Embed Size (px)
Citation preview

A cyclin-dependent protein kinase, CDKC2, colocalizes withand modulates the distribution of spliceosomal componentsin Arabidopsis
Georgios Kitsios†, Konstantinos G Alexiou, Max Bush, Peter Shaw and John H Doonan*
John Innes Centre, Colney Lane, Norwich NR4 7UH, UK
Received 10 October 2007; revised 3 December 2007; accepted13 December 2007.*For correspondence (fax +44 1603 450045; e-mail [email protected]).†Present address: Agricultural University of Athens, Department of Agricultural Biotechnology, Iera Odos 75, 11855 Athens, Greece.
Summary
Cyclin-dependent kinases (CDKs) play key regulatory roles in diverse cellular functions, including cell-cycle
progression, transcription and translation. In plants, CDKs have been classified into several groups, named
A through to G, but the functions of most are poorly characterized. CDKCs are known to phosphorylate the
C-terminal domain (CTD) of RNA polymerase II (RNAP II), and therefore the CDKC-cyclinT (CycT) complex may
have a role similar to the animal CDK9-CycT complex of the positive transcription elongation factor b (P-TEFb).
However, we found that the predicted structure of the Arabidopsis CDKC2 protein is more similar to the
mammalian cdc2-related kinase, CRK7, than to CDK9. CRK7 is proposed to link transcription with splicing, and
CDKC2 contains all the structural features of CRK7 that make the latter distinct from CDK9. Consistent with
this, we show that GFP-CDKC2 fusion proteins co-localize with spliceosomal components, that the expression
of CDKC2 modifies the location of these components, and that co-localization was dependent on the
transcriptional status of the cells and on CDKC2-kinase activity. We propose, therefore, that the Arabidopsis
CDKC2 combines the functions of both CRK7 and CDK9, and could also couple splicing with transcription.
Keywords: cyclin-dependent kinases (CDK), splicing factors, transcription, RNA polymerase II, Arabidopsis,
cell cycle.
Introduction
Cyclin-dependent kinases (CDKs) are a large family of serine/
threonine protein kinases that belong to the CMGC group
(Hanks, 2003). By definition, CDKs require binding with a
cyclin for their activity. CDKs were first discovered in yeasts,
and their function in cell-cycle control was defined during
genetic studies of the yeast cell division cycle, resulting in
their designation as cdc genes. Thus, the cdc2 gene for
Schizosaccharomyces pombe and CDC28 for Saccharomy-
ces cerevisiae (Hartwell et al., 1974; Hindley and Phear, 1984;
Nasmyth and Reed, 1980; Nurse and Thuriaux, 1980) encode
the prototypical CDK.
Several cdc2/CDC28-related genes have been identified
subsequently, thereby defining a much larger family of
proteins. CDK-related proteins from plants have been
categorized into eight groups, CDKA–CDKG (Mironov et al.,
1999; Vandepoele et al., 2002), and a newly defined CDK-
like group (CKLs) with 15 members (CKL1–CKL15; Menges
et al., 2005). The biological processes that require CDK
activity in plants are beginning to emerge. CDKA1 is the
homolog of the prototypic cdc2 protein from yeast, and is
a key regulator of the plant cell cycle (Dewitte and Murray,
2003; Nowack et al., 2006; Iwakawa et al., 2006). The CDKB
family is unique to plants and contains two subclasses, B1
and B2. This family is cell-cycle regulated at the level of
both transcript and protein accumulation. Members of the
B1 subclass are expressed from the onset of the S phase
until mitosis, whereas those belonging to the B2 subclass
are expressed during the G2–M transition (Fobert et al.,
1996; Segers et al., 1996). Members of CDKD belong to the
family of CDK-activating kinases or CAKs, one role of
which is to regulate the activation of CDKs through
phosphorylation of a conserved residue within the T-loop
(Thr161 in CDKA1). There are four potential CAK orthologs
in Arabidopsis. The three members of the D class are
closely related to the human CDK-activating kinase CDK7,
and the fourth potential CDKD gene has been recently
ª 2008 The Authors 1Journal compilation ª 2008 Blackwell Publishing Ltd
The Plant Journal (2008) doi: 10.1111/j.1365-313X.2008.03414.x

proposed to define a distinct class, designated as CDKF1
(Shimotohno et al., 2003, 2004, 2006).
The plant CDKC genes have been proposed to be homo-
logs of mammalian CDK9, and the plant CDKC-CycT com-
plex was suggested to have a role similar to the mammalian
CDK9/CycT complex of the positive transcription elongation
factor b (P-TEFb; Barroco et al., 2003; Fulop et al., 2005). In
mammalian cells, P-TEFb regulates transcription elongation
by phosphorylating the C-terminal domain (CTD) of RNA
polymerase II (RNAP II; Price, 2000). The functions of other
CDK and CKL genes remain to be investigated.
We compared the amino acid sequences of Arabidopsis
CDKs with CDKs from other species, particularly from
animals. Our results indicated that the protein sequence of
CDKC2 was related to both CRK7 and CDK9, but more closely
to CRK7. CRK7 is a novel cdc2-related kinase that links
transcription with splicing, as it phosphorylates the CTD of
RNAP II and associates with SR-splicing factors. Addition-
ally, CRK7 has structural characteristics that are not found in
CDK9 (Ko et al., 2001). To investigate if CDKC2 was likely to
have a role in splicing, we made GFP fusions and charac-
terized their locations in the cell, relative to known splicing
factors and in response to drugs that perturb transcription.
Our results indicate that CDKC2 dynamically associates with
spliceosomal components in a transcription- and kinase-
dependent manner. These data are consistent with the idea
that this plant CDK has a role similar to CRK7 in linking
transcription with the splicing apparatus.
Results
Bioinformatic analysis suggests that the Arabidopsis
CDKC2 is closely related to the mammalian CRK7
As a first step towards understanding the possible function
of the Arabidopsis CDKC genes, we compared their struc-
tural features with other classes of CDK, particularly those
from animals. The Arabidopsis genome contains two CDKC
genes, CDKC1 and CDKC2, which share a variant cyclin-
binding motif, PITAIRE. The entire protein sequence of
CDKC2 was used in a BLAST search against the human
genome (NCBI database, 15-9-07), which revealed that the
two proteins most similar to CDKC2 were members of
either the cholinesterase-related cell division kinases
(CHED-kinases) or the cdc2-related protein-RS (CrkRS)
family. CHED-kinases are involved with megakaryocyte
differentiation in hematopoiesis (Lapidot-Lifson et al.,
1992), and the relation of plant CDKCs with these mam-
malian proteins has been reported previously (Lessard
et al., 1999). The other structurally-related mammalian
protein is CrkRS, also known as CRK7, which is a novel
cdc2-related kinase proposed to link transcription with
splicing, as it phosphorylates the CTD of RNAP II and
associates with components of the SR-splicing factors
(Ko et al., 2001). Although CDKC2 does share significant
homology with mammalian CDK9, the BLAST score with
CDK9 is significantly lower than with CRK7. Reciprocal
BLAST searches using CDK9 and CRK7 both identified
CDKC2 as the most similar plant protein (data not shown).
This suggested to us that CDKC2 may have other roles in
addition to its previously reported functions in transcrip-
tional elongation (Barroco et al., 2003; Fulop et al., 2005).
Similar results were obtained when the protein sequence of
CDKC1 was used as a probe in a BLAST search against the
human genome (data not shown), suggesting that both
CDKC proteins were more similar to CRK7 than to CDK9.
Furthermore, both CDKC2 and CRK7 have N- and
C-terminal extensions on either side of the core kinase
domain, whereas CDK9 lacks such extensions (Figure 1a).
The N- and C-terminal amino acid extensions on CRK7 are
much longer than those of CDKC2 (723 and 452 residues
respectively, vs 22 and 181), but they share several features.
The hydropathic profiles of the extensions of both CDKC2
and CRK7 indicated that they both contain mainly hydro-
philic residues (data not shown). Amino acid inserts and
extensions within the sequence of protein kinases are usually
composed of hydrophilic residues, and represent surface
elements that do not disrupt their basic core structure (Hanks
and Hunter, 1995). CRK7 also contains an RS-rich domain, a
prominent feature of the pre-mRNA splicing factors. Twenty-
one RS motifs are present within the N-terminal amino
extension of the CRK7 protein, and only one in the remaining
sequence (Ko et al., 2001). On the other hand, analysis of the
CDKC2 sequence revealed the presence of only two RS
motifs (one within the N-terminal extension of the protein
and the other within the core kinase), whereas CDK9 had only
one (Figure 1a). A characteristic feature of CRK7, which is
also present in Arabidopsis CDKC2, is that no RS motifs are
found after the ‘PPE’ sequence, corresponding to the ‘APE
motif’ of protein kinases (Ko et al., 2001). The ‘APE’ motif
represents the boundary of the kinase central core, as almost
all of the characterized functional residues are found
before that motif (Hanks and Hunter, 1995). This indicates
the importance of RS motifs in the functional properties of
those proteins. In CDK9, however, the only RS motif present
is located after the ‘PPE’ triplet, and is consequently outside
of the central core of protein kinases (Figure 1a). This
suggests that the RS motif of CDK9 is unlikely to be
implicated in the functional properties of the protein. The
lower number of RS motifs in CDKC2 compared with CRK7
could be a plant-specific characteristic.
Comparison of central core regions indicated that the
region between the ‘GxGxxG’ and the ‘APE’ motifs of CDKC2
was significantly more similar to CRK7 (60.7%) than to CDK9
(51.1%). CDKC2 and CRK7 also share a key regulatory motif,
T14/Y15, which CDK9 lacks. Y15 is substituted by Phe in
CDK9 (Figure 1b), suggesting that this residue is not
involved in the regulation of protein function (Betts and
2 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
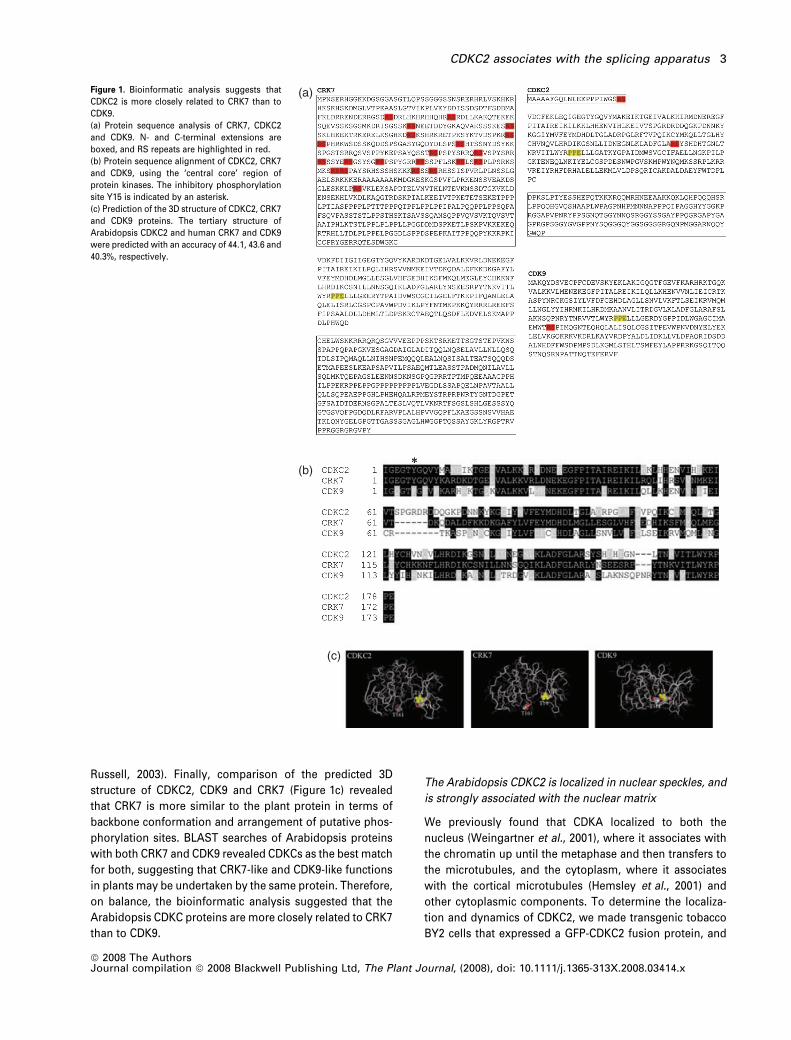
Russell, 2003). Finally, comparison of the predicted 3D
structure of CDKC2, CDK9 and CRK7 (Figure 1c) revealed
that CRK7 is more similar to the plant protein in terms of
backbone conformation and arrangement of putative phos-
phorylation sites. BLAST searches of Arabidopsis proteins
with both CRK7 and CDK9 revealed CDKCs as the best match
for both, suggesting that CRK7-like and CDK9-like functions
in plants may be undertaken by the same protein. Therefore,
on balance, the bioinformatic analysis suggested that the
Arabidopsis CDKC proteins are more closely related to CRK7
than to CDK9.
