Embed Size (px)
Citation preview

JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR
THE MATCHING LAW IN AND WITHIN GROUPSOF RATS1
D. A. GRAFT, S. E. G. LEA, AND T. L. WHITWORTH
UNIVERSITY OF CAMBRIDGE
In each of the two experiments, a group of five rats lived in a complex maze containingfour small single-lever operant chambers. In two of these chambers, food was available onvariable-interval schedules of reinforcement. In Experilnent I, nine combinations of varia-ble intervals were used, and the aggregate lever-pressing rates (by the five rats together)were studied. The log ratio of the rates in the two chambers was linearly related to the logratio of the reinforcenment rates in them; this is an instance of Herrnstein's matching law,as generalized by Baum. Suinming over the two food chambers, food consumption de-creased, and response output increased, as the time required to earn each pellet increased.In Experiment II, the behavior of individual rats was observed by time-sampling on se-lected days, while different variable-iinterval schedules were arranged in the two chamberswhere food was available. Individual lever-pressing rates for the rats were obtained, andtheir median bore the same "matching" relationship to the reinforcement rates as the groupaggregate in Experimenit I. There were differences between the rats in their distributionof time and responses between the two food chambers; these differences were correlatedwith differences in the proportions of reinforcements the rats obtained from each chamber.Key words: matching law, group operant, imulti-operant, economic behavior, demand
curve, shopping centers, variable interval, lever press, rats
In its simplest form, Herrnstein's (1970)matching law states that if two schedules ofreinforcement are concurrently available, ratesof responding on them are in the same propor-tion as reinforcement rates. The law summa-rizes much data on choice, especially choiceby pigeons between two concurrently availableschedules of reinforcement, each imposing avariable interval between reinforcements (concVI VI schedules). There are exceptions, butthey mainly concern choice between schedulesof different types (e.g., Nevin, 1971; Trevett,Davison, and Williams, 1972). At least some ofthe exceptions can be accommodated by a gen-
"The apparatus was constructed by H. M. Pamphilon.The ONLI system used in Experiment II was developedin collaboration with C. K. Crook, with the support ofGrant Number G970/297/B to A. J. Watson from theU.K. Medical Research Council. Thanks are due toD. J. Nicholas for help in conducting the experimentsand to R. R. Miller, M. J. Morgan, and T. J. Roperfor comments on the manuscript. Results from theseexperiments were presented to the meetings of the Ex-perimental Analysis of Behaviour Group at Exeter (Eas-ter 1975), and of the Experimental Psychology Societyat Durham (Easter 1976). D. A. Graft was supported bythe H. E. Durham Fund of King's College, Cambridge.Reprints may be obtained from S. E. G. Lea, Univer-sity of Exeter, Department of Psychology, Washington-Singer Laboratories, Exeter EX4 4QG, England.
eralization of the matching law proposed byBaum (1974b), according to which
log (R1/R2) = a log (r,/r2) + b (1)
where Ri and ri are the response and reinforce-ment rates on the ith schedule and a and b arefitting constants. Lobb and Davison (1975) sug-gested that a is usually in the range 0.8 to 1.0;the simple matching law would require it toequal 1.0 always.We are concerned with the relation between
psychological experiments on choice, and thechoices made by consumers in the course oftheir economic behavior. The present paperdescribes experiments with conc VI VI sched-ules under conditions that we tried to makesomewhat like those of the real economy. Morethan one subject had access to the apparatusat a time, the subjects had to obtain all theirfood by working on the schedules, and accessto the schedules was unrestricted. Our purposewas to see whether the matching law wouldstill be useful in this "real-life" (or nearer-to-real-life) situation.Should the matching law be expected to
hold in these experiments? On the one hand,some schedule effects survive relaxation ofstandard experimental conditions. Grott and
183
1977, 25, 183-194 NUMBER I (JANUARY)

184
Neuringer (1974) showed that a group of ratsbehaves roughly like a single rat on some
standard schedules of reinforcement; Morgan,Fitch, Holman, and Lea (1976) used multiple-interval schedules to teach higher-order con-
cepts to free-living pigeons with unrestrictedaccess to the schedules; and Baum (1972,1974a) obtained matching from a single pi-geon with unrestricted access to conc VI VI,and from free-living pigeons with virtuallyunrestricted access (only one pigeon could oc-
cupy the apparatus at a time). On the otherhand, if different subjects can command twomanipulanda, the situation might approxi-mate a simple, rather than a concurrent sched-ule: as soon as reinforcement is available on
each schedule, it might be taken by the sub-ject commanding the corresponding manipu-landum. Under these conditions there wouldbe little reason to expect the interaction be-tween schedules that gives rise to matching.
EXPERIMENT I
In the first experiment, the aggregate be-havior of a group of rats was studied undera range of conc VI VI schedules.
METHODSubjects
Five female hooded rats were randomly se-
lected from a group of 10 obtained from Ani-mal Suppliers Ltd, London, aged threemonths. The remainder were kept as reserves.
ApparatusFigure 1 shows a scale drawing of a vertical
cross-section through the apparatus, and de-fines the names used for its parts. The stair-case and chambers were made of sheet alumi-num, the floors were of 1.3-cm galvanizedmesh, and the tubes were cylinders of clearplastic. The floors and tubes could be liftedout for washing; when this was done, spareswere substituted. The staircase was lit by0.6-W bulbs in flush mountings fixed to therear wall at the heights indicated. The frontof the staircase was formed of a neutral den-sity plastic door, divided horizontally at thelevel of stair 4, and hinged at the left. Thefront of each chamber was a neutral densityplastic door, hinged at the right. Sheet alum-inum panels could be inserted to cut off thetubes from the staircase or the chambers. The
D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
rstalr" tu a chamberS
ir6- ..s
staircase
ftar2_
..........
laz t z l z z r
-~~~~~~~~~~~~~~~~I=. ........
