Embed Size (px)
Citation preview

0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 269, No. 2, Issue of January 14, pp. 1375-1379, 1994
Printed in U.S.A.
Two Lectins from the Marine Sponge Halichondria okadai AN N-ACETYL-SUGAR-SPECIFIC LECTIN (HOL-I) AND AN N-ACE’IYLLACTOSAMINE-SPECIFIC LECTIN (HOL-II)*
(Received for publication, July 21, 1993, and in revised form, September 9, 1993)
Hirokazu Kawagishi+, Masamichi Yamawaki, Satoko Isobe, Taichi Usui, Atsuo KimuraO, and Seiya ChibaO From the Department of Applied Biological Chemistry, Faculty of Agriculture, Shizuoka University, 836 Ohya, Shizuoka 422, Japan and the §Department of Applied Bioscience, Faculty of Agriculture, Hokkaido University, Sapporo 060, Japan
Two lectins (HOL-I and HOL-11) were isolated from the marine sponge Halichondria okadai by affinity chroma- tography on a bovine submaxillary mucin (BSM)-Toyo- pearl and an acid-treated Sepharose 4B columns, respec- tively. In hemagglutination inhibition assays, GlcNAc, GalNAc, and their methyl glycosides were the most po- tent inhibitors among the monosaccharides tested against the HOL-I-mediated hemagglutination, suggest- ing that HOL-I can especially recognize the N-acetyl groups of the sugars. This N-acetyl specificity was sup- ported by ‘H NMR analyses; the highest field-shifts of the signal of the N-acetyl group among all the signals in Me PGlcNAc were observed in ‘H NMR spectra of mix- tures of HOL-I and the sugar. Among the oligosacchari- des tested, GlcNAcpl+4(GlcNAc~l+2)Manc~1-O(CH~)~ CHs was the most potent inhibitor, and the inhibitory potency of the oligosaccharide was z4 times greater than those of GlcNAc and GalNAc. On the other hand, N-acetyllactosamine (Galpl-rlGlcNAc) and its analogs were the strongest inhibitors toward HOL-11-induced hemagglutination. The agglutination was completely inert to Galpl+3GlcNAc, GalPl-rGGlcNAc, Galpl- SGalNAc, GalPl+lGalNAc, and Galpl+GGalNAc. Fur- thermore, HOL-I1 exhibited no binding ability to BSM, asialo-BSM, fetuin, asialofetuin, a,-acid glycoprotein, and human transferrin. These results indicate that HOL- I1 strictly recognizes simple Galpl-r4GlcNAc unit.
Many lectins have been isolated from various sources. How- ever, information is relatively sparse concerning lectins from marine origins as compared with the large variety of purified lectins of the other sources. It is well known that marine or- ganisms produce unique low molecular compounds possessing unique structures and biological activities. It is also possible that novel lectins having unique properties will be found from the organisms.
Halichondria okadai is a toxic marine sponge, and potent toxins, okaidaic acid (11, and halichondrins (2, 3) were isolated from this sponge. However, there is no report about high mo- lecular substances from the sponge. Herein the isolation and characterization of two unique lectins from the sponge are de- scribed.
* This work was supported by a Grant-in-Aid for Scientific Research on Priority Areas “Dynamic Aspects of Natural Product Chemistry” from the Ministry of Education, Science and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of
ment” in accordance with 18 U.S.C. Section 1734 solely to indicate this page charges. This article must therefore be hereby marked “aduertise-
fact. $. To whom correspondence should be addressed.
