Embed Size (px)
Citation preview

The Prenatal Development of the Human Femur ' ERNEST GARDNER AND D. J. GRAY Departments of Anatomy, Wayne State University School of Medicine, Detroit, Michigan, and Stanford University School of Medicine, Stanford, California 94305
ABSTRACT The study reported here is the result of a detailed investigation of the changes in shape of the femur with growth and torsion, the type, rate and character of ossification, and the onset, subsequent course, and general features of remodeling.
Forty pairs of femurs, from a series of embryos and fetuses ranging from 26 to 342 mm in crown-rump length, were measured, radiographed, and sectioned for microscopic study.
A primary bony collar was present before the end of the embryonic period, and in a 27 mm embryo it extended for about one-fifth of the length of the femur. Erosion of the collar was evident at 34 mm, and invasion and destruction of calcified cartilage were occurring by 37 mm. Cartilage canals first appeared in the proximal epiphysis at 57 mm and in the distal epiphysis at 61 mm.
Along with the progression of endochondral ossification proximally and distally and the establishment of growth zones, periosteal bone formation also proceeded in both directions, and, until 275 mm, extended about 1 mm beyond the zones of cartilage destruction. After 275 mm, the extents of periosteal and endochondral ossification were the same and at term occupied almost four-fifths of the length of the femur.
Trabeculation of the bony collar was first noted at 37 mm. Fusion of endochondral trabeculae with the inner aspect of the periosteal shell began by 61 mm. A central marrow cavity free of trabeculae was present at 86 mm and thereafter. Evidence of reconstruction appeared in both proximal and distal ends by 92 mm, and was con- sistently present in both ends in all specimens of 111 mm and larger.
Although a number of studies of various aspects of the prenatal development and growth of the femur have been reported by many investigators, a comprehensive and detailed account is not available. The study reported here was focused mainly upon the changes in shape with growth and torsion, the first appearance, type, and rate of os- sification, and on the onset, subsequent course, and general features of remodeling.
Early development and shape. The most important of the early studies were by Bardeen and Lewis ('01) and Bardeen ('05). Bardeen and Lewis noted that in a 9 mm embryo the femur consisted of con- densed mesenchyme, which faded gradu- ally into the undifferentiated mesenchyme of the distal portion of the limb. Bardeen found that at 11 mm the femur was chon- drifying, and that at 14 mm it had a simple, rounded head. A slight projection, distal and dorsal to the head, formed the beginning of the greater trochanter, but a neck was not evident until 20 mm, when the lesser trochanter and the intertro- chanteric ridge were indicated by blastemal extensions. Cartilage had extended into the
AM. J. ANAT., 129: 121-140.
lesser trochanter and the intertrochanteric ridge by 33 mm, and, although the neck was proportionately short and very thick, the main characteristics distinguishing the proximal end of the femur had become es- tablished. The head was more rounded and the neck relatively more slender at 50 mm. The linea aspera consisted of perichon- drium during early development. Up to 50 mm cartilage did not extend into this area, and, since the shaft was ossified by this time, it was evident that no cartilaginous linea aspera was ever formed.
Pitzen ('23) included the femurs of a somewhat older group of fetuses in his study. He found the diaphysis of a femur 11 mm long to be entirely ossified, and re- ported that the linea aspera consisted of periosteum in the smaller specimens.
Strayer ('43) published the results of an extensive study of the early development of the hip joint, in which he discussed the blood supply of the head and neck of the
1 This investigation was supported by research grants HD 00377 from the National Institute of Child Health and Human Development and AM6705 from the National Institute of Arthritis and Metabolic Dis- eases of the National Institutes of Health, Public Health Service.
121

122 ERNEST GARDNER AND D. J. GRAY
femur as well as the development of the anlagen which form the acetabulum. IIaines ('47) and Gray and Gardner ('50) found that chondrification of the skeletal blastema was present at 12 mm, and that by 13 to 15 mm the femoral condyles and the intercondylar fossa were indicated. Cardner and Gray ('50) observed that the head of the femur was rounded and the neck constricted at 20 mm, and that the greater trochanter was present as a slight elevation at 22 mm. A pit for the ligament of the head was first found at 37 to 39 mm.
A number of investigators have cor- related embryonic development with Streeter's ('49) stages. O'Rahilly, Gardner and Gray ('56), O'Rahilly, Gray and Gardner ('57), and Gardner and ORahilly ('68) published reports on the development of the lower limb and on the time of ap- pearance and the initial ossification of the femur in staged human embryos. They found that the lower limb buds first ap- pear at stage 13, that is, in embryos about 5 mm long and about four weeks after fertilization. By about five weeks after fertilization, the skeletal parts of the lower limb are present in blastemal form and are beginning to chondrify in a proximo- distal sequence. By stage 18 (usual crown- rump length of 12-17 mm, about 6 weeks), the femur is beginning to chondrify. By stage 19 (usual length of 16-19 mm, about 7 weeks), the femoral condyles may be chondrifying. By stage 22 (usual length of 23-28 mm, about 7.5-8 weeks), the femur has a clear-cut cartilaginous form and a primary bony collar is forming. At the end of the embryonic period (stage 23, usual length of 27-31 mm, about 8 weeks), the knee and hip joints clearly resemble those of the adult in form and arrange- ment. The primary bony collar is well- defined, but is not yet invaded by a perio- steal bud.
In reviewing the early development of the femur, Olivier ('62) included illustra- tions of embryonic femurs reconstructed to show changes in size and shape.
Several investigators have paid atten- tion to the angle of torsion of the femur during prenatal development (Friedlander, '01; Le Damany, '03; Pitzen, '23; Altmann, '25; Nishizuka, '26; Brandt, '28; de Cuve- land, '50; Felts, '54). There seems to be
fairly general agreement that torsion oc- curs mainly during the last half of pre- natal development and reaches its maxi- mum near term.
Ossification. Many investigators have concerned themselves with the histo- genesis and ossification of the femur dur- ing prenatal development.
Gray and Gardner ('50) found that a primary bony collar was forming in an embryo of 20 mm, and O'Rahilly, Gray and Gardner ('57) and Gardner and ORahilly ('68) found i t present in Streeter's stages 22 and 23. Bade ('00) was the first to apply radiographic meth- ods in a study of the development of the femur, and noted an ossific center (endo- chondral) in the shaft after nine weeks of development ( a similar finding noted by Gray and Gardner ('50) and Gardner and Gray ('50) in their histological studies). He published a table in which he listed the lengths of the ossified parts of the shaft in a series of fetuses ranging from 3.4 to 39.0 cm in crown-rump length. He also observed that epiphysial centers were present in the distal epiphyses of two fetuses at term.
After a radiographic study of 140 fetuses ranging in age from 10.5 to 38 weeks, Pryor ('23) concluded that the bones of the female ossify in advance of those of the male. He found centers in the distal epiphyses of femurs at from 25 to 30 weeks in female fetuses and at from 30 to 40 weeks in male.
Halonen ('29) noted the first radio- graphic appearance of bone in the shaft at 5.0 cm crown-heel length in a male fetus and at 4.0 cm crown-heel length in a fe- male. He found the proximal unossified part of the femur to be longer than the distal unossified portion in fetuses up to 286 mm crown-heel; in specimens larger than this the ratio was variable. The con- vexity of the ossific zones toward the un- ossified parts of the bone was first evident at 30 cm crown-heel. Halonen also re- corded the percentages of total length oc- cupied by the bony shaft at 10 cm inter- vals in fetuses ranging from 10 to 50 cm crown-heel length. He noted that in several cases periosteal bone extended beyond the endochondral. Bidder ('06) had reported this extension previously.

PRENATAL DEVELOPMENT OF FEMUR 123
Gardner and Gray ('50) observed that ossification had almost reached the neck in fetuses from 85 to 95 mm crown-rump length, that it extended proximal to the trochanters at 285 mm, and that it reached the level of the inferior synovial reflection of the hip joint at 308 mm.
In a report of a study of transverse sec- tions of the long bones of a series of fetuses ranging from 34 to 335 mm in crown-rump length, Knese ('57) claimed to have seen a border line between the periosteal and endochondral bone. He regarded this line as a remnant of cartilage tissue, and stated that it was present until shortly after birth. Knese also reported that the marrow cavity, which consisted of several cavities at 58 and at 74 mm, was still subdivided at 105 mm.
Along with his studies of torsion, Nishizuka ('26) studied the growth of the femur by measuring its greatest length in a series of fetuses ranging from four months of age to term. Konig and Korn- feld ('27) found that about 10% of paired femurs were asymmetrical in length in fetuses between four and six months. These asymmetries decreased in number in older fetuses and disappeared in those at term.
Felts ('54) made 15 measurements on each of 53 whole femurs from fetuses and infants ranging from 31 to 480 mm in crown-rump length. He found that the total length of the femur increases by 0.21 mm for every millimeter increase in crown- rump length, and that the length of the ossified part of the shaft, as shown by staining with alizarin red S, increases very rapidly relative to the total length of the shaft. Bone formed about 80% and carti- lage about 20% of the length of the femur at term. Felts also described the changing shape of the chondro-osseous junction with the growth of the femur.
