Embed Size (px)
Citation preview

i
Post-partum Anoestrus in Bali Cattle under Low-Input Animal Production Systems in Eastern Indonesia
Mohamad Ilyas Mumu
S.Pt., M.Sc.Ag.
A thesis submitted for the degree of Doctor of Philosophy at
The University of Queensland in 2017
School of Agriculture and Food Sciences

ii
Abstract
Bali cows (Bos javanicus) are a common breed in Indonesia and are favoured by
smallholders for their size and ease of husbandry. However reproduction rates can be
low in some areas and poor nutritional and reproductive management including poor
mating management, low availability of genetically superior bulls, poor oestrous
detection, poor Artificial Insemination (AI) procedures and poor availability of
inseminators at the proper time are crucial issues which are likely to affect this. This
thesis examined factors affecting post-partum anoestrus (PPA) and inter-calving
interval (ICI) in Bali cows and heifers. This was examined by using large data sets to
identify the factors which affect pregnancy rates in villages, developing methods to
detect oestrus and pregnancy in villages, examining the effect of body condition score
(BCS) on hormonal patterns and by quantifying milk production in cows.
A large scale analysis of records from across the eastern islands of Indonesia
indicated that there was a median inter-calving interval of 380 days but that post-
partum anoestrus could not be determined precisely because of poor observations of
first return to oestrus and mating. These records used observational data over 2-4
years where cows under various village management procedures were observed for
weight, BCS and at least dates of calving from which various reproductive parameters
could be calculated. ICI proved to be most useful and accurate given the quality of
observed data. The range in ICI was very large and low annual calving % were
recorded. There was an association of high reproduction rate with body condition
score (BCS) at calving such that a BCS >3.5 (out of 5) was associated with inter-
calving intervals of approximately 380 days. The median ICI across all sites was 380
days but some sites recorded 75 percentile values in excess of 500 days.
In controlled pen experiments, faecal progestogens and vaginal electrical conductivity
were examined as techniques to detect oestrus and pregnancy in the field. Faecal
progestogens could be used successfully but there were problems with vaginal
electrical conductivity in routine application and this requires more work. Cycling
heifers were observed over complete cycles and plasma progesterone and faecal
progestogens examined every 2 days. The pattern of faecal progestogen

iii
concentration over an oestrous cycle mirrored that of plasma progesterone and hence
could be used to monitor the oestrous cycle and pregnancy but the sensitivity was low
and a number of samples would need to be taken to detect the cycling pattern and/or
pregnancy. Similarly cows were monitored over the oestrous cycle and vaginal
electrical conductivity measurements made but the association was not strong or
sensitive enough to be used routinely. Associations between plasma leptin or plasma
IGF-1 concentration and BCS were examined in another controlled pen experiment
and plasma IGF-1 concentration was increased in response to higher BCS but plasma
leptin concentration was not affected. Heifers were fed two diets to result in two groups
of moderate (<3.1 out of a scale of 5) or high (>4) BCS. Plasma IGF-1 reflected level
of nutrition (higher concentration in high BCS heifers) and the resulting BCS (high or
moderate) had an effect on cycling in cows and PPA whereby moderate BCS first calf
cows showed complete anoestrus up to 100 days post-calving.
In a village based experiment, milk production was measured for 12 weeks after
calving and found to be low (1.7kg/d) and not related to stage of lactation or first calf
cow or mature cow status. There was no evidence of lactation anoestrus for extended
periods in this experiment in all cows but some cows had not exhibited oestrus after
84 days.
Bali cows and heifers are inherently highly fertile and a simple management system
of a target BCS at calving (at least ≥3), access to a bull or AI from 40 days post-calving
and weaning of calves at 5-6 months of age (or earlier if BCS is declining fast due to
poor nutrition) will enable annual calving or weaning % of approximately 80-90% to be
achieved.

iv
Declaration by author
This thesis is composed of my original work, and contains no material previously
published or written by another person except where due reference has been made
in the text. I have clearly stated the contribution by others to jointly-authored works
that I have included in my thesis.
I have clearly stated the contribution of others to my thesis as a whole, including
statistical assistance, survey design, data analysis, significant technical procedures,
professional editorial advice, and any other original research work used or reported
in my thesis. The content of my thesis is the result of work I have carried out since
the commencement of my research higher degree candidature and does not include
a substantial part of work that has been submitted to qualify for the award of any
other degree or diploma in any university or other tertiary institution. I have clearly
stated which parts of my thesis, if any, have been submitted to qualify for another
award.
I acknowledge that an electronic copy of my thesis must be lodged with the
University Library and, subject to the policy and procedures of The University of
Queensland, the thesis be made available for research and study in accordance with
the Copyright Act 1968 unless a period of embargo has been approved by the Dean
of the Graduate School.
I acknowledge that copyright of all material contained in my thesis resides with the
copyright holder(s) of that material. Where appropriate I have obtained copyright
permission from the copyright holder to reproduce material in this thesis.

v
Publications during candidature
No publications included.
Publications included in this thesis
No publications included.
Contributions by others to the thesis
The author’s principal supervisor is Professor Dennis Poppi who contributed to the
design and discussion of the current study which formed part of an ACIAR project in
Lombok and Southeast Sulawesi.
Professor Michael McGowan initiated the use of the VEC (vaginal electrical
conductivity) probe and taught the author its use. Dr. Sophia Edward also trained the
author in the use of the probe.
Dr. Stephen Anderson provided supervision and technical expertise in hormonal
analysis particularly blood and faecal samples through RIA (radioimmuno assay),
ELISA (enzyme-linked immunosorbent assay) and EIA (enzyme Immunoassay).
Dr. Simon Quigley advised and contributed to data collection in the field.
Dr. Kieren McCosker provided statistical analysis in particular the descriptive
analysis of the longitudinal datasets of reproduction in villages of cows and heifers.
Statement of parts of the thesis submitted to qualify for the award of another
degree
None.

vi
Acknowledgements
Special and countless thanks are given by the author to my beloved wife, Aini
Febriana, SE, SKom, MEcSt, and son, Hadi Indraprasti, who became a teenager
during this study, for their countless love and valuable times accompanying the
author during this long and exhaustive PhD Journey in UQ Gatton and with my long
stay in Gatton town.
My mother, Hj. Hamida Edward Mumu and my brothers and sister, Emsalwati Mumu,
SE, Irfan Rivai Mumu, SH, Drs. H. Moh. Ridwan Mumu, MSi, Zamroni Mumu, SE for
their strong support and motivation. My father in law, Drs. H. Ahmad Hamid and my
mother in law, Hj. Siti Ginawati and all my wife’s family for also their strong support
and motivation during my exhaustive and very long PhD journey in UQG.
I am grateful to thank to my original sponsorship – DIKTI (Indonesian Higher
Education scholarship) for giving me a chance to pursue my study at The University
of Queensland.
The author would also like to express his thanks to ACIAR Project for supporting and
funding my experiments and to the Graduate School of The University of
Queensland, Professor Alastair McEwan (Dean, UQ Graduate School) and
Professor Neal Menzies (Head, UQ School of Agriculture & Food Sciences) for
supporting my complicated extension, resolving such related administrative matters,
as well as financial help and administrative support.
Special thanks to all of my supervisors, Professor Dennis Poppi for his support in
designing and funding my experimental research, Professor Michael McGowan, for
his excellent lessons in teaching me how to operate and use an ultrasound scanner
and VEC machine, Dr. Stephen Anderson for his excellent lab skills of steroid and
protein hormonal assays, RIA and ELISA, Dr. Simon Quigley for his excellent skill in
organising required tools for data collection and Dr Kieren McCosker for his
statistical support and analysis. Without the support and encouragement of all these
people I would not have been able to complete this PhD. Dr Sophia Edwards also

vii
taught me how to use the VEC machine so that I was competent in its use and I
appreciated her help.
The author would like also thank to his milestone seminar reviewers, Dr. John
Gaughan, Dr. Judy Cawdell-Smith, and Dr. Karen Harper. Thanks also to Dr. Rafat
Al-Jassim, Professor Wayne Bryden, Associate Professor Peter Murray, Dr. Mark
Hohenhaus and Dr. Doug George (Post Graduate Coordinator) for their friendly
relationship and warm regards during my study at The UQ.
The author also would like to express his thanks you to the School of Agriculture and
Food Sciences, School of Biomedical Science and School of Veterinary Science at
The University of Queensland for providing facilities.
Dr. Diane Mayberry was a great support in accessing literature and reports from
ACIAR projects that she led. Dr. Frances Cowley and Dr. Dianne Stephens provided
advice and editorial corrections into some of my thesis chapters.
Special thanks also to Peter Isherwood for his kindness and helpful manner,
especially in the lab and settling into Gatton town. Kerri Tyller provided basic lessons
in Endocrinology lab at UQ St Lucia.
This project interacted with a lot of different people in Indonesia some of whom
provided access to data and animals for use in this thesis. Large Ruminant Research
Consortium Head, Dr. Dahlanuddin and its staffs, Fachrul Irawan, S.Pt., MP,
Muhammad Supriyadi, S.Pt, MP, Baiq T. Yuliana, S.Pt., MP (Uthie) and all of the
members provided overall co-ordination within Indonesia. Dr. Dahlanuddin and Dean
of Animal Science Faculty of Mataram University for allowing me to access facilities
and the Teaching Farm staff of Mataram University: Sap, Jul, Edi and Yadi and the
Karang Kendal North Lombok research site staff, Sahrul Gunadi, S.Pt and
Kurniawan and especially Dr. Tanda Panjaitan at BPTP NTB West Nusa Tenggara
Indonesia. The author also wants to thank experimental farm staff at the Indonesian
Beef Cattle Research Station (IBCRS) Grati for their helpfulness and kindness to
provide facilities and support my data collection. The Head of IBCRS at Grati
Pasuruan East Java Indonesia, Dr. Dicky Pamungkas and all of his staff including

viii
Dicky Dikman and all staff and technicians for providing me access to do my VEC
data and blood collection and the related supporting data. Professor Takdir Saili,
Dean of Animal Science Faculty Haluoleo University Kendari Southeast Sulawesi
(Sultra) and his staffs for providing data from the Kendari - Lapangisi ACIAR project
as part of my longitudinal datasets.
I want to thank Professor Marsetyo Head of International Office (IO) Tadulako
University for his strong support and guidance. I also thank him for helping me with
administrative matters for my PhD scholarship. My colleagues: Professor Damry for
his kindness and technical help in analysing my data and Professor Rusdi both for
their motivation and strong support. Professor Muhammad Basir as Tadulako
University Rector provided strong support for my PhD for which I am very grateful
and appreciative.
This long PhD journey was also supported by the Indonesian community in Gatton
(Indoga) during my stay and I thank all the students and families for their social
interaction, friendship and support.
Lastly, the author wants to especially dedicate this thesis work to my father, Edward
Mumu, who passed away before I commenced my study and was not able to see my
PhD journey. This thesis work was also for my brothers, Dirwan E. Mumu, SH and
Moh. Imran Mumu who also passed away.

ix
Keywords
Keywords: post-partum anoestrus, inter-calving interval, oestrous detection,
reproductive performance, progesterone, leptin, IGF-1, Body Condition Score, milk
production, Vaginal Electrical Conductivity.
Australian and New Zealand Standard Research Classifications (ANZSRC)
ANZSRC code: 070206, Animal Reproduction, 60%
ANZSRC code: 070204, Animal Nutrition, 30%
ANZSRC code: 100199, Agricultural Biotechnology not elsewhere classified, 10%
Fields of Research (FoR) Classification
FoR code: 0702, Animal Production, 80%
FoR code: 0707, Veterinary Science, 20%

x
Table of Contents
Chapter 1 Introduction ............................................................................................... 1
1.1 Background ................................................................................................... 1
1.2 Research importance .................................................................................... 3
1.3 Research objectives ...................................................................................... 3
Chapter 2 Literature review ........................................................................................ 5
2.1 Beef farming in Indonesia ............................................................................. 5
2.1.1 Overview ............................................................................................. 5
2.1.2 Small holder beef farming ................................................................... 5
2.1.3 Reproductive performance issues in Eastern Islands Indonesia ......... 6
2.1.4 Bali cattle ............................................................................................ 8
2.2 Factors affecting reproductive performance of heifers and cows .................. 9
2.3 Reproduction in female cattle ...................................................................... 12
2.3.1 Oestrous cycles ................................................................................ 12
2.3.2 Follicle development ......................................................................... 12
2.3.3 Oestrus and Ovulation ...................................................................... 13
2.4 Factors affecting reproductive performance of heifers and cows ................ 13
2.4.1 Mating and after mating management............................................... 13
2.4.2 Age at puberty ................................................................................... 14
2.4.3 Calving intervals ................................................................................ 16
2.4.4 Post-partum anoestrus ...................................................................... 16
2.4.5 Oestrous detection ............................................................................ 19
2.4.6 Puberty .............................................................................................. 21
2.4.7 Heifer fertility ..................................................................................... 22

xi
2.4.8 Return to oestrus .............................................................................. 23
2.4.9 Anoestrus .......................................................................................... 24
2.5 Factors affecting outcome of AI and pregnancy rate ................................... 25
2.6 Nutritional effects on reproductive performance .......................................... 25
2.6.1 Leptin concentration .......................................................................... 25
2.6.2 IGF-1 concentration .......................................................................... 26
2.6.3 Hormonal concentration effects on reproductive functions................ 27
2.6.4 Effect of nutrition ............................................................................... 31
2.6.5 The role of kisspeptin in metabolic regulation of reproduction .......... 33
2.6.6 Body condition score ......................................................................... 34
2.7 Faecal progestogens ................................................................................... 35
2.8 Conclusions................................................................................................. 36
Chapter 3 The relationship between plasma progesterone and faecal
progestogens in cyclic Bali heifers and post-calving Bali cows......... 37
3.1 Background ................................................................................................. 37
3.2 Aim of experiment ....................................................................................... 38
3.3 Materials and methods ................................................................................ 38
3.3.1 Location ............................................................................................ 38
3.3.2 Animals and diets .............................................................................. 39
3.3.3 Sample collection and importation .................................................... 40
3.3.4 Laboratory analysis ........................................................................... 42
3.4 Results ........................................................................................................ 44
3.4.1 Experiment 3a Cyclic Animals ........................................................... 44
3.4.1.1 Plasma progesterone ..................................................................... 44
3.4.1.2 Oestrous behaviour ........................................................................ 46

xii
3.4.1.3 Faecal progestogens ...................................................................... 47
3.4.2 Experiment 3b Post-calving cows ..................................................... 52
3.4.2.1 Plasma Progesterone ..................................................................... 52
3.4.2.2 Faecal progestogens ...................................................................... 55
3.5 Discussion ................................................................................................... 59
3.5.1 Cyclic heifers ..................................................................................... 59
3.5.2 Post-calving cows ............................................................................. 61
3.6 Conclusion .................................................................................................. 62
Chapter 4 The effect of nutrition on plasma leptin and IGF-1 concentrations in Bali
first calf cows during lactation ............................................................... 63
4.1 Background ................................................................................................. 63
4.2 Aim of experiment ....................................................................................... 64
4.3 Materials and Methods ................................................................................ 64
4.3.1 Location ............................................................................................ 64
4.3.2 Animals, diets and measurements .................................................... 64
4.3.3 Laboratory analysis ........................................................................... 65
4.3.4 Statistical analysis ............................................................................. 66
4.4 Results ........................................................................................................ 66
4.5 Discussion ................................................................................................... 70
4.6 Conclusion .................................................................................................. 72
Chapter 5 Investigation of measurement of Vaginal Electrical Conductivity to
determine stage of the oestrous cycle in Bali, Ongole and Madura cattle.
.................................................................................................................. 73
5.1 Background ................................................................................................. 73
5.2 Materials and Methods ................................................................................ 74
5.2.1 Location and time of experiments ..................................................... 74

xiii
5.2.2 Animals and animal management ..................................................... 74
5.3 Results ........................................................................................................ 76
5.4 Discussion ................................................................................................... 83
5.5 Conclusion .................................................................................................. 84
Chapter 6 Milk production in Bali cows and heifers in a village system ............. 86
6.1 Background ................................................................................................. 86
6.2 Aim of experiment ....................................................................................... 87
6.3 Materials and methods ................................................................................ 87
6.4 Results ........................................................................................................ 88
6.5 Discussion ................................................................................................... 90
6.6 Conclusion .................................................................................................. 91
Chapter 7 Reproductive performance of heifers and cows within villages ......... 93
7.1 Background ................................................................................................. 93
7.2 Aim of experiment ....................................................................................... 94
7.3 Materials and methods ................................................................................ 94
7.4 Results ........................................................................................................ 97
7.5 Discussion ................................................................................................. 102
7.6 Conclusion ................................................................................................ 105
Chapter 8 General discussion ............................................................................... 107
8.1 General comments .................................................................................... 107
8.2 Reproductive parameters .......................................................................... 109
8.3 Methodology to study reproduction in the field .......................................... 111
8.4 Conclusions............................................................................................... 111
References ........................................................................................ 113

xiv
List of Figures Page
2.1 Patterns of growth and regression of individual follicles (solid lines)
during two complete bovine oestrous cycles with two follicular waves per cycle are shown in upper graphs. Lower graphs show plasma progesterone and LH concentrations.
11
2.2 The interacting effects of Bali heifer fertility 22 2.3 Reproductive cycle of beef cow 23 2.4 The interactions between plasma leptin, cortisol and insulin in
ruminants during underfeeding and re-feeding 29
3.1 Hormone concentration during the 21 day of bovine oestrous cycle 37 3.2 Oestrous behaviour with bull detection 40 3.3 Blood collection procedure 41 3.4 Faecal sample collection procedure 41 3.5 Individual cow progesterone profiles for animals with high BCS.
Arrows indicate days on which oestrous behaviours were noted. The dates of sampling are given in the Materials and methods section
45
3.6 Individual cow progesterone profiles for animals with moderate BCS. Arrows indicate days on which oestrous behaviours were noted. The dates of sampling are given in the Materials and methods section
46
3.7 Plasma progesterone concentrations in cows with high (n=4) and moderate (n=8) BCS aligned around the nadir in progesterone (designated day 0) determined from individual profiles
47
3.8 Plasma progesterone (red) and faecal progestogen (blue) concentrations from matched samples on individual heifers across a 30 day sampling period. The dates of sampling are given in the Materials and methods section.
49-51
3.9 Relationship between plasma progesterone and faecal progestogen concentrations in matched samples from all cyclic heifers (n=6 Mod-BCS and n=6 High-BCS, and total samples n=178). Data was analysed by Pearson correlation
52
3.10 Plasma progesterone concentration in post-calving animals with high BCS
53
3.11 Plasma progesterone concentration in post-calving animals with moderate BCS
54
3.12 Individual animal profiles for plasma progesterone (red) and faecal progestogen (blue) on matched samples taken during post-calving across a 124 day sampling period
56-58
3.13 Relationship between plasma progesterone and faecal progestogen concentrations in matched samples from post-calving cows (n=8 Mod-BCS and n=4 High-BCS, and total sample number n=105). Data was analysed by Pearson correlation
59
4.1 The plasma concentration of insulin-like growth factor-1 (IGF-1) of lactating Bali cows in moderate (Mod-BCS, n=6) or high (High-BCS, n=2) body condition score for 100 days after calving. Results are expressed as mean ± SEM. * P <0.05, ** P < 0.01, *** P < 0.001
67

xv
4.2 The plasma concentration of leptin of lactating Bali cows in moderate (Mod-BCS, n=8) or high (High-BCS, n=2) body condition score for 100 days after calving. Results are expressed as mean ± SEM. No significant difference between BCS groups was observed at any time point
68
4.3 The relationship between liveweight of lactating Bali cows and the plasma concentration of insulin-like growth factor-1 (IGF-1) (a.; n=6 Mod-BCS and n=2 High-BCS) and leptin (b.; n=8 Mod-BCS and n=3 High-BCS) of lactating Bali cows in moderate (Mod-BCS) or high (High-BCS) body condition score for 100 days after calving
69
5.1 Vaginal electrical conductivity (VEC) measurement procedure 76 5.2 Vaginal electrical conductivity (VEC) for all breeds vs plasma
progesterone concentration on Days 0-3 of the experiment 77
5.3 Vaginal electrical conductivity (VEC) for all breeds vs plasma progesterone concentration on Day 42 to 44 of the experiment
77
6.1 Weigh-suckle-weigh method at Karang Kendal North Lombok research site
88
6.2 Milk production profiles of Bali heifers at Karang Kendal North Lombok research site (Numbers refers to animal ID)
89
6.3 Milk production profiles of Bali cows at Karang Kendal North Lombok research site (Numbers refers to animal ID)
89
7.1 Inter-calving interval (ICI) in relation to Body Condition Score (BCS) at calving for all villages
98
7.2 Estimated percentage of cows pregnant within 100 days of calving in relation to Body Condition Score (BCS) at calving. Data from Central Lombok only
99
7.3 Percentage of total calves born by site within each bi-monthly period
100

xvi
List of Tables
Page
2.1 Effect of the number of oestrous cycles exhibited by dairy cows in the period before AI on the subsequent reproductive performance (Reproduced from Obese et al. (2011)
7
2.2 Reproductive parameters of beef suckler cows in the early post-partum period (from Crowe 2008a)
8
2.3 Summary data profiles on reproductive and productive performances of Bali cows in Nusa Tenggara Indonesia (1990-2012)
18
2.4 Relatively frequency of oestrous symptoms in dairy cows 20 2.5 Mounting activity is influenced by the number of cows in heat 20 2.6 Total number of oestrous activities recorded followed by the
subsequent behaviour 21
2.7 Effect of oestrous detection rate on pregnancy rate 25 4.1 Response equations describing relationships between cow
liveweight and the plasma concentration of IGF-1 and leptin 68
5.1 Vaginal electrical conductivity (VEC) measurement, plasma progesterone concentration and estimated cycling status/stage of the oestrous cycle for Ongole, Bali and Madura females at Day 0 - 3 and Day 42 - 44 of the study
79-81
5.2 Mean (± SD) VEC measurements and plasma progesterone (Prog.) concentrations (ng/mL) by estimated cycling status/stage of the oestrous cycle
82
6.1 Calving date and Post-Partum Anoestrus (PPA, days) of heifers and cows with Body Condition Score (BCS, 1-5 scale) and milk production (kg/d) at Karang Kendal village of North Lombok research site. BCS and milk production were the average over 12 weeks of measurement
90
7.1 The years of observation and the average time cows were monitored for each project
96
7.2 Reproductive parameters by site with Body Condition Score (BCS, scale 1-5) at calving and inter-calving interval (ICI, days). ICI was calculated as the time difference between sequential calving dates
97
7.3 Estimated percentage of cows pregnant within 100 days of calving within village sites
97

xvii
List of Abbreviations AA Anovulatory anoestrus
ABTS 2, 2'-Azino-Bis-3-Ethylbenzothiazoline-6-Sulfonic Acid
ACIAR Australian Centre for International Agricultural Research
AI Artificial insemination
AQIS Australian Quarantine Inspection Service
BCS Body condition score
BD Becton, Dickinson and Company
BPTP Balai Penelitian Teknologi Pertanian
°C degrees Celsius
C Central
CBW Calf birth weight
CIDR Controlled internal drug releases
CL Corpus luteum
CP Crude protein
CPM Count per minute
CR Conception rate
csv comma-separated values
CV Coefficient of variation
d day(s)
DGLVS Directorate General Livestock and Veterinary Services
DNA Deoxyribonucleic acid
DM Dry matter
DW Distilled water
E East
eg exempli gratia
etc et cetera
EIA Enzyme Immunoassay
ELISA Enzyme-linked immunosorbent assay
Fig. Figure
FSH Follicle stimulating hormone
g gram
GAMG Goat anti-mouse gamma globulin
GH Growth hormone
GLM General linear models
GnRH Gonadotropin releasing hormone
GoI Government of Indonesia
h hour(s)
H2O2 Hydrogen peroxide
HF Holstein-Friesian
HPLC High-performance liquid chromatography
HRP Horseradish peroxidase
IBCRS Indonesian Beef Cattle Research Station

xviii
ICI Inter-calving Interval
ID Identification
i.e. id est
IGF-1 Insulin-like growth factor-1
IFN Interferon
IO International Office
IPRD Intravaginal progesterone releasing device
IRMA Immunoradiometric Assay
kg kilogram
kGY KiloGray
km kilometre
L Litre
LH Luteinizing Hormone
LW Liveweight
M Missing data
MCF Malignant catarrhal fever
ME Metabolizable energy
min minutes
MJ Megajoules
mL Milliliters
Mod Moderate
µl Microliter
MP Milk production
MSD Mating starting date
N North
NA Not available
NC North Carolina
NDF Neutral detergent fibre
NEB Negative energy balance
NEFA Non-Esterified Fatty Acids
ng Nanogram
NM Natural mating
NSB Non-specific binding
NTB Nusa Tenggara Barat
NTT Nusa Tenggara Timur
ODB Oestradiol benzoate
OM Organic matter
OVNE Ovulation without overt oestrus
PBS Phosphate buffer saline
pers. comm. Personal communication
PG Prostaglandin
PGF2α Prostaglandin –F2α
PhD Philosophy Doctoral

xix
Poss. oestrus Possessive oestrus
PPA Post-partum Anoestrus
Prog. Progesterone
QCs Quality controls
RIA Radioimmunoassays
Root MSE Root-mean-square error
SAS Statistical analysis system
SEM Standard error mean
Sultra Sulawesi Tenggara
TCs Total counts
TN Tanpa nama
TSH Thyroid stimulating hormone
TX Texas
UQG University of Queensland in Gatton
USA United States of America
VEC Vaginal electrical conductivity
vs versus

xx
List of Definitions
Cow A female cattle after first mating, whether non-pregnant or
pregnant
Heifer A young cohort of female cattle up to the time of first
calving and after that the cohort is classed as first-lactation
cows.

1
Chapter 1 Introduction
1.1 Background
Poor nutritional and reproductive management as well as inefficiency in implementing
assisted reproductive technology of artificial insemination (AI) are crucial issues in
Indonesian beef cattle production. Reproductive issues include poor mating
management, low availability of genetically superior bulls, poor oestrous detection and
poor availability of inseminators at the proper time. These factors contribute to the
observed decrease in national meat production (4.37%) (DGLVS 2010). In order to
be self-sufficient, Indonesia needs to have a national herd of at least 20 million cattle
(Krisnamurthi 2011). With the estimated consumption of 2 kg per person per year of
beef, the country has a deficit of 5.2 million animals; the present population of beef
cattle being 14.8 million (DGLVS 2012).
Bali cattle are one of the most popular domesticated animals reared by smallholder
farmers especially in the eastern islands of Indonesia. The geographical distribution
is historically based on the establishment of breeds by Dutch colonisers. Bali cattle
are well adapted to these islands; they are small with high fertility and reproduction in
regions where nutrition is poor (McCool 1992). Calving rates range from 75-90%
(Devendra et al. 1973; Copland 1974; Everitt 1978; Kirby 1979; Wirdahayati &
Bamualim 1990) but more recent surveys indicate values much lower than this (Bakry
1994).
A major issue is the low liveweight of Bali cattle currently offered for sale which is much
lower than historical weights at slaughter (Winter 2011). Cash for education for
children or hospital access are the main reasons for farmers selling their cattle even
though they may be in poor condition (Teleni et al. 1993; Priyanti et al. 2013). Poor
body condition will also limit reproductive performance (Richards et al. 1986;
Laflamme & Connor 1992; DeRouen et al. 1994; Syahniar et al. 2012).
Other aspects that influence cow management in low-input systems are oestrous
detection and long weaning periods (Poppi 2009). Poor oestrous detection will
contribute to low reproductive performance, especially when AI is used. Together with
a possible extended post-partum anoestrous interval, this will contribute to low

2
reproductive performance. Therefore, by implementing good management of
reproduction, including correct oestrous detection and early weaning, reproductive
performance should improve to biologically and economically acceptable levels.
In assessing the causes of poor reproductive performance, the major factors are body
condition score of the cow at calving, and timing of first post-partum ovulation and
weaning. Traditionally in beef production systems, first calf heifers have a low
pregnancy rate because of low body condition score and cows have a long post-
partum anoestrus. Variation between cows can be high. Heifer pregnancy rate in
villages has not yet been assessed in Bali cattle. Similarly, the appearance of first
oestrus in cows after parturition appears variable, but this could be due to
inexperienced observers or a real post-partum anoestrous problem. The pattern of
plasma progesterone can be used to determine if heifers and cows are cycling, and
these changes have been described in Bali cattle (Isobe et al. 2005; Belli & Jelantik
2011; Astiti & Panjaitan 2013).
The major problems affecting the reproductive performance of Bali heifers and cows
include:
Poor oestrous detection
Poor AI management and/or access to bulls
Long lactation and late weaning leads to lactation anoestrus, although there
is some evidence that anoestrus is not an issue (Fordyce et al. 2003). The
issue of post-partum anoestrous interval (PPA) has been reduced by
improving nutrition of breeding females via supplements that increase
utilisation of available poor quality forages and provide additional nutrients
directly since nutrition was the primary limiting factor of calving to conception
intervals in this case (Fordyce et al. 2003). This was also supported by good
mating management which provided a mating pen or special area into which
oestrous females were introduced during the desired mating season to
maximise the chance of mating during the optimal period. (Fordyce et al.
2003).
In well managed, fertile herds it is expected that 95% of cows will wean a calf each
year. A period after calving where cows do not return to oestrus is known as post-

3
partum anoestrus (Montiel & Ahuja 2005). Post-partum anoestrus was identified as a
problem in the cattle industry 60 years ago (Short et al. 1990). In anoestrous cows,
growing follicles do not mature, preventing ovulation (Roche et al. 1992; Montiel &
Ahuja 2005). The normal period of anoestrus is about 60 days, but long periods of
anoestrus (>150 days) have been reported in Bos indicus breeds in the tropics (Montiel
& Ahuja 2005), and this is a main cause of infertility in the tropics. Nutritional status
measured by body condition score and suckling management are the two major
factors affecting the duration of post-partum anoestrus in cattle (Short & Adams 1988;
Randel 1990).
1.2 Research importance
Smallholder beef farming systems dominate in Indonesia and comprise more than 4
million households that constitute almost 70% of the national beef cattle population
(Boediyana 2007). A key to improving the beef industry in Indonesia is to develop
smallholder systems (Hadi et al. 2002). Being a smallholder farmer is a way of life
and of cultural importance for the farmer. This system confers benefits, such as
income generating activity, social status, and household security. In addition, cattle
commonly become a buffer or insurance for the farmers (Siegmund-Schultze et al.
2007; Stroebel et al. 2008; Le Thi Thanh et al. 2010). As previously mentioned, the
farmers can sell their cattle whenever they need cash for education or health.
Therefore, improving the performance and productivity of smallholder farmers will be
an important step to alleviate farmers’ household welfare, and will in turn also support
the national beef supply.
The outcome of this research is to design a system of reproductive management
suitable for Bali cattle in smallholder farmer systems in Eastern Islands of Indonesia.
1.3 Research objectives
To define the reproductive pattern of Bali genotype heifers and cows by
studying their plasma and faecal progesterone profiles.
To study the relationship of level of nutrition and BCS with the hormones leptin
and IGF-1.
To study the relationship between progesterone (and ovulation) and vaginal
electrical conductivity

4
To measure milk production in heifers and cows
To use large observational data sets on reproductive performance of cows and
heifers to identify factors which impact on reproductive performance such as
BCS at calving

5
Chapter 2 Literature review
2.1 Beef farming in Indonesia
2.1.1 Overview
Bali cattle are a popular cattle breed that play an important role in the livelihoods of
smallholder farmers in Indonesia, and they are the main breed type in Eastern
Indonesia. In 2011, the Bali cattle population reached 4.8 million animals (Kementan-
BPS 2011). The Government of Indonesia (GoI) is attempting to increase beef cattle
production each year. One program aims to increase the local population of Nusa
Tenggara Barat (NTB) to one million cattle. In order to achieve this goal, there must
be sufficient nutrition. With proper management, cow reproductive cycles and
parturition can be synchronised with the seasonal availability and quality of feed.
2.1.2 Small holder beef farming
The current beef cattle population of Indonesia is 14.8 million (DGLVS 2012).
Approximately 70% of this is household scale. Bali cattle, an indigenous breed,
comprise about 32% of the total national cattle population (Kementan-BPS 2011) and
are farmed in smallholder farming systems (Diwyanto 2003); they are considered to
be the most suitable indigenous cattle breed for a low-input system (Martojo 2012),
and the most suitable for smallholder farming systems especially in Eastern Islands of
Indonesia.
Bali cattle are typically managed in cut and carry systems (Talib et al. 2003). Weaning
typically occurs at about 12 months of age, and this prolonged lactation may result in
cows of low body condition with long inter-calving intervals (Bakry 1994). Weaning of
the calf, at about 5-6 months of age will reduce the nutrient demand on the cow,
allowing her to recover body condition for the subsequent lactation and should
facilitate a quick return to oestrus, thus reducing the inter-calving interval (Fordyce et
al. 1997; Burns et al. 2010).
Studies to improve beef farming in Indonesia have been conducted by several workers
(Hadi et al. 2002; Lisson et al. 2010; Lisson et al. 2011; Poppi et al. 2011). Hadi et al.
(2002) implemented input-output modelling and proposed several scenarios for the
developing beef industry; Lisson et al. (2010) & (2011) applied a participatory

6
approach to generate a model for developing smallholder Bali cattle farming in Eastern
Indonesia; and Poppi et al. (2011) focused on the implementation of a simple
integrated village management system with a technical extension package in nutrition
and reproduction to improve Bali cattle productivity in Eastern Indonesia. This latter
study combined early weaning, controlled natural mating, and improved nutrition to
achieve an optimal body condition score at calving and to increase weaning
percentages. By weaning calves at 5-6 months of age, 90% of cows and 60% of first
calf heifers were pregnant within 90 days of commencement of mating. This suggests
that post-partum anoestrus in Bali cattle is shorter than Bos indicus cows.
Limited studies to date suggest the 40 day first oestrous rate of Bali cattle is 15% (Lucy
et al. 2001a), and heifers have 20% higher conception rate at the third oestrus than
the first (Byerley et al. 1987). However, detailed studies of post-partum anoestrus
have not been reported with primiparous and multiparous Bali cattle.
2.1.3 Reproductive performance issues in Eastern Islands Indonesia
In the beef industry, one of the major sources of economic loss is reproductive failure.
The main goal of beef breeding programs (AI or natural mating) is to maximize the
number of pregnant females. Failure of an animal to become pregnant at one or more
matings will prolong the calving interval and lead to inefficiency in beef production.
Long calving intervals (> 19 months) and low calving rates (<60%) are the two main
reproductive problems identified in eastern Indonesian beef cattle, and these problems
are amplified by the low-input systems in which they are managed. Traditional beef
management systems are usually implemented by poorly skilled farmers, with limited
technology, limited genetic quality and resources, and reproduction and nutrient
management problems.
Since its development in the 1940s, AI has become a routinely used commercial
procedure especially for farmers in developed countries. One of the advantages of
using this technology is the ability to store and widely disseminate genetic material
from superior animals. AI is the method of choice for genetic improvement programs,
as the dissemination of good quality male genetics can occur more quickly compared
to natural mating. The success of AI in eastern Indonesia, however, is generally lower
than natural mating, although a thorough evaluation of AI program efficacy has not
7
been conducted (Diwyanto 2008). A key to the success of AI implementation is
through intensively reared cattle, which facilitates better oestrous detection and
properly timed AI.
Adequate knowledge of cattle reproduction is essential to good herd management.
Life-time reproduction in cattle is likely constrained by three main factors: 1) age at
puberty and 2) age at first calving and 3) post-partum conception intervals.
The effect of the number of oestrous cycles exhibited by dairy cows in the period
before AI on the subsequent reproductive performance has been studied by Obese et
al. (2011) as presented in Table 2.1.
Table 2.1. Effect of the number of oestrous cycles exhibited by dairy cows in the period before AI on the subsequent reproductive performance (Reproduced from Obese et al. (2011).
Reproductive performance
Oestrous cycle group (Mean ± SEM) ≥ 2 cycles
(n=23) 1 cycle (n=26)
OVNE2 (n=10)
AA3 (n=13)
Calving to first oestrus (days)
38.7c ± 2.0
53.8b ± 2.5
72.1a ± 3.3
75.8a ± 2.1
Calving to first ovulation (days)
34.1b ± 2.5
44.0b ± 2.7
41.5b ± 3.9
76.8a ± 2.0
Calving to MSD (days) 75.5a ± 1.2
73.2ab ± 1.1
67.7b ± 3.6
71.5ab ± 2.0
MSD to conception (days)
22.3 ± 5.2
25.1 ± 4.8 18.8 ± 5.3 26.3 ± 6.0
Calving to conception (days)
98.1 ± 5.3
97.1 ± 4.8 87.9 ± 6.8 97.8 ± 5.8
Conception rate (%) 56.5 53.8 40.0 46.2 Pregnancy at 3 weeks (%)
69.6 50.0 50.0 46.2
Pregnancy at 6 weeks (%)
73.9 76.9 80.0 84.6
Failure to conceive (%) 4.3 15.4 10.0 0.0
ab Means within the same row with different with different superscripts differ (P < 0.05). 1 Group F-test probability (3 df) oestrous cycle groups. 2 Ovulation without overt oestrus (OVNE). 3 Anovulatory anoestrus (AA). MSD = Mating starting date Source: Obese et al. (2011).
Fertility and post-partum infertility parameters of beef cows observed in the early post-
partum period are summarized in Table 2.2.