EXPERIMENTAL PROCEDURES Materials-H. okadai was collected at Aburatsubo, Kanagawa, Ja-
pan, frozen upon collection, and stored at -20 “C. Sepharose 4B, Supe- rose 12 HR10/30, and ProRPC HR5/10 columns were products of Phar- macia (Sweden). Amino-Toyopearl and Toyopearl 55F were purchased from Tosoh (Japan). SpectraPor 1 was a product of Spectrum Medical Industries. All glycoproteins and lactulose for the hemagglutinating inhibition tests were products of Sigma. Galpl+3GlcNAc, Galpl-4GlcNAc, Galpl-GGlcNAc, Galpl+3GalNAc, Galplk 4GalNAc, Galpl-GGalNAc, Galpl-4GlcNAc-PMe, Galpl+4GlcNAc- pPNP, Galpl-6GlcNAc-PPNP, GalplhGGal-PPNP, and Galpl-, 4GlcNA~(pl-4GlcNAc)~ were synthesized by enzymatic methods (4-6). The other synthetic oligosaccharides, Galpl-.4GlcNAcpl- 2Man, GlcNAcpl-2Mana1-6Man~l-O(CH2)8CO&H3, GlcNAcpl-, 2Mana1-3Manp1-O(CH2),C02CH3, GlcNAc~l-6(GlcNAc~l- 2)Manal-O(CHZ)&H3, GlcNAcp1+6(GlcNAcp1~2)Mana1-3Manpl- O(CH2),C02CH3, GlcNAcpl-4(GlcNAcpl-2)Manal-O(CH2)2CH3, GlcNAcp1-4(GlcNAcp1-2)Mana1+3Man~1-O(CH2)8CO2CH3, Glc- NAc~1+2Manal-6(GlcNAc~1-2Manal-3~Man~1-O~CH2~,CO,CH3 were kindly provided by Towa Chemical Industry Co., Ltd. (Japan).
All the other sugars for the tests were obtained from Nacalai Tesque (Japan). All the other chemicals were reagent grade.
Preparation of Afinity Adsorbent-Acid-treated Sepharose 4B was prepared by the method of Errson et al. (7). BSM’ was conjugated to Amino-Toyopearl by following the instructions of the manufacturer.
Isolation of HOLs-All the procedures were carried out at 4 “C. The frozen sponges (42.0 g) were defrosted at room temperature, immedi- ately homogenized with saline (100 ml) in a blender, and extracted with stirring overnight. The resulting suspension was filtered with gauze and the filtrate was centrifuged (10,000 x g , 15 min) to remove insoluble residues. The supernatant was directly applied to a BSM-Toyopearl column (4 1.5 x 8.5 em) linked to an acid-treated Sepharose 4B (7) column (4 1.5 x 8.5 cm) equilibrated with PBS. After extensive washing, the two columns were disconnected. The fraction containing HOL-I was desorbed with 0.5 N acetic acid from the former column. After dialysis against PBS and concentration by ultrafiltration with PM-IO, the elu- ates were further applied to a Toyopearl 55F column (+ 1.6 x 80 cm). Purified HOL-I was obtained by dialysis against distilled water and then liophilization (1.86 mg). Purified HOL-I1 was obtained by elution with distilled water from the acid-treated Sepharose 4B affinity column and dialysis against distilled water followed by liophilization (1.98 mg).
Hemagglutination Test-Erythrocytes and Pronase-treated erythro- cytes for agglutinating test were prepared as reported previously (8). Agglutination of erythrocytes and inhibition of the agglutination by sugars and glycoproteins were done in microtiter U-plates. The titer was defined as reciprocal of the end-point dilution causing hemagglu- tination. Inhibition was expressed as the minimum concentration of each sugar or glycoprotein required for inhibition of hemagglutination of titer 4 of the lectin.
SDS-Polyacrylamide Gel Electrophoresis-SDS-polyacrylamide gel electrophoresis was carried out by the method of Laemmli (9). Samples were heated in the presence or absence of 2-mercaptoethanol for 10 min at 100 “C. Gels were stained with Coomassie Brilliant Blue. The mo- lecular weight standards (Pharmacia, Sweden) used were phosphoryl-
’ The abbreviations used are: BSM, bovine submaxillary mucin; PBS, phosphate-buffered saline. All the sugars used are of the D configuration unless otherwise stated.
