Embed Size (px)
Citation preview

The current state of knowledge:
genotypic vs phenotypic drug-susceptibility testing (DST)
Daniela M Cirillo
Emerging Bacterial Pathogens Unit,
San Raffaele Scientific Institute Milan

Outline
• Concordance between molecular based and phenotypic DST
• Rifampicin
• Fluoroquinolones
• Isoniazide
• Pyrazinamide
• SLDs
• Second line LPA preliminary data
• Conclusions
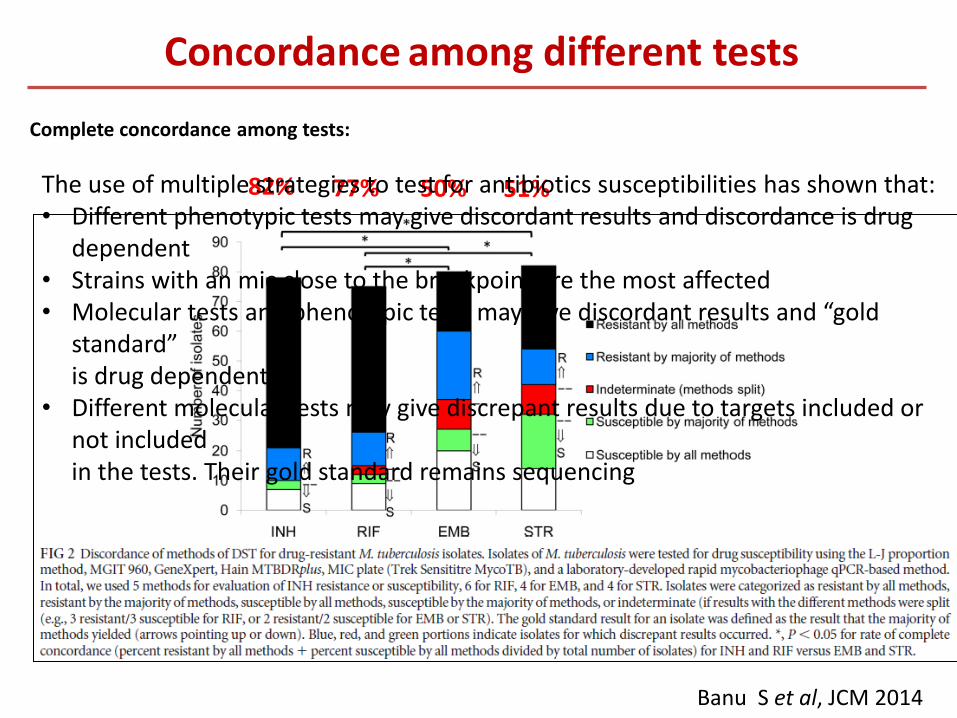
Banu S et al, JCM 2014
82% 77% 50% 51%
Complete concordance among tests:
Concordance among different tests
The use of multiple strategies to test for antibiotics susceptibilities has shown that: • Different phenotypic tests may give discordant results and discordance is drug
dependent • Strains with an mic close to the breakpoint are the most affected • Molecular tests and phenotypic tests may give discordant results and “gold
standard” is drug dependent • Different molecular tests may give discrepant results due to targets included or
not included in the tests. Their gold standard remains sequencing

Genotypic and phenotypic methods provide different pictures
Phenotypic tests :”in vitro” growth in the presence of the drug
• Drug activation/concentration
• Testing media
• Inoculum-related effects
• Reading time
Genotypic tests: identification of mutations associated to impairment of the mechanism of action
• Presence of a mixed population
• Lack of knowledge of all DR determinants
• Unclear association due to lack of gold standard or errors in the gold standard performance
• Cumulative effects
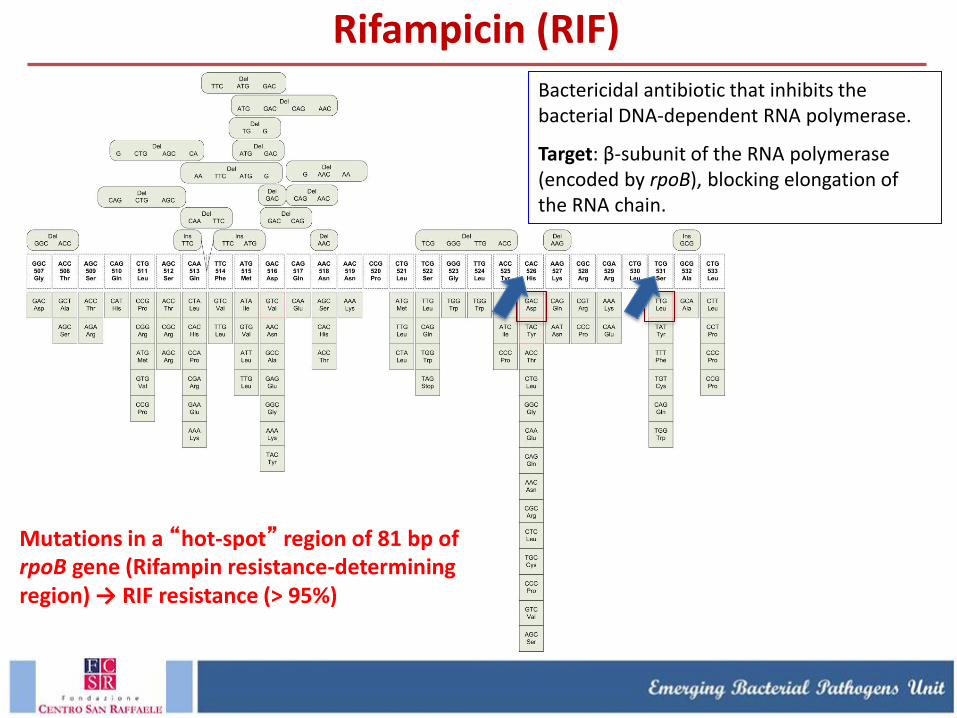
Bactericidal antibiotic that inhibits the bacterial DNA-dependent RNA polymerase.
Target: β-subunit of the RNA polymerase (encoded by rpoB), blocking elongation of the RNA chain.
Mutations in a “hot-spot” region of 81 bp of rpoB gene (Rifampin resistance-determining region) → RIF resistance (> 95%)
Rifampicin (RIF)
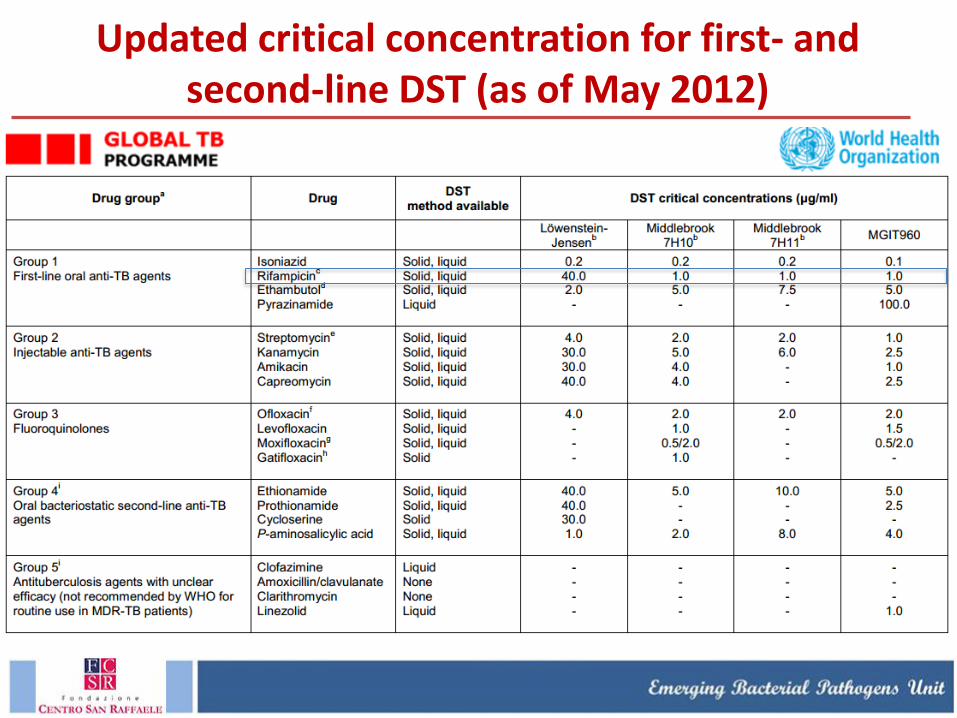
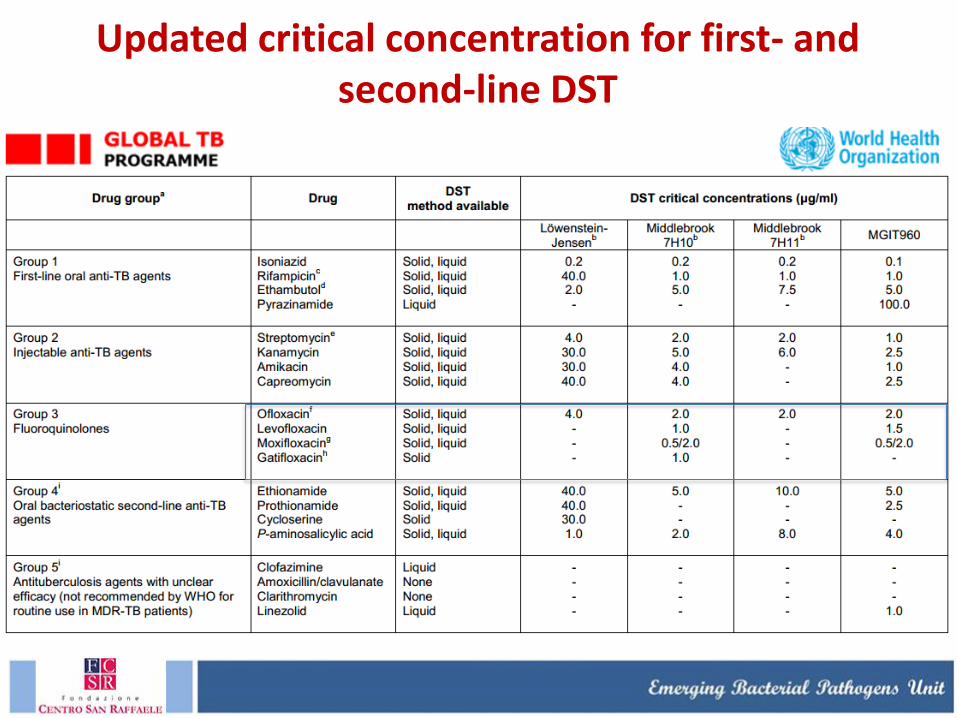
Updated critical concentration for first- and second-line DST (as of May 2012)
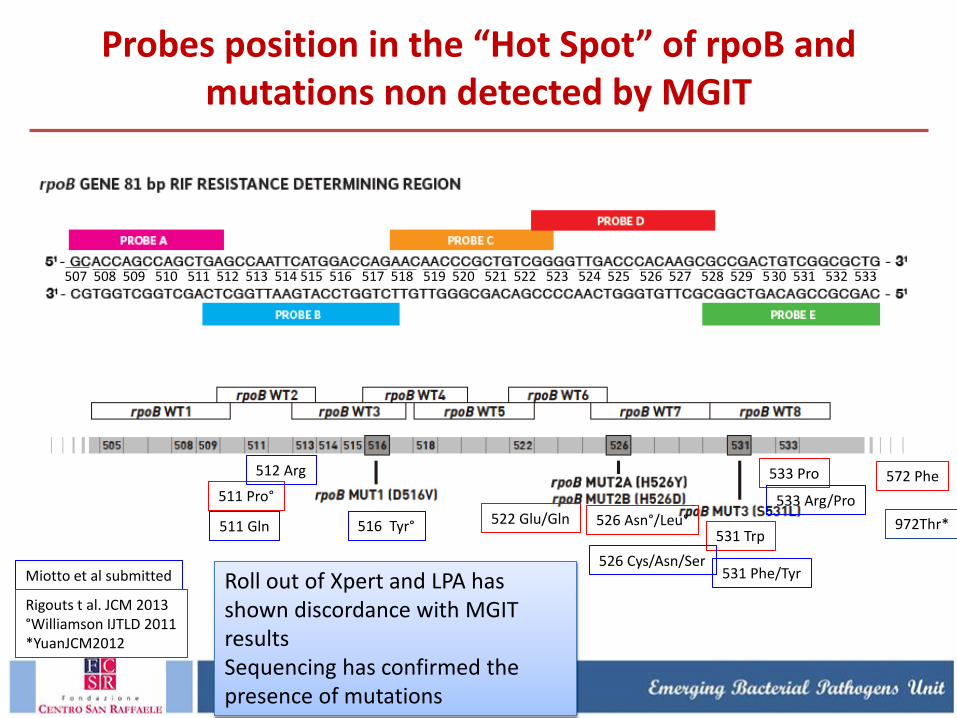
Probes position in the “Hot Spot” of rpoB and mutations non detected by MGIT
507 508 509 510 511 512 513 514 515 516 517 518 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533
511 Pro°
522 Glu/Gln 526 Asn°/Leu° 531 Trp
533 Pro 572 Phe
511 Gln
531 Phe/Tyr
533 Arg/Pro
526 Cys/Asn/Ser
512 Arg
516 Tyr°
Miotto et al submitted
Rigouts t al. JCM 2013 °Williamson IJTLD 2011 *YuanJCM2012
972Thr*
Roll out of Xpert and LPA has shown discordance with MGIT results Sequencing has confirmed the presence of mutations