8
Table 2.2. Reproductive parameters of beef suckler cows in the early post-partum period (Reproduced from Crowe 2008a).
Reproductive parameters Beef cows
Emergence of the 1st follicle wave (days post-partum)
5–10
% Cows that ovulate the 1st dominant follicle 20–35 Post-partum interval to first oestrus (days) 30–130 Nature of 1st ovulation Silent % Short cycles after 1st ovulation >70 Regulation of LH pulse frequency Suckling, Maternal bond,
Declining energy balance, and BCS at calving
Source: Crowe (2008a).
2.1.4 Bali cattle
Bali cattle, which is derived from a major species of wild banteng (Bos banteng)
(Mohamad et al. 2012), is a breed of evolutionary importance (Purwantara et al. 2012),
and is one of the superior cattle breeds for fertility and conception rates. Mitochondrial
DNA sequencing has confirmed that Bali cattle have a different ancestor from that of
European and Zebu cattle, and this is supported by their dissimilar characteristics
(Kikkawa et al. 1995). Therefore, breeding programs for Bali cattle should not only aim
to produce commercial stock, but also contribute to genetic improvement (Talib et al.
2003).
This breed was first domesticated in Bali and has been maintained by Balinese
ancestors for centuries. Bali cattle are considered the pillar breed for smallholder
farmers in Indonesia (Purwantara et al. 2012), and one of the important beef cattle
breeds contributing to livestock industry development in Indonesia (Talib et al. 2003).
They are popular (Handiwirawan & Subandriyo 2004; Winaya et al. 2011) and raised
in almost all Indonesian provinces under smallholder rearing systems as they easily
adapt to the many variations of the tropical harsh environment (Talib 2002). Notably,
they are also raised in other tropical areas, such as Malaysia (Sukanata 2008), and
the Cobourg Peninsula, Northern Territory, Australia (Hodges 1997). This breed is the
most suitable for intensive village-based management, as its deer-like temperament
allows it to be used for plowing rice paddy fields (Mohamad et al. 2009). The four
major cattle resource areas in Indonesia include South Sulawesi, West and East Nusa
Tenggara (NTB and NTT), and Lampung provinces (Talib 2002; Martojo 2012). Due

9
to the incidence and susceptibility of Bali cattle to Jembrana and MCF (malignant
catarrhal fever), respectively, Bali cattle are unable to be raised together with sheep
in Bali and Java islands (Mohamad et al. 2009). The Indonesian Government has
placed a high priority and policy on developing the beef cattle population in the eastern
islands of Indonesia in order to achieve its beef self sufficiency program.
Bali cattle have several advantages to help ensure the success of this program, such
as (1) high fertility rate (83%) (McCool 1992; Bandini 1997; Talib 2002), (2) shorter
inter-calving interval than European breeds, (3) high carcass percentage (56% of
liveweight), and (4) easily adapted to new environments (Bandini 1997), although
many of these characteristics are not achieved at the village level. These cattle are
also efficient workers (Sukanata 2008), have an excellent ability to grow on low-quality
fodder (Sukanata 2008; Mohamad et al. 2009), and are responsive to improved
management (Bandini 1997).
2.2 Factors affecting reproductive performance of heifers and cows
The success of any breeding program is indicated by the herd fertility rate. Several
factors affecting herd fertility include cycling status, calf survival, body condition
(nutrition level), and disease (Perry et al. 2011). In ruminants, prolificacy, fertility and
fecundity vary by breed, season, age, nutritional status, health, breeding management
and farm supplies (Duricic et al. 2012).
Ovulation after calving is one of the normal cycles that supports animal reproductive
status. Ovarian follicular dynamics was defined by (Lucy et al. 1992) as “The process
of continual growth and regression of antral follicles that leads to the development of
the preovulatory follicle”. Normal follicular dynamics are indicated by the successful
development of the dominant follicle. Several follicles are selected for growth in
regular waves, and one large follicle is recruited to become the ovulatory follicle
(Fortune 1993). The dominant follicle that is recruited and selected during a follicular
wave becomes a large ovarian follicle (>10 mm) (Lucy et al. 1992). This follicle
ultimately matures, and with adequate pituitary gonadotropic stimulation, it progresses
to ovulation (Sirois & Fortune 1988).
The whole process of recruitment, selection, and dominance is associated with
hormonal cycling. This process is stimulated by increasing plasma FSH

10
concentrations (Walters & Schallenberger 1984). The failure of the emergence of a
dominant follicle is determined by the level of plasma FSH (Adams et al. 2008). The
development of the dominant follicles is followed by increased LH pulse frequency
(Fortune 1993). When LH pulse frequency is increased, the dominant follicles grow
larger and remain dominant for a longer interval (Adams et al. 2008). Cyclic patterns
of gonadotropic and steroid hormones in cattle are determined by the patterns of
follicular development (Fortune 1993). Bovine follicle growth occurs in a regular
pattern in which the three-wave pattern of follicular development is exhibited during
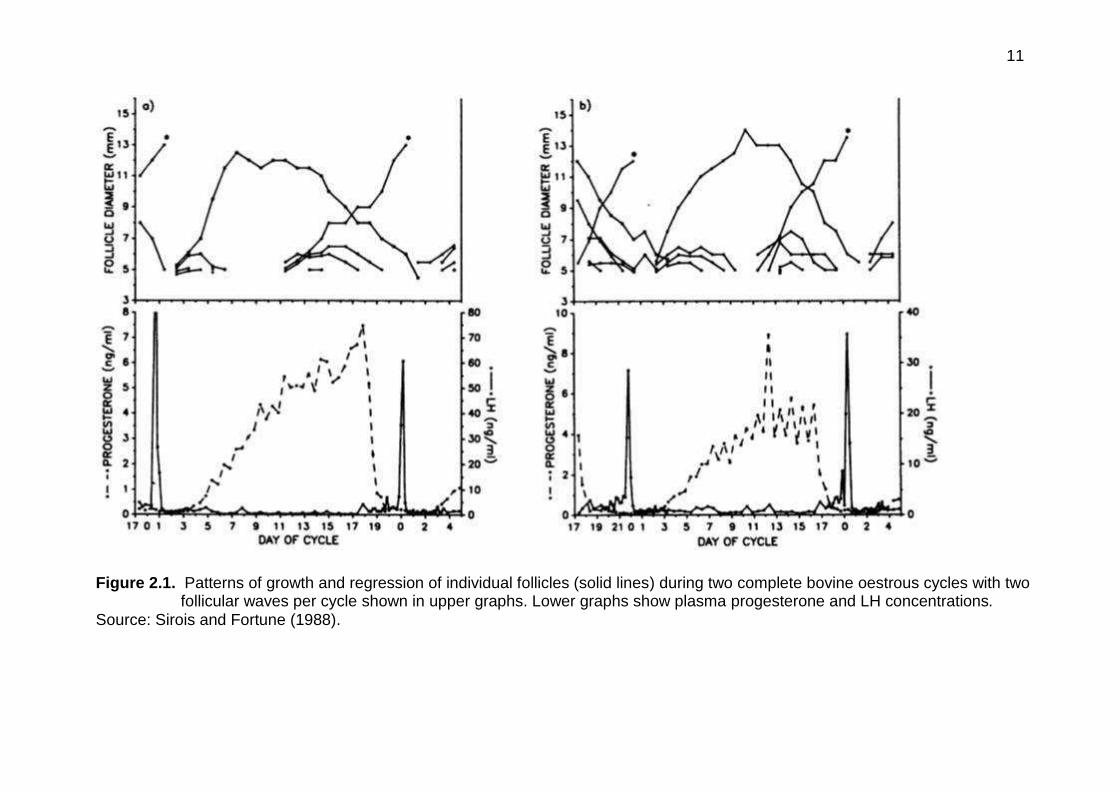
the oestrous cycle (Figure. 2.1).
Suckling is also a major factor involved in delaying post-partum return to oestrus
(Chenoweth 1994; Montiel & Ahuja 2005). Suckling depresses the development of
dominant ovarian follicles (Murphy et al. 1990; Stagg et al. 1998) due to reduced
hypothalamic release of GnRH and LH (Williams 1990) which is required for
development and final maturation of pre-ovulatory follicles (Montiel & Ahuja 2005).
This will lead to the occurrence of an anovulation and in turn extend post-partum return
to oestrus in suckled beef cows. Therefore, restricting calf suckling to once or twice
per day after the calf reaches a certain age is a beneficial approach to reduce the
length of return to oestrus, and increase pregnancy rates (Bastidas et al. 1984).
Reduced suckling is a simple, non-invasive and easily applied intervention to shorten
the interval for post-partum return to oestrus (Stagg et al. 1998).
11
Figure 2.1. Patterns of growth and regression of individual follicles (solid lines) during two complete bovine oestrous cycles with two follicular waves per cycle shown in upper graphs. Lower graphs show plasma progesterone and LH concentrations.
Source: Sirois and Fortune (1988).

12
2.3 Reproduction in female cattle
Reproduction is controlled by the endocrine system. The hypothalamus produces
gonadotrophin-releasing hormone (GnRH), which stimulates the anterior pituitary
gland hormones; Follicle-Stimulating Hormone (FSH) and Luteinizing Hormone (LH)
Luteinizing Hormone and Follicle Stimulating Hormone (FSH) are the two main
gonadotrophin hormones that regulate the function of the ovaries (Fortune 1994). This
includes the development of the follicle and oocyte, together with production of the
steroid hormones progesterone and oestradiol 17beta, and the protein hormone
inhibin. These ovarian hormones regulate reproduction locally in the ovary, but also
via feedback mainly negative, but also positive feedback to control GnRH and the
gonadotrophin hormones (Forde et al. 2011a). This complex hormonal system
controls the cyclical development of the ovarian follicles and corpora lutea (Stobart
1994).
2.3.1 Oestrous cycles
An oestrous cycle in cattle occurs over 18-24 days and consists of two phases, the
luteal and follicular phases (Forde et al. 2011b). The luteal phase (14-18 days) is
divided into two periods, metoestrus (formation of the corpus luteum period) and
dioestrus (the cyclic period when the corpus luteum is fully functional), and this is the
period following ovulation when the corpus luteum (CL) is formed. The follicular phase
(4-6 days) is also divided into two periods and designated as pro-oestrus (the
beginning period of corpus luteum regression) and oestrus (the period of time when
the female is receptive to the male and will stand for mating), and the follicular phase
is the period following the demise of the corpus luteum (luteolysis) until ovulation,
when final maturation and ovulation of the oocyte occurs into the oviduct allowing the
potential for fertilization (Crowe & Mullen 2013).
2.3.2 Follicle development
The whole process of follicle development, maturation, selection, and follicle
dominance is initially stimulated by an increase in FSH (Adams et al. 1992; Sunderland
et al. 1994; Stagg et al. 1998). FSH stimulates follicle development and in turn the
follicle produces more oestradiol. Oestradiol normally regulates LH secretion (Short et
al. 1979) through negative feedback by inhibiting the release of GnRH (Chandler 2001),

13
and inhibiting the LH pulse amplitude induced by GnRH (Goodman & Karsch 1980).
The pre-ovulatory LH surge is induced by oestradiol, once an adequate concentration
of oestradiol in blood is reached, in a positive feedback mechanism (Lyimo et al. 2000).
Also at the same time, the primary signal to the hypothalamus that induces oestrous
behavior is oestradiol. The duration of oestrus is controlled by increasing progesterone
concentrations early in the luteal phase (Vailes et al. 1992).
2.3.3 Oestrus and Ovulation
Ovulation in Bos indicus cattle occurs approximately 25 to 29 hours after the onset of
oestrus (Cavalieri et al. 1997; Pinheiro 1998). In general, Bos indicus breeds ovulate
sooner after the first sign of oestrus and display a shorter oestrus compared to Bos
taurus animals (Randel 1990; Chenoweth 1994). The duration of oestrus varies, from
7 hours for Brahman cows to about 14 hours for both Bos indicus and Bos taurus cows,
(Plasse et al. 1970; Orihuela 2000; Alves et al. 2009). It is also shorter in heifers than
in mature cows (De Silva et al. 1981; Mukasa-Mugerwa 1989). Other factors which
include nutrition, age, management and cow production status (Mukasa-Mugerwa
1989) will also impact the condition and reproductive status of animals.
Ovulation is a term describing the process whereby a mature follicle releases an
oocyte. The ovarian follicle develops and matures, under the influence of
gonadotropins from the anterior pituitary gland, together with steroid hormones and
follicle derived substances, culminating in the ovulation of the oocyte from the ovaries
(Stobart 1994).
The pre-ovulatory LH surge can be inhibited by progesterone via preventing positive
feedback by oestradiol (Scaramuzzi et al. 1971; Karsch et al. 1980). Progesterone
exerts negative feedback via the hypothalamus on GnRH and therefore reduces the
frequency of LH pulses (Chandler 2001). Other studies showed that the response to
positive feedback by the oestradiol is postponed by progesterone (Martin et al. 1983).
2.4 Factors affecting reproductive performance of heifers and cows
2.4.1 Mating and after mating management
The key factors in reproductive management of beef cattle include maximizing fertility
in females, provision of high-quality semen delivered at the appropriate time and place,

14
and good nutrition. Reproductive management also involves the provision of
replacement beef heifers, selected for early puberty and adequately grown to minimize
dystocia (Patterson et al. 1992).
Dystocia, or calving difficulty, results in direct and indirect losses. High birth weight is
the major cause of dystocia in cattle. This can be minimized by good heifer
management and selection of calving-ease sires.
Poppi et al. (2011) reported that earlier mating of Bali heifers resulted in a better
reproductive rate, shorter calving interval and higher growth rates of calves leading to
a better cash flow and market opportunities for households. In order to ensure that
heifers reach a pre-breeding target weight, management strategies of heifers or first
calf heifers should be designed to support the optimum age for such periods
(Patterson et al. 1992).
Although herd fertility can be influenced by bull fertility, little attention has been given
to the bull management in Indonesia, as in most cases farmers tend to focus more on
heifers and cows. Bull fertility is assessed by total progressive motile sperm
production, and this can be measured as it is highly correlated with the scrotal
circumference (Coulter et al. 1976; Bailey et al. 1996; Brito et al. 2002). Despite
genetic constitution of breed, climate, nutrition, age and disease all influencing
reproductive performance of bulls, bull management is primarily about the ratio of
males to females. The recommended ratio for natural mating of beef bulls to non-
synchronized beef cows ranges from 1:10 to 1:60 (Perry et al. 2011); the optimal ratio
was influenced by the bull (the age, experience, and semen quality) and the pasture
(the size and terrain). In many parts of Indonesia, however, pasture is not very
relevant as the cattle are all intensively housed (not free-grazed). The problems of
bull management are, therefore, more likely to be eliminated if care is given to the
design of the breeding program under housed conditions.
2.4.2 Age at puberty
Age at puberty is one of the important early-life reproduction traits that can determine
lifetime reproductive performance of an animal and genetic and environmental factors
are major influences affecting the attainment of puberty. Age at puberty differs among
breeds of cattle; Bos indicus cattle tend to reach puberty later than Bos taurus (16-40

15
and 12 months of age, respectively) (Plasse et al. 1965; Wiltbank et al. 1969; Tran et
al. 1988), and this reproductive trait has been shown to be highly heritable in Zebu
heifers (Nogueira 2004). A more recent report claimed that Bos indicus cattle are
unable to achieve puberty until they reach 60% of adult body weight
(Abeygunawardena & Dematawewa 2004). In order to increase lifetime reproductive
performance with no harmful effects on longevity or weaning weights, it is an
advantage if heifers calve at 2 years old compared to 3 years old (Tran et al. 1988).
Although age at puberty is an important characteristic of reproductive performance, it
can be difficult to determine based on behaviour associated with oestrus, and in
extensive management systems age at first calving serves as a useful proxy measure.
In Bos indicus heifers in Australia, puberty has been defined by plasma progesterone
levels reaching 1.0 ng/mL (Post & Reich 1980).
Bakry (1994) reported that male and female Bali cattle reach puberty between 12 and
24 months of age at weight 100–150 kg. In village-based Bali heifers with low growth,
first oestrus occurs at approximately 20 months of age (Poppi 2009). This study
suggested that weight was the main driver of first oestrus in heifers.
Werre (1980) measured the correlation between animal growth and age at puberty.
Earlier puberty in heifers was positively correlated with growth rate - faster growing
heifers presumably reaching a target weight for earlier initiation of puberty. Several
studies have shown that variations in feed intake affected the age at which heifers
reach puberty (Reid 1960; Bellows et al. 1965; Wiltbank et al. 1966; Wiltbank et al.
1969; Fajersson et al. 1991; Grings et al. 1999; Macdonald et al. 2005; Mohamed et
al. 2010; Randel & Welsh 2013). Poor nutrition significantly delays puberty in Bos
indicus breeds (Mancio et al. 1982; Oyedipe et al. 1982) and taurine cattle (Joubert
1954; Sorensen et al. 1959; Short & Bellows 1971). Body weight and growth, instead
of age, are the two major factors controlling the onset of puberty in these breed types
(Sorensen et al. 1959; Macdonald et al. 2005; Montiel & Ahuja 2005). Heifers growing
faster are likely to be younger and heavier at puberty (Arije & Wiltbank 1971; Werre
1980; Steffan et al. 1983; Martin et al. 1992b).

16
2.4.3 Calving intervals
Inter-calving interval is one of the main factors that affect the overall reproductive
efficiency of breeding herds. Bos indicus cattle tend to have a longer inter-calving
interval than Bos taurus cattle (Dobson & Kamonpatana 1986). The inter-calving
interval itself is influenced by many factors, principal among which are length of
gestation and the interval between parturition and subsequent conception (post-
partum anoestrus) (Montiel & Ahuja 2005). Late puberty in Bos indicus cattle is also
associated with a longer gestation length compared to Bos taurus (Paschal et al. 1991).
A long inter-calving interval caused by an extended post-partum anoestrus results in
low calf production in beef breeding herds, and is one of the major causes of economic
loss (Dziuk & Bellows 1983; Mwaanga & Janowski 2000; El-Wishy 2007). Inter-calving
interval is associated with fertility, and the fertility of heifers and cows will reflect their
reproductive efficiency. The ideal inter-calving interval for efficient breeding is 12
months: one calf every year. In Bali cattle, the inter-calving interval can be longer than
19 months (and up to 36 months) (Bamualim & Wirdahayati 2003).
Nutrition, body condition score (BCS), and genotype will also affect the post-partum
anoestrous interval. Adequate nutrition, particularly during the pre-partum period,
reduces the post-partum anoestrus and calving intervals (Montiel & Ahuja 2005).
Adequate BCS at calving and during the early post-partum period as well as suckling
twice a day will shorten the post-partum anoestrous interval (Montiel & Ahuja 2005).
Cyclic ovarian activity will re-establish earlier in beef cows when body condition at
calving is at a moderate or good level (Richards et al. 1986).
With better management and nutrition, it is possible to achieve the most efficient time
from calving to re-conception. The optimal time for cows to conceive after parturition
(post-partum conception) is one of the best criteria for measuring this reproductive
performance (Plasse et al. 1969) and percentage of cows pregnant within 100 days of
calving is a useful parameter for this.
2.4.4 Post-partum anoestrus
Post-partum anoestrus encompasses the interval between calving and first mating
after parturition. In Indonesia, 62% of reproductive disorders in heifers and cows are
attributed to post-partum anoestrus. Direct causes include nutrition and suckling

17
factors. Macronutrient deficiencies are usually reflected in poor BCS, but deficiencies
of essential minerals, such as zinc, magnesium and calcium, will also affect
anoestrous periods (Lall et al. 2000). Other influences include breed, age, pregnancy
rate, milk production, bull availability, late uterus involution, dystocia, and health status.
Cow management also affects measurement of anoestrus. Bos indicus cattle tend to
exhibit overt signs of oestrus mostly during the night time or early morning hours
(Pinheiro et al. 1998b), which are missed by most farmers (Mattoni et al. 1988b;
Pinheiro 1998). Regular observation may be insufficient to detect the signs of oestrus,
and better management strategies are needed. Advances in new techniques for
reducing the post-partum interval include non-hormonal remedies, such as physical
massage of the reproductive system (Mwaanga & Janowski 2000).
There are little published data on Bali cattle since most farmers keep poor livestock
records. This study will use data from other monitoring experiments to examine
reproduction in cows and heifers and in particular the influence of BCS. A summary of
the literature reporting key features of reproductive and productive traits of Bali cows
in Nusa Tenggara Indonesia is provided in Table 2.3.
18
Table 2.3. Summary data profiles on reproductive and productive performances of Bali cows in Nusa Tenggara Indonesia (1990-2012).
Key features of reproductive and productive traits of Bali cows in Nusa Tenggara Indonesia
Age at puberty (years)
Puberty weight
(kg)
Oestrous post-
partum (days)
Calving rate (%)
Calf mortality
(%)
Weaning weight (kg)
Birth weight
(kg)
Mature weight
(kg)
Concepti-on rate
(%)
First calving (years)
Inter-calving interval (months)
References
79 (Wirdahayati &
Bamualim 1990)
1.5 - 2 100 - 150
20 - 30 60 - 83 2.9 13 - 15 (McCool 1992)
1-2 100-150 62.8 (Bakry 1994)
2 – 2.5 192.6 – 179.8
51.7 – 66.6
15 - 48 79.2 – 83.9 11.9 – 12.7
221.5 – 241.9
3-3.4 15.4 - 16
(Entwistle et al. 2001)
60-90 80-90 15-18 (Talib 2002)
67.2 (Bamualim & Wirdahayati 2003)
2 182.6 ± 48.0
51.7 15 83.9 ± 25.9 12.7 ± 0.7
221-303 16 (Talib et al. 2003)
14.2 ± 2.4
11.9 ± 1.9
(Panjaitan et al. 2003)
93.80 18.60 (Chamdi 2004)
1.25 77.9 ± 7.53 15.12 ±
1.44 247 ±
39 1.5 (Sukanata 2008)
1.6 18-24 (Poppi 2009)
up to 30% (Poppi et al. 2011)
2 - 40 9 - 17 (Purwantara et al. 2012)

19
2.4.5 Oestrous detection
Oestrous-related behaviours in cows are affected by species, seasons, and stress. Galina
et al. (1982) reported a mounting behaviour of 1 and 2.8 per hour during oestrus for Brahman
and Charolais cows, respectively. Less frequent mounting is generally observed for Bos
indicus cows in the tropics. Plasse et al. (1970) and Llewelyn et al. (1987) reported that only
27% of mounting behaviour was observed in a herd of Bos indicus. In the tropics, an
extended period of high ambient temperatures is stressful to animals and leads to reduced
intensity of oestrous behaviours (Orihuela 2000; De Rensis & Scaramuzzi 2003). In the
temperate regions, however, the onset of oestrus seems to be better in summer than other
seasons. Zakari et al (1981) showed the oestrous behaviours of Zebu cows increased
during the hottest months while the oestrous manifestation was limited during winter (Alves
et al. 2009).
With regard to reproductive performance in general, the ovarian cycle sets reproductive
function (Berisha & Schams 2005). Detection of oestrus is important for a successful AI
program (Pinheiro 1998). The three conventional methods used to detect ovulation are daily
observation of sexual behavior, examination of vaginal temperature, and smear of cervical
mucus (Fallon & Crofts 1959; Kanitz & Becker 2005). More recent methods under
development include several related hormonal analyses.
Accurate oestrous detection, although time-consuming and sometimes difficult to do, is one
of the most important aspects of herd management (Anzar et al. 2003), and failure to detect
oestrus represents a serious management problem (Larson & Ball 1992),
The physiological expression of oestrus and the frequency of their occurrence in cows are
shown in Table 2.4.

20
Table 2.4. Relative frequency of oestrous symptoms in dairy cows.
Symptoms of oestrus Score
Mucous vaginal discharge 3 Cajoling 3 Restlessness 5 Sniffing the vagina of other cow 10 Chin resting 15 Mounted but not standing 10 Mounting (or attempt) other cows 35 Mounting headside of other cow 45 Standing heat 100
Source: Reproduced from VanEerdenburg et al. (1996).
One study demonstrated that mounting activity is influenced by the number of cows in heat
at the same time, as shown in Table 2.5, and can influence the rate of oestrous detection.
The number of mounts per cow increased to a maximum when three cows were in heat
simultaneously (Diskin & Sreenan 2000).
Table 2.5. Mounting activity is influenced by the number of cows in heat.
Number of cows in heat simultaneously
Average mounts per cow in heat
1 11.2 2 36.6 3 52.6
4 + 49.8
Source: Reproduced from Hurnik et al. (1975).
Mounting activity is considered the most reliable sign of oestrus (Orihuela 2000), but the
probability of directly observing this activity is fairly low (Cortes et al. 1999). However,
mounting activity is associated with a number of other behaviours, such as sniffing and
head-butting, which may alert the farmer to the presence of oestrus. Results of a study
investigating other behaviours associated with oestrus are shown in Table 2.6. Although the
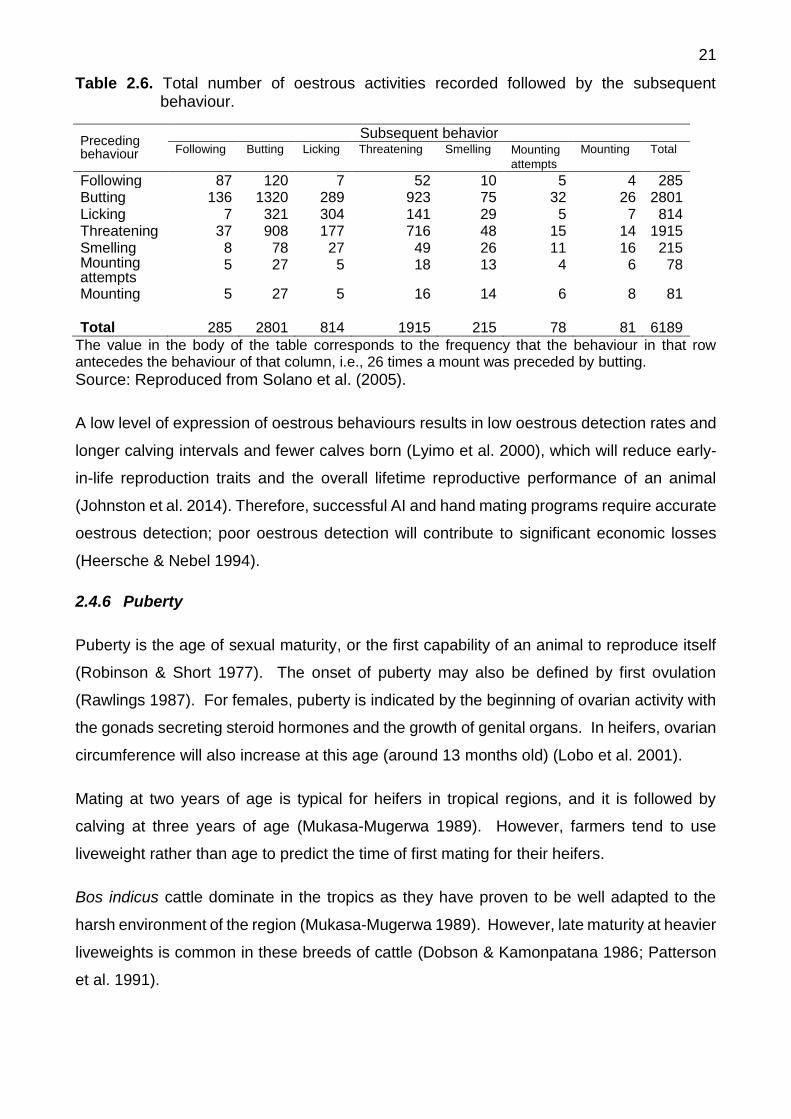
most frequent sign following mounting attempts or mounts was butting, these authors could
not establish a specific pattern of behaviours that defined oestrus (Solano et al. 2005).
21
Table 2.6. Total number of oestrous activities recorded followed by the subsequent behaviour.
Preceding behaviour
Subsequent behavior Following Butting Licking Threatening Smelling Mounting
attempts
Mounting Total
Following 87 120 7 52 10 5 4 285 Butting 136 1320 289 923 75 32 26 2801 Licking 7 321 304 141 29 5 7 814 Threatening 37 908 177 716 48 15 14 1915 Smelling 8 78 27 49 26 11 16 215 Mounting attempts
5 27 5 18 13 4 6 78
Mounting 5 27 5 16 14 6 8 81 Total 285 2801 814 1915 215 78 81 6189
The value in the body of the table corresponds to the frequency that the behaviour in that row antecedes the behaviour of that column, i.e., 26 times a mount was preceded by butting. Source: Reproduced from Solano et al. (2005).
A low level of expression of oestrous behaviours results in low oestrous detection rates and
longer calving intervals and fewer calves born (Lyimo et al. 2000), which will reduce early-
in-life reproduction traits and the overall lifetime reproductive performance of an animal
(Johnston et al. 2014). Therefore, successful AI and hand mating programs require accurate
oestrous detection; poor oestrous detection will contribute to significant economic losses
(Heersche & Nebel 1994).
2.4.6 Puberty
Puberty is the age of sexual maturity, or the first capability of an animal to reproduce itself
(Robinson & Short 1977). The onset of puberty may also be defined by first ovulation
(Rawlings 1987). For females, puberty is indicated by the beginning of ovarian activity with
the gonads secreting steroid hormones and the growth of genital organs. In heifers, ovarian
circumference will also increase at this age (around 13 months old) (Lobo et al. 2001).
Mating at two years of age is typical for heifers in tropical regions, and it is followed by
calving at three years of age (Mukasa-Mugerwa 1989). However, farmers tend to use
liveweight rather than age to predict the time of first mating for their heifers.
Bos indicus cattle dominate in the tropics as they have proven to be well adapted to the
harsh environment of the region (Mukasa-Mugerwa 1989). However, late maturity at heavier
liveweights is common in these breeds of cattle (Dobson & Kamonpatana 1986; Patterson
et al. 1991).
22
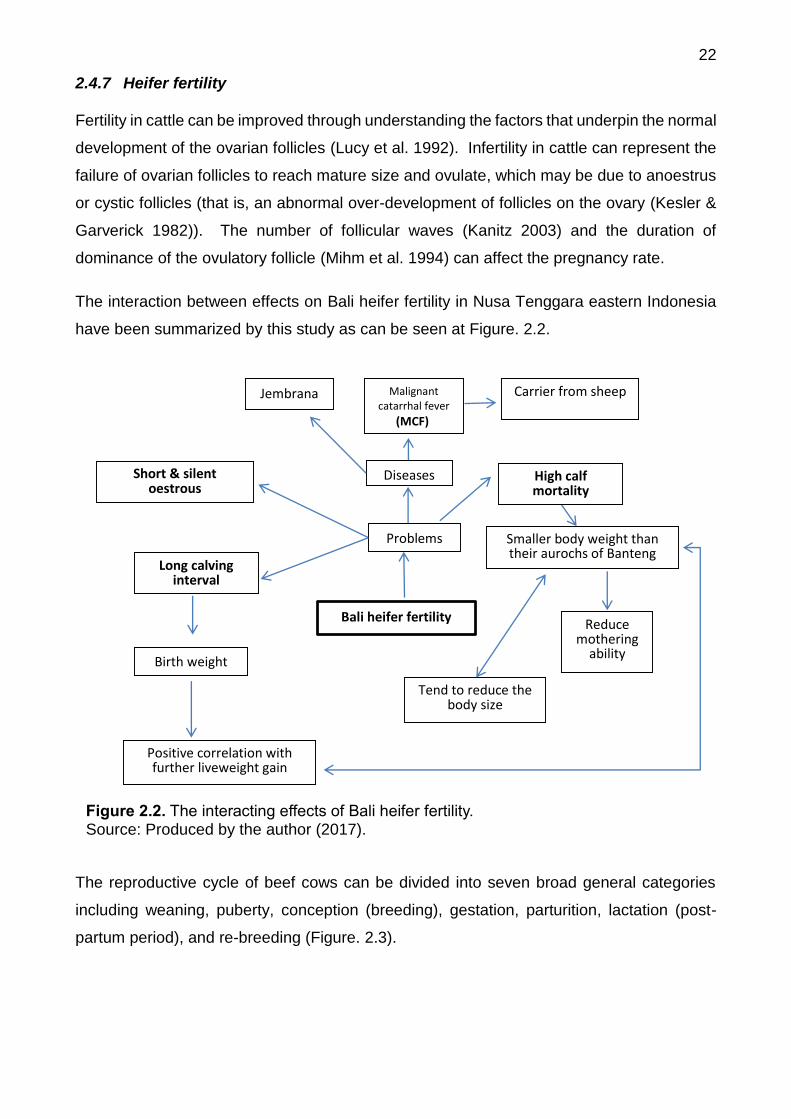
2.4.7 Heifer fertility
Fertility in cattle can be improved through understanding the factors that underpin the normal
development of the ovarian follicles (Lucy et al. 1992). Infertility in cattle can represent the
failure of ovarian follicles to reach mature size and ovulate, which may be due to anoestrus
or cystic follicles (that is, an abnormal over-development of follicles on the ovary (Kesler &
Garverick 1982)). The number of follicular waves (Kanitz 2003) and the duration of
dominance of the ovulatory follicle (Mihm et al. 1994) can affect the pregnancy rate.
The interaction between effects on Bali heifer fertility in Nusa Tenggara eastern Indonesia
have been summarized by this study as can be seen at Figure. 2.2.
The reproductive cycle of beef cows can be divided into seven broad general categories
including weaning, puberty, conception (breeding), gestation, parturition, lactation (post-
partum period), and re-breeding (Figure. 2.3).
Bali heifer fertility
Birth weight
Positive correlation with further liveweight gain
Tend to reduce the body size
Problems
Diseases High calf mortality
Long calving interval
Malignant catarrhal fever
(MCF)
Jembrana
Smaller body weight than their aurochs of Banteng
Short & silent oestrous
Carrier from sheep
Reduce mothering
ability
Figure 2.2. The interacting effects of Bali heifer fertility. Source: Produced by the author (2017).

23
Figure. 2.3. Reproductive cycle of a beef cow. Source: Reproduced from Patterson et al. (1992).
2.4.8 Return to oestrus
To achieve annual calving, cows need to become pregnant within 90 days after calving, as
mean gestation interval is 285 days (McDougall et al. 1998; Montiel & Ahuja 2005; Wright
et al. 2010). Thus initiation of first ovulation after parturition is important and this defines the
length of the post-partum anoestrus. In tropical climates, a long inter-calving period is
common (Ahuja & Montiel 2005).
As previously mentioned, a normal interval of return to oestrus for Bos indicus cows is
considered to be 18 to 24 days (Garcia et al. 1990). It usually takes longer for heifers after
their first calf (Mukasa-Mugerwa 1989); a minimum return to oestrous period is normally
reached on their third oestrus (Byerley et al. 1987).
Return to oestrus is physiologically influenced by hormonal mechanisms, nutrient intake and
energy reserves. Changes to hypothalamic, anterior pituitary and ovarian activity and
subsequent ovulation rate inhibit follicular development (Scaramuzzi et al. 2006). This is
closely related to nutrient requirements at certain periods. This is also associated with
energy availability in an animal. In most cases, nutrition has a direct effect on the animal’s
energy balance and the timing of the post-partum return to cyclic ovarian function (Canfield
& Butler 1991).
On a micro-nutrient scale, it is also known that nutritional deficiency of phosphorous can
cause oestrous suppression (Lopez et al. 2004). Phosphorus deficiency causes lowered

24
conception rates, irregular oestrus, anoestrus, reduced ovarian activity, increased incidence
of cystic follicles, and generally depressed fertility (Morrow 1980; Pugh et al. 1985).
However, Lopez et al. (2004) reported that excessive levels of dietary P (0.48%) above the
National Research Council (2001) of USA recommendation (0.38%) did not improve
duration or intensity of oestrus.
2.4.9 Anoestrus
Anoestrus is a period where an animal has an absence of ovarian cyclic activity and exhibits
no overt oestrous signs or sexual activity (Wright & Malmo 1992). During anoestrus, serum
progesterone concentrations are low (< 0.5 ng/mL) (Walker et al. 1984).
In cattle, this has long been considered as the most common single cause of infertility
(Lopez-Gatius et al. 2008). Anoestrus occurs because of the negative feedback mechanism
of oestradiol on LH secretion (Legan et al. 1977; Goodman et al. 1981; Karsch et al. 1993).
When a normal follicle fails to develop, oestradiol production is removed, which affects LH
secretion (Bartlewski et al. 1999).
Prolactin concentration can also contribute to the length of anoestrus. Increased prolactin
during lactation is known to inhibit GnRH secretion and consequently reduce oestrogen
secretion, resulting in a long anoestrus (McNeilly 1987). This lactational anoestrus
effectively causes a loss of positive oestradiol feedback signal to the pre-ovulatory LH surge,
and therefore prevents ovulation and oestrus. Other causes of prolonged anoestrus include
giving birth to twin calves, having a retained placenta, and having uterine infections or milk
fever (Mwaanga & Janowski 2000).
There are two main drivers of anoestrus: physiological and pathological states.
Physiological states of anoestrus occur before puberty and during pregnancy for up to 60
days following parturition, whereas pathological anoestrus is defined when insufficient
oestrus occurs beyond 60 days post-calving (Mwaanga & Janowski 2000). Clinically, there
are four different types of anoestrus in cows: (1) silent heat, (2) cystic ovarian disease, (3)
ovarian afunction or hypofunction, and (4) corpus luteum pseudograviditatis (Jeong et al.
1996). Common symptoms of anoestrus are having small, flaccid uteri and small, inactive
ovaries with no palpable corpus luteum or follicle (Mukasa-Mugerwa 1989).
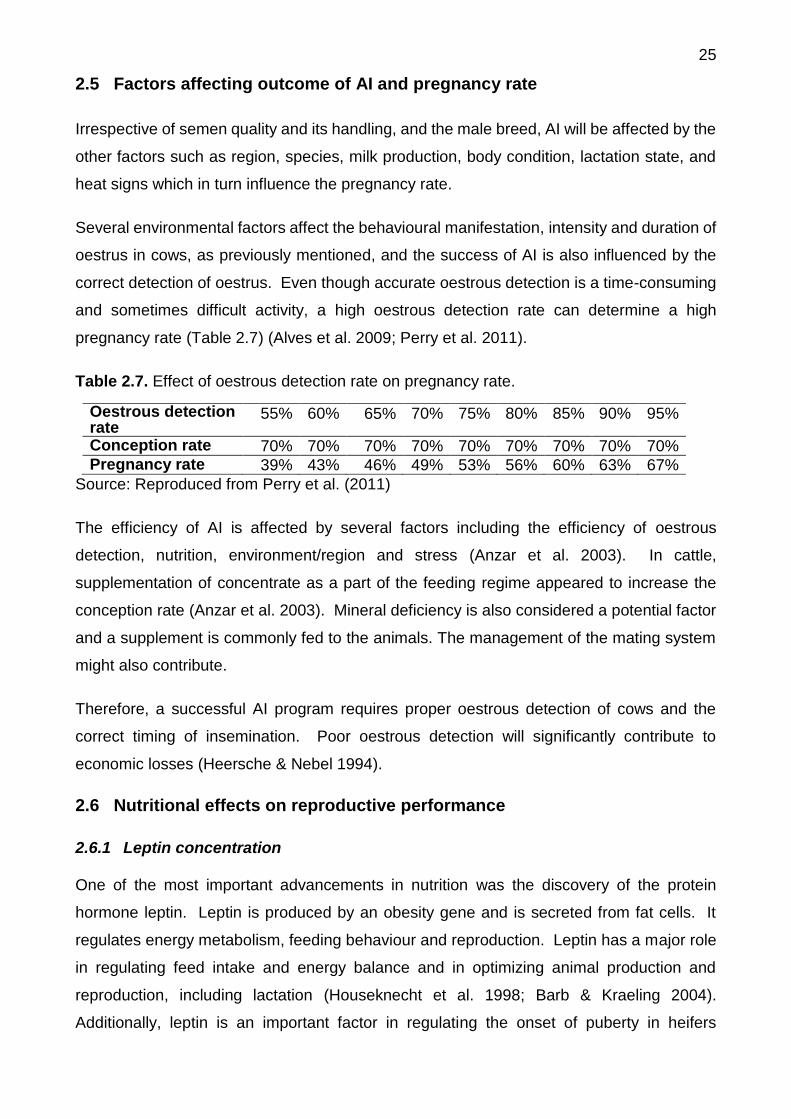
25
2.5 Factors affecting outcome of AI and pregnancy rate
Irrespective of semen quality and its handling, and the male breed, AI will be affected by the
other factors such as region, species, milk production, body condition, lactation state, and
heat signs which in turn influence the pregnancy rate.
Several environmental factors affect the behavioural manifestation, intensity and duration of
oestrus in cows, as previously mentioned, and the success of AI is also influenced by the
correct detection of oestrus. Even though accurate oestrous detection is a time-consuming
and sometimes difficult activity, a high oestrous detection rate can determine a high
pregnancy rate (Table 2.7) (Alves et al. 2009; Perry et al. 2011).
Table 2.7. Effect of oestrous detection rate on pregnancy rate.
Oestrous detection rate
55% 60% 65% 70% 75% 80% 85% 90% 95%
Conception rate 70% 70% 70% 70% 70% 70% 70% 70% 70% Pregnancy rate 39% 43% 46% 49% 53% 56% 60% 63% 67%
Source: Reproduced from Perry et al. (2011)
The efficiency of AI is affected by several factors including the efficiency of oestrous
detection, nutrition, environment/region and stress (Anzar et al. 2003). In cattle,
supplementation of concentrate as a part of the feeding regime appeared to increase the
conception rate (Anzar et al. 2003). Mineral deficiency is also considered a potential factor
and a supplement is commonly fed to the animals. The management of the mating system
might also contribute.
Therefore, a successful AI program requires proper oestrous detection of cows and the
correct timing of insemination. Poor oestrous detection will significantly contribute to
economic losses (Heersche & Nebel 1994).
2.6 Nutritional effects on reproductive performance
2.6.1 Leptin concentration
One of the most important advancements in nutrition was the discovery of the protein
hormone leptin. Leptin is produced by an obesity gene and is secreted from fat cells. It
regulates energy metabolism, feeding behaviour and reproduction. Leptin has a major role
in regulating feed intake and energy balance and in optimizing animal production and
reproduction, including lactation (Houseknecht et al. 1998; Barb & Kraeling 2004).
Additionally, leptin is an important factor in regulating the onset of puberty in heifers

