Embed Size (px)
Citation preview

THE HUMAN ENHANCER-BLOCKER CTCF INTERACTSWITH THE TRANSCRIPTION FACTOR KAISO*
Pierre-Antoine Defossez‡§¶, Kevin F. Kelly**, Guillaume Filion§, Roberto Pérez-Torrado§,Frédérique Magdinier‡, Hervé Menoni‡, Curtis L. Nordgaard**, Juliet M. Daniel**, and Eric
Gilson‡From the ‡CNRS UMR5161, Ecole Normale Supérieure de Lyon, 46 allée d’Italie, 69364 Lyon
Cedex 07, France; §CNRS UMR218, Institut Curie Section Recherche, 26 rue d’Ulm, 75248 ParisCedex 05, France and ** Department of Biology, McMaster University, 1280 Main St. West,
Hamilton, ON L8S 4K1, Canada.Running Title: Inhibition of CTCF by Kaiso
¶ To whom correspondence should be addressed. Tel. +33 1 42 34 67 00; Fax +33 1 46 33 30 16; E-mail:[email protected]
CTCF is a DNA-binding protein ofvertebrates that plays essential roles inregulating genome activity through its capacityto act as an enhancer-blocker. We performed ayeast-two hybrid screen to identify proteinpartners of CTCF that could regulate itsactivity. Using full-length CTCF as bait werecovered Kaiso, a POZ-zinc fingertranscription factor, as a specific bindingpartner. The interaction occurs through a C-terminal region of CTCF and the POZ domainof Kaiso. CTCF and Kaiso are co-expressed inmany tissues and CTCF was specifically co-immunoprecipitated by several Kaisomonoclonal antibodies from nuclear lysates.Kaiso is a bimodal transcription factor thatrecognizes methylated CpG dinucleotides, or ac o n s e r v e d u n m e t h y l a t e d s e q u e n c e(TNGCAGGA - the Kaiso binding site). Weidentified one consensus unmethylated Kaisobinding site in close proximity to the CTCFbinding site in the human 5’ beta-globininsulator. We found that the presence of thisKaiso binding site reduces the enhancer-blocking activity of CTCF in an insulationassay. These data suggest that the Kaiso-CTCFinteraction negatively regulates CTCF insulatoractivity.
The genome of eukaryotes is partitionedinto transcriptionally active and transcriptionallyinactive domains (1). Insulators are DNAelements that maintain this partition, and they canbe subdivided into two functional classes: barrierelements, that stop the spread of heterochromatin,and enhancer blockers, that prevent an enhancerfrom activating transcription in a neighboringrepressed region (2). One of the best-studied loci
regarding long-range transcriptional regulation isthe beta-globin locus of vertebrates (3). This locuscontains an enhancer, the LCR (Locus ControlRegion ), that acts on the globin genes. Theactivity of the LCR is confined by two insulators,one at the 5’ boundary of the locus, and another atthe 3’ boundary. Both insulators depend on thesame protein, CTCF (4,5).
CTCF was originally isolated as a zinc-finger transcription factor that recognized a CTC-rich sequence in the c-myc promoter (6). Over theyears, CTCF has been shown to have complex andimportant roles in the control of gene expression(7). CTCF binds many different DNA targetsequences through the combinatorial use of its 11zinc fingers, and it is capable of both activatingand repressing gene transcription. An additionalrole of CTCF is to act as an enhancer blocker thatprevents communication between an enhancer anda target gene. This process is known astranscriptional insulation. CTCF and YY1 (8) arethe only two vertebrate proteins known to act asenhancer-blockers. CTCF exerts this criticalfunction at many loci (2). For example, enhancerblocking by CTCF permits correct expression ofthe imprinted genes H19 and IGF2. At this locus,CTCF is only active on the maternal chromosome.The CTCF target sites on the paternal chromosomeare methylated, and this modification completelyprecludes CTCF binding (9-11).
Recently, significant advances were madein our understanding of how CTCF functions as anenhancer-blocker at the 5’ chicken beta-globininsulator (12,13). At this insulator site, CTCFinteracts with nucleophosmin, a nuclear matrixprotein that is concentrated at the surface of the
http://www.jbc.org/cgi/doi/10.1074/jbc.M510802200The latest version is at JBC Papers in Press. Published on October 17, 2005 as Manuscript M510802200
Copyright 2005 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

nucleolus. This is thought to result in theformation of physically separated DNA loops,which would then prevent an enhancer element inone chromatin loop from acting on a gene in theneighboring chromatin loop. Interestingly, thisfinding is consistent with results obtained inDrosophila, where a crucial link between nucleararchitecture and transcriptional insulation wasdiscovered (1). It is likely that this model alsoaccounts for CTCF action at other insulators, yetalternative mechanisms cannot be ruled out at thispoint.
The activity of enhancer blockers is notstatic, but can turned on and off. Recentexperiments with CTCF have shown that post-translational modification of the protein plays animportant regulatory role (14). In addition, thefunction of CTCF can also be regulated throughinteracting proteins (15). In this study, we soughtto identify binding partners of CTCF that couldinfluence its activity as an enhancer-blocker. Wereport the identification of the protein Kaiso as aspecific binding partner for CTCF. Kaiso is amember of the POZ (Pox virus and zinc finger)family of zinc finger (ZF) transcription factors thatare implicated in cancer and development (16). Todate, Kaiso is the only POZ-ZF protein that hasbeen shown to have dual-specificity DNA-binding:it can bind methylated CpG dinucleotides (17), ora specific non-methylated DNA sequence(TNGCAGGA) (18). Indeed, we identified onenon-methylated Kaiso consensus site near theCTCF binding site on the human 5’ beta globininsulator. We show that the presence of an intactKaiso binding site near the CTCF binding site onthe 5’ beta globin insulator inhibits the enhancer-blocking activity of CTCF. This raises thepossibility that Kaiso is a negative regulator ofCTCF enhancer-blocking activity.
MATERIALS AND METHODS
Plasmids-The two-hybrid bait plasmid wasconstructed by cloning the full-length chickenCTCF cDNA (provided by Rainer Renkawitz)between the EcoRI and BglII sites of vectorpGBDU-C(1) (URA3-marked, multicopy, ADH1promoter driving expression of the bait fused toGAL4p1-147) (19). The Gal4-CTCF deletionconstructs used in Fig. 3 have been previously
described (20). To construct the insulationreporters described in Fig. 4, we first cloned therelevant oligonucleotides into the EcoRV site ofpBluescriptII KS. They were then PCR amplifiedwith primers containing MluI sites, digested withMluI, and inserted into the AscI site of pNI (21).All constructs were sequenced.