Liquid DST methods can miss cases of RIF-R
Adapted from Van Deun A et al, J Clin Microbiol 2009; 47(11)3501-6
Missing rate for RIF low-level R : • 81% MGIT960 • 58% Bactec radiometric • 30% agar proportion method
10% RIF-R isolates (mut. at cod. 511, 516, or 533) missed by the
Bactec radiometric method
Van Deun A et al, J Clin Microbiol 2009; 47(11)3501-6 Traore H et al, Int J Tuberc Lung Dis 2000; 4:481-84
Most of the borderline R strains are associated with bacteriologically unfavorable treatment outcomes

Non canonical rpoB mutations were identified in >10% of cases The presence of unconventional mutations correlated with a poor outcome

Silent mutations in the RRDR and mutations outside the RRDR
Silent mutations can be detected by molecular assays but do not modify the aa and the protein structure, and they are NOT relevant for drug resistance. Silent mutations may cause false positivity in molecular tests Silent mutations observed: F506, T508, Q510, L511, Q513, F514, T525, A532, L533, P535 Alonso et al, 2011: silent SNP at cod. F514 was registered in 0.8% of cases Van Deun et al 2013: silent RRDR mutations in retreatment cases occurred in <0.5% of cases Mani et al 2001: silent mutations observed in 4% of cases Yuan et al 2012: silent mutation observed in 1.5% of cases
Mutations outside the RRDR: the transformation of V146F or I572F mutated rpoB into the wild-type M. tuberculosis strains causes RIF-R phenotype Frequency of these mutations: Siu et al 2011: 4% Ahmad et al 2005: 6% Ahmad et al 2012: 11% Van Deun et al 2013: 1-2% Van Deun et al 2009: 5% Rigouts et al 2013: 5% Miotto et al in preparation: 2%
Alonso M et al, J Clin Microbiol 2011; 49(7):2688-90 Moure R et al, J Clin Microbiol 2011; 49(10):3722
Ocheretina O et al, PLoS One 2014; 9(3):e90569 Van Deun A et al, J Clin Microbiol 2013; 51(8):2633-40
Kim BJ et al, J Clin Microbiol 1997; 35(2):492-4 Yuan X et al, J Clin Microbiol 2012; 50(7):2404-13
Siu GK et al, J Antimicrob Chemother 2011; 66(4):730-3 Ahmad S et al, Int J Antimicrob Agents 2005; 26(3):205-12
Van Deun A et al, J Clin Microbiol 2009; 47(11)3501-6 Ahmad S et al, Indian J Med Res 2012; 135(5):756-62
Mani C et al, J Clin Microbiol 2001; 39(8):2987-90 Williamson DA et al, Diagn Microbiol Infect Dis 2011; 74(2):207-9
Kapur V et al, J Clin Microbiol 1994; 32(4):1095-8
Approx. 2%
Approx. 5%