26
(Williams et al. 2002; Barb & Kraeling 2004; Zieba et al. 2005). During sexual maturation in
heifers, this protein significantly increases when heifers reach puberty in late spring or early
summer (Williams et al. 2002).
In growing peripubertal heifers, fasting causes marked reductions in leptin concentration in
circulation, and a coincident reduction in LH pulse frequency (Williams et al. 2002). The
neurotransmitter neuropeptide Y has been recognised as the primary regulator of the action
of leptin in the hypothalamus to regulate the secretion of LH and growth hormone (Barb &
Kraeling 2004). Leptin stimulates both the release of GnRH and LH, by acting directly at
both the hypothalamus and the pituitary respectively (Yu et al. 1997a; Yu et al. 1997b; Woller
et al. 2001). The way that leptin affects the hypothalamic–hypophyseal axis in ruminants is
determined by both sexual maturation and nutritional status (Amstalden et al. 2002;
Amstalden et al. 2003; Zieba et al. 2003) as feed intake is influenced by leptin secretion that
is related to body fat mass and its circulation increases during pubertal development in
heifers (Zieba et al. 2005).
2.6.2 IGF-1 concentration
Growth hormone from the anterior pituitary gland stimulates the liver, and other tissues, to
produce insulin-like growth factor 1 (IGF-1). This protein hormone which is similar in
structure to insulin, may act together with insulin at the hypothalamus, pituitary and ovary
resulting in reduced LH secretion and restricted oestradiol production (Sheldon 2004).
These events will contribute to lengthen post-partum anovulatory anoestrus. Therefore, in
order to shorter the anovulatory period, supplementation of the diet with fat that reverses
the effect of negative energy balance leading to larger first dominant follicles and higher
maximum oestradiol concentrations may be a successful strategy (Sheldon 2004).
IGF-1 is often associated with leptin as a metabolic hormone. Short-term nutrient changes
can affect leptin, insulin, IGF-1, and LH pulsatility in prepubertal heifers (Amstalden et al.
2000). Increased expression of the leptin gene occurs in pubertal heifers associated with
increases in serum IGF-1 and body weight (Garcia et al. 2002). In underfed cows, plasma
IGF-1 and insulin levels are consistently depressed (Jolly et al. 1996; Cassady et al. 2009).
During pre-partum and at calving, the association between BCS with IGF-1, insulin, non-
esterified fatty acids (NEFA) and urea reflect a better nutritional status (Soca et al. 2013).
Moreover, in anovulatory beef heifers given better nutrition to induce resumption of
ovulatory cycles, increased concentrations of IGF-1, LH, and oestradiol were associated

27
with increased diameter, growth rate, and persistence of the dominant follicle (Bossis et al.
2000).
2.6.3 Hormonal concentration effects on reproductive functions
The effects of gonadotropic and steroid hormones on BCS (body condition score) will be
reviewed in section 2.6.5. This section will focus on reviewing their effects on reproductive
functions. Metabolic hormones such as insulin, insulin-like growth factor-1 (IGF-1), growth
hormone, cortisol and TSH (thyroid stimulating hormone) will also be briefly addressed.
It is assumed that delay of first post-partum oestrus is related to negative energy balance.
The protein leptin is produced by white fat cells. Leptin affects the central hypothalamic
nuclei, and alters food uptake and energy-spending processes (Halaas et al. 1995;
Houseknecht et al. 1998; Houseknecht & Portocarrero 1998; Ahima & Flier 2000).
Increased body fat content will cause increased leptin production, which in turn decreases
feed intake and increases oxygen expenditure (Bartha et al. 2005). Leptin helps an animal
to survive in (severe) energy deficiency (Chilliard et al. 2001), and regulates nutritional status
and reproductive function (Liefers et al. 2005). As energy balance becomes more negative,
depressed appetite causes a decrease in energy expenditure, and this may affect
reproduction and the immune system (Bartha et al. 2005).
Growth hormone (GH) is produced from the anterior pituitary gland under the control of
Growth Hormone Releasing Hormone (GHRH) from the hypothalamus. The production of
GH can be modulated by steroids (Devesa et al. 1991), and GH appears to promote
gonadotropin action (Webb et al. 2004; Scaramuzzi et al. 2006).
Growth hormone has effects on feed intake, milk yield, and body composition. In dairy cows,
milk production can be increased by the direct action of GH (Burton et al. 1994), but also
GH together with IGF-1 can control growth and lactation in beef cattle (Lucy 2008). Feed
intake of growth hormone-treated cows can also be affected for several weeks after the
response in milk yield (McBride et al. 1988; Bauman et al. 1994). Body condition can also
be affected by GH, with GH being protein anabolic and fat catabolic (Burton et al. 1994).
Insulin growth factor (IGF-1) is produced in response to GH, mainly from the liver. IGF-1
normally acts in concert with other related hormones. IGF-1 mRNA levels in the liver can be
increased by treating animals with GH and this also increases adipose tissue concentration
of leptin (Houseknecht et al. 2000). IGF-1 together with GH controls post-natal growth and
metabolism in mammals. These hormones influence growth rate, body composition, health,

28
and aging by modulating the expression of many genes (Sumantran et al. 1992; Ho &
Hoffman 1993; Lincoln et al. 1995). Beside growth these hormones control lactation (Lucy
2008). IGF-1 and GH increase mammary gland availability of glucose and NEFAs through
providing chronic lipolytic, diabetogenic, and gluconeogenic signals to target tissues
(Houseknecht et al. 2000)..
Energy expenditure is closely correlated with glucose availability in blood plasma, and this
availability is related to the concentration of insulin. Low insulin concentration causes low
energy status (Staples et al. 1998). Insulin regulates glucose uptake and is thought to
stimulate the development of follicles, thus providing a mechanism for nutrition to affect
ovulation (Somchit-Assavacheep 2011).
The mechanisms of metabolic adaptation to underfeeding in ruminants can be seen in
Figure 2.4. The adaptation of ruminants to under-nutrition reduces leptin concentration, in
turn increasing cortisol (Bornstein et al. 1997). Cortisol contributes to the condition of
underfeeding through metabolic adaptations such as protein mobilization and
gluconeogenesis, and stimulates re-feeding behavior (Chilliard et al. 2001). Therefore, the
interactions between cortisol, insulin and leptin in ruminants might serve an important role
in the adaptation of under-feeding and re-feeding (Chilliard et al. 2001).
Leptin appears to act as an important nutritional signal to the reproductive axis (Nagatani et
al. 2000). In under-fed animals, decreased leptin stimulates appetite and this is associated
with increased circulating glucocorticoids, decreased thyroid gland activity, decreased
energy expenditure and protein synthesis, and the inhibition of reproduction (Ahima et al.
1996; Chilliard et al. 2001). A suppression of pulsatile LH and cessation of gonadal activity
is noted in ruminant animals (Nagatani et al. 2000). Increased cortisol drives metabolic
adaptations including protein mobilization, gluconeogenesis, and stimulates re-feeding
behavior (Chilliard et al. 2001). Cortisol also appears to influence progesterone metabolism
(Fitko et al. 1996). During the luteal phase of the oestrous cycle, cortisol concentration is
increasing during early lactation (Dieleman et al. 1986; Hockett et al. 2000; Lyimo et al.
2000). Increasing cortisol concentrations suppressed LH in cows (Echternkamp 1984; Peter
et al. 1990; Stahringer et al. 1994).

29
Figure 2.4. The interactions between plasma leptin, cortisol and insulin in ruminants during underfeeding and re-feeding.
Source: (Bornstein et al. 1997; Chilliard et al. 1998; Chilliard et al. 2001).
Oestradiol is an active form of oestrogen and it is high in concentration in cycling cows. The
form of oestradiol is oestradiol benzoate, and this hormone together with a progesterone
releasing device has been shown to improve reproductive performance in suckled Bos
indicus cows because of their beneficial effect on LH pulse frequency, follicle growth and
ovulation (Baruselli et al. 2004). Similarly, ovulation is driven by the actions of oestrogen
(Wintermantel et al. 2006). An increased level of oestrogen has been shown to be able to
trigger the ovulatory surge of LH in the reproductive cycle (Labhsetwar 1970). In addition,
during caloric restriction, the set-point for gonadotropin suppression could be altered by
oestrogen (Nagatani et al. 2000). The biosynthetic and secretory behavior of the GnRH
neurons, and in turn fertility, is likely to be regulated by oestrogen (Herbison & Pape 2001).
Progesterone and oestrogen concentrations in blood plasma are important metabolic
parameters associated with a female’s reproductive cycle, and it has been reported that
these vary within breeds during different stages of oestrous cycle. In Bali cows, for example,
Belli & Jelantik (2011) reported a plasma progesterone concentration 0.11 ng/mL for animals
fed on a concentrate and 0.06 ng/mL for those not fed on concentrate. A much higher
concentration during the oestrous cycle (8.95 ng/mL) was reported by (Bakry 1994) for Bali
cows reared in West and East Nusa Tenggara, Indonesia. In Holstein heifers, the
progesterone concentration was 2.4 ng/mL from day 4 after oestrus and continued to
increase on day 6 and 8 to 5.2 and 7.7 ng/mL, respectively (Valdez et al. 2005). Lammoglia
et al. (1997) reported an average level of less than 1 ng/mL at oestrus and between 5 to 9
ng/mL at the peak of the luteal phase for normal beef and dairy cows.

30
For plasma oestrogen level, the study of Valdez et al (2005) found a peak concentration of
320 ng/mL for dairy cows. Arimbawa et al. (2012) stated recently that research reporting
reproductive hormone levels in Bali cattle is lacking and is worthwhile pursuing.
A significant difference in progesterone concentration between pregnant and non-pregnant
cows has been repeatedly demonstrated. In non-pregnant Bali cows, the plasma
concentration was 0.52 ng/mL (Astiti & Panjaitan 2013) which was higher than the level
previously reported for non-pregnant Zebu cows (Mukasa-Mugerwa & Tegegne 1989). Astiti
& Panjaitan (2013) concluded that a Bali cow is considered to be not pregnant when its
plasma progesterone concentration is less than 3.0 ng/mL. A similar progesterone
concentration was reported for non-pregnant dams in a different study (Ricoy et al. 2001).
In Bali cattle, an increase in plasma progesterone concentration to 1.9 ng/mL was observed
on day 4 of the oestrous cycle and it continued to increase to as high as 5.4 ng/mL before
returning to its basal concentration of 1 ng/mL (Belli & Jelantik 2011).
A hormonal treatment using progesterone to stimulate the manifestation of oestrus
particularly for animals in a herd, has gained wide use as physical and electronic oestrous
detection methods are most likely to be unable to be efficiently used in the field among the
animals where oestrus occurred in different times. Treatment with progesterone stimulates
the normal ovulatory response to the introduction of male animals (Hunter et al. 1971;
Oldham et al. 1980). In an oestrous synchronization program, the use of low dosages of
progestogens can result in persistence of the dominant follicle (Sirois & Fortune 1990; Savio
et al. 1993). Greater frequency of the LH pulse occurred in cows given one half of a
progesterone intravaginal device (Roberson et al. 1989), which implies that low
concentration of progestogens resulted in increasing LH pulse frequency.
There are four categories of progesterone concentration described by Mann et al. (2005) as
follows: (1) normal cyclicity in which the progesterone concentration remains below 3 ng/mL
for less than one week followed by progesterone concentrations exceeding 3 ng/mL for more
than two weeks, or high levels of progesterone concentration (exceeding 3 ng/mL) in
association with confirmed pregnancy, (2) cessation of luteal activity in which progesterone
concentration is less than 3 ng/mL for more than two weeks followed by a period of luteal
activity, (3) prolonged luteal activity in which the concentration of progesterone is greater
than 3 ng/mL for more than three weeks with the absence of pregnancy, and (4) erratic
phases, which fail to conform with 1, 2 or 3.

31
Oestradiol acts contrary to progesterone; low concentration of progesterone is associated
with high concentration of oestradiol (Chandler 2001). Increased serum concentrations of
oestradiol-17ß have been found with the induction of low concentrations of progestogen
using controlled internal drug releasing devices (CIDRs) containing progesterone (Sirois &
Fortune 1990). Low levels of progesterone cause an increase of LH secretion, resulting in
increased oestradiol-17ß production by follicles (Chandler 2001).
2.6.4 Effect of nutrition
It is well established that nutrition can influence reproductive performance. The most
common problems associated with poor nutrition are decreasing sperm quality in bulls and
delayed puberty and disrupted cyclic ovarian function in heifers and cows (Isle et al. 2007).
Nutritional factors may also exacerbate or inhibit the incidence of oestrous behaviours in
cows (Bolanos et al. 1998; Orihuela 2000).
Nutrition can influence gonadotropin-releasing hormone (GnRH) secretion and the
frequency of the LH pulse (Bolanos et al. 1998) by stimulating the release of LH from pituitary
cells (Naor & Catt 1981). Nutrition affects FSH and LH pulse frequency through short term
modulation of ovarian function (Martin et al. 1992a). This will trigger the development of
follicles and will influence the diameter of the dominant follicle (Murphy et al. 1990). Feed
intake may act on steroid hormones and folliculogenesis (Dawuda et al. 2002). Furthermore,
levels of metabolic hormones and substrates will also be affected by nutrition (Stobart 1994).
Moreover, it indirectly affects conception rate by influencing progesterone metabolism and
its dominant follicle (Crowe 2008a).
Insufficient available energy will delay puberty and lengthen the post-partum anoestrus
interval leading to anoestrus in cyclic females (Short & Adams 1988). These synergetic
effects will also occur when energy intake before calving is insufficient and will also reduce
the rate of appearance of follicles (10 mm) (Perry et al. 1991). The diameter of the dominant
follicle is affected by low feed intake and intake will also affect ovulatory follicle and corpus
luteum sizes of both Bos indicus and Bos taurus heifers (Murphy et al. 1991).
Protein and minerals play an important role in preventing disorders that are related to fertility
(Wilde 2006). Proteins act to protect body functions against several viral infections, and
minerals play a crucial role in supporting many body functions. These functions may
subsequently influence reproductive performance and infertility.

32
Providing higher intake of protein in early lactation will subsequently increase milk
production (McCormick et al. 1999). Metabolisable protein requirements need to be met in
growing and lactating cows by providing them with supplemental undegradable intake
protein (Klopfenstein 1996). However, reproductive efficiency may be depressed by very
high levels of dietary protein intake (Jordan & Swanson 1979; Ferguson et al. 1988;
Blanchard et al. 1990; Ferguson et al. 1993; Butler et al. 1996).
The effect of mineral deficiency on reproductive function has also long been recognised
(Hurley & Doane 1989). Deficiencies of mineral micronutrients are very common during the
dry season in tropical countries with low-input systems such as Indonesia. During the dry
season, the availability of fodder is extremely low in mineral content. This will limit the
reproductive performance, and subsequently the overall productivity of animals (Smith &
Akinbamijo 2000). Selenium (Se) has been one of the major minerals affecting the
reproductive functions of cattle. Together with vitamin E, selenium acts as a cellular
antioxidant that protects cells from the harmful effects of hydrogen peroxide and other
peroxides formed from fatty acids (Smith & Akinbamijo 2000). Deficiency of selenium and
vitamin E will cause the accumulation of free radicals, damage to cell membranes, and the
disruption of several processes linked to the synthesis of steroids (Staats et al. 1988),
prostaglandins (Hemler & Lands 1980), sperm motility (Alvarez & Storey 1989), and the
development of the embryo (Goto et al. 1992). These deficiencies will also influence various
components of reproduction, including ovulation rate (Harrison et al. 1984), uterine motility,
sperm motility and transport (Segerson & Libby 1982; McKenzie et al. 1998), conception
rate and post-partum activities (Arechiga et al. 1994), foetal membrane expulsion (Wichtel
et al. 1996), embryo survival, milk production and post-natal growth (Anke et al. 1989).
Reproductive disorders in cattle caused by dietary Se deficiency include erratic, weak or
silent heat periods, delayed conception, poor fertilization, cystic ovaries (Corah & Ives 1991),
reduced sperm motility (McKenzie et al. 1998), reduced uterine motility (Segerson & Libby
1982), mastitis (Olson 1996), and retained foetal membrane (Trinder et al. 1969; Campbell
& Miller 1998). In contrast, supplementation of Cu, Zn, Mn and Se during lactation at several
months before insemination show improvements in conception rates and days to first service
(Wilde 2006).
Deficiencies of other specific nutrients such as cobalt, manganese, copper, phosphorus and
calcium will also influence reproductive traits. Inadequate cobalt will cause abnormal first
oestrus and irregular oestrous cycle, and insufficient manganese will influence corpus
luteum activity and depress conception rates (Hidiroglou 1979). Insufficient copper (at less

33
than 10 ppm) will result in anoestrus (Hidiroglou 1979). In heifers, inadequate cobalt and
insufficient phosphorous will lead to late puberty in heifers. Manganese deficiency will also
cause infertility with anoestrus and delay the onset of oestrus (Major et al. 2007). In general,
phosphorous deficiency depressed fertility. This deficiency induces a lowered conception
rate, irregular estrus and anestrus, decreased ovarian activity, and increased incidence of
cystic follicles (Morrow 1980; Pugh et al. 1985). Excessive calcium concentration (King
1971) can inhibit the absorption of microelements such as phosphorous, manganese, zinc,
and magnesium in the intestine (Hurley & Doane 1989). This will lead to poor body condition
and will ultimately affect reproduction. Mineral content is low across Indonesia especially in
the crop residues such as rice straw but they are often not the first limiting nutrient in
production systems.
Level of nutrition influences Insulin which in turn regulates glucose uptake and
gluconeogenesis, and it is likely to mediate the effect of nutrient intake on ovulation through
regulating the development of follicle (Somchit-Assavacheep 2011). These reproductive
traits are affected by poor nutrition causing reduced sensitivity of granulosa cells to FSH
stimulation resulting in decreased follicle size (Murphy et al. 1991). Adequate nutrition at
key points in the reproductive cycle is vital for early return to ovarian cyclicity (Dobson &
Kamonpatana 1986).
Knowledge of precise nutritional needs for each stage of the reproductive process and
interaction between metabolic status and reproductive performance is essential to maintain
normal reproductive status and production of cattle (Fatet et al. 2011). Earlier sections have
also outlined effects of nutrition on steroidal function.
2.6.5 The role of kisspeptin in metabolic regulation of reproduction
The major integrator of metabolic signals in the hypothalamus that stimulates GnRH
secretion (Messager et al. 2005) is called kisspeptin. This protein, part of new family of
neuropeptides and its receptors, has been shown to play important roles in GnRH and LH
secretions (Smith et al. 2007; Kadokawa et al. 2008; Ahmadzadeh et al. 2011). Furthermore,
Williams and Amstalden (2010) indicated that it may have an essential role on pubertal
development (Seminara et al. 2003). Moreover, disrupted sexual maturation is most likely
due to the mutated genes encoding kisspeptin and its receptor. Reproduction control for
gonadal steroid hormones such as oestradiol and progesterone is also most likely controlled
by the gene encoding kisspeptin of kiss1 and it is expressed in specific area of the
hypothalamus namely preoptic area and arcuate nucleus (Pielecka-Fortuna et al. 2008). The

34
kisspeptin effect, as a potent stimulator of LH release in mammals including cattle
(Kadokawa et al. 2008), is assumed to be triggered by direct action on GnRH neurons (Smith
et al. 2008).
2.6.6 Body condition score
Estimation of Body condition score (BCS) is one of the easiest and most reliable methods
for assessing nutritional status and its effect on the reproductive cycle (Delgado et al. 2004).
The cow’s nutritional status (reflected in its BCS) and resultant hormonal profile affects
fertility and reproductive performances. BCS can be used to predict growth and reproductive
performance (Soares & Dryden 2011). These scoring methods are very useful to guide cow
management and success in reproduction (Delgado et al. 2004; Morris et al. 2006;
Hoedemaker et al. 2008; Jilek et al. 2008; Hoedemaker et al. 2009).
The optimal BCS for mature cows at their first cycling is expected to be at BCS 5 or greater
(1 to 9 scale) to achieve adequate reproductive function in the breeding season. In order to
make BCS more practical, sometimes a 1-5 scale is also used instead of 1-9, with 1 point
describing thin to 5 points identifying an obese, and 2.5 points reflecting the average body
weight (Montiel & Ahuja 2005).
BCS helps to evaluate the nutritional status of the cow (Short et al. 1990), reflecting the
availability of the energy reserves for activity, basic metabolism, growth, and lactation (Jolly
et al. 1995; Montiel & Ahuja 2005). This method gives a sufficiently reliable estimation of
body energy reserves with ease and low cost (Ezanno et al. 2003). Animals with poor body
condition are generally less likely to reproduce than those in moderate or good body
condition (Dunn & Moss 1992). The attainment of puberty of an animal is predominantly
indicated by its BCS. Furthermore, sufficient energy stores are essential for maintaining
cow’s health, reproductive function, and productivity capacity (Edmonson et al. 1989). Cows
with low body energy reserves will suffer from diseases, metabolic disorders, reproductive
failure, reduction in milk yield, and late puberty in heifers (Edmonson et al. 1989). Several
studies have indicated the negative effects of low BCS on calving, lactation, health and
fertility (Gearhart et al. 1990; Ruegg et al. 1992; Waltner et al. 1993; Ruegg & Milton 1995;
Heuer et al. 1999).
The overall reproductive and economic efficiency in a herd could be maximized by
optimising BCS at calving and weaning. BCS at calving influences calf survival, calf vigor,
and ultimate reproductive performance of a cow. At weaning, BCS is normally maintained

35
or increased when suckling is terminated. There are five important times to evaluate body
condition score in a cowherd (1) thirty days before mating, (2) ninety days after mating, (3)
at weaning, (4) a hundred days before calving, and (5) at calving.
There are several ways to evaluate BCS. Sight and touch are common methods. Palpation
of the back bones and lumbar processes can give an accurate estimate of BCS, as it allows
assessment of fat level (Montiel & Ahuja 2005).
Each type of cattle breed has different body conformation as determined by their genetics.
These genetic traits will contribute to their performance in their surrounding environment.
There are two main types of breeds of cattle, namely, tropical breeds such as Ongole, Zebu,
Brahman, Bali, etc. and temperate breeds (European breeds) such as Simmental, Limousin,
Hereford, Angus, etc. In order to be most useful, scoring systems of BCS should be suited
to the animal type.
BCS evaluation was originally designed for beef cows (Lowman et al. 1973), and, for calves
and weaners: this evaluation is not as reliable as they tend not to have heavy fat deposits
(Ndlovu et al. 2007). Bos indicus cattle deposit more fat internally than subcutaneously,
compared to temperate beef breeds and observers need to be trained in BCS with specific
breed types (Ledger 1959; Kempster 1981).
The measurement of frame size in a cow includes hip height and BCS, which is related to
the carcass composition (percentage crude protein, carcass lipid or empty body fat)
(Thompson et al. 1983; Houghton et al. 1990). At a given age, hip height is highly correlated
with growth rate (Vargas et al. 1999). Fat depth and eye muscle area are related and similar
to the relationship between body composition and energy reserves in beef cows (Bullock et
al. 1991). In reproductively active cows, hip height together with liveweight and subsequent
condition score are moderately heritable (Burrow 2001; Arango et al. 2002). Height by itself,
without reference to BCS, is misleading depending on breed type. Animals with large frame
size (hip height) require more feed to maintain a desired level of BCS.
2.7 Faecal progestogens
Changes in plasma progesterone concentration are associated with the oestrous cycle in
cattle and it has been proposed that a similar pattern will occur in the faeces as steroidal
secretions occur into the digestive tract. Correlations in cattle between plasma and faecal
changes in concentration have been previously reported by Masunda et al. (1999) in tropical
Nkone cows in Zimbabwe, by Rosnina et al. (2012) in tropical Kedah Kelantan heifers and

36
cows in Malaysia, by Hattab et al. (2000) in buffalo cows in Egypt, and Desaulniers et al.
(1989) in Holstein-Freisian dairy cows and even wild muskoxen cows in Canada. These
studies support the hypothesis that faecal sampling and measurement of faecal
progestogens is a valid technique to monitor reproductive cyclicity in cattle in a non-invasive
manner. This could be done with Bali cattle but no reports appear to exist.
2.8 Conclusions
This review has highlighted the hormonal and nutritional control of reproduction in cattle but
there are little data for Bali cattle. Nevertheless they appear to have the potential for high
fertility although several factors may cause low reproductive rates in the field. These are low
BCS, access to a bull or AI, detection of oestrus and long post-partum anoestrus in some
animals. A simple management plan of controlled mating and access to a bull at appropriate
times was successful in improving reproduction rate in villages (Poppi et al. 2011) but the
number of cows studied was low. Oestrous detection was a practical and experimental
problem in all studies especially in the field. Further study of the factors which affect
reproduction in Bali cows and heifers would be useful to better devise management systems
to increase reproduction in village Bali cattle.
In this thesis a number of factors affecting reproduction rate in Bali cattle was examined.
The first issue was to detect oestrus in heifers and cows and the plasma progesterone profile
was a suitable method as outlined in this literature review. However this is not practical and
a less invasive method using faecal progesterone was examined. Faecal sampling could
be used experimentally in villages and on research station to investigate causes of low
reproduction. Another method, vaginal electrical conductivity, was also examined as a
practical method for oestrous detection. These methods were examined in two chapters of
this thesis. The role of body condition on various hormones involved in response to nutrition
was identified as a mechanism controlling oestrus in other cattle species and this was
investigated in Bali cattle in another chapter. Values for milk production in Bali cattle appear
low but values are scarce for heifers and another chapter examines milk production in
heifers and cows and the return to oestrus under lactation. Finally access to a large data set
on reproduction in cows and heifers in various villages under various development projects
were examined to look at key factors which might control reproduction in Bali cattle in
villages so as to give guidelines for managing Bali cattle under village based smallholder
systems of production.
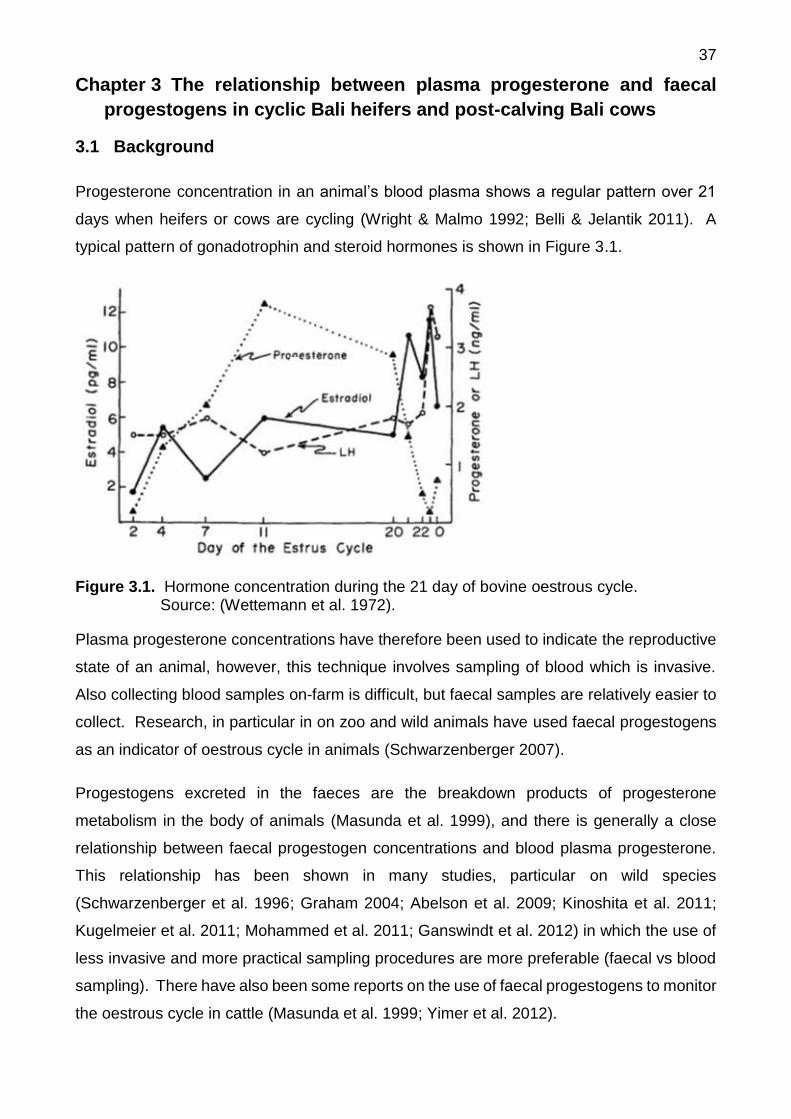
37
Chapter 3 The relationship between plasma progesterone and faecal
progestogens in cyclic Bali heifers and post-calving Bali cows
3.1 Background
Progesterone concentration in an animal’s blood plasma shows a regular pattern over 21
days when heifers or cows are cycling (Wright & Malmo 1992; Belli & Jelantik 2011). A
typical pattern of gonadotrophin and steroid hormones is shown in Figure 3.1.
Figure 3.1. Hormone concentration during the 21 day of bovine oestrous cycle. Source: (Wettemann et al. 1972).
Plasma progesterone concentrations have therefore been used to indicate the reproductive
state of an animal, however, this technique involves sampling of blood which is invasive.
Also collecting blood samples on-farm is difficult, but faecal samples are relatively easier to
collect. Research, in particular in on zoo and wild animals have used faecal progestogens
as an indicator of oestrous cycle in animals (Schwarzenberger 2007).
Progestogens excreted in the faeces are the breakdown products of progesterone
metabolism in the body of animals (Masunda et al. 1999), and there is generally a close
relationship between faecal progestogen concentrations and blood plasma progesterone.
This relationship has been shown in many studies, particular on wild species
(Schwarzenberger et al. 1996; Graham 2004; Abelson et al. 2009; Kinoshita et al. 2011;
Kugelmeier et al. 2011; Mohammed et al. 2011; Ganswindt et al. 2012) in which the use of
less invasive and more practical sampling procedures are more preferable (faecal vs blood
sampling). There have also been some reports on the use of faecal progestogens to monitor
the oestrous cycle in cattle (Masunda et al. 1999; Yimer et al. 2012).

38
The same pattern of plasma progesterone concentration during the oestrous cycle is
expected in heifers, irrespective of BCS (body condition score) and liveweight, once they
have reached puberty and showed first signs of oestrus (Ginther et al. 1989; Hopper et al.
1993).
There has been no study carried out to examine the use of faecal progestogens as an
indicator of oestrous cycles in Bali cattle. Some reports found variations between ruminants
and mammals, such as rhino, sheep, pig and horse, in steroid metabolism and excretion
(Schwarzenberger et al. 1996; Palme et al. 1997; Heistermann et al. 1998; Palme 2005)
indicating that faecal progestogen concentrations in Bali cattle may be different from those
found in sheep (Palme et al. 1996).
Hypotheses:
1. Faecal concentrations of progestogens will correlate with plasma progesterone
concentrations and therefore can be used to monitor oestrous cycles and confirm
pregnancy in Bali heifers and cows.
2. There will be no effect of BCS on plasma progesterone or faecal progestogen
concentrations.
3.2 Aim of experiment
The aim of this experiment was to determine the progesterone pattern during the oestrous
cycle (3a) and post-calving (3b) in Bali cows. The experiments were designed to test
whether it was possible to use faecal sampling with measurement of faecal progestogens to
monitor the reproductive status of Bali cattle.
3.3 Materials and methods
3.3.1 Location
The experiments were carried out between 13th August to 10th September 2013 (part (a)
cyclic animals) and between 29th August 2014 to 28th February 2015 (part (b) post-calving
animals) in Lombok West Nusa Tenggara (NTB) Indonesia. The region of Nusa Tenggara
comprises two provinces namely, East Nusa Tenggara (NTT) and West Nusa Tenggara
(NTB). This experiment was only conducted in NTB. This province is located in eastern
Indonesia and situated between 115° E and 119° E longitude and from 8° S to 9° S latitude.
There are several small islands around the province but the main islands are Sumbawa and
Lombok with the total area is about 20,153 km2. The Nusa Tenggara region is affected by

39
north-west monsoons from November to April with several modifications influenced by the
effects of orographic rainfall. Between December and April is the rainy season with the
wettest months normally being January and February. The dry season is normally from May
to October. The average annual daily minimum and maximum temperatures for rainy and
dry seasons are 21.0°C and 33.3°C, and 24°C and 33.5°C, respectively.
3.3.2 Animals and diets
(a) Cyclic heifer experiment
This experiment used 12 Bali heifers which had reached puberty and shown overt oestrous
cycles. Bali heifers were weighed every month at morning time before feeding and BCS
scale 1 to 5 (Teleni et al. 1993) determined on the day prior to commencement of the
experiment. Six high body condition score (3.67 ± 0.25) and six moderate BCS (2.25 ± 0.11)
heifers were used, The heifers were ranked on liveweight within BCS grouping and randomly
allocated to individual pens with a high and moderate BCS heifer in each pen pairing.
Heifers were fed a diet of 10 g leucaena leaves DM/kg LW.d with rice straw ad libitum. The
heifers were fed twice daily at approximately 0700 and 1700 h and water was available at
all times.
The heifers used in this experiment were a subset from a larger, longer-term experiment
involving Bali heifers (n=28; 217 ± 35 kg LW and 3.1 ± 0.4 BCS) that was designed to
generate heifers in either moderate (Mod-BCS, 2.0 to 2.5) or high (High-BCS, 3.5 to 4.0)
BCS at parturition.
Oestrus was detected prior to feeding and sampling every morning and afternoon throughout
the 30 day experiment, by leading a bull past the heifers. Oestrus was detected by visual
observation and monitoring sexual behaviour which included bull attempts to mount, other
heifers attempts to mount, a decrease in feed intake, or other signs such as vaginal mucus
secretion, reddening of vulvar mucus membrane, enlarged vagina, restlessness and
vocalisations.
(b) Post-calving cow experiment
For this experiment 12 Bali first calf cows were examined for 100 days during the post-
calving period. Again these animals were from the larger, long-term experiment, with 4 High-
BCS (4.00 ± 0.20) and 8 Mod-BCS (3.13 ± 0.16) cows being used. Cows were fed King
grass (Pennisetum purpureum; 857 g OM, 77 g CP, 753 g NDF/kg DM) ad libitum throughout
pregnancy and lactation. Only King grass was fed to Mod-BCS cows. Any Mod-BCS cows

40
that had two consecutive BCS measurements of 2 were offered 10 g maize grain (Zea mays;
880 g OM, 83 g CP, 245 g NDF/kg DM) DM/kg LW.per day until BCS returned to 2.5. High-
BCS cows were fed leuceana hay (Leucaena leucocephala; 917 g OM, 239 g CP, 463 g
NDF/kg DM) ad libitum for the first 5 to 7 months of pregnancy, and during the final 2 to 4
months of pregnancy and throughout lactation. High-BCS cows were fed 10 g maize grain
DM/kg LW.per day and leucaena hay ad libitum which equated to approximately 1 kg
corn/1.5 kg leucaena hay. All cows were fed twice daily at approximately 0700 and 1700 h,
with the roughage component offered before maize grain, and water was available at all
times.
Cows were exposed to a single Bali bull prior to feeding every morning and again in the
afternoon. Cows were re-exposed to bulls for 90 days after an initial mating was recorded
to allow re-mating after failed conceptions or early embryonic losses that may have occurred.
Cows that failed to conceive or maintain a pregnancy were removed from the experiment
and the cows maintained for this experiment were 12 cows.
Figure 3.2. Oestrous behaviour with bull detection.
3.3.3 Sample collection and importation
(a) Blood samples
Blood samples were collected every 2 days over a 29 day period at puberty and during first
oestrus before mating for cyclic animals experiment (3a), and from the first calf cows 10
days after calving and every 10 days thereafter for a further 90 days (i.e. last sample was
collected 124 days after calving) for post-calving animals experiment. Samples were
collected from the jugular vein into lithium heparin vacutainers (BD) which were inverted
slowly 6 to 8 times and stored on ice prior to centrifugation at 3,000 g for 10 min at 4°C.
Plasma was collected and stored at -20°C prior to analysis.

41
Figure 3.3. Blood collection procedure.
(b) Faecal samples
Faecal samples were grabbed directly from the ground ensuring there was no urine or feed
residue contamination. Approximately 5 g of faeces were put into duplicate 5 mL tubes (i.e.
~3/4 fill the tube with faeces) using a small spoon. The lids of the tubes were secured tightly
by hand and then sealed with parafilm. The tubes were then placed in -20°C freezer until
assayed. Total faecal output was collected into a bucket placed at the back of each pen and
total weight measured. Sub-samples of daily faecal output (~100 g) were collected into
individual bags. Individual sub-samples were then oven dried at 65°C to a constant weight.
Figure 3.4. Faecal sample collection procedure.
(c) Sample importation
Plasma and faecal samples were couriered from Indonesia to Australia frozen under dry ice
and were exposed to 50 kGy gamma irradiation prior to release from quarantine under
Australian Quarantine Inspection Service (AQIS) permit IP13013231 for the first samples
set, and IP15011210 for the second one. This dosage of gamma irradiation has previously
been found to have no effect on plasma hormone concentrations (Simon Quigley, pers.
comm.).

