Embed Size (px)
Citation preview

THE JOURNAL cm BIOLOGICAL CHEMISTRY Vol. 233, No. 12, December 1963
Printed in U.S.A.
The Porcine Pancreatic Carboxypeptidase A System*
I. THREE FORMS OF THE ACTIVE ENZYME
J. E. FOLK AND E. W. SCHIRMER
From the National Institute of Dental Research, National Institutes of Health, Public Health Service, United States Department of Health, Education, and Welfare, Bethesda 14 Maryland
(Received for publication, June 20, 1963)
Carboxypeptidase A was first isolated in crystalline form from beef pancreas by Anson in 1935 (2). Extensive enzymatic studies have defined this enzyme as an exopeptidase that is specific for catalyzing the cleavage of certain carboxyl-terminal peptide bonds in peptides and proteins (3, 4). Fresh pancreas tissue and pancreatic juice do not contain the active enzyme, but rather an inactive precursor, procarboxypeptidase A (5, 6). Several reports on the mechanism of procarboxypeptidase A activation indicate that chymotrypsin, as well as trypsin, ac- tivity is necessary for activation (7) and that activation is accompanied by extensive peptide bond cleavage in the zymogen (6, 7). There are also reports that the zymogen is a complex of three subunits (8) and the precursor of two different enzymes, carboxypeptidase A and a chymotrypsin-like endopeptidase (7).
Marchis-Mouren et uZ. (9) have reported that porcine pan- creatic juice is a rich source of procarboxypeptidase A. Pre- liminary studies in our laboratory showed the active enzyme fraction from this species to be extremely water-soluble and resistant to inactivation and, hence, ideally suited for studies on the mechanism of proenzyme activation. During the course of activation studies, it became evident that the activity com- monly ascribed to carboxypeptidase A was associated with three electrophoretically distinct proteins. The details of experi- ments leading to the recognition of these three forms of the porcine enzyme, designated carboxypeptidases Ai, A,, and Aa, are the subject of the present communication. The purification and some of the physical and enzymatic properties of carboxy- peptidases A1 and Az are also given. A mechanism for the trypsin-catalyzed conversion of carboxypeptidase Ai to carboxy- peptidase Az is presented in the accompanying report (10).
EXPERIMENTAL PROCEDURE AND RESULTS
Assay and Analytical Procedures-The method employed to measure carboxypeptidase A activity during the isolation and purification procedures is a differential spectral assay similar to that outlined previously for carboxypeptidase B (11, 12), except that the carboxypeptidase A substrate, N-benzoylglycyl-n- phenylalanine (hippuryl-r.-phenylalanine) , was used in place of the carboxypeptidase B substrates. The substrate was arbi- trarily employed at 1 x 10M4 M concentration in 0.025 M Tris- chloride, pH 7.5, containing 0.5 M NaCl in a total volume of 20 ml. The measurements were carried out at 25” in 50-mm cylindrical cells in a Cary model 11 recording spectrophotometer
* Presented in part before the Forty-seventh Annual Meeting of the American Society of Biological Chemists, Atlantic City, New Jersey, April 16 to 20, 1963 (1).
at 254 rnp. Activity is expressed as change in absorbance per minute, and specific activity, as activity per mg of enzyme protein per ml.
Rate measurements used in kinetic analyses were carried out with the peptide substrate, hippuryl-tphenylalanine, and with the ester substrate, hippuryl-nn-phenyllactic acid, by the above procedure with the use of absorption cells of different light path lengths in the manner outlined previously (11). Rates with carbobenzoxyglycyl-L-phenylalanine and carbobenzoxyglycyl- n-leucine were obtained by means of a direct spectrophotometric assay at wave lengths at which loss in absorbance of readily hydrolyzed bonds is measured (13). With carbobenzoxyglycyl- n-tryptophan, the photometric ninhydrin procedure (14) was employed for rate measurements. Reaction rates were esti- mated and kinetic constants calculated as outlined previously (11).
Procarboxypeptidase A content was measured as the increase in carboxypeptidase A activity produced upon incubation with trypsin. Standard activation mixtures contained 0.05 to 1.0 mg of protein per ml and 0.125 mg of trypsin per ml in 0.025 M Tris-chloride, pH 8.0, at 25’. At this trypsin concentration, full activation was observed in all cases within 2 to 3 minutes, and there was no loss in activity at 30 minutes. Activation mixtures were routinely incubated for 10 minutes to ensure full activation.
The disk electrophoresis method of Ornstein and Davis’ was used to analyze enzyme preparations for homogeneity, to study molecular conversions, and to evaluate the results of fractiona- tion procedures. All migrations were carried out toward the anode on 7.5% cross-lied polyacrylamide gel at pH 9.5. Runs were made at room temperature in columns, 0.5 x 7 cm, for 60 to 80 minutes at 3 ma per column. By varying the polyacryl- amide concentration of the sample gel, 50 to 200 I.rg of sample in volumes of 5 to 200 ~1 could be accommodated.
Mati%&Hippuryl-L-phenylalanine [a]? +39” (c, 5, in 95% ethanol), carbobenzoxyglycyl-L-phenylalanine [cu]? +38” (c, 6, in 95% ethanol), carbobenzoxyglycyl-L-leucine [a]? -9” (c, 6, in 95% ethanol), and carbobenzoxyglycyl-L-tryptophan were obtained from Mann Research Laboratories, Inc., New York. Hippuryl-ncphenyllactic acid was supplied by Cycle Chemical Corporation, Los Angeles, California. Each of the compounds listed gave satisfactory values for elemental compo- sition and showed no gross contamination upon examination by paper chromatography.
1 Procedures, materials, and chemical formulations for disk electrophoresis and details of disk electrophoresis theory are avail- able from Canalco (Canal Industrial Corporation), 4935 Cordell Avenue, Bethesda, Maryland.
3884
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

December 1963 J. E. Folk and E. W. Schirmer 3885
Bovine trypsin, twice crystallized and salt-free, was purchased from Worthington Biochemical Corporation, Freehold, New Jersey.
DEAE-cellulose (standard, 0.94 meq per g) was obtained from Carl Schleicher and Schiill Company, Keene, New Hamp- shire. Sephadex G-25 (medium) was a product of Pharmacia, Uppsala, Sweden.
Preparation of Partially Purified Procarboxypeptidase A
Preparation of Acetone Powders-Freshly excised whole swine pancreas glands were obtained from the slaughterhouse, frozen, and stored at -20”. The solidly frozen glands, 500 to 700 g, were broken into pieces approximately 4 to 7 cm in diameter and were dropped into 2000 ml of acetone at room temperature in a large Waring Blendor operating at full speed. Homogeniza- tion was continued for 1 minute at full speed. The resulting suspension was filtered on a large Buchner funnel. The residue was re-extracted successively in the Blendor at full speed for 1 minute twice with 1200 ml of acetone, once with 1200 ml of acetone-ether (1 :l), and once with 1200 ml of ether. The defatted tissue was spread out and allowed to dry at room temperature and finally stored in plastic bags at 5”.
