Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, JUlY 1987, p. 1573-15790019-9567/87/071573-07$02.00/0Copyright C) 1987, American Society for Microbiology
Purification and Partial Characterization of the Capsular Polymer ofHaemophilus pleuropneumoniae Serotype 5
THOMAS J. INZANAtDepartment of Veterinary Microbiology-Pathology, Washington State University, Pullman, Washington 99164-7040
Received 29 December 1986/Accepted 25 March 1987
The capsular polymer (CP) of Haemophilus pkuropneumoniae serotype 5 was purified, and its chemicalcomposition was analyzed. Radioimmunoassay experiments showed that the maximum amount of CP could beobtained from broth cultures of bacteria in the late stationary phase, rather than from bacteria washed off agarplates. The CP was precipitated from culture supernatant with 5 mM hexadecyltrimethylammonium bromide(Cetavlon) and solubilized with 0.4 M NaCl. Ninety percent of the CP in the culture supernatant wasprecipitated with Cetavlon, although some material remained insoluble after NaCl extraction. The CP wasfurther purified by phenol extraction, ultracentrifugation, and Sepharose CL-4B gel filtration. The Kay of theCP from Sepharose CL-4B chromatography was 0.33. The CP preparation contained 85% hexosamine, 12%hexose, 3% phosphate, 0.17% protein, 0.20% nucleic acid, and 0.01% endotoxin. Thin-layer chromatography,an amino acid analyzer, and a glucose oxidase colorimetric kit were used to identify the sugar components ofthe hydrolyzed CP as glucosamine and glucose. Analysis of the native CP by 13C nuclear magnetic resonance
indicated that amino, N-acetyl, and carboxyl groups were present and that the CP was a disaccharide.
Haemophilus pleuropneumoniae is the etiologic agent ofswine pleuropneumonia, which is characterized by fibrinouspleuritis with hemorrhagic and necrotic lesions. The diseasemay range from acute to chronic and affects healthy,noncompromised pigs of all ages (32, 43). The role ofbacterial virulence factors in the pathogenesis of this diseasehas not been examined. A capsule, lipopolysaccharide(LPS), hemolysin, and a cytotoxin may all contribute topromoting the virulence of the bacteria or to the lesionscharacteristic of the disease (8, 14, 15, 22, 26).Ten serotypes of H. pleuropneumoniae have been pro-
posed (34), and a wide variety of tests have been developedfor serologic diagnosis and serotyping (18, 19, 32, 40, 43).Antiserum to whole cells contains antibodies that areserotype specific, species specific, and cross-reactive withother species or genera (43). Therefore, the use of wholecells or antiserum to whole cells may lead to erroneousdiagnostic results. The serotype-specific antigen has notbeen purified but has been proposed to be capsule, LPS, orboth because serotype-specific activity is greater with anti-gens from the aqueous phase after phenol extraction thanwith those from whole bacteria (18, 20, 40). Pleuropneu-monia vaccines consist of killed whole cell suspensions.These vaccines have limited success in preventing mortality,but do not prevent chronic disease and economic losses (25,33). Purified bacterial antigens that would be efficacious invaccines, however, have not been clearly identified.For some encapsulated bacteria the capsule is responsible
for serotype specificity and antiphagocytic activity, andantibodies to capsular antigens are protective and useful indiagnostic tests (10, 38, 39). Successful vaccines that consistof purified capsule have been developed for encapsulatedbacteria (38, 39). Pijoan (36) has proposed that the capsule ofH. pleuropneumoniae is the primary protective antigen.Studies supporting this concept have shown that onlybacteia from young, encapsulated cultures have any efficacy
t Present address: Department of Pathobiology, Virginia-Maryland Regional College of Veterinary Medicine, Virginia Poly-technic Institute and State University, Blacksburg, VA 24061.
in prevention of disease (33). To determine whether capsulefrom H. pleuropneumoniae is serotype specific and wouldprove useful in a vaccine, we purified a capsular polymer(CP) of H. pleuropneumoniae serotype 5, the most com-monly isolated serotype in the United States (37, 41). Anal-ysis indicated that the serotype 5 CP was a high-molecular-weight polysaccharide consisting of glucosamine, glucose,and phosphate residues that contained N-acetyl and carbox-yl groups. Another study confirmed that this CP was theserotype-specific antigen (21).(A preliminary report of this work was presented at the
87th Annual Meeting of the American Society for Microbi-ology, Atlanta, Ga., 1987.)
MATERIALS AND METHODSBacterial strains and growth conditions. H. pleuropneu-
moniae serotype 5 strains K17 and J45 were obtained fromBradly Fenwick (Department of Pathology, School of Vet-erinary Medicine, University of California, Davis). Serotype5 strain 178 was obtained from Martha Mulks (Department ofMicrobiology and Public Health, Michigan State University,East Lansing). The bacteria were stored at -70°C in skimmilk. For culture, a loopful of the frozen stock was streakedonto brain heart infusion agar containing 5 ,ug ofNAD per ml(BHI-N). Several mucoid, irridescent colonies were pickedand transferred to basal medium. For general purposes, thebasal medium used was BHI-N. For purification of CP, amodification of the buffered Casamino Acids (Difco Labora-tories, Detroit, Mich.)-yeast extract (CY) medium describedby Anderson and Smith (6) was used. The modificationsincluded filtration of the yeast extract through an ultrafiltra-tion membrane (PM10; Amicon Corp., Lexington, Mass.),absence of hemin in the medium, and addition of NAD at 5,ug/ml. Cultures in broth were shaken for maximum aerationat 37°C for 48 h for purification of CP, or for other purposes,to 109 CFU/ml (determined spectrophotometrically aftercorrelation of optical density with viable plate counts).