42
3.3.4 Laboratory analysis
(a) Plasma Progesterone radioimmunoassay
Progesterone concentrations were determined in hexane extracts of the plasma samples
measured by RIA as described by Curlewis et al. (1985), except that progesterone antiserum
C-9817 (Bioquest, North Ryde, NSW, Australia) was used. Briefly, 200 µl of plasma sample
was added to 2 mL HPLC grade hexane and vigorously vortexed for 5 minutes. The solution
was then placed in an ethanol ice bath until aqueous fraction was frozen. The solvent extract
was decanted into glass tubes and dried under a gentle stream of air using a heat block
(45°C). The extract was then reconstituted in 200 µl of assay buffer (PB with 0.9% saline,
and 0.1% gelatin). For RIA, 50 µl of extracted sample, standard and quality controls were
combined with 100 µl of primary antibody (sheep anti-progesterone-11alpha-hemisuccinate,
C9817 at 1:30,000 dilution) and 100 µl of labelled progesterone tracer (1,2,6,7-H3-
progesterone) and incubated overnight at 4°C. For separation 500 µl of dextran-activated
charcoal (0.125% Norit-A charcoal with 0.0125% dextran in PBS without gelatin) was added
to each tube and incubated on ice for 10 min, before refrigerated (4°C) centrifuging for 10
min at 2500 g. The supernatant was decanted into scintillation vials and 1.5 mL of scintillant
(IGRA-Safe Plus, Perkin Elmer) added, before vials were counted on a beta counter for 2
min.
The primary antisera (C-9817) is stated by the manufacturer to exhibit high specificity for
progesterone (100%), some cross-reactivity with 11beta-hydroxyprogesterone (22%), and
minimal cross-reactivity with 20beta-hydroxy-4-pregnene-3-one (3.0%), 20alpha-hydroxy-4-
pregnene-3-one (0.2%), 17alpha-hydroxyprogesterone (0.3%), pregnenolone(2.7%), 5beta-
pregnene-3,20-dione (6.3%), 11-deoxycortisone (3.7%), corticosterone (0.4%), and
oestradiol 17beta (<0.01%).
Extraction efficiency of spiked samples was 75% and the values reported herein were not
corrected for these losses. The assay sensitivity was 0.1 ng/mL and the intra- and inter-
assay coefficients of variation were 5.9 and 8.8% respectively for quality control of 2.8 ng/mL.
(b) Faecal Progestogen ELISA
The faecal extraction protocol was simplified from previously published methods (Graham
et al. 1992). In brief, 0.5 to 0.6 g of thawed wet faeces was mixed with 5 mL of 80% methanol
and extracted overnight (16h) with shaking on a mixing vortex. The following morning an
aliquot of the supernatant (1.5 mL) was removed and centrifuged (12,000 g for 15 mins) to

43
pellet any faecal debris. The supernatant was then removed and stored at -20°C before
assay.
Faecal progestogen hormone concentrations were analysed in duplicate (50 μL aliquots)
using ELISA procedures similar to those previously reported (Munro & Stabenfeldt 1984).
Briefly, 96-well plates (Corning Costar high bind plates) were pre-coated with 125 µl of goat
anti-mouse gamma globulin (GAMG) solution (Arbor Assays A008-25MG) in coating buffer
(2.59 g/L Na2CO3.H2O and 2.936 g/L NaHCO3, adjusted pH 9.6) overnight at room
temperature, before being replaced with 175 µl blocking buffer (20mM Tris, 0.9% NaCl, 1%
BSA, pH 7.5) for at least 4 hours at room temperature. The plate was then stored at 4°C
until use. For assay, the plate was washed 5 times with 250 µl wash buffer (0.02% Tween
20) and then 100 µl of progesterone antibody (CL425 at 1:80,000 dilution) was added before
incubation overnight at room temperature in the dark. The plate was again washed 5 times
with 250 µl wash buffer, before 50 µl of standards (0.0156 to 4 ng/mL progesterone), quality
controls, and samples all diluted in phosphate buffer were added. 100 µl of progesterone-
HRP (1:400,000 dilution) was then immediately added before plates were incubated for 3
hours in the dark. For development, plates were washed 5 times with 250 µl wash buffer
before 200 µl of ABTS substrate (40mM ABTS, 1.9% H2O2, in 50 mM citrate buffer) was
added to each well and incubated between 30 and 60 min until zero standards developed
an OD of 1.0. Plates were read at wavelength of 405 nm, with a reference at 630 nm (Sunrise
TECAN Microplate reader, with XFluor4 software).
Reported cross reactivities of the progesterone antiserum (no. CL425) were 100% with
progesterone, 55% with 5α-Pregnan-3,20-dione, 12.5% with 5β-Pregnan-3β-ol-20-one, 8%
with 5β-Pregnan-3,20-dione, 2.7% with 4-Pregnen-11β-ol-3,20-dione, 2.5% with 5β-
Pregnan-3α-ol-20-one and ≤0.1% with pregnanediol, androstenedione and corticosterone.
The assay sensitivity was 0.031 ng/well and the intra- and inter-assay coefficients of
variation were 1.6% and 4.9% respectively for quality control of 0.44 ng.
(c) Assay analysis
For both plasma progesterone RIA and faecal progestagen ELISA the assay data from the
gamma counter and plate reader respectively were analysed using Assayzap software
(Biosoft, Cambridge, UK).

44
(d) Statistical analysis
To compare plasma progesterone concentrations between high and moderate BCS groups,
the individual animal progesterone profiles were aligned around the nadir in progesterone in
all normal cyclic animals. The data were then compared using a repeat-measures ANOVA
with the main effect of BCS and time.
Pearson’s correlations were used to examine the relationship between plasma progesterone
and faecal progestogen concentrations on matched samples.
3.4 Results
3.4.1 Experiment 3a Cyclic Animals
3.4.1.1 Plasma progesterone
Most of the experimental cows (9 out of 12) showed a normal oestrous cycle pattern of
plasma progesterone concentrations (Figures 3.5 and 3.6). However, there was no increase
of progesterone for two cows (#1 & 7), suggesting these cows did not ovulate and form a
normal corpus luteum. One of the cows (#21) did not show a completely normal blood
progesterone pattern and this cow appeared to have an early regression of the corpus
luteum, before another cycle (Figure 3.5). There did not appear to be any major differences
in plasma progesterone profiles between high and moderate BCS animals (Figures 3.5 and
3.6, respectively).
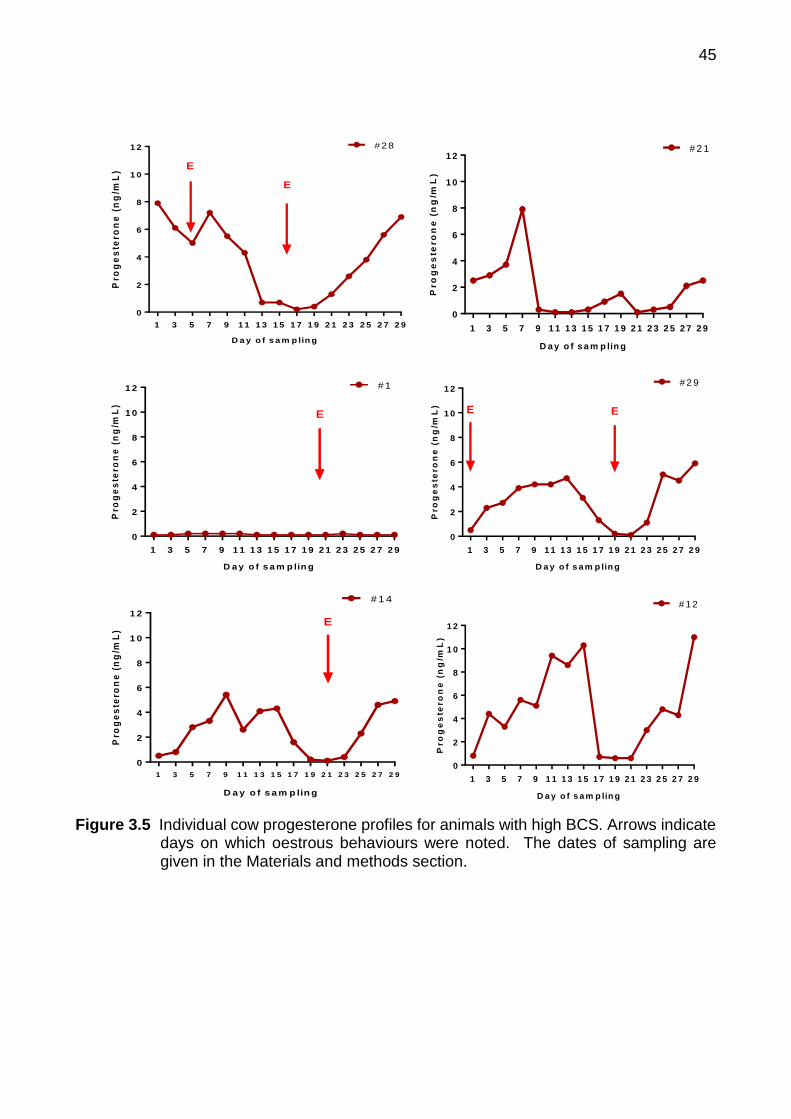
45
Figure 3.5 Individual cow progesterone profiles for animals with high BCS. Arrows indicate days on which oestrous behaviours were noted. The dates of sampling are given in the Materials and methods section.
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ron
e (
ng
/mL
)
#1
E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
Pro
ge
ste
ron
e (
ng
/mL
)
D a y o f s a m p lin g
E
# 1 4
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L)
# 1 2
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ron
e (
ng
/mL
)
# 2 8
E
E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L)
# 2 9
E E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L)
# 2 1
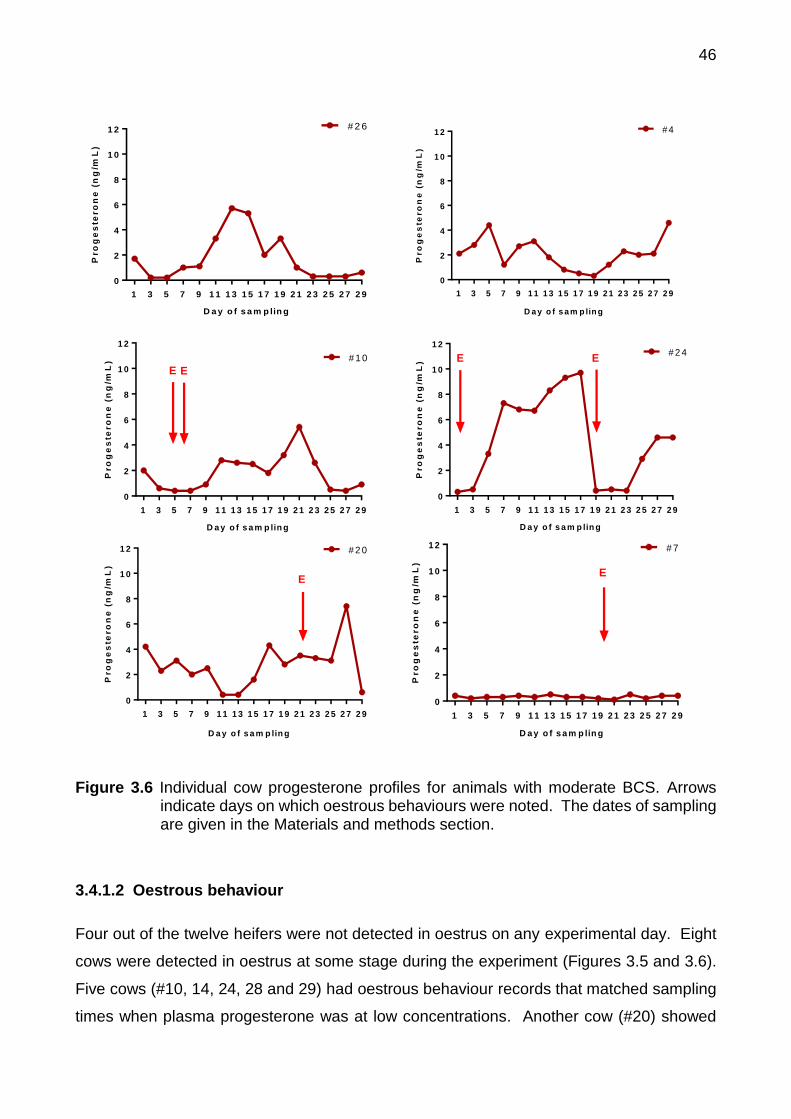
46
Figure 3.6 Individual cow progesterone profiles for animals with moderate BCS. Arrows indicate days on which oestrous behaviours were noted. The dates of sampling are given in the Materials and methods section.
3.4.1.2 Oestrous behaviour
Four out of the twelve heifers were not detected in oestrus on any experimental day. Eight
cows were detected in oestrus at some stage during the experiment (Figures 3.5 and 3.6).
Five cows (#10, 14, 24, 28 and 29) had oestrous behaviour records that matched sampling
times when plasma progesterone was at low concentrations. Another cow (#20) showed
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y o f s a m p lin g
#4
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y o f s a m p lin g
# 2 6
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L) # 1 0
E E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L)
# 2 4E E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
Pro
ge
ste
ro
ne
(n
g/m
L)
# 2 0
D a y o f s a m p lin g
E
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Pro
ge
ste
ro
ne
(n
g/m
L)
#7
E
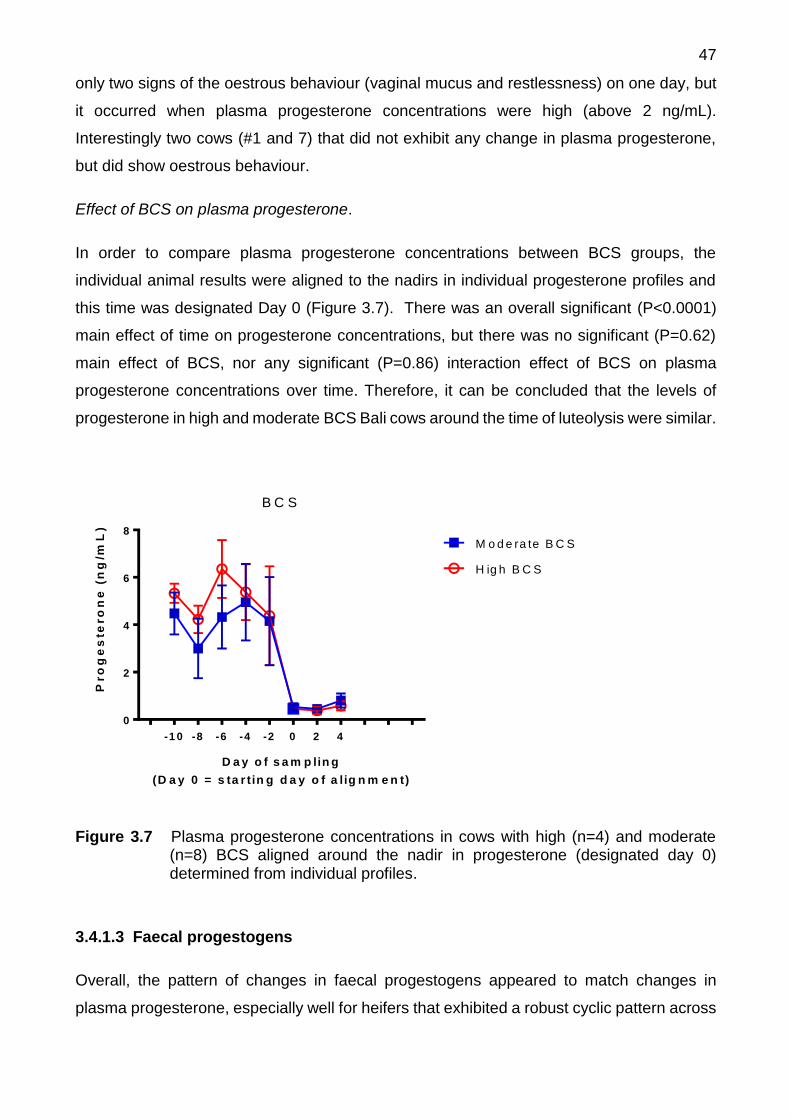
47
only two signs of the oestrous behaviour (vaginal mucus and restlessness) on one day, but
it occurred when plasma progesterone concentrations were high (above 2 ng/mL).
Interestingly two cows (#1 and 7) that did not exhibit any change in plasma progesterone,
but did show oestrous behaviour.
Effect of BCS on plasma progesterone.
In order to compare plasma progesterone concentrations between BCS groups, the
individual animal results were aligned to the nadirs in individual progesterone profiles and
this time was designated Day 0 (Figure 3.7). There was an overall significant (P<0.0001)
main effect of time on progesterone concentrations, but there was no significant (P=0.62)
main effect of BCS, nor any significant (P=0.86) interaction effect of BCS on plasma
progesterone concentrations over time. Therefore, it can be concluded that the levels of
progesterone in high and moderate BCS Bali cows around the time of luteolysis were similar.
B C S
D a y o f s a m p lin g
(D a y 0 = s ta r tin g d a y o f a lig n m e n t)
P
ro
ge
ste
ro
ne
(n
g/m
L)
-1 0 -8 -6 -4 -2 0 2 4
0
2
4
6
8
H ig h B C S
M o d e ra te B C S
Figure 3.7 Plasma progesterone concentrations in cows with high (n=4) and moderate (n=8) BCS aligned around the nadir in progesterone (designated day 0) determined from individual profiles.
3.4.1.3 Faecal progestogens
Overall, the pattern of changes in faecal progestogens appeared to match changes in
plasma progesterone, especially well for heifers that exhibited a robust cyclic pattern across

48
the sampling period; #12, 24, 10, 28, 14 and 21 (Figure 3.8). There were no marked changes
in faecal progestogen concentrations for heifers #10 and #4, unlike plasma progesterone.
Heifers #1 and #7 did not appear to exhibit normally cyclicity, as indicated by plasma
progesterone, generally had low faecal progestogen concentrations, except at times they
were highly variable.
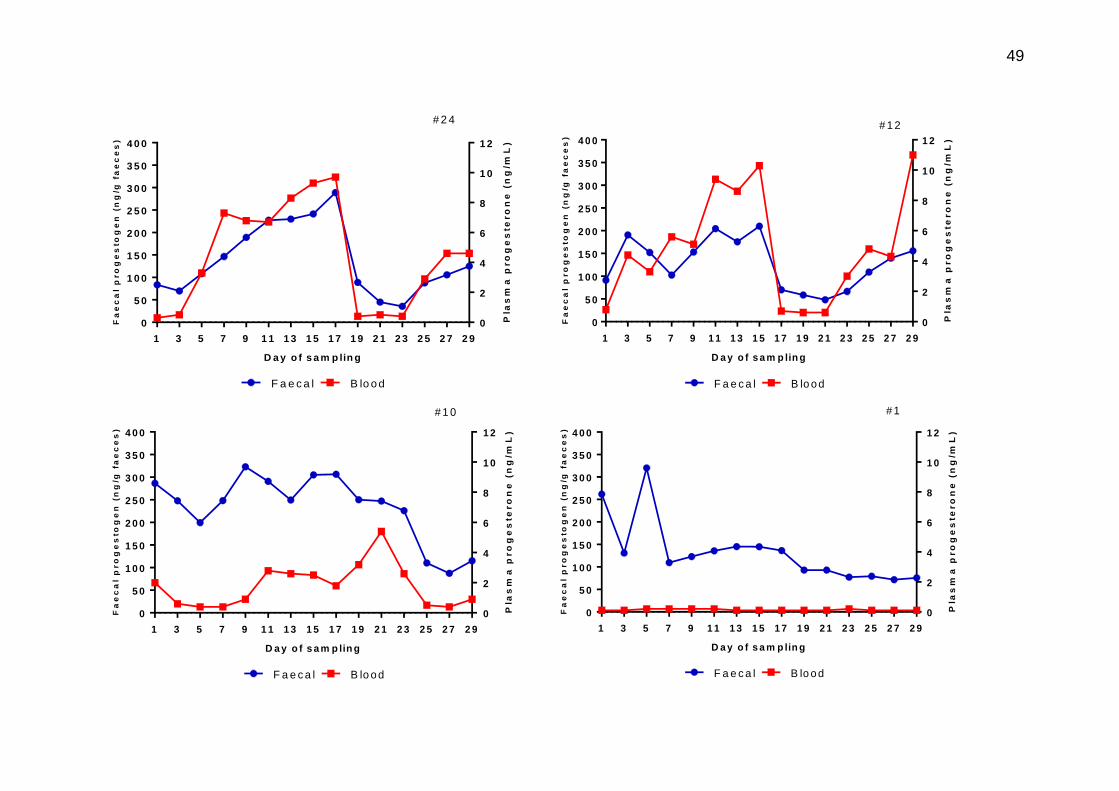
49
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 4
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
#1
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 1 0
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 1 2
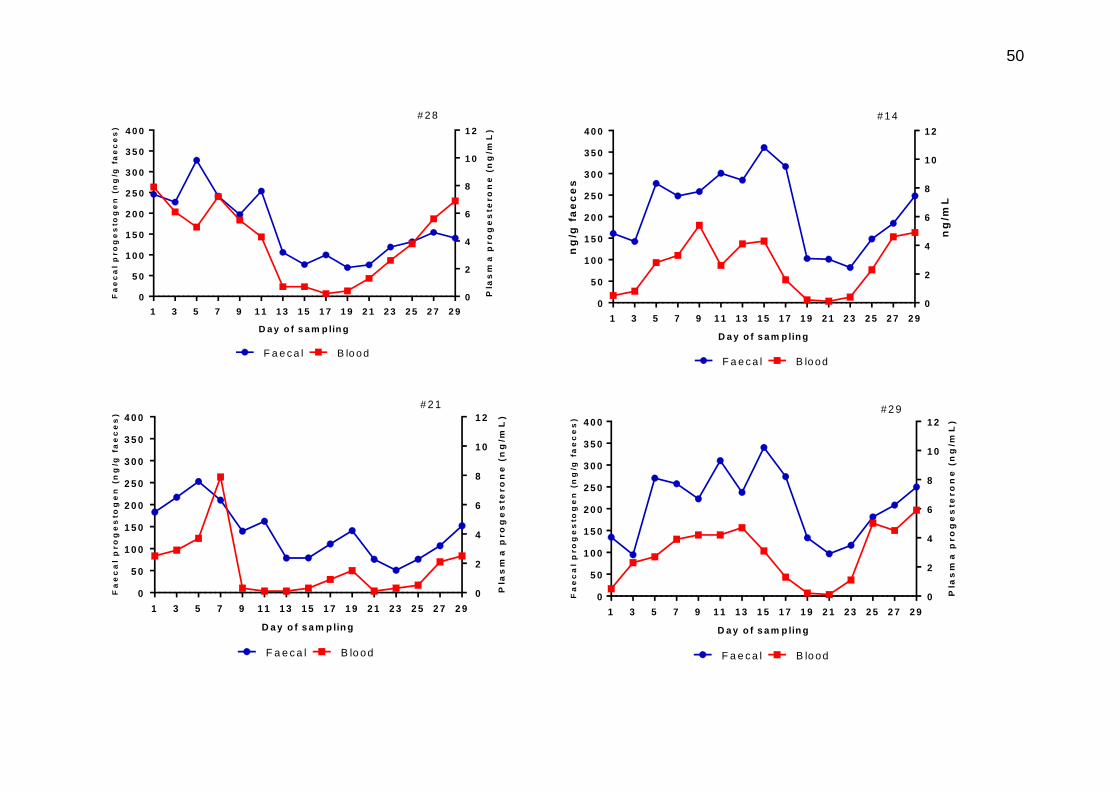
50
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 8
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
ng
/g f
ae
ce
s
ng
/mL
F a e c a l B lo od
# 1 4
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 1
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 9
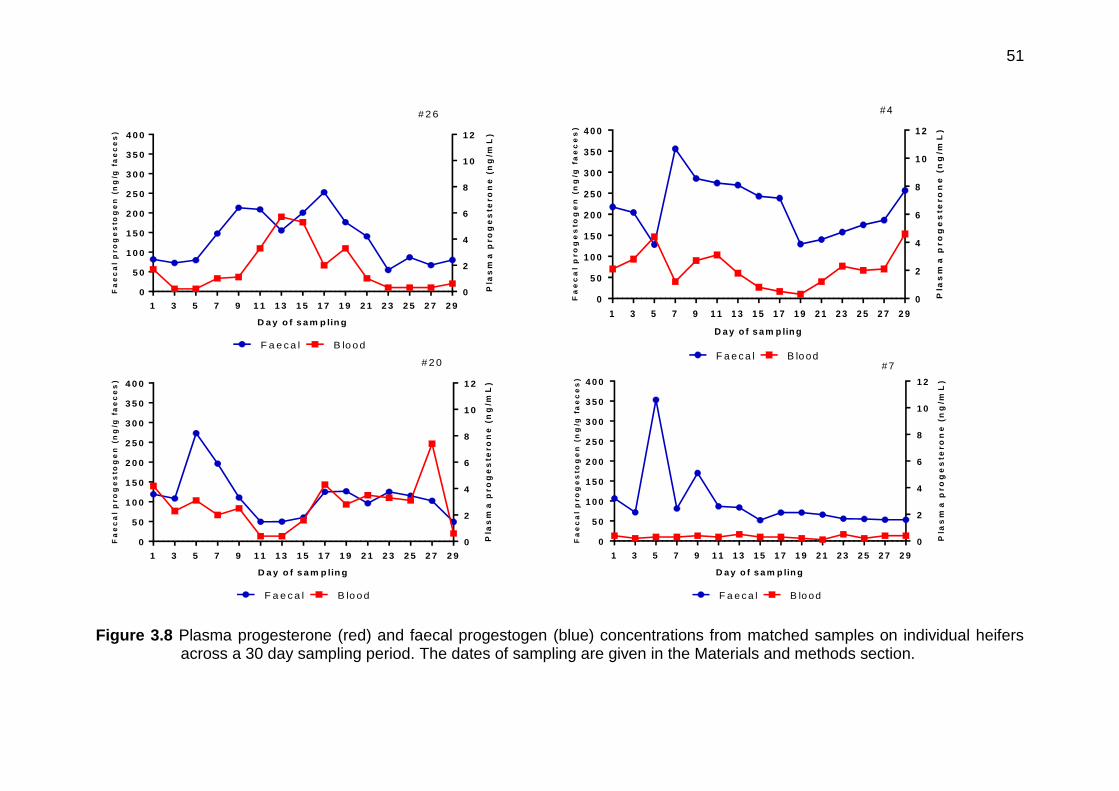
51
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 6
Figure 3.8 Plasma progesterone (red) and faecal progestogen (blue) concentrations from matched samples on individual heifers across a 30 day sampling period. The dates of sampling are given in the Materials and methods section.
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
#4
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 0
1 3 5 7 9 1 1 1 3 1 5 1 7 1 9 2 1 2 3 2 5 2 7 2 9
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
2
4
6
8
1 0
1 2
D a y o f s a m p lin gF
ae
ca
l p
ro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
#7
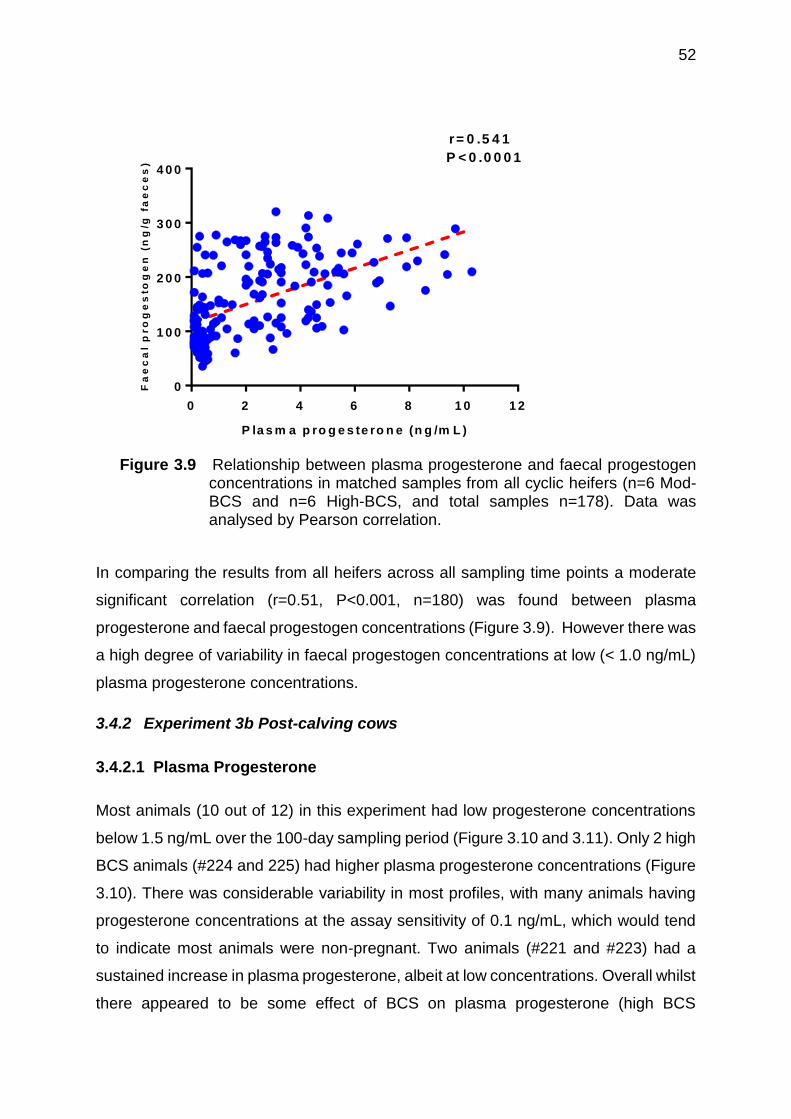
52
0 2 4 6 8 1 0 1 2
0
1 0 0
2 0 0
3 0 0
4 0 0
P la s m a p ro g e s te ro n e (n g /m L )
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
r= 0 .5 4 1
P < 0 .0 0 0 1
Figure 3.9 Relationship between plasma progesterone and faecal progestogen
concentrations in matched samples from all cyclic heifers (n=6 Mod-BCS and n=6 High-BCS, and total samples n=178). Data was analysed by Pearson correlation.
In comparing the results from all heifers across all sampling time points a moderate
significant correlation (r=0.51, P<0.001, n=180) was found between plasma
progesterone and faecal progestogen concentrations (Figure 3.9). However there was
a high degree of variability in faecal progestogen concentrations at low (< 1.0 ng/mL)
plasma progesterone concentrations.
3.4.2 Experiment 3b Post-calving cows
3.4.2.1 Plasma Progesterone
Most animals (10 out of 12) in this experiment had low progesterone concentrations
below 1.5 ng/mL over the 100-day sampling period (Figure 3.10 and 3.11). Only 2 high
BCS animals (#224 and 225) had higher plasma progesterone concentrations (Figure
3.10). There was considerable variability in most profiles, with many animals having
progesterone concentrations at the assay sensitivity of 0.1 ng/mL, which would tend
to indicate most animals were non-pregnant. Two animals (#221 and #223) had a
sustained increase in plasma progesterone, albeit at low concentrations. Overall whilst
there appeared to be some effect of BCS on plasma progesterone (high BCS
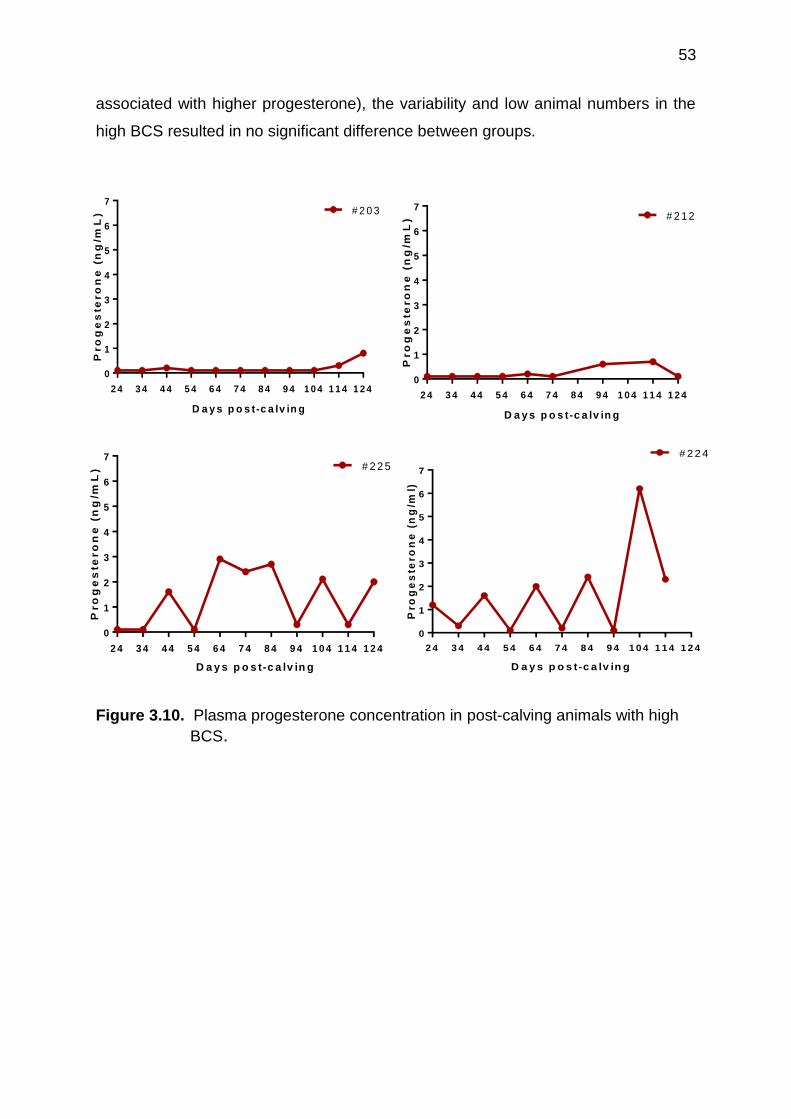
53
associated with higher progesterone), the variability and low animal numbers in the
high BCS resulted in no significant difference between groups.
Figure 3.10. Plasma progesterone concentration in post-calving animals with high
BCS.
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 5
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 0 3
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 1 2
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
l)
D a y s p o s t-c a lv in g
# 2 2 4
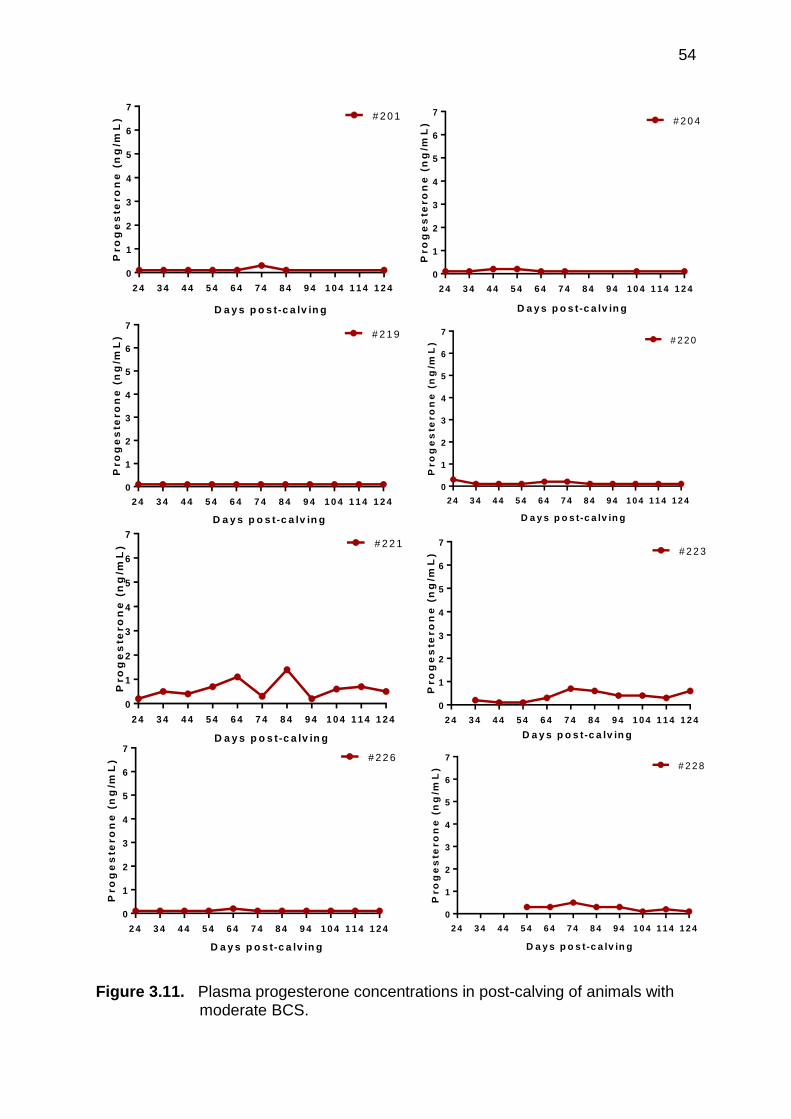
54
Figure 3.11. Plasma progesterone concentrations in post-calving of animals with moderate BCS.
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 8
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 3
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 0 4
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 6
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 1 9
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7P
ro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 0 1
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 0
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
1
2
3
4
5
6
7
Pro
ge
ste
ro
ne
(n
g/m
L)
D a y s p o s t-c a lv in g
# 2 2 1