In a study of the ossified part of the femoral shafts, Moss et al. ('55) used 119 embryos and fetuses, ranging from 14 to 175 mm in crown-rump length, and cleared and stained with alizarin red S. After the specimens had been grouped in 10 mm crown-rump intervals, the mean overall lengths of the ossified part of the shafts were recorded. Plottings of the lengths of the ossified part of the shafts
against crown-rump lengths all showed a characteristic interphase in the crown- rump interval of 80-89 mm, or during the twelfth week. The ossified part of the shaft grew relatively faster than the crown-rump length before the interphase; after this the length of the bony part of the shaft in- creased at a slower specific rate, although it continued to increase relatively faster than crown-rump length.
Vasczrlarizntion. v. Friedlander ('04) studied the course and distribution of cartilage canals in the proximal and distal ends of the femur. Bidder ('06) noted the presence of bone in a cartilage canal of the distal end of the femur of a newborn child. He also described a second kind of canal, different from the cartilage canal, which he termed a longitudinal perforat- ing vascular canal. He found such vascular canals to be present at three and one-half months.
In a study of 14 fetuses, ranging from eight to nine months of age, Bertrand ('23) found that the vessels that run in the cartilage canals arise from the capsule, the synovial membrane, and the ligaments. He maintained that cartilage canals do not anastomose with each other and that they do not communicate with vessels in the djaphysis.
Hintzche ('28) also noted the presence of bone in the cartilage canals of the distal end of the femur. He saw it for the first time at 74 mm, and was able to trace it into the diaphysis. In addition to bone, Hintzsche observed elastic fibers, fat cells, and islands of cartilage in the canals. Al- though he noted connections between the vessels in the cartilage canals and those in the diaphysis, he claimed that the canals play no part in ossification and serve only for nutrition. Harris ('29) did not observe any communication between the vessels of the epiphysis and those of the diaphysis in the newborn.
Gardner and Gray ('50) noted the first appearance of a cartilage canal at 49 to 50 mm, when it entered the femoral head from the perichondrium of the neck. A few vessels from the ligament of the head en- tered canals in the head at from 85 to 95 mm. Gray and Gardner ('50) observed the first appearance of canals in the distal end at 60 mm, when they entered the epi-

124 ERNEST GARDNER AND D . J. GRAY
physis anteriorly, just proximal to the condyle.
Early investigators appear to have as- sumed that the definitive nutrient artery entered the femur at the initial site of erosion of the bony collar by the perio- steal bud, subsequent to the onset of endo- chondral ossification. Humphrey ( 1861 ) attributed the obliquity of the canal to the unequal rate of growth of the two ends of the bone and the direction of the obliquity to the more rapid rate of growth of the distal end.
On the assumption that the first artery to penetrate the bony collar persists as the nutrient artery, Digby ('16) measured the parts of the adult femur proximal and distal to the site of penetration and con- cluded that 31% of the bone is formed from the proximal part and 69% from the distal.
Basing his studies on a similar assump- tion, Halonen ('29) stated that the propor- tions of the parts of the diaphysis proximal and distal to the nutrient foramen changed during fetal life from a ratio of approxi- mately 50:50 to a ratio of 44:56. He first observed a nutrient canal at 8.9 cm crown-heel length, and found one regularly after 100 cm crown-heel length. At 50 cm he found two canals. One entered at the middle of the bone and ran inward and a little distally. The other entered at about the junction of the middle and distal thirds, and ran obliquely and proximally. Three canals were observed exceptionally. Halonen claimed that the positions of the canals were frequently the same on the two sides.
On the other hand, Lutken ('50) found the nutrient foramina to be symmetrical in the left and right femurs that he studied in only about half the cases. Furthermore, he observed no typical position for them on the shafts.
The studies of Brookes ('58) led him to conclude that two principal arteries are normally present in the femoral shaft, and that, after uniting in the medullary cavity, they course toward the juxta-epiphysial zone and repeatedly subdivide, supplying the fetal cortex from the medulla.
Remodeling. Although reports of stud- ies of the reconstruction and remodeling at the ends of long bones have been pub-
lished (by Gardner, '56 and by Enlow, '63, for example), few of these are concerned with the femur specifically. Halonen ('29) first noted resorption in the femur at 10 cm crown-heel length, and observed it regularly after 12.5 cm crown-heel.
Amprino and Cattaneo ('37) studied the attachments of muscles to bones dur- ing prenatal development and found two main types. One group attached to bony parts that ossify endochondrally, and the other group to bony parts that ossify periosteally. They discussed the develop- ment of each of these. Hoyte and Enlow ('66) discussed the attachment of muscles to bony surfaces as the latter are being resorbed.
MATERIALS AND METHODS
A series of 40 embryos and fetuses was used for this study. Their crown-rump lengths ranged from 26 to 342 mm when measured in the fresh state. The speci- mens were fixed in neutral formalin for a period of several months, after which their crown-rump lengths were again measured. Few differences were noted in the mea- surements before and after fixation, and these were not appreciable.
After the left and right femurs were removed from each specimen, their lengths were recorded and each bone was radio- graphed. All right bones were then silver- ized with the use of the technique em- ployed by ORahilly and Meyer ('56) and radiographed again. The radiographic technique employed was such that the lengths of the bones appearing on the ra- diograms were the same as those of the bones when first removed from the specimens.
The bones were decalcified in a mixture of equal volumes of 50% formic acid and 20% sodium citrate. The femurs from spe- cimens less than 125 mm were embedded either in paraffin or in celloidin followed by parafin. Those from fetuses larger than 125 mm were embedded in celloidin alone. All bones were serially sectioned in frontal, sagittal, or transverse planes, and then stained with a modified Masson technique.
It has been reported by a number of investigators (see Observations) that his- tological methods are the most sensitive in detecting ossification during early pre-

PRENATAL DEVELOPMENT OF FEMUR 125
'TABLE 1 T h e crown-rump W.R.) lengths of the specimens
By the end of the embryonic period, the femur resembles the adult bone in form and relationships. The shaft is surrounded by a bony collar, which appears first at
in mil l imeters and t h e plane of section of t h e f e m u r s
C.R.
mm 26 2'7 34 37 57 61 63 64 69 73 77 85 86 92 93 97
103 105 110 111
Left Right C.R.
mm L L 113 L L 118 L L 125 F F 127 F ? ' 130 -r F 135 T F 138 F T 146 T F 148 S T 151 F T 165 'r s 180 T F 185 F T 185 S T 211 T F 277 s T 302 F T 316 S T 342 F T 342
~
Left ___
Right
T T T T F T T S F T F T F S T T F T F S
S S F T T F S T T F T F T T F F T F T T ~~ -
L, longitudinal; S, sagittal; F, frontal; T, transverse.
natal development. Ordinary radiographic techniques are the least sensitive and show neither periosteal or endochondral ossifi- cation until several weeks after it appears his tologically.
Because the radiograms of the femurs from the specimens of 26, 27, 34, 37, and 57 mm did not show endochondral ossifi- cation, longitudinal sections were used to measure the lengths of the bones and of the ossified parts of their shafts. Sections of the left femurs were employed for these measurements.
Radiograms of the right femurs of fetuses of 61 mm and larger were used to measure the lengths of the bones and of the ossified parts of their shafts. The per- centages of the femurs occupied by bone were calculated from these measurements.
Table 1 lists the crown-rump lengths of the specimens used and the plane of sec- tion of each femur.
OBSERVATIONS
The early embryonic development of the femur has been described in previous papers (Bardeen and Lewis, '01; Bardeen, '05; Pitzen, '23; Gray and Gardner, '50; Gardner and Gray, '50; O'Rahilly et al., '57; Gardner and ORahilly, '68). It is not reported again here.
Streeter's stages 22 or 23 -(usual crown- rump length of from 23 to 31 mrn).
Shape The general shape of the femur was
quite similar to that of the adult in the youngest specimens examined. The head. neck, trochanters, and condyles were all present. During its early development the shaft was rounded in transverse sections, mainly in its middle part. However, sur- faces and borders soon became evident. The linea aspera (see below) was evident by 61 mm, and the gluteal tuberosity by 69 mm.
The characteristic features of the femur became more sharply accentuated during the fetal period, mainly as a result of ossification, remodeling, and changes in torsion, inclination, and obliquity.
Angular dimensions. No specific stud- ies were made of changes in these di- mensions. Ordinary inspection of a num- ber of femurs corroborated Felts' ('54) finding that torsion is more often nega- tive in the younger Specimens and that it is markedly positive at term.