Rif take home message
• Not all genotypic modification of rpoB gene affects phenotypic resistance to RIF equally
• RIF MIC correlates with the position and nature of the amino-acid substitution in rpoB RRDR
• Correlation between MGIT resistance and mutations is high for rpoB codon 531 and for 526D
• Resistance associated to other mutations at codon 526 are not detected by MGIT at CC of 1mg/ml but is detected by proportion methods on LJ or 7H11
• ‘‘Borderline’’ or resistance to RIF associated to unconventional mutations has been strongly associated with treatment failure
• Xpert, LPA: possible false resistance in case of silent mutations in targeted regions (no wt pattern) → sequencing-based approach to overcome the problem and possible false susceptible if the molecular assay doesn't target the mutation (Swaziland example)
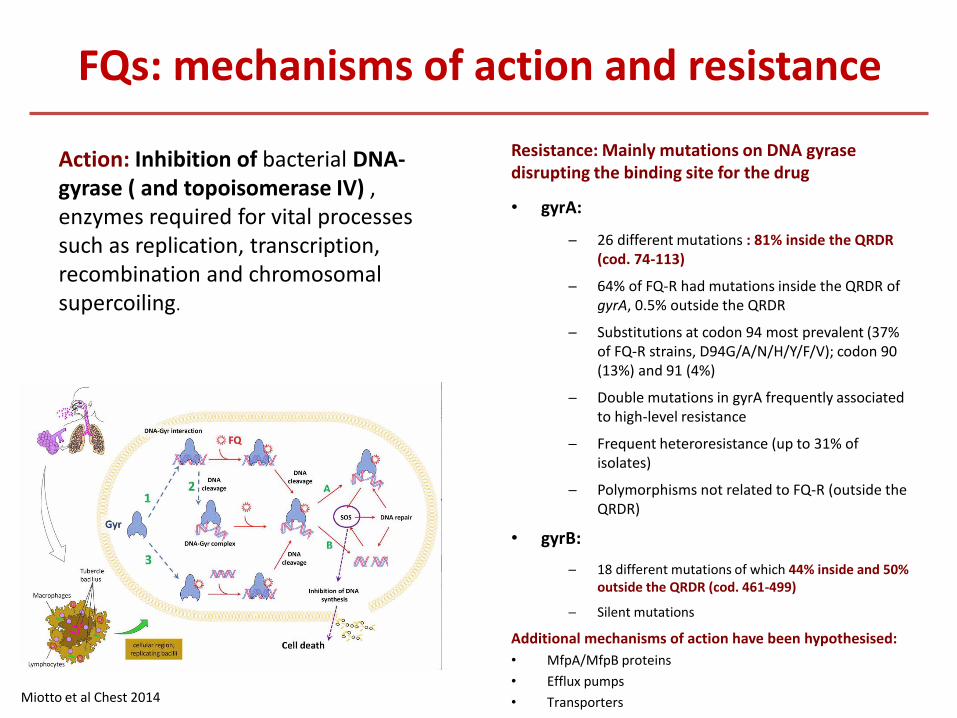
FQs: mechanisms of action and resistance
Resistance: Mainly mutations on DNA gyrase disrupting the binding site for the drug
• gyrA:
─ 26 different mutations : 81% inside the QRDR (cod. 74-113)
─ 64% of FQ-R had mutations inside the QRDR of gyrA, 0.5% outside the QRDR
─ Substitutions at codon 94 most prevalent (37% of FQ-R strains, D94G/A/N/H/Y/F/V); codon 90 (13%) and 91 (4%)
─ Double mutations in gyrA frequently associated to high-level resistance
─ Frequent heteroresistance (up to 31% of isolates)
─ Polymorphisms not related to FQ-R (outside the QRDR)
• gyrB:
─ 18 different mutations of which 44% inside and 50% outside the QRDR (cod. 461-499)
─ Silent mutations
Additional mechanisms of action have been hypothesised:
• MfpA/MfpB proteins
• Efflux pumps
• Transporters Miotto et al Chest 2014
Action: Inhibition of bacterial DNA-gyrase ( and topoisomerase IV) , enzymes required for vital processes such as replication, transcription, recombination and chromosomal supercoiling.
Updated critical concentration for first- and second-line DST

P8A
R68G
H70R low-level (LEV-R) Yin 2010
A74S
T80A FQ-hyperS Aubry 2006
G88A
G88C
D89N
A90E
A90G FQ-hyperS Aubry 2006
A90L
A90V
S91A low-level (OFX-R) Chernyaeva 2013
S91P low-level (OFX-R) Chernyaeva 2013
I92M
D94A low-level (LEV-R) Yin 2010
D94N low-level (LEV-R) Yin 2010
D94G
D94H
D94F
D94T
D94V
P102H
L109V
A126R
A74S + D94G
T80A + A90E
T80A +A90G FQ-hyperS Aubry 2006
T80A + A90G + D94G
G88A + A90V
G88A + D94Y
A90V + D94A
A90V + P102H
A90V + S91P
A90V + D94N
A90V + D94G
S91P + D94G
S91P + D94G + D94A
D94A + D94Y
D94N + D94G
D94N + D94G + D94Y
D94G + D94A
R485C
S447F
D461A MOX/LEV/CIP/OFX-S Malik 2012
D461N OFX/LEV-R only Malik 2012
D461H OFX/LEV-R only Malik 2012
G470A
D494A MOX/LEV/CIP/OFX-S Malik 2012
N499D MOX/LEV/CIP/OFX-R Malik 2012
N499K MOX-R only Malik 2012
N499T
T500N low-level (OFX-R) Chernyaeva 2013
T500P unclear Malik 2012
E501D MOX-R only Malik 2012
E501V MOX/LEV/CIP/OFX-R Malik 2012
E501A low-level (OFX-R) Chernyaeva 2013
A504T
A504V low-level (OFX-R) Chernyaeva 2013
Q538H
D461H + G470A
N499T + T507M MXF/LEV/CIP/OFX-S Malik 2012
A90V + D461A
A90V + N499T
A90V + D94A + N499T
A90V + S91P + D94G + D94A + N499T
A90V + T500P
D94A + D461N
D94G + N499K
D94G + N499T
D94N + A504V
Q
R
D
R
Q
R
D
R
GyrA mutation
GyrB mutation
GyrA + gyrB mutation
• Confidence defined based on: DST results, biochemical/genetic validation, n° of studies reporting association to FQ-R • Maruri et al 2012: 42 studies 2482 clinical isolates (1220 FQ-R, 1262 FQ-S) • Total number of studies considered: gyrA 45; gyrB 16 • Few data associating gyrA-gyrB mutations and clinical outcome are available
Li J et al, Emerg Microb Inf 2014; Maruri F et al, J Antimicrob Chemother 2012;
Aubry A et al, Antimicrob Agents Chemother 2006; Chernyaeva E et al, Tuberculosis 2013
Malik S et al, PLoS One 2012; Lau RW et al, Antimicrob Agents Chemother 2011;
Yin X et al, J Infect 2010 Jo KW et al, Int J Tuberc Lung Dis 2014;
Defining high and low confidence mutations for FQ-R: gyrA
Polymorphisms not related to FQ-R: T80A, A90G, E21Q, S95T, G668D, G247S, A384V Silent mutations: I614I, A830A
81% of mutations inside the QRDR 64% of FQ-R had mutations inside the QRDR of GyrA
Substitutions: cod. 94 37% cod. 90 13% cod. 91 4%
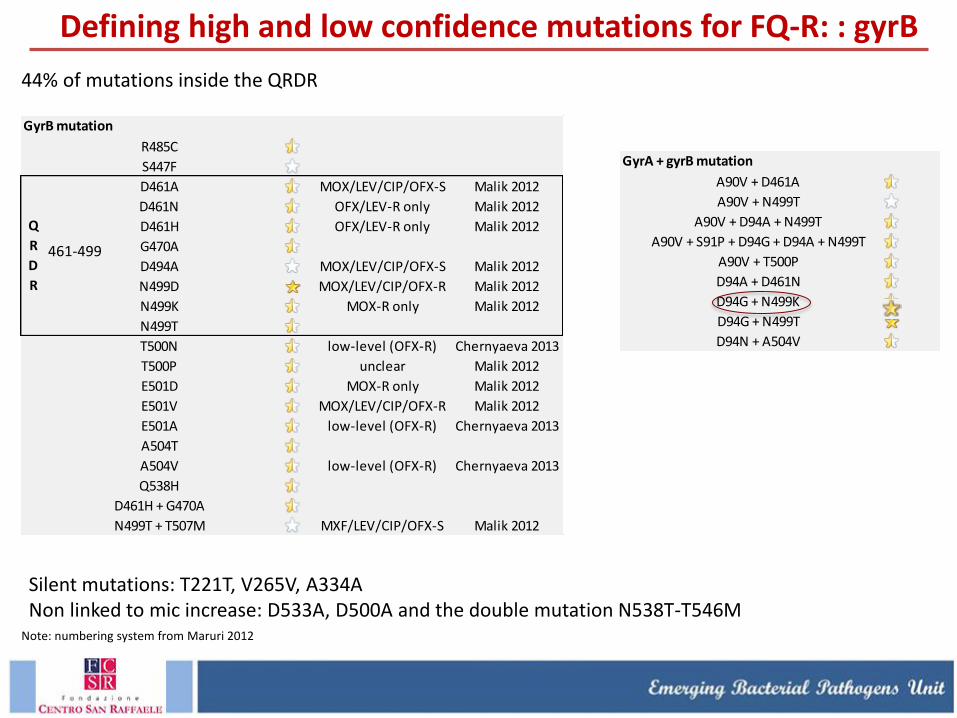
R485C
S447F
D461A MOX/LEV/CIP/OFX-S Malik 2012
D461N OFX/LEV-R only Malik 2012
D461H OFX/LEV-R only Malik 2012
G470A
D494A MOX/LEV/CIP/OFX-S Malik 2012
N499D MOX/LEV/CIP/OFX-R Malik 2012
N499K MOX-R only Malik 2012
N499T
T500N low-level (OFX-R) Chernyaeva 2013
T500P unclear Malik 2012
E501D MOX-R only Malik 2012
E501V MOX/LEV/CIP/OFX-R Malik 2012
E501A low-level (OFX-R) Chernyaeva 2013
A504T
A504V low-level (OFX-R) Chernyaeva 2013
Q538H
D461H + G470A
N499T + T507M MXF/LEV/CIP/OFX-S Malik 2012
Q
R
D
R
GyrB mutation
A90V + D461A
A90V + N499T
A90V + D94A + N499T
A90V + S91P + D94G + D94A + N499T
A90V + T500P
D94A + D461N
D94G + N499K
D94G + N499T
D94N + A504V
GyrA + gyrB mutation
461-499
44% of mutations inside the QRDR
Note: numbering system from Maruri 2012
Silent mutations: T221T, V265V, A334A Non linked to mic increase: D533A, D500A and the double mutation N538T-T546M
Defining high and low confidence mutations for FQ-R: : gyrB
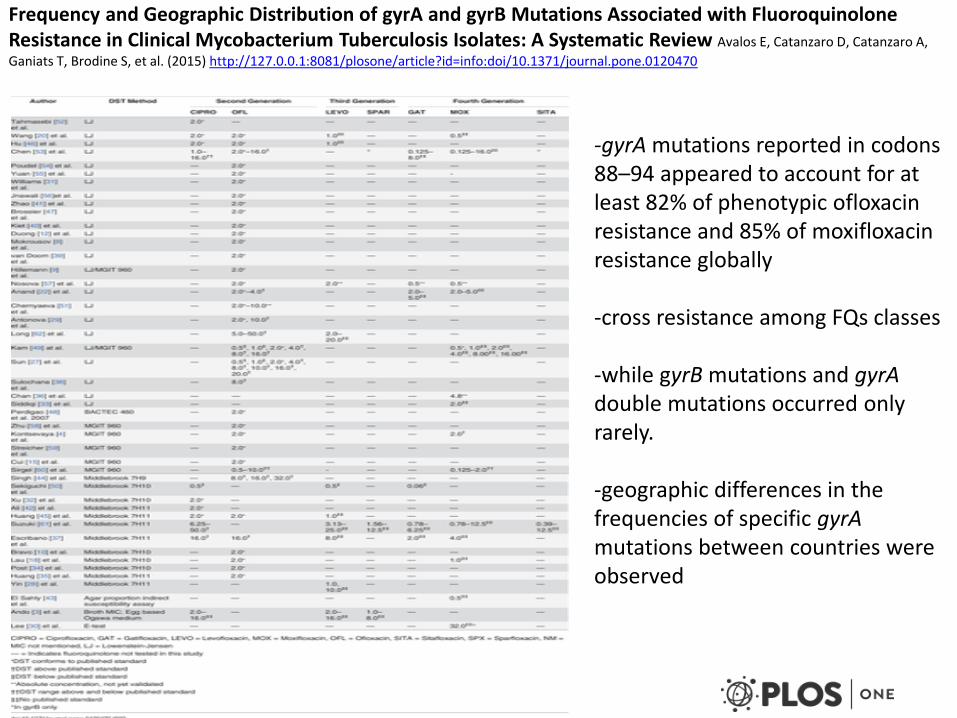
Frequency and Geographic Distribution of gyrA and gyrB Mutations Associated with Fluoroquinolone Resistance in Clinical Mycobacterium Tuberculosis Isolates: A Systematic Review Avalos E, Catanzaro D, Catanzaro A,
Ganiats T, Brodine S, et al. (2015) http://127.0.0.1:8081/plosone/article?id=info:doi/10.1371/journal.pone.0120470
-gyrA mutations reported in codons 88–94 appeared to account for at least 82% of phenotypic ofloxacin resistance and 85% of moxifloxacin resistance globally -cross resistance among FQs classes -while gyrB mutations and gyrA double mutations occurred only rarely. -geographic differences in the frequencies of specific gyrA mutations between countries were observed