55
3.4.2.2 Faecal progestogens
In the post-calving interval, faecal progestogen concentrations were measurable in all
animals despite low plasma progesterone concentrations. Animals #224 and #225 that
exhibited higher plasma progesterone, also had the highest faecal progestogen
concentrations. Overall a moderate significant correlation was observed between
faecal progestogen and plasma progesterone across all animals in the post-calving
period (Figure 3.13).
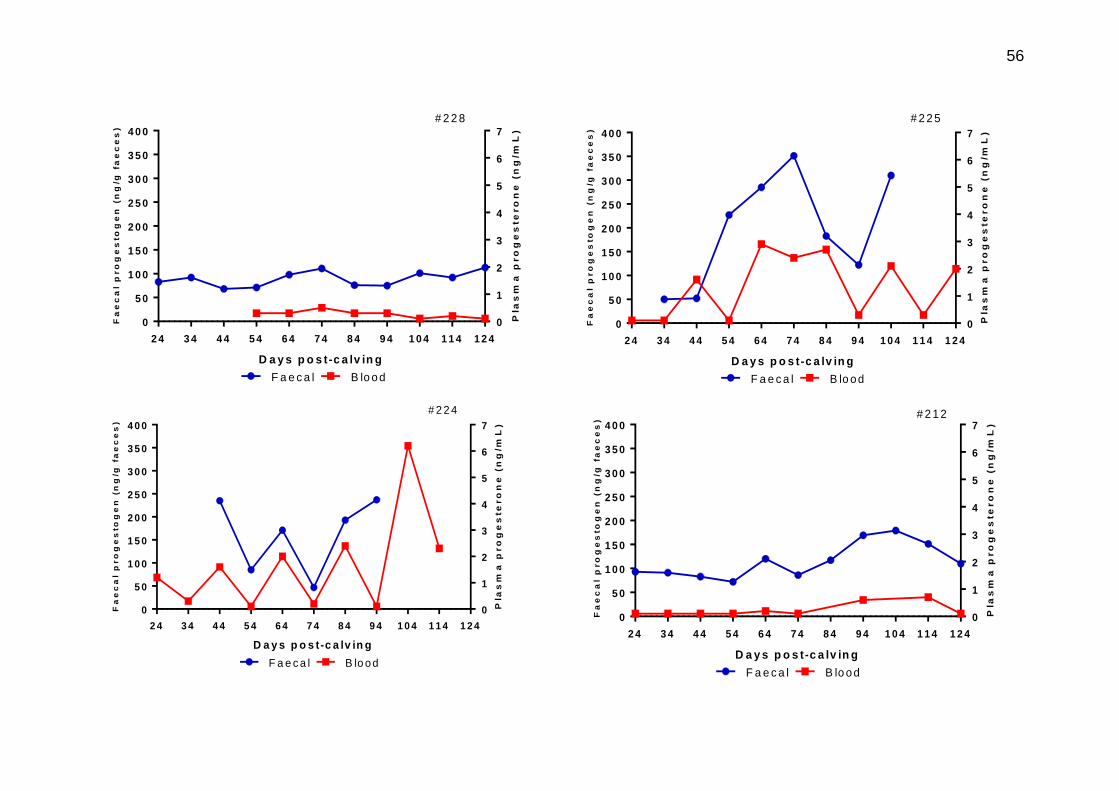
56
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 8
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 5
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 4
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 1 2
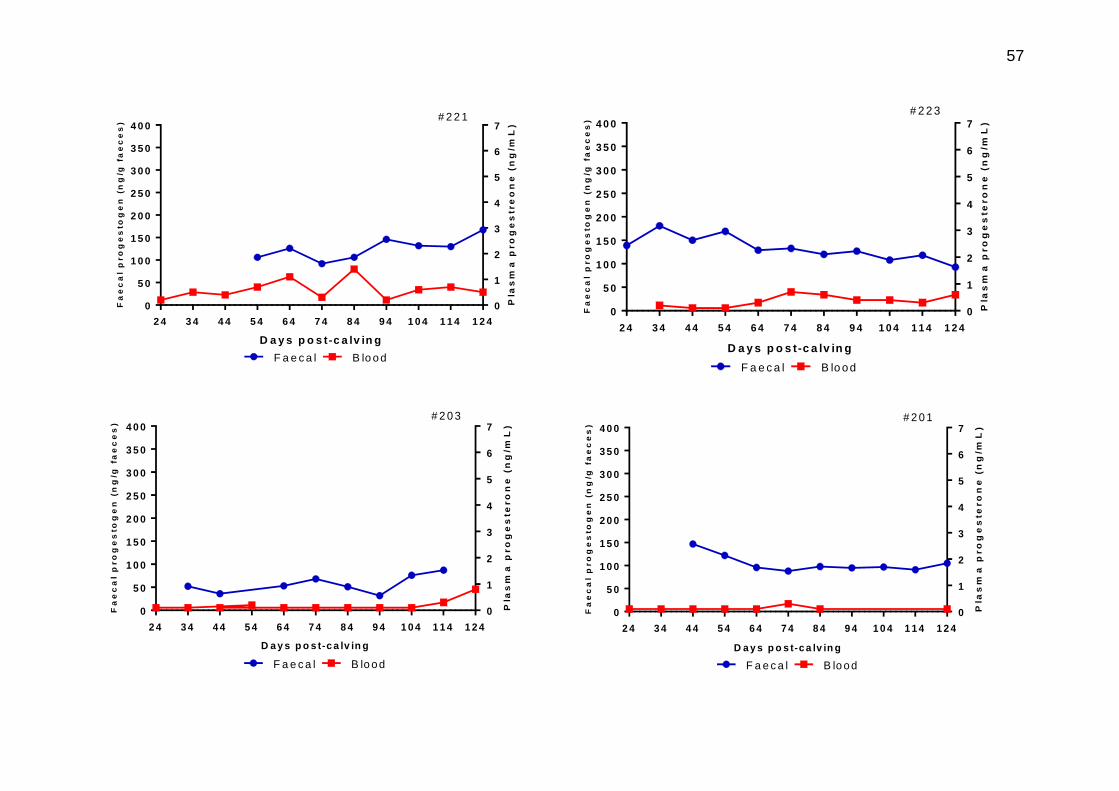
57
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
stre
on
e (
ng
/mL
)
F a e c a l B lo od
# 2 2 1
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 3
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 0 3
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 0 1
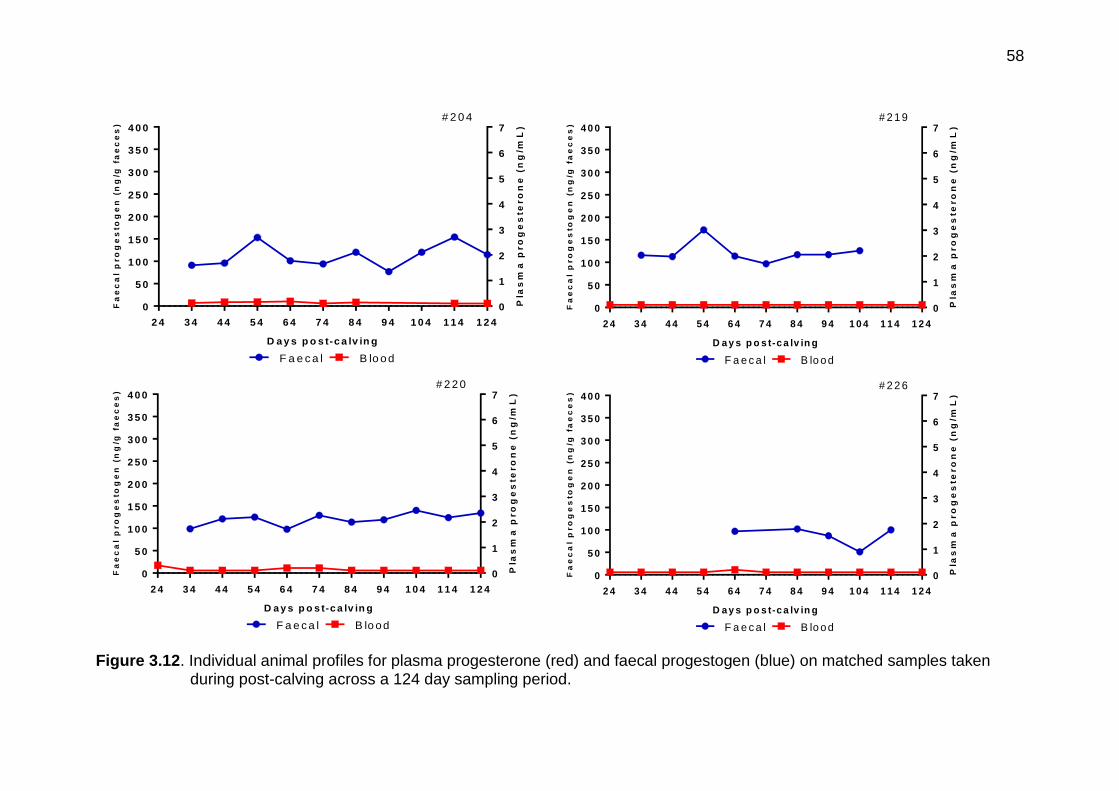
58
Figure 3.12. Individual animal profiles for plasma progesterone (red) and faecal progestogen (blue) on matched samples taken during post-calving across a 124 day sampling period.
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 0 4
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 1 9
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 0
2 4 3 4 4 4 5 4 6 4 7 4 8 4 9 4 1 0 4 1 1 4 1 2 4
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
4 0 0
0
1
2
3
4
5
6
7
D a y s p o s t-c a lv in g
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
Pla
sm
a p
ro
ge
ste
ro
ne
(n
g/m
L)
F a e c a l B lo od
# 2 2 6
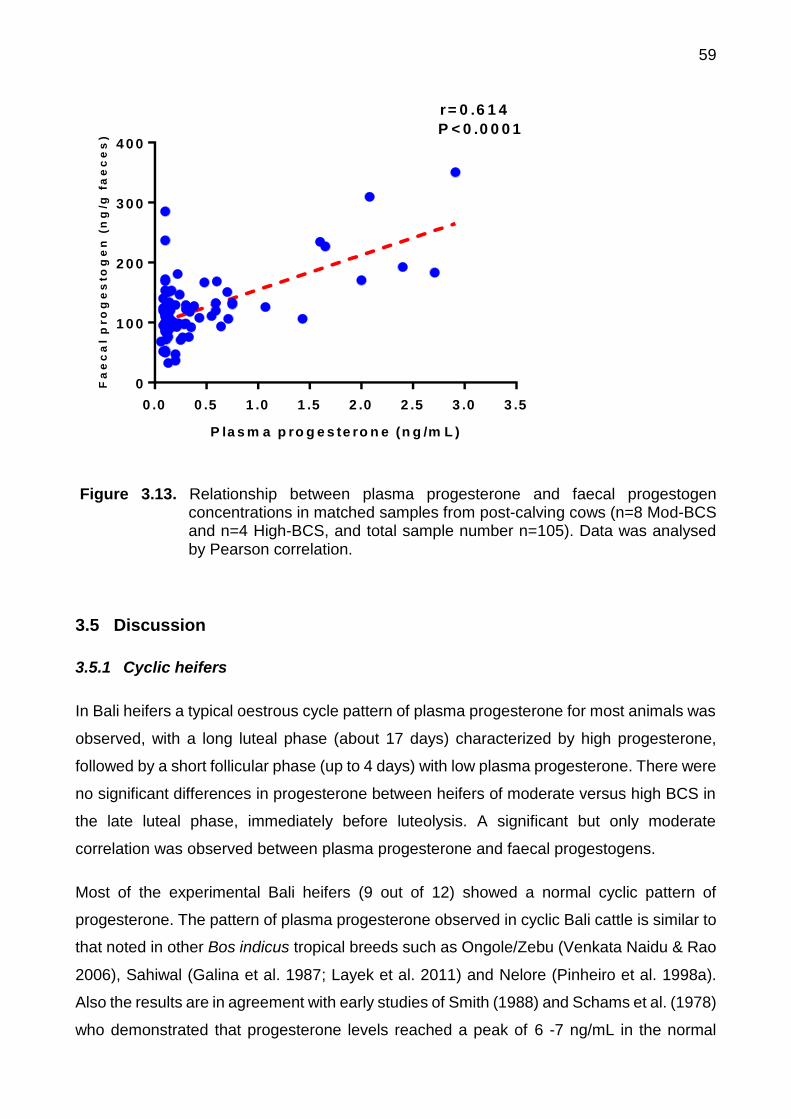
59
0 .0 0 .5 1 .0 1 .5 2 .0 2 .5 3 .0 3 .5
0
1 0 0
2 0 0
3 0 0
4 0 0
P la s m a p ro g e s te ro n e (n g /m L )
Fa
ec
al
pro
ge
sto
ge
n (
ng
/g f
ae
ce
s)
r= 0 .6 1 4
P < 0 .0 0 0 1
Figure 3.13. Relationship between plasma progesterone and faecal progestogen concentrations in matched samples from post-calving cows (n=8 Mod-BCS and n=4 High-BCS, and total sample number n=105). Data was analysed by Pearson correlation.
3.5 Discussion
3.5.1 Cyclic heifers
In Bali heifers a typical oestrous cycle pattern of plasma progesterone for most animals was
observed, with a long luteal phase (about 17 days) characterized by high progesterone,
followed by a short follicular phase (up to 4 days) with low plasma progesterone. There were
no significant differences in progesterone between heifers of moderate versus high BCS in
the late luteal phase, immediately before luteolysis. A significant but only moderate
correlation was observed between plasma progesterone and faecal progestogens.
Most of the experimental Bali heifers (9 out of 12) showed a normal cyclic pattern of
progesterone. The pattern of plasma progesterone observed in cyclic Bali cattle is similar to
that noted in other Bos indicus tropical breeds such as Ongole/Zebu (Venkata Naidu & Rao
2006), Sahiwal (Galina et al. 1987; Layek et al. 2011) and Nelore (Pinheiro et al. 1998a).
Also the results are in agreement with early studies of Smith (1988) and Schams et al. (1978)
who demonstrated that progesterone levels reached a peak of 6 -7 ng/mL in the normal

60
oestrous cycle of dairy cows, and studies by Henricks et al. (1970) who showed that higher
progesterone concentrations occur from day 10 to 14 of the oestrous cycle (with day 0 being
designated as beginning of oestrus). In grazing Bali cows, the peak progesterone
concentration of up to 5.1 ng/mL occurred on day 12 during the luteal phase (2011). In our
current study there were no differences in plasma progesterone concentrations in the late
luteal phase immediately prior to luteolysis between heifers with high and moderate BCS.
Similar findings were made by Belli and Jelantik (2011) who found no effect of concentrate
feeding upon plasma progesterone concentrations in Bali cows.
Oestrous detection measurements in this study were generally inconsistent, with only some
animals being detected in oestrus when plasma progesterone concentrations were low.
Standing oestrus is a most certain sign of oestrus and none of the heifers in this study
exhibited such behavior. Instead the majority of heifers were only sniffed and licked by a
teaser healthy bull around their vagina. Also four heifers in the current study were not
detected in oestrus and showed no oestrus at all on any experimental day. Most likely this
is due to animals exhibiting short oestrous periods, like that observed in dairy cows with
about 25% in cows having an oestrus of less than 8 hr (Duby & Prange 1996). An alternative
possibility is most oestrous behaviour occurred at night when no recordings were made. For
example in East African Zebu cattle some 60% of oestrous mountings occurred during the
night (Mattoni et al. 1988a). Further in the current study it should be noted that only a single
teaser bull was used and some variability in cow oestrous responses could be due to this
common management practice. Variability between individuals in oestrous expression is
common (Orihuela 2000), and the intensity of expression is markedly influenced by
environmental factors particularly the bull, group size and the pen surface type especially
with cows kept indoors (Diskin 2008).
A major aim of this study was to determine if faecal progestogens can be used to monitor
the oestrous cycle in Bali cattle. In general, the results demonstrated a similar pattern in
faecal progestogens as plasma progesterone, with an overall moderate significant
relationship between the measurements. Similar correlative findings in cattle have been
previously reported by Masunda et al. (1999) in tropical Nkone cows in Zimbabwe, by
Rosnina et al. (2012) in tropical Kedah Kelantan heifers and cows in Malaysia, by Hattab et
al. (2000) in buffalo cows in Egypt, and Desaulniers et al. (1989) in Holstein-Freisian dairy
cows and even wild muskoxen cows in Canada. The current results, along with these other
studies support the hypothesis that faecal sampling and measurement of faecal
progestogens is a valid technique to monitor reproductive cyclicity in cattle in a non-invasive

61
manner. However a caveat is that a single faecal sample cannot indicate the stage of the
oestrous cycle with a high degree of precision. There is a moderate degree of variability in
the results that is likely due to issues such as sub-sampling faecal material from the large
quantity of faecal material produced by cattle. Nevertheless, with repeated sampling over
time and observing patterns of faecal progestogens, this method would be informative about
the individual reproductive cyclicity of Bali heifers.
3.5.2 Post-calving cows
In post-calving Bali first-calf cows the plasma progesterone profiles were difficult to interpret
due to very low progesterone concentrations (below 1.5 ng/mL over the 100-day sampling
period) in many cows. Many plasma samples had progesterone concentrations at the assay
sensitivity of 0.1 ng/mL. In cows with higher plasma progesterone concentrations there was
no evidence of a sustained increase in progesterone with blood sampling every 10 days as
might be expected in pregnancy. Therefore it would appear that most cows in this study
were not cyclic and not pregnant.
Plasma progesterone concentrations reach a basal level around the time of parturition in
cows (Crowe 2008b), and often circulating progesterone will remain low due to a lack of
follicle development, oestrus, ovulation and formation of the CL. A continued lack of oestrus
beyond 60 days post-calving, is defined as anoestrus representing a pathological state
(Mwaanga & Janowski 2000). In the current study, since sampling was over 100-day period,
it would appear that most cows were exhibiting post-partum anoestrus. All cows in the
current study were suckling, young and this is known to be the key factor that influences the
length of post-partum anoestrus (Chenoweth 1994; Montiel & Ahuja 2005), and main factor
reducing reproductive efficiency in tropical cattle of Bos indicus and Bos taurus/Bos indicus
cows (Montiel & Ahuja 2005). Nutrient intake and energy reserves, acting via metabolic
hormones, are physiological regulators of the reproductive system (Canfield & Butler 1991).
Ovarian activity is influenced by adequate nutrition (Robinson 1990), and reduced post-
partum anoestrus and calving interval is affected by nutrition particularly during the pre-
partum period (Montiel & Ahuja 2005). Interestingly in the current study all animals calving
in moderate BCS had low plasma progesterone profiles and were likely in anoestrus. In
general these animals had inadequate nutritional intake post-calving, as indicated by their
decreasing IGF-1 profiles (Chapter 4). In contrast only 2 of the 4 high BCS animals had low
progesterone concentrations and might be considered anoestrus. The other two animals
exhibited clear oestrous cyclicity with variable (low to high) plasma progesterone

62
concentrations. One cow #225 even displayed increased progesterone over a 30 day period,
indicating either a possible abnormal extended CL function, or a pregnancy with early
embryo loss and early luteal regression. Nevetheless the predominant outcome from the
plasma progesterone profiles were that most Bali first-calf cows were anoestrus during the
sampling period (up to 124 days post-partum).
Despite minimal plasma progesterone profiles in the post-calving period a relationship
between faecal progestogen and plasma progesterone concentrations was found, and that
relationship was similar to that observed in cyclic Bali heifers. In pregnant cows strong
relationships have been previously noted between plasma progesterone and faecal
progestogens (Desaulniers et al. 1989; Miller & Holtan 1996; Masunda et al. 2002;
Kornmatitsuk et al. 2007). Such strong relationships are not unusual in later pregnancy with
increasing progesterone production by the placenta from about day 45 of pregnancy, in
addition to the continued secretion by the CL (Niswender et al. 2000).
3.6 Conclusion
In conclusion faecal collection and progestogen analysis in Bali cows can be used to monitor
the oestrous cycle and as a method for pregnancy detection, as a moderate correlation
exists between plasma progesterone and faecal progestogens. Further there were no
nutritional influences on progesterone/progestogens concentrations per se in cyclic animals,
rather only nutritional effects on post-partum cyclicity. Therefore faecal sampling and
determination of progesterone metabolites in faeces as a non-invasive technique might be
a simple and practicable tool for animal scientists and extension staff to monitor the
reproductive state of Bali cattle under field condition with low-input production and traditional
management system especially in Eastern Indonesia.

63
Chapter 4 The effect of nutrition on plasma leptin and IGF-1
concentrations in Bali first calf cows during lactation
4.1 Background
Nutrient deficiencies have been identified to have detrimental impacts on animal production,
including reproduction (Rabiee et al. 2001). Overall poor nutrition can impact on lifetime
productivity of breeder cows. If energy restriction is quite pronounced this can result in lower
milk yield, and possibly failure of lactation. More often, low level of nutrient intake is
associated with failure of cows to exhibit overt oestrous behaviour and ovulate, with cows
exhibiting a greater post-partum anoestrus. Whilst weight loss during the early post-partum
period is normal, deficient nutrient intake together with the high nutrient requirements for
lactation produce a situation where reproductive cyclicity may be compromised until the
nutrient deficiency and/or lactation ceases (Roberts 1956; Entwistle 1983).
The effect of nutrition upon reproduction is mediated via the hypothalamus (GnRH) and the
anterior pituitary (FSH and LH) gonadotrophins. Ovarian responses to LH are reduced due
to poor feeding during the post-partum period (Gombe & Hansel 1973; Martinez et al. 1984;
Rutter & Randel 1984; Whisnant et al. 1985). The lack of LH pulses reduces oestradiol
production from the developing follicle leading to less oestrous behaviour and lack of positive
feedback to induce ovulation. Sub-oestrus in post-partum cows is related to lower
concentrations of oestradiol on the day of oestrus, and difficulties in detection (Roche 2006).
Lower conception rates to AI are also another practical outcome from poor nutrition affecting
ovulation (Roche et al. 2000; Roche 2006). Beside a lack of ovulation, there is evidence to
support that poor nutrition in mammals has adverse affects on progesterone production by
the corpus luteum (Niswender et al. 1994). Negative energy balance is associated with low
levels of progesterone and might result in premature luteolysis (Rabiee et al. 2001). Such
affects are most likely mediated by the hypothalamus and pituitary gland, as reductions in
LH pulse frequency and amplitude have been reported on low energy diets (Diskin et al.
2003). Diet supplementation with protected methionine in early lactation has been shown to
enhance luteal activity (Gilmore et al. 2011).
The effects of nutrition upon metabolism and reproduction often involve two metabolic
hormones; Insulin-like growth factor-1 (IGF-1) and leptin (Strauch et al. 2003; Damptey et
al. 2013). Leptin has a predominant role in regulating energy metabolism (Williams et al.
2002), whilst IGF-1 is a major anabolic hormone often mediating the effects of growth
hormone (GH). Both leptin and IGF-1 appear to be affected by energy balance. During early

64
lactation when cows experience a negative energy balance, circulating leptin and IGF-1 are
reduced (Roberts et al. 1997; Liefers et al. 2003), with IGF-1 concentrations being very low
in cows with a restricted energy intake (Roberts et al. 1997) and IGF-1 concentrations
positively correlated with a return to ovulation in the post-partum period (Spicer et al. 1990).
In growing heifers or cows with positive energy balance, there are positive relationships
between IGF-1, insulin and BCS (Bishop et al. 1994; Vizcarra et al. 1998), and leptin with
insulin and glucose (Block et al. 2001).
To date there are no studies examining the effect of nutrition on IGF-1 and leptin in Bali
cattle during lactation. In this experiment Bali first-calf cows were examined during lactation,
after receiving diet treatments during pregnancy that produced diverse BCS at calving.
Hypothesis:
The level of nutrition influences liveweight (LW) and BCS, and these will affect circulating
concentrations of the metabolic hormones leptin and IGF-1 during lactation.
4.2 Aim of experiment
The aim of this experiment was to measure plasma leptin and IGF-1 concentrations during
the first 100 days of lactation in first calf Bali cows, given different planes of nutrition.
4.3 Materials and Methods
4.3.1 Location
The experiment was conducted between August 2014 and February 2015 at the University
of Mataram, Teaching and Research Farm, Lingsar, Nusa Tenggara Barat province in
Indonesia (8°34'20.15"S, 116°10'59.89"E).
4.3.2 Animals, diets and measurements
This study examined the same first calf Bali cows described in experiment 3b (post-calving)
in Chapter 3, with the exception of one animal (#224). Cows were all post-calving and
lactating, in either Mod-BCS (n=8) or high-BCS (n=3).
The description of feeds, feeding and measurements was described in Chapter 3 (section
3.3.2 part b). Blood samples for this experiment were collected 10 days after calving and
every 10 days thereafter until 100 days post-calving. Details of weighing and BCS are given
in Chapter 3 as these are the same animals.

65
4.3.3 Laboratory analysis
(a) Insulin-like growth factor-1 assay
The plasma concentration of IGF-1 was determined using a commercially available
immunoradiometric (IRMA) assay kit (A15729, Immunotech, Beckman Coulter Inc., Prague,
Czech Republic) according to the manufacturer’s instructions. Samples are pre-treated to
remove IGF-1 from binding proteins in plasma, with 25 µl sample being added to 500 µl of
dissociation buffer. For the assay, standards, quality controls and samples (50 µl) are added
with 300 µl tracer (125I-labelled anti-IGF-1 monoclonal antibody) to antibody coated (another
mouse monoclonal) and incubated for 60 min at room temperature. Tubes are then decanted
and washed twice with 2 mL wash solution, before being air dried and counted on a gamma
counter (Perkin Elmer Wizard 2 model 2470). Data were analysed via Assayzap software
(Biosoft; Cambridge, UK) to determine IGF-1 concentrations. The antibodies are reported
by the manufacturer to not cross react GH, insulin, proinsulin or IGF-2. The assay sensitivity
was 1.0 ng/mL, and the inter- and intra-assay coefficients of variation were 8.7% and 12.4%
for a quality control of 381.5 ng/mL.
(b) Leptin assay
The plasma concentration of leptin was determined using a commercially available multi-
species radioimmunoassay kit (XL-85K; Millipore, Darmstadt, Germany) according to the
manufacturer’s instructions. Briefly, 100 µl of sample, standards or quality control was added
to polystyrene tubes with 100 µl of assay buffer, and 100 µl of primary antibody (guinea pig
anti-leptin), vortexed and incubated overnight at 4°C. Then 100 µl of I125-labelled leptin tracer
was added, vortexed and again incubated overnight at 4°C. Lastly for separation, 1.0 mL of
precipitating reagent (goat anti-guinea pig, 3% PEG, 0.05% Triton X-100 in 0.05M
Phosphosaline 0.025M EDTA) was added, incubated for 20 min at 4°C, before centrifugation
at 3000 g for 30 min. The tubes were then carefully decanted and the remaining pellet
counted for 1 min on a gamma counter (Perkin Elmer Wizard 2 Model 2470). The data was
processed using the AssayZap software (Biosoft; Cambridge, UK). Results are expressed
in human equivalents, as the standards and tracer in this assay are human, but the assay
exhibits a broad range of cross-reactivity with many animal species, including bovine. The
antibodies are reported by the manufacturer to not cross react with insulin, proinsulin, c-
peptide or IGF-1. The sensitivity of the assay was 0.8 ng/mL, with intra- and inter-assay
coefficient of variations were 1.9% and 14.5% for a quality control of 14.6 ng/mL.

66
4.3.4 Statistical analysis
Liveweight (LW) and BCS of cows immediately prior to calving and approximately 100 days
after calving were analysed using the GLM procedure in SAS software (v9.4; SAS Institute
Inc., Cary, NC, USA) which included treatment (High-BCS, Mod-BCS) and day (between
day 10 and day 100 of lactation) and their interaction.
The concentration of IGF-1 and leptin in the plasma were analysed using a Mixed model
incorporating repeated measures in SAS software which included treatment and stage (day
of sample collection) and their interaction, with the individual animal included as the subject.
Significant differences or main effects and interactions were accepted when P < 0.05, non-
significant interactions were removed from the statistical model. In addition, polynomial and
linear relationships between the concentration of IGF-1 and leptin in plasma and the LW of
the cows measured on the day nearest to sample collection (within less than 3 days of
sample collection for each data pair) were tested using the GLM procedure in SAS software.
Results are expressed as mean ± standard error of the mean (SEM).
4.4 Results
(a) Cow liveweight and body condition score
The Mod-BCS heifers had significantly lower LW (P < 0.05; 225 ± 10 vs 275 ± 16 kg) and
lower BCS (P < 0.05; 3.1 ± 0.2 vs 4.0 ± 0.3) than the High-BCS heifers immediately prior to
calving, as expected. During the first 100 days of lactation the Mod-BCS cows had a lower
estimated ME intake than the High-BCS cows (40 vs 60 MJ ME intake/day), resulting in
cows that had a significantly lower LW (P < 0.001; 173 ± 8 vs 266 ± 12 kg) and BCS (P <
0.001; 2.6 ± 0.1 vs 4.5 ± 0.2) 100 days after calving. Both LW (P = 0.02) and BCS (P = 0.04)
of Mod-BCS cows declined significantly from immediately prior to calving to 100 days after
calving, whilst the High-BCS cows maintained LW and BCS over the same period.
(b) Concentration of IGF-1 and leptin in the plasma of lactating cows
Mean plasma concentration of IGF-1 was lower in Mod-BCS cows (59 ± 13 ng/mL)
compared with High-BCS cows (172 ± 22 ng/mL) over the entire sampling period (P < 0.01).
A significant interaction existed between cow BCS and days after calving (P < 0.05; Figure
5.1). The concentration of IGF-1 in the plasma of lactating High-BCS cows did not differ
significantly across the 100 day measurement period, whilst in lactating Mod-BCS cows
plasma IGF-1 concentration was significantly lower on days 60 to 100 after calving
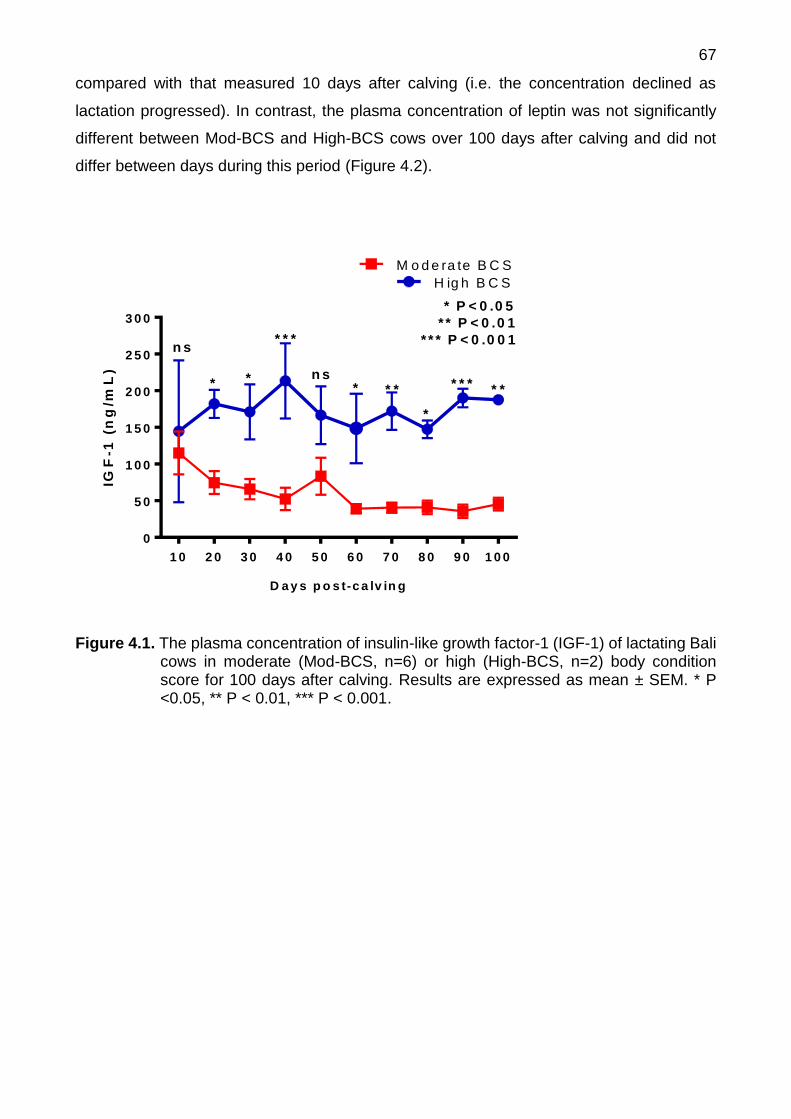
67
compared with that measured 10 days after calving (i.e. the concentration declined as
lactation progressed). In contrast, the plasma concentration of leptin was not significantly
different between Mod-BCS and High-BCS cows over 100 days after calving and did not
differ between days during this period (Figure 4.2).
D a y s p o s t-c a lv in g
IGF
-1 (
ng
/mL
)
1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 1 0 0
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
H ig h B C S
M o d e ra te B C S
* P < 0 .0 5
n s
* *
* * *
n s* * *
*
* * * * *
* * P < 0 .0 1
* * * P < 0 .0 0 1
Figure 4.1. The plasma concentration of insulin-like growth factor-1 (IGF-1) of lactating Bali cows in moderate (Mod-BCS, n=6) or high (High-BCS, n=2) body condition score for 100 days after calving. Results are expressed as mean ± SEM. * P <0.05, ** P < 0.01, *** P < 0.001.
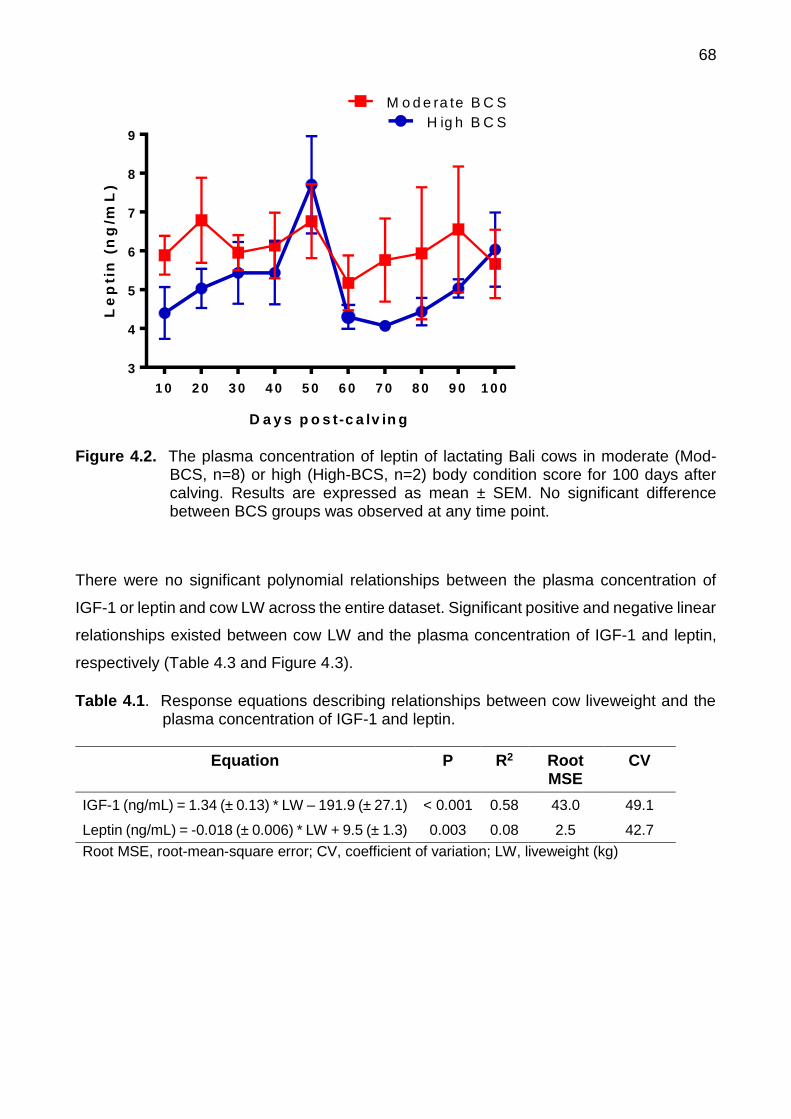
68
D a y s p o s t-c a lv in g
Le
pti
n (
ng
/mL
)
1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 1 0 0
3
4
5
6
7
8
9H ig h B C S
M o d e ra te B C S
Figure 4.2. The plasma concentration of leptin of lactating Bali cows in moderate (Mod-
BCS, n=8) or high (High-BCS, n=2) body condition score for 100 days after calving. Results are expressed as mean ± SEM. No significant difference between BCS groups was observed at any time point.
There were no significant polynomial relationships between the plasma concentration of
IGF-1 or leptin and cow LW across the entire dataset. Significant positive and negative linear
relationships existed between cow LW and the plasma concentration of IGF-1 and leptin,
respectively (Table 4.3 and Figure 4.3).
Table 4.1. Response equations describing relationships between cow liveweight and the plasma concentration of IGF-1 and leptin.
Equation P R2 Root MSE
CV
IGF-1 (ng/mL) = 1.34 (± 0.13) * LW – 191.9 (± 27.1) < 0.001 0.58 43.0 49.1
Leptin (ng/mL) = -0.018 (± 0.006) * LW + 9.5 (± 1.3) 0.003 0.08 2.5 42.7
Root MSE, root-mean-square error; CV, coefficient of variation; LW, liveweight (kg)
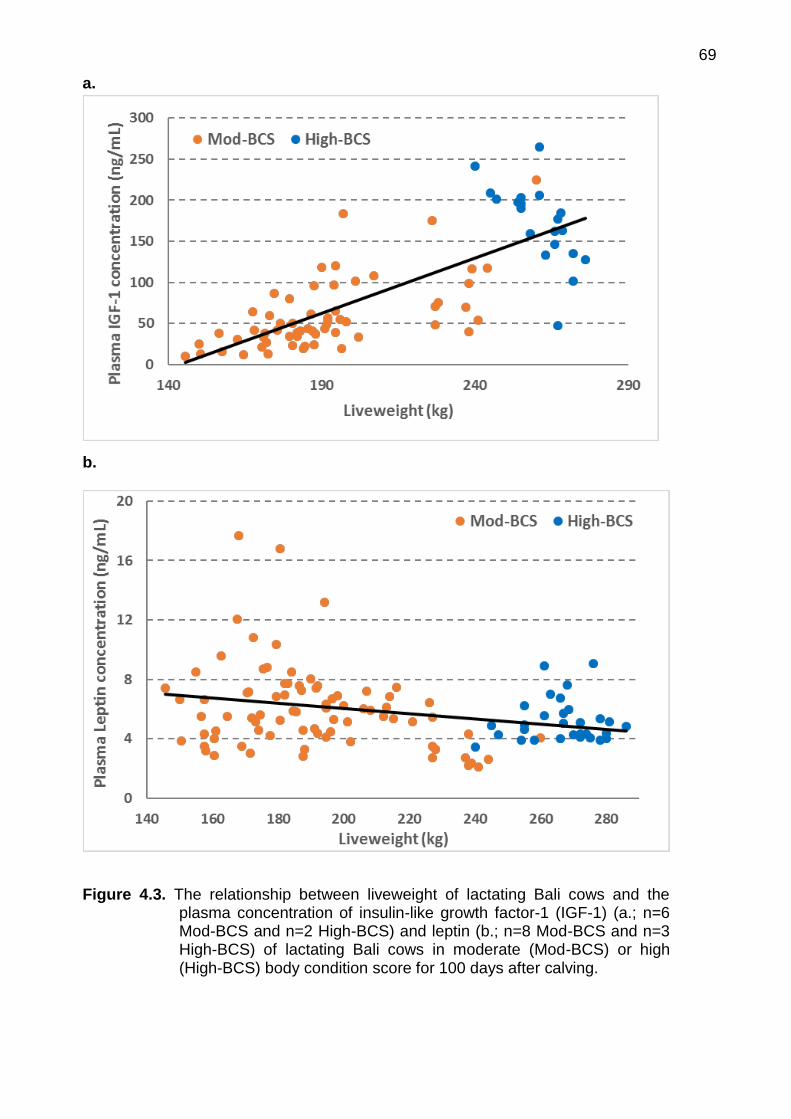
69
a.
b.
Figure 4.3. The relationship between liveweight of lactating Bali cows and the plasma concentration of insulin-like growth factor-1 (IGF-1) (a.; n=6 Mod-BCS and n=2 High-BCS) and leptin (b.; n=8 Mod-BCS and n=3 High-BCS) of lactating Bali cows in moderate (Mod-BCS) or high (High-BCS) body condition score for 100 days after calving.

70
4.5 Discussion
This study examined first calf Bali cows over the first 100 days of lactation. The main findings
were that cows calving in high BCS had significantly increased plasma IGF-1 concentrations
during early lactation compared to cows calving in moderate BCS. In contrast, there was no
difference in plasma leptin concentrations between the BCS groups. Over the 100-day
lactation period cows calving in high BCS maintained LW and BCS, whilst Mod-BCS lost
body condition losing about 20% of LW. Such changes in body condition and utilisation of
energy reserves matched changes in plasma IGF-1 concentrations, with a positive linear
relationship being noted between LW and plasma IGF-1 over the first 100 days of lactation.
Therefore it appears that IGF-1 is a good marker of nutritional status in early lactation in Bali
cows.
Plasma IGF-1 concentrations decreased in early lactation in Bali cows with Mod-BCS, but
IGF-1 remained high in High-BCS cows. At calving IGF-1 concentrations are usually
decreased (Lucy et al. 2001b; Taylor et al. 2003; Meikle et al. 2004; Adrien et al. 2012), and
often remain low throughout early lactation particularly in dairy cows (Hoshino et al. 1991).
In particular reduced BCS in early lactation has been associated with lower IGF-1
concentrations (Douglas 2015). Overall an animal’s nutritional status is one of the various
factors influencing the circulation of IGF-1 (Hoshino et al. 1991). There was a significant
positive linear relationship between cow LW and plasma IGF-1 concentration during the
experiment. As previously mentioned, metabolic hormones such as IGF-1 production might
be influenced by energy/protein intake and its specific signal in the body directly responds
to diet composition (energy/protein). During lactation in dairy cows, circulating IGF-1
concentrations may be particularly associated with its response on protein intake and/or
energy intake (Elsasser et al. 1989; Ronge & Blum 1989). This is also supported by a similar
study by Elsasser et al. (1989) who suggested that IGF-1 might be the primary nutritional
determination of protein in cattle and its response to protein may be escalated by the
availability of increasing metabolisable energy (ME) intake. Negative energy balance
induced by a change in energy partitioning and a lower BCS is associated with low IGF-1
(Veerkamp et al. 2003). In addition, suckling restriction also increased IGF-1 concentrations
in primiparous beef cows of all BCS (Soca et al. 2014), suggesting suckling itself impacts
on IGF-1.
Despite substantial differences in LW and BCS, plasma leptin concentrations of Bali cattle
in lactation were not significantly different. Unlike IGF-1, there was no change in leptin

71
concentrations during the 100-day sampling period. Such results contrast with those in dairy
cows where decreasing leptin concentrations have been noted in early post-partum period
and are thought to be a result of negative energy balance (Block et al. 2001). Indeed
circulating concentrations of leptin are related to body fat mass and the amount of available
energy (Meikle et al. 2004). Leptin acts as an important signal to regulate feed intake in
ruminants (Spicer 2001; Chelikani et al. 2003), and in turn leptin secretion can change with
feed intake (Zieba et al. 2005). In the current study only a small significant negative linear
relationship was observed between LW and plasma leptin concentrations was found in Bali
cows, with both the slope of the linear equation and the coefficient of determination (r2) being
quite low. This result contrasts with the suggestion that leptin has a major role in regulating
energy balance and optimising animal production and reproduction, including lactation
(Houseknecht et al. 1998; Barb & Kraeling 2004), however in the current study only a limited
number of animals was examined, particularly in the high BCS group (n=3). Further work
examining leptin concentrations in Bali cows with different LW and BCS would seem
warranted, particularly as De La Hoya et al. (2015) recently suggested that the amount of
body fat and leptin secretion might be different across cattle breeds.
Poor nutrient intake and lower BCS at the time of calving contributes significantly to a
prolonged calving to first oestrus interval (Montiel & Ahuja 2005). Usually in cows, when
BCS at calving is at a moderate to good level, cyclic ovarian activity will be more quickly re-
established (Richards et al. 1986), and reduce the inter-calving interval. However in the
current study, first-calf Bali cows in moderate BCS as indicated by the plasma progesterone
concentrations did not appear to cycle within the first 100 days of lactation (Chapter 3).
Nevertheless, beside BCS, nutrient intake in the early post-partum lactating period also
affects the length of post-partum anoestrus. In grazing primiparous beef cows “flushing” with
high energy intake, increased both IGF-1 and insulin concentrations and shortened the post-
partum anoestrous interval, but only in cows with moderate BCS (Soca et al. 2014). In the
current study it was observed that moderate BCS cows had declining IGF-1 concentrations
in early lactation, indicating that nutrient intake was insufficient for metabolic demands
during lactation. Overall it is likely that both post-partum nutrient intake and availability of
body energy reserves together contribute to a prolonged post-partum anoestrous interval
(Wettemann et al. 2003). IGF-1 is a key metabolic hormone that affects ovarian function,
even via direct action on the corpus luteum (Einspanier et al. 1990; Sauerwein et al. 1992).
Moreover, as a monitoring signal, IGF-1 allows reproductive events to occur when nutritional
conditions for successful reproduction are reached (Velazquez et al. 2008). In contrast, in

72
the current study cows calving in high BCS sustained LW in the post-calving period, and
maintained IGF-1 concentrations, albeit only half of these cows exhibited oestrous cycles
(Chapter 3). This result highlights the significant influence of suckling on inhibition on the
hypothalamus-pituitary-ovarian axis (Wettemann et al. 2003). As highlighted in Chapter 3
suckling is the main factor reducing reproductive efficiency in tropical cattle of Bos indicus
and Bos taurus/Bos indicus cows (Montiel & Ahuja 2005).
4.6 Conclusion
In lactation first calf Bali cows BCS at the time of calving and nutrition in the post-partum
period affected plasma concentrations of the metabolic hormone IGF-1, but not leptin. Cows
calving in high BCS and fed high energy diets maintained IGF-1 concentrations in lactation
and this was associated with half of these animals exhibited oestrous cycles within the first
100 days post-partum. In contrast cows in moderate BCS, that lost about 20% LW during
the first 100 days of lactation, had significantly lower IGF-1 concentrations, and all appeared
to be anoestrus. Overall there was a positive linear relationship between LW and IGF-1
concentrations in the post-partum period that suggests IGF-1 is a good marker of nutritional
status in early lactation in Bali cows.