1375
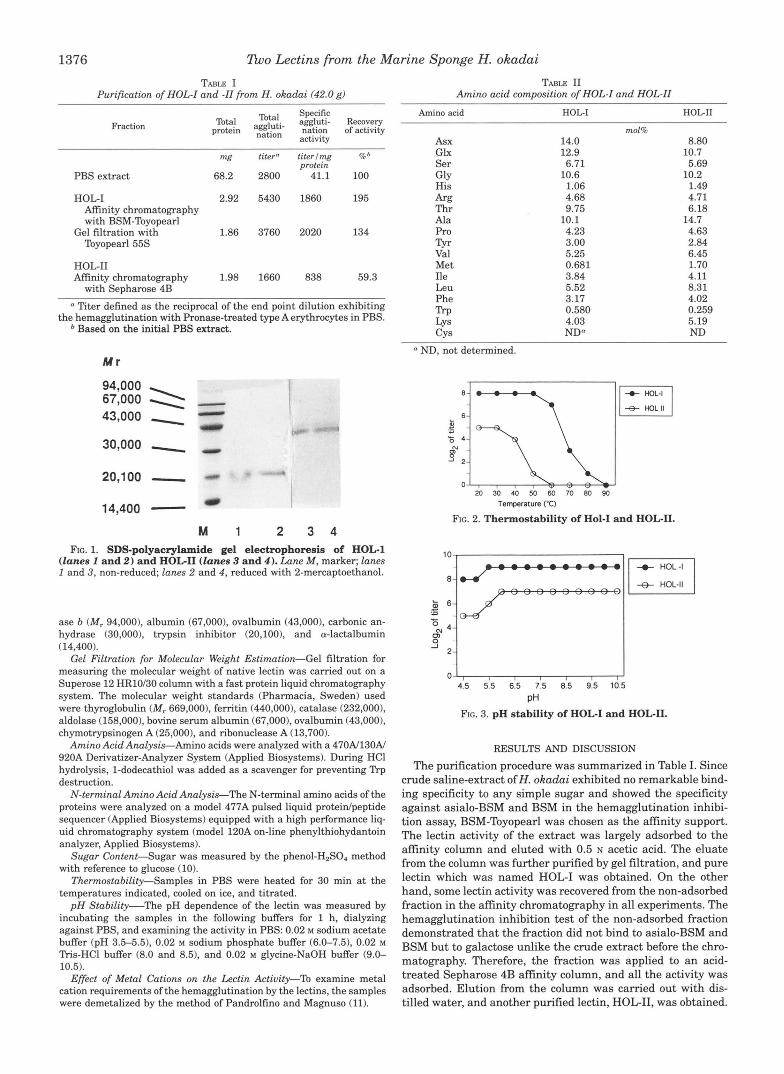
1376 Zbo Lat ins from the Marine Sponge H. okadai
Purification of HOL-I and -11 from H. okadai (42.0 g) TABLE I TABLE I1
Amino acid comDosition of HOL-I and HOL-II
Fraction Total Specific
protein ::$:- nation of activity aggluti- Recovery
activity
Total
mg titer" titerlrng %b
PBS extract 68.2 2800 41.1 100 protein
HOL-I 2.92 5430 1860 195 Affinity chromatography with BSM-Toyopearl
Toyopearl 55s Gel filtration with 1.86 3760 2020 134
HOLII Affinity chromatography 1.98 1660 838 59.3
with Sepharose 4B
a Titer defined as the reciprocal of the end point dilution exhibiting the hemagglutination with Pronase-treated type A erythrocytes in PBS.
Based on the initial PBS extract.
M r
94,000 67,000 a 43,000 - 30,000 - 20,100 - 14,400 -
M 1 2 3 4 FG. 1. SDS-polyacrylamide gel electrophoresis of HOC1
(lanes 1 and 2) and HOG11 (lanes 3 and 4). Lune M, marker; lanes 1 and 3, non-reduced; lanes 2 and 4, reduced with 2-mercaptoethanol.
ase b (M, 94,000). albumin (67,000). ovalbumin (43,000). carbonic an- hydrase (30,000), trypsin inhibitor (20,100), and a-lactalbumin (14,400).
Gel Filtration for Molecular Weight Estimation-Gel filtration for measuring the molecular weight of native lectin was carried out on a Superose 12 HR10/30 column with a fast protein liquid chromatography system. The molecular weight standards (Pharmacia, Sweden) used were thyroglobulin (M, 669,000), ferritin (440,000), catalase (232,000). aldolase (158,000), bovine serum albumin (67,000). ovalbumin (43,000). chymotrypsinogen A (25,000), and ribonuclease A (13,700).
Amino Acid Analysis-Amino acids were analyzed with a 470N130N 920A Derivatizer-Analyzer System (Applied Biosystems). During HCI hydrolysis, 1-dodecathiol was added as a scavenger for preventing Trp destruction.
N-terminal Amino Acid Analysis-The N-terminal amino acids of the proteins were analyzed on a model 477A pulsed liquid proteidpeptide sequencer (Applied Biosystems) equipped with a high performance liq- uid chromatography system (model 120A on-line phenylthiohydantoin analyzer, Applied Biosystems).
Sugar Content-Sugar was measured by the phenol-H2S04 method with reference to glucose (10).