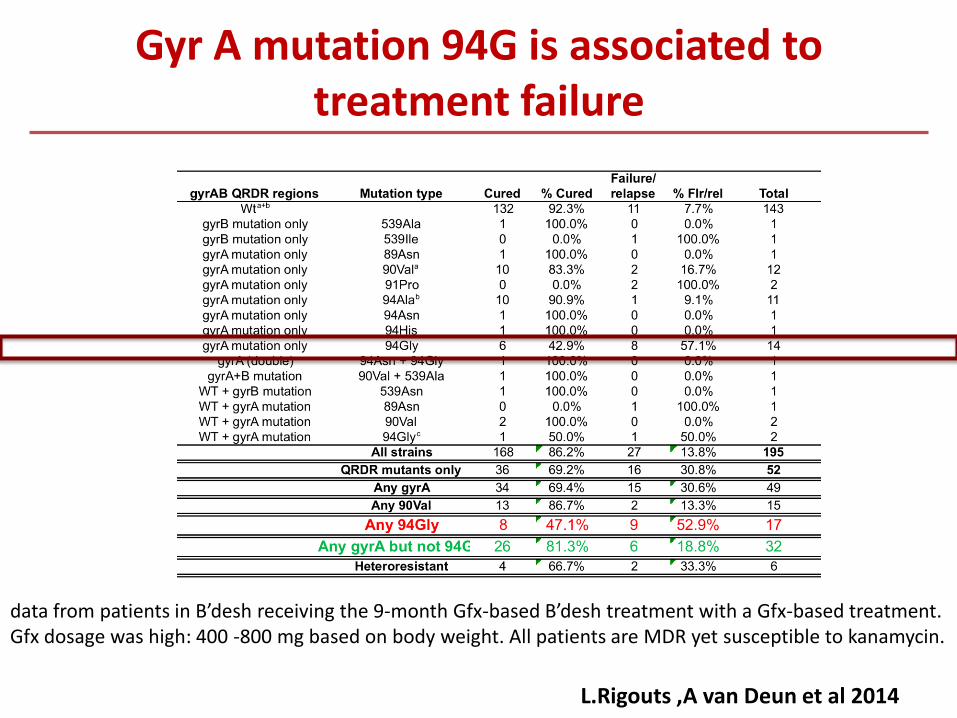
L.Rigouts ,A van Deun et al 2014
Gyr A mutation 94G is associated to treatment failure
data from patients in B’desh receiving the 9-month Gfx-based B’desh treatment with a Gfx-based treatment. Gfx dosage was high: 400 -800 mg based on body weight. All patients are MDR yet susceptible to kanamycin.

