Conc in electron carriersMaterials Cyt. aa3 Cyt. b552 Cyt. b557 Cyt. b561 Cyt. c
niuimoles per lig proteinJerusalem artichoke 0.13 0.16 0.13 0.10 0.16Mung bean 0.14 0.22 0.16 0.09 0.23White potato 0.22 0.34 0.31 01l6 0.37Cauliflower 0.15 0.25 0.22 0A3 0.28Skunk cabbage 0A4 0.33 0.26 0.12 0.38
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.
LANCE AND BONNER-ELECTRON CARRIERS OF PLANT MITOCHONDRIA
All the values reported in tables I and III arebased on difference spectra obtained after reductionby hydrosulfite. While ireduction by succinategives about the same extent of reduction for cyto-chrome oxidase and cytoohrome c, the b cytochromesare less reduced than in ,the presence of hydrosulfite(fig 1, 2, 3). The extra reduction brought aboutby the addition of hydrosulfite to a succinate re-duced preparation is generally in the order of 20 to30 %. However, the real question is whetherhydrosulfite reduces the b cytochromes mnore com-pletely than does anaerobiosis induced by succinate,or if it affects some other comrponent absorbing inthe same region. A olue to thils answer is given infigure 4, which represents a difference spectrumbetween a dithionite reduced and a succinate re-duced preparation of Jerusailem artichoke mitochon-dria. This -spectrum only shows a broad a-bandcentered around 554 m;u, a slight fl-band arouniid525 mju, and a single Soret peak a:t 425 mju. In nocase, however, with all types of mitochondria usedin this study, was it possible to obtain a typical3 'peak absorption band in the at region similar tothat given by an antimycin A spectrum (.fig 2).It t;herefore seems likely that the extra reductionindtuced by dithionite affects some other componentthan the different b cytochromes (7, 14). In thatcase, the vailues reported in tables I and HI for thecytochromes b should be lowered by a factor ofaboult one-third.
The a-a3 Component of Plant Mitochondria.Though the existence of 2 componenbs in cyto-chrome oxidaise has been a controversial quiestionfor a long time, it is now most generalily agreedthat cytochrome oxiidase consists of 2 spectrolpho-tometrically identifiable components, a and a3, evenif it has not been possible to isolate 2 chemicallydif.ferent entities thus far (52, 53). Recently, Ben-dall and Bonner described some optical propertiesof plant cytochromne oxidafse (3). The opticalseparation of the 2 components a and a3 can beachieved under particular conditions in the presenceof cyanide or azide, which form complexes withcytochrome a3 only (3, 34, 49). It has already beennoticed that the a-band of cytochrome oxirdase ofplant mitochondria i's located at 602 mju instead of605 mjf in animal mitochondria (fig 1). Lowtemperature brings a shift toward the blue at598 m,u (fig 2).
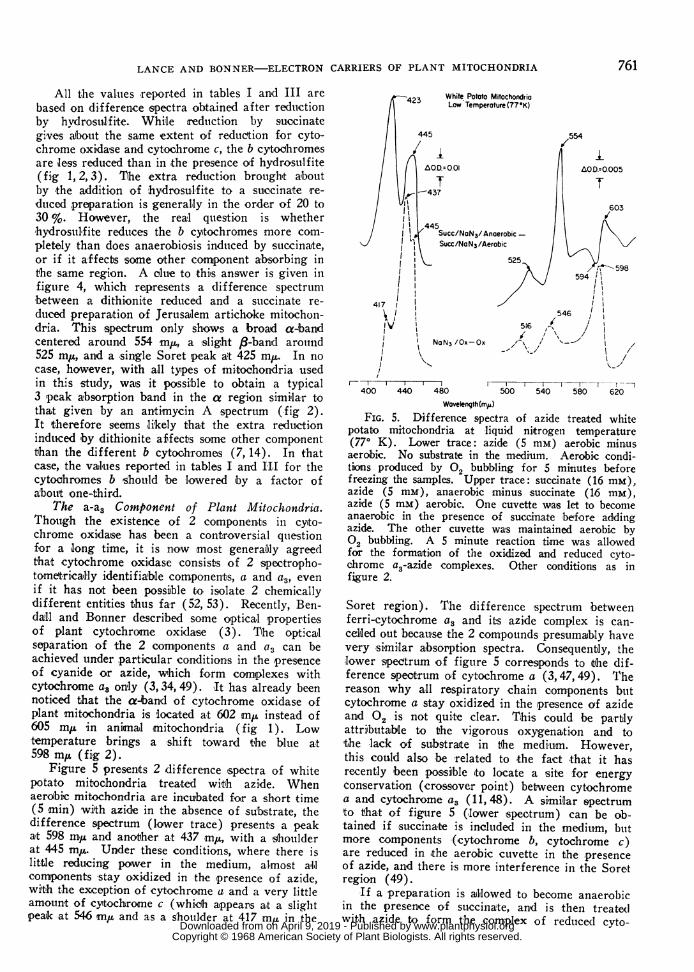
Figure 5 presenits 2 difference spectra of whitepotato mitochondria treated witih azide. Whenaeroibic mitochondria are incubated for a short time(5 min) wilth azide in the absence of substrate, thedifference spectrum (lower trace) presents a peaka't 598 miA and another at 437 m,, with a shoulderat 445 minu. Under these conditions, where there islittle reducing power in the medium, almost all,components stay oxidized in the presence of azide,wit-h the exception of cytochrome a and a very littleamount of cytochrome c (which appears at a slightpeak at 546 in/h and as a shoulder at 417 miu in the