The Arabidopsis CDKC2 is localized in nuclear speckles, and
is strongly associated with the nuclear matrix
We previously found that CDKA localized to both the
nucleus (Weingartner et al., 2001), where it associates with
the chromatin up until the metaphase and then transfers to
the microtubules, and the cytoplasm, where it associates
with the cortical microtubules (Hemsley et al., 2001) and
other cytoplasmic components. To determine the localiza-
tion and dynamics of CDKC2, we made transgenic tobacco
BY2 cells that expressed a GFP-CDKC2 fusion protein, and
(a)
(b)
(c)
Figure 1. Bioinformatic analysis suggests that
CDKC2 is more closely related to CRK7 than to
CDK9.
(a) Protein sequence analysis of CRK7, CDKC2
and CDK9. N- and C-terminal extensions are
boxed, and RS repeats are highlighted in red.
(b) Protein sequence alignment of CDKC2, CRK7
and CDK9, using the ‘central core’ region of
protein kinases. The inhibitory phosphorylation
site Y15 is indicated by an asterisk.
(c) Prediction of the 3D structure of CDKC2, CRK7
and CDK9 proteins. The tertiary structure of
Arabidopsis CDKC2 and human CRK7 and CDK9
were predicted with an accuracy of 44.1, 43.6 and
40.3%, respectively.
CDKC2 associates with the splicing apparatus 3
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

followed the distribution of the fusion protein through the
cell-division cycle. During the interphase, the GFP-CDKC2
protein is localized primarily in the nucleus (Figure 2a)
where it accumulates in small bodies within the nucleo-
plasm. The CDKC2 fusion appeared to be more diffuse in
BY2 cells, probably because of differences in nuclear
organization between the two species or because of vari-
ations at the expression levels of the fusion proteins
(compare Figures 2a and 3a). Unlike CDKA, GFP-CDKC2
is not retained on the chromatin during mitosis (Figure 2b–
d), and appears to be diminished in the chromatin regions
throughout mitosis. GFP-CDKC2 becomes diffusely asso-
ciated with the spindle region during the metaphase
(Figure 2c). During the anaphase (Figure 2d), GFP-CDKC2
begins to accumulate in regions adjacent to the incipient
daughter nuclei, and regains a more discretely nuclear
localization pattern in the telophase (Figure 2e and f).
The localization of CDKC2 to nuclear bodies, and its
dynamic relocalization during the cell cycle, suggested to us
that it may play a role in transcript processing, possibly in
splicing. To further characterize the association of CDKC
with nuclear bodies, we determined its subcellular localiza-
tion by transiently expressing translational fusions between
CDKC genes and GFP in Arabidopsis cell suspensions.
Subsequently, we subjected the expressing cells to various
extractions and drug treatments to test the interaction of
CDKC with nuclear components.
The Arabidopsis GFP-CDKC2 fusion protein localized to
the nucleus, with a reduced level in the nucleolus, and
produced a network of bright speckles and diffuse nucleo-
(a)
(b)
(c)
(d)
(e)
(f)
Figure 2. Subcellular distribution of Arabidopsis
CDKC2-GFP in stably transformed BY2 cells
during the cell cycle.
(a) GFP-CDKC2 subcellular localization during
the interphase. GFP-CDKC2 is localized in the
nucleus and in speckle-like structures. The GFP
image is shown on the left, and the brightfield
image is shown on the right.
(b–f) GFP-CDKC2 subcellular localization during
mitosis. GFP images are shown on the far-left
panels, 4¢,6-diamidino-2-phenylindole (DAPI)-
stained images are shown on the middle-left
panels, the merged images are shown on the
middle-right panels and the brightfield images
are shown on the far-right panels: (b) prophase;
(c) metaphase; (d) anaphase; (e) early telophase;
and (f) late telophase. Scale bar: 5 lm.
4 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

plasmic staining (Figure 3a). The same localization pattern
was obtained for the GFP-CDKC1 fusion protein (data not
shown). Western blot analysis, using an anti-GFP antibody,
revealed a band of approximately 82 kDa, which corre-
sponded to the predicted size of the fusion protein, and
established that the fusion protein remained intact
(Figure 3b).
To determine how tightly CDKC2 interacts with nuclear
components, cells expressing the GFP fusion protein were
extracted with different levels of non-ionic detergents:
(i) 10% (v/v) NP-40 (Nonidet P-40), (ii) 1% (v/v) Triton X-100
and (iii) 10% (v/v) DMSO + 0.05% (v/v) NP-40 + 0.45 M
Sorbitol, referred to hereafter as the ‘extraction buffer’).
Under such conditions, membranes and the nuclear enve-
lope are dissolved, and soluble proteins are differentially
extracted (Weingartner et al., 2001), but the GFP–CDKC2
fusion protein was completely resistant to extraction, even
after prolonged treatment (Figure 3c,e). The signal from
soluble GFP control cells was completely eliminated by
these treatments (Figure 3d). To test if the speckled locali-
zation pattern of CDKC2 was dependent on the integrity of
either DNA or RNA, Arabidopsis cells expressing the
GFP-CDKC2 fusion protein were treated with DNase and/or
RNase, and were then stained with 4¢,6-diamidino-2-pheny-
lindole (DAPI) or ethidium bromide. Nuclease treatment did
not substantially alter the speckled pattern of GFP-CDKC2,
although DNA in the nucleus was digested, as revealed by
DAPI staining (Figure 3f,g). Thus, we concluded that
GFP-CDKC2 was tightly localized to speckles, but this
probably did not require direct association with either RNA
or chromatin.
The Arabidopsis CDKC2 co-localizes in vivo
with spliceosomal components
The plant splicing machinery is located in the nucleus in a
distinctive distribution that is well characterized (Lorkovic
and Barta, 2004a; Tillemans et al., 2005), and if CDKC2 plays
a role in splicing, one might expect the protein to be found
co-located with splicing factors. To test this, we localized
GFP-CDKC2 in cells that were also expressing the splicing
factor SRp34 (Lazar et al., 1995) fused to RFP (Figure 4a).
Co-expression resulted in a very finely dispersed pattern
of speckles for both the GFP and RFP fusion proteins,
suggesting that expression of SRp34 promoted the redis-
tribution of CDKC2 into smaller structures.
Merging of the GFP and RFP signals indicated a similar
distribution pattern for the two fusion proteins (Figure 4a).
To quantify the correlation between the green and the red
channels more precisely, we plotted line-intensity profiles
for each channel with a series of randomly selected lines
through the merged images of nuclei. Comparing the line
profiles from the two channels showed that the positions of
the maxima and minima in the two channels coincided
precisely. A significant level of RFP-SRp34 was also found in
the nucleolus. The RFP-SRp34 fusion protein stained the
nucleolar cavity less intensively when expressed alone in
Arabidopsis cells (Figure 4a,b), as has been reported previ-
ously (Fang et al., 2004; Lorkovic et al., 2004b). Therefore,
CDKC2 localizes in the same regions of the nucleus as
SRp34, but co-expression of the two proteins modifies the
distribution of each protein.
Another protein, a cyclophilin containing RS repeats
(CypRS64), has been proposed to associate with and play a
role in regulating the splicing machinery in Arabidopsis.
CypRS64 was found to associate with the splicing factor
SRp34, and a role for cyclophilins in the regulation of
spliceosomal dynamics was proposed (Lorkovic et al.,
2004b). To determine whether CDKC2 also co-localizes with
CypRS64, we co-transformed Arabidopsis cells with plas-
mids carrying the GFP-CDKC2 and RFP-CypRS64 fusion
constructs. Co-expression of GFP-CDKC2 and RFP-CypRS64
resulted in the co-localization of the two fusion proteins into
large dots (Figure 4c). Both fusion proteins stained the
nucleolus, and areas of co-localization were also found
within the nucleolar cavity (Figure 4d). A more diffuse
pattern and less defined dots were observed when the
RFP-CypRS64 fusion protein was expressed alone in Ara-
bidopsis cells. Also, the nucleolar cavity was less intensively
stained (Figure 4e). The relocalization of CDKC2 upon
co-expression with CypRS64 could result from an interaction
between the two proteins, as was observed previously with
CypRS64 and SRp34 (Lorkovic et al., 2004b). Our results,
combined with previous findings, indicate that CDKC2,
SRp34 and CypRS64 all locate to the same subcompartment
of the nucleus, and that they all influence each other’s
distribution.
Co-localization of splicing components with other mem-
bers of Arabidopsis CDK families demonstrated different
localization profiles (Figures S1 and S2). Co-expression of
CDKB2;1 with SRp34 led to their co-localization in the
nucleus, with CDKB2;1 also being localized in the cytoplasm
(Figure S1). Moreover, co-expression of two proteins did not
alter the other’s localization profile, as compared with the
profile observed when each protein was expressed alone.
Co-expression of SRp34 with CDKD demonstrated that
localization of the splicing factor was not altered (Figure S2,
compare with Figure 4). CDKD was relocalized into fine
speckles, but careful observation of the two protein locali-
zation profiles, in combination with line-intensity profiling,
showed that co-localization between CDKD and SRp34 was
restricted to a few loci (Figure S2). The human homolog of
CDKD, CDK7, was reported to localize in coiled bodies,
structures that contain snRNPs of the U family (Jordan et al.,
1997). Also, SR-splicing factors associate with U-snRNPs,
and facilitate their recruitment to the 5¢ splicing site and
branch-point sequences (Reed, 2000; Hastings and Krainer,
2001). Thus, relocalization of CDKD upon overexpression of
CDKC2 associates with the splicing apparatus 5
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
SRp34 could result from the association of the latter with
U-snRNP. Therefore, the above observations indicate that
co-localization of splicing components with CDKC2 seems to
be specific, and does not result from changes in nuclear
organization.
Localization of CDKC2, SRp34 and CypRS64 is dependent
on the transcriptional status of the cell
Most pre-mRNA splicing occurs co-transcriptionally
(Proudfoot et al., 2002), so that the transcriptional status of
the cell affects the spatial distribution of the SR-splicing
factors. In general, inhibition of transcription causes a
decrease in the numbers of nuclear speckles, and the
redistribution of splicing factors into larger storage bodies
(Spector et al., 1983; Ali et al., 2003; Docquier et al., 2004;
Fang et al., 2004; Tillemans et al., 2005). Therefore, we tes-
ted if the distribution of CDKC2 was influenced by the level of
RNAP II transcription in vivo, and compared this with the
localization of SR-splicing factors. Arabidopsis cell suspen-
sions transiently expressing the GFP-CDKC2, either alone or
with RFP-SRp34 or RFP-CypRS64 fusion proteins, were
incubated with the transcription inhibitor 5,6-dichloro-1-b-D-
ribofuranosylbenzimidazole (DRB; Sehgal et al., 1976).
