Embed Size (px)
Citation preview

STUDIES ON THE ACTION OF CARBOXYPEPTIDASE ON TOBACCO MOSAIC VIRUS*
13~ J. IE;lJAN HARlllSt ASI) (:. A. 1iNIGIlT
(Prom the Department of Biochemistry and the Virus Laboratory, University of California, Berkeley, California)
(Received for publication, August 30, 1954)
The action of carboxypeptidase on tobacco mosaic virus (TMV) results in the exclusive release of threonine from the virus nucleoprotein (1). The enzyme-treated virus retains full virus infectivity, and its disease symptoms on Turkish tobacco leaves are indistinguishable from those caused by the untreated virus. The treated virus, however, does not reproduce in its modified form, since treatment of the progeny with carboxypeptidase results in the release of the same amount of threonine as one obtains from progeny of untreated virus.
In the present communication additional observations on the nature of the enzyme-virus reaction and on the origin of the liberated threonine are presented, together with additional data on the chemical, physical, and serological properties of the carboxypeptidase-treated virus. The reaction of the enzyme with virus which had been subjected to sonic treatment, and also with detergent-dissociated virus, has also been studied. On the basis of these observations evidence is presented for the occurrence in the molecular structure of TMV of a series of repeating fundamental units.
EXPERIMENTAL
Virus and Enzyme Preparations-Highly purified preparations of TMV were obtained by the standard procedure of differential centrifugation and were stored in aqueous solution in the presence of small amounts of added toluene to prevent bacterial or mold contamination. Supernatant solu- tions resulting from the sedimentation of the virus from such preparations were found to be free from ninhydrin-reacting substances (see Spot A, Fig. 5).
Carboxypeptidase-treated virus (CT) was obtained by treating TMV with enzyme until the liberation of its available threonine was complete,
* This investigation was supported in part by a research grant, C-2162 M and I, from the National Cancer Institute, National Institutes of Health, United States Public Health Service, and by grants from the Lederle Laboratories Division, Amer- ican Cyanamid Company, and the Rockefeller Foundation.
t Present address, Carlsberg Laboratorium, Copenhagen, Denmark. This work was carried out while J. I. Harris was a member of the staff of the Hormone Research Laboratory and was supported in part by a grant from the Rockefeller Foundation.
215
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21G ACTION OF CARBOXTPEPTIDASE ON TMV
as described below in the section dealing with rate of release of threonine from TMV. The treated virus was isolated from the reaction mixture in the form of a pellet by means of high speed centrifugation, and subse- quently redissolved in distilled water containing a drop of toluene as pre- servative; such a preparation (CT) was used to inject rabbits in order to obtain CT antiserum. In preparing CT for the electrophoretic experi- ments, the virus was subjected to additional cycles of centrifugation in order to wash out the trace of enzyme occluded in the virus gel. While negligible on a weight basis, this trace of enzyme was sufficient to react with TMV when CT and TMV were mixed and dialyzed for several hours in preparation for certain electrophoresis experiments.
Sonically treated virus, TMV(S), was prepared by subjecting a solution of TMV, containing 20 mg. of virus per ml., to vibrations of 9 kc. for 30 minutes in a magnetostriction oscillator (model S-102A, Raytheon Manu- facturing Company, Waltham, Massachusetts). When the virus solution was subsequently centrifuged at 40,000 r.p.m. for 15 minutes in the Spinco model L ultracentrifuge, a small gelatinous pellet was obtained, and the bulk of the material, which remained in the supernatant fluid, was carefully separated from the residual pellet. TMV(S), like the preparation of TMV from which it was derived, was composed of rod-like particles of varying length as shown in Fig. 1. The size of the rods in these treated prepara- tions averaged, however, about one-third that of the intact virus rods. The particles resulting from sonic treatment of TMV show no evidence of chemical alteration and contain the same proportions of protein and nu- cleic acid as the intact virus (2).
TMV protein, TMV(P), obtained by treating the virus with sodium dodecyl sulfate at pH 8 to 9 and 37” for 24 hours (3), was generously placed at our disposal by Dr. H. Fraenkel-Conrat; the absence of N-terminal groups in the same preparations has already been reported (4). The prep- aration was completely free from virus nucleic acid and was found accord- ing to ultracentrifugal analysis to be composed mainly of material whose molecular weight is 10,000 to 20,000.
The carboxypeptidase preparations used for the enzyme experiments were obtained as crystalline aqueous suspensions from the Worthington Biochemical Sales Company and from the Armour Research Laboratories.* Just before use a portion of enzyme was brought into solution by the addi- tion with stirring at 04” of 1 per cent NaHC03, together with a few drops of 0.1 N NaOH; an alternate procedure, which seemed to work well, was simply to add 0.1 N NaOH and to mix at room temperature. The solution was further diluted with distilled water to a concentration of about 0.004 to 0.005 mg. of enzyme N per ml. Crystalline carboxypeptidase may con-
1 The authors are indebted to Dr. Lawrence L. Lachat of the Biochemical Markrt Development Section for a generous supply of enzyme.