Mutations in gyrA QRDR are up today the major determinants for FQs resistance Most mutations outside of the gyrA and gyrB QRDR do not lead to FQ-R or only
slightly increased the MIC levels for FQs Contribution of mutations in gyrB QRDR to FQ R is 1-6% depending on studies The position and the AA substitution is relevant and needs to be identified D94G and D94N high MICs and D94G associated to treatment failure (with
newer FQ) A90V and D94A Ofloxacyn resistance Double mutations are associated to higher mic to all FQs T80A and A90G mutations are associated to FQ hyper-susceptibility Correlation between the presence of mutations and agar based phenotypic tests
for the different drugs is good. MGIT higher breakpoints may need to be revised Genetic background can influence the relevance of some mutations
FQ – take home message

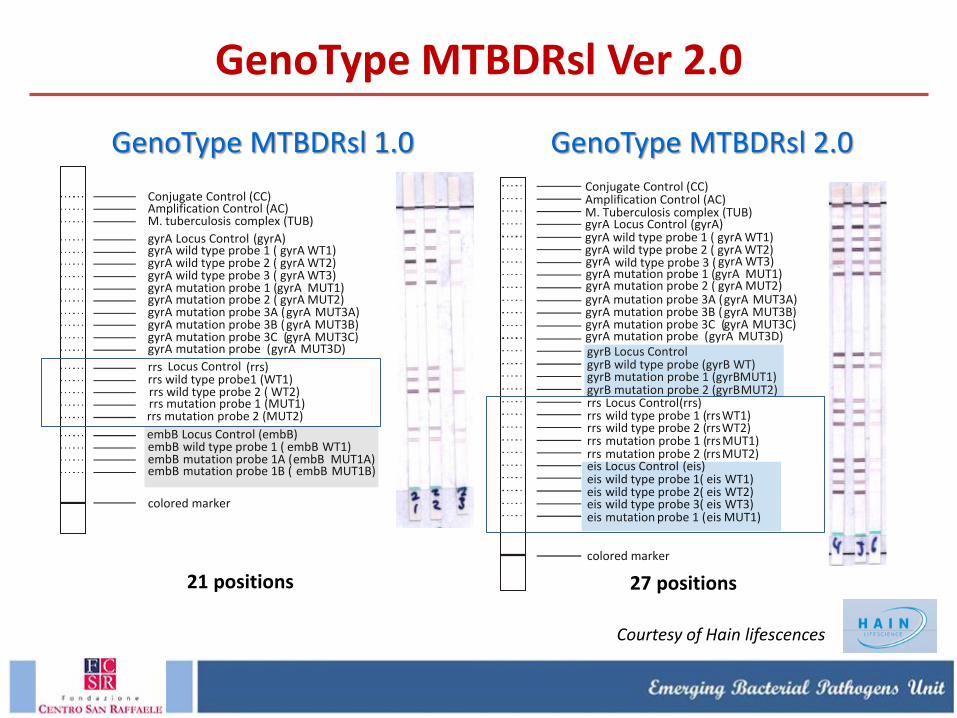
GenoType MTBDRsl 1.0 vs. GenoType MTBDRsl 2.0
Conjugate Control (CC)Amplification Control (AC)
complex (TUB)M. tuberculosis
gyrA (gyrA)gyrA gyrAgyrA gyrAgyrA gyrAgyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA
Locus Control WT1)WT2)WT3)
mutation probe ( MUT )
wild type probe 1 ( wild type probe 2 ( wild type probe 3 ( mutation probe 1 ( MUT1) mutation probe 2 ( MUT2) mutation probe 3A ( MUT3A) mutation probe 3B ( MUT3B) mutation probe 3C ( MUT3C)
3D 3D
rrs (rrs)rrs rrsrrs rrsrrs rrsrrs rrs
Locus Control wild type probe 1 (
wild type probe 2 ( WT2)
mutation probe 2 (
WT1)
mutation probe 1 ( MUT1)MUT2)
embB (embB)embB embBembB embBembB embB
Locus Controlwild type probe 1 (mutation probe 1A (mutation probe 1B (
WT1)
MUT1A)MUT1B)
colored marker
Conjugate Control (CC)Amplification Control (AC)
complex (TUB)M. tuberculosisgyrA (gyrA)gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA
Locus Control wild type probe 1 ( WT1) wild type probe 2 ( WT2) wild type probe 3 ( WT3) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2) mutation probe 3A ( MUT3A) mutation probe 3B ( MUT3B) mutation probe 3C ( MUT3C) mutation probe 3D ( MUT3D)
rrs (rrs)rrs rrsrrs rrsrrs rrs rrs rrs
Locus Control wild type probe 1 ( WT1) wild type probe 2 ( WT2) mutation probe 1 ( MUT1)mutation probe 2 ( MUT2)
colored marker
gyrBgyrB gyrBgyrB gyrBgyrB gyrB
Locus Control wild type probe ( WT) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2)
eis (eis)eis eiseis eiseis eiseis eis
Locus Controlwild type probe 1 ( WT1)wild type probe 2 ( WT2)wild type probe 3 ( WT3)
probe 1 ( MUT1)mutation
GenoType MTBDRsl 1.0 GenoType MTBDRsl 2.0
21 positions 27 positions
Courtesy of Hain lifescences

MTBDRsl Ver 2.0 sensitivity and specificity for FQ resistance
Phenotypic culture-based DST as a reference standard
FQ Culture/DST
Ver 1.0
Ver 2.0
Theron et al compiled data
(Cochrane Rev, 2014)
(95% CI) (95% CI)
Sensitivity 83.1% (78.7-86.7) 83.6% (73.4, 90.3)
Specificity 97.7% (94.3-99.1) 100% (97.6-100)
Tagliani et al submitted to Union 2015
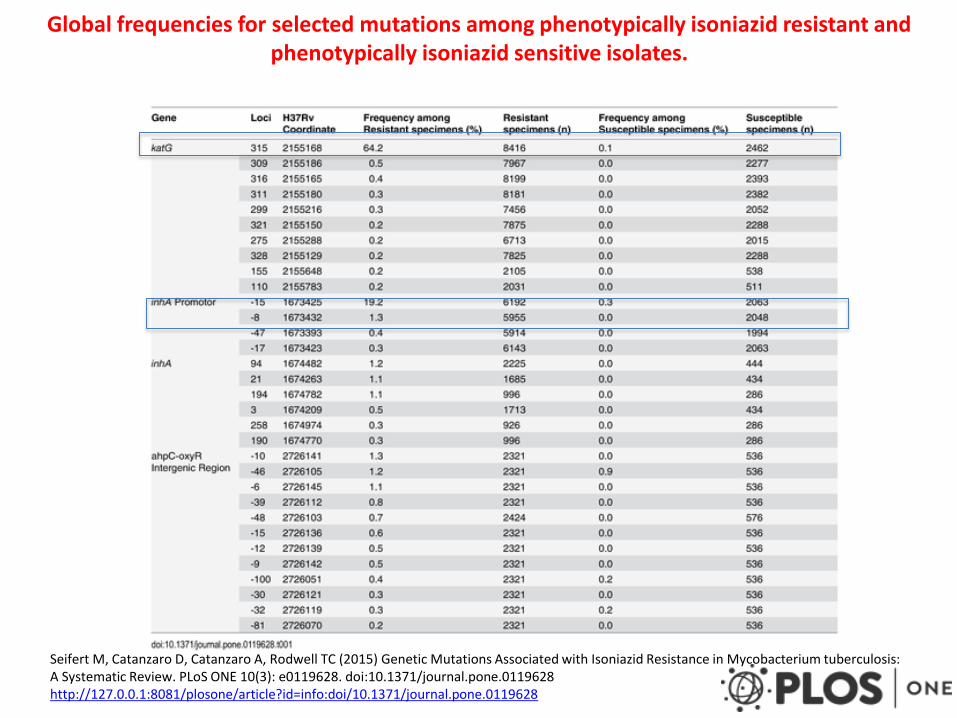
Global frequencies for selected mutations among phenotypically isoniazid resistant and phenotypically isoniazid sensitive isolates.
Seifert M, Catanzaro D, Catanzaro A, Rodwell TC (2015) Genetic Mutations Associated with Isoniazid Resistance in Mycobacterium tuberculosis: A Systematic Review. PLoS ONE 10(3): e0119628. doi:10.1371/journal.pone.0119628 http://127.0.0.1:8081/plosone/article?id=info:doi/10.1371/journal.pone.0119628
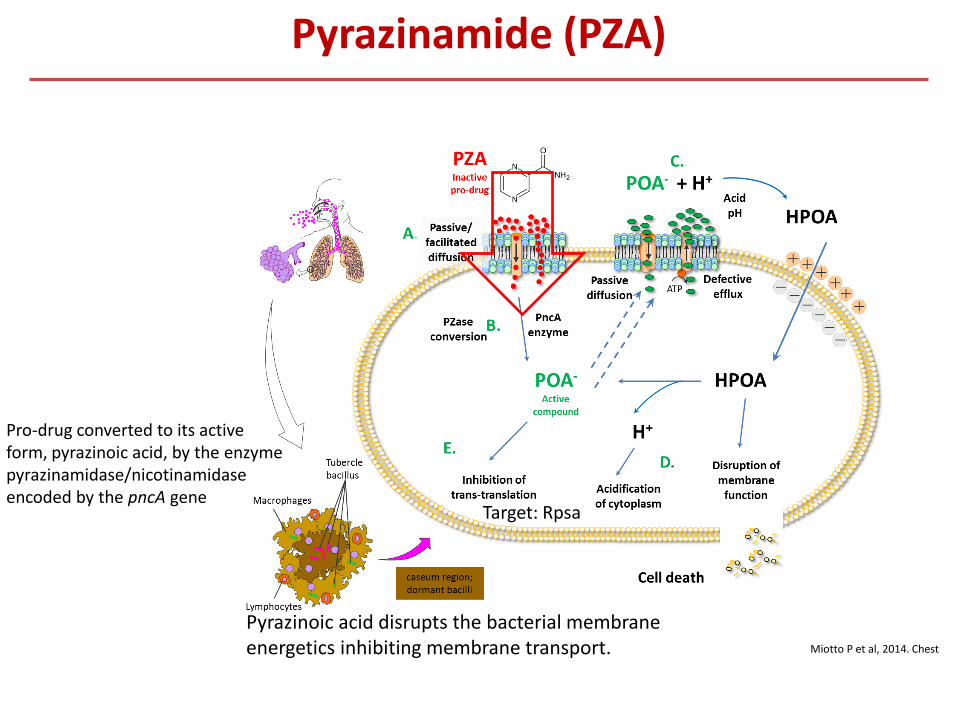
Pyrazinamide (PZA)
Miotto P et al, 2014. Chest
Pro-drug converted to its active form, pyrazinoic acid, by the enzyme pyrazinamidase/nicotinamidase encoded by the pncA gene
Pyrazinoic acid disrupts the bacterial membrane energetics inhibiting membrane transport.
Target: Rpsa