73
Chapter 5 Investigation of measurement of Vaginal Electrical
Conductivity to determine stage of the oestrous cycle in Bali, Ongole
and Madura cattle.
5.1 Background
The success of AI is dependent on accurate detection of oestrus (Canfield & Butler 1989;
Rorie et al. 2002). Where AI is used, improvements in accuracy of oestrous detection should
result in increased submission rate (proportion of cows inseminated or mated) and
conception rate (proportion of inseminations or mating that results in pregnancy). This is
especially important for cattle breeding herds that want to improve their reproductive
management and rate of genetic improvement through use of AI (Heersche & Nebel 1994).
Also, herd profitability can be increased through improvements in herd reproductive
performance (Peralta et al. 2005).
Standing to be mounted is the most commonly used means of detecting oestrus in cattle.
However, where cattle are tethered or have restricted space to move other signs of oestrus
such as mucus discharge from the vulva, swelling and pronounced wrinkling of the vulva,
increased bellowing and restlessness, are used. Measurement of vaginal electrical
conductivity (VEC) has been reported to be a useful method for detecting oestrus in dairy
and beef cattle (Rorie et al. 2002). Further, Wehner et al. (1997) reported that repeated
measurement of VEC during the oestrous period of Gelbvieh and Angus cows using the
probe used in this study (OvatecR) could be used to determine time of AI to produce primarily
male or female calves. When cows were inseminated in early oestrus when VEC measured
35 to 45 on the scale, 92.9% of calves born were female, and when cows were inseminated
in late oestrus when VEC had increased to between 50 to 70 on the scale, 91.7% of calves
born were male. Presumptive oestrus and dioestrus were determined with a high degree of
accuracy with a single measurement of VEC using the Ovatec® probe in Bos indicus cross
heifers that had been treated to synchronise oestrus (Hockey et al. 2010). In anoestrous
females the VEC is similar to that observed in dioestrous females (Gupta & Purohit 2001).
A hypothesis of this experiment was VEC can detect oestrus and early pregnancy. The aim
of this experiment was to determine whether measurement of VEC could be used to
diagnose oestrus, dioestrus and/or early pregnancy in three tropically adapted breeds of
beef cattle (Bali, Ongole and Madura) commonly found in Indonesia.

74
5.2 Materials and Methods
5.2.1 Location and time of experiments
This experiment was conducted at the Indonesian Beef Cattle Research Station (IBCRS) in
Grati, Pasuruan East Java, Indonesia. The experiment was carried out during summer in
July and August 2015. IBCRS is located in eastern Indonesia; latitude 7°30‟S and longitude
113°30‟E with the average minimum and maximum temperatures between rainy and dry
seasons of 23 to 30°C, and 25 to 32°C, respectively and its humidity was 50 to 90% and so
variation was minimal between seasons.
Between December and April is the rainy season with the wettest months normally January
and February. The dry season is normally from May to October. The average annual daily
minimum and maximum temperatures are 21 and 36°C.
5.2.2 Animals and animal management
At the commencement of the experiment (Day 0 to 3) liveweight, BCS and general health
were assessed and all heifers and cows were confirmed to be non-pregnant and had normal
reproductive tracts by transrectal ultrasonography of the reproductive tract. Time of weighing
was between 10 am until midday and each group of cattle consisting of 20 cattle was
weighed daily for 3 sequential days (Ongole, Bali and Madura, respectively). VEC
measurements were conducted at the same days for weighing. Type of scales was TRU-
TEST EW5 with MP 600 (600 mm and 23”) and 1,500 kg live capacity and 2,000 kg total
capacity. Ultrasonography was done using a B-Mode real-time Sonosite M-turbo, Fujifilm
ultrasound scanner with L52X/ 10 to 5 MHz linear array transrectal transducer (Sonosite
Inc., Bothel, WA, USA). A single breed group was examined each day and subsequently
each breed group was examined in the same order on individual days according to the
experimental design. Twenty non-lactating Ongole (Bos indicus) cows aged 3 to 4 years
old (mean (and SEM) weight 239 ± 37.04 kg, mean (and SEM) BCS 3 ± 0.25), 20 four-year-
old Bali (Bos javanicus) heifers (mean (and SEM) weight 179 ± 10.68 kg, mean (and SEM)
BCS 3 ± 0.26) and 20 four-year-old Madura (Bos indicus x Bos javanicus) heifers (mean
(and SEM) weight 193 ± 27.34 kg, mean (and SEM) BCS 3 ± 0.28) were selected for this
experiment. All females were treated with kalbazen (Kalbazen®-SG-Albendazole 19 mg/mL,
PT. Kalbe Farma, Bekasi, Indonesia) orally to eliminate parasites prior to the experiment.
The cattle were held in three open-sided, covered pens with one breed per pen, and
remained in the same pen throughout the study. The cattle were offered the same diet which

75
consisted of 5.7 kg (as fed) concentrate (32% rice bran, 23% palm kernel cake, 20% cassava
flour, 10% corn waste, 7% potato flour, 7% molasses and 1% of limestone) per head per
day, 8.5 kg (as fed) elephant grass (Pennisetum purpurpeum) per head per day and rice
straw ad libitum. The concentrate was offered as a single portion at 0800 h and the elephant
grass was offered in two equal portions at 0800 and 1200 h. Drinking water was available
at all times.
All females were treated with prostaglandin/PGF2α (dinoprost tromethamine 25 mg/injection;
Day 0-3 and Day 14-17) to synchronise oestrus. To assist with oestrous detection a 15 to
20 cm strip of blue tail paint was applied to the tail head of all females at the time of the
second PGF2α treatment, and a bull of the same breed as the females was then placed with
each pen of females. All females again underwent transrectal ultrasonography of the
reproductive tract on Days 42-44 of the study.
(a) VEC Measurement Procedure
Individual females were restrained in a race as shown in Figure 6.1 to enable measurement
of VEC. VEC was measured using the OvatecR probe (Probe Diagnostics Inc) on Day 0-3
and then again at Day 42-44. It was expected that at each day of examination there would
be cattle at different stages of the oestrous cycle and at the second examination potentially
some would be in early pregnancy. Before each measurement session the Ovatec machine
was calibrated by inserting the probe into 0.9% sodium chloride solution and then checking
the reading was less than 100. Between females the probe was rinsed in an antiseptic
solution containing dilute povidone iodine solution. Prior to insertion of the probe in the
vagina the vulva was wiped clean with a moist disposable cloth. After applying a small
amount of Ultrasonic Gel (OneMed) to the tip of the probe it was inserted into the anterior
vagina until it was against the vaginal fornix/cervix. Then slight downward pressure was
applied to the probe and the VEC measurement was taken.

76
Figure 5.1. Vaginal electrical conductivity (VEC) measurement procedure.
(b) Blood sample collection and progesterone assay
Blood samples were collected by jugular venepuncture from each female immediately prior
to measurment of VEC. Blood was collected into a 10 mL lithium heparin coated vacutainer
which was then slowly inverted 6 to 8 times and placed on ice prior to centrifugation at 3500
g for 10 minute at 4°C. All plasma samples were then stored at -20°C until analysis for
progesterone. Samples were couriered from Indonesian to Australia frozen under dry ice
and were exposed to 50 kGy gamma irradiation prior to release from quarantine. This
dosage of gamma irradiation had no effect on the measurement of the concentration of IGF-
1, leptin and progesterone in the plasma of cattle (Simon Quigley, pers. comm.). A full
description of the progesterone assay is also provided in Chapter 3 section 3.3.4.
5.3 Results
Scatter plots of VEC versus plasma progesterone concentration are presented in Figures
5.2 and 5.3.
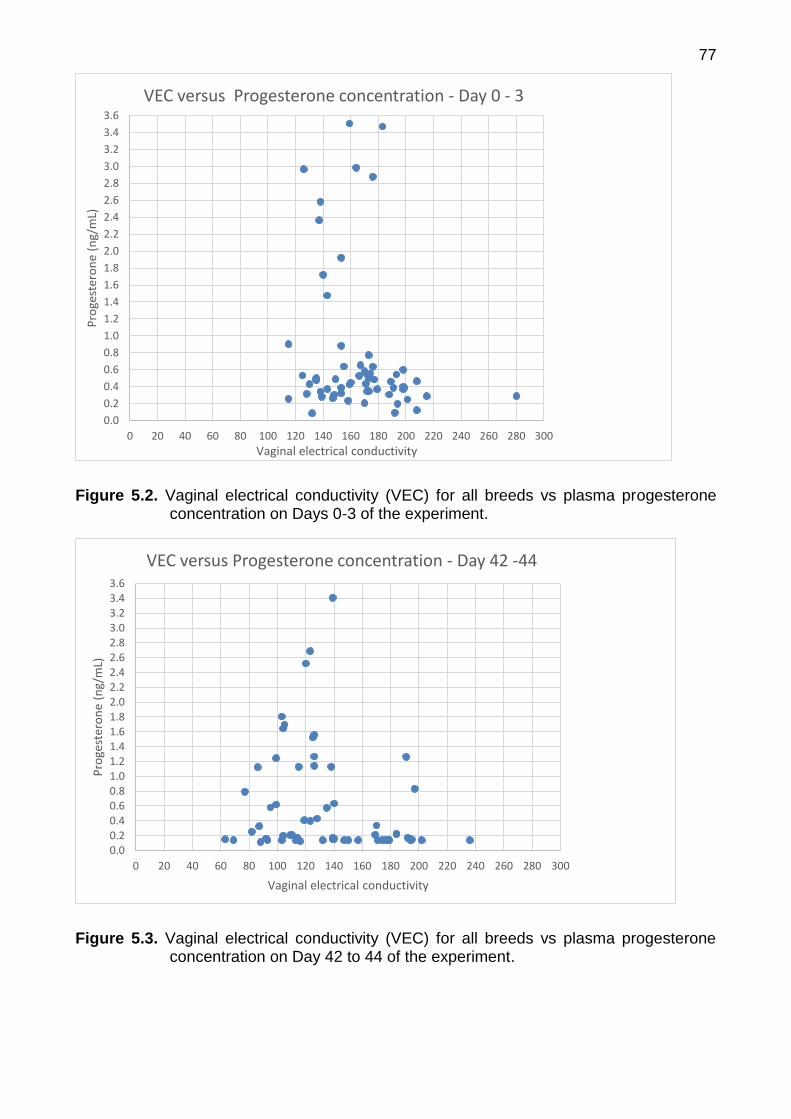
77
Figure 5.2. Vaginal electrical conductivity (VEC) for all breeds vs plasma progesterone concentration on Days 0-3 of the experiment.
Figure 5.3. Vaginal electrical conductivity (VEC) for all breeds vs plasma progesterone concentration on Day 42 to 44 of the experiment.
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
3.0
3.2
3.4
3.6
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Pro
gest
ero
ne
(ng/
mL)
Vaginal electrical conductivity
VEC versus Progesterone concentration - Day 0 - 3
0.00.20.40.60.81.01.21.41.61.82.02.22.42.62.83.03.23.43.6
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Pro
gest
ero
ne
(ng/
mL)
Vaginal electrical conductivity
VEC versus Progesterone concentration - Day 42 -44

78
On initial inspection of the data it appeared that there was a poor relationship between VEC
and plasma progesterone concentration. However, this assumes that all the progesterone
concentrations less than 0.5 ng/mL were from females in oestrus; these female should have
had VEC measurements of less than or equal to 90 (Hockey et al. 2010). Alternatively, the
low progesterone concentrations could have been due to the fact that some females were
in a state of anoestrus. To investigate this further the ultrasonographic ovarian findings and
oestrous detection /mating data as well as the progesterone data for each individual female
at Day 0-3 and day 42-44 were examined and cycling status and stage of oestrous cycle
determined (Table 5.1). The stage of the oestrous cycle was estimated as follows:
females not seen in oestrus throughout the study and for up to 3 months afterwards,
and which had progesterone concentrations less than 0.5 ng/mL were classified as
being anoestrus
females with progesterone concentrations of 1 ng/mL or greater were classified to
be in dioestrus
females which at the time of measurement of VEC ultrasonographically had a CL on
one ovary and had progesterone concentration less than 1 ng/mL were classified to
be in metoestrus, and
females which were observed in oestrus, or had a CL at the time of first VEC
measurement and then subsequently were not seen on heat and had progesterone
concentrations less than 0.5 ng/mL at the 2nd VEC measurement were classified as
having become anoestrus
On Day 0 – 3 of the study 50%, 40%, and 10% of the Ongole, Bali and Madura cattle were
apparently in a state of anoestrum. This is consistent with the observed proportion of
females mated Day 21 – 24. Within 7 days after the second PGF2α treatment, 30%, 15%
and 10% of the Madura, Bali and Ongole cattle, respectively, were observed to have been
mated, and 66%, 0%, 0%, of the mated Madura, Bali and Ongole female subsequently
became pregnant. At both VEC measurements the majority of females were either in
anoestrus or dioestrus with very few estimated to be in oestrus. The high proportion of
females which were determined to be anoestrus at the time of VEC measurement in this
study contributed to the apparent poor relationship between plasma progesterone
concentration and VEC measurement. The mean VEC measurements and progesterone
concentrations by estimated stage of the oestrous cycle/cycling status are summarized in
Table 5.2.
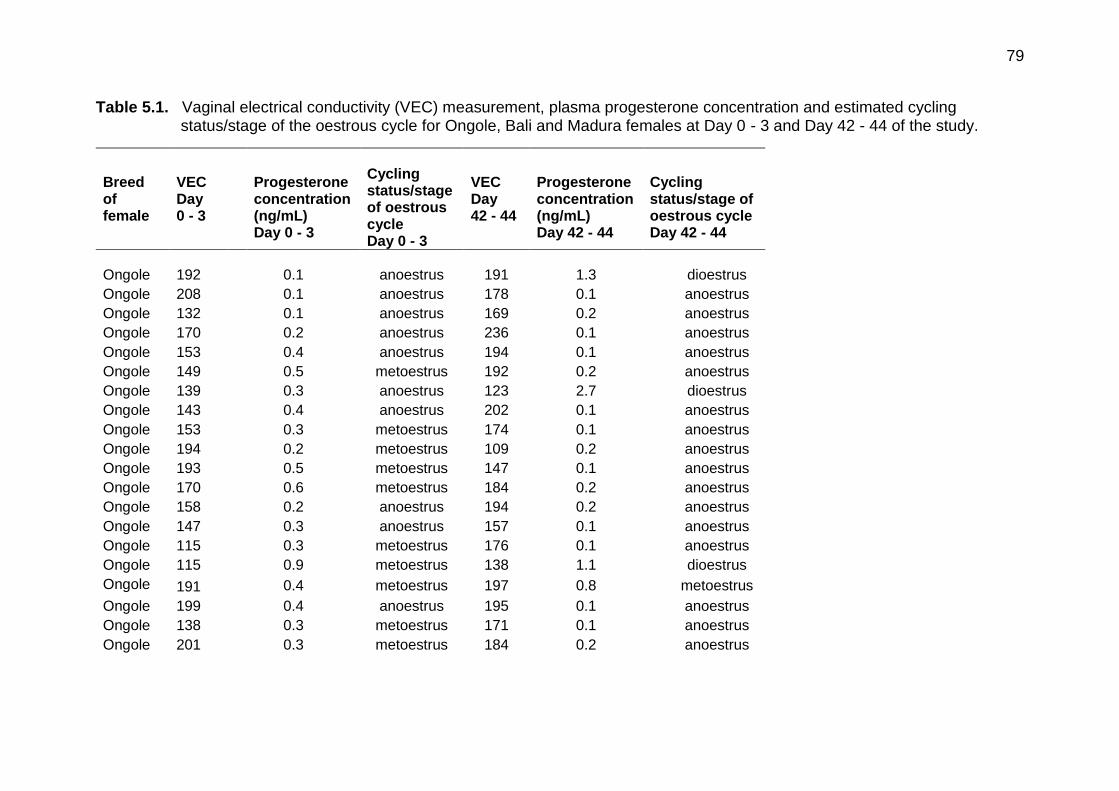
79
Table 5.1. Vaginal electrical conductivity (VEC) measurement, plasma progesterone concentration and estimated cycling status/stage of the oestrous cycle for Ongole, Bali and Madura females at Day 0 - 3 and Day 42 - 44 of the study.
Breed of female
VEC Day 0 - 3
Progesterone concentration (ng/mL) Day 0 - 3
Cycling status/stage of oestrous cycle Day 0 - 3
VEC Day 42 - 44
Progesterone concentration (ng/mL) Day 42 - 44
Cycling status/stage of oestrous cycle Day 42 - 44
Ongole 192 0.1 anoestrus 191 1.3 dioestrus
Ongole 208 0.1 anoestrus 178 0.1 anoestrus
Ongole 132 0.1 anoestrus 169 0.2 anoestrus
Ongole 170 0.2 anoestrus 236 0.1 anoestrus
Ongole 153 0.4 anoestrus 194 0.1 anoestrus
Ongole 149 0.5 metoestrus 192 0.2 anoestrus
Ongole 139 0.3 anoestrus 123 2.7 dioestrus
Ongole 143 0.4 anoestrus 202 0.1 anoestrus
Ongole 153 0.3 metoestrus 174 0.1 anoestrus
Ongole 194 0.2 metoestrus 109 0.2 anoestrus
Ongole 193 0.5 metoestrus 147 0.1 anoestrus
Ongole 170 0.6 metoestrus 184 0.2 anoestrus
Ongole 158 0.2 anoestrus 194 0.2 anoestrus
Ongole 147 0.3 anoestrus 157 0.1 anoestrus
Ongole 115 0.3 metoestrus 176 0.1 anoestrus
Ongole 115 0.9 metoestrus 138 1.1 dioestrus
Ongole 191 0.4 metoestrus 197 0.8 metoestrus
Ongole 199 0.4 anoestrus 195 0.1 anoestrus
Ongole 138 0.3 metoestrus 171 0.1 anoestrus
Ongole 201 0.3 metoestrus 184 0.2 anoestrus
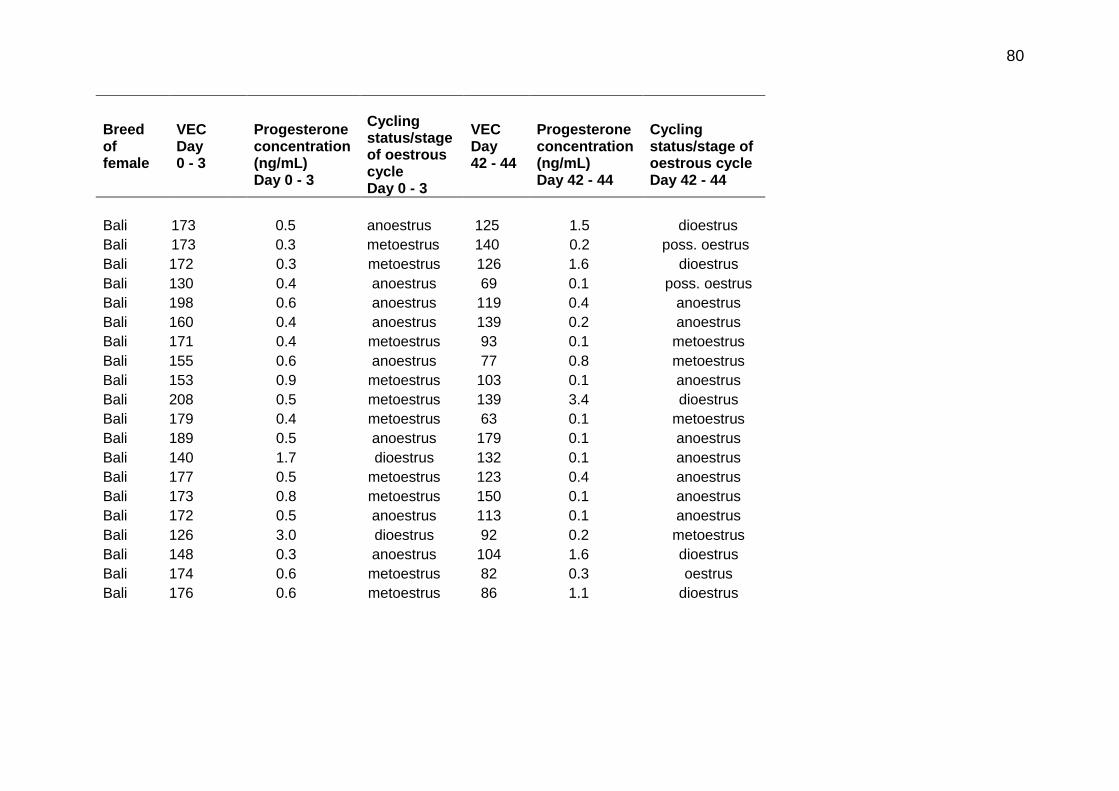
80
Breed of female
VEC Day 0 - 3
Progesterone concentration (ng/mL) Day 0 - 3
Cycling status/stage of oestrous cycle Day 0 - 3
VEC Day 42 - 44
Progesterone concentration (ng/mL) Day 42 - 44
Cycling status/stage of oestrous cycle Day 42 - 44
Bali
Bali
173
173
0.5
0.3
anoestrus
metoestrus
125
140
1.5
0.2
dioestrus
poss. oestrus
Bali 172 0.3 metoestrus 126 1.6 dioestrus
Bali 130 0.4 anoestrus 69 0.1 poss. oestrus
Bali 198 0.6 anoestrus 119 0.4 anoestrus
Bali 160 0.4 anoestrus 139 0.2 anoestrus
Bali 171 0.4 metoestrus 93 0.1 metoestrus
Bali 155 0.6 anoestrus 77 0.8 metoestrus
Bali 153 0.9 metoestrus 103 0.1 anoestrus
Bali 208 0.5 metoestrus 139 3.4 dioestrus
Bali 179 0.4 metoestrus 63 0.1 metoestrus
Bali 189 0.5 anoestrus 179 0.1 anoestrus
Bali 140 1.7 dioestrus 132 0.1 anoestrus
Bali 177 0.5 metoestrus 123 0.4 anoestrus
Bali 173 0.8 metoestrus 150 0.1 anoestrus
Bali 172 0.5 anoestrus 113 0.1 anoestrus
Bali 126 3.0 dioestrus 92 0.2 metoestrus
Bali 148 0.3 anoestrus 104 1.6 dioestrus
Bali 174 0.6 metoestrus 82 0.3 oestrus
Bali 176 0.6 metoestrus 86 1.1 dioestrus
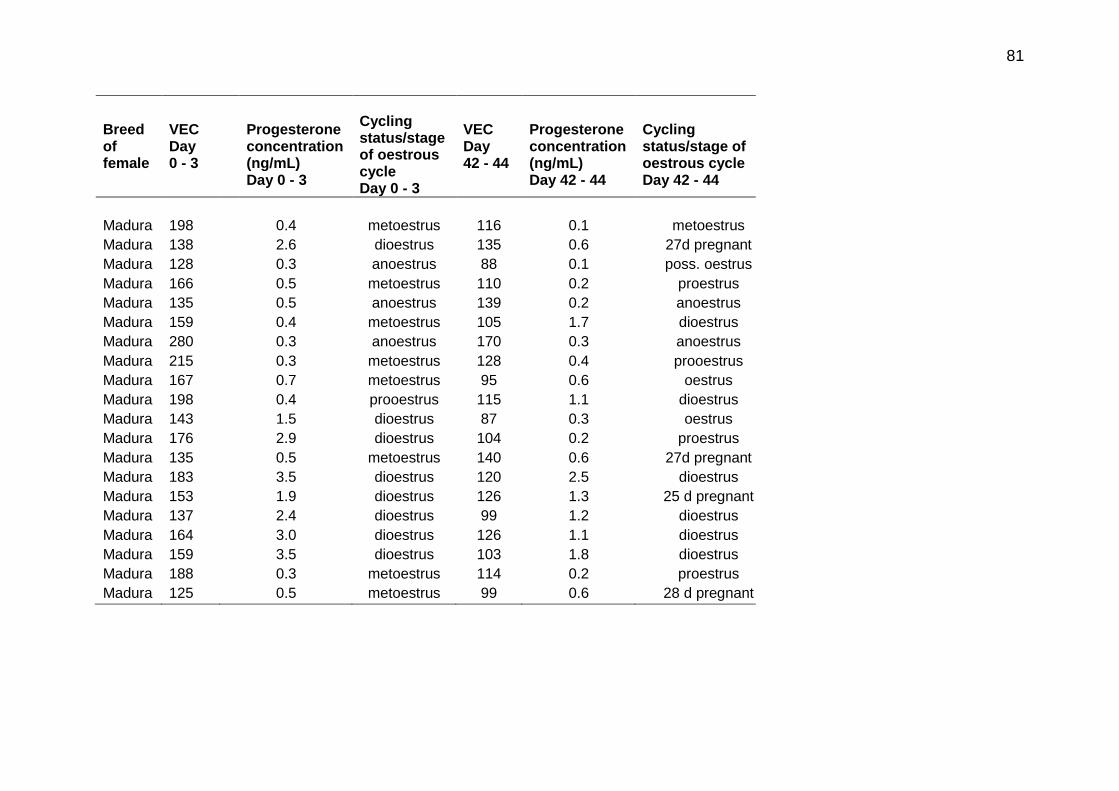
81
Breed of female
VEC Day 0 - 3
Progesterone concentration (ng/mL) Day 0 - 3
Cycling status/stage of oestrous cycle Day 0 - 3
VEC Day 42 - 44
Progesterone concentration (ng/mL) Day 42 - 44
Cycling status/stage of oestrous cycle Day 42 - 44
Madura
Madura
198
138
0.4
2.6
metoestrus
dioestrus
116
135
0.1
0.6
metoestrus
27d pregnant
Madura
Madura
128
166
0.3
0.5
anoestrus
metoestrus
88
110
0.1
0.2
poss. oestrus
proestrus
Madura 135 0.5 anoestrus 139 0.2 anoestrus
Madura 159 0.4 metoestrus 105 1.7 dioestrus
Madura 280 0.3 anoestrus 170 0.3 anoestrus
Madura 215 0.3 metoestrus 128 0.4 prooestrus
Madura 167 0.7 metoestrus 95 0.6 oestrus
Madura 198 0.4 prooestrus 115 1.1 dioestrus
Madura 143 1.5 dioestrus 87 0.3 oestrus
Madura 176 2.9 dioestrus 104 0.2 proestrus
Madura 135 0.5 metoestrus 140 0.6 27d pregnant
Madura 183 3.5 dioestrus 120 2.5 dioestrus
Madura 153 1.9 dioestrus 126 1.3 25 d pregnant
Madura 137 2.4 dioestrus 99 1.2 dioestrus
Madura 164 3.0 dioestrus 126 1.1 dioestrus
Madura 159 3.5 dioestrus 103 1.8 dioestrus
Madura 188 0.3 metoestrus 114 0.2 proestrus
Madura 125 0.5 metoestrus 99 0.6 28 d pregnant
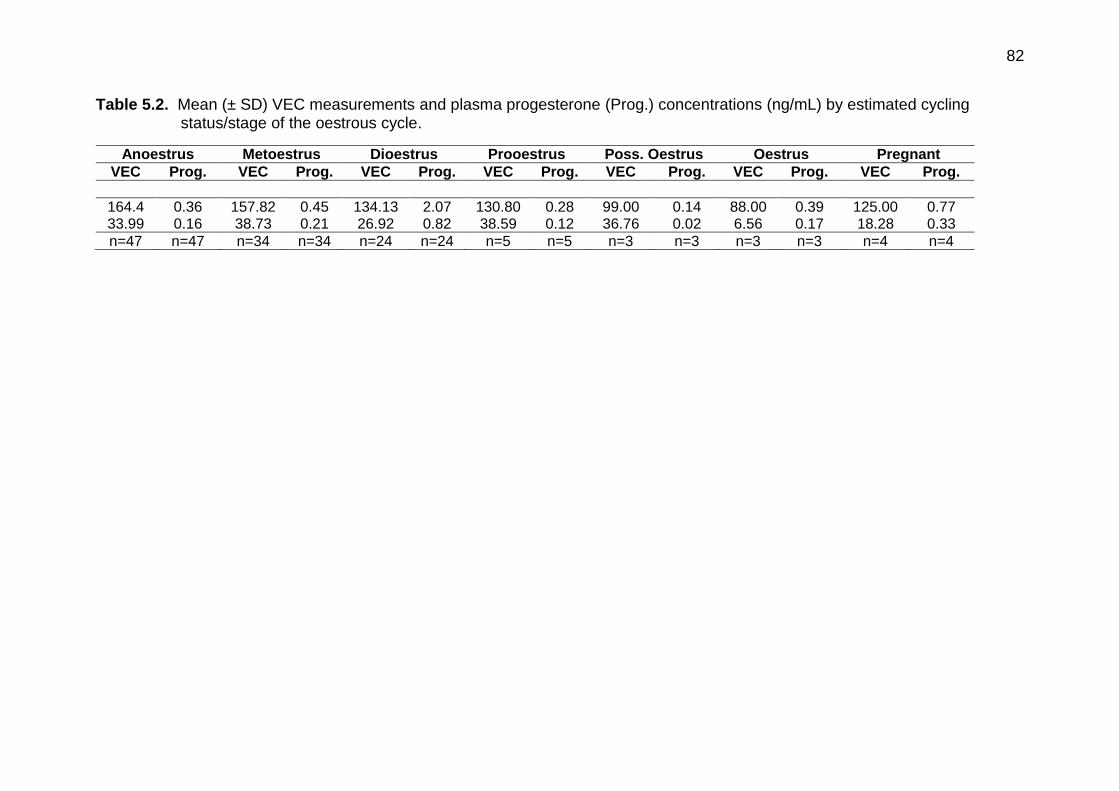
82
Table 5.2. Mean (± SD) VEC measurements and plasma progesterone (Prog.) concentrations (ng/mL) by estimated cycling status/stage of the oestrous cycle.
Anoestrus Metoestrus Dioestrus Prooestrus Poss. Oestrus Oestrus Pregnant
VEC Prog. VEC Prog. VEC Prog. VEC Prog. VEC Prog. VEC Prog. VEC Prog.
164.4 0.36 157.82 0.45 134.13 2.07 130.80 0.28 99.00 0.14 88.00 0.39 125.00 0.77 33.99 0.16 38.73 0.21 26.92 0.82 38.59 0.12 36.76 0.02 6.56 0.17 18.28 0.33
n=47 n=47 n=34 n=34 n=24 n=24 n=5 n=5 n=3 n=3 n=3 n=3 n=4 n=4

83
5.4 Discussion
Initial comparison between VEC measurements and plasma progesterone
concentrations indicated that there was a poor correlation, however this was due to
the fact that a very high proportion of cattle with low progesterone concentrations were
in a state of anoestrus. Unfortunately in this study there were only a small number of
cows considered to be in oestrus on the day of VEC measurements. This study
highlights the critical importance of careful assignment of cattle according to their
reproductive status (i.e stage of the oestrous cycle and cycling status) when
interpreting VEC measurements.
Individual VEC measurements reflect the electrical resistance of the vaginal and
cervical secretions, and are typically lowest when females are in oestrus due to the
oestrogen induced changes in the type and/or quantity of glycoproteins or electrolytes
(Hockey et al. 2010). The concentration of oestrogen during oestrus is increasing
(Hockey et al. 2010) and this induces oedema of the vaginal interstitium and affects
electrolyte concentrations of the vaginal luminal fluid (Aboulela et al. 1983; Ezov et al.
1990) resulting a reduction in the electrical resistance within the vagina of cattle (Lewis
et al. 1989; Smith et al. 1989). Schams et al. (1977) first reported that oestrogen
secretion immediately prior to oestrus decreased the electrical resistance of vaginal
fluids resulting in increased electrical conductivity. Following the preovulatory surge of
luteinizing hormone, oestrogen secretion decreases markedly and VEC
measurements increase rapidly (Rorie et al. 2002) as the corpus luteum forms and
progesterone secretion increases. Thus during metoestrus and dioestrus VEC
measurements are typically high. However, in anoestrous cattle, electrical resistance
of the reproductive tract secretions are also high due to the absence of any significant
follicular growth and hence secretion of oestrogen; thus VEC measurements taken
from these animals are also high. Most publications on use of VEC measurements
focus on use of this technology to detect oestrus and there are only limited reports of
VEC measurements in anoestrous cattle. Gupta and Purohit (2001) reported VEC
measurements in anoestrous buffalo very similar to those for animals in dioestrus.
Further, McCaughey and Patterson (1981) observed that the VEC measurements of
anoestrous dairy cows were very similar to those recorded for cows in the luteal phase
of the oestrous cycle.

84
Examination of the VEC data presented in Table 5.1 also highlights that although in
most cases a low VEC measurement is indicative that the animal is likely to be in
oestrus this is not always the case. Gartland et al. (1976) reported large within and
between animal variation in VEC measurements during oestrus. Some of this variation
may be due to the probe insertion depth in the vagina (Aboulela et al. 1983); the probe
position within the vagina (Foote et al. 1979; Heckman et al. 1979); degree of pressure
against the mucus membrane and pathological conditions of the reproductive tract
(Leidl & Stolla 1976). Further, it is acknowledged that application of a lubricant to the
probe to facilitate insertion in the vagina in this study may have also had some effect
on VEC measurements, however the lubricant was applied for all measurements
recorded in this study.
Some of the heifers and cows in this study were apparently cycling and mated or seen
on heat after the 2nd PGF injection, however they subsequently failed to return to
oestrus, were found to have low plasma progesterone concentrations and were not
detectably pregnant and had high VEC measurements. Further, as part of another
study these cattle were monitored for a further 4 months by weekly transrectal
ultrasonography of the reproductive tract. Based on all of this data the conclusion was
that these cattle had become anoestrus after the 2nd PGF injection. Regular handling
of cattle to conduct procedures such as jugular venipuncture and transrectal
ultrasonography is likely to be stressful for some animals. Cortisol secretion in
response to this stress can suppress GnRH release and hence animals do not have a
normal preovulatory release of LH (Dobson & Smith 2000). As a consequence
ovulation fails to occur or is delayed and cattle are at risk of becoming anoestrus,
particularly if the stressors which initiated this response continue.
5.5 Conclusion
This study supports the findings from previous studies of the use of measurement of
VEC in the reproductive management of cattle. Detection of low VEC measurements
(<90) indicate that the female is likely to be in oestrus but wherever possible this needs
to be supported by observed signs of oestrus and history of hormonal treatments to
synchronise oestrus/ovulation. However interpretation of high values is problematic as
anoestrous and dioestrous females have similar VEC measurements. Single
measurements of VEC to determine pregnancy status, conducted at the time when

85
cattle are expected to return to oestrus, is likely in some cases (as demonstrated in
this study) to be quite inaccurate. The use of this technology for pregnancy diagnosis
in small holder farms in Indonesia is not recommended. Overall it is concluded that
measurement of VEC is likely to be most useful in situations where there is some
uncertainty about whether a heifer or cow is in oestrus.

86
Chapter 6 Milk production in Bali cows and heifers in a village
system
6.1 Background
Milk production in Bali cattle has not been widely reported (Sariubang et al. 2000).
Milk production ranged from 0.8-1.2 kg/d under controlled research station conditions
in South Sulawesi (Sariubang et al. 2000). The mean values for 3 provinces collated
by Talib et al. (2003) was 1.1 kg/d. Cows within a village system can vary widely in
Body Condition Score (BCS) and nutrition. BCS has an effect on milk production in
dairy cows with higher BCS cows having body energy reserves which can be used to
sustain milk production in times of nutritional limitation (Montiel & Ahuja 2005). As a
management tool, BCS has proved to be useful to assess nutritional status of dairy
cows (López-Gatius et al. 2003).The effect of BCS on milk production and calving in
dairy cows has been quantified by Berry et al. (2007).
Some beef breed types, eg Bos indicus genotypes, show lactation anoestrus (post-
partum anoestrus (PPA)) as a result of suckling and hormonal influences on oestrous
activity (Mukasa-Mugerwa 1989; Short et al. 1990; Williams 1990; Fitzpatrick 1994;
Montiel & Ahuja 2005) leading to an occurrence of uterine involution (Maree et al. 1974;
Senger 2003). It was also clearly indicated in the study of genetics of early and lifetime
annual reproductive performance on Brahman and tropical composites cows in
northern Australia that reproductive performances from the second mating were
largely influenced by lactation anoestrus (Johnston et al. 2014). The days to first
ovulation after parturition has not been well described in Bali cattle and earlier chapters
(2, 3 and 4) outlined some of these features. In this experiment, cows from the hamlet
of Karang Kendal (North Lombok) were monitored under controlled field conditions to
measure milk production over twelve weeks of lactation. Time to resume oestrus after
parturition was also recorded.
This study hypotheses were (1) Milk production in Bali cows and heifers is low, and (2)
Nutritional status as reflected in body condition score affects the milk production.

87
6.2 Aim of experiment
The aims of this experiment were (1) to quantify milk production in Bali cows under
different conditions and (2) to examine the relationship between milk production and
Post-Partum Anoestrus (PPA).
6.3 Materials and methods
Measurements were made over 12 weeks of lactation post-parturition with the original
aim to choose two groups of BCS (moderate (<2.5 from range of 5) and high (>3.5
from range of 5) (Teleni et al. 1993) to observe the relationship between BCS, milk
production and time to first oestrus. However there were insufficient numbers within
these ranges and a total of nine animals (5 cows and 4 heifers) were chosen in Karang
Kendal hamlet North Lombok West Nusa Tenggara Indonesia. It was unfortunate that
more animals could not enter the study and these numbers limited the outcomes and
interpretation of the experiment. The mean BCS of the heifers was 2.48 and the cows
was 2.54 with a range of 2.0-3.0 for the whole group. Animals were offered rice straw
(approximately 5 kg per day) with addition of Elephant grass (Pennisetum purpureum),
native grass), Sesbania (Sesbania grandiflora), Gliricidia (Gliricidia sepium)) and
peanut straws available) both prior to and during the study. The estimated CP
contents from the Indonesian data base is 4.2%, 9.1%, 6.7%, 30.1%. 22.7% and
13.8%, respectively. Feed was provided two or three times a day around 6.30 – 8.00
am, 12 – 2 pm and 4.30 – 6.00 pm. Milk production was measured by the weigh-
suckle-weigh method at 4, 8 and 12 weeks post-calving (starting in July 2014).
Milk production was measured starting from 10 days after parturition at 4, 8 and 12
weeks (starting from July 2014) using the weigh-suckle-weigh method as follows:
• This was conducted at 4, 8 and 12 weeks after calving. Calves were
removed from the cows and housed in a separate pen at approximately
18.00 h the day prior to weigh-suckle-weigh.
• At 08.00 h the following morning the calf was returned to the cow and
allowed to suckle to satiety.
• The calf was then removed from the cow for 6 h.