I
I
II
417
Iv
11 NaN3 /Ox-Ox
I \4
,_T II400 440 480
i
T
.*-598594 59
I '1
I546 /
_, ', ,'I1516 -
/ ( I~//
500 540 580 620
Wovelength(ms)FIG. 5. Dilfference spectra of azide treated white
potato mitochondria at liquid nitrogen temperature(770 K). Lower trace: azide (5 mM) aerobic minusaerobic. No substrate in the medium. Aerobic condi-tions produced by 02 bubbling for 5 minutes beforefreezing the samples. Upper trace: succinate (16 mM),azide (5 mM), anaerobic minus succinate (16 mM),azide (5 mm) aerobic. One cuvette was let to becomeanaerobic in the presence of succinate before addingazide. The other cuvette was maintained aerobic by02 bubbling. A 5 minute reaction time was allowedfor the formantion of the oxidized and reduced cyto-chrome a3-azide complexes. Other conditions as infigure 2.
Soret region). The difference spectrum betweenferri-cyto-chrome a3 and its azide complex is can-celled out because the 2 compounds presumably havevery similar absorption spectra. Consequenbly, thelower speetrum of figure 5 corresponds to the dif-ference spectrum of cytochrome a (3,47,49). Thereason why all respiratory chain components butcytochrome a stay oxidized in the presence of azideand 02 is not quite clear. This could be partlyattrilbutable to the vigorous oxygenation and tothe lack of substraite in the medium. However,this could also be related to the fact that it hasrecently 'been possilble 'to locate a site for energyconservation (crossover point) between cytochromea and cytochrome a3 (11,48). A simillar spectrum'to tha-t of figure 5 (lower spectrum) can be ob-tained if succinate is included in the medium, butmore components (cytochrome b, cytochrome c)are redu'ced in the aerobic cuvette in the presenceof azide, and there is more interferen.ce in the Soretregion (49).
-If a preparation is alllowed to become anaerohicin the presence of succinate, and is then treatedwith azide to form the complex of reducedl cyto-
White Potato MitochondrioLow Temperoture (771K)
145
t&OD.-0001Tr
761
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
c!hrome (3, the difference spectrum (fig 5, uppertrace) with a preparation containing both succinateand azide, but maintained aerobic by 02 bub'bling,shows the difference spectrum of the azide complexof cytochrome a3. Cytochrome a(3 a.ppears as asingle broad peak at 445 mj.t in the Soret region.In the a band region, the figture (shows a troutghat 594 m,u and a slight peak around 603 mju. Thesame spectrum has been observed using cyanildeinstead of azide (3, 47, 49). Cytochrome a, whichis reduced in both conditions, i's cancelled ouit. Dtueto tihe high reducing power brought by the presenceof succinalte, all the other components are reducedin the aerobic cuvette with the exception of a bcomponent absorbing a't 554 and 423 mix, which willbe discu,ssed later. Through this technique, bothcytochromes a and a3 can be easily optically sepa-rated from each other. When both componentsare reduced at the sa'me time, the absorption at445 my, which includes both a and a3, becomesprominent, and then the peak of cytochrome a at437 mu appears as a shoutlder (fig 3, middle trace).