Iflo>jrlds
121n.O 30cm
Fig. 1. Vertical cross-section through the apparatus,drawn to scale except for the thicknesses of walls. Notethat the tubes were cylindrical, and that the depth ofthe staircase and chambers was 15 cm. The staircaselamps were mounted on the rear wall.
sliding panels that formed the right-handwalls of the chambers were either blank sheetsof aluminum, or operant test units of the de-sign illustrated in Figure 2. The 3-W lampabove the lever (barlight) was mounted flush.The foodtray was recessed, could be lit frombehind and above by a 3-W traylight, and was
closed by a translucent flap (panel) that oper-
ated a microswitch. The retractable lever op-
erated a sealed-reed switch when depressed
0 4in.i
II
I Io ibcm
Fig. 2. Elevation of the operant panels used in some
chambers, drawn to scale.
MATCHING IN GROUPS OF RATS
with a force adjustable between 0.05 and0.09 N. The foodtray could be supplied eitherwith liquid from a 0.05-ml dipper or with45-mg pellets. The levers, foodtrays, dippers,and pellet feeders were obtained commercially(Campden Instruments Ltd, London), but thepellet feeders were modified to take severalthousand pellets, and hydrostatic systems were
used to keep the dipper reservoirs topped up
from 500-ml flasks.The apparatus was housed in an air-condi-
tioned room, which was lit by fluorescenttubes from 2000 to 0800 hr, and by dim redlight at other times. Ambient temperaturewas maintained at 21°C. Masking noise was
not used. Conventional electromechanical pro-
gramming apparatus was in the same room,
but out of sight from the chambers. It in-cluded filmstrip timers to control the variableintervals; the rats were watched periodicallyfor signs that they were detecting the clicksfrom these timers, but no evidence was foundthat they could discriminate significant noisesamong the general electromechanical clatter.
PretrainingThe rats were not deprived of food or water
before they were first put into the apparatus.On the first day, 10 g of standard laboratorydiet was put in each chamber, and a waterbottle was fixed in Chamber 1; all chambershad blank panels at their right ends. Duringthe next 20 days, feeder panels were installedin Chambers 2 and 5 and dipper panels inChambers 3 and 4, and the rats were trainedto obtain all their food and water by press-
ing the levers in those chambers on a continu-ous reinforcement schedule: a single leverpress operated the feeder, turned the traylighton and the barlight off; the lights changedback again when the tray flap next closed,that is, when a rat next withdrew its headfrom the foodtray. No "shaping" was done,but the contingencies were introduced grad-ually: the operant panels were installed one
at a time, lever training was preceded bymagazine training (in which the feeders were
operated at regular intervals, or whenever thefoodtray panel closed), and the lever forcerequirements were initially set to a minimum.The interchange of signal lights at pellet or
water delivery was used throughout the ex-
periment. At the end of pretraining, all leverforce requirements were set to the maximum.
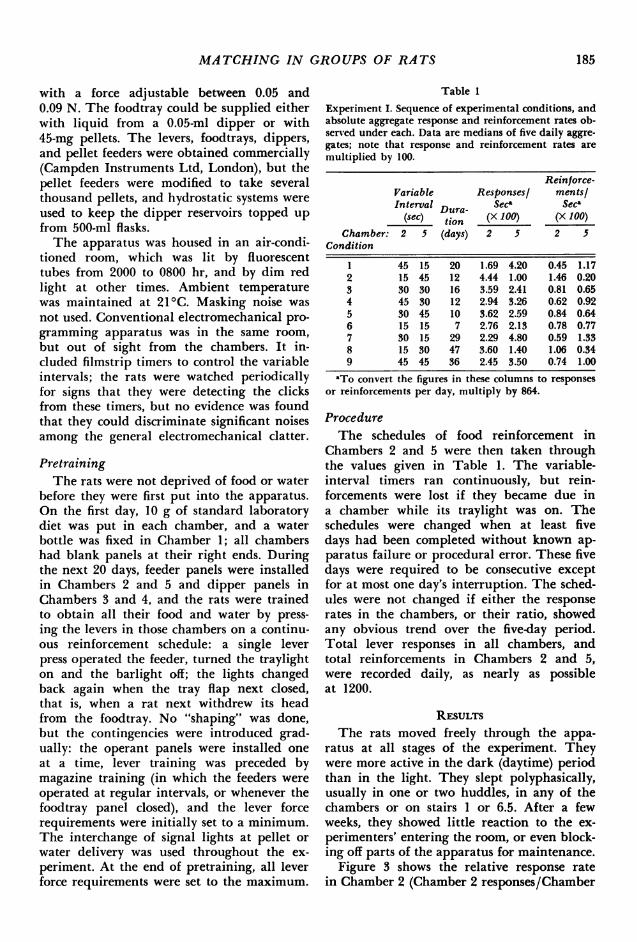
Table 1
Experiment I. Sequence of experimental conditions, andabsolute aggregate response and reinforcement rates ob-served under each. Data are medians of five daily aggre-gates; note that response and reinforcement rates aremultiplied by 100.
Reinforce-Variable Responsesl ments/Interval Dura- Sect Seca
(sec) tion (X 100) (X 100)Chamber: 2 5 (days) 2 5 2 5
Condition
1 45 15 20 1.69 4.20 0.45 1.172 15 45 12 4.44 1.00 1.46 0.203 30 30 16 3.59 2.41 0.81 0.654 45 30 12 2.94 3.26 0.62 0.925 30 45 10 3.62 2.59 0.84 0.646 15 15 7 2.76 2.13 0.78 0.777 30 15 29 2.29 4.80 0.59 1.338 15 30 47 3.60 1.40 1.06 0.349 45 45 36 2.45 3.50 0.74 1.00
aTo convert the figures in these columns to responsesor reinforcements per day, multiply by 864.
ProcedureThe schedules of food reinforcement in
Chambers 2 and 5 were then taken throughthe values given in Table 1. The variable-interval timers ran continuously, but rein-forcements were lost if they became due ina chamber while its traylight was on. Theschedules were changed when at least fivedays had been completed without known ap-paratus failure or procedural error. These fivedays were required to be consecutive exceptfor at most one day's interruption. The sched-ules were not changed if either the responserates in the chambers, or their ratio, showedany obvious trend over the five-day period.Total lever responses in all chambers, andtotal reinforcements in Chambers 2 and 5,were recorded daily, as nearly as possibleat 1200.