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
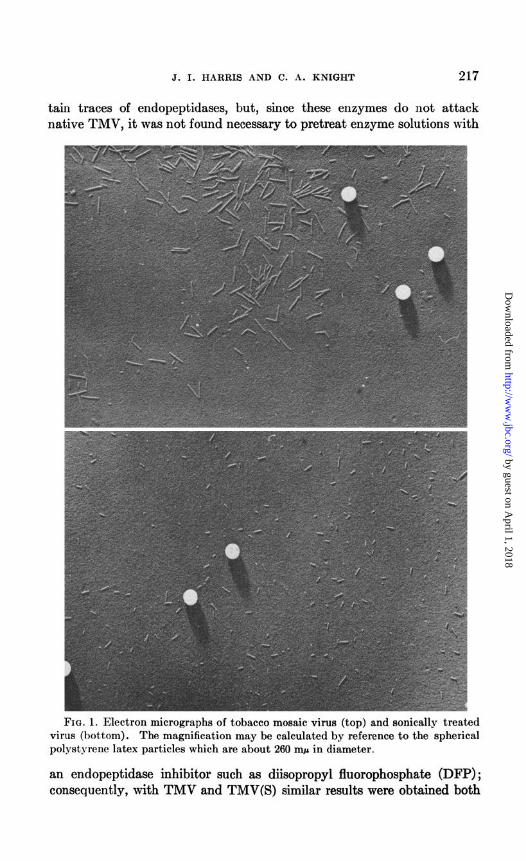
J. I. HARRIS AND C. A. KNIGHT 217
tain traces of endopeptidases, but, since these enzymes do not attack native TMV, it was not found necessary to pretreat enzyme solutions with
FIG. 1. Electron micrographs of tobacco mosaic virus (top) and sonically treated virus (bottom). The magnification may be calculated by reference to the spherical polystyrene latex particles which are about 260 nq.~ in diameter.
an endopeptidase inhibitor such as diiiopropyl fluorophosphate (DFP); consequently, with TMV and TMV(S) similar results were obtained both
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
218 ACTION OF CARBOXYPEPTIDASE ON TMV
with and without DFP. In the case of TMV(P), however, new end- groups in addition to threonine were found to be released in experiments in which the enzyme had not been treated with a 50-fold excess of DFP prior to its addition to the substrate.
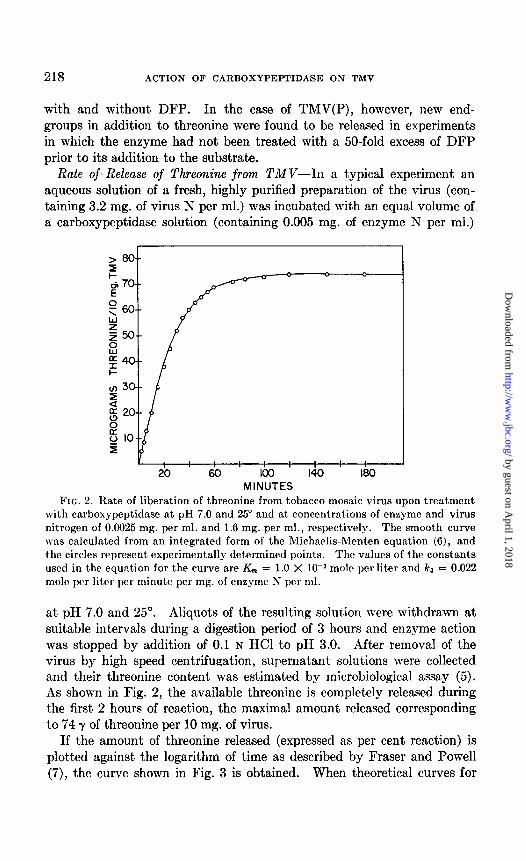
Rate of-Release of Threonine from TMV-In a typical experiment an aqueous solution of a fresh, highly purified preparation of the virus (con- taining 3.2 mg. of virus N per ml.) was incubated with an equal volume of a carboxypeptidase solution (containing 0.005 mg. of enzyme N per ml.)
20 80 loo 140 180 MINUTES
FIG. 2. Rate of liberation of threonine from tobacco mosaic virus upon treatment with carboxypeptidase at pH 7.0 and 25” and at concentrations of enzyme and virus nitrogen of 0.0025 mg. per ml. and 1.6 mg. per ml., respectively. The smooth curve was calculated from an integrated form of the Michaelis-Menten equation (6), and the circles represent experimentally determined points. The values of the constants used in the equation for the curve are K,,, = 1.0 X low3 molt per liter and ka = 0.022 mole per liter per minute per mg. of enzyme N per ml.
at pH 7.0 and 25”. Aliquots of the resulting solution were withdrawn at suitable intervals during a digestion period of 3 hours and enzyme action was stopped by addition of 0.1 N HCl to pH 3.0. After removal of the virus by high speed centrifupation, supernatant solutions were collected and their threonine content was estimated by microbiological assay (5). As shown in Fig. 2, the available threonine is completely released during the first 2 hours of reaction, the maximal amount released corresponding to 74 y of threonine per 10 mg. of virus.
If the amount of threonine released (expressed as per cent reaction) is plotted against the logarithm of time as described by Fraser and Powell (7), the curve shown in Fig. 3 is obtained. When theoretical curves for
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS AND C. A. IiNIGHT 219
zero and first order reactions are constructed on the same scale, the rate of release of threonine from TMV appears to follow zero order kinetics in the initial stages and later to become first order in the region of low substrate concentrat,ion. Release of threonine from the virus is t,hus shown to follow the course \\hich might be prcdictcd from the classical Michaelis-h!Ienten theory for an enzyme-catalyzed reaction at constant enzyme concentration (8).