The b Components of Plant Mitochondria. Be-sides cytochrome a3, figture 5 allso shows that acomponent with a- and fl-bands at 554 and 525 muand a Soret-band at 423 m,u, remains oxidized (ordoes not become reduced) in tihe aerobic sampleeven if all the other carriers, with the exceptionof the oxidized cytochrome (a3-azide complex, arereduced. Due *to the absence of interference withcytochrome c in the a region, anid with cytochromea in the Soret region, this compound is probablyidentical wiith the one which absorbs at 552 anid427 m,u in figures 2 and 3. Most important is thefact that this compound shows a single peak at lowtemperatture. This permits one to consider that the2 peaks at 552 and 557 ml, in figure 2 really belongto 2 different entities and are not the result of thesplitting of the a-band 'of a b cytochrome as isobserved with a microsomal b cytochrome (6, 32).
Plant mit'oohondria differ from animail mito-chondria by the number of their b-type cytochromes.Low temperature spectra clearly show 3 absorptionpeaks (552, 557, 561 mJL) in the a-band, 2 peaks(525, 5.34 m,) in the 8-band and a single Soretpeak (428 mn,) (figs 2 and 3). This indicates thatulp to 3 cytoc'hromes b could be operational in plantmito,chondria. None of them has been isolated inpure form so far, except for tihe coimponent whoselow temperature absorption peak is a't 557 mu, whichhas been partially purified (1) and 2 o'ther b-typecytochromes which have also been purified fromsolutble or particulate fractions from mung beans(32, 42, 43). Therefore it seems likely that theabsorption peaks at 552 and 557 m1u really belongto 2 different b cytochromes.
The absorption peak of the component absorbingat 561 my is consistently ilower than the peaks ofthe 2 other b cytobhrome.s. Moreover, its state ofreduction ifs more dependenvt of the nature of thereducing agents, inhibitors, or uncouplers added to
the medium thain lis the case for the 2 other cyto-chromes (37). It iis very likely that the 561 mJLpeak belongs to a thi!rd b-type cytochrome.
Figure 5 also shows (upper spectrum) that oneof the 'cytochromes b remains oxidized in the pres-ence of azide. Azide or cyanide resistant respira-tion is a typical feattire of some plant materials(9, 37) and a b-type cytochrome (cytochrome b7)ha's been proposed as an inhibitor insensitive catalystto account for the insensitivity to cyaniide of therespiration of the Atroid spa'dices (2,4). Howwever,,low temperatulre spectra of mi,tochondria fromcyanide insensitive tissues such as the skunk cab-bage (6) and determinations of the b cytochromescontent (tables I and II) do not show the presenceof a larger amount of a particular b cytochrome.From this, it appears that tihe b7 component ofBendal-l and Hill (4) shouild be iden'tified with 1 ofthe 3 b ccmponents of figure 2. From evidenceobtained by direct examination with a microspec-troscope of reduced mitochonidrial preparationscooled in liquid nitrogen (2), cytochrome b7 coouldpossibly be identified with cytochrome b557 (8).Whether or not this cytochrome is involved in theinhibitor insensitive respiration is not clear at thepresent time. It has also been suggested that re-sistance to inhibitors could be a general propertyof plant respiration. T'he switch to sensitivity orinsensitivity would be under control mec.hanismsassociated with developmental stages (9).
Cytochrome b3 is 'still'l sometimes considered astaking part in the plant respiratory chain (38,40).The optical and sollubility characteristics of cyto-chrome b3 differ markedly from those of the 3mitochondrial b cytochromes. Cytochrome b3 isextracted from plant itiissuie homogenates 'by di,ltutephosphate buffer (29); the only mitochondrialhemoprotein so extracte'd is cytoc'hrome c, as shownin thi's paper. Cytochrome b3 is reduced withascorbate, no mitochondrial b cytochrome is so re-duced (7). Reduced cytochrome b3 has a broada-band of absorption maximum 559 m,,. Its lowtemperaiture absorption characteristics are notknown, but should the a-banid remain symmetricalat 770K its absorption maximu-m a't this temperaturecould be expected to lie in the region between thoseof b337 and b52.. One deduces from the abovesutmmarized facrts that cytoohrome b3 is not a mem-ber of the plant mitochondrial respiratory chain,and, in fact, the location of b, in the cell remainsunknown.