Large inoculum Suboptimal test media with unreliable pH Critical concentration 100 ug/mL (inconsistent results for isolates with a PZA MIC close to this
concentration)
Intermediate category?
57 isolates showed PZA-R on MGIT 960. Repeat testing of resistant isolates with the Bactec 460 reference method confirmed 33 (58%) of these isolates as resistant, and 24 (42%) were susceptible.
Are we over estimating drug resistance to PZA?
Chedore P et al, 2010. J Clin Microbiol; 48(1):300-1
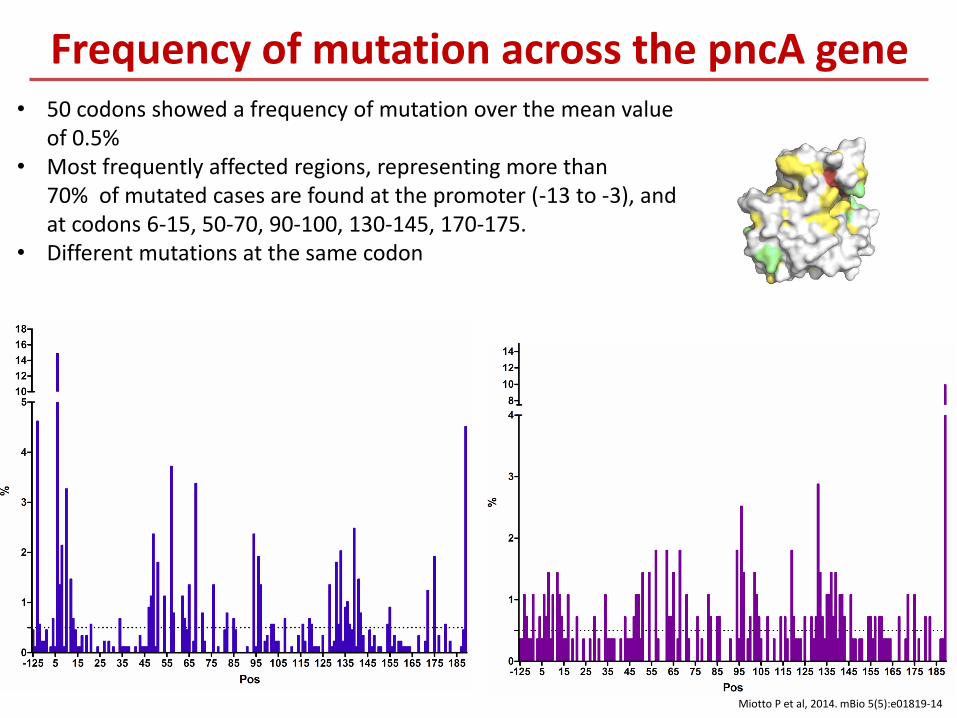
• 50 codons showed a frequency of mutation over the mean value of 0.5%
• Most frequently affected regions, representing more than 70% of mutated cases are found at the promoter (-13 to -3), and at codons 6-15, 50-70, 90-100, 130-145, 170-175.
• Different mutations at the same codon
Frequency of mutation across the pncA gene
Miotto P et al, 2014. mBio 5(5):e01819-14