A direct demonstration of conformity of the TMV-carboxypeptidase re- action to the Michaelis-Menten theory is afforded by application of an integrated form of the Michaelis-Menten equation (6) as follows:
kJet = 2.3K,,, log z + (a0 - a)
As pointed out by Elkins-Kaufman and Neurath (6), Equation 1 is a com- posite first order and zero order expression. When data from the TMV- carboxypeptidase reaction are substituted in this equation, which is then solved for the constants, and the equation is plotted, the smooth curve shown in Fig. 2 is obtained. Application to this plot of experimentally determined values for t.he threonine released from TMV after various rc- action times results in the fit shown in Fig. 2. The excellence of fit thus shown can be considered strong evidence for adherence of the kinetics to the Michaelis-Menten theory. The value for K,, 1.0 X 10e3 mole per liter, is approximately 10 times that found for synthetic model substrates (9).
On the basis of the above considerations, an interpretation of the virus- enzyme reaction can be made as follows. In the early stages of the reac- tion, the effective concentration of the substrate is high; thus, on the assumption that each mole of virus nucleoprotein contains approximately 2900 moles of a reactive subunit, a substrate-enzyme mole ratio of ap- proximately 1000: 1 will apply. Consequently, in the initial stages of the reaction the rate of formation of the virus-enzyme complex is likely to be considerably faster than its rate of dissociation and the rate of release of threonine; in this event the second right-hand term in Equation 1 would predominate; hence the rate of release of threonine from the virus would be expected to approach zero order kinetics.
If, however, as the concentration of TMV diminishes, the rate of forma- tion of the virus-enzyme complex becomes considerably slower than the rate of its decomposition, enzyme-substrate complex formation becomes the rate-determining step; the first right-hand term of Equation 1 will now predominate and as a result release of threonine from the virus should become a first order reaction at low concentrations of TMV.
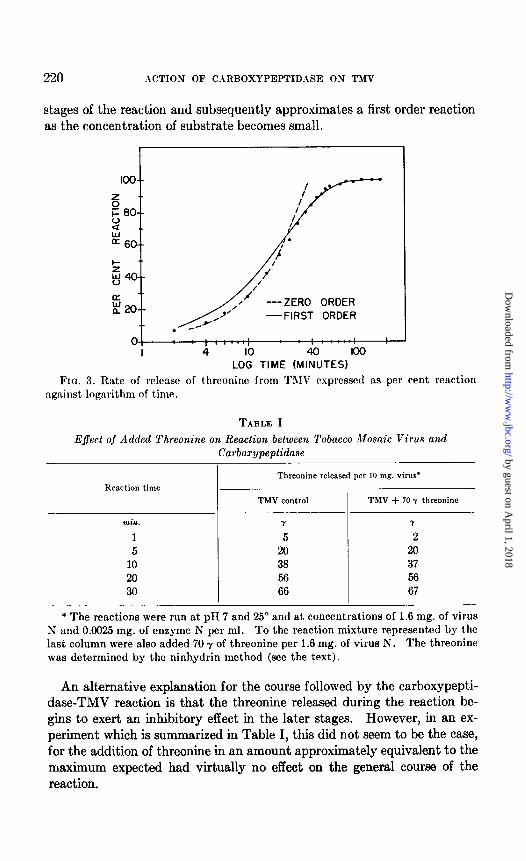
As shown in Fig. 3, these expectations are fulfilled. The rate of release of threonine from the virus approximates zero order kinetics in the initial
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
220 ACTION OF CIRBOXYPEPTID.4SE ON TMV
stages of the reaction and subsequently approximates a first order reaction as the concentration of substrate becomes small.
0-l I ' ' "'1 I 4 IO 40 00
LOG TIME (MINUTES)
Fro. 3. Rate of release of threoninc from TMV expressed as per cent, reaction against logarithm of time.
TABLE I
Effect oj Added Threonine on Reaction between Tobacco Mosaic Virus and Carboxypeptidase
Reaction time
Threonine released per 10 mg. virus’
TMV control TMV + 70 y threonine
min. Y Y
1 5 2 5 20 20
10 38 37 20 56 56 30 66 67
* The reactions were run at pH 7 and 25” and at concentrations of 1.6 mg. of virus N and 0.0025 mg. of enzyme N per ml. To the reaction mixture represented by the last column were also added 70 y of threonine per 1.6 mg. of virus N. The threonine was determined by the ninhydrin method (see the text).
An alternative explanation for the course followed by the carboxypepti- dase-TMV reaction is that the threonine released during the reaction be- gins to exert an inhibitory effect in the later stages. However, in an ex- periment which is summarized in Table I, this did not seem to be the case, for the addition of threonine in an amount approximately equivalent to the maximum expected had virtually no effect on the general course of the reaction.
