Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY, Feb. 2009, p. 714–725 Vol. 29, No. 30270-7306/09/$08.00�0 doi:10.1128/MCB.00825-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
The CTCF Insulator Protein Is Posttranslationally Modified by SUMO�
Melissa J. MacPherson,1 Linda G. Beatty,1 Wenjing Zhou,2 Minjie Du,3 and Paul D. Sadowski1*Department of Molecular Genetics, 1 King’s College Circle, University of Toronto, Toronto, ON M5S 1A8, Canada1; the Campbell
Family Institute for Breast Cancer Research, Princess Margaret Hospital, University Health Network, 620 University Avenue,Toronto M5G 2M9, Canada2; and Microbix Biosystems Inc., 341 Bering Avenue, Toronto M8Z 3A8, Canada3
Received 22 May 2008/Returned for modification 23 June 2008/Accepted 10 November 2008
The CTCF protein is a highly conserved zinc finger protein that is implicated in many aspects of generegulation and nuclear organization. Its functions include the ability to act as a repressor of genes, includingthe c-myc oncogene. In this paper, we show that the CTCF protein can be posttranslationally modified by thesmall ubiquitin-like protein SUMO. CTCF is SUMOylated both in vivo and in vitro, and we identify two majorsites of SUMOylation in the protein. The posttranslational modification of CTCF by the SUMO proteins doesnot affect its ability to bind to DNA in vitro. SUMOylation of CTCF contributes to the repressive function ofCTCF on the c-myc P2 promoter. We also found that CTCF and the repressive Polycomb protein, Pc2, arecolocalized to nuclear Polycomb bodies. The Pc2 protein may act as a SUMO E3 ligase for CTCF, stronglyenhancing its modification by SUMO 2 and 3. These studies expand the repertoire of posttranslationalmodifications of CTCF and suggest roles for such modifications in its regulation of epigenetic states.
CTCF is a highly conserved zinc finger protein that is in-volved in many cellular functions, including gene repression (3,8, 49), gene activation (9), chromatin insulator function (5, 6,9, 36), and the maintenance of genomic imprinting (21, 25, 55).CTCF prevents initiation and/or spreading of DNA methyl-ation of imprinted genes (24, 56, 62, 66). It is also implicated incarcinogenesis, and missense mutations in its zinc finger DNAbinding domain have been described in breast, prostate, andkidney tumors (27). The CTCF protein plays a role in anotherepigenetic process, X chromosome inactivation (11, 58).
Recently, CTCF has been implicated in genomic organiza-tion, mediating intrachromosomal interactions at the �-globinHS4 insulator (64), the H19/IGF2 imprinting control region(44, 74), and the major histocompatibility complex class IIgenes (50), as well as interchromosomal interactions betweenthe IGF2 locus on human chromosome 11 and the Wsb1 locuson chromosome 7 (47). Interestingly, CTCF is required forsite-specific binding of cohesin proteins throughout the ge-nome, and cohesin has been implicated in insulator activity(57, 65, 69). Several bioinformatic and chromatin immunopre-cipitation studies have recently estimated that there are around15,000 CTCF binding sites in the human genome. These CTCFbinding sites have been found to delineate the boundaries ofdifferent chromatin regions, and many of these binding sitespossess insulator activity (4, 41, 53, 71, 72). Despite the impor-tant role that CTCF plays in gene expression, the molecularmechanisms underlying its many functions remain enigmatic.
CTCF is posttranslationally modified by both phosphoryla-tion and poly(ADP) ribosylation (43, 75). Phosphorylation ofCTCF relieves its repressive activity at the c-myc P2 promoter(22, 43), and its poly(ADP) ribosylation is needed for it to actas an insulator protein (42, 75). We have now added another
level of complexity to CTCF’s multiple functions by findingthat it is posttranslationally modified by the SUMO (for smallubiquitin-like modifier) proteins.
The SUMO proteins are members of the family of ubiquitin-like proteins that covalently modify their target proteins (40).SUMO proteins are roughly 100 amino acids long, about 20amino acids longer than ubiquitin. Their three-dimensionalstructure is similar to that of ubiquitin, although they are only18% identical in sequence (28, 29, 60, 63). SUMO 1 differs insequence by about 50% from SUMO 2 and 3, which are about97% identical. The biochemical pathway for the modificationof proteins by SUMO proteins is mechanistically similar to, butdistinct from, that of ubiquitin (28, 29).
Unlike polyubiquitin, SUMO modification of a protein doesnot target it for degradation. The list of SUMO-modified pro-teins is expanding rapidly. Many of them play key roles intranscriptional regulation, chromatin structure, and DNA re-pair. Since SUMOylation is often associated with the repres-sion of gene expression, and genomic imprinting and insulatorfunction involve the silencing of genes, we asked whether theCTCF insulator protein was SUMOylated.
We report that the CTCF protein can be modified by SUMO1, 2, and 3. We have identified two major sites of SUMO-ylation, one in the COOH-terminal domain and one in theNH2-terminal domain. These sites are conserved from fish tomammals. We show that the SUMO modification is function-ally important for the repressive action of CTCF on the c-mycP2 promoter. Furthermore, we found that the Polycomb groupprotein Polycomb 2 (Pc2) likely acts as a SUMO E3 ligase inthe SUMOylation of CTCF. Finally, we report that CTCF iscolocalized with the Pc2 protein to nuclear particles calledPolycomb bodies. Our findings suggest that the SUMOylationof CTCF is involved in the organization of repressive chroma-tin domains involving CTCF-responsive genes.
MATERIALS AND METHODS
Cell culture and transfection. HEK 293 and HeLa cells were maintained inDulbecco’s modified Eagle’s medium (Invitrogen) supplemented with 10% fetal
* Corresponding author. Mailing address: Department of MolecularGenetics, Medical Sciences Building, Room 4284, University of To-ronto, 1 King’s College Circle, Toronto, ON M5S 1A8, Canada. Phone:(416) 978-6061. Fax: (416) 978-6885. E-mail: [email protected].
� Published ahead of print on 24 November 2008.
714
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

bovine serum and 1% penicillin/streptomycin. HEK 293 cells were transfectedusing calcium phosphate as previously described (19, 20). HeLa cells were trans-fected using Fugene 6 transfection reagent (Roche Diagnostic Products), accord-ing to the manufacturer’s instructions.
Plasmids and cloning. HEK 293 cDNA was prepared from total RNA isolatedusing Tri Reagent (Sigma) and was reverse transcribed with Superscript II(Invitrogen) using random hexamers. The coding sequence of Ufm1 (for ubiq-uitin fold-like modifier 1) (NM_016617) and the coding sequences for the pro-cessed forms of SUMO 1 (NM_001005781.1; nucleotides [nt] 149 to 439), SUMO2 (NM_006937.3; nt 170 to 448), and SUMO 3 (NM_006936.2; nt 162 to 437)were PCR amplified from HEK 293 cDNA using the primers listed in Table 1.The PCR products were cloned into the plasmid pCMVmyc, which contains thecytomegalovirus promoter and an N-terminal myc tag (2). CTCF mouse cDNA(36) (a gift from S. Tilghman and C. Schoenherr) was PCR amplified and clonedinto the plasmid pMZS3F (78) (a generous gift of J. Greenblatt) containing aC-terminal calmodulin binding peptide, a tobacco etch virus protease cleavagesite, and a three-FLAG tag. Mutagenesis reactions were carried out using Strat-agene’s QuikChange Multi site directed mutagenesis kit or QuikChange XL sitedirected mutagenesis kit. The sentrin-specific protease 1 (SENP1) and SENP5expression vectors (a generous gift from E. T. Yeh) were previously described(12, 31). The T7-Pc2 expression vector (a generous gift from D. Wotton) waspreviously described (39). The DNA encoding the NH2-terminal and COOH-terminal domains of CTCF (amino acids 1 to 266 and 570 to 736, respectively)was PCR amplified from a cDNA clone and cloned into the vector pGEX4T-1(Amersham) using the primers listed in Table 1 to create NH2-terminal gluta-thione S-transferase (GST) fusions of the two domains.
Purification of NH2- and COOH-terminal domains and in vitro SUMO-ylation. Overnight cultures of Escherichia coli BL21 cells grown in LB containing100 �g/ml ampicillin at 37°C and 300 rpm were diluted 1:10 in fresh LB plusampicillin and grown at 30°C and 250 rpm until an optical density at 600 nmbetween 0.8 and 1.0 was reached. The GST fusion domains were expressed byinduction with 0.1 mM IPTG (isopropyl-�-D-thiogalactopyranoside) at 20°C.Protein was purified using glutathione resin (Amersham) and eluted using re-duced glutathione. Purified protein was dialyzed against 20 mM HEPES, pH 8.0,20% glycerol, 100 mM KCl, 0.2 mM EDTA, 1 mM dithiothreitol and quantifiedusing the Bio-Rad protein assay.
A SUMOylation control kit for SUMO 1 was purchased from LAE BiotechInternational (catalog no. K007) and used according to the manufacturer’s in-structions. Additional SUMO 1, 2, and 3 proteins were purchased from BostonBiochem, Inc. (catalog no. Uh-752 and Uh-762). Briefly, 5 �l of in vitro-trans-lated CTCF or 0.5 �g of the purified GST-tagged CTCF domain was incubatedwith the recommended amounts of E1 and E2 enzymes and 2 mM ATP, with orwithout SUMO proteins, in a 20- or 10-�l reaction mixture for 1 h at 37°C. Thereaction mixture was used in electrophoretic mobility shift assays or analyzed bysodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), fol-lowed by Western blotting and probing with the appropriate antibody (anti-CTCF C-20 or N-17; Santa Cruz Biotechnology). In the case of 35S-labeledCTCF, the gel was dried and exposed to a phosphorimager screen.