PZA-R take-home messages
Need of fast resistance detection PZA DST: frequent problems of false resistance Excellent correlation between PZA resistance and pncA mutations About 85% of PZA-R strains carry mutations in pncA Possible development of rapid molecular tests to detect
resistance (sequencing-based?). No hot-spot regions.
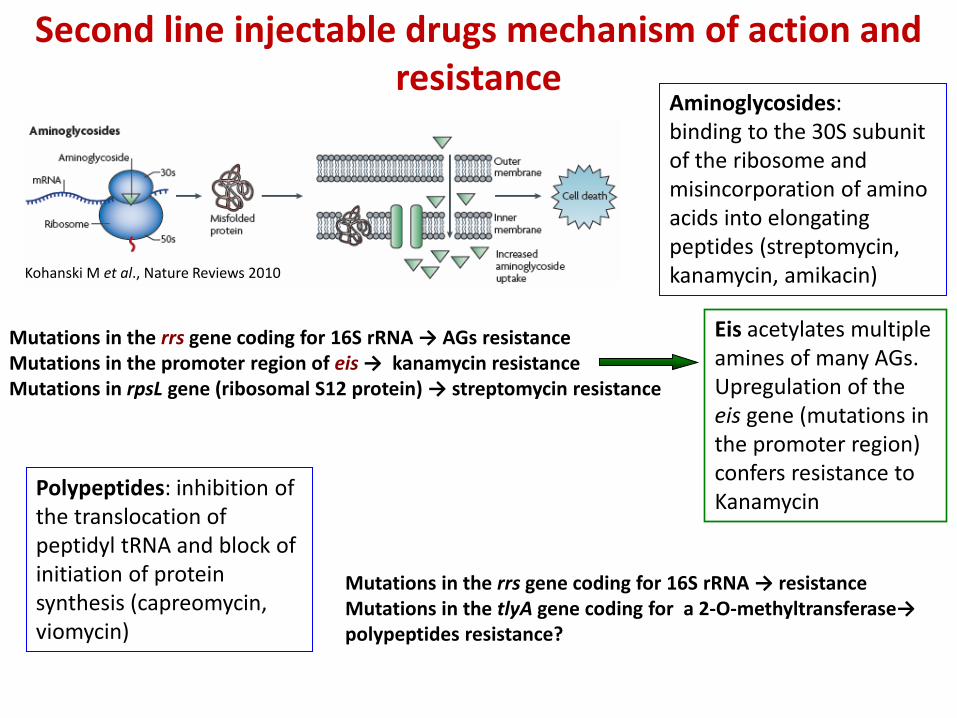
Aminoglycosides: binding to the 30S subunit of the ribosome and misincorporation of amino acids into elongating peptides (streptomycin, kanamycin, amikacin)
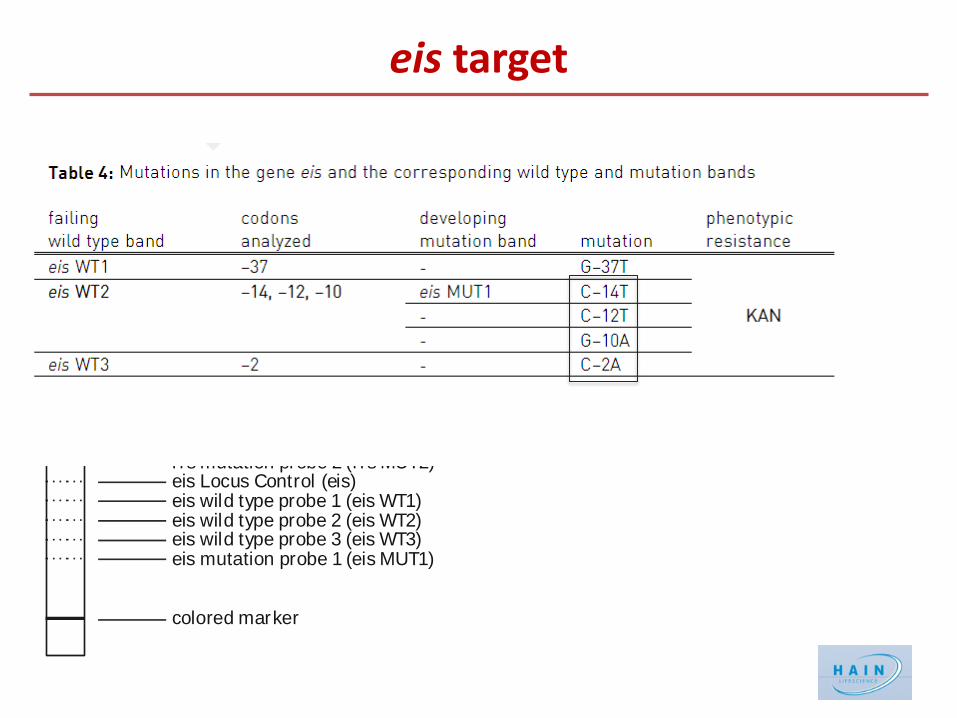
Mutations in the rrs gene coding for 16S rRNA → AGs resistance Mutations in the promoter region of eis → kanamycin resistance Mutations in rpsL gene (ribosomal S12 protein) → streptomycin resistance
Kohanski M et al., Nature Reviews 2010
Eis acetylates multiple amines of many AGs. Upregulation of the eis gene (mutations in the promoter region) confers resistance to Kanamycin
Polypeptides: inhibition of the translocation of peptidyl tRNA and block of initiation of protein synthesis (capreomycin, viomycin)
Mutations in the rrs gene coding for 16S rRNA → resistance Mutations in the tlyA gene coding for a 2-O-methyltransferase→ polypeptides resistance?
Second line injectable drugs mechanism of action and resistance
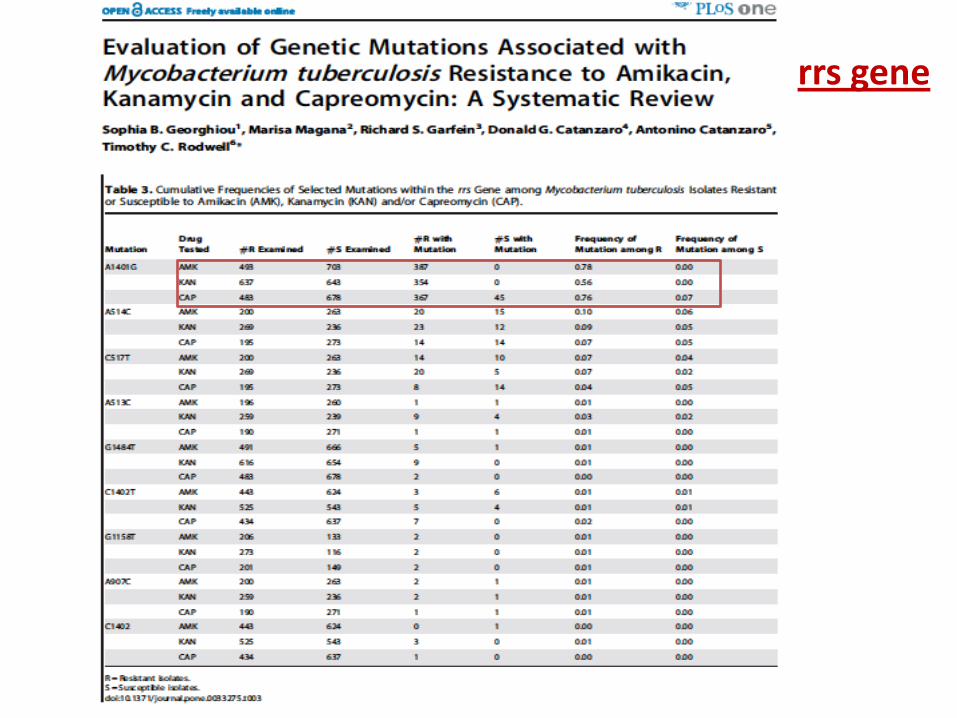
rrs gene
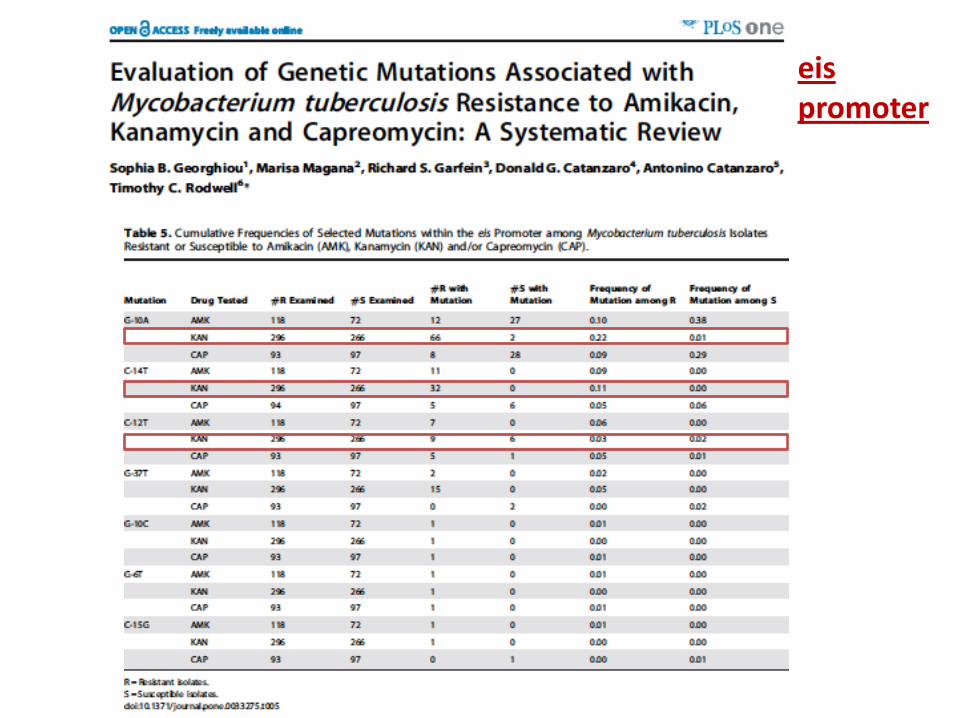
eis promoter
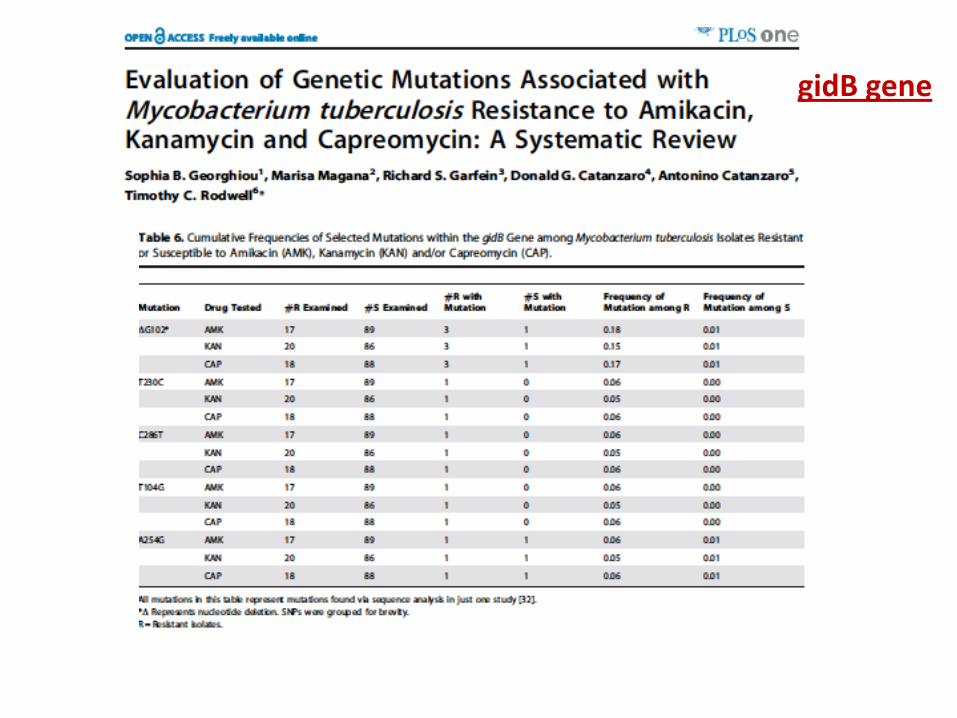
gidB gene