Purification of CP. The CP of H. pleuropneumoniaeserotype 5 was purified from the supernatant of CY mediumby a modification of the procedure described by Anderson
1573
Vol. 55, No. 7
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1574 INZANA
and Smith (6) for Haemophilus influenzae type b. All proce-dures were performed at 0 to 4°C unless described other-wise. Cultures were centrifuged at 10,000 x g for 10 min, andthe supernatant was recentrifuged. Polyanionic material wasprecipitated from the supernatant with hexadecyltrimeth-ylammonium bromide (Cetavlon; Eastman Kodak Co.,Rochester, N.Y.), and the precipitate was extracted withvarious concentrations of NaCl. The solubilized CP from thefinal NaCl extraction was precipitated with 2 volumes of95%ethanol, collected by centrifugation at 10,000 x g for 30 min,and solubilized in distilled water. The solubilized materialwas extracted with 45% phenol at room temperature for 30min followed by centrifugation at 10,000 x g for 15 min. Anequal volume of 90% phenol was added to the aqueousphase, and the extraction was repeated until the interfacewas clear (two to six times depending on the volume ofbacterial culture). The aqueous phase was dialyzed in sev-eral changes of distilled water until phenol could no longerbe detected in the dialysate. Sufficient NaCl was added tothe dialyzed crude CP to make the ionic strength 0.1 M,followed by precipitation of the CP with 2 volumes of 95%ethanol. The precipitate was suspended in distilled water andultracentrifuged at 100,000 x g for 4 h, and the supernatantwas lyophilized. The CP was suspended in a minimumvolume of 0.1 M NaCl and chromatographed on SepharoseCL-4B (420 by 30 mm; Pharmacia, Inc., Piscataway, N.J.)equilibrated in 0.1 M NaCl. Carbohydrate-positive fractionswith an A280 of less than 0.02 were pooled, dialyzed againstdistilled water, and lyophilized.
Radioimmunoassay. Radioimmunoassay for quantitationof CP was done with intrinsically labeled 3H-capsule from H.pleuropneumoniae serotype 5 by a modification of the dou-ble-label (16) procedure described by Anderson (4; T.Inzana, manuscript in preparation). Briefly, H. pleuropneu-moniae serotype 5 strain K17 was intrinsically labeled with[3H]glucose in CY medium, and the CP was purified. Anti-serum to serotype 5 bacteria was diluted in fetal bovineserum so that 50% binding of 2 nCi of 3H-capsule wasobtained. The supernatant of bacteria grown in CY mediumor that of bacteria washed off agar plates with CY mediumwas mixed one to one with fetal bovine serum and preincu-bated with diluted antiserum, followed by the addition of amixture containing 2 nCi of 3H-capsule and 3 nCi of 36CI. Theamount of CP in the culture supernatant was determinedfrom a standard curve, which was established with dilutionsof purified CP in place of culture supernatant.
Chemical analysis. Colorimetric assays were done for thedetermination of protein (28), total carbohydrate (12),hexosamine (44), hexose and heptose (49), phosphate (2),pentose (7), uronic acid (11), and sialic acid (3). Standardsused were bovine serum albumin, glucose, N-acetylglucos-amine, glucose and glucoheptose, KH2PO4, ribose, glucu-ronic acid, and N-acetylneuraminic acid, respectively, allfrom Sigma Chemical Co. (St. Louis, Mo.). The nucleic acidcontent was determined from Adams' nomograph (CaliforniaCorp. for Biomedical Research, Los Angeles) by A260 andA280. The LPS content was measured by the Limulusamoebocyte lysate assay (Associates of Cape Cod, WoodsHole, Mass.), with purified H. pleuropneumoniae serotype 5strain K17 LPS as the standard. The percentage of endotoxinirl the sample was estimated based on the highest concentra-tion of H. pleuropneumoniae LPS that did not clot theLimulus amoebocyte lysate (50 pg/ml).
For analysis by thin-layer chromatography, 5 mg of CPwas hydrolyzed in 1 ml of 4 N trifluoroacetic acid for 2 h at120°C (R. Rurangirwa, T. C. McGuire, and N. Magnuson,
Res. Vet. Sci., in press). Insoluble material was removed bycentrifugation at 8,000 x g for 10 min, and the supernatantwas diluted with 4 ml of distilled water and lyophilized.Resuspension of the hydrolyzed capsule in distilled waterand lyophilization were repeated twice, and the capsule wasfinally resuspended in 0.05 ml of distilled water. A 5-,ulsample of the hydrolysate was chromatographed on cellulosesheets with pyridine-ethyl acetate-acetic acid-water (5:5:1:3,vol/vol) as the solvent (48). Neutral sugars were identifiedwith silver (45), and amino sugars were identified withninhydrin (35). The glucose content of the hydrolysate wasdetermined colorimetrically with a glucose oxidase kit ac-cording to the instructions of the manufacturer (Sigma).Amino sugars were also identified with an autoanalyzer(121-MB; Beckman Instruments, Inc., San Jose, Calif.) afterhydrolysis of the capsule in 4 N HCI for 10 h at 100°C.Hydrochloric acid was removed by repeated rotary evapo-ration, and the sample was suspended in sodium citratebuffer (pH 4.95).
Nuclear magnetic resonance (NMR) spectroscopy wasperformed on 50 mg of capsule in D20 with a NicoletMagnetics (Palo Alto, Calif.) NT200 WT instrument. Diox-ane was used as an internal standard and assigned a chemicalshift of 67.4 ppm. The 13C spectrum was run at 50.3 MHzwith broad-band proton decoupling.
Purification and analysis of LPS. LPS was extracted withhot 45% phenol and purified by ultracentrifugation (47).Analysis of LPS by sodium dodecyl sulfate-polyacrylamidegel electrophoresis was performed as described by Laemmli(27). The stacking gel contained 4% acrylamide, and theseparating gel contained 15% acrylamide and 3 M urea.Samples were mixed one to one in solubilization buffer (46),electrophoresed at 25 mA per gel until the dye reached thebottom of the gel, and stained with ammoniacal silver asdescribed by Tsai and Frasch (46).