Two-hybrid screen-We used the two-hybrid strainPJ69-4a (19). The strain was first transformed withthe bait plasmid, then with a library of cDNAsfrom 6.5-9.5 dpc mouse embryos cloned into thepASV3 vector (LEU2 marker, multicopy, PGK1promoter driving the expression of peptides fusedto the VP16 activation domain) (22). The librarywas a kind gift from Régine Losson. Yeasttransformation was done using Lithium acetate(23). Transformation efficiency was calculated byplating an aliquot of the cells on plates lackinguracil and leucine. Interactors were selected onplates lacking uracil, leucine, histidine, andcontaining 5mM 3-aminotriazole. We then testedwhether growth on plates lacking histidine wasdependent on the bait plasmid. Finally, cellscontaining the candidate interactors were mated toPJ69-4α containing different bait plasmids to testthe specificity of the interaction. Only two clonespassed all the screening procedures.
Cell and tissue culture-Mammalian cells used inthis study were the human cervical carcinoma cellline HeLa and the human erythroleukemia K562cells. HeLa cells were grown at 37ºC, 5% CO2 inDulbecco’s minimal essential medium (DMEM)supplemented with 10% fetal bovine serum, 4 mML-glutamine, penicillin (100 U/ml), streptomycin(100 mg/ml) and fungizone (0.5 mg/ml). K562cells were grown in RPMI1640 medium with 10%fetal bovine serum, penicillin (100 U/ml) andstreptomycin (100 mg/ml).
Co-immunoprecipitation and immunoblot analysis-HeLa cells were washed once with 5 ml PBS (pH7.4) prior to the preparation of cytoplasmic andnuclear fractions (24). Nuclear lysates werequantified by Bradford assay and equal amounts oftotal protein were used for immunoprecipitationwith anti-Kaiso monoclonal antibodies 6F, 2G,12H, 12G, 11D (25) or with rabbit anti-CTCFpolyclonal antibody (Upstate, Ref. 06-917). Theimmune complexes were then subjected to sodium-
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

dodecyl-sulfate polyacrylamide-gel electrophoresis(SDS-PAGE) as previously described (16). Afterelectrophoresis, the proteins were transferred to anitrocellulose membrane. The membrane wasbriefly blocked at room temperature with 3%skimmed milk powder in TBS (pH 7.4) beforeincubating at 4°C overnight with rabbit anti-Kaisopolyclonal antibody at 1/12,000 dilution or rabbitanti-CTCF polyclonal antibody at 1/500 dilution in3% milk/TBS. The primary antibodies wereremoved by rinsing the membranes five times withwater and then once with TBS, for five minuteseach. The membranes were then incubated for 2hours at room temperature with horseradish-peroxidase-conjugated goat anti-rabbit secondaryantibody, diluted 1:40,000 in 3% milk/TBS.Membranes were finally rinsed five times withwater and once with TBS (pH 7.4) for 5 minuteseach, and processed using the enhancedc h e m i l u m i n e s c e n c e ( E C L ) s y s t e m(Amersham/Pharmacia) according to themanufacturer's protocols. The in vitro interactionbetween GST-Kaiso and CTCF was tested aspreviously described (26).
Insulation assay-We used the method developed inthe Felsenfeld lab (21). The various reporters werelinearized with SalI, and DNA was quantified byboth UV spectrophotometry and analysis on anagarose gel. One hundred nanograms of eachlinearized plasmid was then electroporated into1x107 K562 cells by electroporation. After 24hours of recovery, the cells were plated in 0.35%agar medium with 750 µg/ml geneticin (activeconcentration) in two 150-mm dishes. Colonieswere counted after three weeks. Each constructwas tested in duplicate in at least three separateexperiments, using a different DNA preparationeach time. Statistical analysis of the results wasdone using Student’s T-test.
Immunoprecipitation of chromatin- Hela cellswere fixed with 1% formaldehyde for 60 minutesat room temperature. The cells were lysed, and thechromatin was sonicated to an average size of 600base pairs. For each experiment the chromatinprepared from 107 cells was immunoprecipitatedwith 4 micrograms of the relevant antibody, usinga previously described protocol (27). The primersused to amplify the human beta-globin region are
T G A G G A T G C C T C C T T C T C T G a n dCAGCAGCTTCAGCTACCTCTC.
RESULTS