The c Components of Plant Mitochondria.Cytochrome c ipossesses an absorption peak at 550m,u (fig 1) whi'ch shifts to- 547 mpt at liquidnitrogen temperature (fig 2). Many studies havedemonstrate'd that this component is essentiallyidentical, as far as its optical and biochemical'properties are concerned, wit-h mammalian cyto-chrome c (24). Cytoohrome c1 'has also beenpostulated to be presen't in plant mi,tochondria(38, 41, 50). However, the existence of such a
762
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

763LANCE AND BONNER-ELECTRON CARRIERS OF PLANT MITOCHONDRIA
Jerusalem Artichoke Mitochondria' Cytochromes c Low Tempercture(77'K)
597549
AO.D.=0.02
-f
515
I/;t
, Ascorbate-Ferricyanide
r--I1 --
400 440 480Wavelength (mp)
1 'II
\ i
1500 540 580 620
FIG. 6. Difference spectra of the c cytochromescomponents from Jerusalem artichoke mitochondria atliquid nitrogen temperature (770 K). Lower trace:ascorbate reduced minus ferricyanide oxidized phos-pthate buffer extract of mitochondria. Upper trace:same type of difference spectrum with the mitochondrialpellet remaiining after phosphate and cholate extraction.Cyanide (1 mM) added in the ascorbate reduced cuvetteto prevent reoxidation by cytochrome oxidase.
*cytochrome c1-like componen't can only be demon-strated after all the cytochrome c has been removedfrom the mitoahondria by extenisive washing (9).
Mitochondria from Jerusalem artichoke havebeen thoroughily washed for 12 hours in a phosphatebuffer (pH 7.2, 0.1 M) with gentle stirring. Underthese conditions, cytochrome c goes into soilutionand can be separated from the bulk of the mito-chondria by high speed centrifugation (20,000 g).The lower trace of figure 6 represenfts an ascorbatereduced minus a ferricyanide oxidized differencespectrum of such an extract of Jerusalem artichokemitochondria. Thiis spectrum shows a sharp asym-metrical a-banid at 547 m,u, a a-band at 515 my,and a Soret peak at 415 mu. Generally, in mito-chondrial preparations, the Soret peak of cyto-chrome c does not appear because of the strongcytochrome b peak at 427 mju (fig 3) unless specialtreatments are applied to the mitocho-ndria (fig 5,lower trace).
When cytochrome c has been removed fronm themitochondria by extensive washing with the samephosphate buffer, a ,similar difference spectrum canbe performed on the peUlet, providing a little cyanidei,s added in order to inhibit the cytochrome oxidasestill remaining in the particiles. An ascorbate re-*ducible cytochrome, analogous to the cl componentof animal and yeast mitochondria, remains stronglybound to the mitoch-ondria. Attempts to extract itby the use of surface active agents (10 % cholate,1 % Triton X 100) 'have failed. The upper trace
of figure 6 ishows -the difference spectrum olf thecomponent which remains after phosphate bufferand chola-te extraction. It shows a symmetricala-peak at 549 mju, a n-peak at 517 m,u and a Soretpeak at 419 mu. Cytochrome oxidase, which isreduced at *the same time, also appears on this'spectrum. Cy,tochromes b are not reduced byascorbate. By comparison with cytochrome c, allthe peaks of the bound c-type cytochrome areshi,fted a few mUu toward the red. The opticalcharacteriistics, as well as the strong binding to themitochondrial prooteins, relate thius component tocytochro-me cl of animal and yea,st mitochondria(25).
In -this investigation, no quantitative measure-ments have been made on the concentration o'f thesecond c-type cytochrome. Judging fro.m theamoun't of materiall requiired to produce equivaleintdifference spectra, it appears that in Jerusalemartichoke mitochondria -the content of the c1-liikecomponent is less than that of cytochrome c; asituation tha!t might not pertain to all mitochondria.Because of the close proximity of red'uced aabsorp-tion bands, the 2 c-type cytochromes a-ppear in thedifference spectra of intact mitochondria as a singlecomponent (fig 2). In particular, the 552 my(770K) component sho-uld nott ibe confused wi,thcytochrome c1 on rthe 'basis of chemiical and spectralanalogy with the mammalian or yeast cytochromec1 (23,25). The antimycin A sensitivity definitelyrelates the 552 mju component to the b group ofcytoohromes (,fig 2, lower trace).
r ADP/O:I .51 -1.75
FIG. 7. Oxygen electrode trace of succinate oxida-tion by white potato mitochondri(a. The additions areshown as final concentrations. The figures on thetrace are the rates of OO utilization expressed as thedecrease in ium 02 concentration per minute. Mw in-diicates the addition of the mitodhond'ria (350 pg ofinitochondnial nitrogen added to a final volume of 3.0ml)n.