Preparation of Extracts and Ammonium Sulfate Fractionation- Water extracts of acetone powders were prepared by stirring the powder, 100 g, with cold distilled water, 1000 ml, for 45 minutes at O-2”. All further operations were carried out at this tempera- ture. The suspension was centrifuged for 30 minutes at 14,600 x
g, and the light pink turbid supernatant fluid was collected. Extracts prepared in this manner contained only 5 to 150/, of the carboxypeptidase A in active form. The extracts were adjusted to pH 7.2 by the addition of 1 N NaOH, and solid ammonium sulfate was added gradually with stirring to 0.35 saturation (209 g per liter). The mixture was maintained be- tween pH 7.0 and 7.2 with 1 N NaOH during the addition of salt and was allowed to stand for 30 minutes. The suspension was then centrifuged for 30 minutes at 14,600 x g. The super- natant fluid was discarded, and the protein precipitate was dissolved in 250 ml of distilled water. To this protein solution were added gradually with stirring 50 ml of a saturated am- monium sulfate solution (saturated at 0” and adjusted to pH 7.2). After a 30-minute equilibration period, the precipitate was re- moved by centrifugation. To each 100 ml of supernatant fluid were added 22.5 ml of saturated ammonium sulfate solution; the suspension was stirred for 30 minutes and then centrifuged. The precipitate was dissolved in the minimal volume of 0.005 M Tris-chloride buffer, pH 8.0 (total volume, 140 to 200 ml). To this protein solution were added 20 mg of soybean trypsin inhibitor, and the solution was freed of ammonium sulfate by passage in two separate and equal portions through a column, 3.5 X 40 cm of Sephadex G-25 equilibrated with 0.005 M Tris- chloride buffer, pH 8.0. Attempts to desalt the protein solution by dialysis or Sephadex treatment in the absence of trypsin inhibitor resulted in complete conversion of the zymogen to active carboxypeptidase A. Desalted solutions of this protein mixture have been stored frozen at -20’ for as long as 3 months with no change in zymogen content.
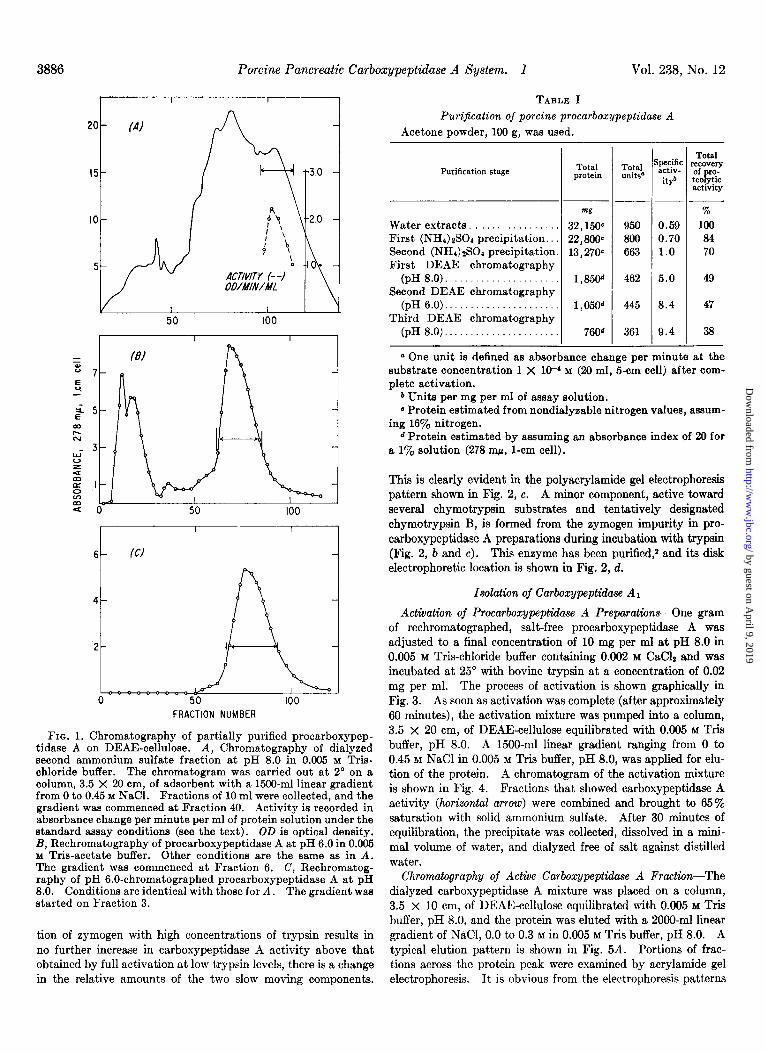
Chromatography on DEAE-cellulose-The combined desalted protein solutions, 350 to 400 ml, were pumped through a column, 3.5 x 20 cm of DEAE-cellulose equilibrated with 0.005 M
Tris-chloride buffer, pH 8.0, at a flow rate of 20 to 30 ml per
minute. A 1500-ml linear gradient ranging from 0 to 0.45 M
NaCl in 0.005 M Tris-chloride buffer, pH 8.0, was applied for elution of the protein. Fractions of 10 ml were collected at a flow rate of 6 to 8 ml per minute. Small portions of the frac- tions were diluted with the Tris buffer in order to measure the absorbance at 278 rnp. Fractions containing the major portion of the procarboxypeptidase A (those fractions containing more than 25% of the activity of the peak fraction) were combined as indicated by the horizontal arrow in Fig. 1A. Protein was precipitated by the addition of solid ammonium sulfate, 43 g per 100 ml. After a 30-minute equilibration period, the precipi- tate was collected by centrifugation, taken up in a minimal volume of water (40 to 50 ml), and dialyzed overnight against distilled water. This salt-free protein solution was rechromat- ographed on a column, 3.5 x 20 cm, of DEAE-cellulose equili- brated with 0.005 M Tris-acetate, pH 6.0. A 1500-ml linear gradient ranging from 0 to 0.45 M NaCl in 0.005 M Tris-acetate, pH 6.0, was applied for elution. A typical chromatogram is shown in Fig. 1B. Those fractions containing the major portion of the zymogen, shown by the horizontal arrow in Fig. lB, were combined. The protein was precipitated with ammonium sulfate, collected, dissolved in water, and dialyzed against water as before. Rechromatography at pH 8.0, under the conditions given above, yielded a single peak, which is pictured in Fig. 1C. The fractions indicated by the arrow were combined, treated with ammonium sulfate, 43 g per 100 ml, and stored at 5” as the ammonium sulfate precipitate. The procarboxypeptidase A has been stored in this manner for 2 to 3 months with no loss in zymogen content. This precipitate of partially purified pro- carboxypeptidase A was collected by centrifugation, dissolved in water, and dialyzed overnight against water before it was converted to active enzyme. In Table I are recorded the total recoveries of activity and enrichment of the eymogen.