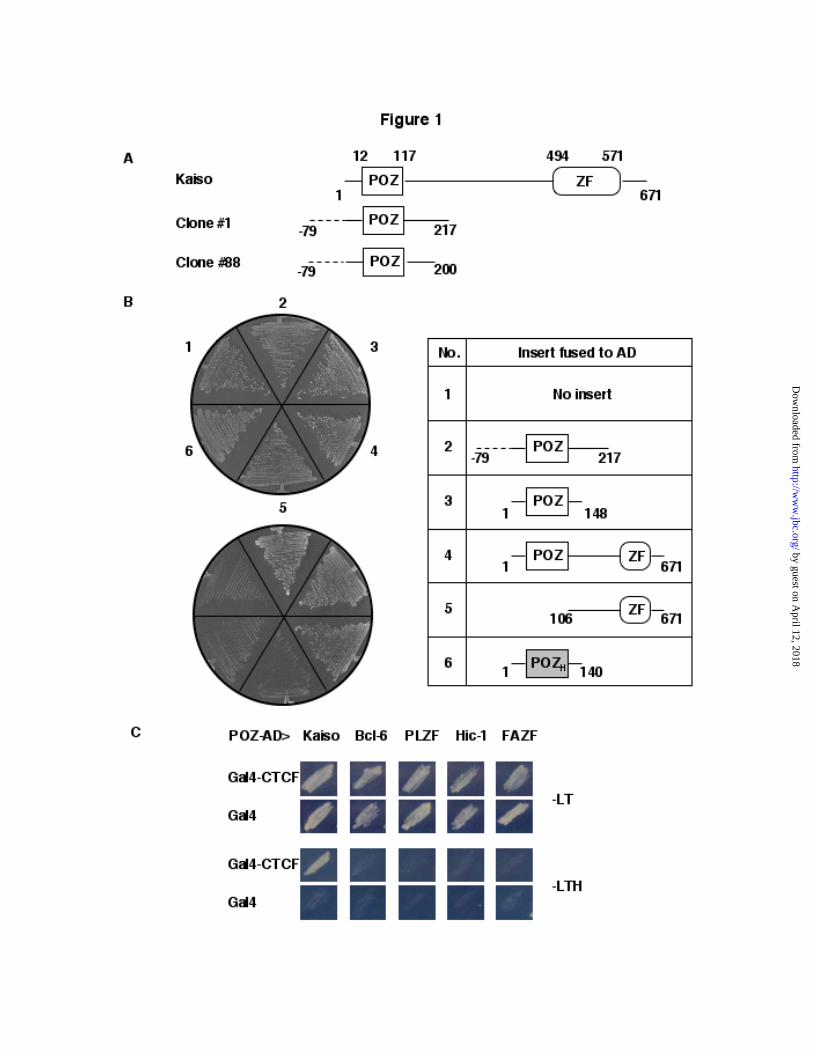
Biochemical approaches have been used toseek CTCF interactors (28). To identify additionalinteractors, and potential regulators of CTCFfunction, we performed a yeast two-hybrid screenusing a mouse embryo cDNA library and full-length CTCF as the bait. Out of 2 milliontransformants we obtained only two positiveclones, #1 and #88. The two clones containedoverlapping fragments of the same cDNA,encoding a fraction of the transcription factorKaiso (Fig. 1A ). Both clones possessed 237nucleotides upstream of the reported Kaiso ATG,that are translated in frame with the rest of thecDNA, and generate a 79 amino-acid extension notnormally present in Kaiso. The clones differed bythe fact that clone #1 encoded the first 217 amino-acids of Kaiso, whereas clone #88 encoded thefirst 200 amino acids. This N-terminal region ofKaiso contains the POZ domain, which is knownto mediate homodimerization and hetero-dimerization with other POZ family proteins, ornon-POZ domain proteins (29,30). This stronglysuggested that the Kaiso POZ domain wasinvolved in mediating the CTCF interaction. Infact, in an independent study, the Daniel labidentified CTCF as a Kaiso-specific bindingpartner using the Kaiso POZ domain as bait (Dr.Juliet Daniel, personal communication). As seen inFig. 1B, we found that CTCF interacted with thefull-length Kaiso protein, and also with the isolatedKaiso POZ domain. When the POZ domain wasdeleted from Kaiso, interaction with CTCF waslost (Fig. 1B). This indicates that the POZ domainis both necessary and sufficient for the CTCF-Kaiso interaction. Since POZ domains are highlyconserved and present in a large number oftranscription factors (30), we tested the specificityof the interaction by asking whether CTCF wouldinteract with the POZ domain of four other POZ-ZF proteins (BCL-6, PLZF, HIC-1, FAZF). Asseen in Fig. 1B and 1C , CTCF interactedspecifically with the Kaiso POZ domain and noother POZ domain tested. We conclude that CTCFrecognizes a specific feature in the POZ domain ofKaiso.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

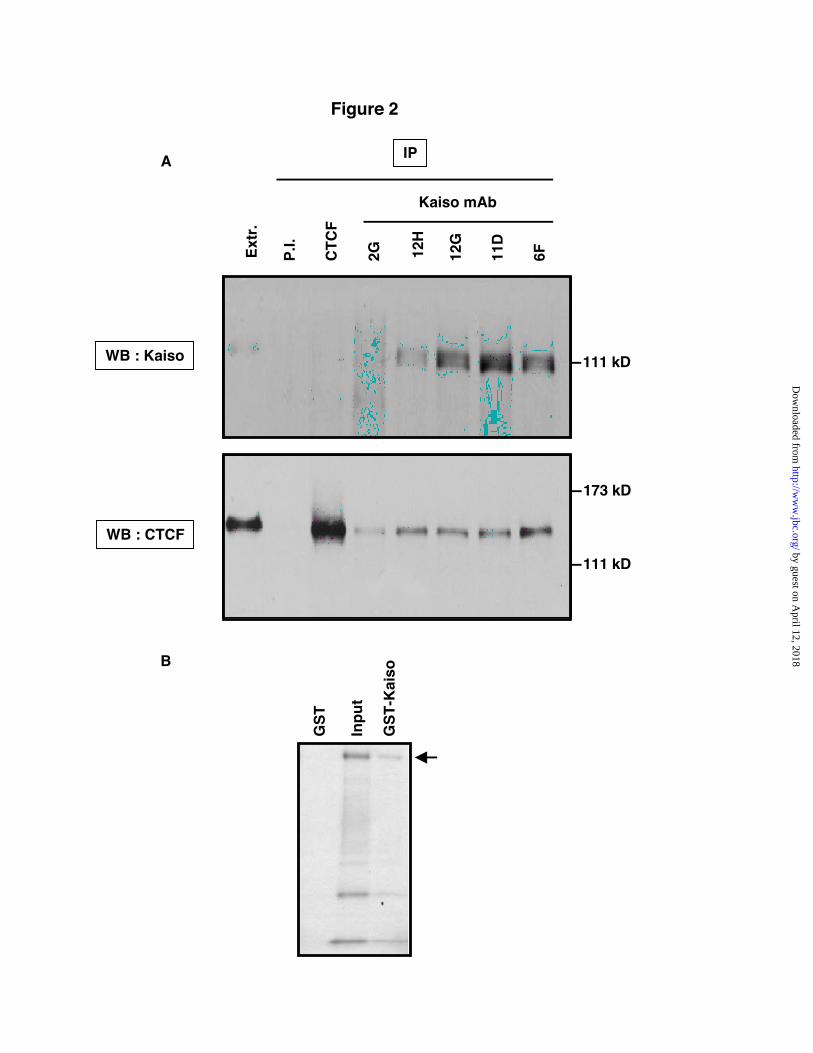
To determine whether CTCF and Kaisointeract in vivo in vertebrate cells, we performedco-immunoprecipitation experiments using humancervical carcinoma (HeLa) cells which expressboth CTCF and Kaiso endogenously at high tomoderate levels. We prepared nuclear extracts andimmunoprecipitated Kaiso using five differentKaiso-specific monoclonal antibodies that havebeen previously characterized (25). Theimmunoprecipitates were subjected to SDS-PAGEand Western blot analysis using CTCF-specificantibodies. As seen in Fig. 2, endogenous CTCFwas specifically co-precipitated by the variousKaiso-specific monoclonal antibodies. In contrast,the pre-immune serum failed to precipitate anyCTCF-containing material. We thus conclude thatCTCF and Kaiso exist in a complex in vertebratecells. To further verify the interaction, weperformed the reciprocal experiment byimmunoprecipitating CTCF and Western blottingwith a Kaiso-specific rabbit polyclonal antibody.However we failed to detect Kaiso co-precipitatingwith CTCF in this reciprocal situation. This maybe due to the fact that the CTCF antiserum wasraised against a region of CTCF that contains theKaiso interaction domain (see following section).The CTCF antibody may recognize onlyuncomplexed CTCF, or the antibody may perturbthe native CTCF-Kaiso interaction. An alternativeexplanation for the failure of the CTCF antiserumto co-immunoprecipitate Kaiso could bestoichiometry: CTCF protein may be in largeexcess relative to Kaiso, with only a minor sub-population of CTCF molecules bound to Kaiso atsteady state. To determine whether the interactionbetween CTCF and Kaiso is direct, we used an invitro interaction assay. Radioactively labeledCTCF was produced by in vitro transcription andtranslation, and was incubated with bacteriallyexpressed Glutathion S-Transferase (GST), or withGST fused to full-length Kaiso (GST-Kaiso). NoCTCF was retained after incubation with GST,whereas CTCF bound the GST-Kaiso protein(Figure 2B). From this we conclude that Kaiso andCTCF interact directly, without the involvement ofa bridging factor.