Immunoprecipitation and Western blotting. Ten-centimeter dishes of conflu-ent HEK 293 cells overexpressing the tagged proteins were lysed in 200 �l ofbuffer containing 62.5 mM Tris-Cl, pH 6.8, 2% SDS, 10% glycerol, proteaseinhibitor cocktail (Sigma), and 1 mM N-ethyl maleimide by boiling them for 5
min. The lysates were then sonicated three times for 5 s each time to reduceviscosity, followed by centrifugation at 15,000 rpm for 15 min to remove cellulardebris. The lysates were diluted fivefold with dilution buffer (50 mM Tris-Cl, pH8.0, 150 mM NaCl, 1% NP-40) incubated with Sepharose Protein G resin (GEHealthcare) and 5 �g of anti-FLAG antibody at 4°C for 3 to 16 h. The immu-noprecipitates were washed three times in wash buffer (50 mM Tris-Cl, pH 8.0,150 mM NaCl, 0.1% NP-40), boiled in 2� SDS-PAGE loading buffer, separatedby SDS-PAGE, and electroblotted onto polyvinylidene difluoride (PVDF) mem-branes (GE Healthcare). The membranes were incubated in mouse anti-FLAGM2 (Sigma; F-3165), rabbit anti-myc (Abcam; ab9106), or goat anti-beta-actin(Abcam; ab8229) primary antibody and horseradish peroxidase-conjugated sec-ondary antibodies (Santa Cruz) and detected using ECL Western Blotting De-tection Reagents (GE Healthcare; RPN2109). In immunoprecipitation experi-ments for Pc2 E3 ligase activity, HeLa cells were transfected using Fugene 6. TheT7-Pc2-transfected HeLa cell lysates were precleared of Pc2 using 20 �l of a 50%slurry of anti-T7 agarose (Abcam; ab1230) prior to immunoprecipitation ofCTCF with anti-FLAG antibody as described above. The detection of the en-dogenous SUMOylation of CTCF was performed by lysing five confluent 15-cmtissue culture dishes of HEK 293 cells in 1 ml of lysis buffer by boiling them ina kettle for 10 min. The lysate was sonicated to reduce the viscosity, centrifugedas before, and diluted 10-fold with dilution buffer containing 1 mM N-ethylmaleimide. In each immunoprecipitation, 2 ml of the diluted lysate was added toSepharose-protein G resin and 10 �g of anti-SUMO 1 (D-11) antibody (SantaCruz) or 10 �g of mouse immunoglobulin G (IgG) (Santa Cruz) as a control andwas incubated at 4°C for 16 h. The immunoprecipitates were washed three timesin wash buffer (50 mM Tris-Cl, pH 8.0, 250 mM NaCl, 0.1% NP-40, and 1 mMN-ethyl maleimide), boiled in 2� SDS-PAGE loading buffer, separated by SDS-PAGE, and electroblotted onto a PVDF membrane as before. The membranewas probed with anti-CTCF (C-20) antibody (Santa Cruz) and horseradish per-oxidase-conjugated secondary antibody and detected using an ECL Plus Westernblotting detection system (GE Healthcare; RPN2132).
Immunofluorescence staining and confocal microscopy. HeLa cells grown onglass coverslips were fixed in 3% (wt/vol) paraformaldehyde in phosphate-buff-ered saline (PBS) for 30 min at 37°C. The cells were then washed two times inPBS at 37°C. The cells were permeabilized with 1% (wt/vol) Triton X-100 in PBSfor 5 min at room temperature and blocked in 3% (wt/vol) bovine serum albuminin PBS for 30 min at 37°C. The coverslips were then incubated with anti-CTCFprimary antibody (1:50 dilution) (BD Biosciences; 612148) or anti-CTCF (1:25 or1:10 dilution) (Santa Cruz Biotechnology; N-17), anti-Pc2 (1:200 dilution) (Ab-cam; ab4189), anti-promyelocytic leukemia protein (PML) (1:50) (Santa CruzBiotechnology; PG-M3), or anti-SC-35 (1:200 dilution) (Sigma) at 37°C for 1 h,followed by two washes in 0.05% Tween 20 in PBS at 37°C. The coverslips wereincubated in secondary antibodies conjugated with Texas Red or fluoresceinisothiocyanate (Jackson ImmunoResearch), Alexa 488, or Alexa 647 (Invitrogen)at 37°C for 1 h, washed with 0.05% Tween 20 in PBS at 37°C, and mounted withProlong Gold antifade reagent (Invitrogen). Cell samples were examined using aZeiss 63� numerical aperture PlanApo objective lens. Images were recordedusing a Zeiss LSM 510 confocal laser microscope. The colocalization of CTCFand Pc2, SC-35, or PML was determined using the ImageJ JaCOP colocalizationplug-in (7; http://rsb.info.nih.gov/ij/).
Luciferase reporter assays. The HEK 293 cells were split and seeded into12-well plates at a density of 2 � 104 to 2.5 � 104 cells per well 24 h prior to
TABLE 1. Primers used for cloning
Primer Orientationa Sequenceb Description
pCMVmyc-SUMO1 F ATCCCC�AAGCTT�ATGTCTGACCAGGAGGCA HindIIIR ATTCGC�GGATCC��CTA�ACCCCCCGTTTGTTCCTG BamHI; stop codon
pCMVmyc-SUMO2 F AATCCG�GAATTC�ATGGCCGACGAAAAGCCCAA EcoRIR TAACGC�GGATCC��TCA�ACCTCCCGTCTGCTGTTGG BamHI; stop codon
pCMVmyc-SUMO3 F AATCCG�GAATTC�ATGTCCGAGGAGAAGCCC EcoRIR AATCGC�GGATCC��CTA�ACCTCCCGTCTGCTGCTG BamHI
pCMV myc-Ufm1 F CACCCG�GAATTC�ATGTCGAAGGTTTCCTTTAAG EcoRIR ATTCGC�GGATCC�TTAACAACTTCCAACACGATC BamHI
pGEX4T-1 N-CTCF(amino acids 1–266)
F TACCG�GAATTC�ATGGAAGGTGAGGCGGTTG EcoRIR TATCCG�CTCGAG�CTGGAATGTTTTCTTTACACC XhoI
pGEX4T-1 C-CTCF(amino acids 570–736)
F TTACGC�GGATCC�ATGGCAAGACATGCAGATAACTG BamHIR TATCCG�CTCGAG�TCACCGGTCCATCATGCTGAG XhoI
a F, forward; R, reverse.b Brackets indicate restriction enzyme sites and stop codons engineered into the primers.
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 715
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

transfection. The cells were transfected using the calcium phosphate-mediatedtransfection technique. The c-myc P2 promoter-luciferase reporter construct,pGL2XNM-Luc, was the generous gift of L. Penn (23). The c-myc P2 minimalpromoter contains 142 bp of DNA, which includes a CTCF binding site at its 3�end (26). For a typical transfection, 0.5 �g of the reporter plasmid DNA wasmixed with different amounts of wild-type or mutant CTCF expression vectorsplus 0.25 ng of pRL-TK plasmid DNA, which contains the Renilla luciferase geneused as a control for transfection efficiency. The total amount of DNA was keptconstant by the addition of pUC18 or pBR322 DNA. The assays were performedas described previously (20). To assess the transcriptional activity of the c-myc P2promoter, the firefly luciferase activity was normalized to the Renilla luciferaseactivity. Each transfection was performed in triplicate, and the experiment wasrepeated with at least three different DNA preparations. DNA was preparedusing the Qiagen Plasmid Midi kit. DNA concentrations were measured by UVspectrophotometry and confirmed by agarose gel electrophoresis.
In vitro transcription/translation of CTCF. Full-length CTCF was prepared byin vitro transcription/translation as described previously (19). A translation re-action in the absence of CTCF mRNA was used as a negative (unprogrammed)control. To make 35S-labeled CTCF, we used a translation kit depleted ofmethionine (Ambion) and supplemented the reaction with L-[35S]methionine(Amersham); 80 �Ci was used per 100-�l reaction mixture.
Electrophoretic mobility shift assay. Assays of in vitro DNA binding werecarried out as described previously (19). The coordinates of the 32P-labeled PCRfragments used for binding are listed in Table 2.
RESULTS
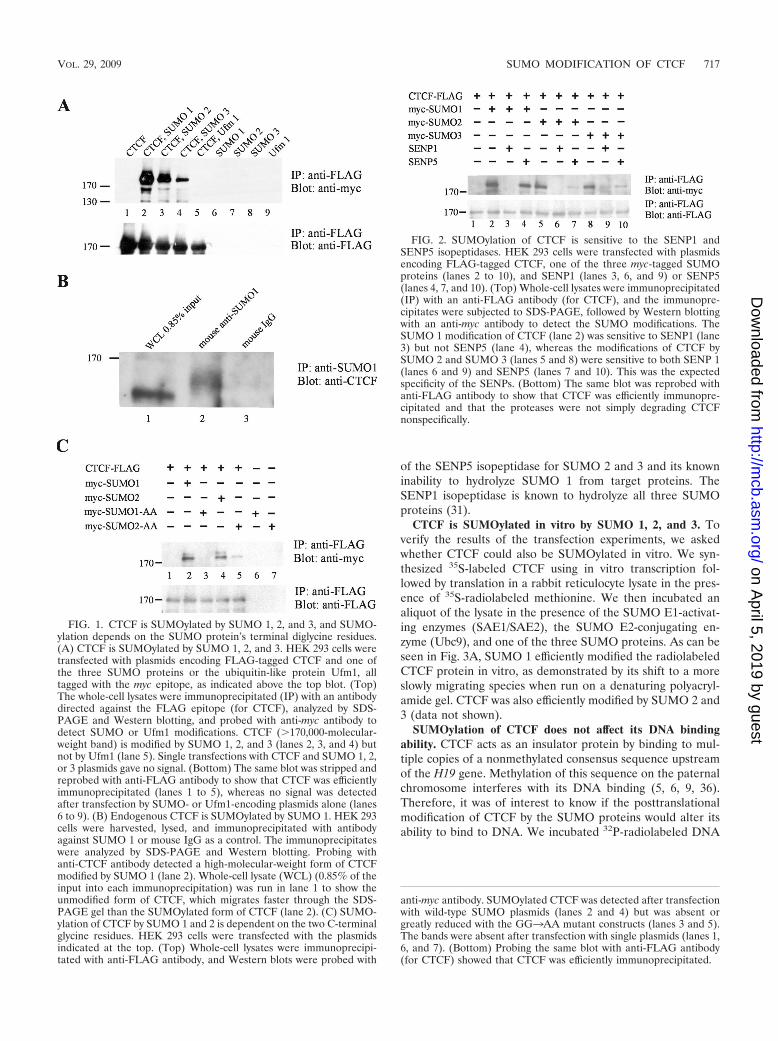
CTCF is modified by SUMO 1, 2, and 3. To show that CTCFis posttranslationally modified by the SUMO proteins, we tran-siently transfected HEK 293 cells with vectors expressing boththree-FLAG-tagged mouse CTCF and one of the three myc-tagged SUMO proteins. Cells were lysed under denaturingconditions, and CTCF was immunoprecipitated using an anti-body against the FLAG epitope. The immunoprecipitate wasrun on a denaturing polyacrylamide gel and electroblottedonto a PVDF membrane. Probing the membrane with an an-tibody against the myc epitope to detect SUMO-modified pro-teins revealed that CTCF was modified by each of the threeSUMO 1, 2, and 3 proteins (Fig. 1A, lanes 2, 3, and 4).SUMOylated CTCF runs at a position corresponding to amolecular mass slightly above 170 kDa. Transfection of cellswith FLAG-tagged CTCF or myc-tagged SUMO proteinsalone did not result in a 170-kDa band (Fig. 1A, lanes 1, 6, 7,and 8). The reciprocal experiment, immunoprecipitation withanti-myc antibody for the SUMO proteins and Western blot-ting for FLAG-tagged CTCF, also showed that CTCF isSUMOylated by all three SUMO proteins (data not shown).To demonstrate that the modification of CTCF is specific tothe SUMO proteins, we used a vector containing myc-taggedUfm1, along with FLAG-CTCF. Ufm1 does not modify CTCF(Fig. 1A, lane 5). To show that the tag itself was not SUMO-
ylated, the same three-FLAG-tag vector encoding FLAG-tagged E. coli beta-galactosidase was cotransfected with theSUMO proteins; LacZ was not modified by the SUMO pro-teins (data not shown). To demonstrate that endogenousCTCF is modified by SUMO 1, HEK 293 cells were harvestedand lysed under denaturing conditions. The lysate was immu-noprecipitated using an anti-SUMO 1 antibody and mouse IgGas a control. The immunoprecipitate was run on a denaturingpolyacrylamide gel and electroblotted onto a PVDF mem-brane. Probing the membrane with an antibody against CTCFrevealed a high-molecular-weight band of SUMOylated CTCFprotein (Fig. 1B, lane 2) that was absent when mouse IgG wasused (lane 3). We performed the reciprocal experiment, but wewere unable to detect SUMOylated CTCF when immunopre-cipitation was done with anti-CTCF antibody due to a lack ofsensitivity in Western blotting with anti-SUMO 1 (data notshown).