Properties of Partially Purified Procarboxypeptidase A
Although all preparations of procarboxypeptidase A chromat- ographed as a single peak at both pH 6 and pH 8 on DEAE- cellulose, it was obvious from disk electrophoresis studies that all of the preparations contained substantial quantities of two proteins (Fig. 2a). Incubation of samples of the partially puri- fied procarboxypeptidase A with trypsin gave rise to activity toward the chymotrypsin substrates, benzoyl-L-tyrosine ethyl es- ter, and acetyl-r-tyrosine ethyl ester, as well as toward carboxy- peptidase A substrates. Gel filtration procedures, to be detailed in a forthcoming communication, have been developed for the separation of the two protein components of this partially purified zymogen preparation2 The faster moving component (Fig. 2a) upon incubation with trypsin gives rise to activity characteristic of chymotrypsin and is probably the anionic chymotrypsinogen B that has been postulated by Rovery et al. (15) to occur in porcine pancreatic tissue extracts. Tryptic activation of the slow mov- ing component (Fig. 2a), purified procarboxypeptidase A, on the other hand is not accompanied by the formation of any detectable chymotrypsin-like activity.2 This is in contradistinction to the reports that chymotrypsin-like activity arises during the tryptic activation of purified bovine procarboxypeptidase A (7, 8).
Full activation of partially purified procarboxypeptidase A preparations with low concentrations of trypsin gives rise to two major components with mobilities lower than those of the zymogen constituents, as shown in Fig. 2b. Although incuba-
2 J. E. Folk and E. W. Schirmer, unpublished data.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
3886 Porcine Pancreatic Carboxypeptidase A System. I Vol. 238, Tu’o. 12
TABLE I
Purification of porcine procarboxypeptidase A Acetone powder, 100 g, was used.
FRACTION NUMBER
FIG. 1. Chromatography of partially purified procarboxypep- tidase A on DEAE-cellulose. A, Chromatography of dialyzed second ammonium sulfate fraction at pH 8.0 in 0.005 M Tris- chloride buffer. The chromatogram was carried out at 2” on a column, 3.5 X 20 cm, of adsorbent with a 1500-ml linear gradient from 0 to 0.45 M NaCl. Fractions of 10 ml were collected, and the gradient was commenced at Fraction 40. Activity is recorded in absorbance change per minute per ml of protein solution under the standard assay conditions (see the text). OD is optical density. B, Rechromatography of procarboxypeptidase A at pH 6.0 in 0.005 Y Tris-acetate buffer. Other conditions are the same as in A. The gradient was commenced at Fraction 6. C, Rechromatog- raphy of pH 6.0-chromatographed procarboxypeptidase A at pH 8.0. Conditions are identical with those for A. The gradient was started on Fraction 3.
tion of zymogen with high concentrations of trypsin results in no further increase in carboxypeptidase A activity above that obtained by full activation at low trypsin levels, there is a change in the relative amounts of the two slow moving components.
Purification stage Total Total protein units”
. _ mg
Water extracts. . 32,15&
First (NHd)zSOd precipitation.. 22,800~ Second (NHI)~SO~ precipitation. 13,270~ First DEAE chromatography
(pH 8.0). l&W Second DEAE chromatography
(pH 6.0).. 1,050d Third DEAE chromatography
(pH 8.0). 760d -
jpecific activ-
it?
Total recovery of ro- tea ytic P activity
950 800
663
462
445
361
%
0.59 100
0.70 84 1.0 70
5.0 49
8.4 47
9.4 38
a One unit is defined as absorbance change per minute at the substrate concentration 1 X 10-d M (20 ml, 5-cm cell) after com- plete activation.
b Units per mg per ml of assay solution. 0 Protein estimated from nondialyzable nitrogen values, assum-
ing 16% nitrogen. d Protein estimated by assuming an absorbance index of 20 for
a 1% solution (278 q, l-cm cell).
This is clearly evident in t&e polyscrylamide gel electrophoresis pattern shown in Fig. 2, c. A minor component, active toward several chymotrypsin substrates and tentatively designated chymotrypsin B, is formed from the zymogen impurity in pro- carboxypeptidase A preparations during incubation with trypsin (Fig. 2, b and c). This enzyme has been purified,2 and its disk electrophoretic location is shown in Fig. 2, d.
Isolation of Carboxypeptidase A1
Activation of Procarboxypeptidase A Preparations-One gram of rechromatographed, salt-free procarboxypeptidase A was adjusted to a final concentration of 10 mg per ml at pH 8.0 in 0.005 M Tris-chloride buffer containing 0.002 M CaClz and was incubated at 25” with bovine trypsin at a concentration of 0.02 mg per ml. The process of activation is shown graphically in Fig. 3. As soon as activation was complete (after approximately 60 minutes), the activation mixture was pumped into a column, 3.5 x 20 cm, of DEAE-cellulose equilibrated with 0.005 M Tris buffer, pH 8.0. A 1500-ml linear gradient ranging from 0 to 0.45 M NaCl in 0.005 M Tris buffer, pH 8.0, was applied for elu- tion of the protein. A chromatogram of the activation mixture is shown in Fig. 4. Fractions that showed carboxypeptidase A activity (horizontal arrow) were combined and brought to 65% saturation with solid ammonium sulfate. After 30 minutes of equilibration, the precipitate was collected, dissolved in a mini- mal volume of water, and dialyzed free of salt against distilled water.
Chromatography of Active Carboxypeptidase A Fraction-The dialyzed carboxypeptidase A mixture was placed on a column, 3.5 x 10 cm, of DEAE-cellulose equilibrated with 0.005 M Tris buffer, pH 8.0, and the protein was eluted with a 2000-ml linear gradient of NaCI, 0.0 to 0.3 M in 0.005 M Tris buffer, pH 8.0. A typical elution pattern is shown in Fig. 5A. Portions of frac- tions across the protein peak were examined by acrylamide gel electrophoresis. It is obvious from the electrophoresis patterns
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

December 1963 J. E. Folk and E. W. Schirmer
FIN. 2. Disk electrophoresis patterns of partially purified por- cine procarboxypeptidase A and its activation products. Experi- mental conditions for electrophoresis are given in the text. The arrow and F designate the positions of the starting points and buffer fronts, respectively. a, Procarboxypeptidase A, 10 pl of a solution containing 20 mg per ml. 5, Procarboxypeptidase A, 10 mg per ml; porcine trypsin, 0.02 mg per ml; 30 minutes at 25”;
WLU 0 40 60
TIME IN MINUTES FIG. 3. Activation of partially purified procarboxypeptidase
A. The conditions of activation: procarboxypeptidase A, 10 mg per ml; bovine trypsin, 0.02 mg per ml in 0.005 M Tris-chloride buffer, pH 8.0, containing 0.002 M CM&; incubation at 25”.