RESULTS
Cetavlon precipitation. Crude CP was initially precipitatedfrom culture supernatant with 10 mM Cetavlon and solubil-ized with 0.5 M NaCl. A variety of colorimetric assays weretested with the solubilized precipitate, but only assays forhexosamine, hexose, phosphate, and reducing carbohydratewere positive. The hexosamine assay was further used tomonitor for CP during additional purification procedures.Various concentrations of Cetavlon were incubated with100-ml samples of culture supernatant to determine theoptimum concentration of Cetavlon for maximum precipita-tion of CP. The precipitated CP from each preparation wassolubilized with 0.5 M NaCl and assayed for hexosamine(Table 1). A concentration of 5 mM Cetavlon precipitatedthe most hexosamine per milliliter, and this concentrationwas further evaluated to determine the percentage ofhexosamine remaining in the culture supernatant after pre-cipitation. The amount of hexosamine in sterile basal me-dium was determined and subtracted from the amount ofhexosamine in the same volume of culture supernatantbefore and after precipitation with 5 mM Cetavlon. Theprecipitate was solubilized with 0.5 M NaCl, equivalent tothe original volume of culture supernatant, and centrifugedat 10,000 x g to remove insoluble material, and the amountof hexosamine in the supernatant was determined. Thehexosamine content of the culture supernatant was 32.0 +0.3 ,ug/ml. The amount of hexosamine remaining in thesupernatant after Cetavlon precipitation was 3.3 ± 0.2,ug/ml. The content of hexosamine in the supernatant of the
INFECT. IMMUN.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
CAPSULAR POLYMER OF H. PLEUROPNEUMONIAE SEROTYPE 5 1575
TABLE 1. Hexosamine content of H. pleuropneumoniaeserotype 5 CP after precipitation by various concentrations
of Cetavlon0Final Cetavlon Hexosamineconcn (mM) content (,ug/ml)
1.0 852.5 1255.0 150
10.0 12925.0 11350.0 25
a H. pleuropneumoniae K17 was grown for 48 h in 1 liter of CY medium.The bacteria were removed by centrifugation, and a different concentration ofCetavlon was added to each 100-ml sample of culture supernatant. Eachprecipitate was extracted with 10 ml of 0.5 M NaCl, and the hexosaminecontent of the extract was determined.
Cetavlon precipitate after 0.5 M NaCl extraction was 16.9 +0.6 p.g/ml. These results indicated that of the original amountof hexosamine in culture supernatant (after subtraction ofhexosamine content from the sterile basal medium), 10.3%remained soluble after precipitation and 52.8% was recov-ered from the precipitate after extraction with 0.5 M NaCl.
Purification Methods. The effect of sequential 0 to 2 MNaCl extraction on solubilization of hexosamine, protein,and nucleic acid was examined (Table 2). Forty-two percentof recoverable hexosamine, as well as 60% protein and 54%nucleic acid, were solubilized by extraction with 0.1 MNaCl. Relatively minor amounts of hexosamine weresolubilized with increasing amounts of NaCl until the con-centration reached 0.4 M, at which point an additional 49%of the hexosamine (including the three previous extractions)was again solubilized. Extraction of the precipitate withconcentrations of NaCI greater than 0.4 M substantiallyincreased the amount of protein and nucleic acid contami-nation, but not the recovery of hexosamine.
After each of the following purification steps, 10% of the.naterial was dialyzed against distilled water, lyophilized,and suspended to 1 mglml in distilled water. When theCetavlon-precipitated supernatant was extracted with 0.4 MNaCl, 10% protein, 3.4% nucleic acid, and 50% endotoxinwere present (Table 3). After extensive phenol extraction,the protein content was substantially reduced to 1.3%,
TABLE 2. Effect of various concentrations of NaCl on extractionof hexosamine, protein, and nucleic acid from precipitates of
culture supernatant mixed with Cetavlona
NaCI concn in Hexosamine Protein Nucleic aciddistilled water content content content
(M) (,ug/ml) (,ug/ml) (,g/ml)
0.00 16 63 240.10 166 41 680.20 57 26 360.25 21 20 <10.30 7 <1 <10.40 131 9 <10.50 25 15 <11.0 8 <1 432.0 6 10 <1
a H. pleuropneumoniae was grown in 1 liter of CY medium, and the bacteriawere removed by centrifugation. Cetavlon (5 mM, final concentration) wasadded to the supernatant, and after 1 h at 4°C, the precipitate was collected bycentrifugation and extracted sequentially with 0 to 2 M NaCl. Hexosamineand protein contents were determined by colorimetric assays, and nucleic acidcontent was determined from a nomograph by A260 and A280.
TABLE 3. Content of protein, nucleic acid, and endotoxin in CPpreparations during various stages of purification
Protein Nucleic Endotoxin EndotoxinSamplea content acd content reciprocal
(%) content () endpoint titer
Supematant from 10.0 3.4 50 2 x 1070.4 M NaClextraction
Aqueous phase after 1.3 2.3 25 1 x 107phenol extraction
Supernatant after 0.7 1.8 0.5 2 x 105ultracentrifugation
Carbohydrate-positive 0.17 0.20 0.01 4 x 103fractions fromSepharose CL-4Ba After each procedure, 10% of the sample was removed, dialyzed against
distilled water, and lyophilized. The material was resuspended to 1 mg/ml indistilled water for assay.