Thermostability-Samples in PBS were heated for 30 min a t the temperatures indicated, cooled on ice, and titrated.
pH Stability-The pH dependence of the lectin was measured by incubating the samples in the following buffers for 1 h, dialyzing against PBS, and examining the activity in PBS: 0.02 M sodium acetate buffer (pH 3.5-5.5), 0.02 M sodium phosphate buffer (6.0-7.5), 0.02 M Tris-HC1 buffer (8.0 and 8 3 , and 0.02 M glycine-NaOH buffer (9.0- 10.5).
Efect of Metal Cations on the Lectin ActiuitpTo examine metal cation requirements of the hemagglutination by the lectins, the samples were demetalized by the method of Pandrolfino and Magnuso (11).
Amino acid HOG1 HOLII
A S X Glx Ser G ~ Y His Arg Thr Ala Pro Tyr Val Met Ile Leu Phe np LYS CYS
14.0 12.9
10.6 6.71
1.06 4.68 9.75
4.23 3.00 5.25 0.681 3.84 5.52 3.17 0.580
NDa 4.03
10.1
mol% 8.80
5.69
1.49 4.71 6.18
4.63 2.84 6.45 1.70 4.11 8.31 4.02 0.259 5.19 ND
10.7
10.2
14.7
ND, not determined.
E l t HOL-I
" HOLII
2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 Temperature ("C)
FIG. 2. Thermostability of Hol-I and HOCII.
I -0- HOL-I
HOL-II
4.5 5.5 6.5 7.5 8.5 9.5 10.5
pH FIG. 3. pH stability of HOC1 and HOL-11.
RESULTS AND DISCUSSION The purification procedure was summarized in Table I. Since
crude saline-extract ofH. okudui exhibited no remarkable bind- ing specificity to any simple sugar and showed the specificity against asialo-BSM and BSM in the hemagglutination inhibi- tion assay, BSM-Toyopearl was chosen as the affinity support. The lectin activity of the extract was largely adsorbed to the affinity column and eluted with 0.5 N acetic acid. The eluate from the column was further purified by gel filtration, and pure lectin which was named HOL-I was obtained. On the other hand, some lectin activity was recovered from the non-adsorbed fraction in the affinity chromatography in all experiments. The hemagglutination inhibition test of the non-adsorbed fraction demonstrated that the fraction did not bind to asialo-BSM and BSM but to galactose unlike the crude extract before the chro- matography. Therefore, the fraction was applied to an acid- treated Sepharose 4B affinity column, and all the activity was adsorbed. Elution from the column was carried out with dis- tilled water, and another purified lectin, HOL-11, was obtained.

lloo Lectins from the Marine Sponge H. okadai 1377
TABLE 111 Inhibition of hemagglutination activity of HOL-I and HOL-II
Minimum inhibitory conc.
HOG1 HOLII Inhibitor"
1 GlcNAc 2 GaWAc 3 Gal 4 GalN 5 Me aGlcNAc 6 Me pGlcNAc 7 Me aGalNAc 8 Me PGalNAc 9 NeuAc 10 11 12 13 14 15 16 17 16
20 19
21 22
24 23
26 26
27
28
29
30
31 32 33
NeuGc Galp 1+4Glc (lactose) Galplh4GlcNAc (LacNAc) Galpl+4GlcNAc-~Me (Me p-LacNAc) Galp1+4GlcNAc-pPNPe (PNP p-LacNAc) Galp1+4GlcNAcpl+2Man Gal p 1+4GlcNAc ( p 1-4GlcNA~)~ GalpljBGlcNAc Galpl+6GlcNAc Galpl4GlcNAc-pPNP Galpl-3GalNAc Galplh4GalNAc Ga lp l4Ga lNAc Galpl-6Gal-pPNP GlcNAcpl+2Manal-.6Man~1-O(CH~)~COzCH~ GlcNAcpl-2Mana1+3Manpl-O(CHz)&O&H3 GlcNAcP1-6
GlcNAc p 1- 2 GlcNAcP14
GlcNAcplh2 GlcNAc/31+4
GlcNAcP1-2 GlcNAcP 1-4
GlcNAcpl-2 GlcNAcpl+2Manal+G
GlcNAcpl-2Manal-3
Manal-O(CHz)zCH3
Mana1-3Manpl-O(CHz)8COzCH3
Manal-O(CHZ)&H3
Mana1-3Manp1-O(CHz)8COzCH3
Manp1-O(CHz)8COzCH3
N-AC-Glu N-AC-Gly N-Ac-Leu
0.098
NIc 0.098
NIc 0.098 0.049 0.049 0.098
12.5 12.5 NIc 0.39 NDd ND 0.39 ND ND NIf ND ND ND ND ND 0.012 0.049
0.024
0.20
0.006
0.024
0.012
6.25 6.25
12.5
mM NIc
50.0 25.0 25.0
NIc NIc NIc NIc NIf NIf 0.39 0.049 0.049 0.049 0.049 NIf NIf NIf NIf NIf NIf NIf NIf ND ND
NIg
ND
NIg
ND
ND
NIc NIc NIc
34 BSMh 0.98 NI' 35 Asialo-BSMh 0.12 NI' 36 Fetuinh 7.81 NI' 37 Asialofetuinh 3.91 NIL
a DL-Arabinose, DL-fucose, ribose, 2-deoxyribose, fructose, glucose, glucosamine, mannosamine, mannose, and xylose exhibited no inhibition a t concentrations up to 0.2 M. Human transfemn and aI acid glycoprotein exhibited no inhibition at concentrations up to 1 mglml.