0.02 M Tris buffer, pH 8.0; 20 ~1 applied. c, Procarboxypeptidase A, 10 mg per ml; porcine trypsin, 0.4 mg per ml; other conditions as in b. d, Purified chymotrypsin B, 10 ~1 of a solution containing 15 mg per ml. e, Purified carboxypeptidase A1 (three times crys- tallized), 10 ~1 of a solution containing 10 mg per ml. 3, Purified carboxypeptidase A?, 10 ~1 of a solution containing 10 mg per ml.
shown in Fig. 5B that partial separation of the two active com- ponents of trypsin-treated procarboxypeptidase A has been obtained. The specific activity of the enzymes in representative fractions across the protein peak was 36 f 2. The fractions rich in the slower migrating component, designated carboxy- peptidase A,, and low in the faster moving carboxypeptidase, designated carboxypeptidase AZ, as shown by the horizontal arrow, were combined, treated with solid ammonium sulfate to 65% saturation, and stored at 5”. The remaining fractions containing carboxypeptidase activity were combined and treated with ammonium sulfate as above. The protein precipitate was collected, dissolved in water, dialyzed against distilled water, and rechromatographed as before. Again those fractions rich in carboxypeptidase A1 and essentially free of the A) form were pooled and treated with ammonium sulfate. This operation was repeated twice more. The fractions rich in carboxypeptidase Az were combined, treated with ammonium sulfate, and set aside at 5” for preparation of the A2 enzyme. The preparations of carboxypeptidase A1 from the individual chromatograms were pooled, and the protein precipitate was collected and dialyzed against distilled water. Finally the enzyme was chromato-
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3888 Porcine Pancreatic Carboxypeptidase A System. I Vol. 238, No. 12
I I I I I
= 6 z t
FRACTION NUMBER
FIG. 4. Chromatography of procarboxypeptidase A activation mixture on DEAE-cellulose. The chromatogram was carried out at 2” with 1 g of activated procarboxypeptidase A on a column, 3.5 X 20 cm, of adsorbent with a 1500-ml linear gradient from 0 to 0.45 M NaCl at pH 8.0. Fractions of 10 ml were collected at a flow rate of 6 to 8 ml per minute, and the gradient was commenced at Fraction 20.
FIG. 5. A, Chromatographic separation of carboxypeptidase AI on DEAE-cellulose. The active carboxypeptidase A fraction from a chromatogram of the procarboxypeptidase A activation mixture (Fig. 4) was rechromatographed at 2” on a column, 3.5 X 10 cm, of DEAE-cellulose with a 200-ml linear gradient from 0 to 0.3 M NaCl at pH 8.0. The gradient was commenced at tube 4; 30-ml fractions were collected in the first 30 tubes, and lo-ml frac- tions in the remaining tubes at a flow rate of 6 to 8 ml per minute. The gradient of NaCl is shown by the dashed line. Specific ac- tivity was determined on fractions designated by the solid points. The fractions indicated by the horizontal arrow were combined for preparation of carboxypeptidase A1 after examination by disk electrophoresis (B). The electrophoresis patterns (B) correspond to the fractions designated by the solid points in the chromato-
gram (A). The pattern at the far right was obtained with a sample of the unfractionated active carboxypeptidase A mixture.
graphed on a column, 1.5 x 15 cm, of DEAE-cellulose equili- brated with 0.005 M Tris-acetate buffer, pH 6.0, and a 300-ml linear gradient ranging from 0 to 0.3 M NaCl in 0.005 M Tris- acetate, pH 6.0, was employed for elution.
Crystallization cf Carboxypeptidase Al-The combined car- boxypeptidase Al fractions from the pH 6.0 chromatogram were brought to 65% saturation with solid ammonium sulfate. The precipitate was taken up in a small volume of water and dialyzed against distilled water with frequent changes to fresh distilled water. After several hours, the enzyme began to crystallize. Dialysis was continued for 24 hours, at which time the crystals were removed by centrifugation and washed with distilled water. For recrystallization, the crystals were dissolved in distilled water by addition of 0.1 N NaOH at pH 6.6 to 6.8. A small amount of insoluble material was removed by centrifugation, and the solution of enzyme was either dialyzed against distilled water or lowered in pH to 5.2 to 5.4. In either case, crystals of the type shown in Fig. 6 were obtained. The yield of thrice crystallized enzyme from 1 g of partially purified proenzyme was approximately 50 mg.
E u
-
EJ2- I
e
jo.2
cd w- 0
Y+ I- f
s /
K,
g -0.1
8 / z
% Q /
/
FRAC%N NUMBER
100
FIG. 5A
FIG. 5B
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

December 1963 J. E. Folk and E. W. Schirmr 3889
FIQ. 6. Crystalline habit of porcine carboxypeptidsse A, (three times crystallized, approximately 275X).
Prqwties of Crystalline Carboxypeptidase A1
Carboxypeptidsse Ai can be stored indefinitely as a suspension of crystals in water at 5” under toluene vapors. The crystals are readily soluble in dilute buffers of pH 6 to 9 or in water upon adjustment of the pH to 6.4 to 6.8 with dilute sodium hydroxide. The enzyme showed a specific activity of 37.0 f 1.0 and dis- played a typical protein absorption spectrum in the ultraviolet range with a maximal extinction at 278 mE.r. The absorbance index at 278 rnp of a 1 Y0 solution was found to be 19.6, and the molar absorbance index was calculated as 6.72 x 104, based on the 34,800 molecular weight value reported below.
The sedimentation pattern illustrated in Fig. 7 is consistent with homogeneity. Four concentrations of carboxypeptidase Ar, ranging from 2 to 8 mg of protein per ml, showed increasing sedimentation coefficients with increasing protein concentration. Linear extrapolation to zero protein concentration yielded an s20,W of 3.34 S. A molecular weight of 34,800 i 600 was deter- mined by means of short column sedimentation equilibrium (16), assuming a partial specific volume of 0.73 cm3 per g.
A typical disk electrophoresis pattern of three times crys- tallized carboxypeptidase A1 is pictured in Fig. 2e, and the schlieren diagram of the ascending boundary of a moving bound- ary electrophoresis run of this material is shown in Fig. 8. The descending boundary of each of t’he active carboxypeptidase A preparat,ions showed estraordinary diffusion after short periods of electrophoresis. This may be seen in Fig. 11. The cause of this spreading of the descending boundary is unknown. The mobility of the ascending boundary at 0’ for carboxypeptidase At was computed to be -2.73 X low6 cm2 volt-r set-’ in 0.05 M
T&-chloride buffer with 0.1 M KaCl, pH 8.0. The kinetic constants obtained with carboxypeptidase A1 for
several peptide substrates and an ester substrate are given in Table II. For comparative purposes, values obtained with other forms of porcine carboxypeptidase A and with crystalline bovine carboxypeptidase A under the same experimental conditions are also recorded in this table. It is of interest that, with the present differential spectral method of assay, rate measurements can be carried out on the ester substrate, hippuryl-nn-phenyllactic acid, at low substrate concentrations that are not inhibitory toward the enzymes. Plots of substrate concentration versus velocity of
FIG. 7. Schlieren diagram of an ultracentrifuge run of carboxy- peptidase A1 in a Kel-F atmlytical cell at 25.0” after 91 minutes at 59,780 r.p.m. Protein was 0.975% in 0.65 M Tris-chloride buffer with 0.1 M NaCl, pH 7.0. Sedimentation proceeded from left to right.
hydrolysis at these low substrate concentrations conformed to the requirements for normal rate equations and showed that, in the case of each enzyme, inhibition by substrate occurred at hip- puryl-nn-phenyllactic acid concentrations above 5 X 10F4 M.