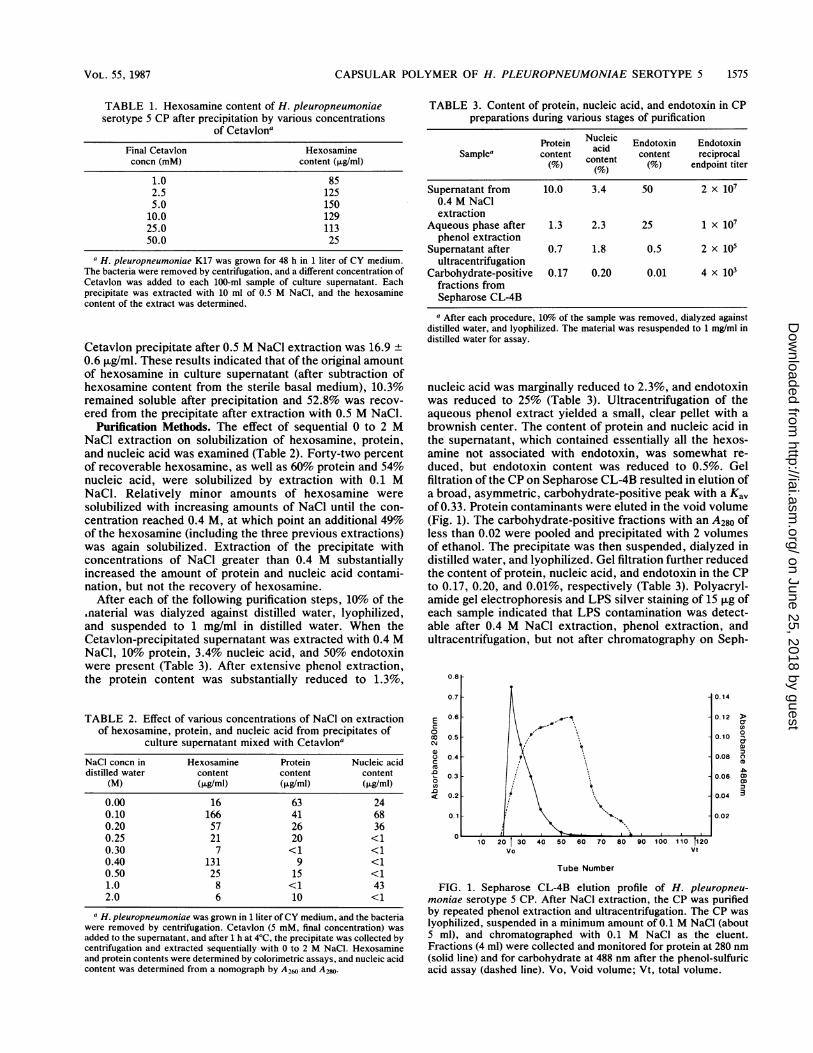
nucleic acid was marginally reduced to 2.3%, and endotoxinwas reduced to 25% (Table 3). Ultracentrifugation of theaqueous phenol extract yielded a small, clear pellet with abrownish center. The content of protein and nucleic acid inthe supernatant, which contained essentially all the hexos-amine not associated with endotoxin, was somewhat re-duced, but endotoxin content was reduced to 0.5%. Gelfiltration of the CP on Sepharose CL-4B resulted in elution ofa broad, asymmetric, carbohydrate-positive peak with a Kavof 0.33. Protein contaminants were eluted in the void volume(Fig. 1). The carbohydrate-positive fractions with an A280 ofless than 0.02 were pooled and precipitated with 2 volumesof ethanol. The precipitate was then suspended, dialyzed indistilled water, and lyophilized. Gel filtration further reducedthe content of protein, nucleic acid, and endotoxin in the CPto 0.17, 0.20, and 0.01%, respectively (Table 3). Polyacryl-amide gel electrophoresis and LPS silver staining of 15 jig ofeach sample indicated that LPS contamination was detect-able after 0.4 M NaCl extraction, phenol extraction, andultracentrifugation, but not after chromatography on Seph-
E0coc'Ja)uc)co
0on
0.14
0.12 >cr
0.10 0*
0.08 CC
(D0.06 0
a00.04 3
0.02
10 20 T 30 40 50 60 70 80 90 100 110 J120Vo Vt
Tube Number
FIG. 1. Sepharose CL-4B elution profile of H. pleuropneu-moniae serotype 5 CP. After NaCl extraction, the CP was purifiedby repeated phenol extraction and ultracentrifugation. The CP waslyophilized, suspended in a minimum amount of 0.1 M NaCl (about5 ml), and chromatographed with 0.1 M NaCl as the eluent.Fractions (4 ml) were collected and monitored for protein at 280 nm(solid line) and for carbohydrate at 488 nm after the phenol-sulfuricacid assay (dashed line). Vo, Void volume; Vt, total volume.