Ossification A primary bony collar was present in
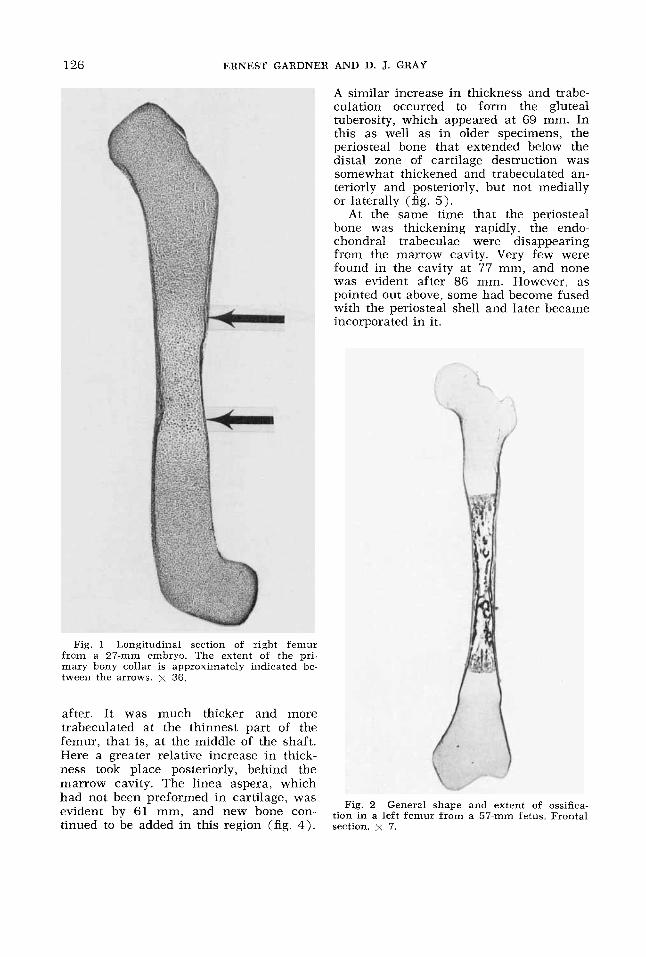
the youngest specimen studied, at 26 mm. In a 27-mm embryo, it extended for about one-fiftieth the length of the femur (fig. 1). Early trabeculation of the collar was first observed at 37 mm.
The erosion of the bony collar by a periosteal bud was evident at 34 mm, and by 37 mm invasion and destruction of calcified cartilage were occurring. Endo- chondral ossification and the formation of endochondral trabeculae were prominent by 57 mm (fig. 2 ) , and, together with destruction of cartilage, progressed rapidly toward each end of the femur. As the trabeculae grew in size and united with one another, some of them fused with the periosteally formed shell. This process was evident by 61 mm, and is illustrated in older specimens (fig. 3).
Regional differences in the periosteal bone were evident at 61 mm and there-
126 ERNEST GARDNER AND D. J. GRAY
A similar increase in thickness and trabe- culation occurred to form the gluteal tuberosity, which appeared at 69 mm. In this as well as in older specimens, the periosteal bone that extended below the distal zone of cartilage destruction was somewhat thickened and trabeculated an- teriorly and posteriorly, but not medially or laterally (fig. 5).
At the same time that the periosteal bone was thickening rapidly, the endo- chondral trabeculae were disappearing from the marrow cavity. Very few were found in the cavity at 77 mm, and none was evident after 86 mm. However, as pointed out above, some had become fused with the periosteal shell and later became incorporated in it.
Fig. 1 Longitudinal section of right femur from a 27-mm embryo. The extent of the pri- mary bony collar is approximately indicated be- tween the arrows. x 36.
after. It was much thicker and more trabeculated at the thinnest part of the femur, that is, at the middle of the shaft. Here a greater relative increase in thick- ness took place posteriorly, behind the marrow cavity. The linea aspera. which
- ,
had not been preformed in was Fig, 2 General shape and extent of ossifica- evident by 61 mm, and new bone ‘On- tinued to be added in this region (fig. 4) .
tion in a left femur from a 57-mm fetus. Frontal section. x 7.
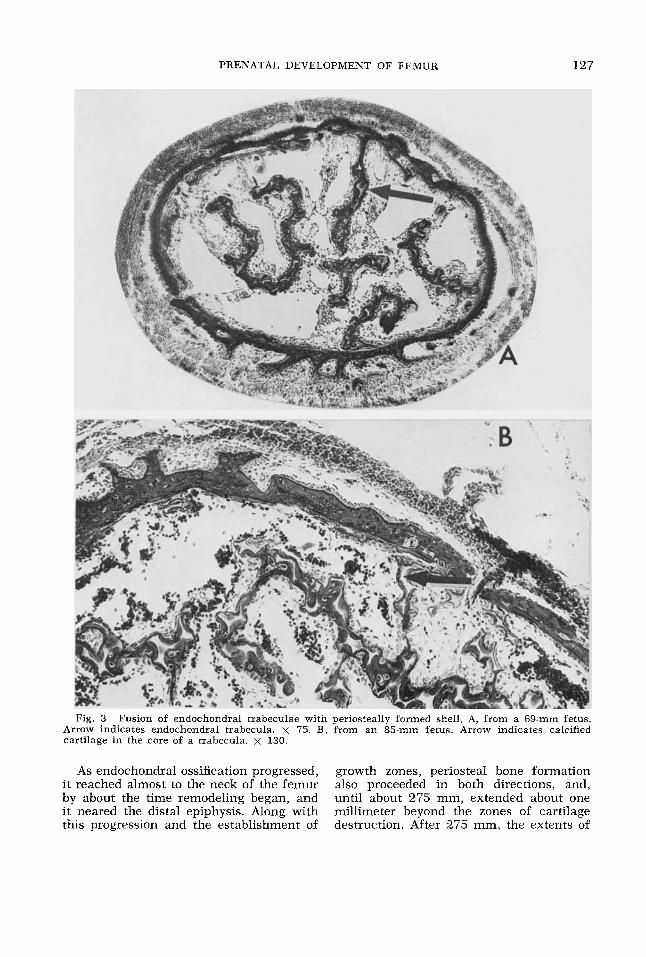
PRENATAL DEVELOPMENT OF FEMUR 127
Fig. 3 Fusion of endochondral trabeculae with periosteally formed shell. A, from a 69-mm fetus. Arrow indicates endochondral trabecula. x 75. B, from an 85-mm fetus. Arrow indicates calcified cartilage in the core of a trabecula. x 130.
As endochondral ossification progressed, growth zones, periosteal bone formation it reached almost to the neck of the femur also proceeded in both directions, and, by about the time remodeling began, and until about 275 mm, extended about one i t neared the distal epiphysis. Along with millimeter beyond the zones of cartilage this progression and the establishment of destruction. After 275 mm, the extents of
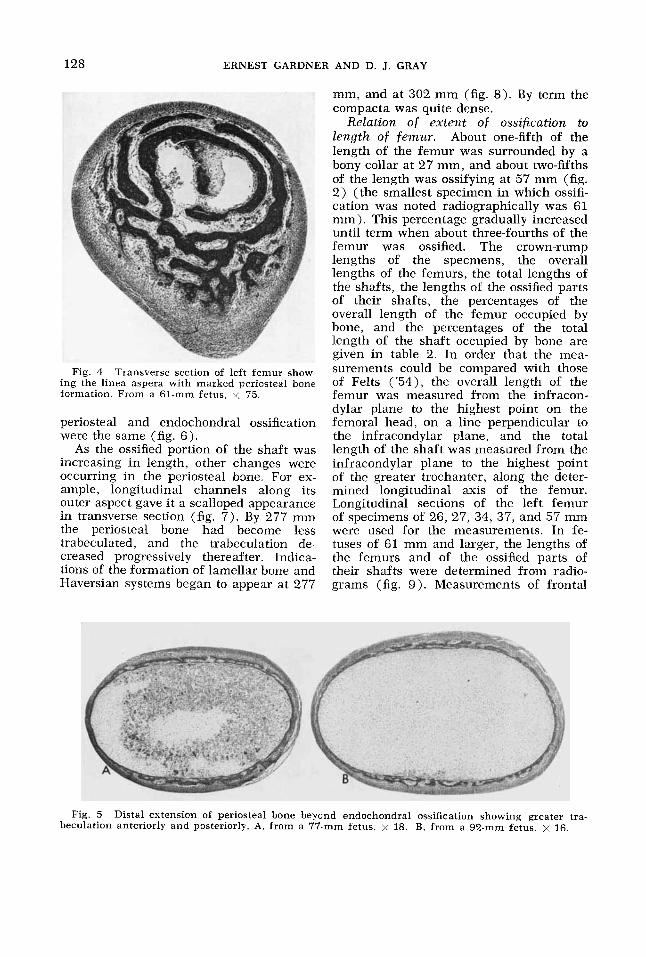
128 ERNEST GARDNER AND D. J. GRAY
Fig. 4 Transverse section of left femur show- ing the linea aspera with marked periosteal bone formation. From a 61-mm fetus, x 75.
periosteal and endochondral ossification were the same (fig. 6 ) .
As the ossified portion of the shaft was increasing in length, other changes were occurring in the periosteal bone. For ex- ample, longitudinal channels along its outer aspect gave it a scalloped appearance in transverse section (fig. 7) . By 277 mm the periosteal bone had become less trabeculated, and the trabeculation de- creased progressively thereafter. Indica- tions of the formation of lamellar bone and Haversian systems began to appear at 277
mm, and at 302 mm (fig. 8). By term the compacta was quite dense.