The SUMOylation reaction requires the presence of aCOOH-terminal diglycine sequence on the SUMO moleculefor the covalent attachment of the SUMO to the side chain ofthe target lysine residue. When we mutated the terminal dig-lycine of SUMO 1 and 2 to dialanine (SUMO 1-AA andSUMO 2-AA), there was a reduction of the SUMOylation ofCTCF, thus demonstrating that the modification of CTCF bythe SUMO proteins is due to the formation of a covalent bondthrough diglycine (Fig. 1C, lanes 2 versus 3, and 4 versus 5).Residual SUMOylation of CTCF by SUMO 2-AA may be dueto the noncovalent interaction of CTCF and SUMO 2.
SUMOylation of CTCF is sensitive to the presence ofSENP1 and SENP5 isopeptidases. SENPs are isopeptidasesthat remove SUMO proteins from their target proteins bycleaving the isopeptide bond between the SUMO and thetarget lysine residue. These enzymes play a critical role inregulating the amount of SUMOylated target protein in thecell. To gain insight into the specificity of the SUMOylation ofCTCF, we cotransfected 293 cells with plasmids that encodedeither the SENP1 or the SENP5 protease (generous gifts fromE. T. Yeh), in addition to those encoding FLAG-tagged CTCFand one of the three myc-tagged SUMO proteins. The over-expression of SENP1 reduces the SUMOylation of CTCF bySUMO 1 (Fig. 2, lane 3), whereas the posttranslational mod-ification of CTCF by SUMO 1 is insensitive to the overexpres-sion of SENP5 (Fig. 2, lane 4). The SUMOylation of CTCF byeither SUMO 2 or SUMO 3 was sensitive to the presence ofboth the SENP1 and SENP5 isopeptidases (Fig. 2, lanes 6, 7, 9,and 10). These results are consistent with the known specificity
TABLE 2. Primers used to amplify DNA fragments used in the electrophoretic mobility shift assays
Primer Accession no. Nucleotidesamplified Orientationa Sequence
H19-B1 AF125183 7893–8079 F GAGGCTTCTCCTTCGGTCTCR AGATGACCCCCGTGAACC
c-myc 5� insulator D10493 292–468 F CCTCGGACGCTCCTGCTCR TCGCTATGCTGGATTTTGCTG
c-myc P2 promoter D10493 2446–2610 F GATCGCGCTGAGTATAAAAGCR CCTATTCGCTCCGGATCTC
Chicken �-globin insulator FII AY040835 20–175 F CCCAAAGCCCCCAGGGATGTR AAGCGATCCCGTGCCACCTT
a F, forward; R, reverse.
716 MACPHERSON ET AL. MOL. CELL. BIOL.
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
of the SENP5 isopeptidase for SUMO 2 and 3 and its knowninability to hydrolyze SUMO 1 from target proteins. TheSENP1 isopeptidase is known to hydrolyze all three SUMOproteins (31).
CTCF is SUMOylated in vitro by SUMO 1, 2, and 3. Toverify the results of the transfection experiments, we askedwhether CTCF could also be SUMOylated in vitro. We syn-thesized 35S-labeled CTCF using in vitro transcription fol-lowed by translation in a rabbit reticulocyte lysate in the pres-ence of 35S-radiolabeled methionine. We then incubated analiquot of the lysate in the presence of the SUMO E1-activat-ing enzymes (SAE1/SAE2), the SUMO E2-conjugating en-zyme (Ubc9), and one of the three SUMO proteins. As can beseen in Fig. 3A, SUMO 1 efficiently modified the radiolabeledCTCF protein in vitro, as demonstrated by its shift to a moreslowly migrating species when run on a denaturing polyacryl-amide gel. CTCF was also efficiently modified by SUMO 2 and3 (data not shown).
SUMOylation of CTCF does not affect its DNA bindingability. CTCF acts as an insulator protein by binding to mul-tiple copies of a nonmethylated consensus sequence upstreamof the H19 gene. Methylation of this sequence on the paternalchromosome interferes with its DNA binding (5, 6, 9, 36).Therefore, it was of interest to know if the posttranslationalmodification of CTCF by the SUMO proteins would alter itsability to bind to DNA. We incubated 32P-radiolabeled DNA
FIG. 1. CTCF is SUMOylated by SUMO 1, 2, and 3, and SUMO-ylation depends on the SUMO protein’s terminal diglycine residues.(A) CTCF is SUMOylated by SUMO 1, 2, and 3. HEK 293 cells weretransfected with plasmids encoding FLAG-tagged CTCF and one ofthe three SUMO proteins or the ubiquitin-like protein Ufm1, alltagged with the myc epitope, as indicated above the top blot. (Top)The whole-cell lysates were immunoprecipitated (IP) with an antibodydirected against the FLAG epitope (for CTCF), analyzed by SDS-PAGE and Western blotting, and probed with anti-myc antibody todetect SUMO or Ufm1 modifications. CTCF (�170,000-molecular-weight band) is modified by SUMO 1, 2, and 3 (lanes 2, 3, and 4) butnot by Ufm1 (lane 5). Single transfections with CTCF and SUMO 1, 2,or 3 plasmids gave no signal. (Bottom) The same blot was stripped andreprobed with anti-FLAG antibody to show that CTCF was efficientlyimmunoprecipitated (lanes 1 to 5), whereas no signal was detectedafter transfection by SUMO- or Ufm1-encoding plasmids alone (lanes6 to 9). (B) Endogenous CTCF is SUMOylated by SUMO 1. HEK 293cells were harvested, lysed, and immunoprecipitated with antibodyagainst SUMO 1 or mouse IgG as a control. The immunoprecipitateswere analyzed by SDS-PAGE and Western blotting. Probing withanti-CTCF antibody detected a high-molecular-weight form of CTCFmodified by SUMO 1 (lane 2). Whole-cell lysate (WCL) (0.85% of theinput into each immunoprecipitation) was run in lane 1 to show theunmodified form of CTCF, which migrates faster through the SDS-PAGE gel than the SUMOylated form of CTCF (lane 2). (C) SUMO-ylation of CTCF by SUMO 1 and 2 is dependent on the two C-terminalglycine residues. HEK 293 cells were transfected with the plasmidsindicated at the top. (Top) Whole-cell lysates were immunoprecipi-tated with anti-FLAG antibody, and Western blots were probed with
anti-myc antibody. SUMOylated CTCF was detected after transfectionwith wild-type SUMO plasmids (lanes 2 and 4) but was absent orgreatly reduced with the GG3AA mutant constructs (lanes 3 and 5).The bands were absent after transfection with single plasmids (lanes 1,6, and 7). (Bottom) Probing the same blot with anti-FLAG antibody(for CTCF) showed that CTCF was efficiently immunoprecipitated.
FIG. 2. SUMOylation of CTCF is sensitive to the SENP1 andSENP5 isopeptidases. HEK 293 cells were transfected with plasmidsencoding FLAG-tagged CTCF, one of the three myc-tagged SUMOproteins (lanes 2 to 10), and SENP1 (lanes 3, 6, and 9) or SENP5(lanes 4, 7, and 10). (Top) Whole-cell lysates were immunoprecipitated(IP) with an anti-FLAG antibody (for CTCF), and the immunopre-cipitates were subjected to SDS-PAGE, followed by Western blottingwith an anti-myc antibody to detect the SUMO modifications. TheSUMO 1 modification of CTCF (lane 2) was sensitive to SENP1 (lane3) but not SENP5 (lane 4), whereas the modifications of CTCF bySUMO 2 and SUMO 3 (lanes 5 and 8) were sensitive to both SENP 1(lanes 6 and 9) and SENP5 (lanes 7 and 10). This was the expectedspecificity of the SENPs. (Bottom) The same blot was reprobed withanti-FLAG antibody to show that CTCF was efficiently immunopre-cipitated and that the proteases were not simply degrading CTCFnonspecifically.
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 717
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
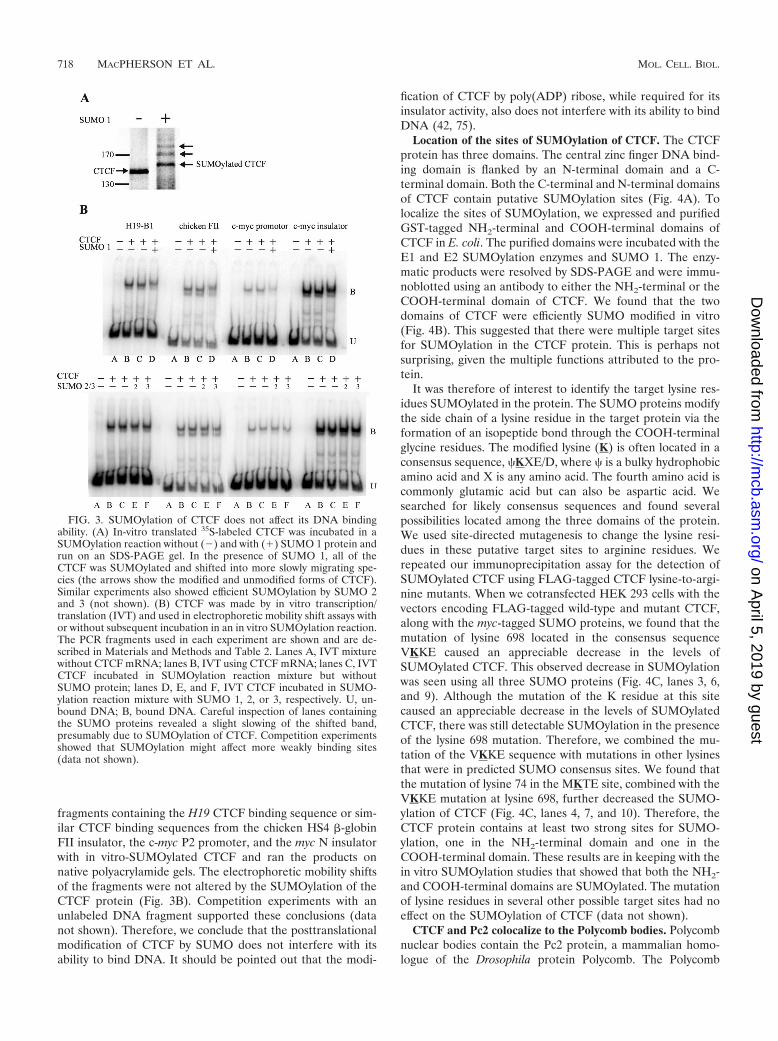
fragments containing the H19 CTCF binding sequence or sim-ilar CTCF binding sequences from the chicken HS4 �-globinFII insulator, the c-myc P2 promoter, and the myc N insulatorwith in vitro-SUMOylated CTCF and ran the products onnative polyacrylamide gels. The electrophoretic mobility shiftsof the fragments were not altered by the SUMOylation of theCTCF protein (Fig. 3B). Competition experiments with anunlabeled DNA fragment supported these conclusions (datanot shown). Therefore, we conclude that the posttranslationalmodification of CTCF by SUMO does not interfere with itsability to bind DNA. It should be pointed out that the modi-
fication of CTCF by poly(ADP) ribose, while required for itsinsulator activity, also does not interfere with its ability to bindDNA (42, 75).