VOL. 55, 1987
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
1576 INZANA
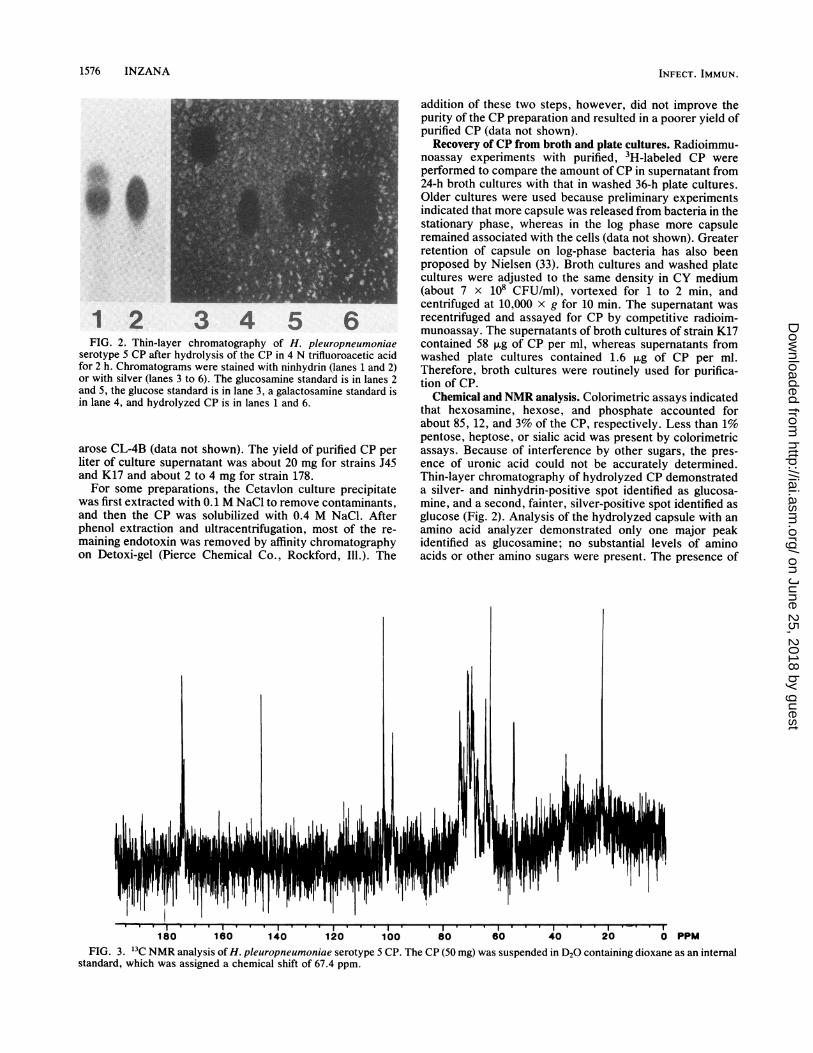
FIG. 2. Thin-layer chromatography of H. pleuropneumoniae
serotype 5 CP after hydrolysis of the CP in 4 N trifluoroacetic acid
for 2 h. Chromatograms were stained with ninhydrin (lanes 1 and 2)or with silver (lanes 3 to 6). The glucosamine standard is in lanes 2
and 5, the glucose standard is in lane 3, a galactosamine standard is
in lane 4, and hydrolyzed CP is in lanes 1 and 6.
arose CL-4B (data not shown). The yield of purified CP per
liter of culture supernatant was about 20 mg for strains J45
and K17 and about 2 to 4 mg for strain 178.
For some preparations, the Cetavlon culture precipitatewas first extracted with 0.1 M NaCl to remove contaminants,and then the CP was solubilized with 0.4 M NaCl. After
phenol extraction and ultracentrifugation, most of the re-
maining endotoxin was removed by affinity chromatographyon Detoxi-gel (Pierce Chemical Co., Rockford, Ill.). The
addition of these two steps, however, did not improve thepurity of the CP preparation and resulted in a poorer yield ofpurified CP (data not shown).
Recovery of CP from broth and plate cultures. Radioimmu-noassay experiments with purified, 3H-labeled CP wereperformed to compare the amount of CP in supernatant from24-h broth cultures with that in washed 36-h plate cultures.Older cultures were used because preliminary experimentsindicated that more capsule was released from bacteria in thestationary phase, whereas in the log phase more capsuleremained associated with the cells (data not shown). Greaterretention of capsule on log-phase bacteria has also beenproposed by Nielsen (33). Broth cultures and washed platecultures were adjusted to the same density in CY medium(about 7 x 108 CFU/ml), vortexed for 1 to 2 min, andcentrifuged at 10,000 x g for 10 min. The supernatant wasrecentrifuged and assayed for CP by competitive radioim-munoassay. The supernatants of broth cultures of strain K17contained 58 ,ug of CP per ml, whereas supernatants fromwashed plate cultures contained 1.6 pLg of CP per ml.Therefore, broth cultures were routinely used for purifica-tion of CP.Chemical and NMR analysis. Colorimetric assays indicated
that hexosamine, hexose, and phosphate accounted forabout 85, 12, and 3% of the CP, respectively. Less than 1%pentose, heptose, or sialic acid was present by colorimetricassays. Because of interference by other sugars, the pres-ence of uronic acid could not be accurately determined.Thin-layer chromatography of hydrolyzed CP demonstrateda silver- and ninhydrin-positive spot identified as glucosa-mine, and a second, fainter, silver-positive spot identified asglucose (Fig. 2). Analysis of the hydrolyzed capsule with anamino acid analyzer demonstrated only one major peakidentified as glucosamine; no substantial levels of aminoacids or other amino sugars were present. The presence of
'111II o r ' 11 -I 1 I I
. I . I I I I I I I I I I II.. I I .
180 160 140 120 100 80 60 40 20 0 PPMFIG. 3. 13C NMR analysis of H. pleuropneumoniae serotype 5 CP. The CP (50 mg) was suspended in D20 containing dioxane as an internal
standard, which was assigned a chemical shift of 67.4 ppm.
INFECT. IMMUN.
II1''
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CAPSULAR POLYMER OF H. PLEUROPNEUMONIAE SEROTYPE 5 1577
glucose in the hydrolyzed CP was confirmed with a colori-metric glucose oxidase kit, which was negative for glucosa-mine and galactose (data not shown).
'3C NMR of CP (Fig. 3) showed a signal at 54.86 ppm thatcorresponded to the C-2 atom of a hexosamine CH-NHresonance. The double-intensity peaks at 22.80 and 175.06ppm were identified as the -CH3 and C=O, respectively, ofN-acetyl groups, indicating that the glucosamine was N-acetylated; peaks corresponding to the -CH3 and C=O ofO-acetyl groups could not be identified. The signal at 173.98ppm was considered to be due to a carboxyl group. Thepeaks in the 50- to 80-ppm region were identified as the C-2to C-6 atoms of a dissaccharide. The peaks at 101.86 and98.54 ppm corresponded to the C-1 atoms of N-acetylglucos-amine and glucose, respectively. The difference in intensityof the peaks corresponding to the C-1 atoms indicated thatN-acetylglucosamine and glucose were not present in equi-molar ratios. The peak at about 146 ppm was not identified.