Minimum concentrations required for inhibition of 4 hemagglutinating doses of the lectins. NI, not inhibited at concentrations up to 0.2 M. ND, not determined.
fNI, not inhibited at concentrations up to 25 m ~ . e PNP; p-nitrophenyl.
g NI, not inhibited at concentrations up to 2 mM.
' NI, not inhibited at concentrations up to 1 mg/ml. Values are pglml.
Each purified lectin was appeared as a single band in SDS- polyacrylamide gel electrophoresis (Fig. 1). HOL-I showed a single band corresponding to a molecular weight of M , = 21,000 regardless of the presence or absence of 2-mercaptoethanol. But without 2-mercaptoethanol the band was somewhat dif- fuse. The reason is not clear but the subunit might have some intramolecular S-S linkage(s1. The native molecular weight of the lectin was 84,000 on the basis of the result of gel filtration in PBS, suggesting that HOL-I consists of four identical sub- units ofM, = 21,000. On the other hand, HOL-I1 showed a band of molecular weight 42,000 in SDS-polyacrylamide gel electro- phoresis, and the estimation of the molecular weight of native
HOL-I1 by gel filtration gave the same molecular weight. This result indicates that HOL-I1 exists as a monomeric molecule in its native state.
A comparison of the amino acid compositions of HOL-I and HOL-I1 (Table 11) indicated that the two lectin forms are simi- lar but differed mainly in their amount of Asx, Thr, Met, Leu, and Trp. Although 2 nmol of each protein was applied to the sequencer, N-terminal amino acid could not be detected. This result suggests that N termini of the proteins may be blocked.
In isoelectric focusing of the lectins, both lectins indicated families of bands in pH zone near 4.5 (data not shown). The carbohydrate contents of HOL-I and HOL-I1 amounted 2.0 and