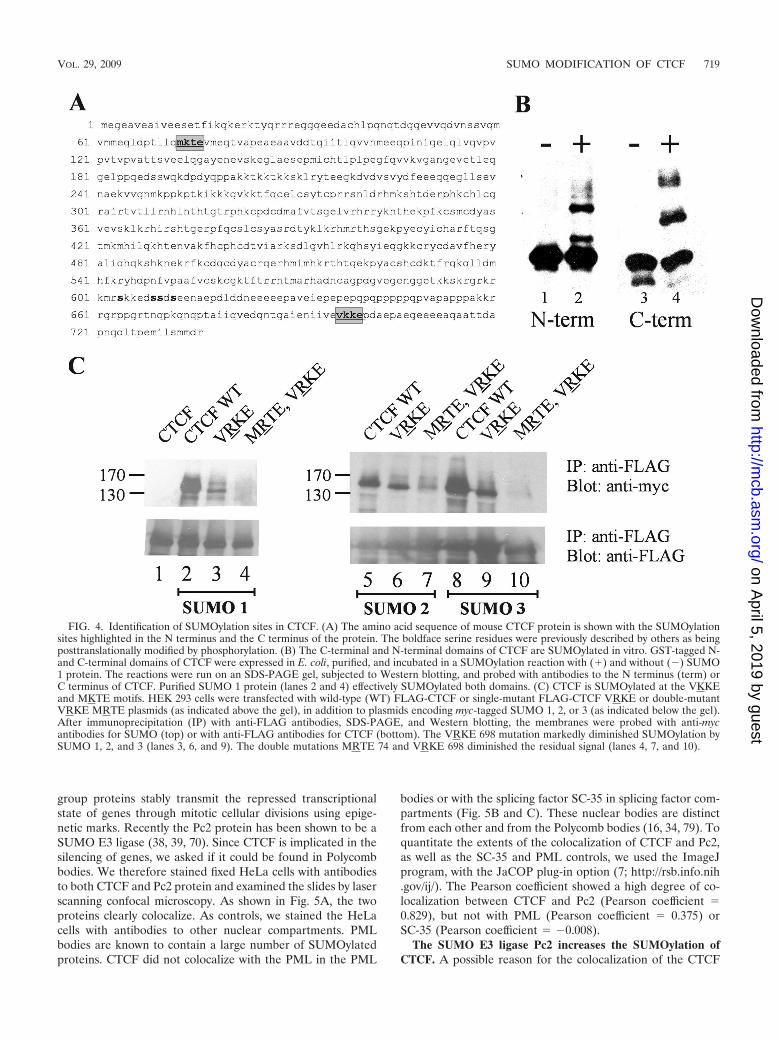
Location of the sites of SUMOylation of CTCF. The CTCFprotein has three domains. The central zinc finger DNA bind-ing domain is flanked by an N-terminal domain and a C-terminal domain. Both the C-terminal and N-terminal domainsof CTCF contain putative SUMOylation sites (Fig. 4A). Tolocalize the sites of SUMOylation, we expressed and purifiedGST-tagged NH2-terminal and COOH-terminal domains ofCTCF in E. coli. The purified domains were incubated with theE1 and E2 SUMOylation enzymes and SUMO 1. The enzy-matic products were resolved by SDS-PAGE and were immu-noblotted using an antibody to either the NH2-terminal or theCOOH-terminal domain of CTCF. We found that the twodomains of CTCF were efficiently SUMO modified in vitro(Fig. 4B). This suggested that there were multiple target sitesfor SUMOylation in the CTCF protein. This is perhaps notsurprising, given the multiple functions attributed to the pro-tein.
It was therefore of interest to identify the target lysine res-idues SUMOylated in the protein. The SUMO proteins modifythe side chain of a lysine residue in the target protein via theformation of an isopeptide bond through the COOH-terminalglycine residues. The modified lysine (K) is often located in aconsensus sequence, KXE/D, where is a bulky hydrophobicamino acid and X is any amino acid. The fourth amino acid iscommonly glutamic acid but can also be aspartic acid. Wesearched for likely consensus sequences and found severalpossibilities located among the three domains of the protein.We used site-directed mutagenesis to change the lysine resi-dues in these putative target sites to arginine residues. Werepeated our immunoprecipitation assay for the detection ofSUMOylated CTCF using FLAG-tagged CTCF lysine-to-argi-nine mutants. When we cotransfected HEK 293 cells with thevectors encoding FLAG-tagged wild-type and mutant CTCF,along with the myc-tagged SUMO proteins, we found that themutation of lysine 698 located in the consensus sequenceVKKE caused an appreciable decrease in the levels ofSUMOylated CTCF. This observed decrease in SUMOylationwas seen using all three SUMO proteins (Fig. 4C, lanes 3, 6,and 9). Although the mutation of the K residue at this sitecaused an appreciable decrease in the levels of SUMOylatedCTCF, there was still detectable SUMOylation in the presenceof the lysine 698 mutation. Therefore, we combined the mu-tation of the VKKE sequence with mutations in other lysinesthat were in predicted SUMO consensus sites. We found thatthe mutation of lysine 74 in the MKTE site, combined with theVKKE mutation at lysine 698, further decreased the SUMO-ylation of CTCF (Fig. 4C, lanes 4, 7, and 10). Therefore, theCTCF protein contains at least two strong sites for SUMO-ylation, one in the NH2-terminal domain and one in theCOOH-terminal domain. These results are in keeping with thein vitro SUMOylation studies that showed that both the NH2-and COOH-terminal domains are SUMOylated. The mutationof lysine residues in several other possible target sites had noeffect on the SUMOylation of CTCF (data not shown).
CTCF and Pc2 colocalize to the Polycomb bodies. Polycombnuclear bodies contain the Pc2 protein, a mammalian homo-logue of the Drosophila protein Polycomb. The Polycomb
FIG. 3. SUMOylation of CTCF does not affect its DNA bindingability. (A) In-vitro translated 35S-labeled CTCF was incubated in aSUMOylation reaction without () and with (�) SUMO 1 protein andrun on an SDS-PAGE gel. In the presence of SUMO 1, all of theCTCF was SUMOylated and shifted into more slowly migrating spe-cies (the arrows show the modified and unmodified forms of CTCF).Similar experiments also showed efficient SUMOylation by SUMO 2and 3 (not shown). (B) CTCF was made by in vitro transcription/translation (IVT) and used in electrophoretic mobility shift assays withor without subsequent incubation in an in vitro SUMOylation reaction.The PCR fragments used in each experiment are shown and are de-scribed in Materials and Methods and Table 2. Lanes A, IVT mixturewithout CTCF mRNA; lanes B, IVT using CTCF mRNA; lanes C, IVTCTCF incubated in SUMOylation reaction mixture but withoutSUMO protein; lanes D, E, and F, IVT CTCF incubated in SUMO-ylation reaction mixture with SUMO 1, 2, or 3, respectively. U, un-bound DNA; B, bound DNA. Careful inspection of lanes containingthe SUMO proteins revealed a slight slowing of the shifted band,presumably due to SUMOylation of CTCF. Competition experimentsshowed that SUMOylation might affect more weakly binding sites(data not shown).
718 MACPHERSON ET AL. MOL. CELL. BIOL.
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
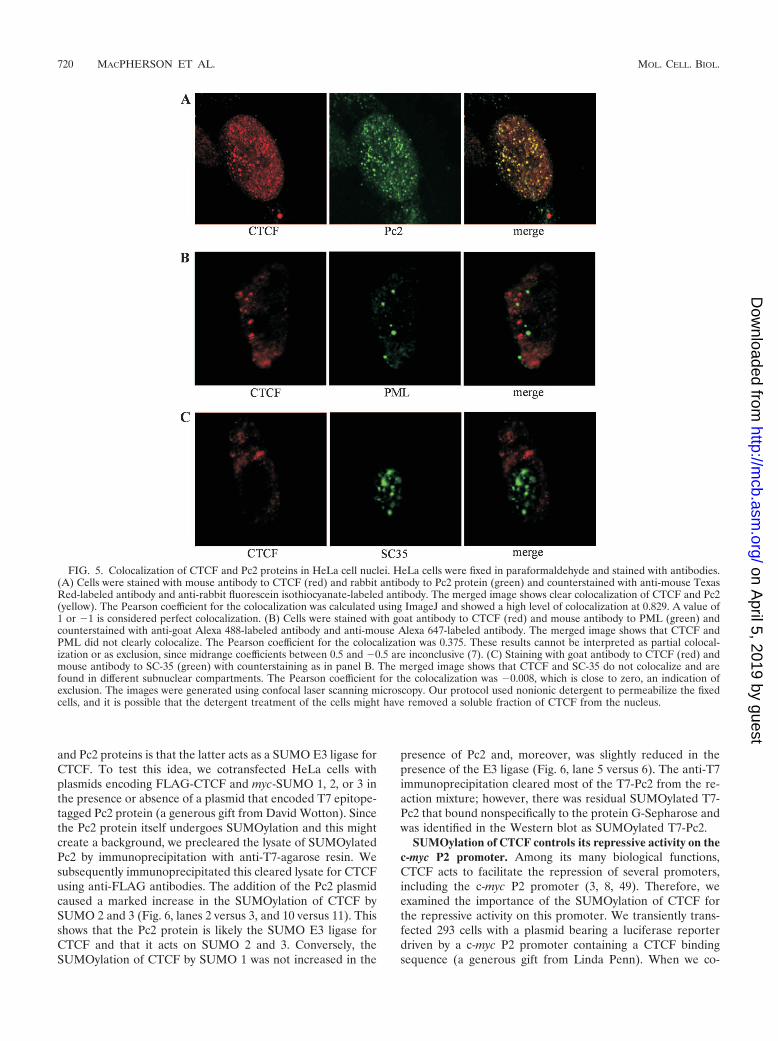
group proteins stably transmit the repressed transcriptionalstate of genes through mitotic cellular divisions using epige-netic marks. Recently the Pc2 protein has been shown to be aSUMO E3 ligase (38, 39, 70). Since CTCF is implicated in thesilencing of genes, we asked if it could be found in Polycombbodies. We therefore stained fixed HeLa cells with antibodiesto both CTCF and Pc2 protein and examined the slides by laserscanning confocal microscopy. As shown in Fig. 5A, the twoproteins clearly colocalize. As controls, we stained the HeLacells with antibodies to other nuclear compartments. PMLbodies are known to contain a large number of SUMOylatedproteins. CTCF did not colocalize with the PML in the PML
bodies or with the splicing factor SC-35 in splicing factor com-partments (Fig. 5B and C). These nuclear bodies are distinctfrom each other and from the Polycomb bodies (16, 34, 79). Toquantitate the extents of the colocalization of CTCF and Pc2,as well as the SC-35 and PML controls, we used the ImageJprogram, with the JaCOP plug-in option (7; http://rsb.info.nih.gov/ij/). The Pearson coefficient showed a high degree of co-localization between CTCF and Pc2 (Pearson coefficient �0.829), but not with PML (Pearson coefficient � 0.375) orSC-35 (Pearson coefficient � 0.008).