Relation o f extent of ossification to length o f femur. About one-fifth of the length of the femur was surrounded by a bony collar at 27 mm, and about two-fifths of the length was ossifying at 57 mm (fig. 2) (the smallest specimen in which ossifi- cation was noted radiographically was 61 mm). This percentage gradually increased until term when about three-fourths of the femur was ossified. The crown-rump lengths of the specmens, the overall lengths of the femurs, the total lengths of the shafts, the lengths of the ossified parts of their shafts, the percentages of the overall length of the femur occupied by bone, and the percentages of the total length of the shaft occupied by bone are given in table 2. In order that the mea- surements could be compared with those of Felts ('54), the overall length of the femur was measured from the infracon- dylar plane to the highest point on the femoral head, on a line perpendicular to the infracondylar plane, and the total length of the shaft was measured from the infracondylar plane to the highest point of the greater trochanter, along the deter- mined longitudinal axis of the femur. Longitudinal sections of the left femur of specimens of 26, 27, 34, 37, and 57 mm were used for the measurements. In fe- tuses of 61 mm and larger, the lengths of the femurs and of the ossified parts of their shafts were determined from radio- grams (fig. 9). Measurements of frontal
Fig. 5 Distal extension of periosteal bone beyond endochondral ossification showing greater tra- beculation anteriorly and posteriorly. A, from a 77-mm fetus. x 18. B, from a 92-mm fetus. x 16.
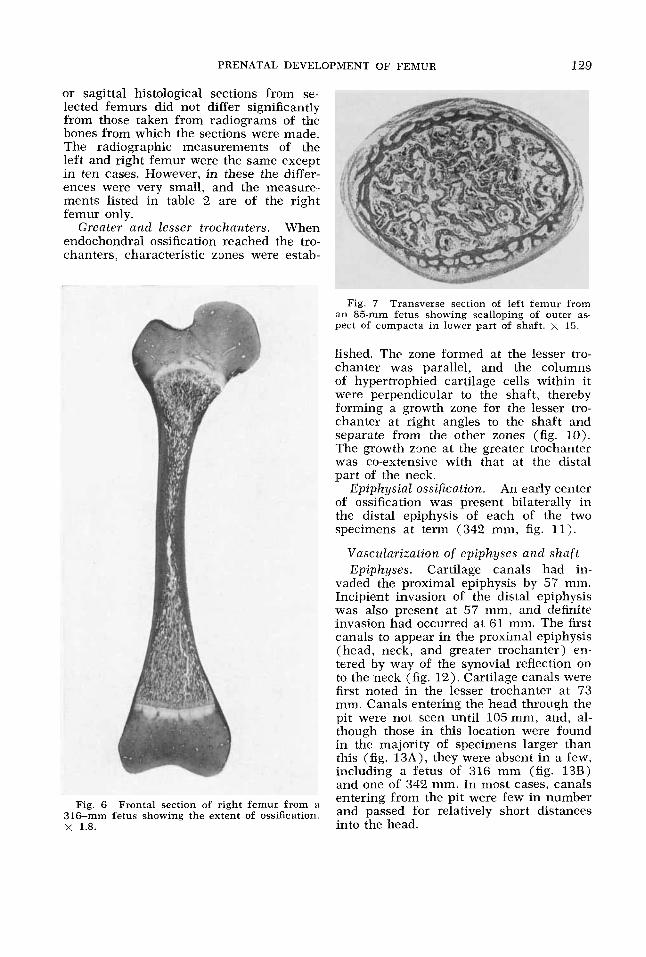
PRENATAL DEVELOPMENT OF FEMUR 129
or sagittal histological sections from se- lected femurs did not differ significantly from those taken from radiograms of the bones from which the sections were made. The radiographic measurements of the left and right femur were the same except in ten cases. However, in these the differ- ences were very small, and the measure- ments listed in table 2 are of the right femur only.
Greater and lesser trochnnters. When endochondral ossification reached the tro- chanters. characteristic zones were estab-
Fig. 7 Transverse section of left femur from an 85-mm fetus showing scalloping of outer as- pect of compacta in lower part of shaft. x 15.
lished. The zone formed at the lesser tro- chanter was parallel, and the columns of hypertrophied cartilage cells within it were perpendicular to the shaft, thereby forming a growth zone for the lesser tro- chanter at right angles to the shaft and separate from the other zones (fig. 10). The growth zone at the greater trochanter was co-extensive with that at the distal part of the neck.
Epiphysial ossification. An early center of ossification was present bilaterally in the distal epiphysis of each of the two specimens at term (342 mm, fig. 11).
Vascularization of epiphyses and shaf t Epiphyses. Cartilage canals had in-
vaded the proximal epiphysis by 57 mm. Incipient invasion of the distal epiphysis was also present at 57 mni, and definite invasion had occurred at 61 mm. The first canals to appear in the proximal epiphysis (head, neck, and greater trochanter) en- tered by way of the synovial reflection on to the neck (fig. 12). Cartilage canals were first noted in the lesser trochanter at 73 mm. Canals entering the head through the pit were not seen until 105mm, and, al- though those in this location were found in the majority of specimens larger than this (fig. 13A) , they were absent in a few, including a fetus of 316 mm (fig. 13B) and one of 342 mm. In most cases, canals entering from the pit were few in number and passed for relatively short distances
Fig. 6 Frontal section of right femur from a 316-mm fetus showing the extent of ossification. x 1.8. into the head.

130 ERNEST GARDNER AND D. J. GRAY
Fig. 8 Sectioiis showing in A, trabeculated compacta in a femur from a 148-mm fetus, x 14; in B, dense compacta and early osteons in a femur from a 277-mm fetus, x 6.3; in C, gross layering of com- pacts and early osteons in a femur from a 302-mmfetus, x 5.5.
By 61 mm and again at 77 mm, the margins of the cartilage canals of the upper end of the femur were calcified, and some of the canals had been invaded by periosteal bone from the region proxi- mal to the level of the zone of cartilage destruction (fig. 14). In some cases, a canal of the upper epiphysis constituted an invasion zone of cartilage destruction, but did not communicate with the endo- chondral area. In other instances, such a canal containing periosteally formed bone was incorporated into the endochondral area. In a fetus of 125 mm, trabeculated bone appeared in one of the canals. Bone was seen in the canals of the distal epi- physis for the first time at 85 mm (fig. 15), and appeared again at 103 and at 125 mm. Bone in cartilage canals was present with decreasing frequency in specimens larger than 150 mm, and none was observed in those larger than 200 mm.
Vascularization of the shaft be- gan with the cellular invasion of the pri- mary bony collar. The sites of the invasion did not coincide with the sites of the nutri- ent foramina and canals, which appeared later. The characteristic pattern of blood supply of the periosteal bone appeared early in the fetal period, with the forma- tion of trabeculae in the periosteal bone and the subsequent enclosure of vessels and associated tissues.
S h a f t .
A nutrient canal was first present at 57 mm. Considerable variation was found in the number, direction, and place of entry of the nutrient canals in the different femurs. The canal usually entered at some point along the linea aspera, but occa- sionally lateral or medial to i t . In the ma- jority of femurs a single canal was present, but in about one-fourth two were observed (fig. 16). Three canals were noted in one specimen and four in another. In more than half of the specimens, and especially in the younger ones, the canal passed obliquely downward to reach the marrow cavity. In less than half of the specimens, and particularly in the older ones, it passed horizontally forward. A canal directed obliquely upward was found in a few fe- murs, and only in those in which a second, descending canal was present. The forma- tion of the definitive canal or canals, which ascend to reach the marrow cavity, presumably occurs during postnatal de- velopment.