When cells expressing GFP-CDKC2 were incubated in DRB
for 2 h, the fusion protein accumulated in the nucleolus
(Figure S3), and the relative level of fusion protein in the
nucleoplasm seemed diminished, although speckles were
(f)
(g)
(a)
(b)(d)
(e)
(c)
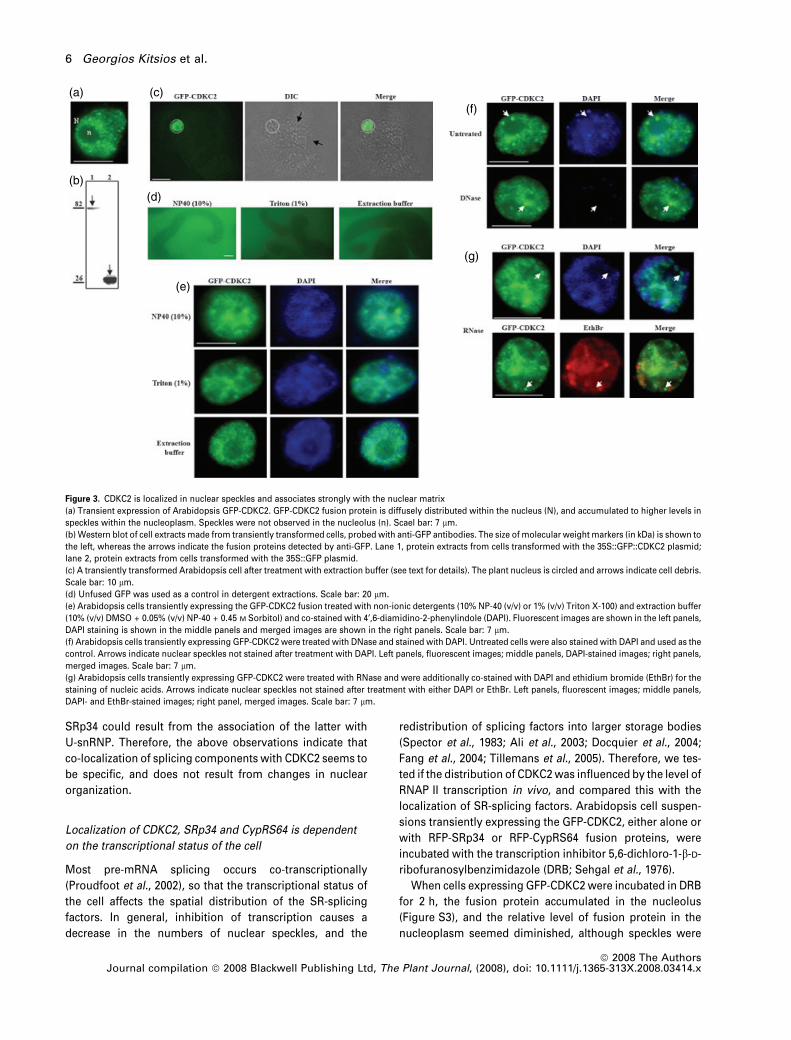
Figure 3. CDKC2 is localized in nuclear speckles and associates strongly with the nuclear matrix
(a) Transient expression of Arabidopsis GFP-CDKC2. GFP-CDKC2 fusion protein is diffusely distributed within the nucleus (N), and accumulated to higher levels in
speckles within the nucleoplasm. Speckles were not observed in the nucleolus (n). Scael bar: 7 lm.
(b) Western blot of cell extracts made from transiently transformed cells, probed with anti-GFP antibodies. The size of molecular weight markers (in kDa) is shown to
the left, whereas the arrows indicate the fusion proteins detected by anti-GFP. Lane 1, protein extracts from cells transformed with the 35S::GFP::CDKC2 plasmid;
lane 2, protein extracts from cells transformed with the 35S::GFP plasmid.
(c) A transiently transformed Arabidopsis cell after treatment with extraction buffer (see text for details). The plant nucleus is circled and arrows indicate cell debris.
Scale bar: 10 lm.
(d) Unfused GFP was used as a control in detergent extractions. Scale bar: 20 lm.
(e) Arabidopsis cells transiently expressing the GFP-CDKC2 fusion treated with non-ionic detergents (10% NP-40 (v/v) or 1% (v/v) Triton X-100) and extraction buffer
(10% (v/v) DMSO + 0.05% (v/v) NP-40 + 0.45 M Sorbitol) and co-stained with 4¢,6-diamidino-2-phenylindole (DAPI). Fluorescent images are shown in the left panels,
DAPI staining is shown in the middle panels and merged images are shown in the right panels. Scale bar: 7 lm.
(f) Arabidopsis cells transiently expressing GFP-CDKC2 were treated with DNase and stained with DAPI. Untreated cells were also stained with DAPI and used as the
control. Arrows indicate nuclear speckles not stained after treatment with DAPI. Left panels, fluorescent images; middle panels, DAPI-stained images; right panels,
merged images. Scale bar: 7 lm.
(g) Arabidopsis cells transiently expressing GFP-CDKC2 were treated with RNase and were additionally co-stained with DAPI and ethidium bromide (EthBr) for the
staining of nucleic acids. Arrows indicate nuclear speckles not stained after treatment with either DAPI or EthBr. Left panels, fluorescent images; middle panels,
DAPI- and EthBr-stained images; right panel, merged images. Scale bar: 7 lm.
6 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
still visible. However, DRB treatment had a more dramatic
effect when GFP-CDKC2 was co-expressed with SRp34
(Figure 5). After 1 h in DRB, the fine, speckled localization
pattern was lost, and both proteins accumulated in larger
bodies, but the extent of co-localization between CDKC2 and
SRp34 temporally decreased, with considerable variation in
the content of the larger bodies. Some bodies contained
primarily one or other protein, but other bodies contained
both proteins (Figure 5a). Treatment for longer periods (3
and 5 h) resulted in the accumulation of GFP-CDKC2 and
RFP-SRp34 in even larger bodies, which decreased in
number and increased in the level of co-localization
(a)
(c)
(e)
(d)
(b)
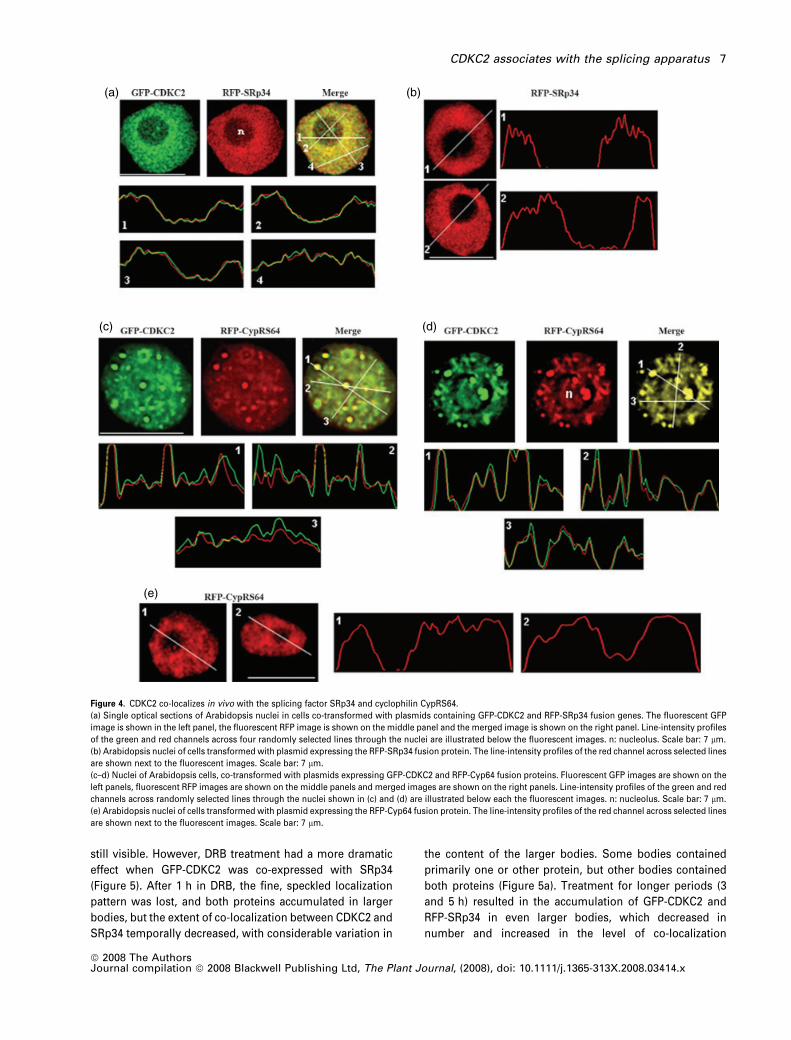
Figure 4. CDKC2 co-localizes in vivo with the splicing factor SRp34 and cyclophilin CypRS64.
(a) Single optical sections of Arabidopsis nuclei in cells co-transformed with plasmids containing GFP-CDKC2 and RFP-SRp34 fusion genes. The fluorescent GFP
image is shown in the left panel, the fluorescent RFP image is shown on the middle panel and the merged image is shown on the right panel. Line-intensity profiles
of the green and red channels across four randomly selected lines through the nuclei are illustrated below the fluorescent images. n: nucleolus. Scale bar: 7 lm.
(b) Arabidopsis nuclei of cells transformed with plasmid expressing the RFP-SRp34 fusion protein. The line-intensity profiles of the red channel across selected lines
are shown next to the fluorescent images. Scale bar: 7 lm.
(c–d) Nuclei of Arabidopsis cells, co-transformed with plasmids expressing GFP-CDKC2 and RFP-Cyp64 fusion proteins. Fluorescent GFP images are shown on the
left panels, fluorescent RFP images are shown on the middle panels and merged images are shown on the right panels. Line-intensity profiles of the green and red
channels across randomly selected lines through the nuclei shown in (c) and (d) are illustrated below each the fluorescent images. n: nucleolus. Scale bar: 7 lm.
(e) Arabidopsis nuclei of cells transformed with plasmid expressing the RFP-Cyp64 fusion protein. The line-intensity profiles of the red channel across selected lines
are shown next to the fluorescent images. Scale bar: 7 lm.
CDKC2 associates with the splicing apparatus 7
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
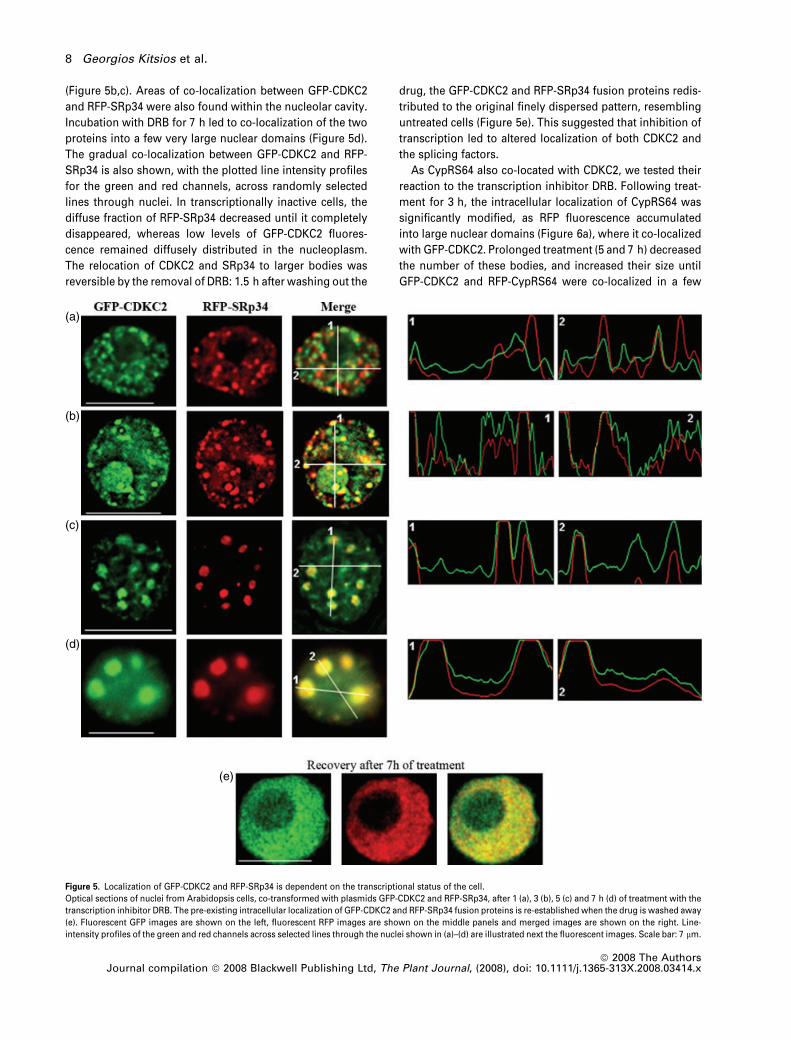
(Figure 5b,c). Areas of co-localization between GFP-CDKC2
and RFP-SRp34 were also found within the nucleolar cavity.