The SUMO E3 ligase Pc2 increases the SUMOylation ofCTCF. A possible reason for the colocalization of the CTCF
FIG. 4. Identification of SUMOylation sites in CTCF. (A) The amino acid sequence of mouse CTCF protein is shown with the SUMOylationsites highlighted in the N terminus and the C terminus of the protein. The boldface serine residues were previously described by others as beingposttranslationally modified by phosphorylation. (B) The C-terminal and N-terminal domains of CTCF are SUMOylated in vitro. GST-tagged N-and C-terminal domains of CTCF were expressed in E. coli, purified, and incubated in a SUMOylation reaction with (�) and without () SUMO1 protein. The reactions were run on an SDS-PAGE gel, subjected to Western blotting, and probed with antibodies to the N terminus (term) orC terminus of CTCF. Purified SUMO 1 protein (lanes 2 and 4) effectively SUMOylated both domains. (C) CTCF is SUMOylated at the VKKEand MKTE motifs. HEK 293 cells were transfected with wild-type (WT) FLAG-CTCF or single-mutant FLAG-CTCF VRKE or double-mutantVRKE MRTE plasmids (as indicated above the gel), in addition to plasmids encoding myc-tagged SUMO 1, 2, or 3 (as indicated below the gel).After immunoprecipitation (IP) with anti-FLAG antibodies, SDS-PAGE, and Western blotting, the membranes were probed with anti-mycantibodies for SUMO (top) or with anti-FLAG antibodies for CTCF (bottom). The VRKE 698 mutation markedly diminished SUMOylation bySUMO 1, 2, and 3 (lanes 3, 6, and 9). The double mutations MRTE 74 and VRKE 698 diminished the residual signal (lanes 4, 7, and 10).
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 719
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
and Pc2 proteins is that the latter acts as a SUMO E3 ligase forCTCF. To test this idea, we cotransfected HeLa cells withplasmids encoding FLAG-CTCF and myc-SUMO 1, 2, or 3 inthe presence or absence of a plasmid that encoded T7 epitope-tagged Pc2 protein (a generous gift from David Wotton). Sincethe Pc2 protein itself undergoes SUMOylation and this mightcreate a background, we precleared the lysate of SUMOylatedPc2 by immunoprecipitation with anti-T7-agarose resin. Wesubsequently immunoprecipitated this cleared lysate for CTCFusing anti-FLAG antibodies. The addition of the Pc2 plasmidcaused a marked increase in the SUMOylation of CTCF bySUMO 2 and 3 (Fig. 6, lanes 2 versus 3, and 10 versus 11). Thisshows that the Pc2 protein is likely the SUMO E3 ligase forCTCF and that it acts on SUMO 2 and 3. Conversely, theSUMOylation of CTCF by SUMO 1 was not increased in the
presence of Pc2 and, moreover, was slightly reduced in thepresence of the E3 ligase (Fig. 6, lane 5 versus 6). The anti-T7immunoprecipitation cleared most of the T7-Pc2 from the re-action mixture; however, there was residual SUMOylated T7-Pc2 that bound nonspecifically to the protein G-Sepharose andwas identified in the Western blot as SUMOylated T7-Pc2.
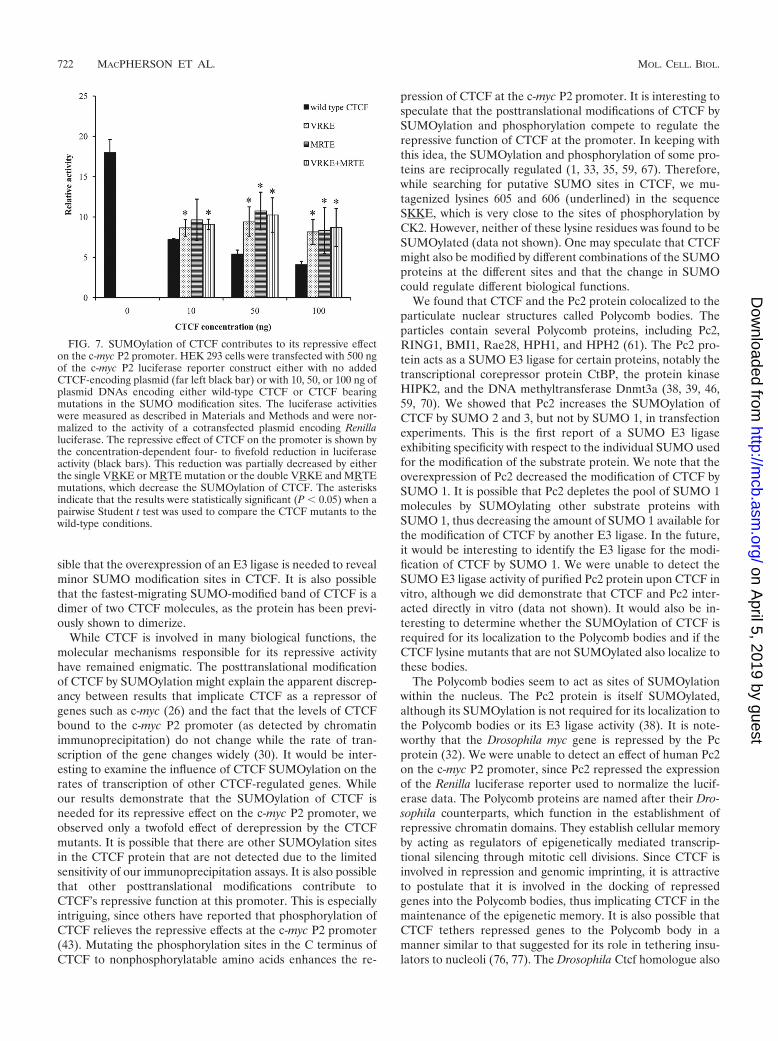
SUMOylation of CTCF controls its repressive activity on thec-myc P2 promoter. Among its many biological functions,CTCF acts to facilitate the repression of several promoters,including the c-myc P2 promoter (3, 8, 49). Therefore, weexamined the importance of the SUMOylation of CTCF forthe repressive activity on this promoter. We transiently trans-fected 293 cells with a plasmid bearing a luciferase reporterdriven by a c-myc P2 promoter containing a CTCF bindingsequence (a generous gift from Linda Penn). When we co-
FIG. 5. Colocalization of CTCF and Pc2 proteins in HeLa cell nuclei. HeLa cells were fixed in paraformaldehyde and stained with antibodies.(A) Cells were stained with mouse antibody to CTCF (red) and rabbit antibody to Pc2 protein (green) and counterstained with anti-mouse TexasRed-labeled antibody and anti-rabbit fluorescein isothiocyanate-labeled antibody. The merged image shows clear colocalization of CTCF and Pc2(yellow). The Pearson coefficient for the colocalization was calculated using ImageJ and showed a high level of colocalization at 0.829. A value of1 or 1 is considered perfect colocalization. (B) Cells were stained with goat antibody to CTCF (red) and mouse antibody to PML (green) andcounterstained with anti-goat Alexa 488-labeled antibody and anti-mouse Alexa 647-labeled antibody. The merged image shows that CTCF andPML did not clearly colocalize. The Pearson coefficient for the colocalization was 0.375. These results cannot be interpreted as partial colocal-ization or as exclusion, since midrange coefficients between 0.5 and 0.5 are inconclusive (7). (C) Staining with goat antibody to CTCF (red) andmouse antibody to SC-35 (green) with counterstaining as in panel B. The merged image shows that CTCF and SC-35 do not colocalize and arefound in different subnuclear compartments. The Pearson coefficient for the colocalization was 0.008, which is close to zero, an indication ofexclusion. The images were generated using confocal laser scanning microscopy. Our protocol used nonionic detergent to permeabilize the fixedcells, and it is possible that the detergent treatment of the cells might have removed a soluble fraction of CTCF from the nucleus.
720 MACPHERSON ET AL. MOL. CELL. BIOL.
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

transfected cells with the reporter plasmid and a plasmid thatexpressed a wild-type version of CTCF, there was a statisticallysignificant and reproducible decrease of promoter activity de-pendent upon the exogenous source of CTCF (Fig. 7). How-ever, when we substituted a plasmid in which the MKTE andthe VKKE SUMOylation sites were mutated singly or in com-bination, the repressive effect was decreased. We show that theSUMOylation of CTCF contributes to its repressive activity onthe c-myc P2 promoter.
DISCUSSION
In this paper, we report that the multifunctional zinc fingerprotein CTCF is posttranslationally modified by each of thethree SUMO proteins. It had been previously reported thatCTCF is modified by phosphorylation and is also poly(ADP)ribosylated (22, 42, 43, 75). The phosphorylation of CTCFrelieves its repressive effects at the c-myc P2 promoter, whilepoly(ADP) ribosylation has been linked to CTCF’s insulatoractivity. Our findings augment the role of posttranslational
modifications in the regulation of CTCF’s many functions byshowing that SUMOylation is involved in the repressive activ-ity of CTCF.