RESULTSThe rats moved freely through the appa-
ratus at all stages of the experiment. Theywere more active in the dark (daytime) periodthan in the light. They slept polyphasically,usually in one or two huddles, in any of thechambers or on stairs 1 or 6.5. After a fewweeks, they showed little reaction to the ex-perimenters' entering the room, or even block-ing off parts of the apparatus for maintenance.
Figure 3 shows the relative response ratein Chamber 2 (Chamber 2 responses/Chamber
185

D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
30~~~~~~~~~
1 00
10 20 30dep
40
Fig. 3. Experiment I. Proportion of all lever pressesmade in food chambers that were made in Chamber 2,for the first three schedule conditions. Each point showsdata from one day: open circles are used for days wherethere was a known apparatus failure or procedural er-ror. The broken horizontal lines show the programmedrelative reinforcement rates in Chamber 2; in order,the schedules in Chamber 2 in the three conditionswere VI 45-, 15-, and 30-sec.
2 + Chamber 5 responses) over the first threeconc VI-VI conditions, illustrating the courseof adaptation to new schedules and the vari-ability of relative response rate within con-ditions.Data are presented below as medians over
the last five errorless days in each condition.Figure 4 shows the ratio of the response ratesin Chambers 2 and 5, as a function of theratio of reinforcement rates. The data areplotted on logarithmic scales as recommendedby Baum (1974b); the figure includes the lin-ear regression of Y on X, and the line pre-dicted by the simplest version of the match-ing law. Ratios of reinforcement rates can bederived in two ways, and results are shown interms of both the programmed ratio, derivedfrom the variable-interval schedules used, andthe observed ratio, derived from the numberof reinforcements actually delivered in the twochambers. The regression lines were obtainedby standard techniques; it could be arguedthat these are not strictly appropriate for theobserved reinforcement ratios, which are sub-ject to errors of observation. A method appro-priate in this case is given by Kendall andStuart (1967, paragraph 29.20), but even un-der the worst-case assumption that the rein-forcement ratio suffers from as much error asthe response ratio, very much the same resultis obtained as by the simple method used here.Both by standard or by more sophisticatedmethods, the regression line for the obtainedreinforcement ratios has a slope significantlyless than 1.0 (p < 0.05).
Figure 5 shows three functions relevant tothe attempt to analyze the situation as a model
.1 .2 .5 1 2 5 10reinforcement rate ratio
Fig. 4. Experiment I. Ratio of lever-press rate inChamber 2 to that in Chamber 5, as a function of theratio of reinforcement rates, on logarithmic coordinates.Data are medians of five daily aggregates. Filled circlesand the broken regression line are for data plottedagainst ratios of programmed reinforcement rates, opencircles and the dotted regression line are for dataplotted against ratios of obtained reinforcement rates.The solid diagonal line gives the prediction from thesimple matching law.
economy. The abscissa is the mean time thathad to elapse, according to the schedules, be-tween successive pellet deliveries anywhere inthe apparatus; it may be thought of as theaverage "price" of a food pellet in time units.Data for continuous reinforcement are plotted
.075
S.05
.50I~02a
,'*-- ,4
-f''''6''DS'o '-8 03 <O* -~; .....A --O
0. .... ........... ..... -
I
15 _\-
i5-
7s 10t25 15 S 22.5mmnmum work Ume4 ebt 'scs'
Fig. 5. Experiment I. Response rates, reinforcementrate, and estimated time committed to working forfood, as a function of the programmed minimum timeb)etween pellet deliveries. Data are medians of five dailyaggregates; the left-most points are disconnected be-cause the abscissa value is a rough estimate.
186

MATCHING IN GROUPS OF RATS
against a "price" of 1 sec, though this is obvi-ously an idealization. The ordinate shows theaggregate rate of earning pellets, the aggre-gate rate of leverpressing, and an estimate ofthe proportion of the day committed to earn-ing food per rat. The first two of these weretotalled across Chambers 2 and 5; correspond-ing data for the individual chambers are re-ported in Table 1. The time committed wasestimated by multiplying the number of pel-lets earned in each chamber by the variableinterval in force there, adding the results forthe two chambers together, and dividing bythe number of rats and the length of the day.
DIscUSSIONThe log ratio of response rates was approxi-
mately equal to the log ratio of programmedreinforcement rates (Figure 4), despite consid-erable day-to-day variation in relative responserates, and not infrequent apparatus trouble(Figure 3). When obtained reinforcement-rateratios were used, response-rate ratios did notmatch them exactly; the slope of the regressionline was less than one, a condition that Baum(1974b) called "undermatching". Lobb andDavison (1975) found a similar degree of un-dermatching in experiments on pigeons withconc VI VI schedules, and they argued thatundermatching of response ratios to obtainedreinforcement ratios is typical. Whether pro-grammed or obtained reinforcement ratioswere used, the coefficients of linear correlationin the present experiment were high (0.95 and0.98 respectively). Baum's (1974b) generaliza-tion of the matching law therefore accountedfor almost all the variance in the data, andthe results were quantitatively and qualita-tively consistent with those from more conven-tional apparatus and more conventional sub-jects (investigations of matching with rats arerare; for an example, see Mazur, 1975).There are two reasons for caution about the
results. First, Herrnstein (1970) pointed outthat when response and reinforcement ratesare similar, matching to obtained reinforce-ment rates may be produced artifactually,since for a reinforcement to be obtained aresponse must occur. Herrnstein investigatedthe limits that this constraint imposes on rela-tive response rate; his formulae can be adaptedfor log response ratios, and the areas wherethe log response ratios could not lie have beenshaded in Figure 4. It is clear from the figure
that this artifact was not responsible for thegood fit of the generalized matching law.A more serious problem is that the obtained
reinforcement rates were an order of magni-tude less than the programmed rates. The pos-sibility therefore exists that the obtained rein-forcement ratios did not cause, but werecaused by, the rats' allocation of time andresponding to the two chambers. For thisreason, we lay more stress on the match ofresponse-rate ratio to programmed reinforce-ment ratio (which is directly manipulable),than on the very nearly linear relationshipof log response ratio to log obtained reinforce-ment ratio. The coincidence between the lat-ter relation and that observed by Lobb andDavison (1975) is undoubtedly striking; butunder plausible assumptions, it can be shownthat exact matching of response ratio to pro-grammed reinforcement ratio, and a low rateof switching between chambers, imply a linearrelationship between log response ratio andlog obtained reinforcement ratio, with a slopebetween 0.5 and 1.0. The observed value (0.80)is in the middle of this range. Even if theorigin of the relationship to observed rein-forcement ratios is uncertain, however, thenear match between response ratios and pro-grammed reinforcement ratios is sufficient toestablish that the present results are consistentwith the published literature on conc VI VI.One reason why the obtained reinforcement
ratio is normally used is that it is the variablemost likely to be controlling behavior; thesubject cannot know the schedules except bythe reinforcements they yield. In the presentsituation, the variable controlling behaviorcould well have been the reinforcement ratesexperienced during long, uninterrupted peri-ods of working in single chambers; since suchperiods would not have been of any standardlength, it is plausible that the rats shouldhave learned about the programmed reinforce-ment rates independently.