Remodeling The beginning of reconstruction, as in-
dicated by the presence of osteoclasts ex- ternal to the compacta, was found first at 92 mm on the upper part of the femur (at about the level of the lesser trochanter) and along the medial supracondylar line. At 103 mm, there were numbers of osteo-
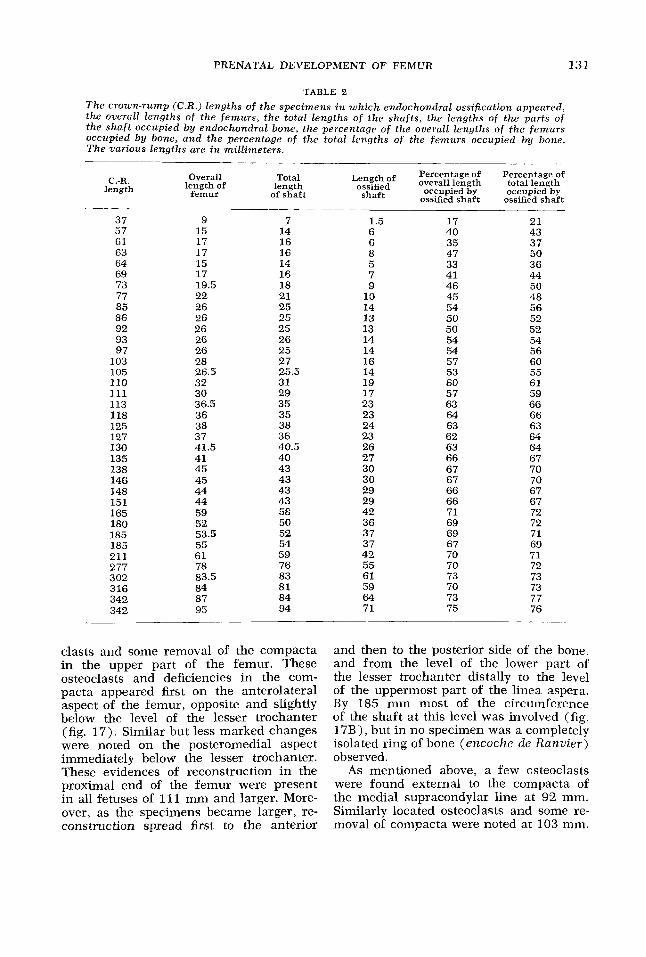
PRENATAL DEVELOPMENT OF FEMUR 131
'TABLE 2
T h e crown-rump (C.R.) lengths o f t h e specimens in w h i c h endochondral ossification appeared, t h e overall lengths o f t h e f e m u r s , t h e total lengths o f t h e shaf t s , t h e lengths o f t h e parts o f the s h a f t occupied b y endochondral bone, the percentage o f the overall lengths o f the f e m u r s occupied b y bone, and t h e percentage o f the total lengths o f the f e m u r s occupied b y bone. T h e various lengths are in mil l imeters .
Length of Percentage of Percentage of ossified overall length total length
occupied by occupied by ossified shaft ossified shaft
Overall Total length of length
femur of shaft shaft C.-R.
length
37 57 61 63 64 69 73 77 85 86 92 93 97
103 105 110 111 113 118 125 127 130 135 138 146 148 151 165 180 185 185 211 277 302 316 342 342
______
9 7 15 14 17 16 17 16 15 14 17 16 19.5 18 22 26 26 26 26 26 28 26.5 32 30 36.5 36 38 37 41.5 41 45 45 44 44 59 52 53.5 55 61 78 83.5 84 87 95
21 25 25 25 26 25 27 25.5 31 29 35 35 38 36 40.5 40 43 43 43 43 58 50 52 54 59 76 83 81 84 94
1.5 6 0 0 5 7 9
10 14 13 13 14 14 16 14 19 17 23 23 24 23 26 27 30 30 29 29 42 36 37 37 42 55 61 59 64 71
1 7 40 35 47 33 41 46 45 54 50 50 54 54 57 53 60 57 63 64 63 62 63 66 67 67 66 66 71 69 69 67 70 70 73 70 73 75
21 43 37 50 36 44 50 48 56 52 52 54 56 60 55 61 59 66 66 63 64 64 67 70 70 67 67 72 72 71 69 71 72 73 73 77 76
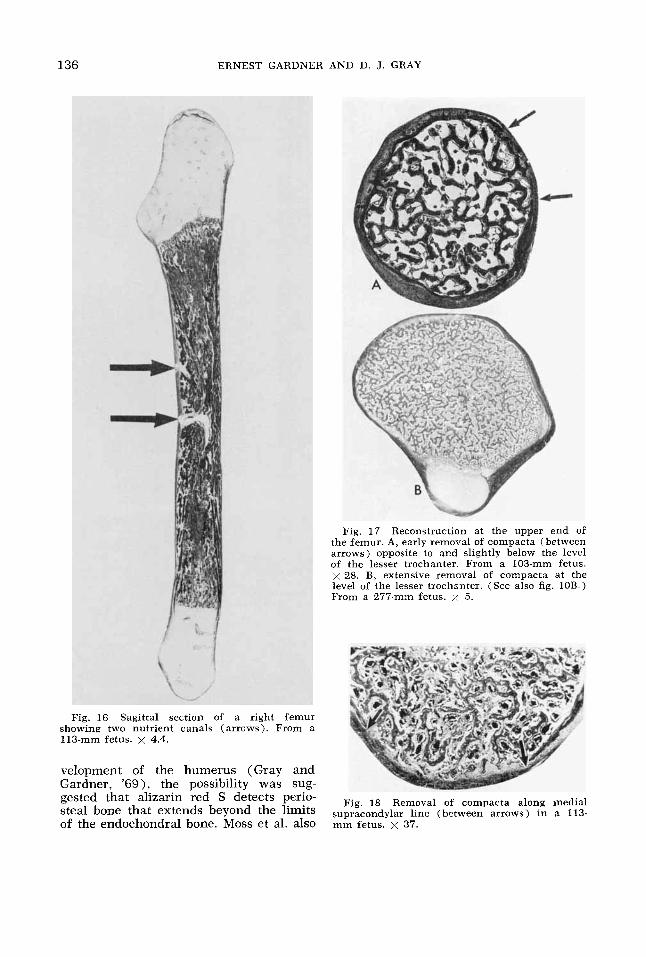
clasts and some removal of the compacta in the upper part of the femur. These osteoclasts and deficiencies in the com- pacts appeared first on the anterolateral aspect of the femur, opposite and slightly below the level of the lesser trochanter (fig. 17). Similar but less marked changes were noted on the posteromedial aspect immediately below the lesser trochanter. These evidences of reconstruction in the proximal end of the femur were present in all fetuses of 111 mm and larger. More- over, as the specimens became larger, re- construction spread first to the anterior
and then to the posterior side of the bone, and from the level of the lower part of the lesser trochanter distally to the level of the uppermost part of the linea aspera. By 185 mm most of the circumference of the shaft at this level was involved (fig. 17B), but in no specimen was a completely isolated ring of bone (encoche de Ranvier) observed.
As mentioned above, a few osteoclasts were found external to the compacta of the medial supracondylar line at 92 mm. Similarly located osteoclasts and some re- moval of compacta were noted at 103 mm.
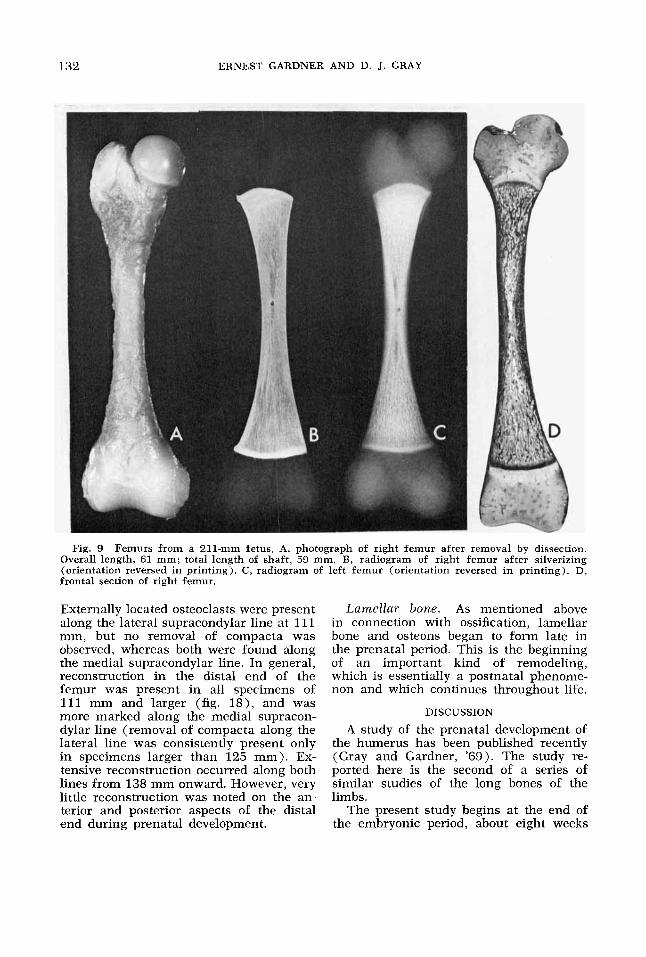
132 ERNEST GARDNER AND D. J. GRAY
Fig. 9 Femurs from a 211-mm fetus. A, photograph of right femur after removal by dissection. Overall length, 61 mm; total length of shaft, 59 mm. B, radiogram of right femur after silverizing (orientation reversed in printing). C, radiogram of left femur (orientation reversed in printing). D, frontal section of right femur.
Externally located osteoclasts were present along the lateral supracondylar line at 11 1 mm, but no removal of compacta was observed, whereas both were found along the medial supracondylar line. In general, reconstruction in the distal end of the femur was present in all specimens of 111 mm and larger (fig. 18), and was more marked along the medial supracon- dylar line (removal of compacta along the lateral line was consistently present only in specimens larger than 125 mm). Ex- tensive reconstruction occurred along both lines from 138 mm onward. However, very little reconstruction was noted on the a n terior and posterior aspects of the distal end during prenatal development.
LamellaT bone. As mentioned above in connection with ossification, lamellar bone and osteons began to form late in the prenatal period. This is the beginning of an important kind of remodeling, which is essentially a postnatal phenome- non and which continues throughout life.