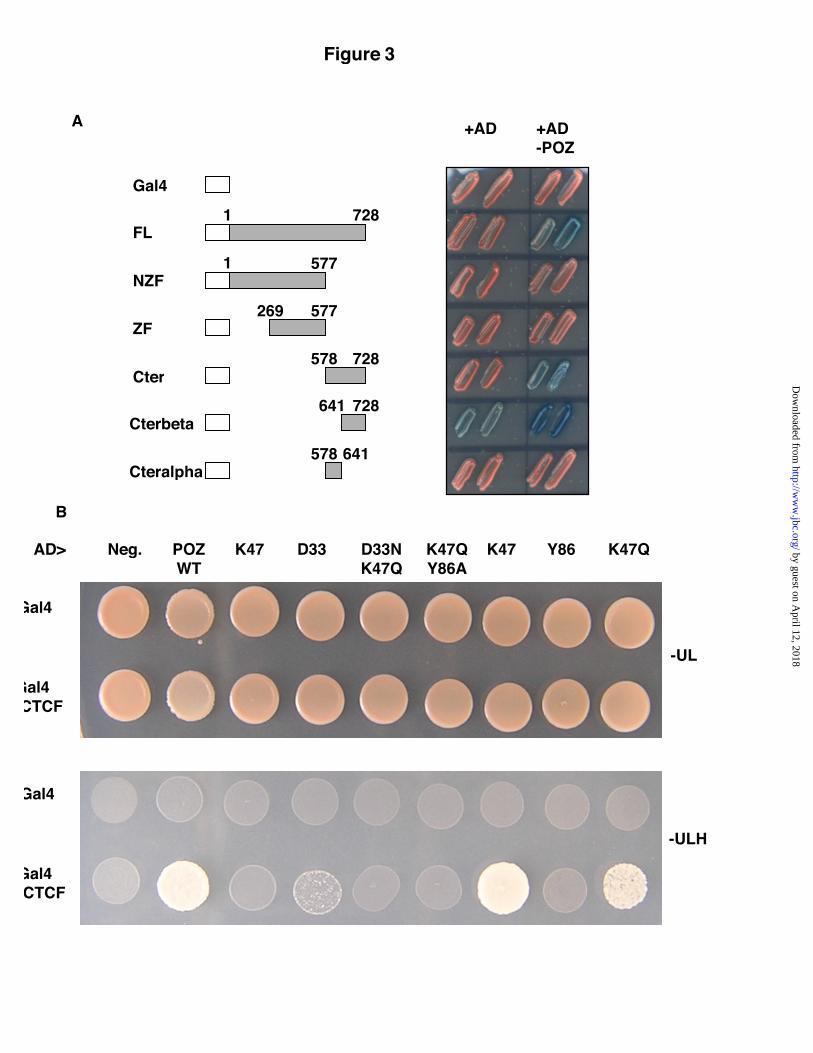
To delineate the region of CTCF thatinteracts with Kaiso, we tested a series of CTCFdeletion mutants for their ability to bind the KaisoPOZ domain in a yeast two-hybrid assay (Fig. 3A).
We delineated the binding domain of Kaiso to a C-terminal region encoding amino-acids 641-728 ofCTCF. This domain proved necessary andsufficient for the CTCF-Kaiso interaction. Sinceprevious studies demonstrated that specific highlyconserved POZ domain residues of BCL-6 andPLZF were crucial for mediating the homo- andheterodimerization capabilities of thesetranscription factors (31), we questioned whetherthe equivalent residues in Kaiso were crucial forthe Kaiso-CTCF interaction. Hence, to moreprecisely define the Kaiso binding site, wegenerated point mutations in the Kaiso POZdomain (D33N, K47Q, K47R, Y86A andD33N/K47Q), and tested the capacity of thesemutants to interact with CTCF in the yeast two-hybrid assay (Fig. 3B ). The lysine residue atposition 47 (present in human Kaiso) could besubstituted with arginine (present in murine Kaiso)with no apparent loss of activity. However, lessconservative substitutions caused partial loss ofinteraction (lysine to glutamine) or totallyabrogated the interaction (lysine to a proline). Wealso tested two other highly conserved amino acidresidues that have been implicated as keydeterminants in POZ domain function (31,32). Wefound that substituting the highly conservedaspartic acid-33 with an asparagine, or tyrosine-86with alanine also disrupted the Kaiso-CTCFinteraction.
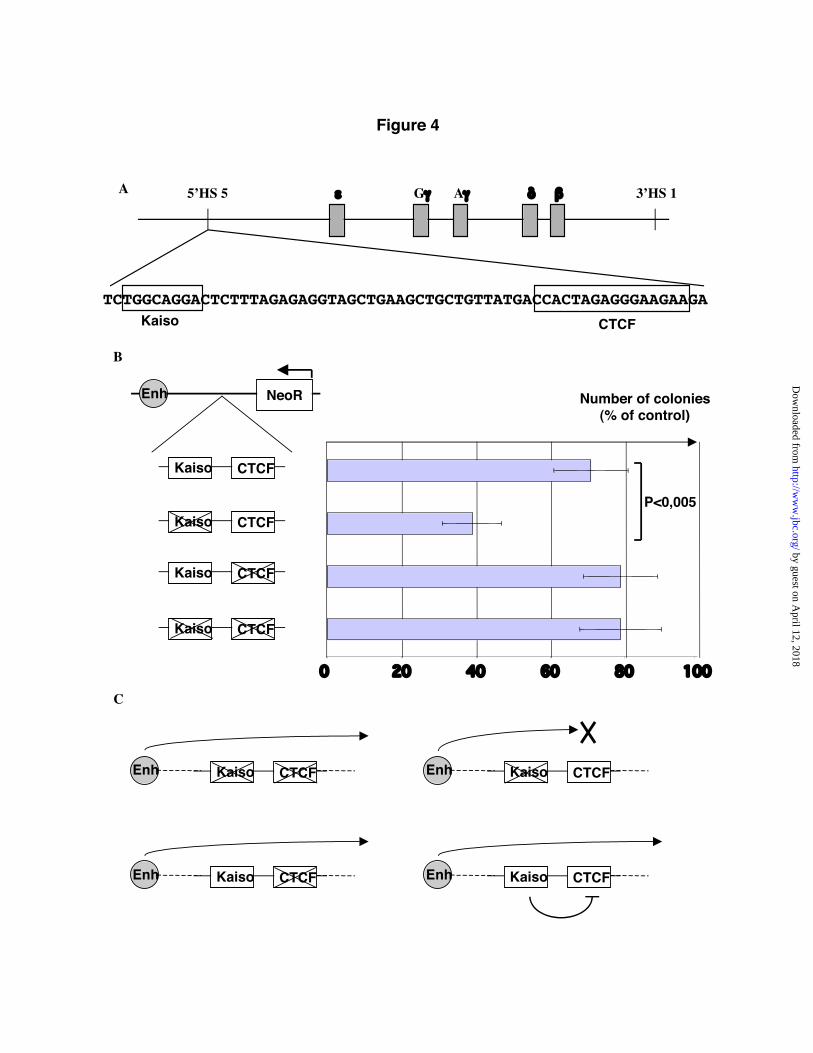
Kaiso is a unique POZ-Zinc finger proteinwith bimodal DNA-binding properties. It canrecognize methyl-CpGs (17), or sequence-specificnonmethylated DNA (18). In site-selectionexperiments, the preferred nonmethylated target ofKaiso is the sequence TNGCAGGA (18). In vitro,Kaiso binds this sequence with higher affinity thanmethylated DNA, but it is not known whether thispreference also exists in vivo (18). If Kaiso is abona fide binding partner of CTCF, we postulatedthat Kaiso may cooperate with, or antagonizeCTCF regulation of target genes. Since CTCFbinding to DNA is abrogated by DNA methylation(9-11), we focused our search on thenonmethylated sequence-specific Kaiso bindingsites (KBS). We examined known insulatorregions for the presence of this consensus site andfound one KBS 34 nucleotides upstream of theCTCF binding site in the human 5’HS5 beta-globin insulator (Fig. 4A). Of note, this region does
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

not contain any CpG dinucleotides and thereforecannot undergo DNA methylation. To test whetherthe presence of this Kaiso binding site affectedinsulation by CTCF, we performed an insulationassay using a characterized reporter construct, pNI(21). This plasmid contains three relevantelements: an enhancer (mouse beta-globin HS2), areporter gene, NeoR, that renders cells resistant toneomycin, and a cloning site between the enhancerand the reporter where test sequences can beinserted. After linearization the plasmid is used tostably transform mammalian cells that are thensubjected to neomycin selection. If the testsequence has no enhancer-blocking activity, NeoRis fully activated and many neomycin-resistantcolonies grow during the selection. In contrast, ifthe test sequence harbors an enhancer-blocker,NeoR is shielded from the enhancer, and fewneomycin-resistant colonies appear.