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
The above remarks show that cytochrome c]cannot be spectroscopically identified and measulredat room temperature in intact plant mitowhondriaas it has been reported in the literaltutre (38, 40,41, 50). The broad aibsorption shoulder around553 mju at room temperature is more likely to becontributed by a b-type cytochrome rather than bya c1-like -cytochrome. Nothing is known o!f therole in electron transport of the c1-like plant cyto-chrome which can 'be identified at low temiperattureafter -the removail of cytochrome c.
-It should also be noted that in their identificationof cytoohrome c in particulate preparations fromhigher plants, Bhagva,t and Hi1l (5) were probablydealing with the c-liike cytochrome rather thanwith the norma(l cytochrome c. The technique ofpreparation of the particles (grinding in 67 mmNa2HPO4) probably extracted mo.st of the cyto-chrome c. In fact, the oxidation of succinate by'such particles was very sensitive to the addition ofexogenous cytochrome c (5).
Turnover of the Cytochromes. The preparativemethod described above yields good quality mito-chondria which exhibit respiratory control (19)when supplied with limiting am-ounts of ADP(fig 7). With stuccinate as a substrate, respiratorycontrol (RC) values of 2 to 4 are generally ob-served as well as ADP/O vadues of 1.5 or higher.The skunk cabbage mnitochondria barely possessedrespiratory control. The highest rate of oxidation,whioh is genera'ldy achieved when mitochondria areoxidizing succinate in the presence of ADP ('state3 rate of oxidation), can be considered as a meastureof the activity of the electron traniisport chain(Qo2 expressed on a protein nitrogen basi's) in aparticular ty.pe of mitochondria. From such meas-urements, and from 'the dalta used to produice ta-ble I,it is also possible to determine the turnover of thedilfferent cytochromes, assuming that 4 electronsare necessary to reduce a molecule of oxygen andthat the oxidation-reduction cycle of each cyto-chrome involve's only 1 electron. Such values aregiven in table IV for the 2 terminal carriers (cyto-chrome c and cytodhrome oxida-se) which are sup-posed to be common to all oxidative pathways.
This talble shows that the oxidative abilities o'fall types of mitochondria are very close to each
other, and in the order of 1000 jl of 02 used perhour per mg of protein nitrogen. For comparison,the respiratory activities of the tissues (,ul O. perhr per g 'fr wt), which have been determined fsepa-rately by standard Warburg manometric techniques(46), are aliso included in tajble IV. It is apparentthat there is a relative independence between theactivities of the mitochondria and the nature ofthe tissues from which they have been extracted.Some of these tissues are storage organs withl alow meta;bolic activity (Jeruisalem artichoke, po-tato), some are actively growing and respiringti'ssues (mung bean, Caulifilower buds). In par-ticular, one of them, the skunk cajbbage flower,belongs to a plant family which exhiJbit one of thehighest respiratory rates in the plant kingdom (24).
Since the concentrations of the cytoc'hromecomponents do not present drastic variations from1 ty-pe of mitochondria to the other (talble I), theturnover number of cytoohrome oxidase and cyto--chrome c are also very close to each other. Thefigures for cytochrome oxidase are about 50 to70 sec-1 and a little smaller for cytochrome c. 'T'hevalues for these 5 types of mitochondria comparevery favoraibly with values found for animal orplant mitochondria (12,16,18,36,51). In fact,when carefullily prepared, plant mitochondria aregenerally more active than mi'tochondria derivedfrom animal tissue (30).
The fact that the electron carrier componentsand the respiratory activities of the plant mito-chondria studied here are 'rather uni,form, wi,th no*relation to the metabolic activities of the tissuesfrom which they are derived, strongly suggests thatthe respiratory activities of most plant tissues de-pend on the ntum-ber of mitochondria per cell ratherthan on some qualitative or quantitative di'fferencein mitochondrial catalysts. Thi's i's also exemplifiedby the fact that the yield in mitochondria of some50 g of skunk cabbage spadices is roughly equiva-lent to those of 2 kg of mung beans or 8 kg olfpotatoes.