It remains to show that the experimentalsituation did function in some respects as amodel economy. Figure 5 shows that, as theaverage cost of a pellet increased, the rats de-creased their total intake of food. The timerequired to earn what food they took in-creased, and so did the number of responsesmade. The first two of these trends were sig-nificant (Kendall's tau values of -0.49 and0.71, p < 0.05), but the striking aspect of the
187

D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
data is that there was very little differencein food intake between the conc VI VI condi-tions. The only substantial drop in intakeoccurred on going from continuous reinforce-ment to conc VI VI.Time required to earn a pellet is chosen
as the analog of price in this situation because,like money, time spent on working for onecommodity cannot be spent on working foranother (cf. Becker, -1965). If the analogy isaccepted, the open circles of Figure 5 repre-sent the economist's demand curve, the func-tion that relates the quantity of a good con-sumed to its price (cf. Lancaster, 1969, p. 13).The fact that the filled squares form an in-creasing function means that demand for foodwas "inelastic" (strictly, had elasticity lessthan one); that is, a given percentage priceincrease caused a smaller percentage decreasein quantity consumed, and hence as price in-creased so did the expenditure of time on food.Inelastic demand for food has been found ina number of studies in which isolated animalslived in operant apparatus and obtained alltheir food on fixed-ratio schedules (Collier,Hirsch, and Hamlin, 1972; Hirsch and Collier,1974; Hogan, Kleist, and Hutchings, 1970,Experiment lb; Logan, 1964). It is also foundin the real economy: if all food is consideredtogether, demand for it is inelastic, and so isdemand for the majority of individual food-stuffs (Stone, 1954, Chapter 20).
Herrnstein (1970) derived matching fromthe following equation for the dependenceof RI, the response rate on the ith schedule,on ri, the corresponding observed reinforce-ment rate:
kr1R 1= ri (2)
where r. and k are constants. At first sight, (2)is inconsistent with inelastic demand, for theequation implies that other things equal theresponse rate will be an increasing function ofreinforcement rate, and inelastic demand im-plies the opposite. However, it is apparentfrom Figure 5 that at least within the concVI VI conditions, both overall response rateand overall observed reinforcement rate wereroughly constant, and (2) was therefore satis-fied. It fails if the continuous reinforcementdata are taken into account, and it would failwith the fixed-ratio data cited above; but that
is part of a wider problem, for the difficultiesin applying matching principles to ratio sched-ules are well known (e.g., Herrnstein andLoveland, 1975).
EXPERIMENT IIExperiment I showed that matching, at least
in the generalized form of Baum (1974b), heldfor the aggregate behavior of a group of rats.But nothing was shown about the behavior ofindividuals. Aggregate matching might resultfrom similar behavior by all members of thegroup. On the other hand, there might beconsistent differences between individuals inundermatching or bias, the constants a and bof Equation 1. These could be associated witha more general dominance or territoriality re-spectively: a dominant rat might be expectedto overmatch, that is to monopolize the betterschedule in a situation where the group as awhole matched or undermatched (on the otherhand, access to a limited resource is not a goodmeasure of dominance in rats, according toSyme, Pollard, Syme, and Reid, 1974). Or itmight be that individuals matched to theirpersonal ratios of reinforcement rates, whichmight not be the same as either the pro-grammed ratios or the ratio obtained by thegroup as a whole.To investigate these possibilities, a similar
group of rats was used in the same apparatus,with two of the concurrent schedule conditionsfrom Experiment I. The rats were observed bytime-sampling for 90-min periods on selecteddays to see whether the aggregate results ofExperiment I were true of individuals. Therats were mildly deprived of food so as toconcentrate activity into the chosen observa-tion periods.
METHODSubjects
Five female hooded rats were selected atrandom from a new batch of ten, obtained asbefore.
ApparatusThe apparatus was the same as in Experi-
ment I, except that the floors of the chamberswere lined with paper towels and sawdust be-neath the grids. The towels, grids, and tubeswere changed, and the doors were washedabout every eight days (one level of the appa-
188

MATCHING IN GROUPS OF RATS
ratus was cleaned on each day, except that onthe seventh day the staircase was cleaned, andno cleaning was done on Sundays). A keyboardand indicator panel was used during observa-tion periods. It was connected to a digitalcomputer and controlled by a general experi-mental control system, ONLI,2 which also col-lected data.