We tested four different sequences derivedfrom the human 5’HS5 insulator. All are 102nucleotides in length. The first sequence simplyreproduces a portion of the insulator containingboth the CTCF and Kaiso binding sites. The sitesare placed in the same orientation as in theendogenous locus: Kaiso is upstream on theenhancer side, and CTCF is on the target geneside. The second sequence differs in that it bearstwo point mutations in the Kaiso binding site.Previous studies have shown that these mutationstotally abrogate Kaiso binding in vitro (18). Thethird sequence contains two mutations that havebeen shown to prevent CTCF binding to its targetsite (5). Finally the fourth sequence contains themutant form of both the CTCF and the Kaisobinding site. All four sequences were inserted intopNI, used for stable transformation, and theneomycin-resistant colonies counted after selection(Fig. 4A). The experiments were performed in thehuman erythroleukemia cell line K562 where theHS2 enhancer is functional, and CTCF is presentand active. We verified by western blotting thatK562 cells also express Kaiso (data not shown).The number of colonies obtained by transfection ofunmodified pNI was the reference point of eachexperiment and was set at 100%. When cells weretransfected with an equivalent amount of pNIcontaining a known CTCF-dependent insulator,chicken 5’HS4, the number of neomycin-resistantcolonies was greatly decreased, to approximately
only 30% of that of pNI. This is in good agreementwith published values (21), and establishes that ourtest conditions are within the expected optimalparameters. The construct containing mutations inboth the CTCF and Kaiso binding sites yielded ahigh number of neomycin-resistant colonies, about80% that of empty pNI. This suggests that there isminimal enhancer-blocking activity in this testsequence. The construct containing a wild-typeKaiso binding site next to an inactivated CTCFbinding site also produced a high number ofneomycin-resistant colonies (80%). This showsthat the Kaiso binding site on its own does nothave enhancer-blocking potential. The constructcontaining a wild type CTCF binding site and amutant Kaiso site decreased the number ofcolonies to about 35% that of pNI. This reflects theknown enhancer-blocking activity of CTCF in thissequence. Again, the effect is of the samemagnitude as the published data (5). Finally wetested the unmodified sequence, bearing functionalCTCF and Kaiso binding sites. Strikingly thissequence had little or no enhancer-blockingpotential and yielded the same proportion (~75%)of neomycin-resistant colonies as the constructslacking an intact CTCF binding site (~80%). Bycomparison with the other constructs we concludethat the presence of an intact Kaiso-binding sitegreatly inhibits the enhancer-blocking activity ofCTCF at this locus.
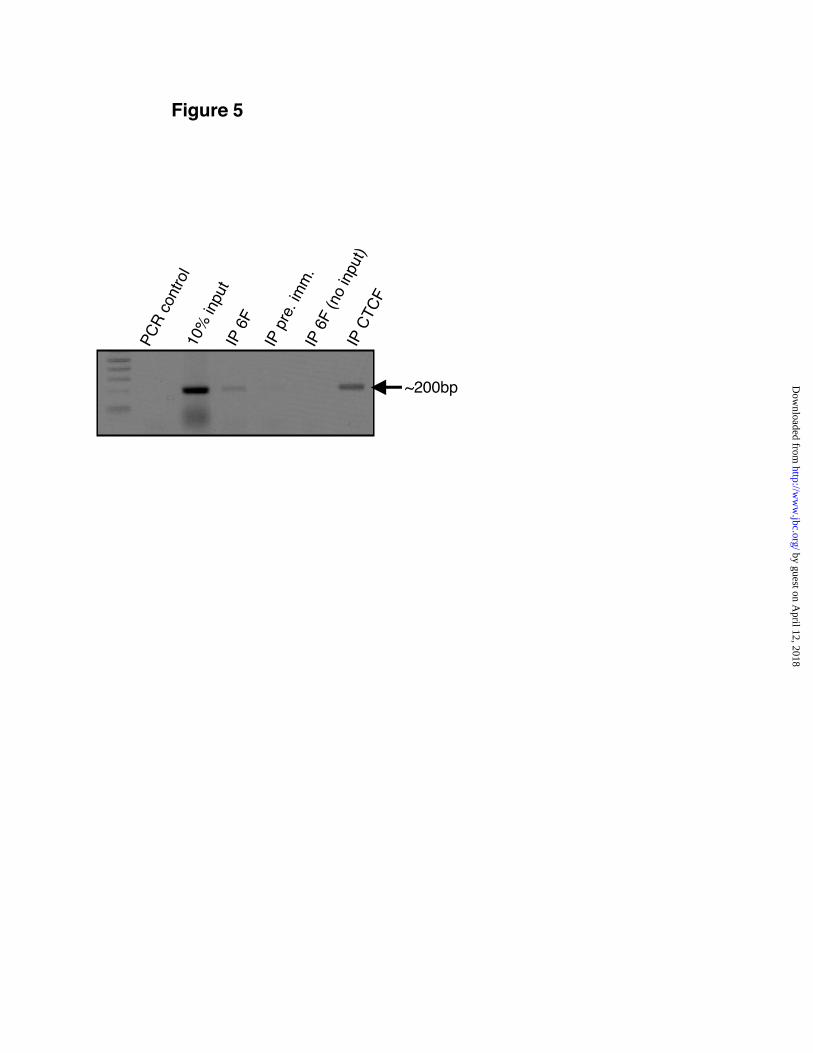
F ina l ly , we used chromat in -immunoprecipitation (ChIP) to test whether Kaisocan recognize its potential binding site in the beta-globin insulator. For these experiments we usedantibodies against CTCF and Kaiso that have beenvalidated for ChIP (15) (33) (27). As expected, thebeta-globin locus was recovered afterimmunoprecipitation with serum directed againstCTCF (Fig. 5). The same locus could also beamplified in Kaiso immunoprecipitates, but not inimmunoprecipitates obtained with a non-specificserum. This shows that Kaiso, like CTCF, bindsthe human 5’HS5 insulator in vivo.