The uniformity of mitochondrial compositionand activities could also be related to some geneticaspect of mitochondrial physiology. It has beenshown that the DNAs of mitochondria preparedfrom a variety of plant tissues are identical though
Tab-le IV. Activity of the Electront Tronsfer Chaitn in Plant MitochondriaVaJues determined from the highest state 3 rate of succinate oxidation for 02 electrode traces (fig 7). Aill
the figures in this table are mean values obtain-ed fromn 8 to 10 different experiments for each type of mitochondria,except for skunk cabbage (4 expts).
Material Qo2 of mitochondria Turnover Q°e of tissuesCyt. aa3 Cyt. c
,A 02 hr-' mg-1 N sec-1 / 02 hr' (g fr wt)-1Jerusalem artichokeMung -beanWhite potatoCauliflowerSkunk cabbage
1750
118011701030
5,179626574
3950404022
5018040240
4000
764
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

LANCE AND BONNER-ELECTRON CARRIERS OF PLANT MITOCHONDRIA
the nuclear DNAs of these tissues are very dif-ferent (45). An identity in mitochondrial DNAcould be related to a similarity in the size, mor-phology, qualitative and quantitative composition ofthese cell organelles.
Acknowledgment
The authors are deeply indebted to Miss Eva Chris-tensen for expert and elegant technical assistance.
Literature Cited
1. BAKER, J. E. AND M. BORCHELT. 1965. Partialpurification of a b-type cytochrome from sweetpotato mitochondria. Plant Physiol. 40: IV.
2. BENDALL, D. S. 1958. Cytochromes and somerespiratory enzymes iin mitochondria from thespadix of Arum linaculatumn. Biochem. J. 70:38'1-90.
3. BENDALL, D. S. AND W. D. BONNER. 1966. Op-tical properties of plant cytochrome oxidase. In:Hemes and Hemoproteins. B. Chance, R. W.Estabrook, and T. Yonetani, eds. Academic Press,New York. p 4854-8.
4. BENDALL, D. S. AND R. HILL. 1956. Cytochronmecomponents in the spadix of Arum wiiaculatumi?.New Phytologist 5i5: 206-12.
5. BHAGVAT, K. AND R. HILL. 1961. Cytochromeoxidase in higher plants. New Phytologiist 50:112-0.
6. BONNER, W. D. 1961. The cytochromes of planttissues. In: Haematin Enzymes. J. E. Falk, R.Lemberg, anid R. K. Morton, eds. PergamonPress, New York. p 479-97.
7. BONNER, W. D. 1963. Higher plant cytochromes.Proc. 5th Intern. Congr. Biochem. PergamonPress, New York. Vol. 2: 50-62.
8. BONNER, W. D. 1964. Plant cytochromes. SixthIntern. Congr. Biochem. New York. abst. IV:p 291-92.
9. BONNER, W. D. 1965. Mitochondria and electrontransport. In: Plant Biochemistry. J. Bonnerand J. E. Varner, eds. Academic Press, NewYork. p 89-123.
10. BONNER, W. D. 1967. A general method of thepreparation of plant mitochondria. In: Methodsin Enzymology. R. W. Estabrook and M. E.Pullman, eds. Academic Press, New York. Vol.X: p 126-33.
11. BONNER, WV. D. AND M. PLESNICAR. 1967. Elec-tron transport carriers in plant mitochondria.Nature 214: 61617.
12. CHANCE, B. 1954. Enzyme mechanisms in livingcells. In: The Mechanism of Enzyme Action.W. D. McElroy and B. Glass, eds. The JohnsHopkins Press, Baltimore, Maryland. p 399-453.
13. CHANcE, B. 1957. Techniques for the assay ofthe respiratory enzymes. In: Methods in Enzy-mology. S. P. Colowick and N. 0. Kaplan, eds.Academic Press, New York. Vol. 4: p 273-329.
14. CHANcE, B. AND D. P. HACKETT. 1959. Theelectron transfer system of skunk cabbage mito-chondria. Plant Physiol. 34: 33-49.