Incubation with DRB for 7 h led to co-localization of the two
proteins into a few very large nuclear domains (Figure 5d).
The gradual co-localization between GFP-CDKC2 and RFP-
SRp34 is also shown, with the plotted line intensity profiles
for the green and red channels, across randomly selected
lines through nuclei. In transcriptionally inactive cells, the
diffuse fraction of RFP-SRp34 decreased until it completely
disappeared, whereas low levels of GFP-CDKC2 fluores-
cence remained diffusely distributed in the nucleoplasm.
The relocation of CDKC2 and SRp34 to larger bodies was
reversible by the removal of DRB: 1.5 h after washing out the
drug, the GFP-CDKC2 and RFP-SRp34 fusion proteins redis-
tributed to the original finely dispersed pattern, resembling
untreated cells (Figure 5e). This suggested that inhibition of
transcription led to altered localization of both CDKC2 and
the splicing factors.
As CypRS64 also co-located with CDKC2, we tested their
reaction to the transcription inhibitor DRB. Following treat-
ment for 3 h, the intracellular localization of CypRS64 was
significantly modified, as RFP fluorescence accumulated
into large nuclear domains (Figure 6a), where it co-localized
with GFP-CDKC2. Prolonged treatment (5 and 7 h) decreased
the number of these bodies, and increased their size until
GFP-CDKC2 and RFP-CypRS64 were co-localized in a few
(a)
(b)
(c)
(d)
(e)
Figure 5. Localization of GFP-CDKC2 and RFP-SRp34 is dependent on the transcriptional status of the cell.
Optical sections of nuclei from Arabidopsis cells, co-transformed with plasmids GFP-CDKC2 and RFP-SRp34, after 1 (a), 3 (b), 5 (c) and 7 h (d) of treatment with the
transcription inhibitor DRB. The pre-existing intracellular localization of GFP-CDKC2 and RFP-SRp34 fusion proteins is re-established when the drug is washed away
(e). Fluorescent GFP images are shown on the left, fluorescent RFP images are shown on the middle panels and merged images are shown on the right. Line-
intensity profiles of the green and red channels across selected lines through the nuclei shown in (a)–(d) are illustrated next the fluorescent images. Scale bar: 7 lm.
8 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
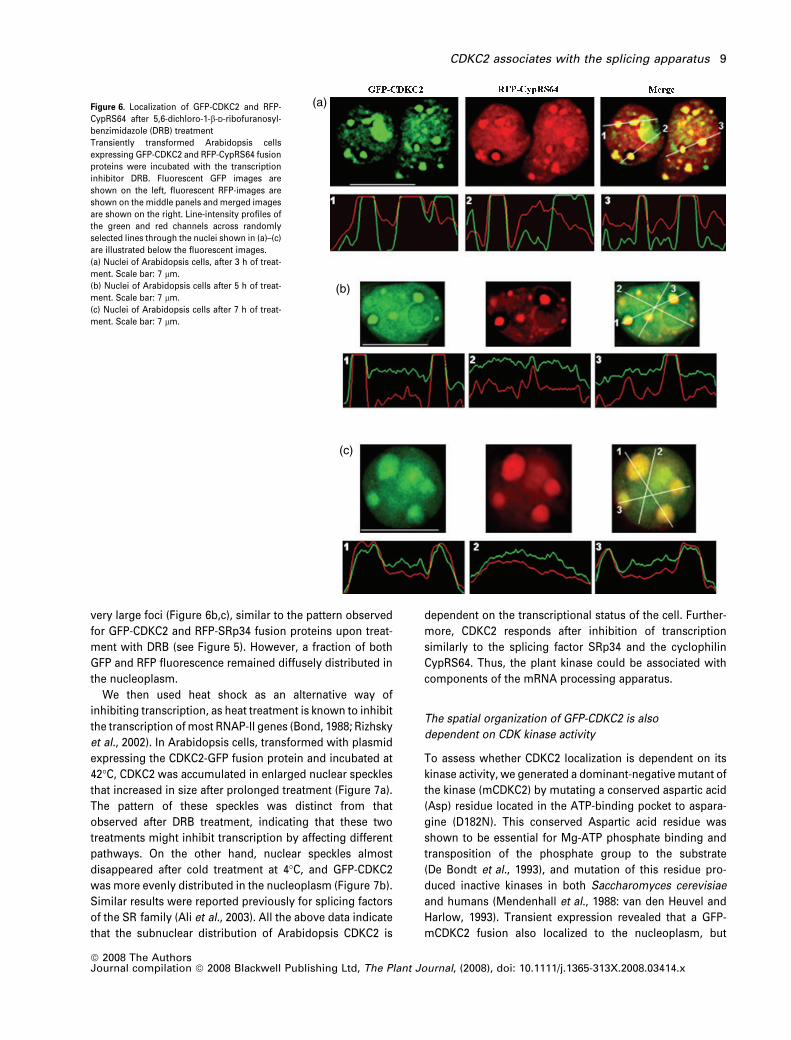
very large foci (Figure 6b,c), similar to the pattern observed
for GFP-CDKC2 and RFP-SRp34 fusion proteins upon treat-
ment with DRB (see Figure 5). However, a fraction of both
GFP and RFP fluorescence remained diffusely distributed in
the nucleoplasm.
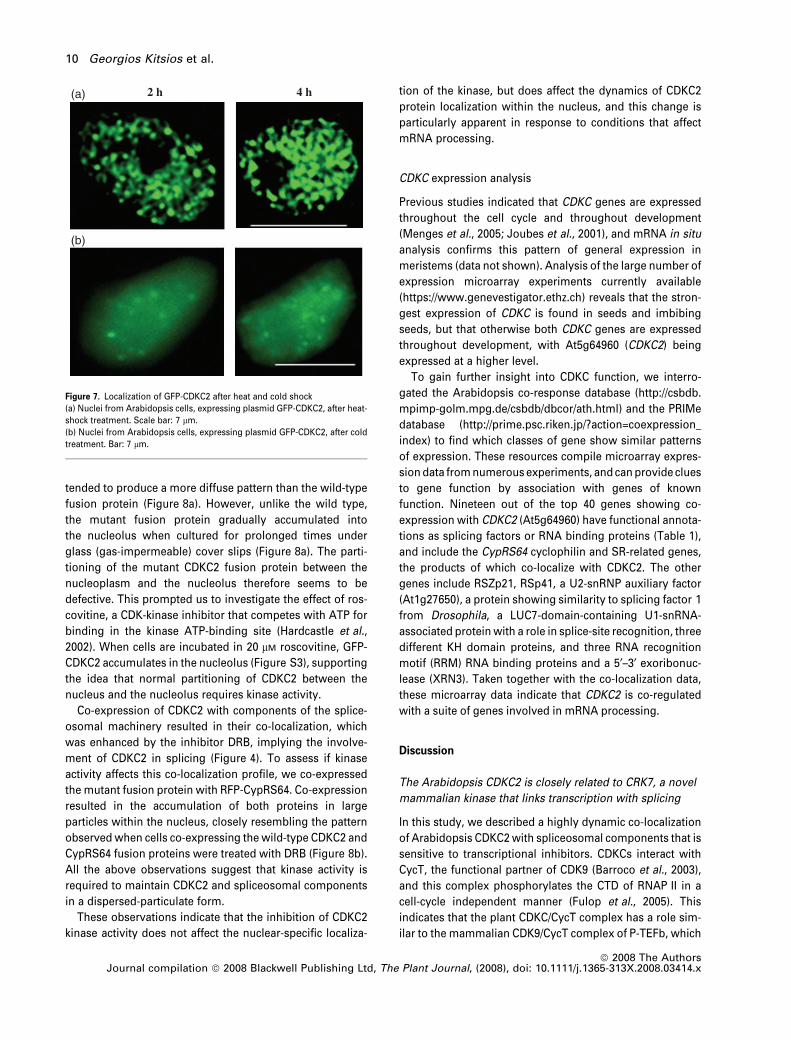
We then used heat shock as an alternative way of
inhibiting transcription, as heat treatment is known to inhibit
the transcription of most RNAP-II genes (Bond, 1988; Rizhsky
et al., 2002). In Arabidopsis cells, transformed with plasmid
expressing the CDKC2-GFP fusion protein and incubated at
42�C, CDKC2 was accumulated in enlarged nuclear speckles
that increased in size after prolonged treatment (Figure 7a).
The pattern of these speckles was distinct from that
observed after DRB treatment, indicating that these two
treatments might inhibit transcription by affecting different
pathways. On the other hand, nuclear speckles almost
disappeared after cold treatment at 4�C, and GFP-CDKC2
was more evenly distributed in the nucleoplasm (Figure 7b).
Similar results were reported previously for splicing factors
of the SR family (Ali et al., 2003). All the above data indicate
that the subnuclear distribution of Arabidopsis CDKC2 is
dependent on the transcriptional status of the cell. Further-
more, CDKC2 responds after inhibition of transcription
similarly to the splicing factor SRp34 and the cyclophilin
CypRS64. Thus, the plant kinase could be associated with
components of the mRNA processing apparatus.
The spatial organization of GFP-CDKC2 is also
dependent on CDK kinase activity
To assess whether CDKC2 localization is dependent on its
kinase activity, we generated a dominant-negative mutant of
the kinase (mCDKC2) by mutating a conserved aspartic acid
(Asp) residue located in the ATP-binding pocket to aspara-
gine (D182N). This conserved Aspartic acid residue was
shown to be essential for Mg-ATP phosphate binding and
transposition of the phosphate group to the substrate
(De Bondt et al., 1993), and mutation of this residue pro-
duced inactive kinases in both Saccharomyces cerevisiae
and humans (Mendenhall et al., 1988: van den Heuvel and
Harlow, 1993). Transient expression revealed that a GFP-
mCDKC2 fusion also localized to the nucleoplasm, but
(a)
(b)
(c)
Figure 6. Localization of GFP-CDKC2 and RFP-
CypRS64 after 5,6-dichloro-1-b-D-ribofuranosyl-
benzimidazole (DRB) treatment
Transiently transformed Arabidopsis cells
expressing GFP-CDKC2 and RFP-CypRS64 fusion
proteins were incubated with the transcription
inhibitor DRB. Fluorescent GFP images are
shown on the left, fluorescent RFP-images are
shown on the middle panels and merged images
are shown on the right. Line-intensity profiles of
the green and red channels across randomly
selected lines through the nuclei shown in (a)–(c)
are illustrated below the fluorescent images.