DISCUSSION
A study of the prenatal development of the humerus has been published recently (Gray and Gardner, '69). The study re- ported here is the second of a series of similar studies of the long bones of the limbs.
The present study begins at the end of the embryonic period, about eight weeks
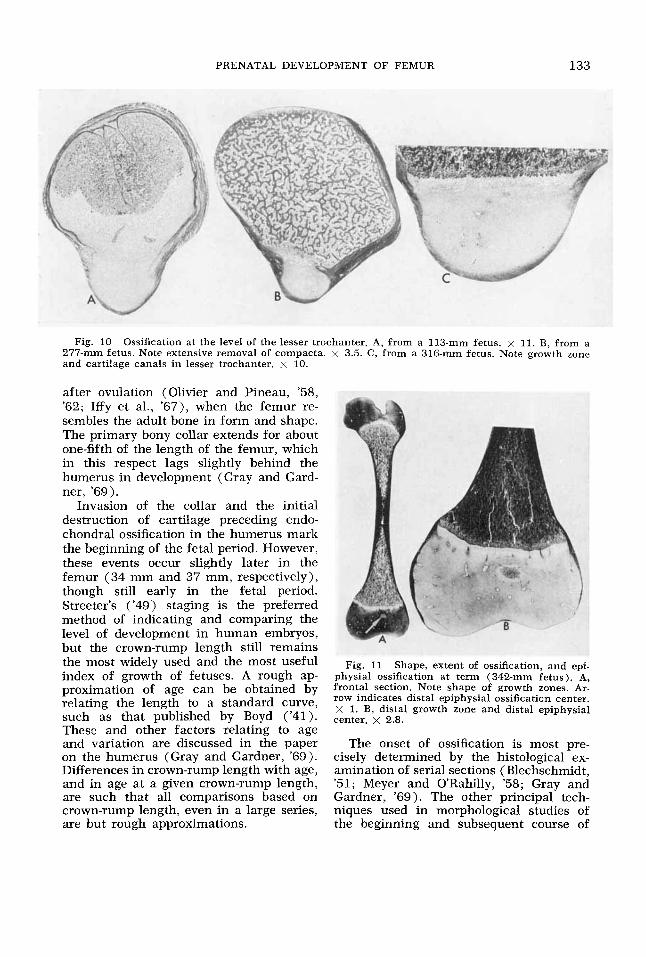
PRENATAL DEVELOPMENT OF FEMUR 133
Fig. 10 Ossification at the level of the lesser trochanter. A, from a 113-mm fetus. x 11. B, from a 277-mm fetus. Note extensive removal of compacta. x 3.5. C, from a 316-mm fetus. Note growth zone and cartilage canals in lesser trochanter. x 10.
after ovulation (Olivier and Pineau, '58, '62; Iffy et al., '67), when the femur re- sembles the adult bone in form and shape. The primary bony collar extends for about one-fifth of the length of the femur, which in this respect lags slightly behind the humerus in development (Gray and Gard- ner, '69 ) .
Invasion of the collar and the initial destruction of cartilage preceding endo- chondral ossification in the humerus mark the beginning of the fetal period. However, these events occur slightly later in the femur (34 mm and 37 mm, respectively), though still early in the fetal period. Streeter's ('49) staging is the preferred method of indicating and comparing the level of development in human embryos, but the crown-rump length still remains the most widely used and the most useful index of growth of fetuses. A rough ap- proximation of age can be obtained by relating the length to a standard curve, such as that published by Boyd ('41). These and other factors relating to age and variation are discussed in the paper on the humerus (Gray and Gardner, '69). Differences in crown-rump length with age, and in age at a given crown-rump length, are such that all comparisons based on crown-rump length, even in a large series, are but rough approximations.
Fig. 11 Shape, extent of ossification, and epi. physial ossification at term (342-mm fetus). A, frontal section. Note shape of growth zones. Ar- row indicates distal epiphysial ossification center. X 1. B, distal growth zone and distal epiphysial center. x 2.8.
The onset of ossification is most pre- cisely determined by the histological ex- amination of serial sections (Blechschmidt, '51; Meyer and ORahilly, '58; Gray and Gardner, '69). The other principal tech- niques used in morphological studies of the beginning and subsequent course of
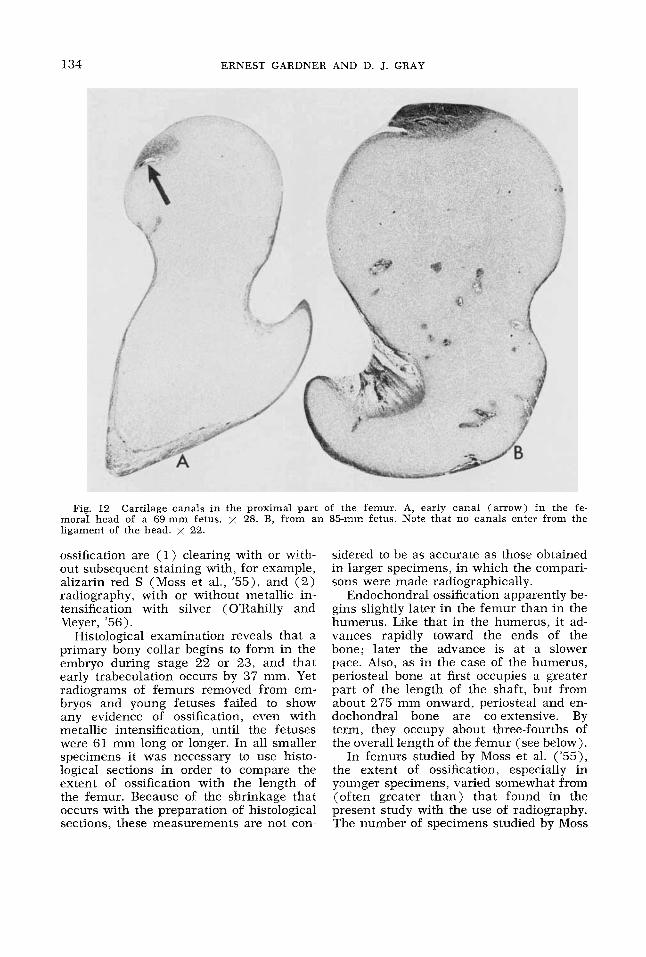
134 ERNEST GARDNER AND D. J. GRAY
Fig. 12 Cartilage canals in the proximal part of the femur. A, early canal (arrow) in the fe- moral head of a 69-mm fetus. x 28. B, from an 85-mm fetus. Note that no canals enter from the ligament of the head. x 22.
ossification are (1) clearing with or .with- out subsequent staining with, for example, alizarin red S (Moss et al., '55), and (2) radiography, with or without metallic in- tensification with silver (ORahilly and Meyer, '56).
Histological examination reveals that a primary bony collar begins to form in the embryo during stage 22 or 23, and that early trabeculation occurs by 37 mm. Yet radiograms of femurs removed from em- bryos and young fetuses failed to show any evidence of ossification, even with metallic intensification, until the fetuses were 61 mm long or longer. In all smaller specimens it was necessary to use histo- logical sections in order to compare the extent of ossification with the length of the femur. Because of the shrinkage that occurs with the preparation of histological sections, these measurements are not con-
sidered to be as accurate as those obtained in larger specimens, in which the compari- sons were made radiographically.
Endochondral ossification apparently be- gins slightly later in the femur than in the humerus. Like that in the humerus, i t ad- vances rapidly toward the ends of the bone; later the advance is at a slower pace. Also, as in the case of the humerus, periosteal bone at first occupies a greater part of the length of the shaft, but from about 275 mm onward, periosteal and en- dochondral bone are co-extensive. By term, they occupy about three-fourths of the overall length of the femur (see below).