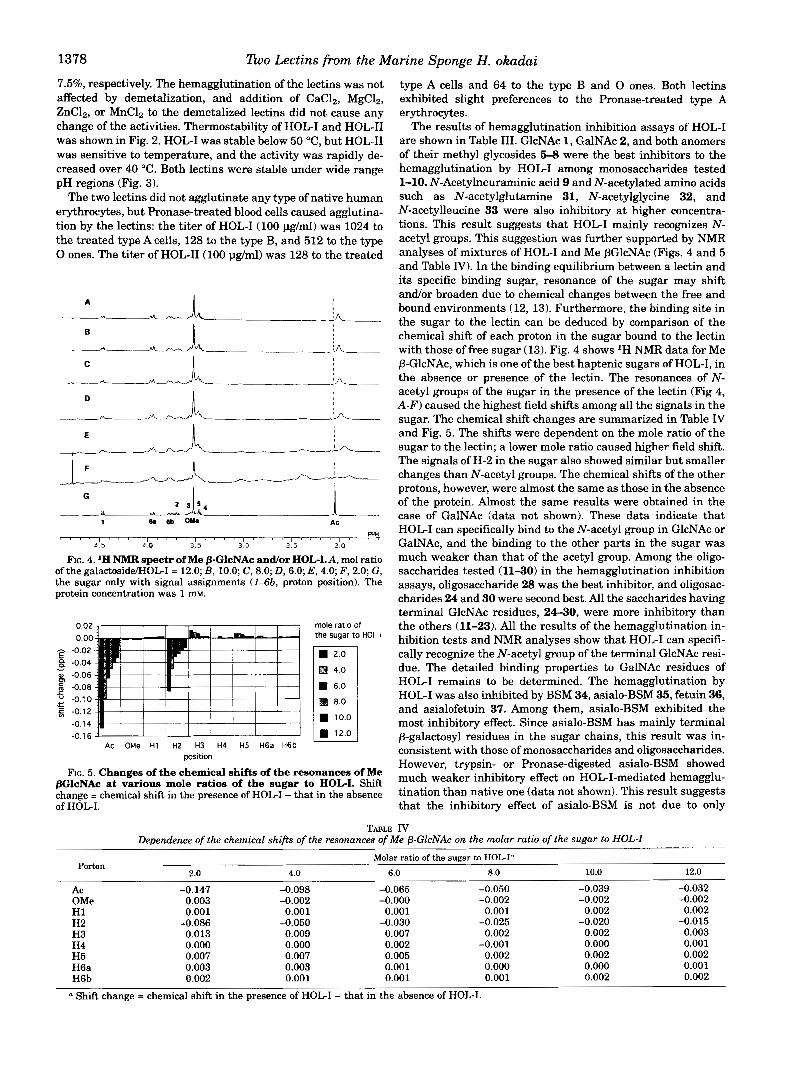
1378 Tho Lectins from the Marine Sponge H. okadai
7.5%, respectively. The hemagglutination of the lectins was not affected by demetalization, and addition of CaCl,, MgCl,, ZnC12, or MnC1, to the demetalized lectins did not cause any change of the activities. Thermostability of HOL-I and HOL-I1 was shown in Fig. 2. HOL-I was stable below 50 "C, but HOL-I1 was sensitive to temperature, and the activity was rapidly de- creased over 40 "C. Both lectins were stable under wide range pH regions (Fig. 3).
The two lectins did not agglutinate any type of native human erythrocytes, but Pronase-treated blood cells caused agglutina- tion by the lectins: the titer of HOL-I (100 pg/ml) was 1024 to the treated type A cells, 128 to the type B, and 512 to the type 0 ones. The titer of HOL-II(100 pg/ml) was 128 to the treated
1 " ~ ' " ' ~ , l C , I , - I ' ~ ' ' l ~ , , ~ Rn
4 5 4 0 3 5 3 0 2.5 2 0
FIG. 4. 'H NMR spectr of Me PGlcNAc andor HOL-LA, mol ratio of the galactosidd-IOL-I = 12.0; B , 10.0; C, 8.0; D, 6.0; E , 4.0; F , 2.0; G , the sugar only with signal assignments (143b, proton position). The protein concentration was 1 m.
0.02 mole ratio of 0.00 the sugar to HOL-I - -0.02 E 8 -0.04
o -0.06
- - : -0.08 c " -0.70 !E f -0.12
-0.1 4
-0.16' ' ' ' ' ' ' ' ' ' AC OMe H1 HZ H3 H4 H5 H6a H6b
position
FIG. 5. Changes of the chemical shifts of the resonances of Me PGlcNAc at various mole ratios of the sugar to HOL-I. Shift change = chemical shift in the presence of HOL-I - that in the absence of HOL-I.
type A cells and 64 to the type B and 0 ones. Both lectins exhibited slight preferences to the Pronase-treated type A erythrocytes.