(a) Nuclei of Arabidopsis cells, after 3 h of treat-
ment. Scale bar: 7 lm.
(b) Nuclei of Arabidopsis cells after 5 h of treat-
ment. Scale bar: 7 lm.
(c) Nuclei of Arabidopsis cells after 7 h of treat-
ment. Scale bar: 7 lm.
CDKC2 associates with the splicing apparatus 9
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
tended to produce a more diffuse pattern than the wild-type
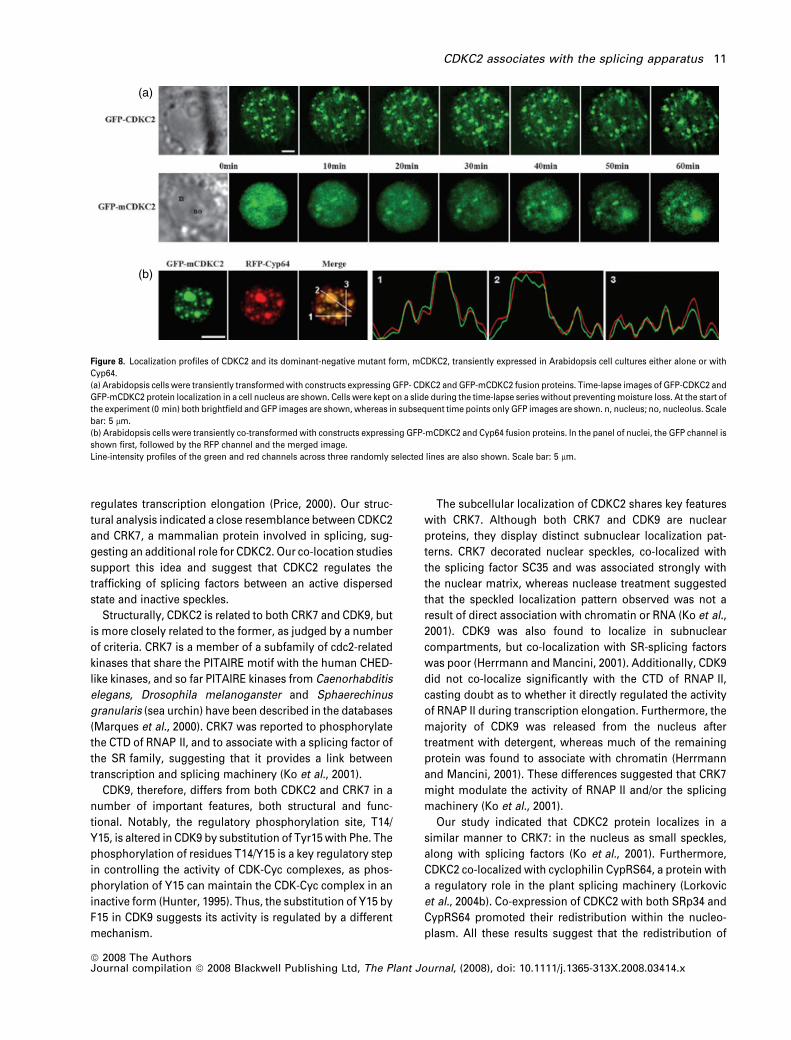
fusion protein (Figure 8a). However, unlike the wild type,
the mutant fusion protein gradually accumulated into
the nucleolus when cultured for prolonged times under
glass (gas-impermeable) cover slips (Figure 8a). The parti-
tioning of the mutant CDKC2 fusion protein between the
nucleoplasm and the nucleolus therefore seems to be
defective. This prompted us to investigate the effect of ros-
covitine, a CDK-kinase inhibitor that competes with ATP for
binding in the kinase ATP-binding site (Hardcastle et al.,
2002). When cells are incubated in 20 lM roscovitine, GFP-
CDKC2 accumulates in the nucleolus (Figure S3), supporting
the idea that normal partitioning of CDKC2 between the
nucleus and the nucleolus requires kinase activity.
Co-expression of CDKC2 with components of the splice-
osomal machinery resulted in their co-localization, which
was enhanced by the inhibitor DRB, implying the involve-
ment of CDKC2 in splicing (Figure 4). To assess if kinase
activity affects this co-localization profile, we co-expressed
the mutant fusion protein with RFP-CypRS64. Co-expression
resulted in the accumulation of both proteins in large
particles within the nucleus, closely resembling the pattern
observed when cells co-expressing the wild-type CDKC2 and
CypRS64 fusion proteins were treated with DRB (Figure 8b).
All the above observations suggest that kinase activity is
required to maintain CDKC2 and spliceosomal components
in a dispersed-particulate form.
These observations indicate that the inhibition of CDKC2
kinase activity does not affect the nuclear-specific localiza-
tion of the kinase, but does affect the dynamics of CDKC2
protein localization within the nucleus, and this change is
particularly apparent in response to conditions that affect
mRNA processing.
CDKC expression analysis
Previous studies indicated that CDKC genes are expressed
throughout the cell cycle and throughout development
(Menges et al., 2005; Joubes et al., 2001), and mRNA in situ
analysis confirms this pattern of general expression in
meristems (data not shown). Analysis of the large number of
expression microarray experiments currently available
(https://www.genevestigator.ethz.ch) reveals that the stron-
gest expression of CDKC is found in seeds and imbibing
seeds, but that otherwise both CDKC genes are expressed
throughout development, with At5g64960 (CDKC2) being
expressed at a higher level.
To gain further insight into CDKC function, we interro-
gated the Arabidopsis co-response database (http://csbdb.
mpimp-golm.mpg.de/csbdb/dbcor/ath.html) and the PRIMe
database (http://prime.psc.riken.jp/?action=coexpression_
index) to find which classes of gene show similar patterns
of expression. These resources compile microarray expres-
sion data from numerous experiments, and can provide clues
to gene function by association with genes of known
function. Nineteen out of the top 40 genes showing co-
expression with CDKC2 (At5g64960) have functional annota-
tions as splicing factors or RNA binding proteins (Table 1),
and include the CypRS64 cyclophilin and SR-related genes,
the products of which co-localize with CDKC2. The other
genes include RSZp21, RSp41, a U2-snRNP auxiliary factor
(At1g27650), a protein showing similarity to splicing factor 1
from Drosophila, a LUC7-domain-containing U1-snRNA-
associated protein with a role in splice-site recognition, three
different KH domain proteins, and three RNA recognition
motif (RRM) RNA binding proteins and a 5¢–3¢ exoribonuc-
lease (XRN3). Taken together with the co-localization data,
these microarray data indicate that CDKC2 is co-regulated
with a suite of genes involved in mRNA processing.
Discussion
The Arabidopsis CDKC2 is closely related to CRK7, a novel
mammalian kinase that links transcription with splicing
In this study, we described a highly dynamic co-localization
of Arabidopsis CDKC2 with spliceosomal components that is
sensitive to transcriptional inhibitors. CDKCs interact with
CycT, the functional partner of CDK9 (Barroco et al., 2003),
and this complex phosphorylates the CTD of RNAP II in a
cell-cycle independent manner (Fulop et al., 2005). This
indicates that the plant CDKC/CycT complex has a role sim-
ilar to the mammalian CDK9/CycT complex of P-TEFb, which
(a) 2 h 4 h
(b)
Figure 7. Localization of GFP-CDKC2 after heat and cold shock
(a) Nuclei from Arabidopsis cells, expressing plasmid GFP-CDKC2, after heat-
shock treatment. Scale bar: 7 lm.
(b) Nuclei from Arabidopsis cells, expressing plasmid GFP-CDKC2, after cold
treatment. Bar: 7 lm.
10 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x
regulates transcription elongation (Price, 2000). Our struc-
tural analysis indicated a close resemblance between CDKC2
and CRK7, a mammalian protein involved in splicing, sug-
gesting an additional role for CDKC2. Our co-location studies
support this idea and suggest that CDKC2 regulates the
trafficking of splicing factors between an active dispersed
state and inactive speckles.
Structurally, CDKC2 is related to both CRK7 and CDK9, but
is more closely related to the former, as judged by a number
of criteria. CRK7 is a member of a subfamily of cdc2-related
kinases that share the PITAIRE motif with the human CHED-
like kinases, and so far PITAIRE kinases from Caenorhabditis
elegans, Drosophila melanoganster and Sphaerechinus
granularis (sea urchin) have been described in the databases
(Marques et al., 2000). CRK7 was reported to phosphorylate
the CTD of RNAP II, and to associate with a splicing factor of
the SR family, suggesting that it provides a link between
transcription and splicing machinery (Ko et al., 2001).
CDK9, therefore, differs from both CDKC2 and CRK7 in a
number of important features, both structural and func-
tional. Notably, the regulatory phosphorylation site, T14/
Y15, is altered in CDK9 by substitution of Tyr15 with Phe. The
phosphorylation of residues T14/Y15 is a key regulatory step
in controlling the activity of CDK-Cyc complexes, as phos-
phorylation of Y15 can maintain the CDK-Cyc complex in an
inactive form (Hunter, 1995). Thus, the substitution of Y15 by
F15 in CDK9 suggests its activity is regulated by a different
mechanism.
The subcellular localization of CDKC2 shares key features
with CRK7. Although both CRK7 and CDK9 are nuclear
proteins, they display distinct subnuclear localization pat-
terns. CRK7 decorated nuclear speckles, co-localized with
the splicing factor SC35 and was associated strongly with
the nuclear matrix, whereas nuclease treatment suggested
that the speckled localization pattern observed was not a
result of direct association with chromatin or RNA (Ko et al.,
2001). CDK9 was also found to localize in subnuclear
compartments, but co-localization with SR-splicing factors
was poor (Herrmann and Mancini, 2001). Additionally, CDK9
did not co-localize significantly with the CTD of RNAP II,
casting doubt as to whether it directly regulated the activity
of RNAP II during transcription elongation. Furthermore, the
majority of CDK9 was released from the nucleus after
treatment with detergent, whereas much of the remaining
protein was found to associate with chromatin (Herrmann
and Mancini, 2001). These differences suggested that CRK7
might modulate the activity of RNAP II and/or the splicing
machinery (Ko et al., 2001).
Our study indicated that CDKC2 protein localizes in a
similar manner to CRK7: in the nucleus as small speckles,
along with splicing factors (Ko et al., 2001). Furthermore,
CDKC2 co-localized with cyclophilin CypRS64, a protein with
a regulatory role in the plant splicing machinery (Lorkovic
et al., 2004b). Co-expression of CDKC2 with both SRp34 and
CypRS64 promoted their redistribution within the nucleo-
plasm. All these results suggest that the redistribution of
(a)
(b)
Figure 8. Localization profiles of CDKC2 and its dominant-negative mutant form, mCDKC2, transiently expressed in Arabidopsis cell cultures either alone or with
Cyp64.
(a) Arabidopsis cells were transiently transformed with constructs expressing GFP- CDKC2 and GFP-mCDKC2 fusion proteins. Time-lapse images of GFP-CDKC2 and
GFP-mCDKC2 protein localization in a cell nucleus are shown. Cells were kept on a slide during the time-lapse series without preventing moisture loss. At the start of
the experiment (0 min) both brightfield and GFP images are shown, whereas in subsequent time points only GFP images are shown. n, nucleus; no, nucleolus. Scale
bar: 5 lm.
(b) Arabidopsis cells were transiently co-transformed with constructs expressing GFP-mCDKC2 and Cyp64 fusion proteins. In the panel of nuclei, the GFP channel is
shown first, followed by the RFP channel and the merged image.
Line-intensity profiles of the green and red channels across three randomly selected lines are also shown. Scale bar: 5 lm.
CDKC2 associates with the splicing apparatus 11
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

CDKC2 upon co-expression with spliceosomal components
probably results from a functional interaction.