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS AND C. A. KNIGHT 221
Quantitative Estimation of Threonine-As indicated above, many of the threonine determinations were made by means of microbiological assay (5). Some determinations were also made by the quantitative ninhydrin method of Moore and Stein (10) with threonine as a standard. Finally, in order to obtain an accurate and independent check of the amount of threonine released in the TMV-carboxypeptidase reaction, the dinitrophenyl (DNP) procedure (11) was applied to a supernatant solution obtained as described above.
A 1 ml. aliquot of the supernatant solution from a 3 hour digest of TMV was allowed to react with dinitrofluorobenzene (DNFB) at pH 9.0 and 40’ for 90 minutes; excess reagent was removed by extracting the reaction solution with ether. The aqueous solution was then acidified to pH 1.0 with HCl and again extract,ed with ether to remove the DNP derivative (or derivatives) of the products of the carboxypeptidase reaction. The combined ether extracts were concentrated in vacua, and the dinitrophenol formed by hydrolysis of DNFB was removed by sublimat.ion; the residue was then examined by two-dimensional chromatography on paper (11). As expected, only one spot, corresponding to DNP threonine, was obtained. The DNP amino acid was eluted with 1 per cent NaHC03 (4 ml.) and the optical density of the solution at 360 rnp was measured in a Beckman spectrophotometer.
In a separate experiment the above procedure was applied to a known amount (1 pmole) of pure m-threonine, and the optical density readings of such standard solutions were used to calculate the amount of threonine (in micromoles) present in supernatant solut.ions obtained from carboxy- peptidase-TMV reaction mixtures. This’ procedure automatically com- pensates for experimental losses, and furthermore an accurate value for the molecular extinction coefficient of DNP threonine need not be known.
In this way it was estimated that 69.2 (f2) y of threonine are released from 10 mg. of TMV by the action of carboxypeptidase; this value is in good agreement with that of 70 f 4 y obtained by microbiological assay on the same solution and is equivalent to 1 threonine residue per unit of molecular weight of 17,200 on the basis of a molecular weight of about 50 x 10” for TMV (12).
Action of Carboxypeptidase on TMV(S) and TMV(P)-In order to ob- tain further evidence concerning the possible existence of a chemical sub- unit in TMV a study was made of the action of carboxypeptidase on disso- ciated fragments obtained from TMV by sonic treatment and by the action of sodium dodecyl sulfate as described earlier; these procedures as applied here are presumed not to break covalent bonds.
Solutions containing TMV, TMV(S), and TMV(P) (containing the equivalent of 20 mg. of virus per ml.) were each mixed with equal volumes of a carboxypeptidase solution (containing 0.064 mg. of enzyme per ml.,
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

222 ACTION OF CARBOXYPEPTIDASE ON TMV
which had been pretreated with a 50-fold excess of DFP at room tempera- ture). The TMV and TMV(S) reaction solutions were maintained at pH 7.0, but the TMV(P) solution was allowed to react at pH 8.0 in order to keep the substrate in solution during the course of the enzyme reaction. Aliquots of the respective reaction solutions were withdrawn at suitable intervals during a 4 hour digestion period and enzyme action in each ali- quot was terminated by the addition of 0.1 N HCl to pH 3.0. TMV and TMV(S) were removed by high speed centrifugation and the threonine content of the supernatant solutions was estimated by microbiological assay. Although TMV(P) did not form a pellet in the ultracentrifuge,
-K. TMV=3.Ox lO-3 MG/MIN
-K, TMV(S)= I.6 x lO-3 ”
D-K, TMV(P)=O.9 x 1O-3 ”
0 15 30 60 90 120 180 240 M I NUTES
FIG. 4. Rates of liberation of threonine from TMV, TMV(S), and TMV(P). The reactions were run at 22” and at pH 7 except for TMV(P), which was held at pH 8. The reaction mixtures contained 10 mg. of substrate and 0.032 mg. of enzyme per ml. and the enzyme was pretreated with a N-fold excess of diisopropyl fluorophosphate.
assay for threonine was successfully performed in the presence of the pro- tein substrate. The results showing the rates of release of threonine from the three substrates are presented in Fig. 4.
Release of threonine from TMV was complete after 1 hour and, as ex- pected, amounted to 67 (+3) y from 10 mg. of virus. From the sonically treated virus, threonine was released at a slower rate, but after about 2 hours reached a maximal value of 66 (+3) y from 10 mg. of TMV(S). In the case of TMV(P) the reaction rate was still slower despite the fact that the reaction was carried out at a pH which is considered nearer the optimum for the action of the enzyme. Thus, a maximum corresponding to 67 f 3 y of threonine from 10 mg. of substrate was reached with TMV(P) only after 4 hours of digestion.
For paper chromatographic studies aliquots of the supernatant solutions
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS AND C. A. KNIGHT 223
taken after 3 hours digestion, and corresponding to about 5 mg. of virus, were freed from residual protein by the “molecular sieve” procedure (13, 14). Thus after high speed centrifugation the supernatant solutions were shaken mechanically for 1 hour with Amberlite IR-120 resin in the H+ form; the protein solutions were removed by means of a pipette, and the
FIG. 5. Chromatogram obtained with supernatant solutions from carboxypepti- d--treated virus preparations with butanol-acetic acid-water, 4:1:5, as the de- veloping solvent and Whatman No. 1 paper. Spot A, TMV control, without, enzyme; Spot B, TMV-carboxypeptidase; Spot C, threonine; Spot D, TMY(P)-carboxypepti- dase; Spot E, TMV(S)-carboxypeptidase.
residual resin was washed several times with distilled water to remove last traces of protein. The resin was then eluted with 5 N NHbOH and aliquots of the ammonia eluates were chromatographed on Whatman No. 1 paper with butanol-acetic acid-water (4:1:5) as the developing solvent. As shown in Fig. 5, threonine exclusively is released from all three substrates, indicating that no new C-terminal groups are revealed in the virus molecule after sonic or detergent treatment.