DISCUSSION
Here we report an interaction between thevertebrate enhancer-blocker CTCF and thetranscription factor Kaiso. The interaction was alsodetected in an independent, reciprocal two-hybrid
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

screen that used Kaiso as the bait (C.L. Nordgaardand J.M.Daniel, unpublished results). There are atleast two prerequisites for this interaction to havephysiological meaning: CTCF and Kaiso should beco-expressed in some cell types, and they shouldhave an overlapping intracellular distribution.These two criteria are fulfilled by Kaiso andCTCF: both proteins predominantly localize to thenucleus and both proteins appear to beubiquitously expressed (18,34,35). Although thesetwo proteins may be coexpressed in many cells,this does not necessarily mean that they alwaysfunction in a complex. It is likely that only afraction of all CTCF proteins present in a cell areengaged in a complex with Kaiso, and vice versa.Furthermore, it is possible that the Kaiso-CTCFinteraction is regulated in a temporal manner.Clearly, further study is required to gain insightinto the upstream events or signals that regulatetheir in vivo interaction.
The interaction we have detected is highlyspecific: CTCF interacts with the POZ domain ofKaiso, but not with the POZ domain of four otherproteins (Bcl-6, PLZF, Hic-1 and FAZF), despitethe relatively high degree of conservation amongtheir respective POZ domains. The regionnecessary for interaction with Kaiso is located atthe CTCF C-terminus. This region is absent fromBoris, the closest paralogue of CTCF (36,37). It isthus likely that Kaiso interacts with specificallywith CTCF, but not with Boris. In addition, thisdomain of CTCF is particularly rich inphosphorylation sites that have been postulated toplay a regulatory role in CTCF-mediatedtranscriptional repression and possibly protein-protein interactions (38,39). Hence one possibilityis that the phosphorylation status of CTCFregulates its interaction with Kaiso.
Kaiso is a bi-modal transcription factorthat can bind methylated DNA (17), ornonmethylated targets containing the sequenceTNGCAGGA (18). To date, several putative Kaisotarget genes have been identified on the basis ofsuch Kaiso binding sites in their natural promoters.Kaiso activates the transcription of one target gene,rapsyn (27), but it represses the transcription of theother target genes, matrilysin, Wnt-11, and MTA2(33,40,41). For rapsyn, matrilysin, and Wnt-11,transcriptional regulation is mediated via the
conserved TNGCAGGA binding site whereas theMTA2 locus is regulated through Kaiso recognitionof methylated CpG sites. If Kaiso does indeed playa role in CTCF-mediated enhancer blocking, wepostulated that some CTCF target genes maypossess sequence-specific Kaiso binding sites ormethylated CpG dinucleotides. Indeed, we foundone conserved Kaiso binding site in the human 5’HS5 insulator at the beta-globin gene cluster. Thepresence of the Kaiso Binding Site stronglydecreases the enhancer-blocking effect of CTCFand implicates Kaiso as a regulator of CTCFfunction.
Our data could explain the in vivo findingsof other investigators. Grosveld and co-workershave investigated the behavior of the human beta-globin insulator in transgenic mice (42). Theirresults show that the insulator is active inembryonic erythroid tissues, but is inactive in othertissues. In tissues where the insulator is inactive,the footprint of a protein is detected over thepotential Kaiso binding site TGGCAGGA; intissues where the insulator is active, this footprintis missing. We have detected binding of Kaiso tothe beta-globin insulator in human cells, and wesuggest that Kaiso is the protein that inhibits theactivity of CTCF in these in vivo experiments.
Kaiso could act on CTCF by variousmechanisms. The simplest possibility would bethat Kaiso inhibits the binding of CTCF to itstarget site. Such a situation has been described inthe case of FBI-1, a POZ domain transcriptionalrepressor. The POZ domain of FBI-1 binds theZinc fingers of Sp1, and impedes its DNA-bindingactivity (43). We do not believe this mechanismapplies here, as CTCF is readily detected at the 5’HS5 insulator both in cultured cells (this work),and in vivo (42). An alternative hypothesis relatesto the proposed mode of action of CTCF.Felsenfeld and co-workers have shown that CTCFrecruits insulated loci to the nuclear matrix at thesurface of the nucleolus (12,13). It may be thatKaiso interferes with this recruitment step,possibly by redirecting the locus to another nuclearsite.
What could be the biological relevance ofthis regulation? We hypothesize that Kaiso mightbe used to downregulate the activity of CTCF at
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

certain times or in certain cell types whenenhancer-blocking is not desirable. Interestingly,the Kaiso target site is not conserved in the mousegenome. It is possible that Kaiso binds a differentsequence on the mouse chromosome, or thatregulation of the insulator by the Kaiso site isspecific to humans. This could relate to the factthat the human 5’HS5 sequence functions as aninsulator in vivo, while the mouse sequence doesnot (5,44,45). Finally, we note that the interactiondescribed here, in addition to its effect on CTCFfunction, could also regulate the activity of Kaiso.The adhesion molecule p120ctn can translocatefrom the cytosol to the nucleus, where it inhibitstranscriptional repression by Kaiso (18,26). ThePOZ domain of Kaiso mediates interaction withCTCF, and also permits homodimerization (16).
Interaction with CTCF could possibly influenceKaiso homodimerization, which in turn couldmodify the transcriptional activity of Kaiso.
The regulation of CTCF activity by Kaisois reminiscent of another situation : it has beenshown that a subset of CTCF target sites areadjacent to binding sites for the Thyroid HormoneReceptor (TR), and that, in this context, theinsulating activity of CTCF is modulated by theactivity of TR (15). While the mechanism of thisregulation is not understood, it could indicate thatCTCF-dependent insulators are not generallyconstitutive, but can be fine-tuned to regulate thetranscriptional activity of the genome.