Kinase activity is required for the spatial organization and
dynamics of CDKC2, and of spliceosomal components
Previous studies with mammalian and plant cells have
demonstrated that changes in the transcriptional status
cause the redistribution of splicing factors within the nucleus,
and their accumulation into larger bodies (Ali et al., 2003;
Docquier et al., 2004; Grande et al., 1997; Misteli et al., 1997;
Tillemans et al., 2005). Our results indicated that CDKC2 dis-
played a similar response upon DRB treatment with the
Arabidopsis splicing components SRp34 and CypRS64.
The ability of DRB to diminish RNAP-II-directed transcrip-
tion has been attributed to the inhibition of casein kinase-II
Table 1 Co-response analysis of microarray expression data for the Arabidopsis CDKC2 gene
RankSpearmanvalue Gene Gene description
1 0.8765 At4g00830 RNA recognition motif (RRM)-containing protein
2 0.873 At5g64610 Histone acetyltransferase3 0.8622 At1g23860 Splicing factor RSZp21
4 0.8598 At1g15340 Methyl-CpG-binding domain-containing protein5 0.8534 At5g16260 RNA recognition motif (RRM)-containing protein
6 0.8471 At5g04280 Glycine-rich RNA-binding protein|
7 0.8467 At3g22990 Expressed protein plant specific family8 0.841 At1g01160 SSXT protein-related/transcription co-activator-related
9 0.8335 At1g27650 U2-snRNP auxiliary factor small subunit, putative
10 0.8272 At3g15590 DNA-binding protein, putative11 0.8231 At1g29350 Unknown protein12 0.8226 At5g16505 Mutator-like transposase family13 0.8175 At3g19630 Radical SAM domain-containing protein catalytic/iron ion binding14 0.8164 At5g51300 Splicing-factor related, similar to SF1 protein from Drosophila melanogaster
15 0.8161 At3g18380 Expressed protein16 0.8158 At5g51410 LUC7 N-terminus domain-containing protein; U1-snRNA associated protein with
a role in splice-site recognition
17 0.8133 At5g52040 Arginine/serine-rich splicing factor RSP41
18 0.8121 At1g04080 Hydroxyproline-rich glycoprotein pre-mRNA processing protein PRP39 gb L29224
from Saccharomyces cerevisiae
19 0.8099 At5g50320 Radical SAM-domain-containing protein20 0.808 At5g58130 RNA recognition motif (RRM)-containing protein
21 0.8074 At3g06480 DEAD box RNA helicase, putative, similar to RNA helicase DRH1 from Arabidopsis thaliana
22 0.8074 At1g80020 hAT-like transposase family (hobo/Ac/Tam3)23 0.8064 At3g12130 KH domain-c/K homology RNA-binding domain, type I; KH binds single-stranded RNA or DNA
24 0.8019 At1g77470 Putative replication factor C similar to replication factor C GB:P40937 from Homo sapiens25 0.8018 At2g25970 KH-domain-containing protein
26 0.7991 At1g16610 Arginine/serine-rich protein, putative (SR45); similar to arginine/serine-rich protein
GI:6601502 from (Arabidopsis thaliana)
27 0.7987 At2g38420 Pentatricopeptide (PPR)-repeat-containing protein
28 0.7932 At3g17450 hAT dimerisation domain-containing protein29 0.7911 At2g05120 Expressed protein30 0.7905 At1g47970 Expressed protein31 0.7898 At3g04610 KH-domain-containing protein, similar to putative nucleic acid binding protein
GB:CAB39665 from Arabidopsis thaliana
32 0.7897 At3g55200 Splicing factor, putative, similar to splicing factor 3B subunit 3 (SF3b130)
33 0.7891 At2g06510 Putative replication protein A134 0.789 At2g17870 Putative glycine-rich, zinc-finger DNA-binding protein35 0.7887 At4g02460 DNA mismatch repair protein, putative, similar to SP|P54278 PMS1 protein
homolog 2 from Homo sapiens36 0.7884 At4g31880 Expressed protein|37 0.7882 At3g18035 Histone H1/H5 family protein, similar to HMR1 protein (GI:4218141) from Antirrhinum majus38 0.788 At4g31430 Expressed protein39 0.7877 At3g63400 Cyclophilin CypRS64 peptidyl-prolyl cis-trans isomerase cyclophilin-type family protein
40 0.787 At1g75660 5¢–3¢ exoribonuclease (XRN3)
The expression pattern of CDKC2 was compared in silico with all other Arabidopsis genes available in Affymetrix microarray experiments usingco-response databases. The 40 genes that are most closely co-regulated with CDKC2 are presented in rank order with the Spearman correlationcoefficient. Genes with functional annotations related to mRNA processing are illustrated in bold.
12 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

(CKII) activity, as the drug competes with the ATP and GTP
molecules required for the phosphorylation of the CTD of
RNAP II by the kinase (Zandomeni et al., 1986; Zandomeni,
1989; Trembley et al., 2003). Furthermore, inhibition of CKII
activity by DRB occurs during the transcription elongation
process (Chodosh et al., 1989). DRB also targets the kinase
activity of the CAK component of TFIIH and of the catalytic
subunit of P-TEFb, both of which can phosphorylate the CTD
of RNAP II, with the latter functioning during transcription
elongation (Marshall and Price, 1995; Marshall et al., 1996;
Yankulov et al., 1996; Pinchero et al., 2004). All the above
indicate that DRB inhibits transcription by reducing protein
phosphorylation, although the target kinase is unclear
(Yamaguchi et al., 1998).
The catalytic subunit of plant P-TEFb is CDKC (Barroco
et al., 2003; Fulop et al., 2005; Cui et al., 2007), and DRB
affects the localization dynamics of the fusion protein in a
similar manner to that caused by the introduction of a
mutation that is predicted to eliminate kinase activity. Also,
roscovitine treatment of cells co-expressing CDKC2 and
SRp34 affected the spatial organization of the proteins, and
resulted in their accumulation in nucleolar and perinucleolar
regions (data not shown). Protein phosphorylation is impor-
tant in the regulation of plant SR-protein dynamics and
localization (Tillemans et al., 2005, 2006). Taken together,
these data suggest that CDKC kinase activity is required for
its normal dynamic association with spliceosomal speckles,
and for normal partitioning between the nucleoplasm and
the nucleolus.
Accumulation of nucleoplasmic proteins into or around
the nucleolus occurs under conditions of transcriptional
arrest (Malatesta et al., 2000; Shav-Tal and Zipori, 2002).
Recent data suggest that the nucleolus is sensitive to the
transcriptional profile of the cell, and that the status of
transcriptional activity is reflected in nucleolar structure
(Shav-Tal et al., 2005). Hypophosphorylated SR-splicing
factors localize transiently for 15–20 min in perinucleolar
regions during the telophase, before their accumulation into
nuclear speckles in G1 (Bubulya et al., 2004). Furthermore,
inhibition of transcription results in co-localization between
SR proteins and SR-protein kinases in the above regions,
suggesting that phosphorylation is required for the release
of splicing factors from perinucleolar regions into tran-
scription sites. This nucleolar reorganization during low
transcriptional activity can also be impersonated by drug-
induced inhibition of transcription (Zinszner et al., 1997;
Dousset et al., 2000; Andersen et al., 2002; Fox et al., 2002;
Ospina and Matera, 2002). Treatment of mammalian cells
with actinomycin-D resulted in nucleolar/perinucleolar accu-
mulation of several proteins: among them ASF/SF2 (Shav-
Tal et al., 2005), which is the mammalian homolog of SRp34,
and CDK2 (Liu et al., 2000). Furthermore, the plant splicing
factor SRp31 was concentrated in areas around the nucleo-
lus after treatment with staurosporine. All the above could
explain the nucleolar/perinucleolar profiles that we
observed for CDKC2 and spliceosomal components after
the inhibition of transcription.
In summary, our data suggest a role for the Arabidopsis
CDKC2 in spliceosomal dynamics. CDKC2 is co-regulated
with genes involved in mRNA processing, and association of
CDKC2 with spliceosomal components is dependent both on
kinase activity and on the transcriptional status of the cell. To
our knowledge, an association between a plant CDK and the
splicing machinery has not been reported previously. The
mobility of SR-splicing factors from nucleolar/perinucleolar
regions into nuclear speckles, and from there into active
transcription sites, is dependent on phosphorylation (Liu
et al., 2000; Lamond and Spector, 2003), and CDKC2 might
participate in such processes. Interestingly, SRp34 was
recently identified in the Arabidopsis nucleolus (Pendle et al.,
2005). Our data, combined with previous findings that CDKC
phosphorylates the CTD of RNAP II (Barroco et al., 2003;
Fulop et al., 2005; Cui et al., 2007), suggest novel functions
for the plant CDKC family, and an attractive scenario would be
that CDKC links transcription with splicing.z
Experimental procedures
Plasmids used, and transformation of Agrobacterium
and plant cells
The Arabidopsis CDKC2 and CDKB2;1 genes were obtained as trim-med open reading frames (ORFs) from the SSP consortium collection(http://signal.salk.edu/SSP/index.html), in the form of ‘U clones’(CDKC2-clone number 09114 and CDKB2;1-clone number 12632)and in a pUNI51 expression vector (accession AY260846). The ORFswere cloned into the Gateway�-compatible expression vectorpGWB6 (a gift from Tsuyoshi Nakagawa, Shimane University,Japan), which is a 35S::N-terminus GFP-fusion expression vector, asdescribed in Koroleva et al. (2005). The Agrobacterium strain usedwas GV3101 (Koncz and Schell, 1986), and bacterial cells weretransformed by the freeze-thaw method. Approximately 1 lg ofplasmid DNA was added to 100 ll of bacterial cells. The mixture wasfrozen in liquid nitrogen, thawed at a 37�C for 5 min and incubated at28�C for 2–4 h. Transformed cells were selected on agar plates con-taining the appropriate antibiotic. Colonies appeared followingincubation at 28�C for 2–3 days. The Arabidopsis cell suspensionused was the Arabidopsis Colombia-0 (Mathur et al., 1998). TheTobacco cell suspension was BY2 (Nagata et al., 1992). Plant cellswere transformed as described in Koroleva et al. (2004, 2005).
Generation of a dominant-negative mutant of CDKC2
Mutation at the ATP-binding pocket of CDKC2 (D182N; GA-T fi AAT) was generated using the GeneTailorR site-directedmutagenesis kit (Invitrogen, http://www.invitrogen.com) followingthe manufacturer’s instructions. Primer sequences used forthe mutagenesis were: 5¢-AGGGAAATTTAAAGCTAGCGAATT-TTGGGCTTGC-3¢ (forward; the mutagenic nucleotide is under-lined) and 5¢-CGCTAGCTTTAAATTTCCCTCATTGTCAAT-3¢(reverse). Constructs containing the mutated CDKC2 weresequenced to confirm the presence of the desired mutation and
CDKC2 associates with the splicing apparatus 13
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

the absence of any other changes, transformed into Agrobacte-rium and were then transiently expressed into Arabidopsis cells,as described above.
RNase, DNase and detergent treatments of
Arabidopsis cell suspensions
Transformed cells were centrifuged for 1 min at 208 g, and the plantpellet was resuspended in the following detergent solutions: (i) 10%(v/v) Nonidet P-40 (NP-40; Sigma, http://www.sigmaaldrich.com), (ii) 1% (v/v) Triton X-100 (ACROS, http://www.acros.com) and(iii) extraction buffer [10% (v/v) DMSO + 0.05% (v/v) NP-40 + 0.45 M
Sorbitol in PEM buffer (50 mM PIPES, 5 mM MgSO4, 5 mM EGTA)].Before DNaseI and RNaseA treatments, Arabidopsis cells wereconverted into protoplast, as described in Chan et al. (1996). Theobtained protoplasts were washed twice with PEM plus 0.025 M
Sorbitol (PEMS), and were then incubated in extraction buffer for15 min. The extracted protoplasts were washed twice with 10%DMSO in PEM and 5–50 ll of DNase (10 mg ml)1; Sigma), and100–200 ll of RNaseA (10 mg ml)1; Sigma) was added to 1 ml of theextracted protoplasts. For nuclei staining, 1 lg ml)1 DAPI and1 lg ml)1 ethidium bromide was used.