DISCUSSION
Previous reports of CP preparations from H. pleuropneu-moniae used harvested bacteria or aqueous extracts of cellswashed off agar plates (29-31). For some preparations, theaqueous extracts were further extracted with 45% phenol(18, 40; R. Nielsen, Ph.D. thesis, Royal Veterinary andAgricultural University, Copenhagen, Denmark, 1982) andultracentrifuged (14, 22). None of the antigen preparationsthat have previously been reported were confirmed to be apure capsular antigen by immunologic methods. In a sepa-rate study, the CP reported here was shown to be theserotype-specific antigen of H. pleuropneumoniae by indi-rect fluorescent-antibody, immunodiffusion, and enzyme-linked immunosorbent assays with whole cell serum andmonospecific serum to the CP (21).Anderson et al. (5) showed that the total amount of H.
influenzae type b capsule per unit volume was severalfoldgreater from broth cultures than from cells washed off plates.Furthermore, in stationary-phase broth cultures of H. influ-enzae most of the capsule was released from the cell.Radioimmunoassay experiments in our laboratory indicatedthat greater release of CP occurred during the stationaryphase and that there was about 50 times more CP in thesupernatant of broth cutures than in an equal volume ofsupernatant from washed plate cultures. This large differ-ence in CP content may have been due to enhanced sheddingor recovery of CP in broth culture. Therefore, to obtainmaximum yields of purified CP, the supernatant of station-ary-phase broth cultures was used rather than the superna-tant of cells washed off plates.The cationic detergent Cetavlon has been useful in the
purification of polyanionic capsular polysaccharides fromthe broth supernatants of H. influenzae type b (6), Neisseriameningitidis (17), Klebsiella species (9), and Escherichia coli(23). Many of the contaminants precipitated with capsule arein the culture medium. Therefore, the CY medium (supple-mented with NAD only) used for purification of capsule fromH. influenzae type b (6) was used because the medium iseasy to prepare, all the components are of low molecularweight, and the bacteria grow quickly with a short lag phase.The use of yeast extract dialysate or ultrafiltrate aids inreducing the amount of nucleic acid in the culture superna-tant (6). Maximum precipitation of anionic capsules withcationic detergents is concentration dependent and is basedon the overall charge and structure of the polyanion, as wellas the carbon chain length of the quaternary ammonium salt
(42). The maximum recovery of CP (determined as hexos-amine) with minimum protein and nucleic acid contamina-tion occurred in the presence of 5 mM Cetavlon, whichprecipitated 90% of the serotype 5 CP. Hexosamine-containing material that was not extracted with NaCl prob-ably remained in the precipitate as part of unsolubilized CPand LPS.Anderson and Smith (6) used progressively higher NaCl
concentrations to remove protein and nucleic acid contami-nants from the precipitated capsule of H. influenzae type band to solubilize the capsule. This procedure was alsosuccessful with the CP preparation of H. pleuropneumoniaeserotype 5, but a substantial amount of CP was lost byextraction with 0.1 M NaCl. Therefore, only 0.4 M NaCl wassubsequently used for solubiization of CP.
Serotype-specific antigens of H. pleuropneumoniae havebeen reported to follow the aqueous phase after hot aqueousphenol extraction (18, 40). Therefore, for some serologicassays, investigators have used cultures that have beenphenol extracted once (14, 19, 22, 40). For this study, the CPwas extracted with aqueous phenol at room temperature tominimize the amount of LPS in the aqueous phase (6).Although extraction with hot phenol may be more efficientthan that with cold phenol for removal of protein (47),repeated cold phenol extractions do remove most of theprotein from CP preparations (6). To reduce the proteincontent of the CP to minimum levels in this study, the CPrequired two to six phenol extractions (for 1 to 8 liters ofculture, respectively). My results indicated, however, thatphenol-extracted preparations still contained some proteinand as much as 25% endotoxin. In some cases, ultracentri-fugation has been used to remove LPS (14, 22). Results fromthis study and those by Fenwick et al. (14), using theLimulus amoebocyte lysate assay and sodium dodecyl sul-fate-polyacrylamide gel electrophoresis, showed that sub-stantial amounts of endotoxin remained in ultracentrifugedphenol extracts. After chromatography on SepharoseCL-4B, however, only 0.01% endotoxin was present in theCP preparation. Protein and nucleic acid contamination wasalso further reduced.Altman et al. (1) and Egan et al. (13) have successfully
used 13C NMR to obtain structural information on thecapsular polysaccharides of H. pleuropneumoniae serotype1 and H. influenzae type c, respectively. Of interest is thatthe composition of the CPs from H. influenzae type c and H.pleuropneumoniae serotype 1 is identical but the linkagesare different (1, 13). Both CPs are composed of a disaccha-ride of 2-acetamido-2-deoxy-D-glucose and D-galactose inequimolar ratios and contain phosphate and O-acetyl groups.The serotype 5 CP was composed of N-acetylglucosamine,glucose, and phosphate, but N-acetylglucosamine and glu-cose did not appear to be in equimolar ratios. Hexosamineaccounted for the majority of the serotype 5 CP, andtherefore an assay for hexosamine was the most sensitiveand convenient method to monitor for CP. The phosphatecontent of H. pleuropneumoniae serotype 5 CP was three tofive times lower than the phosphate content of the CP ofserotypes 1 to 3 (unpublished data). This deficiency inphosphate could be explained if only the glucose was phos-phorylated. Unlike for the serotype 1 CP (1), O-acetylgroups could not be identified in the serotype 5 CP by '3CNMR. A signal at 173.98 ppm was consistent with acarboxylic acid group that would enhance the negativecharge of the capsule since phosphate, the common anionicmoiety in Haemophilus and Neisseria capsules (24), waspresent in relatively low concentrations. The large number
VOL. 55, 1987
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1578 INZANA
of peaks in the region of 50 to 80 ppm (the C-2 to C-6 atoms)and the peaks at 98.54 and 101.86 ppm (the C-1 atoms)confirmed that the CP was a disaccharide. Additional anal-ysis will be required to determine the complete structure ofthe serotype 5 CP. The purified serotype 5 CP described inthis report should prove useful for determining the role ofcapsule in the virulence of H. pleuropneumoniae andwhether antibody to capsule is protective.