Efect of Elevated Temperature 012 Carboxypeptidase-TMV Reaction-h
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

224 ACTION OF CARBOXYPEPTIDASE ON TMV
order to test the possibility that more threonine or that other amino acids might be released from TMV by carboxypeptidase at an elevated tempera- ture, duplicate reactions were run at 23” and 37”. The relative concen- trations of enzyme and virus and other conditions were those previously described.
Paper chromatography of the supernatant fluids from the reaction mix- tures showed threonine to be the only amino acid present in significant amount in either case. Quantitative analysis of the threonine, made by the ninhydrin method of Moore and Stein (lo), gave identical values for the supernatant fluids at 23” and 37”, namely, the equivalent of 66 y of threonine per 10 mg. of virus.
In other experiments made in this laboratory: elevated temperature, 10 times the concentration of enzyme, and several times the reaction period were employed, still without appreciably changing the outcome.
These results further strengthen the conclusion that threonine is the only product of the reaction of carboxypeptidase on TMV and that the maximal amount of threonine is released under the mild conditions used in most of our experiments.
Influence of Host on Nature of C-Terminal Groups of TMV-In order to investigate the possibility that the threonine released from TMV by the action of carboxypeptidase might represent an unusual type of impurity associated with the culture of the virus in Turkish tobacco, highly purified preparations of TMV were obtained from Turkish and white Burley tobacco and from phlox plants. Each preparation was treated with car- boxypeptidase at pH 7 to 7.5 for 2 hours at room temperature. After removal of the virus by high speed centrifugation, the supernatant fluids were assayed for threonine in the usual manner. By using two separate preparations of each, values of 65 and 61, 61 and 58, and 61 and 63 y of threonine3 per 10 mg. of virus were obtained for TMV from Turkish to- bacco, white Burley tobacco, and phlox, respectively. From these results it can be concluded that the C-terminal threonine associated with TMV is characteristic of the virus and not of the host from which the TMV is obtained.
Efect of Endopeptidases on Carboxypeptidase-Treated TMV-Although it is well known that trypsin and chymotrypsin do not attack native TMV, it seemed of interest to investigate whether the virus becomes more sus- ceptible to proteolytic degradation after its initial reaction with carboxy- peptidase.
Virus which had undergone complete reaction with carboxypeptidase
2 Dr. H. Fraenkel-Conrat, personal communication. 3 These results are slightly low, presumably owing to decreased activity of the
enzyme preparation used, since other experimental evidence also indicated this to be the case.
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS AND C. A. KNIGHT 225
was adjusted to a pH of 7.5 and treated with crystalline trypsin and chymotrypsin, respectively. Each reaction mixture contained 10 mg. of CT and 0.01 mg. of enzyme, and the reaction was allowed to proceed at room temperature (22”) for 6 hours. CT was removed from the reaction solutions by centrifugation at 40,000 r.p.m. for 1 hour and the supernatant fluids were withdrawn, concentrated, and chromatographed on paper with butanol-acetic acid-Hz0 (4: 1: 5) as the developing solvent. No ninhydrin- reacting material was detected; hence it is concluded that CT, like native TMV, is not susceptible to hydrolysis by trypsin and chymotrypsin.
Physicochemical Properties of Carboxypeptidase-Treated TMV-Prepara- tions of TMV and CT were kindly examined in the electron microscope by Dr. R. C. Williams using the spray droplet technique in preparation of the mounts (15). The electron micrographs obtained showed nothing distinctive in the morphology of the CT particles, since the bulk of the material was represented by rod-like particles about 300 rnp in length with shorter and longer rods each present in approximately the same numbers commonly observed in purified preparations of TMV.
The sedimentation behavior and electrochemical properties of TMV and CT in 0.1 M cacodylate buffer at pH 6.8 were kindly investigated by Dr. H. K. Schachman, using the Spinco model E centrifuge and the Perkin- Elmer and Spinco electrophoresis apparatus.