PretrainingPretraining was carried out as in Experi-
ment I, but more quickly and systematically.Two food panels and two water panels were
introduced at once, but at first, water bottleswere fixed so that their spouts poked throughthe foodtrays of the dipper chambers (num-bers 3 and 5), with the foodtray flaps removed.Magazine and lever training for food rein-forcement were then carried out in the twofood chambers (numbers 2 and 6), making allchanges in contingencies simultaneously in thetwo chambers. The water bottles were thenremoved from Chambers 3 and 5, and dipperand lever training for water reinforcementcompleted, again making changes simulta-neously in the two chambers. In pretrainingand throughout the experiment, the presenceof a reinforcer in a foodtray was indicated bythe interchange of tray- and barlights, as in Ex-periment I.
ProcedureFour days after lever pressing had been
established in all four chambers, variable-interval (VI) 15-sec schedules were introducedin the food chambers. These schedules werein force for 23 days, and were followed byseven days with VI 45-sec in Chamber 6 andVI 15-sec in Chamber 2; 46 days with the re-
verse; and 29 days following a second reversal.The last three conditions were those of theexperiment proper. The decision to changeconditions was based on the observer's im-pression that group interaction had stabilized.
Except for the first five days with VI 15-secin both food compartments, the two foodchambers were darkened, their levers retractedand their schedules made inoperative, at 1000hr each day. The chambers returned to normalat some time between 1600 and 1715, and on
some days a 90-min observation then began.An hour before each observation session, the
2See Footnote 1.
rats were marked with distinct symbols onboth flanks. Observations were made by timeand activity sampling. A light flashed on theobserver's keyboard to indicate which of fivechambers (numbers 6 to 2 in that order) andwhich of three activities (present in chamber,contacting lever, eating/drinking) should beobserved. The observer responded by pressinga button for each rat that was engaged in thespecified activity in the specified chamber.Errors could be corrected until a "data com-plete" button was pressed, after which thenext activity or chamber was selected after ahalf-second pause. The whole cycle was re-peated every 30 sec; if a cycle took longer than30 sec to complete, the next one started assoon as the extended one finished, and the se-quence was brought back into phase as soonas possible. Note that the "contacting lever"observation was made visually; no attempt wasmade to gate it with the switch operated bythe lever.There were 5, 12, and 11 observation peri-
ods in the three conditions with unequal VIschedules. Some observation periods were alsoundertaken while the VI-15 sec schedules werein force in both chambers, but these were ex-ploratory: the conditions of observation werevaried to find the most suitable procedure.Data from these conditions are not reported.Their major result was that 90 min was a longenough session to observe each rat have a sub-stantial "meal" and then go to sleep.
In some observation sessions, a closed-circuittelevision camera, adapted for low illumina-tions, was used to assist the observer. The cam-era could be swung to as to show any chamberon a monitor placed where the observer couldwatch it and the apparatus together.
RESULTSTable 2 gives the aggregate'response and
reinforcement rates, both absolute and rela-tive, observed during the observation sessionsand during the periods that began with ob-servations sessions and ended with the with-drawal of the levers in the food chambers at1000 next day. Table 3 gives, for each rat, thenumber of occasions on which each type ofbehavior monitored (present in chamber, leverpressing, eating) was observed in Chambers 2and 6. All these data are reported as mediansover all the observation sessions in each sched-ule condition.
189

D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
Table 2Experiment II. Aggregate data from the observation sessions and the days that started withthem; all data are medians across all observation sessions in a schedule condition.
Schedule Condition
1 2 3
Chamber: 2 6 2 6 2 6VI (sec): 15 45 45 15 15 45
Observation sessionsLever-press rate (/sec) 0.129 0.074 0.089 0.239 0.226 0.122Reinforcement rate (/sec) 0.0298 0.0065 0.0111 0.0515 0.0443 0.0124Relative lever-press ratea 0.70 0.28 0.66Relative reinforcement rate' 0.82 0.19 0.78
Periods from the start of an observation session until1000 next day
Lever-press rate (/sec) 0.076 0.034 0.025 0.084 0.082 0.021Reinforcement rate (/sec) 0.0155 0.0030 0.0044 0.0200 0.0182 0.0035Relative lever-press rate 0.68 0.25 0.77Relative reinforcement ratea 0.82 0.18 0.85
"Chamber 2 rate/(Chamber 2 rate + Chamber 6 rate).
Figure 6 shows the relative lever contactscore in Chamber 2 for each rat (number oftimes the rat was observed in contact with thelever in Chamber 2/number of times it was
observed in contact with the lever in Cham-ber 2 or 6). Data are shown for all observationsessions with unequal VI schedules. The con-
cordance between sessions in ordering the ratsby relative lever-contact score was significant
Table 3
Experiment II. Median number of each type of obser-vation made of each rat in each schedule condition, outof a maximum of 180.
Schedule Condition
Rat Activity 1 2 3Chamber: 2 6 2 6 2 6
VI(sec): 15 45 45 15 15 45
1 Present 15 15 19 51 36 29Eating 13 15 3 12 11 3Lever-contact 13 15 4 12 12 11
2 Present 14 10 10 84 65 33Eating 15 10 1 24 17 3Lever-contact 15 10 2 29 21 5
3 Present 22 6 21 57 44 11Eating 21 5 2 16 10 1
Lever-contact 15 5 6 14 14 2
4 Present 25 1 32 45 28 40Eating 21 1 4 12 7 4Lever-contact 22 1 13 21 17 14
5 Present 21 7 20 50 48 9Eating 22 3 4 16 13 1Lever-contact 22 3 4 20 16 4
(1d
25
x1 5 1 1i 1
om mbnn1
Fig. 6. Experiment II. Proportion of all observationsof lever contact by each rat that were made in Cham-ber 2. Data are shown for each observation session inthe second and third schedule conditions of the experi-ment. Solid lines connect the medians for each session;the broken horizontal lines show relative programmedreinforcement rates in Chamber 2.
in the first two conditions but not the third(Kendall W = 0.52, p < 0.05; W = 0.28, p <0.01; W = 0.13, p > 0.05). Averaging over daysby summing ranks, the order of the rats byrelative lever-contact score in Chamber 2 was,from lowest to highest: Rat 1, 2, 3, 4, 5 inCondition 1; Rat 2, 5, 1, 3, 4 in Condition 2;Rat 1, 4, 5, 2, 3, in Condition 3. The concord-ance between these three orders is not signifi-cant, whether the second order is used directly(so as to test for consistent differences in rela-tive response rate in Chamber 2: W = 0.38,p = 0.41) or in reverse (so as to test for con-
sistent differences in relative response rate inthe preferable chamber: W = 0.38, p = 0.41,identity coincidental). Very similar results, butwith slightly more day-to-day variation, were
obtained for the present-in-chamber observa-tions.