The results of hemagglutination inhibition assays of HOL-I are shown in Table 111. GlcNAc 1, GalNAc 2, and both anomers of their methyl glycosides 5-8 were the best inhibitors to the hemagglutination by HOL-I among monosaccharides tested 1-10. N-Acetylneuraminic acid 9 and N-acetylated amino acids such as N-acetylglutamine 31, N-acetylglycine 32, and N-acetylleucine 33 were also inhibitory at higher concentra- tions. This result suggests that HOL-I mainly recognizes N- acetyl groups. This suggestion was further supported by NMR analyses of mixtures of HOL-I and Me PGlcNAc (Figs. 4 and 5 and Table IV). In the binding equilibrium between a lectin and its specific binding sugar, resonance of the sugar may shift andlor broaden due to chemical changes between the free and bound environments (12, 13). Furthermore, the binding site in the sugar to the lectin can be deduced by comparison of the chemical shift of each proton in the sugar bound to the lectin with those of free sugar (13). Fig. 4 shows 'H NMR data for Me P-GlcNAc, which is one of the best haptenic sugars of HOL-I, in the absence or presence of the lectin. The resonances of N- acetyl groups of the sugar in the presence of the lectin (Fig 4, A-F) caused the highest field shifts among all the signals in the sugar. The chemical shift changes are summarized in Table nT and Fig. 5. The shifts were dependent on the mole ratio of the sugar to the lectin; a lower mole ratio caused higher field shift. The signals of H-2 in the sugar also showed similar but smaller changes than N-acetyl groups. The chemical shifts of the other protons, however, were almost the same as those in the absence of the protein. Almost the same results were obtained in the case of GalNAc (data not shown). These data indicate that HOL-I can specifically bind to the N-acetyl group in GlcNAc or GalNAc, and the binding to the other parts in the sugar was much weaker than that of the acetyl group. Among the oligo- saccharides tested (1130) in the hemagglutination inhibition assays, oligosaccharide 28 was the best inhibitor, and oligosac- charides 24 and 30 were second best. All the saccharides having terminal GlcNAc residues, 24-30, were more inhibitory than the others (11-23). All the results of the hemagglutination in- hibition tests and NMR analyses show that HOL-I can specifi- cally recognize the N-acetyl group of the terminal GlcNAc resi- due. The detailed binding properties to GalNAc residues of HOL-I remains to be determined. The hemagglutination by HOL-I was also inhibited by BSM 34, asialo-BSM 35, fetuin 36, and asialofetuin 37. Among them, asialo-BSM exhibited the most inhibitory effect. Since asialo-BSM has mainly terminal P-galactosyl residues in the sugar chains, this result was in- consistent with those of monosaccharides and oligosaccharides. However, trypsin- or Pronase-digested asialo-BSM showed much weaker inhibitory effect on HOL-I-mediated hemagglu- tination than native one (data not shown). This result suggests that the inhibitory effect of asialo-BSM is not due to only
TABLE IV Dependence of the chemical shifts of the resonances of Me p-GlcNAc on the molar ratio of the sugar to HOL-I
Porton Molar ratio of the sugar to HOGI"
2.0 4.0 6.0 8.0 10.0 12.0
Ac OMe
-0.147 -0.098 -0.065 -0.050 -0.002 -0.000
H1 0.001 0.001 0.001 -0.002 0.001
H2 -0.086 -0.050 H3 0.013 0.007 0.002
-0.030 0.009
-0.025
H4 0.000 H5
0.000 0.007 0.007
0.002 -0.001 0.005 0.002
H6a 0.003 0.003 0.001 0.000 H6b 0.002 0.001 0.001 0.001
0.003
a Shift change = chemical shift in the presence of HOL-I - that in the absence of HOL-I.
-0.039 -0.002 0.002
-0.020 0.002
0.002 0.000
0.000 0.002
-0.032 -0.002 0.002
-0.015 0.003 0.001 0.002 0.001 0.002

?too Lectins from the Marine Sponge H . okadai 1379
simple interaction between the lectin and the sugar chains of the glycoprotein.
Binding reactivity of HOL-I1 is limited to PGal(l+l)GlcNAc (LacNAc) 12 and its analogs 13-15 (Table 111). Interestingly, Gal~1+4GlcNAc(~1+4GlcNAc)P 16 was inert to the lectin at 25 mM. Two regioisomers of LacNAc, Galplh3GlcNAc and GalplhGGlcNAc, did not affect HOL-11-mediated hemaggluti- nation at all even at 25 mM. Similar disaccharides having Gal- NAc as subterminal residues, pGal(1+3)GalNAc, pGal(l-.4) GalNAc, and pGal(l+G)GalNAc, were also inactive to the hemagglutination. These results show the binding specificity of the lectin is strictly LacNAc unit only. In general, G a l p l b 4GlcNAc-specific lectins can bind to Galpl+SGlcNAc at higher concentrations (14), and Gal~l+3GlcNAc-specific lectins can also recognize Galpl+4GlcNAc (15). To our knowledge, this is the first lectin which exhibits such strict specificity to Galplh4GlcNAc linkage. In addition, the hemagglutination caused by HOL-I1 was not inhibited by BSM, asialo-BSM, fe- tuin, asialofetuin, al-acid glycoprotein, and human transferrin even a t 1 mg/ml. Hemagglutination by known lectins is usually inhibited by at least one of the glycoproteins used in this study. Similar lectins (PFA) have been isolated from larvae of the moth Phalera flauescens (16). Glycoproteins such as fetuin, asi- alofetuin, human transferrin, ovalbumin, and porcine stomach mucin showed no inhibitory activity to the hemagglutination by PFA, and LacNAc was the best inhibitor like HOL-11.