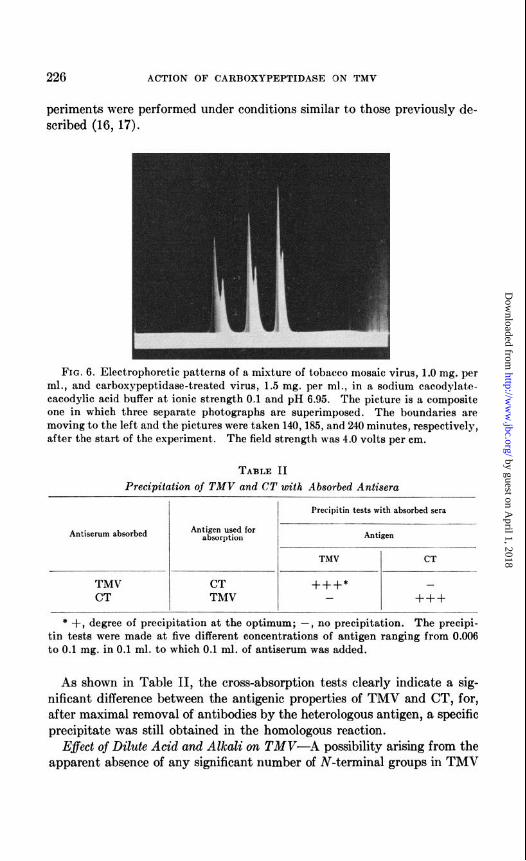
Both the ultracentrifuge patterns and the sedimentation coefficients were in agreement with the results of electron microscopy in indicating that treatment of TMV with carboxypeptidase effects no substantial change in the size, shape, or density of the virus particles. The individual prepara- tions of TMV and CT exhibited single boundaries in the electrophoresis apparatus in a sodium cacodylate-cacodylic acid buffer at an ionic strength of 0.1 and pH 6.9. Although the electrophoretic mobilities of TMV and CT were similar, -7.4 X 1O-5 and -7.9 X 10-5 cm. per second per volt per cm. for TMV and CT, respectively, resolution was obtained, as illus- trated in Fig. 6, when the two were studied in a mixture. These results show that treatment of TMV with carboxypeptidase leads to a material with greater electrophoretic mobility at pH 6.9.
Serological Properties of Carboxypeptidase-Treated TMV-Antisera to TMV and CT were obtained by subjecting rabbits to a course of eight spaced intravenous injections. A total of 58 mg. of each substance was injected into the appropriate rabbit. About 5 days after the last injection, the rabbits were bled by cardiac puncture and the sera were separated from the clotted blood. Four rabbits were employed, two for each antigen.
Strong reactions and cross-reactions were observed in precipitin tests with the various sera and antigens, thus demonstrating strong serological relationships. In order to detect possible qualitative differences in the antigens, TMV and CT, as reflected in their antisera, cross-absorption ex-
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from
226 ACTION OF CARBOXYPEPTIDASE ON TMV
periments were performed under conditions similar to those previously de- scribed (16, 17).
FIG. 6. Electrophoretic patterns of a mixture of tobacco mosaic virus, 1.0 mg. per ml., and carboxypeptidaee-treated virus, 1.5 mg. per ml., in a sodium cacodylate- cacodylic acid buffer at ionic strength 0.1 and pH 6.95. The picture is a composite one in which three separate photographs are superimposed. The boundaries are moving to the left and the pictures were taken 140,185, and 240 minutes, respectively, after the start of the experiment. The field strength was 4.0 volts per cm.
TABLE II Precipitation of TMV and CT with Absorbed Antisera
I I Precipitin tests with absorbed sera
Antiserum absorbed Antigen used for absorption Antigen
TMV CT
TMV CT +++* - CT TMV - +++
* +, degree of precipitation at the optimum; -, no precipitation. The precipi- tin tests were made at five different concentrations of antigen ranging from 0.006 to 0.1 mg. in 0.1 ml. to which 0.1 ml. of antiserum was added.
As shown in Table II, the cross-absorption tests clearly indicate a sig- nificant difference between the antigenic properties of TMV and CT, for, after maximal removal of antibodies by the heterologous antigen, a specific precipitate was still obtained in the homologous reaction.
Eflect of Dilute Acid and Alkali on TMV-A possibility arising from the apparent absence of any significant number of iv-terminal groups in TMV
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS AND C. A. KNIGHT 227
(4) is that TMV is made up of cyclic peptide chains and that the threonine residues removed by carboxypeptidase do not really represent C-terminal groups, but rather are side chain residues which are linked in the virus either singly or as polythreonine chains to y-carboxyl groups of glutamic acid residues. To test these possibilities, the stability of TMV at pH 2.8 and 11 was investigated.4
An aqueous solution of TMV (2 ml., containing 10 mg. of virus per ml.) was brought to pH 2.8 with 0.1 N HCl and kept at 100” for 1 hour. The coagulated virus was centrifuged at 20,000 r.p.m. for 1 hour, and the super- natant solution was separated from the pellet of denatured virus.
A similar solution of TMV was brought to pH 11 with 0.1 N NaOH and heated at 100” for 1 hour. The solution remained clear and no pellet was recovered after centrifugation at 20,000 r.p.m. for 1 hour; in order to sepa- rate degradation products of low molecular weight the solution was sub- jected to the “molecular sieve” resin procedure already described.
The supernatant solution at pH 2.8 and the material adsorbed by the resin from the solution at pH 11 were chromatographed on Whatman No. 1 paper in butanol-acetic acid-water (4: 1: 5). By this means, aspartic acid was found to be the only amino acid released from the virus at pH 2.8 and 1OOo,6 while at pH 11 small amounts of free serine, glycine, and threo- nine could be detected. Their combined amount, however, represents less than 5 per cent of the threonine which would be released from an equiv- alent amount of virus under the action of carboxypeptidase. In view of these results, the presence of polythreonine units or of r-glutamyl peptide linkages in TMV is considered to be unlikely
DISCUSSION
In our preliminary communication (1) it was reported that approxi- mately 80 y of threonine were released from 10 mg. of TMV by the action of carboxypeptidase. However, more extensive studies, also involving reevaluation of threonine standards, have yielded a revised value reported herein of 69 (f2) y of threonine per 10 mg. of virus. On this basis it may be calculated that 2900 (f100) residues of threonine are released per mole of TMV, assuming a molecular weight of 50,000,OOO for the virus.