ACKNOWLEDGMENTS
I thank Porter Anderson for helpful comments and for review ofthe manuscript and Ma Jianneng and Brian Iritani for technicalassistance. I also thank S. Gurusiddaiah for doing the amino sugaranalysis and Donald Appel for the '3C NMR analysis.
This work was supported by a grant from Praxis Biologics andfrom the Washington State University Grant-in-Aid program. Sup-port for use of the NMR facility was provided by funds from theBoeing Co., Seattle, Wash.
LITERATURE CITED
1. Altman, E., J.-R. Brisson, and M. B. Perry. 1986. Structuralstudies of the capsular polysaccharide from Haemophiluspleuropneumoniae serotype 1. Biochem. Cell Biol. 64:707-716.
2. Ames, B. N. 1966. Assay of inorganic phosphate, total phos-phate, and phosphatases. Methods Enzymol. 8:115-118.
3. Aminoff, D. 1961. Methods for the quantitative estimation ofN-acetylneuraminic acid and their application to hydrolysates ofsialomucoids. Biochem. J. 81:384-391.
4. Anderson, P. 1978. Intrinsic tritium labeling of the capsularpolysaccharide antigen of Haemophilus influenzae type b. J.Immunol. 120:866-870.
5. Anderson, P., J. Pitt, and D. H. Smith. 1976. Synthesis andrelease of polyribophosphate by Haemophilus influenzae type bin vitro. Infect. Immun. 13:581-589.
6. Anderson, P., and D. H. Smith. 1977. Isolation of the capsularpolysaccharide from culture supernatant of Haemophilus influ-enzae type b. Infect. Immun. 15:472-477.
7. Ashwell, G. 1957. Colorimetric analysis of sugars. MethodsEnzymol. 3:73-105.
8. Bendixen, P. H., P. E. Shewen, S. Rosendal, and B. W. Wilkie.1980. Toxicity of Haemophilus pleuropneumoniae for porcinelung macrophages, peripheral blood monocytes, and testicularcells. Infect. Immun. 33:673-676.
9. Cryz, S. J., Jr., E. Furer, and R. Germanier. 1985. Purificationand vaccine potential of Klebsiella capsular polysaccharides.Infect. Immun. 50:225-230.
10. Dirks-Go, S. I. S., and H. C. Zanen. 1978. Latex agglutination,counterimmunoelectrophoresis, and protein A coagglutinationin the diagnosis of bacterial meningitis. J. Clin. Pathol. 31:1167-1171.
11. Dische, Z. 1947. A new specific color reaction for hexuronicacids. J. Biol. Chem. 167:189-198.
12. Dubois, M., K. A. Hamilton, P. A. Rebers, and F. Smith. 1956.Colorimetric method for determination of sugars and relatedsubstances. Anal. Chem. 167:350-356.
13. Egan, W., F.-P. Tsui, P. A. Climenson, and R. Schneerson.Structural and immunological studies of the Haemophilus influ-enzae type c capsular polysaccharide. Carbohydr. Res. 80:305-316.
14. Fenwick, B. W., B. I. Osburn, and H. J. Olander. 1986. Isolationand biological characterization of two lipopolysaccharides and acapsular-enriched polysaccharide preparation from Haemoph-ilus pleuropneumoniae. Am. J. Vet. Res. 47:1433-1441.
15. Fenwick, B. W., B. I. Osburn, and H. J. Olander. 1986.Resistance of C3H/HeJ mice to the effects of Haemophiluspleuropneumoniae. Infect. Immun. 53:474-479.
16. Gotschlich, E. C. 1971. A simplification of the radioactiveantigen-binding test by a double label technique. J. Immuol.107:910-911.
17. Gotschlich, E. C., T. Y. Liu, and M. S. Artenstein. 1969.Preparation and immunochemical properties of the group A,group B, and group C meningococcal polysaccharides. J. Exp.Med. 129:1349-1365.
18. Gunnarsson, A. 1979. Serologic studies on porcine strains ofHaemophilus parahaemolyticus (pleuropneumoniae): extrac-tion of type-specific antigens. Am. J. Vet. Res. 40:469-472.
19. Gunnarsson, A. 1979. Evaluation of different antigens in thecomplement-fixation test for diagnosis of Haemophiluspleuropneumoniae (parahaemolyticus) infections in swine. Am.J. Vet. Res. 40:1564-1567.
20. Gunnarsson, A., B. Hurvell, and E. L. Biberstein. 1978.Serologic studies of porcine strains of Haemophilus para-haemolyticus (pleuropneumoniae): antigenic specificity and re-lationship between serotypes. Am. J. Vet. Res. 39:1286-1292.
21. Inzana, T. J., and B. Mathison. 1987. Serotype specificity andimmunogenicity of the capsular polymer of Haemophiluspleuropneumoniae serotype 5. Infect. Immun. 55:1580-1587.
22. Jensen, A. E., and T. A. Bertram. 1986. Morphological andbiochemical comparison of virulent and avirulent isolates ofHaemophilus pleuropneumoniae serotype 5. Infect. Immun. 51:419-424.
23. Kaiser, B. 1977. A simple method for typing of acidic polysac-charide K antigens of E. coli. FEMS Microbiol. Lett. 1:285-288.
24. Kenne, L., and B. Lindberg. 1983. Bacterial polysaccharides, p.287-363. In G. 0. Aspinall (ed.), The polysaccharides, vol. 2.Academic Press, Inc., New York.