In femurs studied by Moss et al. ('55), the extent of ossification, especially in younger specimens, varied somewhat from (often greater than) that found in the present study with the use of radiography. The number of specimens studied by Moss
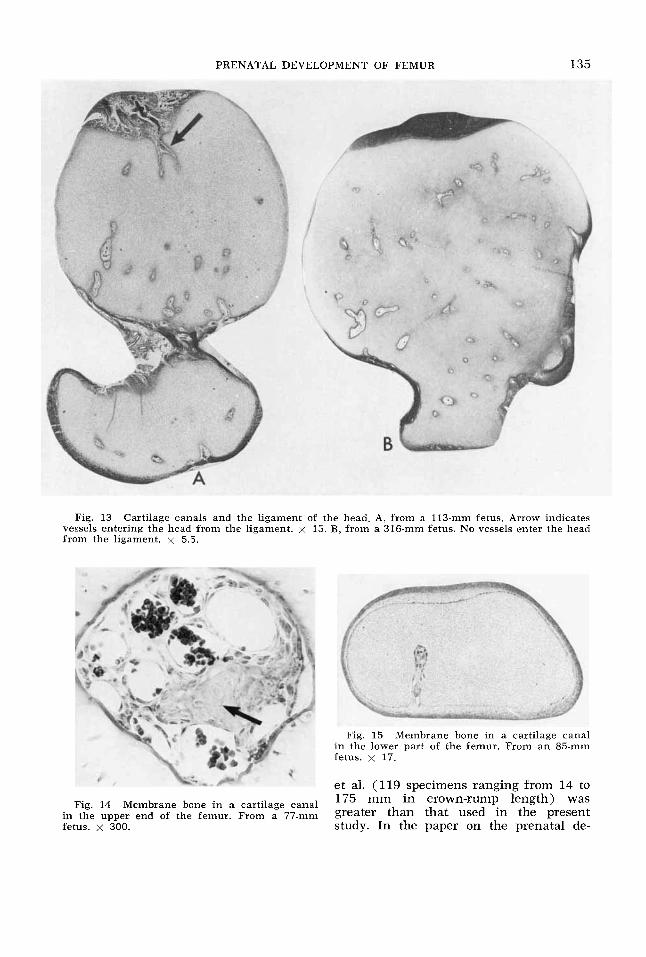
PRENATAL DEVELOPMENT OF FEMUR 135
Fig. 13 Cartilage canals and the ligament of the head. A, from a 113-mm fetus. Arrow indicates vessels entering the hcad from the ligament. x 15. B, from a 316-nim fetus. No vessels enter the head from the ligament. x 5.5.
Fig. 15 Membrane bone in a cartilage canal in the lower part of the femur. From an 85-mm fetus. x 17.
et al. (119 specimens ranging from 14 to 175 mm in crown-rump length) was
fetus. x 300. study. In the paper on the prenatal de-
Fig. 14 Membrane bone i n a cartilage canal in the upper end of the femur. From a 77-mm greater than that used in the present
136 ERNEST GARDNER AND D. J. GRAY
Fig. 17 Reconstruction at the upper end oE the femur. A, early removal of compacta (between arrows) opposite to and slightly below the level of the lesser trochanter. From a 103-mm fetus. x 28. B, extensive removal of compacta at the level of the lesser trochanter. (See also fig. 10B.) From a 277-mm fetus. x 5.
Fig. 16 Sagittal section of a right femur showing two nutrient canals (arrows). From a 113-mm fetus. x 4.4.
velopment of the humerus (Gray and Gardner, '691, the possibility was sug- gested that alizarin ;ed S detects perio-
of the endochondral bone. Moss et al. also
Fig, 18 Removal of compacts along medial steal bone that extends beyond the limits supracondylar line (between arrows) in a 113-
mm fetus. x 37.

PRENATAL DEVELOPMENT OF FEMUR 137
reported that pooling of data from all the long bones studied revealed that by 80 to 90 mm, ossification had reached the meta- physes, orderly growth zones had begun to form, and remodeling was under way. These findings were similar to those re- ported previously by Gray and Gardner ('50) and by Gardner and Gray ( '50) , and were corroborated in the present study. It is clear that major changes occur at this age, which is a t about the end of the first trimester or, in the case of the femur, early in the second trimester. Subse- quently, growth continues at a somewhat slower rate, but in an orderly and sym- metrical manner, contrary to the findings of Konig and Kornfeld ('27). The present series was not large enough, nor did it contain enough female specimens, to make possible the assessment of the sexual dif- ferences in the extent of diaphysial ossifi- cation reported by Halonen ('29).
By the time when growth rate slows and reconstruction begins, the femur contains a cavity, usually eccentrically placed, that is nearly or completely devoid of endo- chondral trabeculae. The compacta con- sists almost entirely of periosteal bone, and the metaphysis comprises a network of endochondral trabeculae derived from the growth zone. Many osteoclasts lie ad- jacent to the trabeculae, each of which has a core of calcified cartilage matrix. The fusion of a number of trabeculae with the compacta of the shaft results in a fusion of the shaft with the metaphysis and the presence of calcified cartilaginous matrix in the compacta.
As in the case of the humerus, perio- steal bone of the primary bony collar be- comes trabeculated soon after the collar forms. The characteristic markings of the femur, such as borders and elevations for muscle attachments, are at first thicken- ings of the periosteum that later become converted into periosteal bone. Lamellar bone and osteons begin to form when the fetus is well into the third trimester (Am- prino and Bairati, '36).
The complexity of femoral growth is illustrated by the orientation of the zones that will eventually become epiphysial plates, including the zone of the lesser tro- chanter. It is possible to assess their rela- tive contributions to growth in length, but
not their possible role in the development of angular features, including torsion. Most growth in length occurs a t the distal end, and epiphysial ossification has us- ually started there by term.
Vascularization of the epiphysial carti- lages consistently begins long before the onset of epiphysial ossification. Some of these vascular channels communicate with the diaphysis by penetrating the growth zone and area of replacement by bone. It is an interesting but unexplained finding that for a period of time, some channels contain bone. This bone may be traced to become continuous with periosteal bone and is sometimes found in a communicat- ing channel of the metaphysis.
The number and location of nutrient canals were similar to those in the adult bone, but even in older fetuses the canals seldom ascended to reach the marrow cavity. This change in direction takes place during postnatal development.
The process of remodeling is similar to that which occurs in the humerus. It is characterized by the appearance of osteo- clasts external to compacta, followed by the removal of compacta, where the shaft widens into the metaphysis. In the upper end of the femur, it begins first on the aspect of the bone opposite to and slightly below the level of the lesser trochanter, and also on the aspect of the bone im- mediately below the lesser trochanter. The process of removal of bone then spreads circumferentially, although a ring of perio- steal bone (encoche de Ranvier) is seldom completely isolated. In the lower end of the femur, reconstruction occurs first along the medial supracondylar line. Shortly thereafter, it appears along the lateral supracondylar line. In both loca- tions it spreads longitudinally and does not involve the anterior and posterior as- pects to any significant extent.
Remodeling in the femur therefore is under way by the end of the first trimester or early in the second, when the rate of ossification becomes slower and orderly growth zones begin to form. The first his- tological evidence is the appearance of osteoclasts external to the compacta on the upper part of the femur, at about the level of the lesser trochanter, and along the medial supracondylar line. The pres-

138 ERNEST GARDNER AND D. J. GRAY
ence of osteoclasts is regarded as the indi- cation of removal of bone, and their appearance is soon followed by loss of periosteal bone where they are grouped, a phenomenon first reported by Kolliker (1873). The appearance of osteoclasts, followed by loss of periosteal bone soon occurs elsewhere in the femur, for exam- ple, along the lateral supracondylar line. In general, this process is characterized by the external removal and internal de- position of bone in the metaphysis, the reverse of the process by which the shaft grows in width (external deposition and internal removal of bone). This process is necessary in order for the funnel-shaped metaphysis to become reduced to the width of the shaft (Lacroix, '51; Gardner, '56; Enlow, '63). Experimental confirmation of this process has been provided by Brash ('34), Leblond et al. ('50), and Leblond and Greulich ('56).
This kind of remodeling does not occur where the shaft does not widen upon reaching the metaphysis, as, for example, on the anterior and posterior aspects of the distal end of the femur. It may be modified where local features and muscu- lar and ligamentous attachments are de- veloping or changing (Enlow, '63; Hoyte and Enlow, '66). Moreover, the femur ex- hibits angular dimensions and shape that complicate matters. In his excellent study of femoral shape and growth before birth (53 femurs ranging in total shaft length from 8.4 mm to 110.6 mm, from fetuses and neonatal infants ranging from 31 mm to 485 mm crown-rump length), Felts ('54) reported detailed data on a number of dimensions. He distinguished between total length (from the infracondylar plane to the highest point on the greater tro- chanter) and overall length (from the infracondylar line to the highest point on the femoral head). He also used the terms proximal cartilage, distal cartilage, and ossified shaft, rather than epiphysis, meta- physis, and diaphysis. He divided the car- tilages in to in tracapsular , ex trac apsular , and juxta-osseous parts. He reported that the total length of the shaft increased by 0.21 mm for every millimeter increase in crown-rump length in embryos more than 31 mm crown-rump length and in femurs more than 8.4 mm long. Our data indicate
that the total length of the shaft increases by 0.285 mm for every millimeter increase in crown-rump length. Felts also reported that the length of the ossified portion of the shaft increased rapidly relative to total length (0.84 mm per 1.00 mm). By term, bone constituted about 80% and cartilage about 20%. The findings in the present study, as well as those of Bade ('00) and Nishizuka ('26), were similar. The above ratios refer to the total period of growth studied, and do not indicate changes in rate, such as Moss et al. reported. In general, Felts found that the ends of the femur became increasingly robust relative to the shaft (see his fig. 4). The greatest change during prenatal development was in the extent of ossification. The second greatest change was the increase in the transverse diameter across the proximal and distal cartilages. The least change was in the diameter of the ossified part of the shaft. Similar data, along with drawings, had been published by Nishizuka ('26).