SLIDs – take home message
Selected rrs mutations are relevant for SLIDs resistance 1401G is the most relevant mutation associated to high
resistance to all SLIDs Mutations in the eis promoter region are relevant
determinants for resistance to kanamycin. Positions and AAs substitutions are relevant
MTBDRsl Ver2.0 has highly increased his sensitivity compared to version1by the addition of eis promoter region; specificity will need to be addressed with more data on specific mutations
GenoType MTBDRsl Ver 2.0
GenoType MTBDRsl 1.0
21 positions
Courtesy of Hain lifescences
Conjugate Control (CC) Amplification Control (AC)
complex (TUB) M. tuberculosis
gyrA (gyrA) gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA
Locus Control WT1) WT2) WT3)
mutation probe ( MUT3D)
wild type probe 1 ( wild type probe 2 ( wild type probe 3 ( mutation probe 1 ( MUT1) mutation probe 2 ( MUT2) mutation probe 3A ( MUT3A) mutation probe 3B ( MUT3B) mutation probe 3C ( MUT3C)
rrs (rrs) rrs wild type probe1 (WT1)
Locus Control
rrs wild type probe 2 ( WT2)
rrs mutation probe 2 (MUT2) rrs mutation probe 1 (MUT1)
embB embB embB embB embB embB
embB Locus Control (embB) wild type probe 1 ( mutation probe 1A ( mutation probe 1B (
WT1)
MUT1A) MUT1B)
colored marker
GenoType MTBDRsl 2.0
27 positions
Conjugate Control (CC) Amplification Control (AC)
complex (TUB) M. Tuberculosis gyrA (gyrA) Locus Control
rrs (rrs) rrs rrs rrs rrs rrs rrs rrs rrs
Locus Control wild type probe 1 ( WT1) wild type probe 2 ( WT2) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2)
colored marker
gyrB gyrB gyrB gyrB gyrB gyrB
Locus Control wild type probe (gyrB WT) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2)
eis (eis) eis eis eis eis eis eis eis eis
Locus Control wild type probe 1( WT1) wild type probe 2( WT2) wild type probe 3( WT3)
probe 1 ( MUT1) mutation
gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA mutation probe ( MUT3D)
mutation probe 3A ( MUT3A) mutation probe 3B ( MUT3B) mutation probe 3C ( MUT3C)
gyrA gyrA gyrA gyrA gyrA gyrA
WT3) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2)
wild type probe 3 (
gyrA gyrA gyrA gyrA
WT1) WT2)
wild type probe 1 ( wild type probe 2 (
Conjugate Control (CC)Amplification Control (AC)
complex (TUB)M. tuberculosisgyrA (gyrA)gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA gyrA
Locus Control wild type probe 1 ( WT1) wild type probe 2 ( WT2) wild type probe 3 ( WT3) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2) mutation probe 3A ( MUT3A) mutation probe 3B ( MUT3B) mutation probe 3C ( MUT3C) mutation probe 3D ( MUT3D)
rrs (rrs)rrs rrsrrs rrsrrs rrs rrs rrs
Locus Control wild type probe 1 ( WT1) wild type probe 2 ( WT2) mutation probe 1 ( MUT1)mutation probe 2 ( MUT2)
colored marker
gyrBgyrB gyrBgyrB gyrBgyrB gyrB
Locus Control wild type probe ( WT) mutation probe 1 ( MUT1) mutation probe 2 ( MUT2)
eis (eis)eis eiseis eiseis eiseis eis
Locus Controlwild type probe 1 ( WT1)wild type probe 2 ( WT2)wild type probe 3 ( WT3)
probe 1 ( MUT1)mutation
First Data on the new MTBDRsl VER2.0 eis target
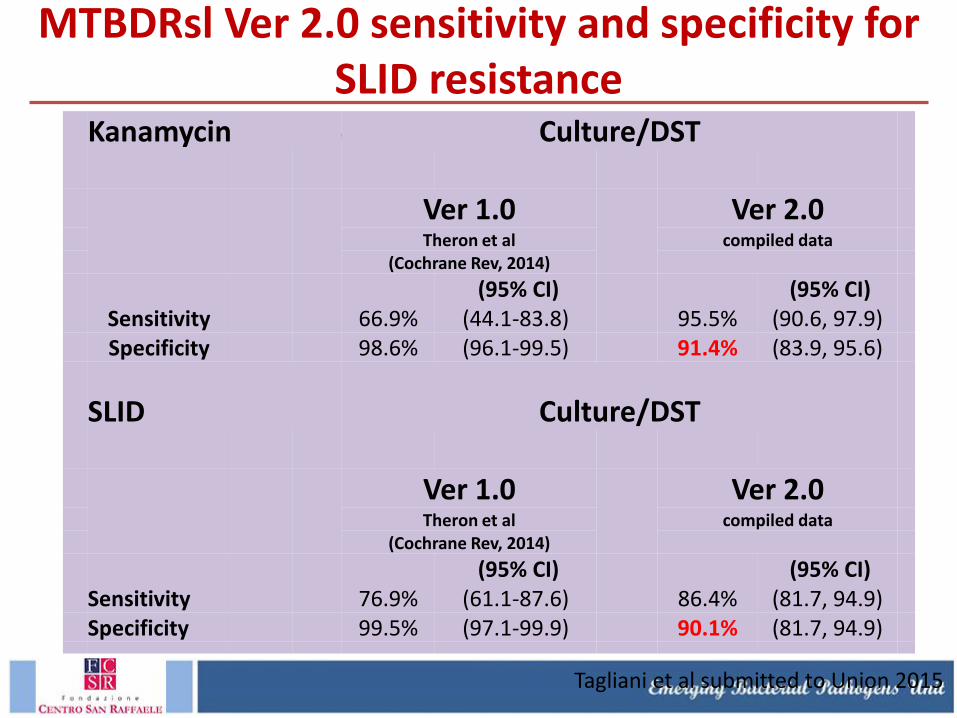
MTBDRsl Ver 2.0 sensitivity and specificity for SLID resistance
Phenotypic culture-based DST as a reference standard
Kanamycin Culture/DST
Ver 1.0
Ver 2.0
Theron et al compiled data
(Cochrane Rev, 2014)
(95% CI) (95% CI)
Sensitivity 66.9% (44.1-83.8) 95.5% (90.6, 97.9)
Specificity 98.6% (96.1-99.5) 91.4% (83.9, 95.6)
SLID Culture/DST
Ver 1.0
Ver 2.0
Theron et al compiled data
(Cochrane Rev, 2014)
(95% CI) (95% CI)
Sensitivity 76.9% (61.1-87.6) 86.4% (81.7, 94.9)
Specificity 99.5% (97.1-99.9) 90.1% (81.7, 94.9)
Tagliani et al submitted to Union 2015

Performance characteristics of Genotype®MTBDRsl V2.0 for detection of FQ resistance is comparable to Genotype®MTBDRsl V1.0.
MTBDRsl test can correctly ID the relevant mutations in gyr A (and gyr B).
Contribution of mutations in gyrB QRDR to FQ R is 1-6% depending on studies.
MTBDRsl Ver2.0 has an increased sensitivity for SLID resistance compared to Ver 1.0 due to the addition of eis promoter region; specificity will need to be addressed with more data on specific mutation.
Mutations in the eis promoter region are relevant determinants for resistance to kanamycin. Positions and Ns substitutions are relevant.
Use of Hain sl– take home message

Conclusion • The identification of the nature of the mutations is needed for accurate diagnosis of
resistance
• There are high confidence genetic markers of resistance that can replace conventional DST
• RIF, rpoB gene: mutations at cod. 531 and specific mutations at codons 513 and 526; multiple mutations
• INH, katG gene: mutations at cod. 315 • FQ, gyrA-gyrB genes: specific mutations within the QRDRs • PZA, pncA gene: specific mutations (85%) • SLID, rrs gene: a1401g
• There are genetic markers for low-level resistance that can be used to improve clinical management of the patients • RIF, rpoB gene: 511P, 515I, 516Y, 526L, 526N, 526C, 526S, 532V, 533P, and 572F • INH, inhA gene: c-15t • FQ, gyrA gene: T80A, A90G, A90V, D94A
• Identification of the type of mutation is relevant for patient management

A common platform to investigate the relationship between mutations, phenotypic, surveillance and clinical data If interested please contact:
Dr David Dolinger: [email protected] Dr Paolo Miotto: [email protected]
A joint effort toward a common goal: providing effective diagnostic tools where are mostly needed
More data are needed

IRCCS San Raffaele
R Alagna
B Asimewe
R Baldan
S.Battaglia
E Borroni
AM Cabibbe
L Furci
P Miotto
E Schena
E Tagliani
E Tortoli
A Trovato
Acknowledgements
Hain lifesciences
FZB Borstel:
S. RueshGerdes
D. Hilleman
SRL Stockolm:
Sven Hoffner
NTP/NRL Belarus
Alena Skrahina
Aksana Zalutskaya
SRLN and TBPANNET Consortium





