25. Kume, K., T. Nakai, and S. Sawata. 1985. Efficacy ofHaemophilus pleuropneumoniae vaccine in pigs. Jpn. J. Vet.Sci. 47:201-206.
26. Kume, K., T. Nakai, and A. Sawata. 1986. Interaction betweenheat-stable hemolytic substance from Haemophilus pleuropneu-moniae and porcine pulmonary macrophages in vitro. Infect.Immun. 51:563-570.
27. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
28. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
29. Mittal, K. R., R. Higgins, and S. Lariviere. 1983. Determinationof antigenic specificity and relationship among Haemophiluspleuropneumoniae serotypes by an indirect hemagglutinationtest. J. Clin. Microbiol. 17:787-790.
30. Mitui, T., H. Onaga, Y. Nagasawa, Y. Nomura, and S. Kurama-su. 1981. Studies on Haemophilus infections in swine. I. Appli-cations of the latex agglutination test to the diagnosis ofHaemophilus pleuropneumoniae (H. parahaemolyticus) infec-tions. Vet. Microbiol. 6:339-349.
31. Nicolet, J. 1971. Sur l'hemophilose du porc. III. Diffdrenciationserologique de Haemophilus parahaemolyticus. Zentralbl.Bakteriol. Parasitenkd. Infektioskr. Hyg. Abt. 1 Orig. Reihe B216:487-495.
32. Nicolet, J. 1986. Haemophilus infections, p. 426-436. In A. D.Leman, B. Straw, R. D. Glock, W. L. Mengeling, R. H. C.Penny, and E. Scholl, (ed.), Diseases of swine, 6th ed. IowaState University Press, Ames.
33. Nielsen, R. 1976. Pleuropneumonia in swine caused by Haemo-philus parahaemolyticus. Studies on the protection obtained byvaccination. Nord. Vet. Med. 28:337-348.
34. Nielsen, R. 1985. Serological characterization of Haemophiluspleuropneumoniae (Actinobacillus pleuropneumoniae) strainsand proposal of a new serotype: serotype 10. Acta Vet. Scand.26:581-585.
35. Patton, A. R., and P. Chism. 1951. Quantitative paper chroma-tography of amino acids. Anal. Chem. 23:1683-1685.
36. Pijoan, C. 1982. Haemophilus pleuropneumoniae: pathogenesisand control. Annu. Proc. Am. Assoc. Swine Pract. 1982:95-101.
37. Rapp, V. J., R. F. Ross, and B. Z. Erickson. 1985. Serotyping ofHaemophilus pleuropneumoniae by rapid slide agglutinationand indirect fluorescent antibody tests in swine. Am. J. Vet.Res. 46:185-192.
INFECT. IMMUN.
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CAPSULAR POLYMER OF H. PLEUROPNEUMONIAE SEROTYPE 5 1579
38. Robbins, J. B. 1978. Vaccines for the prevention of encapsu-lated bacterial diseases: current status, problems and prospectsfor the future. Immunochemistry 15:839-854.
39. Robbins, J. B., R. Schneerson, W. B. Egan, W. Vann, and D. T.Liu. 1980. Virulence properties of bacterial polysaccharides-unanswered questions, p. 115-132. In H. Smith, J. J. Skehel,and M. J. Turner (ed.), The molecular basis of microbialpathogenicity. Verlag Chemie GmbH, Weinheim.
40. Rosendal, S., and D. Boyd. 1982. Haemophilus pleuropneu-moniae serotyping. J. Clin. Microbiol. 16:840-843.
41. Schultz, R. A., R. F. Ross, A. Gunharsson, and R. Nielsen. 1983.Serotyping of 50 different isolates of Haemophilus pleuropneu-moniae from swine pleuropneumonia in Iowa and surroundingstates. Vet. Med. Small Anim. Clin. 78:1451-1453.
42. Scott, J. E. 1960. Aliphatic ammonium salts in the assay ofacidic polysaccharides from tissues. Methods Biochem. Anal.8:145-197.
43. Sebunya, T. N. K., and J. R. Saunders. 1983. Haemophiluspleuropneumoniae infections in swine: a review. J. Am. Vet.
Med. Assoc. 182:1331-1337.44. Smith, R. L., and E. Gilkerson. 1979. Quantitation of glycos-
aminoglycan hexosamine using 3-methyl-2-benzothiazalonehydrazone hydrochloride. Anal. Biochem. 98:478-480.
45. Trevelyan, W. E.j D. P. Procter, and J. S. Harrison. 1950.Detection of sugars on paper chromatograms. Nature (London)166:444 445.
46. Tsai, C. M., and C. E. Frasch. 1982. A sensitive silver stain fordetecting lipopolysaccharides in polyacrylamide gels. Anal.Biochem. 119:115-119.
47. Westphal, O., and K. Jann. 1965. Bacterial lipopolysaccharides.Extraction with phenol-water and further applications of theprocedure. Methods Carbohydr. Chem. 5:83-87.
48. Wolfrom, M. L., D. L. Patin, and R. M. De Lederkremer. 1965.Thin-layer chromatography on microcrystalline cellulose. J.Chromatogr. 17:488-494.
49. Wright, B., and P. A. Rebers. 1972. Procedure for determiningheptose and hexose in lipopolysaccharides. Anal. Biochem.49:307-319.
VOL. 55, 1987
on June 25, 2018 by guesthttp://iai.asm
.org/D
ownloaded from














![Cultivation andPartial Characterization ofSpiroplasmas in ... · Spiroplasma citri and unidentified strains (corn stunt organism, 277F [tick isolate], powderpuff, BNR-1, honeybee,](https://img.pdfslide.us/doc/110x75/5e52f91966b58e76ac372278/cultivation-andpartial-characterization-ofspiroplasmas-in-spiroplasma-citri.jpg)