Assays for carbosypeptidase B in samples of carboxypeptidase A1 showed no trace of this activity under conditions that would measure as little as 0.01 y0 of this enzyme in the crystalline sam- ples.
The zinc content of carbosypeptidase A1 was determined on the assumption that the porcine enzymes, like bovine carboxy- peptidase A, are zinc metallo-enzymes. Values of 1.07 and 0.91 moles of zinc per mole of enzyme (mol. wt., 34,800) were found with the use of two samples of the crystalline enzyme and a chem- ical procedure (17) with samples prepared by the wet ashing technique. The metallo-enzyme nature of porcine carboxypep- tidase A1 was further substantiated by inhibition studies with the metal-chelating agent, 1, IO-phenanthroline. Preincubation of the enzyme in the presence of 1 x lo-” M 1, IO-phenanthroline for 40 minutes at 25” and measurement in substrate solution, 10 x IO-’ M, 0.5-cm cell containing the same inhibitor concen- tration resulted in 40% inhibition of activity, whereas no in- hibition was observed in the presence of t’he same inhibitor con- centration without preincubation.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3890 Porcine Pancreatic Carboxypertidase A System. I Vol. 238, Ko. 12
FIG. 8. Schlieren diagram of the ascending boundary in an electrophoresis run of carboxypeptidase Ai in an Aminco-Stern instru- ment. The conditions were: 1.0% protein in 0.05 M Tris-chloride buffer with 0.1 M NaCl, pH 8.0; 2”; 4.1 volts per cm for 209 minutes. Movement was from left to right.
TABLE II Kinetic constants for carboxypeptidase A
Assays were conducted at 25” in 0.025 M Tris-chloride buffer, pH 7.5, containing 0.5 M NaCl. Values for ko were calculated with the assumption that there was one active site per molecular weight of 34,800 where V,,. = koEl and K,,, = (k-1 + ko)/kl in the general
4 scheme for enzyme catalysis E + S & ES --+ E i- products
1 - I Porcine carboxypeptidase A Bovine carboxypeptidase A
AZ-As mixture This work’ Literature6
ka I Km --
mdl
16.6
1.75 33.0
7.15
0.15
ko I K, ko
I- -i- Al A2 Substrate
Carbobenzoxyglycyl-L-phenylalanine N-Benzoylglycyl-n-phenylalanine (hippuryl-n-phenyl-
alanine)................................,.,...,,..,. Carbobenaoxyglycyl-L-leucine. . . .
Carbobenzoxyglycyl-n-tryptophan.. . . N-Benzoylglycyl-nn-phenyllactic acid (hippuryl-m-
phenyllactic acid) . . . .
&
mlr
16.6
4.7 35.0
14.3
0.12 -
set-1 mdl SCC-I
195 33” 178”
131 27.2
73.4
110 17oc 27d 106d 93c 54c 5.ld 89d
1060 -__
QO” 138” -
-
-
ko ko
WC-
139
91.5 30
30
1000
nnf
16.6
35.0
14.3
0.12
SCC’
139
30
30
1000
mrr WC-
16.1 139
33.0 29
14.3 29.8
0.11 995
a Carboxypeptidase, five times crystallized, was purchased from the Worthington Biochemical Corporation. b See Green and Neurath (3). c In 0.04 M phosphate buffer, pH 7.5, containing 0.1 M LiCl. d In 0.007 M Veronal, pH 7.5; ionic strength to 0.5 with KCI.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

December 1963 J. E. Folk and E. W. Schirmer 3891
Isolation of Carboxypeptidase A2
Conversion of Mixture of Carboxypeptidases Al and A2 to Carboxypeptidase AZ-Samples of the mixture of carboxypep- tidases A1 and A? remaining from the preparation of carboxy- peptidase A1 were found upon esamination by disk electrophoresis to be readily and quantitatively converted to the AZ form by incubation with high concentrations of trypsin. The ammonium sulfate precipitate of the Al-A2 misture remaining from the A1 preparation was dissolved in a small volume of water and dialyzed free of salt against distilled water. The enzyme mixture was adjusted to a final concentration of 10 mg per ml at pH 8.0 in 0.005 M Tris buffer containing 0.002 M CaCI2 and was incubated for 2 hours at 25” with bovine trypsin at a concentration of 1 mg per ml.
Chromatography of Carboxypeptidase .A Z-The incubation mix- ture was pumped into a column, 3.5 x 10 cm, of DEAE-cellulose equilibrated with 0.005 M Tris buffer, pH 8.0, and protein was eluted with a lOOO-ml linear gradient ranging from 0 to 0.3 M
NaCl in 0.005 M Tris buffer, pH 8.0. The fractions containing carboxypeptidase AZ were combined, and the enzyme was salted out by addition of solid ammonium sulfate to 0.65 saturation. After dialysis against distilled water, the enzyme was rechro- matographed at pH 6.0 under the above conditions of column volume and buffer concentration. Finally, the enzyme fractions were combined and treated with ammonium sulfate (0.65 satura- tion). The protein precipitate was dissolved in a small volume of water (20 to 50 mg per ml) and dialyzed overnight against several changes of distilled water. The clear or slightly turbid enzyme solutions generally became milky but did not sediment or crystallize upon standing at 5”. The yield of twice chro- matographed enzyme from 1 g of partially purified proenzyme was 175 to 250 mg.
Properties of Purijied Carboxypeptidose AZ
Carbosypeptidase AZ prepared by the described procedure can be stored indefinitely under toluene vapors at 5”. The enzyme is readily soluble in water at pH values above 6. Freezing solu- tions of the enzyme resulted in partial loss of activity, and the lyophilized enzyme displayed only 20 to 40% of the original activity. A number of the characteristics of this form of car- boxypeptidase A were found to be identical with those reported above for the A1 form, i.e. specific activity, 37 f 1; absorbance index, 19.6; SZO,~, 3.34 S; molecular weight, 34,800 f 600, as determined by short column sedimentation equilibrium (16) based on an assumed partial specific volume of 0.73 cm3 per g. One solution of carboxypeptidase AZ, 0.43% in 0.05 M Tris- chloride buffer with 0.1 M NaCI, pH 7.0, gave a DUO,,,, of 8.63 X lo-’ cm2 set-I, as determined by the procedure of Longsworth (18). The molecular weight calculated from this D2+ and the above s~~,~, assuming a partial specific volume of 0.73 cm3 per g, is 35,250 f 600 and is in good agreement with that obtained by means of sedimentation equilibrium.
Carboxypeptidase A2 differs significantly from the Al form in its electrophoretic mobility. The higher mobility is reflected in the disk electrophoresis pattern shown in Fig. 2f (compared to carboxypeptidase Al, Fig. 2e) and in the value of -3.13 x 10m6 cm2 volt? set-l (0.05 M Tris-chloride buffer, 0.1 M NaCl, pH 8.0) calculated for the mobility of the ascending boundary in a moving boundary electrophoresis run.