Some lectins have been isolated from sponge (17-20). A lectin was obtained from Halichondria panicea which is the same genus as H. okadai. The lectin also recognizes p-galactosides like HOL-11. But the lectin activity was dependent on metal cation whereas HOL-I1 did not need the cation for the activity.
All the data mentioned above allowed us to conclude that
HOL-I and HOL-I1 can be used as valuable tools for sugar chain research.
Acknowledgments-We are grateful to Prof. D. Uemura, Shizuoka University (Japan) for supplying the sponges and Towa Chemical In- dustry Co., Ltd. (Japan) for supplying synthetic oligosaccharides. We are also indebted to Biopolymer Analysis Laboratory, Faculty of Agn- culture, Hokkaido University (Japan) for the use of the protein se- quencer and the system.
REFERENCES 1. Tachibana, K, Scheuer, P. J., Tsukitani, Y., Kikuchi, H., Engen, D. V., Gopi-
2. Uemura, D., Takahashi, K., Yamamoto, T., Katayama, C., Tanaka, J.. Oku-
3. Hirata, Y., and Uemura, D. (1986) Pure Appl . Chem. 58, 701-710 4. Hedbys, L., Johansson, E., Mosbach, K. , Larsson, P.-O., Gunnarsson, A.,
5. Sakai, K., Katsumi, K., Ohi, H., Usui, T., and Ishido, Y. (1992) J. Carbohydx
6. Usui, T., Kubota, S., and Ohi, H. (1993 Carbohydr: Res. 244,315-323 7. Errson, B., Asberg, K., and Porath, J. (1973) Biochim. Biophys. Acta 310,
8. Kawagishi, H., Nomura, A., Mizuno, T., Kimura, A., and Chiba, S. (1990)
9. Laemmli, U. K. (1970) Nature 227,680-685
chand, Y., and Schmits, E J. (1981) J. Am. Chem. Soc. 103,2469-2471
mura, Y., and Hirata, Y. (1985) J. Am. Chem. Soc. 197,47964798
Svensson, S., and Liinn, H. (1989) Glycoconjugate J. 6,161-168
Chem. 11,553-565
446-452
Biochim. Biophys. Acta 1034, 247-252
10. Duhis, M., Gilles, K. A,, Hammilton, J. K., P. A,, R., and Smith, F. (1956)Aml.
11. Pandolfino, E. R., and Magnuson, J. A. (1980) J. Biol. CHem. 225, 87M73 12. Kronis, K. A,, and Carver, J. P. (1982) Biochemistry 21, 305CL3057 13. Kawagishi, H., and Mori, H. (1991) Biochim. Biophys. Acta 1076,179-186 14. Sparrow, C. P., Leffers, H., and Barondes, S. H. (1987) J. Biol. Chem. 262,
15. Lotan, R., Skutelsky, E., Danan, D., and Sharon, N. (1975) J. Biol. Chem. 250,
16. Umetsu, K., Yamashita, K, Suzuki, J., Yamashita, T., and Suzuki, T. (1993)
17. Bretting, H., and Kabat, E. V. (1976) Biochemistry 15,322%3236 18. Vaith, P., Uhlenbruck, C . , Miiller, W. E. G., and Holz, G. (1979) Deu. Comp.
19. Kamiya, H., Muramoto, K., and Goto, R. (1990) Bull. Jpn. Soc. Sci. Fish. 56,
20. Kamiya, H., Muramoto, K., Hoshino, T., Yamazaki, M., and Raj, U. (1986) Bull.
Chem. 280,35&356
7383-7390
8518-8523
Arch. Bioehem. Biophys. 301,20C~205
Immunol. 3,399416
1159-1159
Jpn. Soc. Sci. Fish. 52,2205-2206





