9,4~ ratIo -- a~o :13Pv~~~~~.. lo <I I~~~~~~~*0 V 0D*0 a' V7 70Vwo' 'ooI 00
I 0~~~~v00s~~~~~~~~_~~~~~~~~~~~f.4
190

MATCHING IN GROUPS OF RATS
DISCUSSIONThe median relative lever-contact scores for
Chamber 2 are connected in Figure 6. Theydid not differ significantly from the pro-grammed relative reinforcement rates (alsoshown in the figure) in any condition (two-tailed signs tests over sessions, p = 1.0 in eachcondition). Thus, the matching law held be-tween visual observations of lever-pressing andprogrammed reinforcement rates; this resultagrees with that of Experiment I, and suggeststhat the observations of lever pressing werereasonably accurate. The aggregate relative re-sponse and reinforcement rates (Table 2) arealso consistent with Experiment I, and againsuggest matching to programmed relative rein-forcement rates, but undermatching to ob-served relative reinforcement rates.
Despite some significant differences betweenthe rats in relative lever-contact score, the con-cordance among the three sets of conditionsgave no indication that individuals differed indominance or had consistent chamber biasses.Some rats at some times were seen to defenda chamber, or a lever, but there was nothingin the observations or the quantitative datato suggest that this was territorial defense; atsome time or other, every rat was seen to dis-place every other from both chambers. Theonly obvious consistency in individual behav-ior concerned Rat 1. It undermatched consist-ently, and was also the smallest and lightestrat; it may have been displaced from the morefavorable chamber unusually often, or mayhave obtained an unusually low percentageof the reinforcements delivered when it wasthere. But even Rat 1 was often seen to dis-place other rats.The differences between rats within condi-
tions were therefore not attributable to any-thing that could be traced from condition tocondition. Might they have been caused bydifferences in the ratio of reinforcement ratesobtained by the rats under each condition?Because of competition, a single rat would notnecessarily obtain the reinforcement rate pro-grammed for a chamber even if it were con-tinuously present there. The "eating" obser-vation scores were used to estimate each rat'sobtained ratio of reinforcement rates in thetwo chambers. The lever-contact scores werenumerically so close to the eating scores thatan artifactual match was inevitable (see Ta-
ble 3); but the present-in-chamber scores werean order of magnitude larger, so they couldsensibly be plotted against the eating scores.Figure 7 shows, on log scales, the ratio ofpresent-in-chamber scores as a function of theratio of eating scores in the two food cham-bers. Each data point gives median data fromone rat in one schedule condition; no datafrom the first schedule condition are shown,because the criteria used for eating and levercontact were too liberal in that condition, sothat all three scores were virtually identical.Regression lines and correlation coefficientswere obtained for the other two schedule con-ditions separately. The correlation betweenlog present-in-chamber ratio and log eatingratio was significant in schedule Condition 2,but not in schedule Condition 3 (r = 0.89,p = 0.045, and r = 0.47, p = 0.47, respectively).
Thus, at least in schedule Condition 2, thelikelihood that a rat would be seen in onechamber rather than the other was signifi-cantly correlated with the likelihood that itwould be seen to eat in that chamber, takingthe correlation across the group of rats, all ofwhom were working at a constant programmedratio of reinforcement rates. But, as in Experi-
.0ES
U
C.w0.S
0
0YA3X+lnl.8
r1 .5 Iratio of eating observations 5 10
Fig. 7. Ratio of present-in-chamber observations ofeach rat (number of observations in Chamber 2/num-ber in Chamber 6) as a function of the correspondingratio of observations of eating, on logarithmic scales.Each data point gives the median for one rat over allthe observation sessions in one schedule condition; thepoints and the regression line in the lower-left quad-rant are from schedule Condition 3. The different sym-bols indicate different rats, using the same code as inFigure 6.
191

D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
ment I, it is unclear which is the independentvariable here. To be sure that the rats werereacting to differences in the contingenciesthey experienced, we need a measure thatcould not be determined by time allocation,and yet reflects the different rats' success inobtaining pellets from each chamber, giventhe competitive situation. The following mea-sure was devised. Each observation session wasdivided into 5-min blocks. Within each ofthese, each rat's competitive success in eachchamber was assessed as the proportion of ob-servations of eating in that chamber that wereattributed to that rat. This success proportionwas then averaged over all the 5-min blocksin the session, excluding blocks where the ratwas never observed in the chamber (the ex-clusion is important; without it, a relativecompetitive success score would reduce to asimple transform of the relative eating scoreused above). An adjusted reinforcement ratefor each rat in each chamber was then ob-tained by mutliplying the programmed rein-forcement rate in the chamber by the rat'smean competitive success there. Figure 8 showsthe ratio of present-in-chamber scores as afunction of the ratio of programmed reinforce-ment rates thus adjusted; we call the latter"socially programmed" reinforcement rates,because they are derived from the schedulearranged by the experimenter together withthe rats' social interactions. The details ofFigure 8 are the same as for Figure 7. Thecorrelation between log ratio of present-in-chamber scores and log ratio of socially pro-grammed reinforcement rates was not signifi-cant for either schedule condition (r = 0.65,p = 0.24 for schedule Condition 2; r = 0.62,p = 0.27 for schedule Condition 3). However,the regression lines (shown in Figure 8) werequite close to matching, and noticeably closerthan those shown in Figure 7. This differenceis consistent with the better match to pro-grammed than to observed reinforcement ra-tios observed for aggregate data in both Ex-periments I and II. The implication is that thesame processes operated between rats within acondition as operated for the aggregate be-tween conditions; but the conclusion from thisanalysis must be that there is no firm evidencethat the rats' individual experiences of thetwo chambers affected their tendencies to visitthem under constant programmed schedules.The converse process may have occurred.