Drug treatments
CDK-kinase activity was inhibited with roscovitine (Sigma). Tran-scription was inhibited with DRB (Sigma). Arabidopsis cells weretreated with 20 lM roscovitine, 20 lM Microcystin-LR and50 lg ml)1 DRB. To remove the drugs, treated cells were washedwith ATN growth medium [4.42% (w/v) MS + GB5 (Duchefa, http://www.duchefa.com), 3% (w/v) sucrose and 0.2 mg l)1 2,4-dichloro-phenoxyacetic acid (Duchefa), pH 5.7].
Western blot analysis
Arabidopsis cells expressing the GFP-CDKC2 fusion protein werewashed twice with ATN medium to remove excess bacteria. Plantpellets were homogenized to a powder with liquid nitrogen, andwere resuspended in ·2 EB buffer [50 mM Tris–HCl, 150 mM NaCl,5 mM EDTA, 5 mM NaF, 0.1% (v/v) Triton X-100, one plant proteaseinhibitor cocktail tablet per 10 ml of buffer (Roche, http://www.roche.com)]. Thawed samples were centrifuged at 14 000 rpm for20 min and at 4�C. The supernatant was removed and the pellet wasresuspended in 5 ml of EB buffer and subjected to sonication usinga SANYO� Soniprep 150 sonicator (SANYO, http://www.sanyo.com). The sonicated plant lysates were resolved by SDS-PAGE,transferred to nitrocellulose [PROTRAN BA 85 CELLULOSENI-TRAT(E); Schleicher and Schuell; http://www.whatman.com/protrannitrocellulosemembranes.aspx]; and incubated with rabbitanti-GFP (Abcam, http://www.abcam.com) and anti-rabbit HRP(Pierce, http://www.piercenet.com) antibodies.
Microscopic techniques and imaging
Arabidopsis cells expressing green and red fluorescent proteinfusions were visualized with an epifluorescence microscope(Eclipse E600; Nikon, http://www.nikon.com) and the Cain CCDsystem (Hamamatsu Orca HQ cooled CCD digital camera;Hamamatsu, http://www.hamamatsu.com). Images were pro-cessed using PHOTOSHOP
� Software (version CS2; Adobe Systems,http://www.adobe.com) and IMAGEJ software (http://rsb.info.nih.gov/ij).
Acknowledgements
We gratefully acknowledge Andrea Barta for providing the SRp34and CypRS64 template plasmids, John Brown for providing thepROK2-mRFP plasmid and to Matt Tomlinson for his help with theGateway� cloning system. We also thank Olga Koroleva for pro-viding the RFP fusion constructs and for her critical reading of themanuscript. Georgios Kitsios was funded by a CASE PhD student-ship from the Biotechnology and Biological Sciences ResearchCouncil and Syngenta. The work was funded in part by theEU-Framework VI AGRO-OMICS program (Contract number LSHG-CT-2006-037704).
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. Co-expression of GFP-CDKB2;1 and RFP-SRp34 fusionproteins in Arabidopsis cell suspensions.Figure S2. Co-expression of GFP-CDKD and RFP-SRp34 fusionproteins in Arabidopsis cell suspensions.Figure S3. Treatment of Arabidopsis cells, expressing the GFP-CDKC2 fusion protein, with 5,6-dichloro-1-b-D-ribofuranosylbenz-imidazole (DRB) and roscovitine.This material is available as part of the online article from http://www.blackwell-synergy.com.Please note: Blackwell publishing are not responsible for thecontent or functionality of any supplementary materials suppliedby the authors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
References
Ali, G.S., Golovkin, M. and Reddy, A.S. (2003) Nuclear localizationand in vivo dynamics of a plant-specific serine/arginine-richprotein. Plant J. 36, 883–893.
Andersen, J.S., Lyon, C.E., Fox, A.H., Leung, A.K., Lam, Y.W., Steen,
H., Mann, M. and Lamond, A.I. (2002) Directed proteomic analysisof the human nucleolus. Curr. Biol. 12, 1–11.
Barroco, R.M., De Veylder, L., Magyar, Z., Engler, G., Inze, D. and
Mironov, V. (2003) Novel complexes of cyclin-dependent kinasesand a cyclin-like protein from Arabidopsis thaliana with a functionunrelated to cell division. Cell Mol. Life Sci. 60, 401–412.
Betts, M.J. and Russell, R.B. (2003) Amino acid properties andconsequences of substitutions. In Bioinformatics for Geneticists.(Barnes, M.R. and Gray, I.C., eds). Chichester, UK: John Wiley &Sons, pp. 289–316.
Bond, U. (1988) Heat shock but not other stress inducers leads to thedisruption of a subset of snRNPs and inhibition of in vitro splicingin HeLa cells. EMBO J. 7, 3509–3518.
Bubulya, P.A., Prasanth, K.V., Deerinck, T.J., Gerlich, D., Beaudouin,
J., Ellisman, M.H., Ellenberg, J. and Spector, D.L. (2004) Hypo-phosphorylated SR splicing factors transiently localize aroundactive nucleolar organizing regions in telophase daughter nuclei.J. Cell Biol. 167, 51–63.
Chan, J., Rutten, T. and Lloyd, C. (1996) Isolation of microtubule-associated proteins from carrot cytoskeletons: a 120 kDa mapdecorates all four microtubule arrays and the nucleus. Plant J. 10,251–259.
Chodosh, L.A., Firell, A., Samuels, M. and Sharp, P.A. (1989) 5,6-Dichloro-1-b-D-ribofuranosylbenzimidazole inhibits transcriptionelongation by RNA polymerase II in vitro. J. Biol. Chem. 264,2250–2257.
14 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

Cui, X., Fan, B., Scholz, J. and Chen, Z. (2007) Roles of Arabidopsiscyclin-dependent kinase C complexes in cauliflower mosaic virusinfection, plant growth and development. Plant Cell, 19, 1388–1402.
De Bondt, H.L., Rosenblatt, J., Jancarik, J., Jones, H.D., Morgan,
D.O. and Kim, S.-H. (1993) Crystal structure of cyclin-dependentkinase 2. Nature, 363, 595–602.
Dewitte, W. and Murray, J.A. (2003) The Plant Cell Cycle. Annu. Rev.Plant Physiol. Plant Mol. Biol. 54, 235–264.
Docquier, S., Tillemans, V., Deltour, R. and Motte, P. (2004) Nuclearbodies and compartmentalization of pre-mRNA splicing factors inhigher plants. Chromosoma, 112, 255–266.
Dousset, T., Wang, C., Verheggen, C., Chen, D., Hernandez-Verdun,
D. and Huang, S. (2000) Initiation of nucleolar assembly is inde-pendent of RNA polymerase I transcription. Mol. Biol. Cell, 11,2705–2717.
Fang, Y., Hearn, S. and Spector, D.L. (2004) Tissue-specific expres-sion and dynamic organization of SR splicing factors in Arabid-opsis. Mol. Biol. Cell, 15, 2664–2673.
Fobert, P.R., Gaudin, V., Lunness, P., Coen, E.S. and Doonan, J.H.
(1996) Distinct classes of cdc2-related genes are differentiallyexpressed during the cell division cycle in plants. Plant Cell, 8,1465–1476.
Fox, A.H., Lam, Y.W., Leung, A.K., Lyon, C.E., Andersen, J., Mann,
M. and Lamond, A.I. (2002) Paraspeckles: a novel nuclear domain.Curr. Biol. 12, 13–25.
Fulop, K., Pettko-Szandtner, A., Magyar, Z., Miskolczi, P., Kondo-
rosi, E., Dudits, D. and Bako, L. (2005) The Medicago CDKC;1-CYCLINT;1 kinase complex phosphorylates the carboxy-terminaldomain of RNA polymerase II and promotes transcription. Plant J.42, 810–820.
Grande, M.A., van der Kraan, I., de Jong, L. and van Driel, R. (1997)Nuclear distribution of transcription factors in relation to sites oftranscription and RNA polymerase II. J. Cell Sci. 110 (Pt 15), 1781–1791.
Hanks, S.K. (2003) Genomic analysis of the eukaryotic protein kinasesuperfamily: a perspective. Genome Biol. 4, 111.
Hanks, S.K. and Hunter, T. (1995) Protein kinases 6. The eukaryoticprotein kinase superfamily: kinase (catalytic) domain structureand classification. FASEB J. 9, 576–596.
Hardcastle, I.R., Golding, B.T. and Griffin, R.J. (2002) Designinginhibitors of cyclin-dependent kinases. Annu. Rev. Pharmacol.Toxicol. 42, 325–348.
Hartwell, L.H., Culotti, J., Pringle, J.R. and Reid, B.J. (1974) Geneticcontrol of the cell division cycle in yeast. Science, 183, 46–51.
Hastings, M.L. and Krainer, A.R. (2001) Pre-mRNA splicing in thenew millennium. Curr. Opin. Cell Biol. 13, 302–309.
Hemsley, R., McCutcheon, S., Doonan, J. and Lloyd, C. (2001) P34(cdc2) kinase is associated with cortical microtubules from higherplant protoplasts. FEBS Lett. 2001, 508.
Herrmann, C.H. and Mancini, M.A. (2001) The Cdk9 and cyclin Tsubunits of TAK/P-TEFb localize to splicing factor-rich nuclearspeckle regions. J. Cell Sci. 114, 1491–1503.
van den Heuvel, S. and Harlow, E. (1993) Distinct roles for cyclin-dependent kinases in cell cycle control. Science, 262, 2050–2054.
Hindley, J. and Phear, G.A. (1984) Sequence of the cell division geneCDC2 from Schizosaccharomyces pombe; patterns of splicingand homology to protein kinases. Gene, 31, 129–134.
Hunter, T. (1995) Protein kinases and phosphatases: the yin andyang of protein phosphorylation and signaling. Cell, 80, 225–236.
Iwakawa, H., Shinmyo, A. and Sekine, M. (2006) ArabidopsisCDKA;1, a cdc2 homologue, controls proliferation of generativecells in male gametogenesis. Plant J. 45, 819–831.
Jordan, P., Cunha, C. and Carmo-Fonseca, M. (1997) The cdk7-cyclinH-MAT1 complex associated with TFIIH is localized in coiledbodies. Mol. Biol. Cell, 8, 1207–1217.
Joubes, J., Lemaire-Chamley, M., Delmas, F., Walter, J., Hernould,
M., Mouras, A., Raymond, P. and Chevalier, C. (2001) A new C-type cyclin-dependent kinase from tomato expressed in dividingtissues does not interact with mitotic and G1 cyclins. PlantPhysiol. 126, 1403–1415.
Ko, T.K., Kelly, E. and Pines, J. (2001) CrkRS: a novel conservedCdc2-related protein kinase that colocalises with SC35 speckles.J. Cell Sci. 114, 2591–2603.
Koncz, C. and Schell, J. (1986) The promoter of TL-DNA gene 5controls the tissue-specific expression of chimaeric genes carriedby a novel type of Agrobacterium binary vector. Mol. Gen. Genet.204, 383–396.
Koroleva, O.A., Tomlinson, M., Parinyapong, P., Sakvarelidze, L.,
Leader, D., Shaw, P. and Doonan, J.H. (2004) CycD1, a putativeG1 cyclin from Antirrhinum majus, accelerates the cell cycle incultured tobacco BY-2 cells by enhancing both G1/S entry andprogression through S and G2 phases. Plant Cell, 16, 2364–2379.