Kinetic constants for carboxypeptidase AZ toward several sub- strates are recorded in Table II. No trace of carboxypeptidase
TABLE 111
Effects of heat, pH, and urea on enzymatic activities of porcine carboxypeptidase AZ and bovine
carboxypeptidase A
The porcine and bovine enzymes were exposed to the various treatments at identical concentrations (0.5 mg per ml) and were assayed under standard conditions at the level of 0.5 to 10 x lOma mg per ml.
Treatment
Original ,a+vity rema,n,ng
Porcine AZ
%
Bovine A
% Heat (0.05 M Tris buffer, 0.5 M NaCl, pH 7,
for 5 minutes) 50”....................................... 92 15 60”..................... .., 52 0
pH (0.5 M NaCl at 25” for 30 minutes) 4.0 (0.05 M acetate buffer). 7.0 (0.05 M Tris buffer). 8.0 (0.05 M Tris buffer)
11.6 (0.1 M NazC03).
65 0 100 100 100 100 87 70
Urea (8 M, 0.05 M Tris buffer, pH 8.0, 25”) 6 minutes.
11 minutes. . 20 minutes. .
100 32 100 12 97 5
-
B activity was observed in samples of purified carboxypeptidase A2.
The zinc contents of two samples of this form of the enzyme were found to be 0.90 and 1.18 moles of zinc per mole of enzyme (mol. wt., 34,800). Inhibition studies with 1, lo-phenanthroline paralleled those reported above for carboxypeptidase Al.
Preliminary comparisons of the effects of heat, pH, and urea on porcine carboxypeptidase A2 and bovine carboxypeptidase A showed that the enzymatic activity of the porcine enzyme is strikingly more stable to these influences than is that of the bovine enzyme. The results of several of these studies are re- corded in Table III.
Preparation of Mixtures of Carboxypeptidases A2 and AS
Chromatographically purified preparations of the active car- boxypeptidase A fraction from acetone powders of autolyzed porcine pancreas glands displayed specific activities similar to those of carboxypeptidases A1 and AP. Electrophoretic examina- tion of these fractions, however, revealed that they were com- posed of two proteins, one of which, although fully active en- zymatically, is distinct from carboxypeptidases A1 and A2. This third carboxypeptidase, designated Aa, has as yet not been ob- tained in a form free of carboxypeptidase AZ.
Preparation of Extracts and Ammonium Sulfate Fractionatiun- Water extracts of an acetone powder of autolyzed porcine pan- creas glands (5) were prepared by gently stirring the powder, 100 g, with distilled water, 1000 ml, at room temperature for 30 to 45 minutes. The suspension was centrifuged for 30 minutes at 14,600 x g, and the clear yellow supernatant fluid was col- lected. These extracts were cooled to O-2”, and the pH was adjusted to 7.0 to 7.2 with 1 N NaOH. All further operations were carried out at this temperature. Solid ammonium sulfate
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3892 Porcine Pancreatic Carboxypeptidase A System. I Vol. 238, No. 12
was added slowly with stirring to 0.4 saturation (243 g per liter). The pH was maintained at 7.0 to 7.2 during this operation by the addition of NaOH. The suspension was allowed to stand for 30 minutes and was centrifuged for 30 minutes at 14,600 X .g. The protein precipitate was taken up in 150 to 250 ml of water and dialyzed against distilled water overnight. The volume of the salt-free protein solution was adjusted to 400 ml with distilled water, and the pH was adjusted to 7.0 to 7.2. To this solution were added gradually with stirring 124 ml of saturated am- monium sulfate solution, saturated at 0” and adjusted to pH 7.2, to bring the salt to approximately 0.24 saturation. After 30 minutes the precipitate was removed by centrifugation, and sat- urated ammonium sulfate solution was added to the supernatant solution at a level of 16.9 ml per 100 ml to bring the salt to approximately 0.35 saturation. After 30 minutes the precipitate was collected by centrifugation, dissolved in 100 to 200 ml of 0.005 M Tris buffer, pH 8.0, and dialyzed against the same buffer overnight.
Batch Absorption and El&on from DEAE-cellulose--The di- alyzed protein solution was brought to a volume of 400 ml with 0.005 M Tris buffer, pH 8.0. Forty grams of DEAE-cellulose equilibrated with 0.005 M Tris buffer, pH 8.0, and sucked to damp dryness on a Buchner funnel were added, and the mixture was stirred for 20 minutes. The adsorbent was collected by suction filtration and washed four times by stirring with 250~ml portions of the Tris buffer and once by stirring with 100 ml of 0.08 M NaCl in Tris buffer. Finally, the enzyme was eluted from the ad- sorbent by stirring for 2 to 3 minutes with one 200-ml and two lOO-ml portions of 0.25 M NaCl in Tris buffer. The combined eluates were brought to 0.65 saturation with solid ammonium sulfate, and the precipitate was collected by centrifugation, dis- solved in 10 to 30 ml of 0.005 M Tris buffer, pH 8.0, and dialyzed against the same buffer.
Chromatography on DEAE-cellulose--The dialyzed solution was introduced into a column, 3.5 X 10 cm, of DEAE-cellulose equilibrated with 0.005 M Tris-chloride buffer, pH 8.0. The
FRACTION NUMBER
FIG. 9. Chromatography of carboxypeptidase A fraction from autolyzed porcine pancreas on DEAE-cellulose. Protein from the batch DEAE-cellulose treatment, 500 mg, was placed on a column, 3.5 X 10 cm, of DEAE-cellulose at pH 8.0 (see the text for details). Protein was eluted at 2” with a 1500-ml linear gradient from 0 to 0.3 Y NaCl in 0.995 M Tris buffer, pH 8.0. Gradient was com- menced at tube 3, and l&ml fractions were collected. A small forepeak of inactive unabsorbed material is not shown in the figure.
TABLE IV Preparation of mixtures of carboxypeptidases AZ and Aa
from acetone powder of autolyzed porcine pancreas Acetone powder, 100 g, was used.
Putification stage
mg %
Water extracts . 18,100~ 1,562 1.7 100 First (NHI)~SO, precipitation 3,900” 1,236 6.6 82 Second (NH&SO, precipitation 1,112c 760 14.6 49 DEAE-cellulose batch treatment 510d 690 27.0 44 First DEAE chromatography
(pH 8.0). 324d 525 32.4 34 Second DEAE chromatography
(pH 6.0). . . . . 274d 482 35.2 31
Total protein
Total units0
-
o One unit is defined as absorbance change per minute at the substrate concentration 1 X 10-” M (20 ml, 5-cm cell).
* Units per mg of protein per ml of assay solution. E Protein estimated from nondialyeable nitrogen value, assum-
ing 16% nitrogen. d Protein estimated with the use of an absorbance index of 19.6
for a 1% solution (278 rnp, l-cm cell).