15. CHANCE, B. AND B. HAGIHARA. 1963. Directspectroscopic measurements of interaction of
components of the respiratory chain with ATP,ADP, phosphate, and uncoupling agents. Proc.5th Intern. Congr. Biochem. Pergamon Press,New York. Vol. 5: 3-37.
16. CHANCE, B. AND B. HESS. 1959. Metabolic con-trol mechanisms. I. Electron transfer in themammalian cell. J. Biol. Chem. 234: 2404-12.
17. CHANCE, B., D. F. PARSONS, AND G. R. WILLIAMS.1964. Cytochrome content of mitochondria strip-ped of inner membrane structure. Science 143:135-39.
18. CHANCE, B. AND B. SACKTOR. 1958. Respiratorymetabolism of insect flight muscle. II. Kineticsof respiratory enzymes in flight musele sarco-somes. Arch. Biochem. Biophys. 76: 509-31.
19. CHANCE, B. AND G. R. WILLIAMS. 1955. Respira-tory enzymes in oxidative phosphorylation. I.Kinetics of oxygen utilizatiDn. J. Biol. Chem.217: 383-93.
20. CHANCE, B. AND G. R. WILLIAMS. 1955. Respira-tory enzymes in oxidative phosphorylation. II.Differen-ce spectra. J. Biol. Chem. 217: 395-407.
21. ESTABROOK, R. W. 1956. The low temperaturespectra of hemoproteins. I. Apparatus and itsapplication to a study of cytochrome c. J. Biol.Chem. 223: 781-94.
22. ESTABROOK, R. W. AND A. HoLowINSKY. 1961.Studies on the content and organization of therespiratory enzymes of mitochondria. J. Biophys.Biochem. Cytol. 9: 19-28.
23. ESTABROOK, R. W. AND B. MACKLER. 1957. Thelow temperature spectra of hemoproZeins. II.Cytochromes of heart muscle preparations. J.Biol. Chem. 224: 637-46.
24. GODDARD, D. R. AND W. D. BONNER. 1960. Cel-lular respiration. In: Plant Physiology, A Trea-titse. F. C. Steward, ed. Academic Press, NewYork. Vol. 1A: 209-312.
25. GOLDBERG, R. AND D. E. GREEN. 1963. Propertiesand function of mammalian cytochromnes b and c].In: The Enzymes. P. D. Boyer, H. Lardy, andK. Myrbick, eds. Academic Press, New York. Vol.8: 81-95.
26. HARMEY, M. A., H. IKUMA, AND W. D. BONNER.1966. Near ultra-violet spectrum of white po-tato mitochondria. Nature 209: 174-75.
27. HERK, A. WV. H. VAN. 1937. Die chemischen Vor-gange im Saurornatum-Kolben. Rec. Trav. Bot.Neerl. 34: 69-156.
28. HILL, R. AND F. BENDALL. 1960. Function of thetwo cytochrome comnponents in chloroplasts: Aworking hypothesis. Nature 186: 136-37.
29. HILL, R. AND R. SCARISBRICK. 1951. The hae-matin compounds of leaves. New Phytologist 50:96-1r11.
30. IKUMA, H. AND W. D. BONNER. 1967. Propertiesof higher plant mitochondria. I. Isolation andsomie characternistics of tightly-coupled mitochon-dria from dark-grown mung bean hypocotyls.Plant Physiol. 42: 67-75.
31. JAMES, W. 0. AND H. BEEVERS. 1950. The res-piration of Arum spadix. A rapid respiration,resistant to cyanide. New Phytologist 49: 353-74.
32. KASINSKY, H. E., H. SHICHI, AND D. P. HACKETT.11966. Particulate cytochromes of mung beanseedlings. Plant Physiol. 41: 73948.
765
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
33. KEILIN, D. 1925. Oni cytochrome, a respiratorypigment, commoin to ani,mals, yeasts, and higherplants. Proc. Roy. Soc. London B 98: 312-39.
34. KEILIN, D. AND E. F. HARTREE. 1939. Cyto-chrome and cytochrome oxidase. Proc. Roy. Soc.London B 127: 167-91.
35. LANCE, C. AND W. D. BONNER. 1966. Quantitativedetermination of sonme electron carrier componentsin plant mitochondria. Plant Physiol. 41: XXVI.