By analogy with the action of carboxypeptidase on synthetic substrates of known chemical structure (9), and in view of its successful application to the C-terminal group analysis of several protein substrates (al), it is
* These conditions were chosen because y-peptide linkages are known to be more labile to acid than normal a-peptide bonds (18) and because it would be predicted t,hat polythreonine would behave similarly to polyserine which has been shown to bc particularly labile in alkaline solution (19).
6 The release of aspartic acid upon treatment with dilute acid has also been noted with other proteins (20).
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

228 ACTION OF CARBOXYPEPTIDASE ON TMV
reasonable to assume that the threonine residues released from the virus represent the C-terminal amino acids of a corresponding number of poly- peptide chains in the virus molecule.6 Accordingly, it may be postulated that each constituent peptide chain represents a chemical subunit, of mo- lecular weight 17,000, within the virus molecule. If a fundamental sub- unit is calculated on the basis of 1 cysteine residue per peptide chain’ of the virus, a value of approximately 18,000 is derived for its molecular weight. The presence of such a repeating subunit in the virus molecule is also consistent with the results of x-ray diffraction studies (22, 23), which indicate a great degree of regularity in the virus structure and the presence of regular repeat units.
Although the precise molecular size of the protein moiety obtained by treating TMV with sodium dodecyl sulfate is not known, the material ap- pears to behave like a protein of low molecular weight (3) ; in fact, prelim- inary data from this laboratory indicate that it has a molecular weight in the range of 10,000 to 20,000 (4). A molecule of this size is compatible with the calculated subunit proposed above, and, further, the fact that threonine is released from such a protein preparation in an amount which corresponds to 1 mole of threonine per unit of molecular weight 17,000 provides strong support for the concept that the virus molecule is com- posed of a series of repeating fundamental units.
Additional support for the subunit is provided by the kinetics of the virus-carboxypeptidase reaction, which by conformity to the Michaelis- Menten theory indicate a uniformity of the threonine released and hence imply an origin from identical subunits.
The fact that TMV is dissociated by treatment with sonic vibrations and by detergents indicates that the structural units of TMV are probably held together by relatively weak bonds such as hydrogen bridges. This view is strengthened by the present findings because no new C-terminal groups appear to be created by sonic and detergent treatments of the vi- rus,* and in all cases essentially the same quantity of threonine is released per unit weight of virus.
The nature of the amino acid residues which occur adjacent to C-ter- minal threonine in the virus molecule has not been established, but it appears that the residual structure resulting from the enzymatic cleavage of threonine does not meet the conditions required for the continued action
6 It is possible that the reaction olxxxvetl with TMV is not due to c:~rl~os~l~c~l~titlase but to a contaminating enzyme not susceptible to inhibition by diisopropyl fluoro- phosphate. However, the few anomalous reactions reported with c:~rl~os~peptitlasc so far (21) have proved to be inhibited by DFP, and we feel that the idea of a contami- nant is a rather unlikely explanation of the pronounced reaction observed with TXIV.
7 Cysteine is the amino acid which occurs in least amount in TMV (5). 8 Likewise, no N-terminal groups appear to be produced in detergent-treated virus
(4) *
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. I. HARRIS .4ND C. .4. KNIGHT 229
of carboxypeptidase. Some of the specificity requirements of this enzyme have been established by studies on synthetic substrates (9), but it has become apparent from recent studies on proteins and polypeptides (21, 24, 25) that the specificity of carboxypeptidase is dependent not only on the nature of the amino acid sequences but also on over-all environmental conditions. Nevertheless, by analogy with the B chain of insulin, in which alanine is the only amino acid released from the terminal sequence . Pro.Lys.Ala, an analogous sequence (viz. .Pro.Lys.Thr) can be postu- lated to explain the specific release of threonine from TMV.g
The observed changes in serological specificity and in electrophoretic mobility of the carboxypeptidase-treated virus both indicate a significant alteration of the surface chemistry of the virus, and it seems likely that determination of the actual nature of the penultimate residues in TMV will contribute something to the understanding of both electrophoresis and serology.
The present structural impasse with TMV illustrates the limitations of the carboxypeptidase method for the elucidation of amino acid sequences from the carboxyl end of polypeptide chains. An unequivocal answer to this question must await the development of other methods applicable to stepwise degradation from the carboxyl end.
Finally, in considering t,he peptide chains of TMV, it is apparent that the C-terminal end is only half the picture. The N-terminal groups of TM\’ and its dissociated protein moiety have been sought by the DNFl3 aud phenylisothiocyanate procedures (4). No appreciable amounts of such groups, however, were found in any of the preparations studied; furthermore, the c-amino groups of the lysine residues in the virus appear to account for its total amino nitrogen content when estimated by the Van Slyke procedure (26). In view of these findings it was postulated (4) that the a-NH2 groups of the N-terminal residues in the virus molecule may be masked by combination with the w-carboxyl groups of aspartic and glu- tamic acid residues, thus forming 6-shaped peptide chains containing C-ter- minal threonine. A similar hemicyclic structure has been proposed for tropomyosin by Bailey (27).