S pecitic acti-
vity*
Total ecovery of IO- tea ytic P uztivity
FIG. 10. Disk electrophoresis patterns of porcine carboxypep- tidase Ai, AZ, and A3. Experimental conditions for electrophoresis are given in the text. The arrow and F designate the positions of the starting points and buffer fronts, respectively. a, Purified carboxypeptidase AZ, 10 gl of a solution containing 10 mg per ml. b, Mixture of carboxypeptidases Ai and AZ, 10 pl each of solutions containing 10 mg per ml. c, Mixture of carboxypeptidases & and As isolated from acetone powder of autolyzed pancreas, 10 ~1 of a solution containing 20 mg of protein per ml.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
December 1963 J. E. Folk and E. W. Schirrner
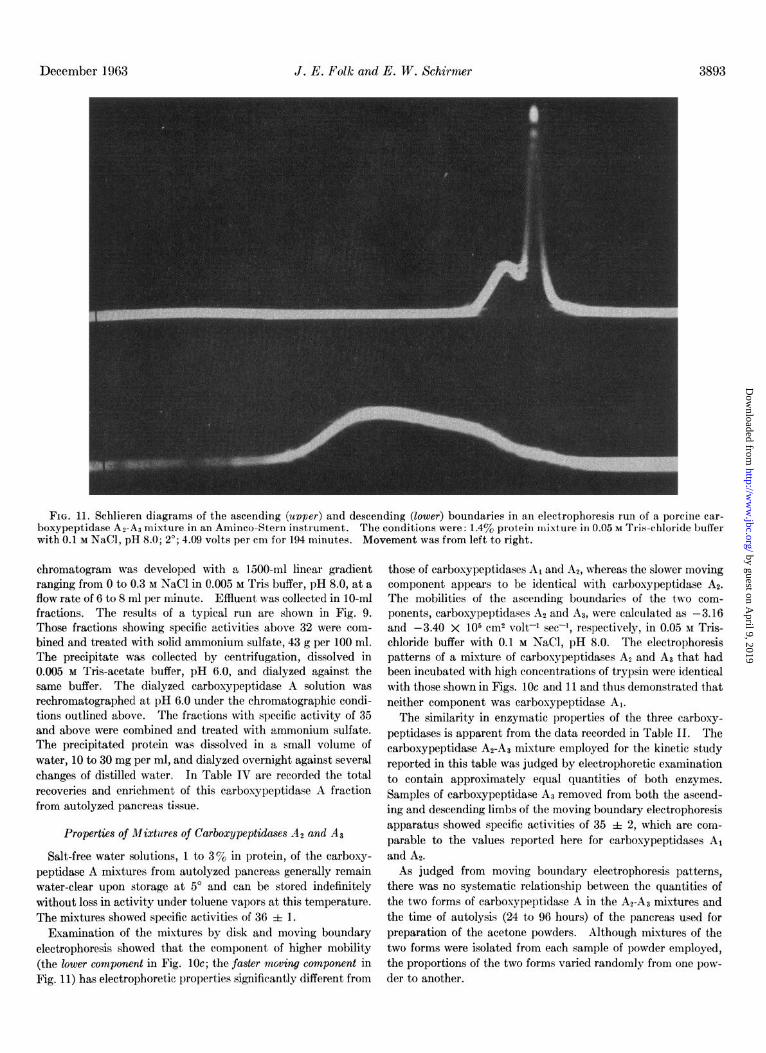
FIG. 11. Schlieren diagrams of the ascending (upper) and descending (lower) boundaries in an electrophoresis run of a porcine car- boxypeptidase AZ-AS mixture in an Aminco-Stern instrument. The conditions were: 1.4yo protein mixture in 0.05 M Tris-chloride buffer with 0.1 M NaCl, pH 8.0; 2’; 4.09 volts per cm for 194 minutes. Movement was from left to right.
chrometogram was developed with a 1500-ml linear gradient ranging from 0 to 0.3 M NaCl in 0.005 M Tris buffer, pH 8.0, at a flow rate of 6 to 8 ml per minute. Effluent was collected in lo-ml fractions. The results of a typical run are shown in Fig. 9. Those fractions showing specific activities above 32 were com- bined and treated with solid ammonium sulfate, 43 g per 100 ml. The precipitate was collected by centrifugation, dissolved in 0.005 M Tris-acetate buffer, pH 6.0, and dialyzed against the same buffer. The dialyzed carboxypeptidase A solution was rechromatographed at pH 6.0 under the chromatographic condi- tions outlined above. The fractions with specific activity of 35 and above were combined and treated with ammonium sulfate. The precipitated protein was dissolved in a small volume of water, 10 to 30 mg per ml, and dialyzed overnight against several changes of distilled water. In Table IV are recorded the total recoveries and enrichment of this carboxypeptidase A fraction from autolyzed pancreas tissue.
Properties of Mixtures of Carboxypeptidases A:! and Aa
Salt-free water solutions, 1 to 3% in protein, of the carboxy- peptidase A mixtures from autolyzed pancreas generally remain water-clear upon storage at 5” and can be stored indefinitely without loss in activity under toluene vapors at this temperature. The mixtures showed specific activities of 36 f 1.
Examination of the mixtures by disk and moving boundary electrophoresis showed that the component of higher mobility (the lower component in Fig. 10~; the faster mwing component in Fig. 11) has electrophoretic properties significantly different from
those of carboxypeptidases A1 and AS, whereas the slower moving component appears t’o be identical with carboxypept.idase AZ. The mobilities of the ascending boundaries of the two com- ponents, carboxypeptidases At and Al, were calculated as -3.16 and -3.40 X lo5 cm2 volt-l set-I, respectively, in 0.05 M Tris- chloride buffer with 0.1 M NaCl, pH 8.0. The electrophoresis patterns of a mixture of carbosypeptidases A2 and A) that had been incubated with high concentrations of trypsin were identical with those shown in Figs. 1Oc and 11 and thus demonstrated t,hat neither component was carboxypeptidase Al.
The similarity in enzymatic properties of the three carboxy- peptidases is apparent from the data recorded in Table II. The carboxypeptidase AZ-AS mixture employed for the kinetic study reported in this table was judged by electrophoretic examination to contain approximately equal quantities of both enzymes. Samples of carboxypeptidase AS removed from both the ascend- ing and descending limbs of the moving boundary electrophoresis apparatus showed specific activities of 35 f 2, which are com- parable to the values reported here for carbosypeptidases A1 and At.
As judged from moving boundary electrophoresis patterns, there was no systematic relationship between the quantities of the two forms of carboxypeptidase A in the At-A3 mixtures and the time of autolysis (24 to 96 hours) of the pancreas used for preparation of the acetone powders. Although mixtures of the two forms were isolated from each sample of powder employed, the proportions of the two forms varied randomly from one pow- der to another.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3894 Porcine Pancreatic Carboxypeptidase A System. I Vol. 238, Ko. 12
DISCUSSION
There have been numerous reports on the chemical and physi- cal characteristics of bovine pancreatic carboxypeptidase A (3,4). There have been no formal reports, however, that this enzyme exists in active forms other than that of the crystalline protein first described by Anson (2). The data presented in this com- munication show that active carboxypeptidase A of porcine origin may exist in at least three physically distinguishable forms and that these three carbosypeptidases have equivalent en- zymatic properties. These observations were made possible by the fact that the carboxypeptidases of porcine pancreas are much more water-soluble than the bovine euglobulin carboxypeptidase A and hence lend themselves more readily to physical separation and examination.