O f
0Y=.91X+ln.73
, v
v
.1 .5 1 5 10socially programmed, reinforcement rate ratio
Fig. 8. Ratio of present-in,chamber observations ofeach rat (number of observations in Chamber 2/num-ber in Chamber 6) as a function of the correspondingratio of "socially programmed" reinforcement rates.The socially programmed reinforcement rates were de-rived from the programined reinforcement rates bydiscounting for each rat's competitive success in eachchamber. The details of the figure are as for Figure 7.
In summary, then, there is no conclusive evi-dence for individual differenecs due to terri-toriality or dominance between conditions, ordue to differences in competitive success withinconditions. In fact, if the lever-contact scoresfrom Table 3 are regarded as response ratescorresponding to relative programmed rein-forcement rates of 0.25 (Condition 2) and 0.75(Condition 3), the individual deviations frommatching are only twice as great as those ob-served in straightforward conc VI VI experi-ments with pigeons by Catania (1963, Figure 3,left column), using the same relative reinforce-ment rates.
GENERAL DISCUSSIONThe matching law, as generalized by Baum
(1974b), provided a useful summary of thebehavior of rats choosing between alternativesources of food reinforcement on variable-interval schedules. This was true both of theaggregate behavior of the group, and of thebehavior of individuals within the group. Be-havior differed only slightly (though signifi-cantly) from that predicted by the simplematching law of Herrnstein (1961, 1970).These results extend the generality of the
matching law, but they also have implicationsfor the origins of matching. Shimp (1966, 1969)
192

MA TCHING IN GROUPS OF RA TS
argued that matching occurs because animalsmake whatever response is instantaneouslymost likely to lead to reinforcement; Mackin-tosh (1974) appealed to Shimp's argument,and asserted that matching is an instance ofa general principle of reinforcement maximi-zation akin to the principle of consumer ratio-nality used in the microeconomic theory ofdemand (e.g., Lancaster, 1969). But it is not atall clear that matching could result from in-stantaneous maximizing in the present experi-ments. If a rat left one chamber to obtain apellet that had been set up in the other cham-ber, it took a considerable risk of failing toget that pellet (because another rat was con-currently occupying the other chamber) andlosing the next one in its original chamber(because another rat had come and taken it).It is more plausible to explain matching interms of the different response rates supportedby the different schedules used; in terms, thatis, of the psychology of learning and habit,rather than the psychology of rational de-cision.
In one respect, the rats did behave like con-sumers in the economy: they showed inelasticdemand for food, an essential commodity.Such behavior is conventionally deduced fromconsumer rationality, but it would be wrongto conclude that the rats were, therefore, partlyrational. Real consumers, as distinct from theideal consumers of economic theory, are at bestapproximately rational (Reynaud, 1954, p. 118ff.); in many respects, their behavior is a mat-ter of habit (Katona, 1953).
Since matching was found in a situation thatwas constructed to be a little like a real econ-omy, and since real economic behavior wassuccessfully mimicked in at least one respect,it seems worth asking whether any phenom-enon like matching can be found in the econ-omy. A possible analog is in Equation 3, whichgives a law formulated by Huff (1962) to pre-dict consumers' probabilities of visiting differ-ent shopping centers:
RJ/~RJ = S( /(TIX)) (3)
In (3), Rj is the number of visits to the jthshopping center, Sj is its size, and Tj is thetravel time to it. For two shopping centers ofthe same size, we have
RI/R LiT-] (4).
If we look on 1 / Tj as a reinforcement rate,(4) is a form of Equation 1, Baum's (1974b)generalization of the matching law; and of thematching law itself if X (a fitting constant)equals one. Huff found values of 2.6 to 3.7for X when the commodity in question wasclothing, and 2.1 to 3.3 when it was furniture.
For two shopping centers of different size,we have
RIRj _- Si/(TIX)Sj/(Tjl) (5).The size of a shopping center might be pro-portional to the quantity of goods soughtthere, and thus analogous to amount of re-ward. Equation 5 would then not be consist-ent with Baum's proposals for choice betweenschedules offering different amounts of reward,for according to Baum the entire quotient(Sj/ Tj) should be raised to the power X (or ain Baum's terminology). However, Todorov(1973) found that when the rate and theamount of reinforcement were both varied inconc VI VI schedules, pigeons' choices wereless affected by the relative amount than bythe relative rate of reinforcement. This resultis consistent with the amount being raised toa lower power than the rate in an extensionof Equation 1, and it thus has something atleast in common with Huff's formula.The empirical base of Huff's law is much
less secure than that of the matching law (forsome criticisms of it, and of related models, seeMason and Moore, 1970). The point of intro-ducing it here, and the point of the combinedmatching and "economic" analysis of the re-sults of Experiment I, is to suggest that con-sumer behavior, and economic behavior ingeneral, is a proper field for the applicationof operant psychology. Skinner (1953, p. 100ff.) compared different manners of payingwages with different schedules of reinforce-ment, but there has been little serious attemptto make an experimental analysis of economicphenomena. The most important developmenthas been the use of token economies (e.g.,Ayllon and Azrin, 1968), sometimes explicitlylinked to an economic analysis (Winkler,1971), and of multi-operant environments (e.g.,Findley, 1966). But much more could be done,along the lines of applying economic concepts
193

194 D. A. GRAFT, S. E. G. LEA, and T. L. WHITWORTH
to standard operant paradigms (as in Greenand Rachlin, 1975, or Kagel, Battalio, Rachlin,Green, Basmann, and Klemm, 1975), or show-ing that phenomena known in individuals alsoapply to groups (as in Grott and Neuringer,1974). The present experiments have shownthat both these last strategies can be pursuedsimultaneously.