We have identified two major sites of SUMOylation. TheCOOH-terminal VKKE sequence is the principal site, whereasmodification of the NH2-terminal MKTE site is detectableonly when the VKKE site has been mutated. The observationthat the repressive effect on the c-myc P2 promoter occurs bymutation of a single CTCF SUMO site suggests that theSUMOylation of either site is sufficient to produce the repres-sive effect. This is in keeping with findings of repressive do-mains in both the N and C termini of the protein (13, 17, 55).Both sites are modified by all three SUMO proteins.
Unlike polyubiquitin, which targets proteins for proteolyticdegradation by the 26S proteasome, SUMO modifications invivo are usually not polymeric. Our experiments show that themodification of CTCF results in several CTCF SUMO-modi-fied species that migrate as multiple high-molecular-weightbands during denaturing PAGE. These higher-molecular-weight species are enriched in the presence of Pc2. It is pos-
FIG. 6. The SUMO E3 ligase Pc2 increases the SUMOylation of CTCF. HeLa cells were transfected using Fugene 6 transfection reagent withthe indicated plasmids. The whole-cell lysates were precleared with anti-T7-agarose resin prior to immunoprecipitation (IP) with anti-FLAGantibodies. After SDS-PAGE and Western blotting, the membrane was probed with anti-myc antibody to detect SUMO 1, 2, or 3 (top) or withanti-FLAG antibody to detect CTCF (middle). SUMO E3 ligase activity was indicated by the increase in the SUMOylation signal in lanes 3 and11. Pc2 exhibited no SUMO E3 ligase activity for the SUMOylation of CTCF by SUMO 1 (lane 6). CTCF degradation products were alsoSUMOylated by SUMO 2 and 3 in the presence of Pc2. Flowthrough from the immunoprecipitation was blotted for �-actin to show equal loading(bottom).
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 721
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
sible that the overexpression of an E3 ligase is needed to revealminor SUMO modification sites in CTCF. It is also possiblethat the fastest-migrating SUMO-modified band of CTCF is adimer of two CTCF molecules, as the protein has been previ-ously shown to dimerize.
While CTCF is involved in many biological functions, themolecular mechanisms responsible for its repressive activityhave remained enigmatic. The posttranslational modificationof CTCF by SUMOylation might explain the apparent discrep-ancy between results that implicate CTCF as a repressor ofgenes such as c-myc (26) and the fact that the levels of CTCFbound to the c-myc P2 promoter (as detected by chromatinimmunoprecipitation) do not change while the rate of tran-scription of the gene changes widely (30). It would be inter-esting to examine the influence of CTCF SUMOylation on therates of transcription of other CTCF-regulated genes. Whileour results demonstrate that the SUMOylation of CTCF isneeded for its repressive effect on the c-myc P2 promoter, weobserved only a twofold effect of derepression by the CTCFmutants. It is possible that there are other SUMOylation sitesin the CTCF protein that are not detected due to the limitedsensitivity of our immunoprecipitation assays. It is also possiblethat other posttranslational modifications contribute toCTCF’s repressive function at this promoter. This is especiallyintriguing, since others have reported that phosphorylation ofCTCF relieves the repressive effects at the c-myc P2 promoter(43). Mutating the phosphorylation sites in the C terminus ofCTCF to nonphosphorylatable amino acids enhances the re-
pression of CTCF at the c-myc P2 promoter. It is interesting tospeculate that the posttranslational modifications of CTCF bySUMOylation and phosphorylation compete to regulate therepressive function of CTCF at the promoter. In keeping withthis idea, the SUMOylation and phosphorylation of some pro-teins are reciprocally regulated (1, 33, 35, 59, 67). Therefore,while searching for putative SUMO sites in CTCF, we mu-tagenized lysines 605 and 606 (underlined) in the sequenceSKKE, which is very close to the sites of phosphorylation byCK2. However, neither of these lysine residues was found to beSUMOylated (data not shown). One may speculate that CTCFmight also be modified by different combinations of the SUMOproteins at the different sites and that the change in SUMOcould regulate different biological functions.
We found that CTCF and the Pc2 protein colocalized to theparticulate nuclear structures called Polycomb bodies. Theparticles contain several Polycomb proteins, including Pc2,RING1, BMI1, Rae28, HPH1, and HPH2 (61). The Pc2 pro-tein acts as a SUMO E3 ligase for certain proteins, notably thetranscriptional corepressor protein CtBP, the protein kinaseHIPK2, and the DNA methyltransferase Dnmt3a (38, 39, 46,59, 70). We showed that Pc2 increases the SUMOylation ofCTCF by SUMO 2 and 3, but not by SUMO 1, in transfectionexperiments. This is the first report of a SUMO E3 ligaseexhibiting specificity with respect to the individual SUMO usedfor the modification of the substrate protein. We note that theoverexpression of Pc2 decreased the modification of CTCF bySUMO 1. It is possible that Pc2 depletes the pool of SUMO 1molecules by SUMOylating other substrate proteins withSUMO 1, thus decreasing the amount of SUMO 1 available forthe modification of CTCF by another E3 ligase. In the future,it would be interesting to identify the E3 ligase for the modi-fication of CTCF by SUMO 1. We were unable to detect theSUMO E3 ligase activity of purified Pc2 protein upon CTCF invitro, although we did demonstrate that CTCF and Pc2 inter-acted directly in vitro (data not shown). It would also be in-teresting to determine whether the SUMOylation of CTCF isrequired for its localization to the Polycomb bodies and if theCTCF lysine mutants that are not SUMOylated also localize tothese bodies.
The Polycomb bodies seem to act as sites of SUMOylationwithin the nucleus. The Pc2 protein is itself SUMOylated,although its SUMOylation is not required for its localization tothe Polycomb bodies or its E3 ligase activity (38). It is note-worthy that the Drosophila myc gene is repressed by the Pcprotein (32). We were unable to detect an effect of human Pc2on the c-myc P2 promoter, since Pc2 repressed the expressionof the Renilla luciferase reporter used to normalize the lucif-erase data. The Polycomb proteins are named after their Dro-sophila counterparts, which function in the establishment ofrepressive chromatin domains. They establish cellular memoryby acting as regulators of epigenetically mediated transcrip-tional silencing through mitotic cell divisions. Since CTCF isinvolved in repression and genomic imprinting, it is attractiveto postulate that it is involved in the docking of repressedgenes into the Polycomb bodies, thus implicating CTCF in themaintenance of the epigenetic memory. It is also possible thatCTCF tethers repressed genes to the Polycomb body in amanner similar to that suggested for its role in tethering insu-lators to nucleoli (76, 77). The Drosophila Ctcf homologue also
FIG. 7. SUMOylation of CTCF contributes to its repressive effecton the c-myc P2 promoter. HEK 293 cells were transfected with 500 ngof the c-myc P2 luciferase reporter construct either with no addedCTCF-encoding plasmid (far left black bar) or with 10, 50, or 100 ng ofplasmid DNAs encoding either wild-type CTCF or CTCF bearingmutations in the SUMO modification sites. The luciferase activitieswere measured as described in Materials and Methods and were nor-malized to the activity of a cotransfected plasmid encoding Renillaluciferase. The repressive effect of CTCF on the promoter is shown bythe concentration-dependent four- to fivefold reduction in luciferaseactivity (black bars). This reduction was partially decreased by eitherthe single VRKE or MRTE mutation or the double VRKE and MRTEmutations, which decrease the SUMOylation of CTCF. The asterisksindicate that the results were statistically significant (P � 0.05) when apairwise Student t test was used to compare the CTCF mutants to thewild-type conditions.
722 MACPHERSON ET AL. MOL. CELL. BIOL.
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

acts as an insulator protein (15, 37, 51, 52, 68). A recentpublication has shown that the Drosophila Ctcf homologuemediates looping between two Fab-8 insulators and one of theAbd-B promoters. This looping is thought to involve enhancersthat activate transcription in the bithorax complex, overcomingthe repressive effects of the Polycomb proteins (45).
Numerous publications have also highlighted CTCF’s role inorganizing the nuclear architecture of mammalian cells. CTCFis thought to mediate its insulator function by organizing chro-matin into looped domains (47, 64) that occur even betweenchromosomes. A role for CTCF binding sites has been dem-onstrated in X chromosome inactivation (11, 54), and CTCFhas been implicated in the pairing between the two X inacti-vation centers that precedes the choice for X inactivation (18,73). Three recent publications have demonstrated another rolefor CTCF in nuclear architecture, showing that cohesins bindspecifically to CTCF binding sequences (57, 65, 69). It will beimportant to determine the molecular mechanisms responsiblefor CTCF’s role in genomic organization and the role that theSUMOylation of CTCF plays in these functions.
The CTCF protein has multiple functions in several eukary-otic organisms. Our observations considerably expand the rep-ertoire of posttranslational modifications that may play roles inregulating this multiplicity of functions. It is not knownwhether CTCF’s nuclear localization changes depending uponwhether it is acting as a repressor, activator, or insulator pro-tein. It is noteworthy that the SUMO sites we have identifiedin the mouse protein are conserved in vertebrates from fish tomammals. Although the actual SUMO sites in the vertebrateproteins are not conserved in the Drosophila protein, there areseveral high-probability putative SUMO consensus sites inboth its NH2 and COOH termini. Since SUMOylation hasrecently been found to attenuate the function of the gypsyinsulator in Drosophila (10) and since Drosophila Ctcf is in-volved in insulator function (45), it would be of interest tolearn whether the CTCF of flies is also SUMOylated. It wouldalso be valuable to learn the effects of SUMOylation on insu-lator activity in vertebrates.
In this paper, we have focused on the role of SUMOylationin CTCF function. However, it has not escaped our attentionthat basic residues, such as lysine and arginine, that are com-monly modified in histones can also be methylated and acety-lated and that such modifications can also affect the activityand stability of nonhistone proteins, for example, p53 (14, 48).Further studies of the roles of posttranslation modifications inthe molecular mechanisms of CTCF functions are warranted.
ACKNOWLEDGMENTS
This work was supported in part by funds from the Canadian Insti-tutes for Health Research. M.J.M. was the recipient of a University ofToronto Open Fellowship.
We thank E. T. Yeh, D. Wotton, L. Frappier, J. Greenblatt, and L.Penn for vectors. We thank L. Frappier and A. Sakwe for theirthoughtful advice, A. Cochrane for the SC-35 antibody, and HowardLipshitz and Marc Shulman for their interest and support. We thank L.Frappier for her careful reading of the manuscript.
REFERENCES
1. Anckar, J., and L. Sistonen. 2007. SUMO: getting it on. Biochem. Soc.Trans. 35:1409–1413.
2. Andersson, S., D. L. Davis, H. Dahlback, H. Jornvall, and D. W. Russell.1989. Cloning, structure, and expression of the mitochondrial cytochrome
P-450 sterol 26-hydroxylase, a bile acid biosynthetic enzyme. J. Biol. Chem.264:8222–8229.