REFERENCES
1. Labrador, M., and Corces, V. G. (2002) Cell 111, 151-1542. West, A. G., Gaszner, M., and Felsenfeld, G. (2002) Genes Dev 16, 271-288.3. Li, Q., Peterson, K. R., Fang, X., and Stamatoyannopoulos, G. (2002) Blood 100, 3077-30864. Burgess-Beusse, B., Farrell, C., Gaszner, M., Litt, M., Mutskov, V., Recillas-Targa, F., Simpson,
M., West, A., and Felsenfeld, G. (2002) Proc Natl Acad Sci U S A 99 Suppl 4, 16433-164375. Farrell, C. M., West, A. G., and Felsenfeld, G. (2002) Mol Cell Biol 22, 3820-3831.6. Klenova, E. M., Nicolas, R. H., Paterson, H. F., Carne, A. F., Heath, C. M., Goodwin, G. H.,
Neiman, P. E., and Lobanenkov, V. V. (1993) Mol Cell Biol 13, 7612-7624.7. Ohlsson, R., Renkawitz, R., and Lobanenkov, V. (2001) Trends Genet 17, 520-527.8. Kim, J., Kollhoff, A., Bergmann, A., and Stubbs, L. (2003) Hum Mol Genet 12, 233-2459. Bell, A. C., and Felsenfeld, G. (2000) Nature 405, 482-485.10. Hark, A. T., Schoenherr, C. J., Katz, D. J., Ingram, R. S., Levorse, J. M., and Tilghman, S. M.
(2000) Nature 405, 486-489.11. Kanduri, C., Pant, V., Loukinov, D., Pugacheva, E., Qi, C. F., Wolffe, A., Ohlsson, R., and
Lobanenkov, V. V. (2000) Curr Biol 10, 853-856.12. Yusufzai, T. M., and Felsenfeld, G. (2004) Proc Natl Acad Sci U S A 101, 8620-862413. Yusufzai, T. M., Tagami, H., Nakatani, Y., and Felsenfeld, G. (2004) Mol Cell 13, 291-29814. Yu, W., Ginjala, V., Pant, V., Chernukhin, I., Whitehead, J., Docquier, F., Farrar, D.,
Tavoosidana, G., Mukhopadhyay, R., Kanduri, C., Oshimura, M., Feinberg, A. P., Lobanenkov,V., Klenova, E., and Ohlsson, R. (2004) Nat Genet 36, 1105-1110
15. Lutz, M., Burke, L. J., LeFevre, P., Myers, F. A., Thorne, A. W., Crane-Robinson, C., Bonifer, C.,Filippova, G. N., Lobanenkov, V., and Renkawitz, R. (2003) Embo J 22, 1579-1587
16. Daniel, J. M., and Reynolds, A. B. (1999) Mol Cell Biol 19, 3614-3623.17. Prokhortchouk, A., Hendrich, B., Jorgensen, H., Ruzov, A., Wilm, M., Georgiev, G., Bird, A., and
Prokhortchouk, E. (2001) Genes Dev 15, 1613-1618.18. Daniel, J. M., Spring, C. M., Crawford, H. C., Reynolds, A. B., and Baig, A. (2002) Nucleic Acids
Res 30, 2911-2919.19. James, P., Halladay, J., and Craig, E. A. (1996) Genetics 144, 1425-1436.20. Defossez, P. A., and Gilson, E. (2002) Nucleic Acids Res 23, 5136-514121. Bell, A. C., West, A. G., and Felsenfeld, G. (1999) Cell 98, 387-396.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22. Le Douarin, B., Pierrat, B., vom Baur, E., Chambon, P., and Losson, R. (1995) Nucleic Acids Res23, 876-878
23. Gietz, R. D., and Woods, R. A. (2002) Methods Enzymol 350, 87-9624. Klenova, E., Chernukhin, I., Inoue, T., Shamsuddin, S., and Norton, J. (2002) Methods 26, 254-
259.25. Daniel, J. M., Ireton, R. C., and Reynolds, A. B. (2001) Hybridoma 20, 159-166.26. Kelly, K. F., Spring, C. M., Otchere, A. A., and Daniel, J. M. (2004) J Cell Sci 117, 2675-268627. Rodova, M., Kelly, K. F., VanSaun, M., Daniel, J. M., and Werle, M. J. (2004) Mol Cell Biol 24,
7188-719628. Chernukhin, I. V., Shamsuddin, S., Robinson, A. F., Carne, A. F., Paul, A., El-Kady, A. I.,
Lobanenkov, V. V., and Klenova, E. M. (2000) J Biol Chem 275, 29915-2992129. Bardwell, V. J., and Treisman, R. (1994) Genes Dev 8, 1664-167730. Collins, T., Stone, J. R., and Williams, A. J. (2001) Mol Cell Biol 21, 3609-3615.31. Melnick, A., Carlile, G., Ahmad, K. F., Kiang, C. L., Corcoran, C., Bardwell, V., Prive, G. G., and
Licht, J. D. (2002) Mol Cell Biol 22, 1804-181832. Melnick, A., Ahmad, K. F., Arai, S., Polinger, A., Ball, H., Borden, K. L., Carlile, G. W., Prive,
G. G., and Licht, J. D. (2000) Mol Cell Biol 20, 6550-656733. Yoon, H. G., Chan, D. W., Reynolds, A. B., Qin, J., and Wong, J. (2003) Mol Cell 12, 723-73434. Filippova, G. N., Lindblom, A., Meincke, L. J., Klenova, E. M., Neiman, P. E., Collins, S. J.,
Doggett, N. A., and Lobanenkov, V. V. (1998) Genes Chromosomes Cancer 22, 26-36.35. Filippova, G. N., Fagerlie, S., Klenova, E. M., Myers, C., Dehner, Y., Goodwin, G., Neiman, P.
E., Collins, S. J., and Lobanenkov, V. V. (1996) Mol Cell Biol 16, 2802-2813.36. Loukinov, D. I., Pugacheva, E., Vatolin, S., Pack, S. D., Moon, H., Chernukhin, I., Mannan, P.,
Larsson, E., Kanduri, C., Vostrov, A. A., Cui, H., Niemitz, E. L., Rasko, J. E., Docquier, F. M.,Kistler, M., Breen, J. J., Zhuang, Z., Quitschke, W. W., Renkawitz, R., Klenova, E. M., Feinberg,A. P., Ohlsson, R., Morse, H. C., 3rd, and Lobanenkov, V. V. (2002) Proc Natl Acad Sci U S A99, 6806-6811
37. Klenova, E. M., Morse, H. C., 3rd, Ohlsson, R., and Lobanenkov, V. V. (2002) Semin Cancer Biol12, 399-414
38. Klenova, E. M., Chernukhin, I. V., El-Kady, A., Lee, R. E., Pugacheva, E. M., Loukinov, D. I.,Goodwin, G. H., Delgado, D., Filippova, G. N., Leon, J., Morse, H. C., 3rd, Neiman, P. E., andLobanenkov, V. V. (2001) Mol Cell Biol 21, 2221-2234.