Koroleva, O.A., Tomlinson, M.L., Leader, D., Shaw, P. and Doonan,
J.H. (2005) High-throughput protein localization in Arabidopsisusing Agrobacterium-mediated transient expression of GFP-ORFfusions. Plant J. 41, 162–174.
Lamond, A.I. and Spector, D.L. (2003) Nuclear speckles: a model fornuclear organelles. Nat. Rev. Mol. Cell Biol. 4, 605–612.
Lapidot-Lifson, Y., Patinkin, D., Prody, C.A., Ehrlich, G., Seidman, S.,
Ben-Aziz, R., Benseler, F., Eckstein, F., Zakut, H. and Soreq, H.
(1992) Cloning and antisense oligodeoxynucleotide inhibition of ahuman homolog of cdc2 required in hematopoiesis. Proc. NatlAcad. Sci. USA, 89, 579–583.
Lazar, G., Schaal, T., Maniatis, T. and Goodman, H.M. (1995) Iden-tification of a plant serine-arginine-rich protein similar to themammalian splicing factor SF2/ASF. Proc. Natl Acad. Sci. USA,92, 7672–7676.
Lessard, P., Bouly, J.P., Jouannic, S., Kreis, M. and Thomas, M.
(1999) Identification of cdc2cAt: a new cyclin-dependent kinaseexpressed in Arabidopsis thaliana flowers. Biochim. Biophys.Acta, 1445, 351–358.
Liu, J., Hebert, M.D., Ye, Y., Templeton, D.J., Kung, H. and
Matera, A.G. (2000) Cell cycle-dependent localization of theCDK2-cyclin E complex in Cajal (coiled) bodies. J. Cell Sci. 113
(Pt. 9), 1543–1552.Lorkovic, Z.J. and Barta, A. (2004a) Compartmentalization of the
splicing machinery in plant cell nuclei. Trends Plant Sci. 9, 565–568.
Lorkovic, Z.J., Lopato, S., Pexa, M., Lehner, R. and Barta, A. (2004b)Interactions of Arabidopsis RS domain containing cyclophilinswith SR proteins and U1 and U11 small nuclear ribonucleopro-tein-specific proteins suggest their involvement in pre-mRNASplicing. J. Biol. Chem. 279, 33890–33898.
Malatesta, M., Gazzanelli, G., Battistelli, S., Martin, T.E., Amalric,
F. and Fakan, S. (2000) Nucleoli undergo structural andmolecular modifications during hibernation. Chromosoma, 109,506–513.
Marques, F., Moreau, J.-L., Peaucellier, G., Lozano, J.-C., Schatt, P.,
Picard, A., Callebaut, I., Perret, E. and Geneviere, A.-M. (2000) ANew Subfamily of High Molecular Mass CDC2-Related Kinaseswith PITAI/VRE Motifs. Biochem. Biophys. Res. Commun. 279,832–837.
Marshall, N.F. and Price, D.F. (1995) Purification of P-TEFb, a tran-scription factor required for the transition into productive elon-gation. J. Biol. Chem. 270, 12335–12338.
CDKC2 associates with the splicing apparatus 15
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x

Marshall, N.F., Peng, J., Xie, Z. and Price, D.H. (1996) Control of RNApolymerase II elongation potential by a novel carboxyl-terminaldomain kinase. J. Biol. Chem. 271, 27176–27183.
Mathur, J., Szabados, L., Schaefer, S., Grunenberg, B., Lossow, A.,
Jonas-Straube, E., Schell, J., Koncz, C. and Koncz-Kalman, Z.
(1998) Gene identification with sequenced T-DNA tags generatedby transformation of Arabidopsis cell suspension. Plant J. 13,707–716.
Mendenhall, M.D., Richardson, H.E. and Reed, S.I. (1998) Dominantnegative protein kinase mutations that confer a G1 arrest phe-notype. Proc Natl Acad Sci USA, 85, 4426–4430.
Menges, M., de Jager, S.M., Gruissem, W. and Murray, J.A. (2005)Global analysis of the core cell cycle regulators of Arabidopsisidentifies novel genes, reveals multiple and highly specific pro-files of expression and provides a coherent model for plant cellcycle control. Plant J. 41, 546–566.
Mironov, V.V., De Veylder, L., Van Montagu, M. and Inze, D. (1999)Cyclin-dependent kinases and cell division in plants – the nexus.Plant Cell, 11, 509–522.
Misteli, T., Caceres, J.F. and Spector, D.L. (1997) The dynamics ofa pre-mRNA splicing factor in living cells. Nature, 387, 523–527.
Nagata, T., Nemoto, Y. and Hasezawa, S. (1992) Tobacco BY2 cellline as the ‘HeLa’ cell in cell biology of higher plants. Int. Rev.Cytol. 132, 1–30.
Nasmyth, K.A. and Reed, S.I. (1980) Isolation of genes by comple-mentation in yeast: molecular cloning of a cell-cycle gene. Proc.Natl Acad. Sci. USA, 77, 2119–2123.
Nowack, M.K., Grini, P.E., Jakoby, M.J., Lafos, M., Koncz, C. and
Schnittger, A. (2006) A positive signal from the fertilization of theegg cell sets off endosperm proliferation in angiospermembryogenesis. Nat. Genet. 38, 63–67.
Nurse, P. and Thuriaux, P. (1980) Regulatory genes controllingmitosis in the fission yeast Schizosaccharomyces pombe.Genetics, 96, 627–637.
Ospina, J.K. and Matera, A.G. (2002) Proteomics: the nucleolusweighs in. Curr. Biol. 12, R29–R31.
Pendle, A.F., Clark, G.P., Boon, R., Lewandowska, D., Lam, Y.W.,
Andersen, J., Mann, M., Lamond, A.I., Brown, J.W. and Shaw,
P.J. (2005) Proteomic analysis of the Arabidopsis nucleolussuggests novel nucleolar functions. Mol. Biol. Cell, 16, 260–269.
Pinchero, R., Liaw, P., Bertens, K. and Yankulov, K. (2004) Threecyclin-dependent kinases preferentially phosphorylate differentparts of the C-terminal domain of the large subunit of RNApolymerase II. Eur. J. Biochem. 271, 1004–1014.
Price, D.H. (2000) P-TEFb, a cyclin-dependent kinase controllingelongation by RNA polymerase II. Mol. Cell. Biol. 20, 2629–2634.
Proudfoot, N.J., Furger, A. and Dye, M.J. (2002) Integrating mRNAprocessing with transcription. Cell, 108, 501–512.
Reed, R. (2000) Mechanisms of fidelity in pre-mRNA splicing. Curr.Opin. Cell Biol. 12, 340–345.
Rizhsky, L., Liang, H. and Mittler, R. (2002) The combined effect ofdrought stress and heat shock on gene expression in tobacco.Plant Physiol. 130, 1143–1151.
Segers, G., Gadisseur, I., Bergounioux, C., de Almeida Engler, J.,
Jacqmard, A., Van Montagu, M. and Inze, D. (1996) The Arabid-opsis cyclin-dependent kinase gene cdc2bAt is preferentiallyexpressed during S and G2 phases of the cell cycle. Plant J. 10,601–612.
Sehgal, P.B., Darnell, J.E. and Tamm, I. (1976) The inhibition by DRB(5,6-dichloro-1-b-D-ribofuranosylbenzimidazole) of hnRNA andmRNA production in HeLa cells. Cell, 9, 473–480.
Shav-Tal, Y. and Zipori, D. (2002) PSF and p54(nrb)/NonO – multi-functional nuclear proteins. FEBS Lett. 531, 109–114.
Shav-Tal, Y., Blechman, J., Darzacq, X., Montagna, C., Dye, B.T.,
Patton, J.G., Singer, R.H. and Zipori, D. (2005) Dynamic sorting ofnuclear components into distinct nucleolar caps during tran-scriptional inhibition. Mol. Biol. Cell, 16, 2395–2413.
Shimotohno, A., Matsubayashi, S., Yamaguchi, M., Uchimiya, H.
and Umeda, M. (2003) Differential phosphorylation activities ofCDK-activating kinases in Arabidopsis thaliana. FEBS Lett. 534,69–74.
Shimotohno, A., Umeda-Hara, C., Bisova, K., Uchimiya, H. and
Umeda, M. (2004) The plant-specific kinase CDKF;1 is involved inactivating phosphorylation of cyclin-dependent kinase-activatingkinases in Arabidopsis. Plant Cell, 16, 2954–2966.
Shimotohno, A., Ohno, R., Bisova, K., Sakaguchi, N., Huang, J.,
Koncz, C., Uchimiya, H. and Umeda, M. (2006) Diverse phos-phoregulatory mechanisms controlling cyclin-dependent kinase-activating kinases in Arabidopsis. Plant J. 47, 701–710.
Spector, D.L., Schrier, W.H. and Busch, H. (1983) Immunoelectronmicroscopic organization of snRNPs. Biol. Cell, 49, 1–10.
Tillemans, V., Dispa, L., Remacle, C., Collinge, M. and Motte, P.
(2005) Functional distribution and dynamics of Arabidopsis SRsplicing factors in living plant cells. Plant J. 41, 567–582.
Tillemans, V., Leponce, I., Rausin, G., Dispa, L. and Mottea, P. (2006)Insights into nuclear organization in plants as revealed by thedynamic distribution of Arabidopsis SR splicing factors. PlantCell, 18, 3218–3234.
Trembley, J.H., Hu, D., Slaughter, C.A., Lahti, J.M. and Kidd, V.J.
(2003) Casein kinase 2 interacts with cyclin-dependent kinase 11(CDK11) in vivo and phosphorylates both the RNA polymerase IIcarboxyl-terminal domain and CDK11 in vitro. J. Biol. Chem. 278,2265–2270.
Vandepoele, K., Raes, J., De Veylder, L., Rouze, P., Rombauts, S.
and Inze, D. (2002) Genome-wide analysis of core cell cycle genesin Arabidopsis. Plant Cell, 14, 903–916.
Weingartner, M., Binarova, P., Drykova, D., Schweighofer, A.,
David, J.P., Heberle-Bors, E., Doonan, J. and Bogre, L. (2001)Dynamic recruitment of Cdc2 to specific microtubule structuresduring mitosis. Plant Cell, 13, 1929–1943.
Yamaguchi, Y., Tadashi, W. and Handa, H. (1998) Interplay betweenpositive and negative elongation factors: drawing a new view ofDRB. Genes Cells, 3, 9–15.
Yankulov, K.Y., Pandes, M., McCracken, S., Bouchard, D. and
Bentley, D.L. (1996) TFIIH functions in regulating transcriptionalelongation by RNA polymerase II in Xenopus oocytes. Mol. Cell.Biol. 16, 3291–3299.
Zandomeni, R.O. (1989) Kinetics of inhibition by 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole on calf thymus casein kinase II.Biochem. J. 262, 469–473.
Zandomeni, R., Zandomeni, M.C., Shugar, D. and Weinmann, R.
(1986) Casein kinase type II is involved in the inhibition by 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole of specific RNApolymerase II transcription. J. Biol. Chem. 261, 3414–3419.
Zinszner, H., Immanuel, D., Yin, Y., Liang, F.X. and Ron, D. (1997) Atopogenic role for the oncogenic N-terminus of TLS: nucleolarlocalization when transcription is inhibited. Oncogene, 14, 451–461.
16 Georgios Kitsios et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), doi: 10.1111/j.1365-313X.2008.03414.x













![Elevated Cyclins and Cyclin-dependent Kinase Activity in ...[CANCER RESEARCH 58, 2042-2049, May I, 1998] Elevated Cyclins and Cyclin-dependent Kinase Activity in the Rhabdomyosarcoma](https://img.pdfslide.us/doc/110x75/5e4e63ca3358114ff2317f00/elevated-cyclins-and-cyclin-dependent-kinase-activity-in-cancer-research-58.jpg)