3. Arnold, R., M. Burcin, B. Kaiser, M. Muller, and R. Renkawitz. 1996. DNAbending by the silencer protein NeP1 is modulated by TR and RXR. NucleicAcids Res. 24:2640–2647.
4. Barski, A., S. Cuddapah, K. Cui, T. Y. Roh, D. E. Schones, Z. Wang, G. Wei,I. Chepelev, and K. Zhao. 2007. High-resolution profiling of histone meth-ylations in the human genome. Cell 129:823–837.
5. Bell, A. C., and G. Felsenfeld. 2000. Methylation of a CTCF-dependentboundary controls imprinted expression of the Igf2 gene. Nature 405:482–485.
6. Bell, A. C., A. G. West, and G. Felsenfeld. 1999. The protein CTCF isrequired for the enhancer blocking activity of vertebrate insulators. Cell98:387–396.
7. Bolte, S., and F. P. Cordelieres. 2006. A guided tour into subcellular colo-calization analysis in light microscopy. J. Microsc. 224:213–232.
8. Burcin, M., R. Arnold, M. Lutz, B. Kaiser, D. Runge, F. Lottspeich, G. N.Filippova, V. V. Lobanenkov, and R. Renkawitz. 1997. Negative protein 1,which is required for function of the chicken lysozyme gene silencer inconjunction with hormone receptors, is identical to the multivalent zincfinger repressor CTCF. Mol. Cell. Biol. 17:1281–1288.
9. Burton, T., B. Liang, A. Dibrov, and F. Amara. 2002. Transforming growthfactor-beta-induced transcription of the Alzheimer beta-amyloid precursorprotein gene involves interaction between the CTCF-complex and Smads.Biochem. Biophys. Res. Commun. 295:713–723.
10. Capelson, M., and V. G. Corces. 2006. SUMO conjugation attenuates theactivity of the gypsy chromatin insulator. EMBO J. 25:1906–1914.
11. Chao, W., K. D. Huynh, R. J. Spencer, L. S. Davidow, and J. T. Lee. 2002.CTCF, a candidate trans-acting factor for X-inactivation choice. Science295:345–347.
12. Cheng, J., N. D. Perkins, and E. T. Yeh. 2005. Differential regulation ofc-Jun-dependent transcription by SUMO-specific proteases. J. Biol. Chem.280:14492–14498.
13. Chernukhin, I., S. Shamsuddin, S. Y. Kang, R. Bergstrom, Y. W. Kwon, W.Yu, J. Whitehead, R. Mukhopadhyay, F. Docquier, D. Farrar, I. Morrison,M. Vigneron, S. Y. Wu, C. M. Chiang, D. Loukinov, V. Lobanenkov, R.Ohlsson, and E. Klenova. 2007. CTCF interacts with and recruits the largestsubunit of RNA polymerase II to CTCF target sites genome-wide. Mol. Cell.Biol. 27:1631–1648.
14. Chuikov, S., J. K. Kurash, J. R. Wilson, B. Xiao, N. Justin, G. S. Ivanov, K.McKinney, P. Tempst, C. Prives, S. J. Gamblin, N. A. Barlev, and D. Rein-berg. 2004. Regulation of p53 activity through lysine methylation. Nature432:353–360.
15. Ciavatta, D., S. Rogers, and T. Magnuson. 2007. Drosophila CTCF is re-quired for Fab-8 enhancer blocking activity in S2 cells. J. Mol. Biol. 373:233–239.
16. Dahle, O., O. Bakke, and O. S. Gabrielsen. 2004. c-Myb associates with PMLin nuclear bodies in hematopoietic cells. Exp. Cell Res. 297:118–126.
17. Defossez, P. A., and E. Gilson. 2002. The vertebrate protein CTCF functionsas an insulator in Saccharomyces cerevisiae. Nucleic Acids Res. 30:5136–5141.
18. Donohoe, M. E., L. F. Zhang, N. Xu, Y. Shi, and J. T. Lee. 2007. Identifica-tion of a Ctcf cofactor, Yy1, for the X chromosome binary switch. Mol. Cell25:43–56.
19. Du, M., L. G. Beatty, W. Zhou, J. Lew, C. Schoenherr, R. Weksberg, andP. D. Sadowski. 2003. Insulator and silencer sequences in the imprintedregion of human chromosome 11p15.5. Hum. Mol. Genet. 12:1927–1939.
20. Du, M., W. Zhou, L. G. Beatty, R. Weksberg, and P. D. Sadowski. 2004. TheKCNQ1OT1 promoter, a key regulator of genomic imprinting in humanchromosome 11p15.5. Genomics 84:288–300.
21. Dunn, K. L., and J. R. Davie. 2003. The many roles of the transcriptionalregulator CTCF. Biochem. Cell Biol. 81:161–167.
22. El-Kady, A., and E. Klenova. 2005. Regulation of the transcription factor,CTCF, by phosphorylation with protein kinase CK2. FEBS Lett. 579:1424–1434.
23. Facchini, L. M., S. Chen, W. W. Marhin, J. N. Lear, and L. Z. Penn. 1997.The Myc negative autoregulation mechanism requires Myc-Max associationand involves the c-Myc P2 minimal promoter. Mol. Cell. Biol. 17:100–114.
24. Fedoriw, A. M., P. Stein, P. Svoboda, R. M. Schultz, and M. S. Bartolomei.2004. Transgenic RNAi reveals essential function for CTCF in H19 geneimprinting. Science 303:238–240.
25. Filippova, G. N. 2008. Genetics and epigenetics of the multifunctional pro-tein CTCF. Curr. Top. Dev. Biol. 80:337–360.
26. Filippova, G. N., S. Fagerlie, E. M. Klenova, C. Myers, Y. Dehner, G.Goodwin, P. E. Neiman, S. J. Collins, and V. V. Lobanenkov. 1996. Anexceptionally conserved transcriptional repressor, CTCF, employs differentcombinations of zinc fingers to bind diverged promoter sequences of avianand mammalian c-Myc oncogenes. Mol. Cell. Biol. 16:2802–2813.
27. Filippova, G. N., C. F. Qi, J. E. Ulmer, J. M. Moore, M. D. Ward, Y. J. Hu,D. I. Loukinov, E. M. Pugacheva, E. M. Klenova, P. E. Grundy, A. P.Feinberg, A. M. Cleton-Jansen, E. W. Moerland, C. J. Cornelisse, H. Suzuki,A. Komiya, A. Lindblom, F. Dorion-Bonnet, P. E. Neiman, H. C. Morse III,S. J. Collins, and V. V. Lobanenkov. 2002. Tumor-associated zinc finger
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 723
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

mutations in the CTCF transcription factor selectively alter its DNA-bindingspecificity. Cancer Res. 62:48–52.
28. Gill, G. 2005. Something about SUMO inhibits transcription. Curr. Opin.Genet. Dev. 15:536–541.
29. Gill, G. 2004. SUMO and ubiquitin in the nucleus: different functions,similar mechanisms? Genes Dev. 18:2046–2059.
30. Gombert, W. M., S. D. Farris, E. D. Rubio, K. M. Morey-Rosler, W. H.Schubach, and A. Krumm. 2003. The c-Myc insulator element and matrixattachment regions define the c-Myc chromosomal domain. Mol. Cell. Biol.23:9338–9348.
31. Gong, L., and E. T. Yeh. 2006. Characterization of a family of nucleolarSUMO-specific proteases with preference for SUMO-2 or SUMO-3. J. Biol.Chem. 281:15869–15877.
32. Goodliffe, J. M., E. Wieschaus, and M. D. Cole. 2005. Polycomb mediatesMyc autorepression and its transcriptional control of many loci in Drosoph-ila. Genes Dev. 19:2941–2946.
33. Gregoire, S., A. M. Tremblay, L. Xiao, Q. Yang, K. Ma, J. Nie, Z. Mao, Z.Wu, V. Giguere, and X. J. Yang. 2006. Control of MEF2 transcriptionalactivity by coordinated phosphorylation and sumoylation. J. Biol. Chem.281:4423–4433.
34. Guo, A., P. Salomoni, J. Luo, A. Shih, S. Zhong, W. Gu, and P. P. Pandolfi.2000. The function of PML in p53-dependent apoptosis. Nat. Cell Biol.2:730–736.
35. Gutierrez, G. J., and Z. Ronai. 2006. Ubiquitin and SUMO systems in theregulation of mitotic checkpoints. Trends Biochem. Sci. 31:324–332.
36. Hark, A. T., C. J. Schoenherr, D. J. Katz, R. S. Ingram, J. M. Levorse, andS. M. Tilghman. 2000. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature 405:486–489.
37. Holohan, E. E., C. Kwong, B. Adryan, M. Bartkuhn, M. Herold, R. Renka-witz, S. Russell, and R. White. 2007. CTCF genomic binding sites in Dro-sophila and the organisation of the bithorax complex. PLoS Genet. 3:e112.
38. Kagey, M. H., T. A. Melhuish, S. E. Powers, and D. Wotton. 2005. Multipleactivities contribute to Pc2 E3 function. EMBO J. 24:108–119.
39. Kagey, M. H., T. A. Melhuish, and D. Wotton. 2003. The polycomb proteinPc2 is a SUMO E3. Cell 113:127–137.
40. Kerscher, O., R. Felberbaum, and M. Hochstrasser. 2006. Modification ofproteins by ubiquitin and ubiquitin-like proteins. Annu. Rev. Cell Dev. Biol.22:159–180.
41. Kim, T. H., Z. K. Abdullaev, A. D. Smith, K. A. Ching, D. I. Loukinov, R. D.Green, M. Q. Zhang, V. V. Lobanenkov, and B. Ren. 2007. Analysis of thevertebrate insulator protein CTCF-binding sites in the human genome. Cell128:1231–1245.
42. Klenova, E., and R. Ohlsson. 2005. Poly(ADP-ribosyl)ation and epigenetics.Is CTCF PARt of the plot? Cell Cycle 4:96–101.
43. Klenova, E. M., I. V. Chernukhin, A. El-Kady, R. E. Lee, E. M. Pugacheva,D. I. Loukinov, G. H. Goodwin, D. Delgado, G. N. Filippova, J. Leon, H. C.Morse III, P. E. Neiman, and V. V. Lobanenkov. 2001. Functional phosphor-ylation sites in the C-terminal region of the multivalent multifunctionaltranscriptional factor CTCF. Mol. Cell. Biol. 21:2221–2234.
44. Kurukuti, S., V. K. Tiwari, G. Tavoosidana, E. Pugacheva, A. Murrell, Z.Zhao, V. Lobanenkov, W. Reik, and R. Ohlsson. 2006. CTCF binding at theH19 imprinting control region mediates maternally inherited higher-orderchromatin conformation to restrict enhancer access to Igf2. Proc. Natl. Acad.Sci. USA 103:10684–10689.