88
• The calf was then weighed, returned to the cow and allowed to suckle to
satiety and reweighed. The difference in calf weight = milk production
over that time period of 6 h.
• This milk production was converted to a 24 h value by multiplying by 4.
Calves were observed during the weigh-suckle-weigh activity for any excretion of
faeces and urine; if excretion was observed the procedure was repeated in 48 h time.
A PPA period was calculated by noting the calving date and the signs of first oestrus
in the cow or heifer after calving with the cow being mated at this time and this was
accompanied by the farmer mating the animal at that point.
Figure 6.1. Weigh-suckle-weigh method at Karang Kendal North Lombok research site.
6.4 Results
The BCS of the 9 heifers and cows are shown in Table 6.1. It ranged from 2.0-3.0 with
no difference between heifers and cows. Milk production was low over the 12 weeks
(1.45 and 1.68 kg/day, for heifers and cows, respectively, not significantly different).
There was little change over the 12 weeks of lactation (Figures 6.2 and 6.3). The mean
highest production for heifers occurred at week 12 (1.70 ± 0.19) and cows occurred at
week 8 (1.84 ± 0.27). There was no significant difference between heifers and cows
in average milk production over the 12 weeks or at any specific time point (4, 8 or 12
weeks).
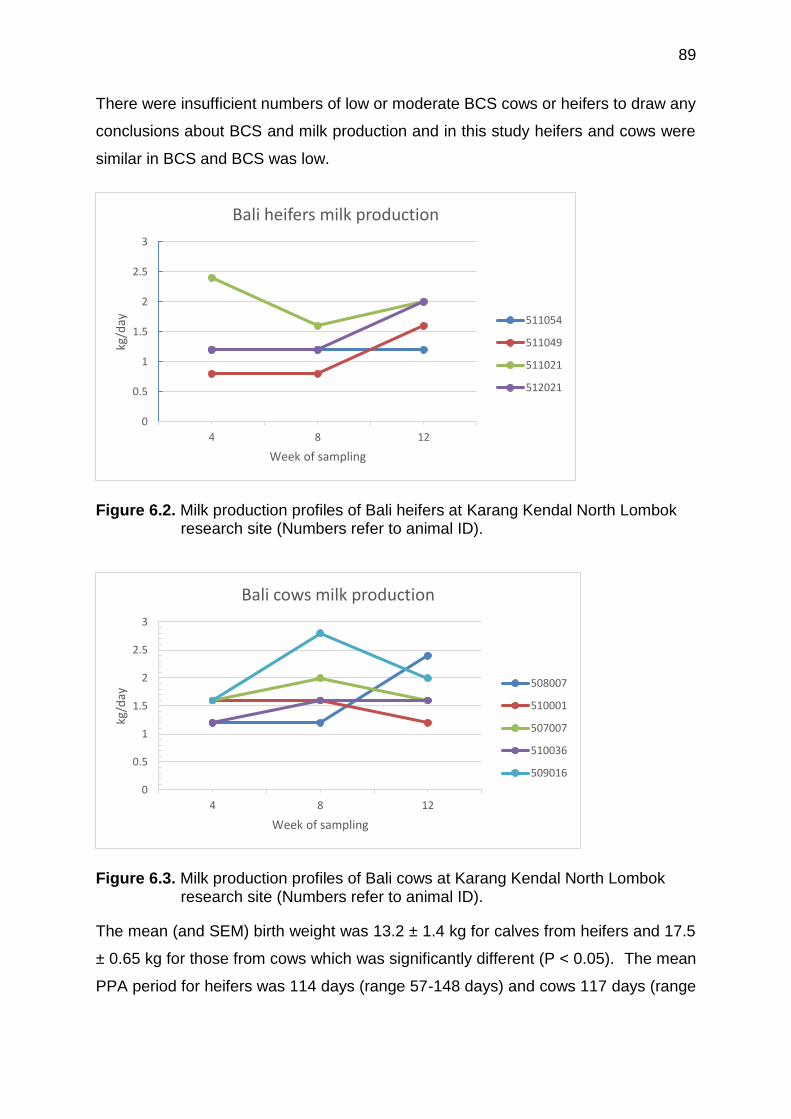
89
There were insufficient numbers of low or moderate BCS cows or heifers to draw any
conclusions about BCS and milk production and in this study heifers and cows were
similar in BCS and BCS was low.
Figure 6.2. Milk production profiles of Bali heifers at Karang Kendal North Lombok research site (Numbers refer to animal ID).
Figure 6.3. Milk production profiles of Bali cows at Karang Kendal North Lombok research site (Numbers refer to animal ID).
The mean (and SEM) birth weight was 13.2 ± 1.4 kg for calves from heifers and 17.5
± 0.65 kg for those from cows which was significantly different (P < 0.05). The mean
PPA period for heifers was 114 days (range 57-148 days) and cows 117 days (range
0
0.5
1
1.5
2
2.5
3
4 8 12
kg/d
ay
Week of sampling
Bali heifers milk production
511054
511049
511021
512021
0
0.5
1
1.5
2
2.5
3
4 8 12
kg/d
ay
Week of sampling
Bali cows milk production
508007
510001
507007
510036
509016
90
53-262 days) but the number of observations was low with only 3 heifers and 4 cows
observed and one heifer and one cow dropping out of the system with no observations
(Table 6.1). Feed and supplements that were directly provided such as rice straw
(approximately 5 kg per day), Elephant grass (Pennisetum purpureum) (approximately
20 kg per day), native grass (approximately 8-10 kg per day), Sesbania grandiflora
(approximately 7-10 kg per day), Gliricidia (Gliricidia sepium) (approximately 4-5 kg
per day) and peanut straw (approximately 2-4 kg per day) were offered to the cows
and animals both prior to and during the study, and given two or three times a day
around 6.30 – 8.00 am, 12 – 2 pm and 4.30 – 6.00 pm.
Table 6.1. Calving date and Post-Partum Anoestrus (PPA, days) of heifers and cows with Body Condition Score (BCS, 1-5 scale) and milk production (kg/d) at Karang Kendal village of North Lombok research site. BCS and milk production were the average over 12 weeks of measurement.
6.5 Discussion
These results are in broad agreement with those of Sariubang et al. (2000) and Talib
et al. (2003) and a study of Jelantik et al. (2008) who also measured it with various
supplements of grass hay and concentrate (rice bran, cornmeal, leucaena leaf and
fish meal) under a village based system in Kupang villages East Nusa Tenggara (1.50
± 0.09 kg/d). While in Besipae village West Timor, such measurement conducted with
No Animal
ID
Animal
Type
Calving
Date
Calf Birth
Weight (kg)
PPA
(Days) BCS (1-5)
Milk Production
(kg/day) 2014
1 511054 Heifer 22-Jun 16 57 2.2 1.2
2 511049 Heifer 22-Jun 11 139 2.5 1.1
3 511021 Heifer 26-May 15 148 2.5 2.0
4 512021 Heifer 21-Aug 10.7 M 2.7 1.5
5 508007 Cow 14-Aug 16 75 2.0 1.6
6 510001 Cow 19-Aug 17.5 78 3.0 1.5
7 507007 Cow 4-Sep 19.9 262 2.3 1.7
8 510036 Cow 9-Sep 17 M 2.7 1.5
9 509016 Cow 11-Sep 17 53 2.7 2.1

91
cows grazing natural pasture and given supplements consisted of 750 g rice bran (Milk
production, 1.48 ± 0.04 kg/d, and 1.42 ± 0.02 for control cows (Belli et al. 2008)). They
confirm the low milk production of Bali cows and suggest little difference between
heifers and cows (Table 6.1). Initially larger numbers were planned for this experiment
but insufficient numbers of cows and heifers calved in the time period and the
experiment was curtailed to these numbers.
The values for milk production are important in assessing energy and protein
requirements of Bali cows in lactation. They suggest that energy and protein
requirements do not increase markedly in lactation and provide a reason why Bali
cattle are able to perform well under adverse conditions (Wirdahayati & Bamualim
1990; Talib 1998; Talib et al. 1998). Sariubang et al. (2000) in South Sulawesi,
Indonesia reported that the mean milk production of Bali cows aged >6 years with a
minimum of four lactations was 1.2 vs 0.8 kg/day for cows that had a high and low
survival rate of calves respectively. This demonstrated that milk production was
important in calf survival and that the value recorded in the present experiment should
be adequate. Mean calf birth weight was higher in cows than heifers (Table 6.1), as
might be expected, but none of the calves had such low birth weight that there was a
potential risk of mortality.
There was quite a range in PPA among heifers and cows. The numbers were not high
enough to draw meaningful statistical analysis and this is done on a broader data base
in Chapter 7. Nevertheless with a mean PPA of 114-117 days for the current animals
(but a much wider range from 53-262 days), the ability of Bali heifers and cows to
conceive close to the 100 day post-calving period appears high. Lactation did not
produce complete anoestrus and there were individuals (4 out of 9 animals) in this
experiment that returned to oestrus in <80 days further supporting the observation that
there is potential for high reproductive rates in Bali cattle.
6.6 Conclusion
Milk production in Bali cows and heifers is low (approximately 1.7 kg/d) with no marked
lactation curve over the 12 weeks of lactation. BCS of these cows ranged from 2.0-
3.0/5 which is not high. PPA was approximately 114-117 days but 4/9 cows returned
to oestrus in less than 80 days supporting the observation that Bali cows are highly

92
fertile even while lactating. There was no difference between heifers and cows in any
of the parameters related to milk production and reproduction.

93
Chapter 7 Reproductive performance of heifers and cows within
villages
7.1 Background
Previous surveys have shown that reproduction rate varies widely across Eastern
Indonesia affected by breed type and region. For example Bakry (1994) has shown
that calving rate varies from 63 to 78 %. These studies were done by survey and not
long term monitoring. Where long term monitoring was done by Panjaitan et al. (2014)
calving percentage was low (62%), while Wiryosuhanto (1997) reported calving rates
of Bali cattle were 64%, 78%, 72%, 74% in Timor, Flores, Sumbawa and Lombok,
respectively. A lower rate of calving percentage than those has also been found by
Darmadja (1980) in Bali (52%). Bali cattle appear to have better reproduction rates in
villages with 49% pregnant within 100 days of calving compared to 12% pregnant with
Ongole cows (Mayberry et al. 2016). None of the studies has looked at first calving
heifers and Schatz and Hearnden (2008) have shown that managing the heifer is
important in overall productivity of the cow as poor body condition score of the first calf
heifer markedly reduces calving to <20% in the subsequent year. Longitudinal
monitoring provides a much better way to look at factors which affect reproduction rate
than short term studies. The opportunity was taken in this chapter to use a large
number of longitudinal monitor data from villages which had followed cow and heifer
(where possible) reproduction over a number of years. Statistical analysis of such data
provides the means to investigate cow and heifer reproduction and with a large
number of animals there is greater statistical power than in previous studies.
Body condition score at calving has been shown to be an important parameter to
explain calving and weaning percentage in Brahman cows across northern Australia
(McGowan et al. 2014). From this study, BCS at calving needed to be >3.5/5 for high
calving percentage of >75%. BCS is a simple way to assess cattle production as
nutritional status and animal parity are evaluated through BCS (Montiel & Ahuja 2005).
Bali cattle have not been studied in this way with most reproduction studies conducted
with surveys or limited cow numbers over 3-5 years eg. Fordyce et al. (2002). The
collation of data outlined here provides an opportunity to develop relationship between
BCS and reproduction rate for Bali cows and heifers.

94
Other reproductive performance parameters that are important in studying
reproduction rate are post-partum anoestrus (PPA), inter-calving interval (ICI) and
percentage of cows pregnant within 100 days. The reproductive performances of ICI
and PPA values are strongly related to the productivity of cattle, and this can reduce
production efficiency of beef production systems (Short et al. 1990). The ICI of Bali
cattle in Eastern Indonesia has not been comprehensively measured using large data
sets so the current study provides an opportunity to study this. Time to resumption of
post-partum ovarian activity or return to oestrus can be calculated and measured
through PPA. The length of PPA and ICI of Bali cattle need to be identified as these
reproductive performance parameters can be used in breeding and mating
management programs. Poppi et al. (2011) reported that PPA in Bali cattle is shorter
than in Bos indicus cows and this is reflected in the percentage of cows pregnant within
100 days of calving which is much higher for Bali cattle (Mayberry et al. 2016). The
ideal length for ICI is <365 days with the assumption of having a calf each year which
means that cows need to become pregnant within 100 days of calving, hence the value
of that parameter. This ideal ICI length is for optimal fertility (Reynolds et al. 1990).
All these parameters reflect different ways of describing reproductive performance but
PPA provides a reason as to why ICI and percentage cows pregnant within 100 days
may not reach targets to achieve a calf each year. Other reasons such as poor mating
management (recognition of oestrus, access to a bull or AI in a timely fashion, bull
fertility etc) and foetal loss can thus be identified. In several longitudinal studies, data
were collected for most of these parameters and a combined analysis will elucidate
reproductive performance in heifers and cows and the reasons for the results.
7.2 Aim of experiment
This experiment aims to determine reproductive performance parameters in Bali cattle
and elucidate reasons for the values obtained. Factors such as BCS at calving, village
site (region of Indonesia) and cow or heifer status were investigated as contributors to
the reproductive performance.
7.3 Materials and methods
Large data sets of heifer and cow reproduction from several sites and projects were
collated, namely: (1) Central Lombok – SMAR 2009 036 Project, (2) Kelebuh village

95
(Lombok) – AS2 2000 2013 Project, (3) Tandek village (Lombok) - AS2 2000 2013
Project, (4) N. Lombok – LPS/2008/038 Project, (5) Kendari Southeast Sulawesi -
Lapangisi village - LPS/2008/038 Project, and (6) Central Lombok – Scale Out Project.
Most of the studies were from Lombok but the sites vary in rainfall and feed supply. In
general North Lombok has a long dry season, Central Lombok varies widely with
village type from long dry season to short dry season. Kendari, South East Sulawesi
has a short dry season. These seasonal conditions impact on feed supply and the
requirement of farmers to buy external feed of variable quality. No attempt is made to
relate reproductive data to climatic conditions as the consequences of the extent of
the dry season impact on the nutrition of the cow and this is reflected in BCS. Nine
thousand five hundred and seventy one individual animal production year data were
recorded in the study representing seven hundred and twenty one individual animals
who contributed to the final data set. In this study, criteria were imposed as to the
quality of the data and whether they could be accepted for analysis. This culling of
animal data was based on the quality of the records related to animal ID, dates and
biological possibility (eg mating just before calving etc) and the cow to have at least 2
calves. There were insufficient heifer data from Central Lombok but Kendari had a
high proportion of first calving heifers in the data set (86%).
The data were collected for other purposes but for this study the data on cow BCS at
calving, time of calving over the 2-5 years of each study and other associated
parameters were collected. Some sites measured more frequently but all sites had a
measure of BCS at calving, time of calving, some aspects of weight and also a
description of cow or heifer. The latter definition had problems as descriptions were
not complete at all sites and this limited the analysis based on heifers or cows.
Nevertheless longitudinal reproductive data could be interrogated to determine
reproductive rate (as measured by inter-calving interval, calving percentage, post-
partum anoestrus etc) in a way that has not previously been described by survey data
in Indonesia. One site, Kendari – Lapangisi, were mostly heifers (86%) and inferences
are drawn about heifer reproduction from this site as most other sites had too few
animals classed as heifers or uncertainty about heifer or cow status. Some sites had
data in a format to enable specific parameters to be investigated. These are identified
as site specific in Tables and Figures compared to combined site analysis.

96
The basic reproductive and regular data in each site/village such as Animal ID, Year,
Date of Measurement, Date Mating, Date Calving, Birth Date, Sex, Calf ID, Weaning
Date, Weaning Weight, Weight, Inter-calving Interval, PPA, BCS, Stage of Pregnancy,
Lactation Status etc. have been collated to investigate reproductive performances of
Bali cattle. Central Lombok site data from the Scale-Out project from year 2008 to
2010 were collated for measuring inter-calving interval, return to oestrus, and mating
procedure. The basic data were Date of Measurement, Year, Weight, BCS, Inter-
calving interval (ICI), Date Mating (multiple mating dates - mating to re-mating – might
be categorised as return to oestrus), Date Calving, Gestation Length, Weaning Date,
Weaning Weight, Post-partum anoestrus (PPA), etc.
A descriptive statistical method using Microsoft Excel has been applied to analyse
these large data sets. The data sets have been categorised based on the quartile
percentage categories (eg. 4 out of 10). These categories have been applied to
analyse all of the data. The starting data set included 9,571 rows of data with a single
row representing an individual animal production year. The final data set included
1,737 animal production years and represented 721 individual cows. On average an
individual cow contributed 2.08 animal production years of data and ranged between
1-5 years. Each village was represented by 347 animal production years of data and
ranged between 90-1143 records. Longitudinal data analysis typically included a table
giving the overall sample size, and demographic, etc.
From the large initial data set the above final data were analysed as these satisfied
criteria relating to data quality (missing or wrong ID, cows being sold etc) and
measurement of parameters of interest. Some studies were scale out studies of a
technology, eg cow management, and as a consequence only basic information was
collected eg BCS at calving, calf birth date etc from which reproductive parameters
could be estimated. Other studies had more detailed and frequent measurement eg
monthly weights and BCS. The data were analysed based on what was considered
important parameters for reproduction, eg BCS at calving, and this enabled a much
larger data set to be analysed to get a more comprehensive analysis of reproduction
in Bali cattle.
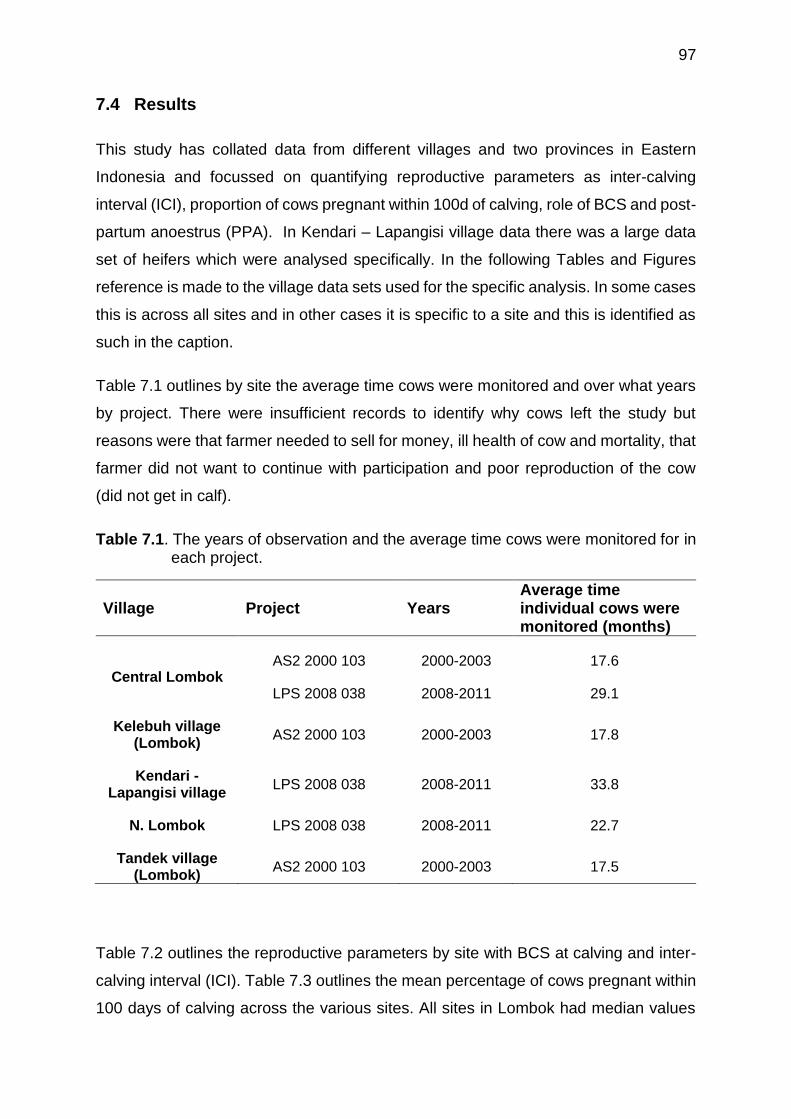
97
7.4 Results
This study has collated data from different villages and two provinces in Eastern
Indonesia and focussed on quantifying reproductive parameters as inter-calving
interval (ICI), proportion of cows pregnant within 100d of calving, role of BCS and post-
partum anoestrus (PPA). In Kendari – Lapangisi village data there was a large data
set of heifers which were analysed specifically. In the following Tables and Figures
reference is made to the village data sets used for the specific analysis. In some cases
this is across all sites and in other cases it is specific to a site and this is identified as
such in the caption.
Table 7.1 outlines by site the average time cows were monitored and over what years
by project. There were insufficient records to identify why cows left the study but
reasons were that farmer needed to sell for money, ill health of cow and mortality, that
farmer did not want to continue with participation and poor reproduction of the cow
(did not get in calf).
Table 7.1. The years of observation and the average time cows were monitored for in each project.
Village Project Years Average time individual cows were monitored (months)
Central Lombok AS2 2000 103
LPS 2008 038
2000-2003
2008-2011
17.6
29.1
Kelebuh village (Lombok)
AS2 2000 103 2000-2003 17.8
Kendari - Lapangisi village
LPS 2008 038 2008-2011 33.8
N. Lombok LPS 2008 038 2008-2011 22.7
Tandek village (Lombok)
AS2 2000 103 2000-2003 17.5
Table 7.2 outlines the reproductive parameters by site with BCS at calving and inter-
calving interval (ICI). Table 7.3 outlines the mean percentage of cows pregnant within
100 days of calving across the various sites. All sites in Lombok had median values
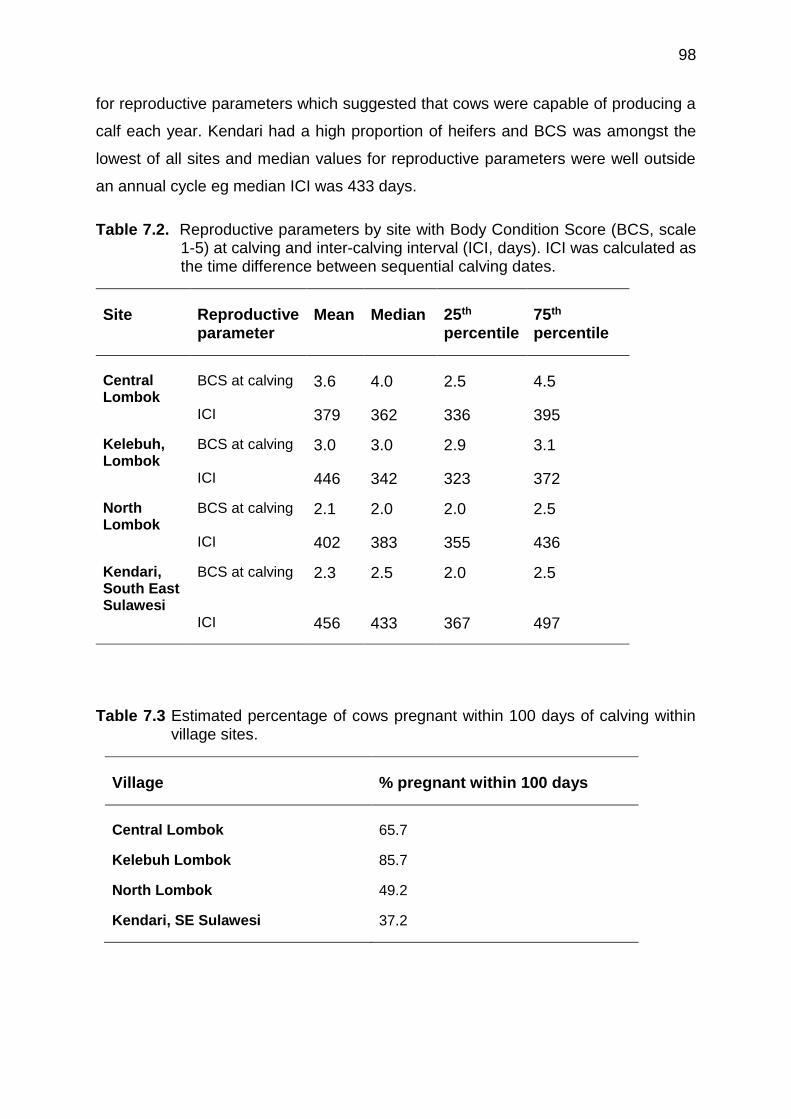
98
for reproductive parameters which suggested that cows were capable of producing a
calf each year. Kendari had a high proportion of heifers and BCS was amongst the
lowest of all sites and median values for reproductive parameters were well outside
an annual cycle eg median ICI was 433 days.
Table 7.2. Reproductive parameters by site with Body Condition Score (BCS, scale 1-5) at calving and inter-calving interval (ICI, days). ICI was calculated as the time difference between sequential calving dates.
Site Reproductive parameter
Mean Median 25th percentile
75th percentile
Central Lombok
BCS at calving 3.6 4.0 2.5 4.5
ICI 379 362 336 395
Kelebuh, Lombok
BCS at calving 3.0 3.0 2.9 3.1
ICI 446 342 323 372
North Lombok
BCS at calving 2.1 2.0 2.0 2.5
ICI 402 383 355 436
Kendari, South East Sulawesi
BCS at calving 2.3 2.5 2.0 2.5
ICI 456 433 367 497
Table 7.3 Estimated percentage of cows pregnant within 100 days of calving within village sites.
Village % pregnant within 100 days
Central Lombok 65.7
Kelebuh Lombok 85.7
North Lombok 49.2
Kendari, SE Sulawesi 37.2
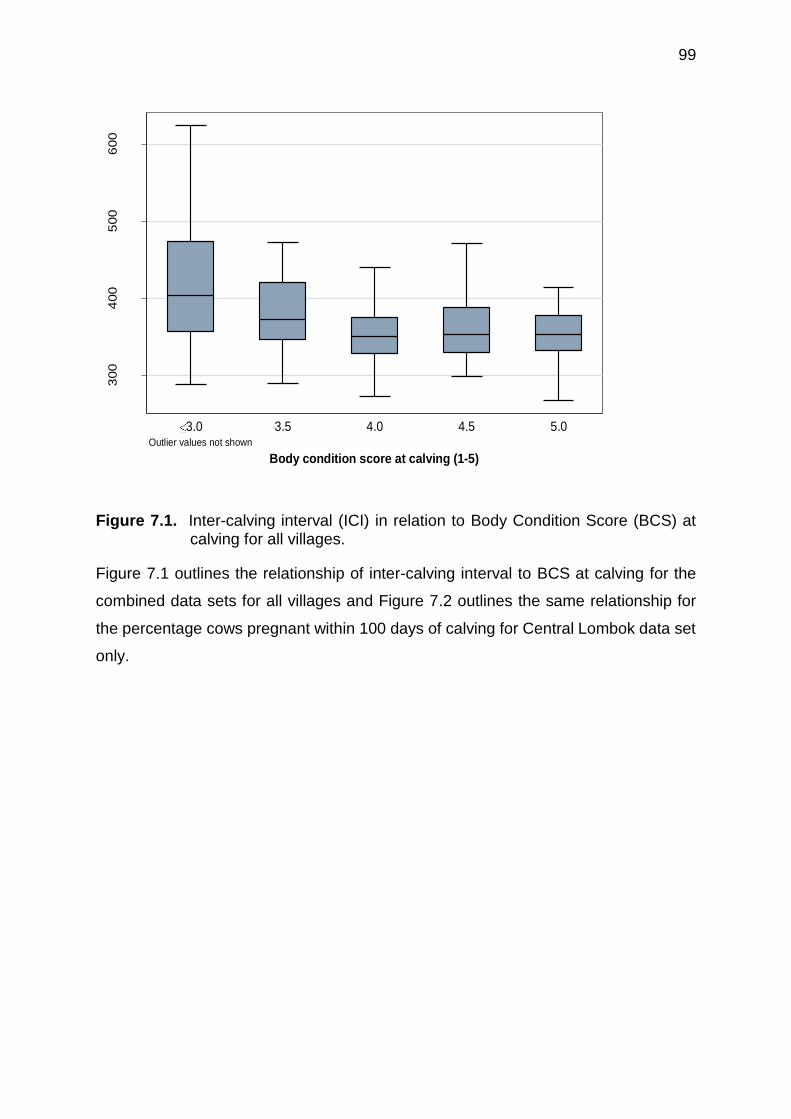
99
Figure 7.1. Inter-calving interval (ICI) in relation to Body Condition Score (BCS) at calving for all villages.
Figure 7.1 outlines the relationship of inter-calving interval to BCS at calving for the
combined data sets for all villages and Figure 7.2 outlines the same relationship for
the percentage cows pregnant within 100 days of calving for Central Lombok data set
only.
300
400
500
600
Inte
r-c
alv
ing
in
terv
al (d
ay
s)
3.0 3.5 4.0 4.5 5.0Outlier values not shown
Body condition score at calving (1-5)
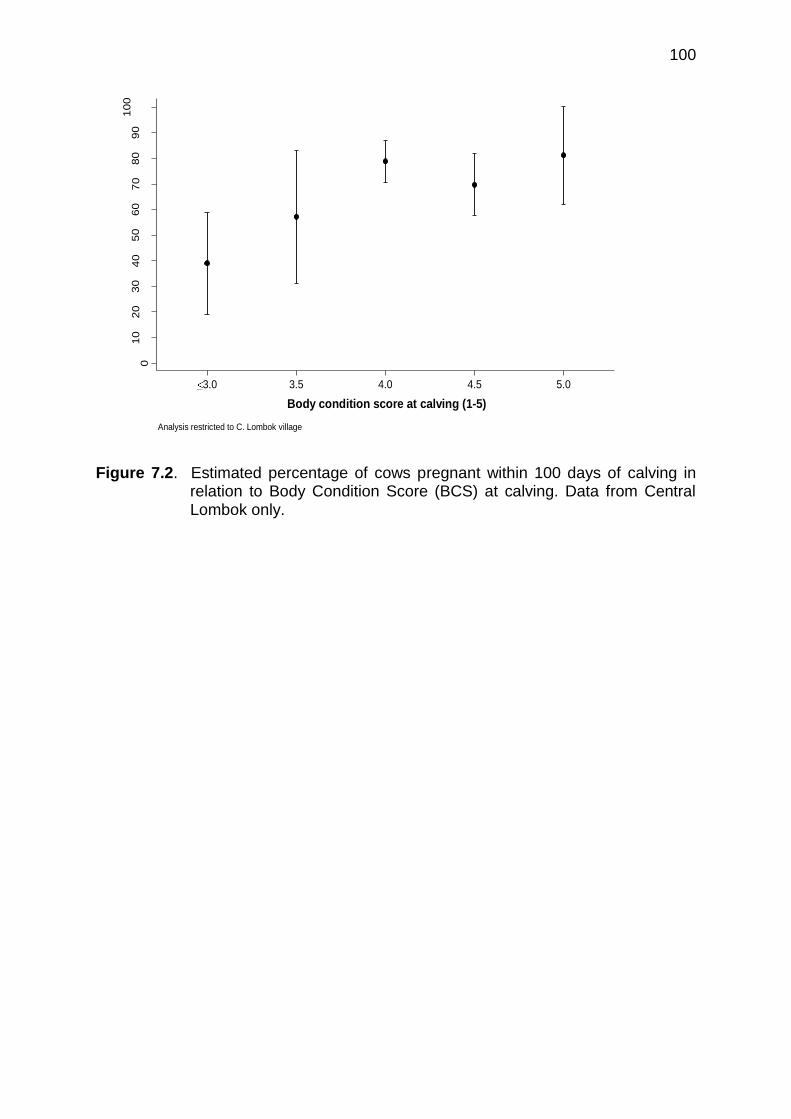
100
Figure 7.2. Estimated percentage of cows pregnant within 100 days of calving in relation to Body Condition Score (BCS) at calving. Data from Central Lombok only.
010
20
30
40
50
60
70
80
90
100
Perc
en
t p
reg
na
nt
wit
hin
10
0 d
ay
s o
f c
alv
ing
(%
)
3.0 3.5 4.0 4.5 5.0
Body condition score at calving (1-5)
Analysis restricted to C. Lombok village
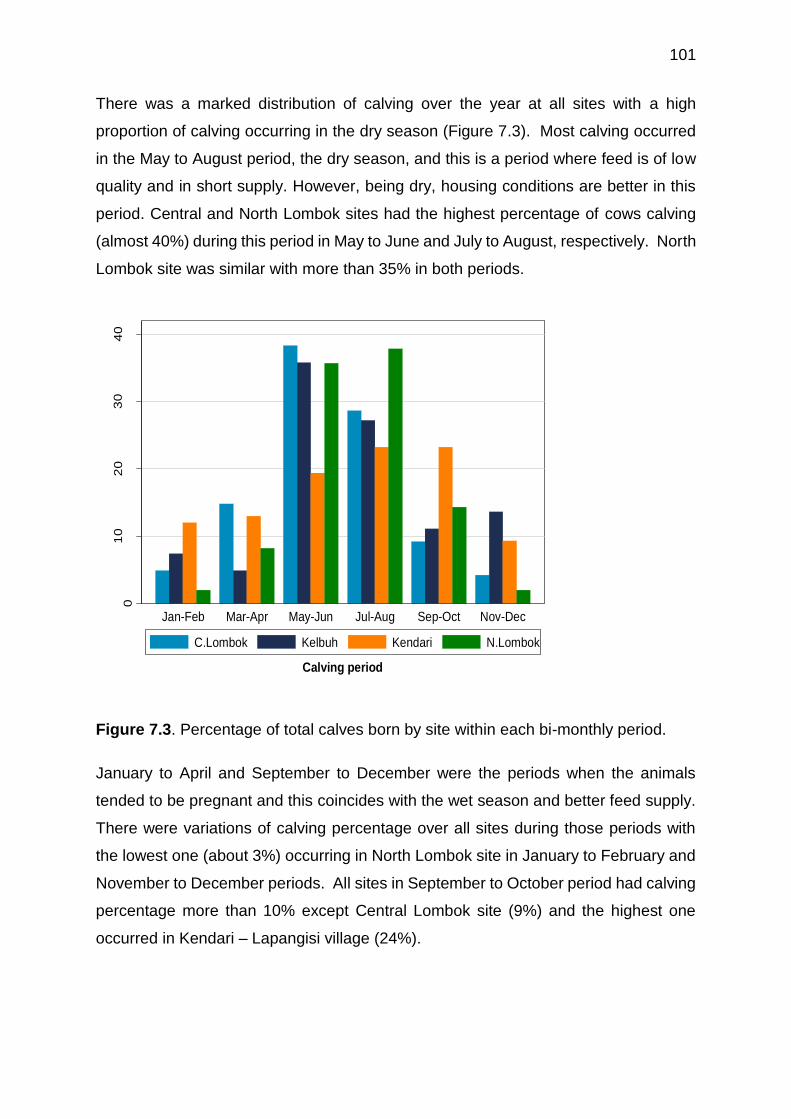
101
There was a marked distribution of calving over the year at all sites with a high
proportion of calving occurring in the dry season (Figure 7.3). Most calving occurred
in the May to August period, the dry season, and this is a period where feed is of low
quality and in short supply. However, being dry, housing conditions are better in this
period. Central and North Lombok sites had the highest percentage of cows calving
(almost 40%) during this period in May to June and July to August, respectively. North
Lombok site was similar with more than 35% in both periods.
Figure 7.3. Percentage of total calves born by site within each bi-monthly period.
January to April and September to December were the periods when the animals
tended to be pregnant and this coincides with the wet season and better feed supply.
There were variations of calving percentage over all sites during those periods with
the lowest one (about 3%) occurring in North Lombok site in January to February and
November to December periods. All sites in September to October period had calving
percentage more than 10% except Central Lombok site (9%) and the highest one
occurred in Kendari – Lapangisi village (24%).
01
02
03
04
0
Pe
rcen
tag
e o
f ca
lvin
gs
(%
)
Jan-Feb Mar-Apr May-Jun Jul-Aug Sep-Oct Nov-Dec
Calving period
C.Lombok Kelbuh Kendari N.Lombok

102
7.5 Discussion
Even though several studies have been conducted to measure reproductive
performance in Bos indicus cattle (eg (McGowan et al. 2014), the current study was
the first study to analyse large longitudinal data sets from different research sites so
as to measure the reproductive performance of Bali cattle in the Eastern Indonesian
tropical region.
This study has shown that Bali cattle potentially can have very high reproduction rates
as measured by ICI or the percentage of cows pregnant within 100 days and
biologically should be able to deliver high reproduction rates with an expectation that
calving percentage of approximately 80-90% is achievable under good management.
This does not occur widely at present as observed in Figures 7.1 and 7.2 and Tables
7.2 and 7.3. The terms, ICI and percentage cows pregnant within 100 days of calving,
describe overall cow reproduction and provide a means of examining the ability of the
system to produce a calf each year, an important parameter for a smallholder
(Rahman et al. 2001). These values are derived from manipulation of the limited data
which were collected in longitudinal monitoring of the herd. ICI was simply calculated
as the difference between birth dates and the percentage of cows pregnant at 100
days post-calving was similarly calculated from birth date and subtracting an assumed
gestation length to derive conception date which is then related to previous calf birth
date and so calculating when pregnant from calving. There is confidence in these
dates from the monitor sites. The large number of longitudinal studies collated here
came from various ACIAR and related projects. These projects largely set out to
implement a mating scheme of observation of oestrus, a mating policy of access to a
bull and a weaning process (Poppi et al. 2011) and, as such, observation by the farmer
and extension officer was much higher than would be the normal situation and the
values derived would more closely represent what could be achieved rather than what
is being achieved across the eastern islands now. Survey data indicate a wide
variation in reproduction rates (as defined by various parameters) in Bali cattle with
ICI ranging from 374 to 592 days (Bakry 1994).
The main factor limiting reproductive efficiency in cattle is post-partum anoestrus
(Montiel & Ahuja 2005). A PPA period less than 85-100 days is required if cows are
to have the opportunity to produce a calf each year (Peters & Lamming 1986; Wright

103
et al. 1987). Prolonged post-partum anoestrus will prevent achievement of a 12 month
calving interval as commonly occurs with Bos indicus and Bos taurus/Bos indicus cows
from tropical regions. It was not possible to determine PPA accurately from the
observational data as it relied on good observations of cows cycling and there was
less confidence in these observations so these calculations were not pursued. One of
the challenges in effective mating of cows is the detection of oestrus. Nevertheless the
high percentage of cows pregnant within 100 days at some sites suggests that
lactation PPA is not an issue with Bali cattle in good BCS and that other factors such
as access to a bull or AI were more important.
Nutrition is the key limitation to reproduction as differences in nutritional status of
animals across the various sites involved with this study were observed as reflected
in the differences in BCS. The main feed types used by farmers are rice straw, native
grass, elephant grass, Sesbania, Gliricidia and peanut straw. Observations of feed
types and amounts were not made in all studies and BCS was used the parameter to
summarise level of nutrition. Similarly BCS was measured by different operators at
each site and over different years but all used a 1-5 scale according to Teleni et al.
(1993). It was not possible to normalize scores between operators and years but all
were trained under the same system.
Level of nutrition, that is reflected in BCS, and suckling management are the two main
factors contributing to the length of post-partum anoestrus (Montiel & Ahuja 2005).
Additionally, extended-suckling during lactation will also prevent ovulation and thus
prolong post-partum anoestrus in some breeds. Bali cows appear to have low PPA
due to suckling as reflected in the Kelebuh data (Tables 7.2 and 7.3), Fordyce et al.
(2003) and Chapter 6 and the high ICI appears more due to BCS at calving (Figure
7.2). The data are not extensive enough to examine this across all sites except that,
where good management is practised, the ICI is low. An appropriate weaning time will
be of benefit, mainly by conserving BCS in cows. Weaning at four months of age (from
7 months of weaning age) can advance the next calving date by 39 days in other cattle
breeds (Schottler & Williams 1975). There was no evidence of a marked lactation
anoestrus in lactating heifers and cows (Chapter 6).
The ICI and percentage of cows pregnant at 100 days in this study showed that some
sites are able to achieve high reproduction rates (Tables 7.2, 7.3 and Figures 7.1, 7.2).