There are some indications that bcvine carbosypeptidase A, like the pcrcine enzymes, may exist in more than one active molecular form. Thus, Keller, Cohen, and Neurath (6) reported a slight but significant difference in the electrophoretic mobility of crystalline carboxypeptidase A prepared by tryptic activation of procarboxypeptidase A and that of the crystalline enzyme prepared according to the procedure of Anson. They point out that “the significance of the slight difference in electrophoretic mobilities is not yet known.” Poljak, moreover, has observed two chromatographically separable active components in samples of commercial crystalline bcvine carbosypeptidase A.3 Varied quantities of amino-terminal asparagine and serine were re- ported in samples of crystalline bovine carboxypeptidase A by Thompson (19). These NHz-terminal amino acids were identi- fied as their dinitrophenyl derivatives in acid hydrolysates of the dinitrophenylated protein samples. Although Thompson at- tributed his finding to peptide bond cleavage occurring during the initial step of the end group determinations and assumed that the enzyme had a single NHz-terminal residue, asparagine, and the NHz-terminal sequence asparaginylserine, it is quite possible that these two end groups represented two forms of active en- zyme in the crystalline preparations employed for analyses. This possibility gains added emphasis from the’recent report of Coombs and Omote (20) that the NH&erminal sequence of bovine carboxypeptidase A is asparaginyltyrosylalanine, not as- paraginylserine, and from the observation reported in the accom- panying communication (10) that porcine carboxypeptidases A1 and AP have different NH*-terminal amino acid residues.
It is evident from the present studies that porcine carboxypep- tidase A2 is the product of tryptic action on carboxypeptidase A,. Whether carboxypeptidase AS arises directly from the proenzyme through tryptic digestion or by conversion from the Al form only is not known at present. It is also not clear from the data pre- sented here whether carbosypeptidase A1 is formed from the zymogen by the action of trypsin or through the catalytic in- fluence of the chymotrypsin-like material formed during activa- tion of partially purified procarbosypeptidase A preparations. Carbosypeptidase AS has been found only in autolysates of por- cine pancreas. The mechanism of its formation and the enzyme or enzymes involved are unknown. Experiments designed to clarify these questions are in progress.
SUMMARY
1. Three electrophoretically distinct active forms of porcine pancreatic carboxypeptidase A have been recognized. These three enzymes, which have essentially identical enzymatic prop- erties, have been designated carbosypegtidases A,. A.. and Al.
3 R. J. Poljak, personal communication.
2. Procedures have been developed for the isolation of car- boxypeptidases Al and AP using, as a starting material, partially purified procarboxypeptidase A. These procedures involve ac- tivation of the zymogen with trypsin and chromatographic sep- aration of the active carboxypeptidases on diethylaminoethyl cellulose. Carboxypeptidase At has been obtained in crystalline form.
3. Carboxypeptidases A1 and AP behave as homogeneous pro- teins with respect to several physical chemical criteria. The molecular weight of each has been calculated as 34,800 f 600. Each enzyme contains 1 atom of zinc per molecule.
4. Carboxypeptidase Al is converted, through the action of trypsin, to the Az enzyme.
5. Carboxypeptidase Aa has been obtained as a mixture of the AZ and Aa forms from extracts of acetone powders of autolyzed porcine pancreas glands and has not been fully characterized.
6. Several kinetic constants and some of the physical proper- ties of the porcine enzymes have been compared with those of crystalline bcvine carboxypeptidase A.
Acknozuledgments-Our thanks are due to Dr. Marc S. Lewis for molecular weight determinations by the short column sedi- mentation equilibrium method, to Mr. E. R. Mitchell for per- forming the moving boundary electrophoretic and ultracentrifu- gal analyses, to Mr. H. L. Wolff for the diffusion analysis, to Mr. R. C. Braunberg for espert technical assistance, and to Dr. William R. Carroll for his cooperation during the initial phase of this work.
REFERENCES
1. FOLK, J. E., AND SCHIRMER, E. W., Federation Proc., 22, 594 (1963).
2. ANSON, M. L., Science, 81, 467 (1935). 3. GREEN, N. M., AND NEURATH, H., in H. NEURATH AND K.
BAILEY (Editors), The proteins, Vol. ZZ, Academic Press, Inc., New York, 1954, Part B, p. 1057.
4. NEURATH, H., in P. D. BOYER, H. LARDY, AND K. MYRBXCK (Editors), The enzymes, Vo’ol. 4, Academic Press, Inc., New York, 1960, p. 11.
5. ANSON, M. L., J. Gen. Physiol., 20, 777 (1937). 6. KELLER, P. J., COHEN, E., AND NEURATH, H., J. Biol. Chem.,
223, 457 (1956). 7. KELLER, P. J., COHEN, E., AND NEURATH, H., J. Biol. Chem.,
230, 905 (1958). 8. BROIVN, J. R., Cox, D. J., GREENSHIELDS, R. N., WALSH, K.
A., YAMASAKI, M., AND NEURATH, H., Proc. Natl. Acad. Sci. I/‘. S., 47, 1554 (1961).
9. MARCHIS-MOUREN, G., CHARLES, M., BEN ABDELJLIL, A., AND DESNUELLE, P., Biochim. et Biophys. Actu, 60, 186 (1961).
10. FOLK, J. E., J. Biol. Chem., 238, 3895 (1963). 11. WOLFF, E. C., SCHIRMER, E. W., AND FOI,K, J. E., J. Biol.
Chem., 23’7, 3094 (19G2). 12. FOLK, J. E., PIEZ, K. A., CARROLL, W. R., AND GLADNER, J.
A., J. Biol. Chem., 236,2272 (19GO). 13. FOLK, J. E., AND GLADNER, J. A., J. Biol. Chem., 236,60 (1960). 14. MOORE, S., AND STEIN, W. H., J. Biol. Chem., 176, 367 (1948). 15. ROVERY, M., CHARLES, M., GTJY, O., GUIDONE, A., AND DES-
NUELLE, P., Bull. sot. chim. biol., 42,1235 (1960). 16. RICHARDS, E. G., AND SCHACHMAN, H. K., J. Phys. Chem., 63,
1578 (1959). 17. MALMSTROM, B. G., in D. GLICK (Editor), Methods of biochemi-
cal analysis, Vol. ZZZ, Interscience Publishers, Inc., New York, 1956, p. 327.
18. LONGSWORTH, L. G., J. Am. Chem. Sot., 74, 4155 (1952). 19. THOMPSON, E. 0. P., Biochim. et Biophys. Acta, 10, 633 (1953). 20. COOMBS, T. L., AND OMOTE, Y., Federation Proc., 21, 234
(1962).
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

J. E. Folk and E. W. SchirmerTHE ACTIVE ENZYME
The Porcine Pancreatic Carboxypeptidase A System: I. THREE FORMS OF
1963, 238:3884-3894.J. Biol. Chem.
http://www.jbc.org/content/238/12/3884.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/238/12/3884.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