45. Kyrchanova, O., S. Toshchakov, Y. Podstreshnaya, A. Parshikov, and P.Georgiev. 2008. Functional interaction between the Fab-7 and Fab-8 bound-aries and the upstream promoter region in Drosophila Abd-B gene. Mol.Cell. Biol. 28:4188–4195.
46. Li, B., J. Zhou, P. Liu, J. Hu, H. Jin, Y. Shimono, M. Takahashi, and G. Xu.2007. Polycomb protein Cbx4 promotes SUMO modification of de novoDNA methyltransferase Dnmt3a. Biochem. J. 405:369–378.
47. Ling, J. Q., T. Li, J. F. Hu, T. H. Vu, H. L. Chen, X. W. Qiu, A. M. Cherry,and A. R. Hoffman. 2006. CTCF mediates interchromosomal colocalizationbetween Igf2/H19 and Wsb1/Nf1. Science 312:269–272.
48. Liu, L., D. M. Scolnick, R. C. Trievel, H. B. Zhang, R. Marmorstein, T. D.Halazonetis, and S. L. Berger. 1999. p53 sites acetylated in vitro by PCAFand p300 are acetylated in vivo in response to DNA damage. Mol. Cell. Biol.19:1202–1209.
49. Lobanenkov, V. V., R. H. Nicolas, V. V. Adler, H. Paterson, E. M. Klenova,A. V. Polotskaja, and G. H. Goodwin. 1990. A novel sequence-specific DNAbinding protein which interacts with three regularly spaced direct repeats ofthe CCCTC-motif in the 5�-flanking sequence of the chicken c-myc gene.Oncogene 5:1743–1753.
50. Majumder, P., J. A. Gomez, B. P. Chadwick, and J. M. Boss. 17 March 2008.The insulator factor CTCF controls MHC class II gene expression and isrequired for the formation of long-distance chromatin interactions. J. Exp.Med. doi10.1084/jem.20071843.
51. Mohan, M., M. Bartkuhn, M. Herold, A. Philippen, N. Heinl, I. Bardenha-gen, J. Leers, R. A. White, R. Renkawitz-Pohl, H. Saumweber, and R. Ren-kawitz. 2007. The Drosophila insulator proteins CTCF and CP190 link en-hancer blocking to body patterning. EMBO J. 26:4203–4214.
52. Moon, H., G. Filippova, D. Loukinov, E. Pugacheva, Q. Chen, S. T. Smith, A.
Munhall, B. Grewe, M. Bartkuhn, R. Arnold, L. J. Burke, R. Renkawitz-Pohl, R. Ohlsson, J. Zhou, R. Renkawitz, and V. Lobanenkov. 2005. CTCFis conserved from Drosophila to humans and confers enhancer blocking ofthe Fab-8 insulator. EMBO Rep. 6:165–170.
53. Mukhopadhyay, R., W. Yu, J. Whitehead, J. Xu, M. Lezcano, S. Pack, C.Kanduri, M. Kanduri, V. Ginjala, A. Vostrov, W. Quitschke, I. Chernukhin,E. Klenova, V. Lobanenkov, and R. Ohlsson. 2004. The binding sites for thechromatin insulator protein CTCF map to DNA methylation-free domainsgenome-wide. Genome Res. 14:1594–1602.
54. Navarro, P., D. R. Page, P. Avner, and C. Rougeulle. 2006. Tsix-mediatedepigenetic switch of a CTCF-flanked region of the Xist promoter determinesthe Xist transcription program. Genes Dev. 20:2787–2792.
55. Ohlsson, R., R. Renkawitz, and V. Lobanenkov. 2001. CTCF is a uniquelyversatile transcription regulator linked to epigenetics and disease. TrendsGenet. 17:520–527.
56. Pant, V., S. Kurukuti, E. Pugacheva, S. Shamsuddin, P. Mariano, R. Ren-kawitz, E. Klenova, V. Lobanenkov, and R. Ohlsson. 2004. Mutation of asingle CTCF target site within the H19 imprinting control region leads to lossof Igf2 imprinting and complex patterns of de novo methylation upon ma-ternal inheritance. Mol. Cell. Biol. 24:3497–3504.
57. Parelho, V., S. Hadjur, M. Spivakov, M. Leleu, S. Sauer, H. C. Gregson, A.Jarmuz, C. Canzonetta, Z. Webster, T. Nesterova, B. S. Cobb, K. Yokomori,N. Dillon, L. Aragon, A. G. Fisher, and M. Merkenschlager. 2008. cohesinsfunctionally associate with CTCF on mammalian chromosome arms. Cell132:422–433.
58. Pugacheva, E. M., V. K. Tiwari, Z. Abdullaev, A. A. Vostrov, P. T. Flanagan,W. W. Quitschke, D. I. Loukinov, R. Ohlsson, and V. V. Lobanenkov. 2005.Familial cases of point mutations in the XIST promoter reveal a correlationbetween CTCF binding and pre-emptive choices of X chromosome inacti-vation. Hum. Mol. Genet. 14:953–965.
59. Roscic, A., A. Moller, M. A. Calzado, F. Renner, V. C. Wimmer, E. Gresko,K. S. Ludi, and M. L. Schmitz. 2006. Phosphorylation-dependent control ofPc2 SUMO E3 ligase activity by its substrate protein HIPK2. Mol. Cell24:77–89.
60. Saitoh, H., and J. Hinchey. 2000. Functional heterogeneity of small ubiq-uitin-related protein modifiers SUMO-1 versus SUMO-2/3. J. Biol. Chem.275:6252–6258.
61. Saurin, A. J., C. Shiels, J. Williamson, D. P. Satijn, A. P. Otte, D. Sheer, andP. S. Freemont. 1998. The human polycomb group complex associates withpericentromeric heterochromatin to form a novel nuclear domain. J. CellBiol. 142:887–898.
62. Schoenherr, C. J., J. M. Levorse, and S. M. Tilghman. 2003. CTCF maintainsdifferential methylation at the Igf2/H19 locus. Nat. Genet. 33:66–69.
63. Schwartz, D. C., and M. Hochstrasser. 2003. A superfamily of protein tags:ubiquitin, SUMO and related modifiers. Trends Biochem. Sci. 28:321–328.
64. Splinter, E., H. Heath, J. Kooren, R. J. Palstra, P. Klous, F. Grosveld, N.Galjart, and W. de Laat. 2006. CTCF mediates long-range chromatin loop-ing and local histone modification in the beta-globin locus. Genes Dev.20:2349–2354.
65. Stedman, W., H. Kang, S. Lin, J. L. Kissil, M. S. Bartolomei, and P. M.Lieberman. 2008. Cohesins localize with CTCF at the KSHV latency controlregion and at cellular c-myc and H19/Igf2 insulators. EMBO J. 27:654–666.
66. Szabo, P. E., S. H. Tang, F. J. Silva, W. M. Tsark, and J. R. Mann. 2004. Roleof CTCF binding sites in the Igf2/H19 imprinting control region. Mol. Cell.Biol. 24:4791–4800.
67. Vanhatupa, S., D. Ungureanu, M. Paakkunainen, and O. Silvennoinen.2007. MAPK-induced Ser727 phosphorylation promotes sumoylation ofSTAT1. Biochem. J. 409:179–185.
68. Wallace, J. A., and G. Felsenfeld. 2007. We gather together: insulators andgenome organization. Curr. Opin. Genet. Dev. 17:400–407.
69. Wendt, K. S., K. Yoshida, T. Itoh, M. Bando, B. Koch, E. Schirghuber, S.Tsutsumi, G. Nagae, K. Ishihara, T. Mishiro, K. Yahata, F. Imamoto, H.Aburatani, M. Nakao, N. Imamoto, K. Maeshima, K. Shirahige, and J. M.Peters. 2008. Cohesin mediates transcriptional insulation by CCCTC-bindingfactor. Nature 451:796–801.
70. Wotton, D., and J. C. Merrill. 2007. Pc2 and SUMOylation. Biochem. Soc.Trans. 35:1401–1404.
71. Xi, H., H. P. Shulha, J. M. Lin, T. R. Vales, Y. Fu, D. M. Bodine, R. D.McKay, J. G. Chenoweth, P. J. Tesar, T. S. Furey, B. Ren, Z. Weng, and G. E.Crawford. 2007. Identification and characterization of cell type-specific andubiquitous chromatin regulatory structures in the human genome. PLoSGenet. 3:e136.
72. Xie, X., T. S. Mikkelsen, A. Gnirke, K. Lindblad-Toh, M. Kellis, and E. S.Lander. 2007. Systematic discovery of regulatory motifs in conserved regionsof the human genome, including thousands of CTCF insulator sites. Proc.Natl. Acad. Sci. USA 104:7145–7150.
73. Xu, N., M. E. Donohoe, S. S. Silva, and J. T. Lee. 2007. Evidence thathomologous X-chromosome pairing requires transcription and Ctcf protein.Nat. Genet. 39:1390–1396.
74. Yoon, Y. S., S. Jeong, Q. Rong, K. Y. Park, J. H. Chung, and K. Pfeifer. 2007.Analysis of the H19ICR insulator. Mol. Cell. Biol. 27:3499–3510.
75. Yu, W., V. Ginjala, V. Pant, I. Chernukhin, J. Whitehead, F. Docquier, D.
724 MACPHERSON ET AL. MOL. CELL. BIOL.
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

Farrar, G. Tavoosidana, R. Mukhopadhyay, C. Kanduri, M. Oshimura, A. P.Feinberg, V. Lobanenkov, E. Klenova, and R. Ohlsson. 2004. Poly(ADP-ribosyl)ation regulates CTCF-dependent chromatin insulation. Nat. Genet.36:1105–1110.
76. Yusufzai, T. M., and G. Felsenfeld. 2004. The 5�-HS4 chicken beta-globininsulator is a CTCF-dependent nuclear matrix-associated element. Proc.Natl. Acad. Sci. USA 101:8620–8624.
77. Yusufzai, T. M., H. Tagami, Y. Nakatani, and G. Felsenfeld. 2004. CTCF
tethers an insulator to subnuclear sites, suggesting shared insulator mecha-nisms across species. Mol. Cell 13:291–298.
78. Zeghouf, M., J. Li, G. Butland, A. Borkowska, V. Canadien, D. Richards, B.Beattie, A. Emili, and J. F. Greenblatt. 2004. Sequential peptide affinity(SPA) system for the identification of mammalian and bacterial proteincomplexes. J. Proteome Res. 3:463–468.
79. Zhong, S., P. Salomoni, and P. P. Pandolfi. 2000. The transcriptional role ofPML and the nuclear body. Nat. Cell Biol. 2:E85–E90.
VOL. 29, 2009 SUMO MODIFICATION OF CTCF 725
on April 5, 2019 by guest
http://mcb.asm
.org/D
ownloaded from





