39. El-Kady, A., and Klenova, E. (2005) FEBS Lett 579, 1424-143440. Spring, C. M., Kelly, K. F., O'Kelly, I., Graham, M., Crawford, H. C., and Daniel, J. M. (2005)
Exp Cell Res In press41. Kim, S. W., Park, J. I., Spring, C. M., Sater, A. K., Ji, H., Otchere, A. A., Daniel, J. M., and
McCrea, P. D. (2004) Nat Cell Biol 6, 1212-122042. Wai, A. W., Gillemans, N., Raguz-Bolognesi, S., Pruzina, S., Zafarana, G., Meijer, D., Philipsen,
S., and Grosveld, F. (2003) Embo J 22, 4489-450043. Lee, D. K., Suh, D., Edenberg, H. J., and Hur, M. W. (2002) J Biol Chem 277, 26761-2676844. Tanimoto, K., Sugiura, A., Omori, A., Felsenfeld, G., Engel, J. D., and Fukamizu, A. (2003) Mol
Cell Biol 23, 8946-895245. Li, Q., Zhang, M., Han, H., Rohde, A., and Stamatoyannopoulos, G. (2002) Nucleic Acids Res 30,
2484-2491
FOOTNOTES
Acknowledgements-We thank the following colleagues for the gift of reagents: Dominique Leprince, GaryFelsenfeld, Claire Francastel, Elena Klenova, David Shore, Régine Losson, Phillip James, RainerRenkawitz. PA Defossez is supported by an ATIP from CNRS. JM Daniel is supported by a grant fromthe Canadian Institutes of Health Research (MOP 42045), and K.F. Kelly is the recipient of an NSERC
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D2 Doctoral scholarship. Work in the Gilson lab is supported by Ligue Nationale contre le Cancer (Equipelabéllisée).
1 The abbreviations used are: CTCF, CTC-binding factor; POZ, Poxvirus and Zinc finger; KBS, Kaiso-binding site.
FIGURE LEGENDS
Fig. 1. Two-hybrid interaction between CTCF and the BTB/POZ domain of Kaiso. A, Clones recoveredin the two-hybrid screen with CTCF. Mouse Kaiso contains a BTB/POZ domain (boxed), and three Zincfingers (ZF). Two plasmids containing overlapping inserts were obtained in the screen. The peptidescontain 79 additional amino acids encoded by the 5’UTR of Kaiso (dashed line). B, The POZ domain ofKaiso mediates interaction with CTCF. Transformants containing Gal4-CTCF and the indicated preyplasmids were selected on medium lacking uracil and leucine (-UL, top). Interaction between the proteinspermits growth on medium lacking histidine (-ULH, bottom). POZH is the POZ domain of the humanprotein HIC1. C, CTCF interacts with Kaiso, but not with other POZ domain proteins. Two-hybridinteraction between Gal4-CTCF and the indicated mouse POZ domains was tested as in B.
Fig. 2. CTCF and Kaiso interact in vivo and in vitro. A, To determine whether Kaiso and CTCF interact invivo, nuclear extracts prepared from HeLa cells were subjected to immunoprecipitation with Kaiso- andCTCF-specific antibodies as indicated. The immune complexes were resolved via SDS-PAGE, and usedfor Western blotting with antibodies directed against Kaiso or CTCF. Kaiso was captured using varioushighly-specific Kaiso monoclonal antibodies. CTCF co-precipitated robustly with endogenous Kaisofrom these extracts, but not with the negative control antibody (pre-immune serum). B, Interactionbetween bacterially expressed GST-Kaiso and in vitro transcribed and translated CTCF. Input : 5% of theinput CTCF. The arrow marks the position of the full-length CTCF protein.
Fig. 3. Mapping the interaction domain in CTCF and Kaiso. A, Various domains of CTCF were fused toGAL4, and their interaction with the POZ domain of Kaiso was assessed in a strain where LacZ is thereporter. B, Point mutations were introduced in the POZ domain of Kaiso. Yeast cells containing theindicated combination of plasmids were plated on control medium (-UL), or on medium that selects forinteraction (-ULH).
Fig. 4. A Kaiso binding site at the beta-globin insulator regulates enhancer blocking by CTCF. A,Structure of the human globin-gene locus (not to scale). A consensus Kaiso binding site is present next tothe CTCF binding site in the 5’HS5 insulator. B, Activity of wild-type and mutant sequences in amammalian insulation assay. Human K562 cells were transfected with the indicated constructs, andseeded in soft agar in the presence of neomycin. The number of colonies was recorded three weeks aftertransformation. C, Model explaining the behavior of the different constructs.
Fig. 5. Kaiso binds the beta-globin insulator in vivo. Hela cells were fixed with formaldehyde, andgenomic DNA was isolated and fragmented by sonication. Kaiso was immunoprecipitated by using the 6Fmonoclonal antibody. A 200-bp fragment of the beta-globin insulator was amplified by PCR from Kaisoimmunoprecipitates (IP 6F) and from CTCF immunoprecipitates (IP CTCF). Negligible amounts of thissequence were amplified from the immunoprecipitation carried out with pre-immune antibody (IP pre.imm.). To verify lack of contamination two other controls were used. First, the PCR reaction was doneusing water as the template (PCR control). Second, an immunoprecipitation reaction was done in theabsence of input chromatin (no input). For the positive control (10% input), the beta-globin insulator wasamplified directly from Hela genomic DNA, which was purified from 1/10 of the amount of lysate thatwas used for each Kaiso immunoprecipitation. IP, immunoprecipitate.
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
WB : Kaiso
WB : CTCF
Extr.
P.I.
CTCF
11D
2G 12H
12G
6F
Kaiso mAb
IP
173 kD
111 kD
111 kD
Figure 2
A
B
GST
Inpu
t
GST
-Kai
so
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
B
POZWT
K47P
D33N
D33NK47Q
K47QY86A
K47R
Y86A
K47QAD>
Gal4-CTCF
Gal4
-UL
-ULH
Neg.
Gal4-CTCF
Gal4
A
1 728
1
269 577
578 728
FL
NZF
ZF
Cter
Gal4
577
578 641Cteralpha
641 728Cterbeta
+AD +AD-POZ
Figure 3
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
A
B
TCTGGCAGGACTCTTTAGAGAGGTAGCTGAAGCTGCTGTTATGACCACTAGAGGGAAGAAGACTCFKaiso
ε Gγ Aγ δ β5’HS 5 3’HS 1
NeoREnh
Kaiso CTCF
Kaiso CTCF
Kaiso CTCF
Kaiso CTCF
Number of colonies(% of control)
0 20 40 60 80 100
Figure 4
Kaiso CTCFEnh
Kaiso CTCFEnh
C
Kaiso CTCFEnh
Kaiso CTCFEnh
P<0,005
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
PCR
contr
ol10
% in
put
IP 6
FIP
pre.
imm.
IP 6F
(no i
nput)
IP C
TCF
~200bp
Figure 5
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

GilsonFrédérique Magdinier, Hervé Menoni, Curtis L. Nordgaard, Juliet M. Daniel and Eric Pierre-Antoine Defossez, Kevin F. Kelly, Guillaume Filion, Roberto Pérez-Torrado,
The human enhancer-blocker CTCF interacts with the transcription factor kaiso
published online October 17, 2005J. Biol. Chem.
10.1074/jbc.M510802200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from