Felts' study of femoral torsion, inclina- tion, and obliquity is also of particular interest. He found that torsion is present in the embryo, where it is often negative. Despite great individual variation, it is, on the average, much more positive at term than postnatally. This represents a con- tinuous change, a kind of remodeling that is still to be explained. Felts believes that torsion is present throughout the shaft. At the present time, there is no histological evidence available which explains the changes in torsion.
This study, like that of the humerus, shows that the time, location, and nature of the onset and subesequent course of ossification and remodeling are remarkably constant, and that the end of the first tri- mester or beginning of the second is a time of critical change.
LITERATURE CITED Altmann, F. 1925 Untersuchungen uber die
Torsio femoris und damit im Zusammenhang stehende Fragen. Z. Anat. EntwGesch., 75:
Amprino, R., and A, Bairati 1936 Processi di recostruzione e di riassorbimento nella sostanza compatta ossa dell'uomo. Richerche su cento sogetti dalla nascita sino a tarda etP. Z. Zell- forsch., 24: 439-511.
Amprino, R., and R. Cattaneo 1937 I1 substrato istologica delle varie modalitl di inserzioni alla
82-126.

PRENATAL DEVELOPMENT OF FEMUR 139
ossa nell’uomo. Z. Anat. EntwGesch., Abt. I, Z. ges. Anat., 107: 680-705.
Bade, P. 1900 Die Entwicklung des mensch- lichen Skelets bis zur Geburt. Arch. mikr. Anat. Entw., 55: 245-290.
Bardeen, C. R. 1905 Studies of the develop- ment of the human skeleton. Am. J. Anat., 4: 265-302.
Bardeen, C. R., and W. H. Lewis 1901 The de- velopment of the limbs, body-wall, and back. Am. J. Anat., I: 1-37.
Bertrand, P. 1923 Notes sur la vascularisation des epiphyses chez le foetus. C. R. Assoc. Anat., 18 Reunion, Lyon, 89-94.
Bidder, A. 1906 Osteobiologie. Arch. mikr. Anat. Entw., 68: 137-213.
Blechschmidt, E. 1951 Die friihembryonale Strukturentwicklung der Gliedmassen. Z. Anat. EntwGesch., 115: 617-657.
Boyd, E. 1941 Outline of Physical Growth and Development. Burgess, Minneapolis.
Brandt, G. 1928 Die Torsion der unteren Ex- tremitat und ihre Bedeutung fur die Deformi- tatenentstehung. Z. orthop. Chir., 49: 481-541.
Brash, J. C. 1934 Some problems in the growth and developmental mechanics of bone. Edinb. med. J., 41: 305-319, and 363-387.
Brookes, M. 1958 The vascularization of long bones in the human foetus. J. Anat., 92: 261- 276.
de Cuveland, E. F. 1950 Ein Beitrag zur Frage der Entstehung der Femurtorquierung im in- trauterinen Leben. Z. orthop. Chir., 44: 213- 223.
Digby, K. H. 1916 The measurement of dia- physial growth in proximal and distal direc- tions. J. Anat., 50: 187-188.
Enlow, D. H. 1963 Principles of Bone Remodel- ing. Charles C. Thomas, Springfield, Illinois.
Felts, W. J. L. 1954 The prenatal development of the human femur. Am. J. Anat., 94: 1 4 4 .
Friedlander, F. R. 1901 Ueber die Entstehung der angeborenen Huftverrenhung. Z. Anat. EntwGesch., 9: 515-543.
v. Friedlander, F. 1904 Beitrag zur Kenntnis der Architektur spongioser Knochen. Anat. Heft, 23: 237-282.
Gardner, E. 1956 Osteogenesis in the human embryo and fetus. In: The Biochemistry and Physiology of Bone. G. H. Bourne, ed. Academic Press, New York.
Gardner, E., and D. J. Gray 1950 Prenatal development of the human hip joint. Am. J. Anat., 87: 163-212.
Gardner, E., and R. O’Rahilly 1968 The early development of the knee joint in staged human embryos. J. Anat., 102: 289-299.
Gray, D. J., and E. Gardner 1950 Prenatal de- velopment of the human knee and superior tibiofibular joints. Am. J. Anat., 86: 235-288. - 1969 The prenatal development of the
human humerus. Am. J. Anat., 124: 431-446. Haines, R. W. 1947 The development of joints.
J. Anat., 81: 33-55. Halonen, L. 1929 Rijntgenologisch-anatomische
Untersuchungen uber die Entwicklung der Knochen der freien Extremiteten beim Men-
schen. I. Die Extremitenknochen der Feten. Acta Societatis Medicorum Fennicae “Duode- cim.” Suonalaisen Laakariseuran Duodecim ’in Toimituksia, Tom XI, Fasc. 3, 1-151, Helsinki.
Harris, H. A. 1929 The vascular supply of bone with special reference to the epiphysial carti- lage. J. Anat., 64: 3-4.
Hintzsche, E. 1928 Untersuchungen an Stiitz- geweben. 11. Uber Knochenbildungsfaktoren, insbesondere iiber den Anteil der Blutgefasse an der Ossifikation. Z. mikr.-anat. Forsch., 14:
Hoyte, D. A. N., and D. H. Enlow 1966 Wolff‘s law and the problem of muscle attachment to resorptive surfaces of bone. Am. J. phys. An- throp., 24: 205-214.
Humphrey, G. J. 1861 Observations on the growth of the long bones and of stumps. Med.- Chir. Tr., London, 44: 117-134.
Iffy, L., T. H. Shepard, A. Jacobovits, R. J. Lemire and P. Kerner 1967 The rate of growth in young human embryos of Streeter’s horizons XI11 to XXIII. Acta anat., 66: 178-186.
Knese, K.-H. 1957 Die diaphysare chondrale Osteogenese bis zur Geburt. Z. Zellforsch., 47: 80-113.
Kijlliker, A. 1873 Die normale Resorption des Knochengewebes und ihre Bedeutung fur die Entstehung der typischen Knochenformen. Vogel, Leipzig.
Konig, K., and W. Kornfeld 1927 Uber Sym- metrieund Langenverhaltnisse der verknocher- ten Skeletteile menschlicher Embryonen. Z . Anat. EntwGesch., 82: 657-693.
Lacroix, P. 1951 The Organization of Bones. Translated from the amended French edition by S. Gelder, Blakiston, Philadelphia.
Leblond, C. P., G. W. Wilkinson, L. F. Bklanger and J. Robichon 1950 Radio-autographic vis- ualization of bone formation in the rat. Am. J. Anat., 86: 289-341.
Leblond, C. P., and R. C. Greulich 1956 Auto- radiographic studies of bone formation and growth. In: The Biochemistry and Physiology of Bone. G. H. Bourne, ed. Academic Press, New York.
Le Damany, P. 1903 Les torsions osseuses. J. Anat., Paris, 39: 426-450.
Liitken, P. 1950 Investigations into the posi- tion of the nutrient foramina and the direction of the vessel canals in the shafts of the hu- merus and femur in man. Acta anat., 9: 57-68.
Meyer, D. B., and R. ORahilly 1958 Multiple techniques in the study of the onset of pre- natal ossification. Anat. Rec., 132: 181-194.
Moss, M. L., C. R. Noback and G. G. Robertson 1955 Critical developmental horizons in human fetal long bones. Am. J. Anat., 97: 155-175.
Nishizuka, T. 1926 Beitrage zur Osteologie der Foten, Neugeborenen, und Kinder nebst Er- wachsenen (Japoner). Knochen der Extremi- taten samt Schulter und Becken. Z. Morph. Anthr., 25: 1-90,
Olivier, G. 1962 Formation du Squelette des Membres chez l’Homme. Vigot Fritres, Bds., Paris.
373-440.

140 ERNEST GARDNER AND D. J. GRAY
Olivier, G . , and H. Pineau 1958 Determination de l’Pge du foetus et de l’embryon. Arch. anat. path., 6: A21-28.
1962 Horizons de Streeter et Lge em- bryonnaire. C. R. Assoc. Anat., 47: 573-576.
ORahilly, R., E. Gardner and D. J. Gray 1956 The ectodermal thickening and ridge in the limbs of staged human embryos. J. Embryol. Exp. Morphol., 4: 254-264.
1957 Chondrification in the hands and feet of staged human embryos. Contrib. Embryol., 36: 183- 192.
O’Rahilly, R., and D. B. Meyer 1956 Roent- genographic investigation of the human skele-
O’Rahilly, R., D. J. Gray and E. Gardner
ton during early fetal life. Am. J. Roentgen., 76: 455-468.
Pitzen, P. 1923 Das menschlichen Femur wahrend seiner Entwicklung. Arch. Anthrop., neue Folge, 19: 57-81.
Pryor, J. W. 1923 Differences in the time of development of centers of ossification in the male and female skeleton. Anat. Rec., 25:
Strayer, L. M. 1943 The embryology of the human hip joint. Yale J. Biol. Med., 16: 13-26.
Streeter, G. L. 1949 Developmental horizons in human embryos (fourth issue). A review of the histogenesis of cartilage and bone. Contrib. Embryol., 33 (220): 149-367. Publication No. 583, Carnegie Institution of Washington.
257-274.