36. LEE, C. P., R. W. ESTABROOK, AND B. CHANCE.1965. Spectrophotometric and kinetic studies ofthe reconstituted succinate oxidase system. Bio-chim. Biophys. Acta 99: 32-45.
37. LIEBERMAN, M. AND J. E. BAKER. 1965. Res-piratory electron transport. Ann. Rev. PlantPhysiol. 16: 343-82.
38. LUNDEGARDH, H. 1958. Spectrophotometric inves-tigations on enzyme systems in living objects.III. Respiratory enzymes in homogenates ofwheat roots. Biochim. Biophys. Acta 27: 355-65.
39. LUNDEGARDH, H. 1'962. The respiratory systemof wheat roots. Biochim. Biophys. Acta 57:352-58.
40. LUNDEGARDH, H. AND G. DUCET. 1966. Le sys-tem;e cytochromique du tebercule et des mitochon-dries de la Pomme de terre. Acad. Sci. Ser. D262: 263-68.
41. MARTIN, E. M. AND R. K. MORTON. 1957. Haempigments of cytoplasmic particles from non-pho-tosynthetic plant tissues. Biochem. J. 65: 40-13.
42. SHICHI, H. AND D. P. HACKETT. 1962. Studieson the b-type cytochromes from mung bean seed-lings. I. Purification of cytochromes b555 andb561. J. Biol. Chem. 237: 2955-58.
43. SHICHi, H. AND D. P. HACKETT. 1966. A pos-sible role for cytochrome b515 in the mung bean
mitochondrial electron transport system. J. Bio-,chem. 59: 84-88.
44. SMITH, L. AND B. CHANCE. 1958. Cytochromesin plants. Ann. Rev. Plant Physiol. 9: 449-82.
45. SUYAMA, Y. AND W. D. BONNER. 1966. DNAfrom plant mitochondria. Plant Physiol. 41:383-88.
46. UMBREIT, W. WV., R. H. BURRIS, AND J. F. STAUF-FER. 1957. Manometric techniques. Burgess Pub-lishing Company. p 274.
47. WILSON, D. F. 1967. Azide inhibition of mito-chondrial electron transport. II. Spectral changesinduced by azide. Biochim. Biophys. Acta 131:431-40
48. WILSON, D. F. AND B. CHANCE. 1967. Azideinhibition of mitochondrial electron transport. I.The aerobic steady state of succinate oxidation.Biochim. Biophys. Acta 131: 421-30.
49. WILSON, D. F. AND M. V. GILAIOUR. 1967. T11elow-temlperature spectral properties of mamma-lian cytochrome oxidase. I. The enzyme in intactrat-liver mitochondria. Biochim. Biophys. Acta143: 52-61.
50. WISKICH, J. T., R. K. MORTON, AND R. N. ROBERT-SON. 1960. The respiratory chain of beetrootmitochondria. Australian J. Biol. Sci. 13: 109-22.
51. YOCUM, C. S. AND D. P. HACKETT. 1957. Par-ticipation of cytochromes in the respiration ofthe aroid spadix. Plant Physiol. 32: 186-91.
52. YONETANI, T. 1960. Studies of cytochrome oxi-dase. I. Absolute and difference absorptionspectra. J. Biol. Chem. 235: 845-52.
53. YONETANI, T. 1963. The a-type cytochromes. In:The Enzymes. P. D. Boyer, H. Lardy, and K.MIyrbdck, eds. Aicademic Press, New York. Vol.8: 41-79.
766
www.plantphysiol.orgon April 9, 2019 - Published by Downloaded from Copyright © 1968 American Society of Plant Biologists. All rights reserved.




![Research Paper Testosterone enhances mitochondrial complex V …€¦ · respiratory chain activity in the SN in adult male rats [14]. The brain is a highly differentiated organ with](https://img.pdfslide.us/doc/110x75/5f9d77aa8fb9867a4221e8a4/research-paper-testosterone-enhances-mitochondrial-complex-v-respiratory-chain-activity.jpg)









![Research Article Grape Powder Improves Age-Related …downloads.hindawi.com/journals/omcl/2016/6135319.pdf · mitochondrial respiratory failure [, ] and mitochondrial ... black California](https://img.pdfslide.us/doc/110x75/5b01e00a7f8b9ad85d8ecac7/research-article-grape-powder-improves-age-related-respiratory-failure-.jpg)