The authors are indebted to Dr. Arthur B. Pardee for helpful suggestions concerning the kinetic aspects of this investigation.
SUMMARY
The action of carboxypeptidase on tobacco mosaic virus (TRW), soni- rally treated TRIV, and detergent-treated TMV has been investigated.
9 Evidence has recently been obtained by Dr. Niu and Dr. Fraenkel-Conrat, of this laboratory, using a hydraziuolysis technique, that the actual sequence is . . . Pro.Ala.Thr.
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

230 ACTION OF CARBOXYPEPTIDASE ON TMV
In all cases, threonine exclusively is released as a result of the enzymatic action. The maximal amount of threonine released is found to be the same for all three types of virus preparations, although the rates of release are different. The reaction with TMV is also shown to conform to the Michaelis-Menten theory for an enzyme-catalyzed reaction at constant enzyme concentration.
The biological properties, and the size, shape, and density of the car- boxypeptidase-treated virus are found to be indistinguishable from those of the untreated virus. However, the electrophoretic mobility and the serological properties of the treated virus are demonstrably different.
From quantitative determination it is calculated that 2900 (&loo) thre- onine residues are released from TMV by the action of carboxypeptidase, and it is concluded that these represent the C-terminal groups of an equal number of peptide chains in the virus structure. From these, and other considerations, it is concluded that TMV is comprised of about 2900 sub- units, each having a molecular weight of approximately 17,000.
BIBLIOGRAPHY
1. Harris, J. I., and Knight, C. A., Nature, 170, 613 (1952). 2. Oster, G., J. Gen. Physiol., 31, 89 (1947). 3. Sreenivasaya, M., and Pirie, N. W., Biochem. J., 32, 1707 (1938). 4. Fraenkel-Conrat, H., and Singer, B., J. Am. Chem. Sot., 76, 180 (1954). 5. Knight, C. A., J. Biol. Chem., 171,297 (1947). 6. Elkins-Kaufman, E., and Neurat,h, H., J. Riol. Chew., 176, 893 (1948). 7. Fraser, D., and Powell, R. E., J. Bio2. Chem., 167,803 (1950). 8. Michaelis, I,., and Menten, M. I,., Biochem. Z., 49, 333 (1913). 9. Neurath, H., and Schwert, G. W., Chem. Rev., 46, 69 (1950).
10. Moore, S., and Stein, W. H., J. Biol. Chem., 176, 367 (1948). 11. Levy, A. I,., Natwe, 174, 126 (1954). 12. Williams, R. C., Backus, R. C., and Steere, It. L., J. Am. Chem. Sot., 73, 2062
(1951). 13. Partridge, S. M., Nature, 169,496 (1952). 14. Thompson, A. R., Nature, 169, 495 (1952). 15. Backus, R. C., and Williams, R. C., J. Appl. Phys., 21. 11 (1950). 16. Bawden, F. C., and Pirie, N. W., Brit. J. Exp. Path., 18, 275 (1937). 17. Knight, C. A., J. Exp. Med., 80,83 (1944). 18. Binkley, F., Fujii, S., and Kimmel, J. R., J. Biol. Chem., 186, 159 (1950). 19. Frankel, M., Cordova, S., and Breuer, M., J. Chem. Sot., 1991 (1953). 20. Partridge, S. M., and Davis, H. F., hratzrre, 166. 62 (1950). 21. Fraenkel-Conrat, H., Harris, J. I., and Levy, A. L., in Glick, D., Methods of bio-
rhcmiral analysis, Nrw York, 2, 359 (1955). 22. Bernal, J. D., and Fankuchcn, I., J. Gen. Physiol., 26, 111 (1941). 23. Watson, J. D., Biochim. et biophys. acta, 13. 10 (1954). 24. Sanger, F., and Thompson, E. 0. P., Biochem. J., 63,366 (1953). 25. Harris, .J. I., and Li, C. H., .I. ilm. Chem. Sot., 76, 3607 (1954). 26. Miller, G. J,., and Stanley, W. hf., J. Biol. Chew., 141, 905 (19-11). 27. Bailey, K., Proc. Roy. Sot. London, Series B, 141, 45 (1953).
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J. Ieuan Harris and C. A. KnightMOSAIC VIRUS
CARBOXYPEPTIDASE ON TOBACCO STUDIES ON THE ACTION OF
1955, 214:215-230.J. Biol. Chem.
http://www.jbc.org/content/214/1/215.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/214/1/215.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on April 1, 2018
http://ww
w.jbc.org/
Dow
nloaded from








![Glutamate carboxypeptidase II gene knockout attenuates ... · metabotropic glutamate receptor (mGluR3) [- 7–9]. Acti vating mGluR3 by NAAG reduces the synaptic glutamate ... (Leica](https://img.pdfslide.us/doc/110x75/5c4d740293f3c34aee567cc7/glutamate-carboxypeptidase-ii-gene-knockout-attenuates-metabotropic-glutamate.jpg)




