REFERENCESAyllon, T. and Azrin, N. The token economy. New
York: Appleton-Century-Crofts, 1968.Baum, W. M. Choice in a continuous procedure. Psy-
chonomic Science, 1972, 28, 263-265.Baum, W. M. Choice in free-ranging wild pigeons.
Science, 1974, 185, 78-79. (a)Baum, W. M. On two types of deviation from the
matching law: bias and undermatching. Journal ofthe Experimental Analysis of Behavior, 1974, 22,231-242. (b)
Becker, G. S. A theory of the allocation of time. Eco-nomic Journal, 1965, 75, 493-517.
Catania, A. C. Concurrent performnances: reinforce-ment interaction and response independence. Jour-nal of the Experimental Analysis of Behavior, 1963,6, 253-263.
Collier, G., Hirsch, E., and Hamlin, P. E. The ecologi-cal determinants of reinforcement in the rat. Physi-ology and Behavior, 1972, 9, 705-716.
Findley, J. D. Programmed environments for the ex-perimental analysis of human behavior. In W. K.Honig (Ed.), Operant behavior. New York: Apple-ton-Century-Crofts, 1966. Pp. 827-848.
Green, L. and Rachlin, H. Economic and biologicalinfluences on the pigeon's key peck. Journal of theExperimental Analysis of Behavior, 1975, 23, 55-62.
Grott, R. and Neuringer, A. Group behavior of ratsunder schedules of reinforcement. Journal of theExperimental Analysis of Behavior, 1974, 22, 311-321.
Herrnstein, R. J. Relative and absolute strength ofresponse as a function of frequency of reinforce-ment. Journal of the Experimental Analysis of Be-havior, 1961, 4, 267-272.
Herrnstein, R. J. On the law of effect. Journal of theExperimental Analysis of Behavior, 1970, 13, 243-266.
Herrnstein, R. J. and Loveland, D. H. Maximizingand matching on concurrent ratio schedules. Jour-nal of the Experimental Analysis of Behavior, 1975,24, 107-116.
Hirsch, E. and Collier, G. The ecological determinantsof reinforcement in the guinea pig. Physiology andBehavior, 1974, 12, 239-249.
Hogan, J. A., Kleist, S., and Hutchings, C. S. L. Dis-play and food as reinforcers in the Siamese fightingfish (Betta splendens). Journal of Comparatitve andPhysiological Psychology, 1970, 70, 351-357.
Huff, D. L. A probabilistic analysis of consumer spa-tial behavior. In W. S. Decker (Ed.), Emerging con-cepts in marketing. Chicago: American MarketingAssociation, 1962. Pp. 443-461.
Kagel, J. H., Battalio, R. C., Rachlin, H., Green, L.,Basmann, R. L., and Klemm, W. R. Experimentalstudies of consumer demand behavior using labora-tory animals. Economic Inquiry, 1975, 13, 22-38.
Katona, G. Rational behavior and economic behavior.Psychological Review, 1953, 60, 307-318.
Kendall, M. G. and Stuart, A. The advanced theoryof statistics, Vol. 2. 2nd ed.; London: Griffin, 1967.
Lancaster, K. Introduction to modern microeconom-ics. Chicago: Rand-McNally, 1969.
Lobb, B. and Davison, M. C. Performance in concur-rent interval schedules: a systematic replication.Journal of the Experimental Analysis of Behavior,1975, 24, 191-197.
Logan, F. A. The free behavior situation. In D. Levine(Ed.), Nebraska Symposium on Motivation, Vol. 12.Lincoln: University of Nebraska Press, 1964. Pp. 99-131.
Mackintosh, N. J. The psychology of animal learning.London: Academic Press, 1974.
Mason, J. B. and Moore, C. T. An empirical reap-praisal of behavioristic assumptions in trading areastudies. Journal of Retailing, 1970, 46 (4), 31-37.
Mazur, J. E. The matching law and quantificationsrelated to Premack's principle. Journal of Experi-mental Psychology: Animal Behavior Processes, 1975,1, 374-386.
Morgan, M. J., Fitch, M. D., Holman, J. G., and Lea,S. E. G. Pigeons learn the concept of an 'A'. Per-ception, 1976, 5, 57-66.
Nevin, J. A. Rates and patterns of responding withconcurrent fixed-interval and variable-interval rein-forcement. Journal of the Experimental Analysis ofBehavior, 1971, 16, 241-247.
Reynaud, P. L. La psychologie dconomique. Paris:Riviere, 1954.
Shimp, C. P. Probabilistically reinforced choice be-havior in pigeons. Journal of the ExperimentalAnalysis of Behavior, 1966, 9, 443-455.
Shimp, C. P. Optimal behavior in free-operant experi-ments. Psychological Review, 1969, 76, 97-112.
Skinner, B. F. Science and human behavior. NewYork: Macmillan, 1953.
Stone, R. The measurement of consumers' expendi-ture and behaviour in the United Kingdom 1920-1938, Vol. 1. Cambridge: Cambridge UniversityPress, 1954.
Syme, G. J., Pollard, J. S., Syme, L. A., and Reid, R. M.An analysis of the limited access measure of socialdominance in rats. Animal Behaviour, 1974, 22,486-500.
Todorov, J. C. Interaction of frequency and magni-tude of reinforcement on concurrent performances.Journal of the Experimental Analysis of Behavior,1973, 19, 451-458.
Trevett, A. J., Davison, M. C., and Williams, R. J.Performance in concurrent interval schedules. Jour-nal of the Experimental Analysis of Behavior, 1972,17, 369-374.
Winkler, R. C. The relevance of economic theory andtechnology to token reinforcement systems. Behav-iour Research and Therapy, 1971, 9, 81-88.
Received 3 May 1976.(Final Acceptance 25 October 1976.)




