104
Bali cattle in this region commonly had an ICI around 400 to 500 days prior to applying
better management systems (Bakry 1994; Winugroho 2002; Fordyce et al. 2010;
Saleh et al. 2011; Pohan & Talib 2014). This was related to BCS (Figures 7.1, 7.2)
and a target BCS of at least >3.5 would ensure high reproduction rates. This is not
dissimilar to observations in the Cash Cow project in Australia with Bos indicus cattle
(McGowan et al. 2013; McGowan et al. 2014) where there was a positive relationship
of cows pregnant within 100 days of calving and BCS. However the median BCS at
calving and the median BCS throughout the year (median 2-2.5) in the current study
was much lower than this recommendation for most sites, reflecting the difficulty in
maintaining a high level of nutrition to cows in this environment. Kelebuh and Central
Lombok project villages achieved a median BCS value of 3-4 (Table 7.2) and much
higher reproduction rates than other village sites even from the same general area.
This suggests that attention to the nutrition of the cow and in particular a focus on BCS
at calving would be a useful management tool for farmers and extension personnel to
implement to achieve high reproduction rates.
Inefficient beef cattle production in Indonesia was mainly due to long ICI (Winugroho
2002). In the current study one village, Kendari – Lapangisi village in SE Sulawesi
had a long ICI of 433 days while the Lombok sites had 342-383 days. This shows the
wide variation between sites. Kendari – Lapangisi village had mainly heifers (86%)
and poor heifer performance is a common feature around the world where there is
poor nutrition (Schatz & Hearnden 2008). Such a phenomenon appears here also
where this site also had low BCS. Both factors would contribute to the poor
performance at this site but emphasizes once again that heifers require better nutrition
and closer attention if they are to become pregnant within 100 days of their first calf
(Winugroho 2002). Although enough heifer data could not be generated across all
sites, the Kendari data suggest that heifer return to pregnancy after first calving is low.
A closer examination of the data from Kendari, South East Sulawesi highlights some
important features. This was a village site that had a high percentage of heifers (86%)
which entered the project in calf and so the reproductive parameters reflect the
influence of first calf heifer reproduction over the course of the monitoring. Table 7.2
clearly shows the low BCS of heifers at this site, a result due to poor nutrition and low
management skills of the participating farmers. Other farm activities were of higher

105
interest at this site. BCS was a median of 2.5 with a median ICI of 433 days clearly
leading to poor reproductive outcomes at this site. This may be contrasted with North
Lombok village sites which is in a more difficult environment (longer dry season) which
had a median BCS of 2 and a median ICI of 383 days. Some of this difference may be
due to different teams scoring BCS but also reflects cow (North Lombok) versus first
calving heifers (Kendari) at the start of the monitoring exercise and the greater
nutritional demand of the heifer which needs to continue to grow but also reproduce.
This suggests that feeding the heifer which has not reached mature size and weight
is very important in overall cow productivity and reproductive outcome over her lifetime.
Such a principle is well established in Bos indicus heifers in northern Australia (Schatz
& Hearnden 2008). Another aspect which affects these results is the management
skills of the farmers where Kendari farmers had limited cow management skills (what
to feed, maintaining BCS, observation of oestrus and mating).
Across all sites, most cows calved in the dry season. Calving early in the dry season
will enable the cow to reach higher BCS at calving as the level of nutrition immediately
preceding calving is the highest for the year. However if lactation is prolonged then
BCS will decline rapidly and it is hard to regain BCS and cows could trend downwards
in BCS eventually reaching low critical BCS which will affect reproduction (Antari et al.
2012; Mayberry et al. 2016). Dry season nutrition and weaning age becomes very
important in this system. Calving in the dry season is preferred as calf mortality is high
in cows calving in the wet season due to poor housing and muddy conditions (Fordyce
et al. 2003).
There was a significant exit of cows from the initial enrolment. This may also skew the
results towards better reproductive performance as one of the reasons given for sale
of cows was poor reproductive performance (Poppi pers. comm.). Some farmers also
chose not to participate for the whole project due to reasons such as village dynamics
and personalities and the effort required to participate.
7.6 Conclusion
Bali cattle are inherently very fertile and could easily achieve high reproductive
performance and improve the current national values for calving and weaning
percentage. Applying the Integrated Village Management System (Poppi et al. 2011)

106
of a medium - high BCS, weaning at 5-6 months and access to a sound bull should
result in high calving % (around 80%) each year. ICI <365 days was achieved in the
well managed herds in this study. There appeared to be a relationship between BCS
at calving and reproductive parameters such that a BCS at calving of 3.0-4/5 would
enable high reproductive performance to be achieved. First calving heifers will require
higher nutrition and BCS than cows in recognition that they are still growing and have
not yet reached mature size which might take another 2-3 years to achieve under this
environment.

107
Chapter 8 General discussion
8.1 General comments
This study concentrated on Bali cattle, an indigenous breed within Indonesia. They are
small cattle (cow weights approximately 220 kg for a standard reference cow in
moderate body condition, Mayberry et al. (2016)) and appear to have high
reproduction rates (McCool 1992) but reproductive performance in villages appears
quite variable. They have many traits that make them a useful breed for smallholder
cattle producers especially the low weight of cows and potential for high reproductive
rates. The liveweight gain of weaned animals within villages appears low (Antari et al.
2014) which is a constraint.
A simple system of cow management (Integrated Village Management System)
appears to achieve high reproduction rates (Poppi et al. 2011; Panjaitan et al. 2014)
and this system is based on access to a bull at 40 days post-calving and weaning of
calves at 5-6 months of age to maintain BCS of cows. Despite this, many anecdotal
reports and other village survey data indicate quite wide variation in reproductive
performance suggesting that other factors such as BCS at calving and heifer
management might be important in influencing region wide reproductive performance.
The Straw Cow project (Antari et al. 2012; Mayberry et al. 2016) suggested that Bali
cattle may not be as sensitive to BCS at calving as Bos indicus breeds. Furthermore
longitudinal monitoring in previous studies have highlighted the difficulties in detecting
oestrus in these timid animals by both farmers and researchers and other methods
such as faecal progesterone and vaginal electrical conductivity may have a role in
detecting oestrus in experiments and in monitoring village cattle.
The opportunity was taken to sample animals involved in other experiments for some
of these parameters. One chapter (Chapter 7) collated a large amount of data from a
variety of sources which monitored reproduction in cows and heifers at the village level.
Key parameters were BCS at calving, time of calving and in some cases return to
oestrus or first mating event after calving. This study highlighted data quality
(observations and records) as a major issue in such analysis and emphasized the
need for closer observation of oestrous behaviour (and training to detect oestrus) and
the deployment of more objective measures or techniques of oestrous activity. In other

108
chapters (Chapter 3, 5 and 6) smaller numbers of cows were observed but oestrous
detection was more closely observed. Nevertheless these studies showed that a high
proportion of cows exhibited oestrus from 40-100 days post-calving ensuring that a
significant proportion of a regional herd (comprising many smallholders) have the
potential to calve each year. This is important for regional policy of breeds and
ensuring high reproduction by the provision of better extension services, AI and bull
access. Methods to improve experimental observation of oestrus and mating
procedures were examined in Chapters 3, 4 and 5 by studying faecal progesterone
profiles, protein hormones (leptin and IGF-1) and vaginal electrical conductivity. Milk
production and interaction with lactation anoestrus was examined in Chapter 6 where
no relationship was found and low milk production of cows in villages confirmed.
This thesis set out to examine post-partum anoestrus in Bali cattle as a contributing
factor to variable and in some cases low levels of reproduction reported across
Indonesia. Difficulties were experienced in data quality from large scale observational
studies and in successfully obtaining data and samples from adequate animal
numbers even in controlled experiments. Post-partum anoestrus was not found to be
a major issue with Bali cattle, although there were examples of long post-partum
anoestrus, but the development of some methods in this thesis, eg faecal progestogen,
would greatly aid such studies in the future. Achieving high reproduction rates at the
level of the village and individual smallholder is essential for high levels of production
and income. An understanding of the factors causing the variable reproduction rates
which are achieved across Indonesia is important to design better cow and heifer
management practices.

109
8.2 Reproductive parameters
The longitudinal monitor data from Chapter 7 clearly show that the median inter-
calving interval was 380 days across a number of sites. This suggests that post-partum
anoestrus is not an issue in Bali cows and that a satisfactory cow management system
can be implemented which potentially allows a calf each year within the normal
boundaries of reproductive function. The results further confirm the wide variability in
inter-calving interval but the observational data relating to time to first oestrus was not
good enough to clearly identify if post-partum anoestrus was an issue accounting for
this. The results could equally have been due to poor reproductive management
(access to a bull or AI) or foetal loss and remating. If some of the variability is post-
partum anoestrus, then this suggests that there is potential for genetic selection to
reduce this in Bali cattle and also nutritional manipulation of BCS. Chapter 6 looked at
milk production and Chapter 4 looked at BCS effects and, although animal numbers
were low, it was clear that lactation anoestrus was not a major issue despite some
animals exhibiting this. The marked exception to this was the first calf cows from
Chapters 3 and 4 where most of the moderate BCS cows showed anoestrus for 100
days after calving suggesting that PPA was an issue and related to BCS. Genetic
selection would be one means to reduce this effect even further although the
proportion of animals in which this appears is low. Management of the cow and heifer
for BCS and access to a bull or AI at timely events would be a simpler and more
effective strategy for the vast majority of cows and heifers.
Inter-calving interval proved to be the simpler parameter to examine reproductive
function in the village based studies because observational data for time to first oestrus
proved less reliable. The main reason for this was lack of confidence in the ability of
farmers and extension personnel to record this accurately in some sites and studies.
Under controlled experimental situations on research stations, these observations are
more easily made but under village conditions with intermittent visits it is not always
possible to have accurate records and so the conclusions about post-partum
anoestrus are tempered by this. Using faecal progestogen and selected sample times
(Chapter 3, see later) would enable village based studies to be conducted better but
at greater cost of time and money.

110
BCS at calving, as with other studies with Bos indicus and Bos taurus genotypes
(McGowan et al. 2014) proved to be the simplest way to manage cows and heifers for
high reproduction and aiming for a BCS of at least >3 would appear to be a good
strategy. This agrees with other studies with other genotypes but there is a suggestion
that some Bali cows can maintain high reproduction at low BCS even in the range 2.5-
3 (Chapter 4 and Chapter 7). A previous study (Straw Cow project, Mayberry et al.
(2016) outlined that it was difficult to maintain high BCS (>3) and very difficult to
increase BCS in lactating and dry cows and heifers. This was supported in Chapter 4
(although with very low animal numbers) where even a high leucaena diet required
some grain supplement to increase BCS to >3.5.
BCS is a simple management tool which is an outcome of level of nutrition. Level of
nutrition and BCS are key factors which influence onset of oestrus in cattle of all breeds
(Jolly et al. 1995; Bolanos et al. 1998; Lamb 1999; Orihuela 2000; Montiel & Ahuja
2005; Isle et al. 2007) and the effects are thought to be mediated by the protein
hormones, leptin and IGF-1. In Chapter 4, IGF-1 increased in the predictable manner
with a higher level of nutrition but with leptin there were no such changes in Bali cattle.
For various reasons the number of cattle were low and this appears to be the first
results for Bali cows. This issue requires further study to confirm the relationship with
Bali cows and the interaction with oestrous activity.
Delayed maturity in Bali heifers was not only all due to poor nutrition especially for the
cow at early stage of pregnancy, but it was also about management of mating after
calving, weaning and BCS in which good BCS at calving with good mating
management were most likely to be believed to accelerate liveweight gain and be able
to attain puberty at a younger age.
Milk production was low and confirms previous studies outlined in Chapter 6. It was
disappointing that the original experimental design to look at heifers and cows in high
and low BCS was not achieved due to availability of heifers and cows within the time
period in the village. Nevertheless the study did confirm that milk production was low
by the standards of other cattle genotypes and that a better controlled experiment
using cows of high and low BCS would be required to demonstrate if milk production
could be increased through management of BCS. Bali cattle have a reputation for high
calf mortality (Jelantik et al. 2008) and low milk production under times of nutritional

111
stress would contribute to this. Hence creep feeding as a strategy would be useful in
these circumstances to overcome the inherently low milk production of Bali cows even
under adequate levels of nutrition (Jelantik et al. 2010).
8.3 Methodology to study reproduction in the field
In this thesis various methods were examined for their usefulness in studying
reproduction in the field. The factors behind low annual reproduction are many eg post-
partum anoestrus, poor mating management, in utero calf loss. It is important to know
which are the main contributing factors if solutions are to be found. Observation of
oestrous activity in Bali cows is a problem as they are timid animals and overt signs of
oestrous activity are not always apparent. This coupled with poor farmer and extension
experience may mask the real reasons behind poor reproduction. Faecal progesterone
profile appears a useful method to identify cycling cows and in Chapter 3 this method
showed promise, albeit with low animal numbers. The method showed that there was
a reasonable correlation between faecal progestogen and plasma progesterone levels
but perhaps more importantly faecal progestogen patterns of change could be used
successfully to identify cycling and pregnant cows in a similar fashion to plasma
progesterone patterns. This method could be usefully employed in wider studies.
Vaginal electrical conductivity is a method used in intensive mating procedures eg AI
schemes (Rorie et al. 2002). In the current study, the method did not appear good
enough to be used routinely with Bali cattle but it would be useful to repeat aspects of
this work to ascertain features specific to Bali cattle.
8.4 Conclusions
This study has shown that Bali cows and heifers have high fertility and that
management in terms of low BCS and poor access to a bull or AI in a timely fashion
most likely accounts for reports of poor reproduction. There was also evidence that a
small proportion of cows and first-calf cows had a long inter-calving interval and in
those intensive studies this appeared to be due to post-partum anoestrus. There are
opportunities for genetic selection to minimize this effect although compared to Bos
indicus cows the problem is not as great and not as significant as first hypothesized.
It may be concluded that Bali cows and heifers are inherently highly fertile and that a
simple management system of a target BCS at calving (>3), access to a bull or AI from

112
40 days post-calving and weaning of calves at 5-6 months of age (or earlier if BCS is
declining fast due to poor nutrition) will enable annual calving or weaning percentage
of approximately 80% to be achieved. Heifers need to continue to grow and hence
they require better nutrition than cows during the latter stages of gestation and first
lactation if they are to achieve high reproduction levels after the first calf.

113
References
Abelson, KSP, Fard, SS, Nyman, J, Goldkuhl, R & Hau, J 2009, 'Distribution of H-3 -Corticosterone in Urine, Feces and Blood of Male Sprague-Dawley Rats after Tail Vein and Jugular Vein Injections', In Vivo, vol. 23, no. 3, pp. 381-6.
Abeygunawardena, H & Dematawewa, CMB 2004, 'Pre-pubertal and postpartum anestrus in tropical Zebu cattle', Animal Reproduction Science, vol. 82, pp. 373-87.
Aboulela, MB, Topps, JH & Macdonald, DC 1983, 'Relationships between intravaginal electrical-resistance, cervicovaginal mucus characteristics and blood progesterone and LH', Animal Reproduction Science, vol. 5, no. 4, pp. 259-73.
Adams, GP, Jaiswal, R, Singh, J & Malhi, P 2008, 'Progress in understanding ovarian follicular dynamics in cattle', Theriogenology, vol. 69, no. 1, pp. 72-80.
Adams, GP, Matteri, RL, Kastelic, JP, Ko, JCH & Ginther, OJ 1992, 'Association between surges of follicle-stimulating-hormone and the emergence of follicular waves in heifers', Journal of reproduction and fertility, vol. 94, no. 1, pp. 177-88.
Adrien, ML, Mattiauda, DA, Artegoitia, V, Carriquiry, M, Motta, G, Bentancur, O & Meikle, A 2012, 'Nutritional regulation of body condition score at the initiation of the transition period in primiparous and multiparous dairy cows under grazing conditions: milk production, resumption of post-partum ovarian cyclicity and metabolic parameters', animal, vol. 6, no. 2, pp. 292-9.
Ahima, RS & Flier, JS 2000, 'Adipose tissue as an endocrine organ', Trends in Endocrinology and Metabolism, vol. 11, no. 8, pp. 327-32.
Ahima, RS, Prabakaran, D, Mantzoros, C, Qu, DQ, Lowell, B, MaratosFlier, E & Flier, JS 1996, 'Role of leptin in the neuroendocrine response to fasting', Nature, vol. 382, no. 6588, pp. 250-2.

114
Ahmadzadeh, A, Carnahan, K & Autran, C 2011, 'Understanding puberty and postpartum anestrus', Proceedings of applied reproductive strategies in beef cattle, vol. 31, pp. 45-60.
Ahuja, C & Montiel, F 2005, 'CO-Synch enhances time to ovulation, cyclicity and pregnancy in anovulatory lactating Bos taurus/Bos indicus cows', Livestock Production Science, vol. 96, no. 2-3, pp. 279-83.
Alvarez, JG & Storey, BT 1989, 'Role of glutathione-peroxidase in protecting mammalian spermatozoa from loss of motility caused by spontaneous lipid-peroxidation', Gamete Research, vol. 23, no. 1, pp. 77-90.
Alves, NG, Pires, MdFA, Silva Filho, JMd, Saturnino, HM, Sa, WFd & Torres, CAA 2009, 'Estrus behavior in Guzera ( Bos taurus indicus) breed cows', Ciencia E Agrotecnologia, vol. 33, no. 5, pp. 1424-30.
Amstalden, M, Garcia, MR, Stanko, RL, Nizielski, SE, Morrison, CD, Keisler, DH & Williams, GL 2002, 'Central infusion of recombinant ovine leptin normalizes plasma insulin and stimulates a novel hypersecretion of luteinizing hormone after short-term fasting in mature beef cows', Biology of Reproduction, vol. 66, no. 5, pp. 1555-61.
Amstalden, M, Garcia, MR, Williams, SW, Stanko, RL, Nizielski, SE, Morrison, CD, Keisler, DH & Williams, GL 2000, 'Leptin gene expression, circulating leptin, and luteinizing hormone pulsatility are acutely responsive to short-term fasting in prepubertal heifers: Relationships z to circulating insulin and insulin-like growth factor I', Biology of Reproduction, vol. 63, no. 1, pp. 127-33.
Amstalden, M, Zieba, DA, Edwards, JF, Harms, PG, Welsh, TH, Stanko, RL & Williams, GL 2003, 'Leptin acts at the bovine adenohypophysis to enhance basal and gonadotropin-releasing hormone-mediated release of luteinizing hormone: Differential effects are dependent upon nutritional history', Biology of Reproduction, vol. 69, no. 5, pp. 1539-44.
Anke, M, Angelow, L, Groppel, B, Arnhold, W & Gruhn, K 1989, 'The effect of selenium deficiency on reproduction and milk performance of goats.', Archiv Fur Tierernahrung-Archives of Animal Nutrition, vol. 39, no. 4-5, pp. 483-90.

115
Antari, R, Syahniar, T, Mayberry, D, Pamungkas, D, Anderson, S & Poppi, D 2014, 'Evaluation of village-based diets for increasing the weight and condition of Ongole (Bos indicus) and Bali (Bos javanicus) cows in Indonesia', Animal Production Science, vol. 54, no. 9, pp. 1368-73.
Antari, R, Syahniar, T, Mayberry, D, Pamungkas, D & Poppi, D 2012, 'Strategies for increasing body condition score of Ongole and Bali cows in Indonesian villages', in 2nd Australian and New Zealand Societies of Animal Production Joint Conference, vol. 29, pp. 65-.
Anzar, M, Farooq, U, Mirza, MA, Shahab, M & Ahmad, N 2003, 'Factors affecting the efficiency of artificial insemination in cattle and buffalo in Punjab, Pakistan', Pakistan Veterinary Journal, vol. 23, no. 3, pp. 106-13.
Arango, JA, Cundiff, LV & Van Vleck, LD 2002, 'Genetic parameters for weight, weight adjusted for body condition score, height, and body condition score in beef cows', Journal of animal science, vol. 80, no. 12, pp. 3112-22.
Arechiga, CF, Ortiz, O & Hansen, PJ 1994, 'Effect of prepartum injection of vitamin-E and selenium on postpartum reproductive function of dairy-cattle.', Theriogenology, vol. 41, no. 6, pp. 1251-8.
Arije, GF & Wiltbank, JN 1971, 'Age and weight at puberty in hereford heifers', Journal of animal science, vol. 33, no. 2, pp. 401-&.
Arimbawa, IWP, Trilaksana, IGNB & Pemayun, TGO 2012, 'Gambaran Hormon Progesteron Sapi Bali selama Satu Siklus Estrus (Profile progestrone hormone Bali cattle in one oestrus cycle)', Indonesia Medicus Veterinus, vol. 1, no. 3, pp. 330-6.
Astiti, LGS & Panjaitan, T 2013, 'Serum Progesterone Concentration in Bali Cow During Pregnancy', Open Science Repository Veterinary Medicine, Online(open-access), vol. e70081943.
Bailey, TL, Monke, D, Hudson, RS, Wolfe, DF, Carson, RL & Riddell, MG 1996, 'Testicular shape and its relationship to sperm production in mature Holstein bulls', Theriogenology, vol. 46, no. 5, pp. 881-7.

116
Bakry, WR 1994, 'Reproductive characteristics and productivity of Bali and Ongole cattle in Nusa Tenggara, Indonesia', Ph.D. thesis, University of Queensland, St. Lucia, Queensland, Australia.
Bamualim, A & Wirdahayati, RB 2003, Nutrition and management strategies to improve Bali cattle productivity in Nusa Tenggara, Strategies to Improve Bali Cattle in Eastern Indonesia, ACIAR Proceedings of a Workshop 4-7 February 2002, Bali Indonesia.
Bandini, Y 1997, Sapi Bali (Bali cattle), PT Penebar Swadaya, Jakarta.
Barb, CR & Kraeling, RR 2004, 'Role of leptin in the regulation of gonadotropin secretion in farm animals', Animal Reproduction Science, vol. 82-3, pp. 155-67.
Bartha, T, Sayed-Ahmed, A & Rudas, P 2005, 'Expression of leptin and its receptors in various tissues of ruminants', Domestic animal endocrinology, vol. 29, no. 1, pp. 193-202.
Bartlewski, PM, Beard, AP & Rawlings, NC 1999, 'Ovarian function in ewes during the transition from breeding season to anoestrus', Animal Reproduction Science, vol. 57, no. 1-2, pp. 51-66.
Baruselli, PS, Reis, EL, Marques, MO, Nasser, LF & Bó, GA 2004, 'The use of hormonal treatments to improve reproductive performance of anestrous beef cattle in tropical climates', Animal Reproduction Science, vol. 82, pp. 479-86.
Bastidas, P, Troconiz, J, Verde, O & Silva, O 1984, 'Effect of restricted suckling on ovarian activity and uterine involution in Brahman cows.', Theriogenology, vol. 21, no. 4, pp. 525-32.
Bauman, DE, McBride, BW, Burton, JL & Sejrsen, K 1994, 'Somatotropin (BST): International Dairy Federation technical report', Bulletin of the International Dairy Federation, no. 293, pp. 2-7.

117
Belli, H, Jelantik, I & Holtz, W 2008, 'Improving Calf Performance by Supplementation in Bali Cattle Grazing Communal Pastures in West Timor', Indonesia Proc. Aust. Soc. Anim. Prod, vol. 27.
Belli, HLL & Jelantik, IGN 2011, 'Progesterone Profiles During the Estrous Cycle and Pregnancy of Grazing Bali Cows Supplemented with Concentrate', Animal Production, vol. 11, no. 2, pp. 79-87.
Bellows, RA, Thomas, OO, Riley, TM, Gibson, RB, Kieffer, NM, Urick, JJ & Pahnish, OF 1965, 'Feed effects on puberty in beef heifers', Journal of animal science, vol. 24, no. 2, pp. 588-&.
Berisha, B & Schams, D 2005, 'Ovarian function in ruminants', Domestic animal endocrinology, vol. 29, no. 2, pp. 305-17.
Berry, DP, Buckley, F & Dillon, P 2007, 'Body condition score and live-weight effects on milk production in Irish Holstein-Friesian dairy cows', animal, vol. 1, no. 9, pp. 1351-9.
Bishop, DK, Wettemann, RP & Spicer, LJ 1994, 'Body energy reserves influence the onset of luteal activity after early weaning of beef cows', Journal of animal science, vol. 72, no. 10, pp. 2703-8.
Blanchard, T, Ferguson, J, Love, L, Takeda, T, Henderson, B, Hasler, J & Chalupa, W 1990, 'Effect of dietary crude-protein type on fertilization and embryo quality in dairy-cattle', American Journal of Veterinary Research, vol. 51, no. 6, pp. 905-8.
Block, SS, Butler, WR, Ehrhardt, RA, Bell, AW, Van Amburgh, ME & Boisclair, YR 2001, 'Decreased concentration of plasma leptin in periparturient dairy cows is caused by negative energy balance', Journal of Endocrinology, vol. 171, no. 2, pp. 339-48.
Boediyana, T 2007, 'Kesiapan dan Peran Asosiasi Industri Ternak Menuju Swasembada Daging Sapi 2010 (Roles of Livestock Industries Association to Support

118
National Beef Self Sufficiency Program 2010)', paper presented to World Food Day National Seminar, Bogor - Indonesia.
Bolanos, JM, Forsberg, M, Kindahl, H & Rodriguez-Martinez, H 1998, 'Biostimulatory effects of estrous cows and bulls on resumption of ovarian activity in postpartum anestrous Zebu (Bos indicus) cows in the humid tropics', Theriogenology, vol. 49, no. 3, pp. 629-36.
Bornstein, SR, Uhlmann, K, Haidan, A, EhrhartBornstein, M & Scherbaum, WA 1997, 'Evidence for a novel peripheral action of leptin as a metabolic signal to the adrenal gland - Leptin inhibits cortisol release directly', Diabetes, vol. 46, no. 7, pp. 1235-8.
Bossis, I, Wettemann, RP, Welty, SD, Vizcarra, J & Spicer, LJ 2000, 'Nutritionally induced anovulation in beef heifers: Ovarian and endocrine function during realimentation and resumption of ovulation', Biology of Reproduction, vol. 62, no. 5, pp. 1436-44.
Brito, LFC, Silva, A, Rodrigues, LH, Vieira, FV, Deragon, LAG & Kastelic, JP 2002, 'Effect of age and genetic group on characteristics of the scrotum, testes and testicular vascular cones, and on sperm production and semen quality in AI bulls in Brazil', Theriogenology, vol. 58, no. 6, pp. 1175-86.
Bullock, KD, Bertrand, JK, Benyshek, LL, Williams, SE & Lust, DG 1991, 'Comparison of real-time ultrasound and other live measures to carcass measures as predictors of beef cow energy stores', Journal of animal science, vol. 69, no. 10, pp. 3908-16.
Burns, BM, Fordyce, G & Holroyd, RG 2010, 'A review of factors that impact on the capacity of beef cattle females to conceive, maintain a pregnancy and wean a calf-Implications for reproductive efficiency in northern Australia', Animal Reproduction Science, vol. 122, no. 1-2, pp. 1-22.
Burrow, HM 2001, 'Variances and covariances between productive and adaptive traits and temperament in a composite breed of tropical beef cattle', Livestock Production Science, vol. 70, no. 3, pp. 213-33.

119
Burton, JL, McBride, BW, Block, E, Glimm, DR & Kennelly, JJ 1994, 'A review of bovine growth-hormone', Canadian Journal of Animal Science, vol. 74, no. 2, pp. 167-201.
Butler, WR, Calaman, JJ & Beam, SW 1996, 'Plasma and milk urea nitrogen in relation to pregnancy rate in lactating dairy cattle', Journal of animal science, vol. 74, no. 4, pp. 858-65.
Byerley, DJ, Staigmiller, RB, Berardinelli, JG & Short, RE 1987, 'Pregnancy rates of beef heifers bred either on puberal or 3rd estrus', Journal of animal science, vol. 65,
no. 3, pp. 645-50.
Campbell, MH & Miller, JK 1998, 'Effect of supplemental dietary vitamin E and zinc on reproductive performance of dairy cows and heifers fed excess iron', Journal of Dairy Science, vol. 81, no. 10, pp. 2693-9.
Canfield, RW & Butler, WR 1989, 'Accuracy of predicting the LH surge and optimal insemination time in Holstein heifers using a vaginal resistance probe', Theriogenology, vol. 31, no. 4, pp. 835-42.
Canfield, RW & Butler, WR 1991, 'Energy-balance, 1st ovulation and the effects of naloxone on LH-secretion in early postpartum dairy-cows', Journal of animal science, vol. 69, no. 2, pp. 740-6.
Cassady, JM, Maddock, TD, DiCostanzo, A & Lamb, GC 2009, 'Initial body condition score affects hormone and metabolite response to nutritional restriction and repletion in yearling postpubertal beef heifers', Journal of animal science, vol. 87, no. 7, pp. 2262-73.
Cavalieri, J, Rubio, I, Kinder, JE, Entwistle, KW & Fitzpatrick, LA 1997, 'Synchronization of estrus and ovulation and associated endocrine changes in Bos indicus cows', Theriogenology, vol. 47, no. 4, pp. 801-14.
Chamdi, AR 2004, 'Karakteristik Sumberdaya Genetik Ternak Sapi Bali (Bos-bibos banteng) dan Alternatif Pola Konservasinya (The Characteristics of Genetic Resource of Bali Cattle (Bos-bibos banteng) and the Alternative of It's Conservation Methods)', Biodiversitas, vol. 6, no. 1, pp. 70-5.

120
Chandler, JB 2001, 'Effect of Concentrations of Progesterone on Follicular Development and Fertility in Beef Cows', Master of Science in Reproductive Physiology thesis, West Virginia University.
Chelikani, PK, Keisler, DH & Kennelly, JJ 2003, 'Response of plasma leptin concentration to jugular infusion of glucose or lipid is dependent on the stage of lactation of Holstein cows', The Journal of nutrition, vol. 133, no. 12, pp. 4163-71.
Chenoweth, PJ 1994, 'Aspects of reproduction in female bos-indicus cattle - A review', Australian Veterinary Journal, vol. 71, no. 12, pp. 422-6.
Chilliard, Y, Bocquier, F & Doreau, M 1998, 'Digestive and metabolic adaptations of ruminants to undernutrition, and consequences on reproduction', Reproduction Nutrition Development, vol. 38, no. 2, pp. 131-52.
Chilliard, Y, Bonnet, M, Delavaud, C, Faulconnier, Y, Leroux, C, Djiane, J & Bocquier, F 2001, 'Leptin in ruminants. Gene expression in adipose tissue and mammary gland, and regulation of plasma concentration', Domestic animal endocrinology, vol. 21, no. 4, pp. 271-95.
Copland, RS 1974, 'Observations on Banteng cattle in Sabah', Tropical animal health and production, vol. 6, no. 2, pp. 89-94.
Corah, LR & Ives, S 1991, 'The effects of essential trace minerals on reproduction in beef-cattle', Veterinary Clinics of North America-Food Animal Practice, vol. 7, no. 1, pp. 41-57.
Cortes, R, Orihuela, JA & Galina, CS 1999, 'Effect of sexual partners on the oestrous behaviour response in Zebu cattle (Bos indicus) following synchronisation with a progestagen (Synchro-mate B)', Asian-Australasian journal of animal sciences, vol. 12, no. 4, pp. 515-9.
Coulter, GH, Rounsaville, TR & Foote, RH 1976, 'Heritability of testicular size and consistency in Holstein bulls', Journal of Animal Science, vol. 43, no. 1, pp. 9-12.

121
Crowe, MA 2008a, 'Resumption of Ovarian Cyclicity in Post-partum Beef and Dairy Cows', Reproduction in Domestic Animals, vol. 43, pp. 20-8.
Crowe, MA 2008b, 'Resumption of Ovarian Cyclicity in Post‐partum Beef and Dairy Cows', Reproduction in Domestic Animals, vol. 43, no. s5, pp. 20-8.
Crowe, MA & Mullen, MP 2013, Regulation and Function of Gonadotropins Throughout the Bovine Oestrous Cycle, INTECH Open Access Publisher.
Curlewis, J, Axelson, M & Stone, G 1985, 'Identification of the major steroids in ovarian and adrenal venous plasma of the brush-tail possum (Trichosurus vulpecula) and changes in the peripheral plasma levels of oestradiol and progesterone during the reproductive cycle', Journal of Endocrinology, vol. 105, no. 1, pp. 53-62.
Damptey, J, Obese, F, Aboagye, G & Ayizanga, R 2013, 'Correlations among concentrations of some metabolic hormones and nutritionally-related metabolites in beef cows', Online J Anim Feed Res, vol. 3, pp. 176-80.
Darmadja, SGND 1980, 'Setengah Abad Peternakan Sapi Tradisional dalam Ekosistem Pertanian di Bali.', Universitas Padjadjaran.
Dawuda, PM, Scaife, JR, Hutchinson, JSM & Sinclair, KD 2002, 'Mechanisms linking under-nutrition and ovarian function in beef heifers', Animal Reproduction Science, vol. 74, no. 1-2, pp. 11-26.
De La Hoya, MPG, Ramirez, JAT, Conteras, PR, Diaz, CE, Saucedo, FOR & Saucedo, JSQ 2015, 'Role of Leptin in Cattle Production: Review', Journal of Animal and Veterinary Advances, vol. 14, no. 4, pp. 81-90.
De Rensis, F & Scaramuzzi, RJ 2003, 'Heat stress and seasonal effects on reproduction in the dairy cow - a review', Theriogenology, vol. 60, no. 6, pp. 1139-51.

122
De Silva, AWMV, Anderson, GW, Gwazdauskas, FC, Lineweaver, JA & McGilliard, ML 1981, 'Interrelationships With Estrous Behavior and Conception in Dairy Cattle', Journal of Dairy Science, vol. 64, no. 12, pp. 2409-18.
Delgado, R, Magana, JG, Galina, C & Segura, JC 2004, 'Effect of body condition at calving and its changes during early lactation on postpartum reproductive performance of Zebu cows in a tropical environment', Journal of Applied Animal Research, vol. 26, no. 1, pp. 23-8.
DeRouen, SM, Franke, DE, Morrison, DG, Wyatt, WE, Coombs, DF, White, TW, Humes, PE & Greene, BB 1994, 'Prepartum body condition and weight influences on reproductive-performance of first-calf beef-cows', Journal of animal science, vol. 72, no. 5, pp. 1119-25.
Desaulniers, DM, Goff, AK, Betteridge, KJ, Rowell, JE & Flood, PF 1989, 'Reproductive hormone concentrations in faeces during the oestrous cycle and pregnancy in cattle ( Bos taurus ) and muskoxen ( Ovibos moschatus )', Canadian Journal of Zoology, vol. 67, no. 5, pp. 1148-54.
Devendra, C, Choo, TLK & Pathmasingam, M 1973, 'The productivity of Bali cattle in Malaysia', Malaysian Agricultural Journal, vol. 49, no. 2, pp. 183-97.
Devesa, J, Lois, N, Arce, V, Diaz, MJ, Lima, L & Tresguerres, JAF 1991, 'The role of sexual steroids in the modulation of growth hormone (GH) secretion in humans', Journal of Steroid Biochemistry and Molecular Biology, vol. 40, no. 1-3, pp. 165-73.
DGLVS 2010, 'Strategic Plan of the Directorate General for Livestock and Veterinary Services Indonesia 2010-2014 Jakarta: Directorate General for Livestock and Veterinary Services, Ministry of Agriculture, Republic of Indonesia'.
DGLVS 2012, 'Statistik Peternakan (Livestock Statistics) Retrieved 3 April, 2012, from http://ditjennak.deptan.go.id/index.php?page=statistikpeternakan&action=info'.
Dieleman, SJ, Bevers, MM, Vantol, HTM & Willemse, AH 1986, 'Peripheral plasma-concentrations of estradiol, progesterone, cortisol, LH and prolactin during the estrous-

123
cycle in the cow, with emphasis on peri-estrous period', Animal Reproduction Science, vol. 10, no. 4, pp. 275-92.
Diskin, MG 2008, 'HeatWatch: a telemetric system for heat detection in cattle', Vet. Q., vol. 30, pp. 37-48.
Diskin, MG, Mackey, DR, Roche, JF & Sreenan, JM 2003, 'Effects of nutrition and metabolic status on circulating hormones and ovarian follicle development in cattle', Animal Reproduction Science, vol. 78, no. 3, pp. 345-70.
Diskin, MG & Sreenan, JM 2000, 'Expression and detection of oestrus in cattle', Reproduction Nutrition Development, vol. 40, no. 5, pp. 481-91.
Diwyanto, K 2003, Strategies to Improve Bali Cattle in Eastern Indonesia - Proceedings of a workshop 4-7 February 2002, Bali, Indonesia - Introduction and opening address, Strategies to Improve Bali Cattle in Eastern Indonesia.
Diwyanto, K 2008, 'Pemanfaatan Sumber Daya Lokal dan Inovasi Teknologi dalam mendukung Pengembangan Sapi Potong di Indonesia', Pengembangan Inovasi Pertanian, vol. 1, no. 3, pp. 173-88.
Dobson, H & Kamonpatana, M 1986, 'A review of female cattle reproduction with special reference to a comparison between buffaloes, cows and zebu', Journal of reproduction and fertility, vol. 77, no. 1, pp. 1-36.
Dobson, H & Smith, RF 2000, 'What is stress, and how does it affect reproduction?', Animal Reproduction Science, vol. 60, pp. 743-52.
Douglas, M 2015, 'Early lactation milk production and energy partitioning in primiparous Holstein-Friesian cows with high or low milk protein concentrations in a seasonally-calving, pasture-based herd', 2015SYMPOSIUM, p. 81.
Duby, R & Prange, R 1996, 'Physiology and endocrinology of the estrous cycle', Dairy Integrated Reproductive Management. University of Massachusetts. IRM-2.

124
Dunn, TG & Moss, GE 1992, 'Effects of nutrient deficiencies and excesses on reproductive efficiency of livestock', Journal of animal science, vol. 70, no. 5, pp. 1580-93.
Duricic, D, Grizelj, J, Dobranic, T, Harapin, I, Vince, S, Kocila, P, Folnozic, I, Lipar, M, Gracner, GG & Samardzija, M 2012, 'Reproductive performance of Boer goats in a moderate climate zone', Veterinarski Arhiv, vol. 82, no. 4, pp. 351-8.
Dziuk, PJ & Bellows, RA 1983, 'Management of reproduction of beef-cattle, sheep and pigs', Journal of animal science, vol. 57, pp. 355-79.
Echternkamp, SE 1984, 'Relationship between LH and cortisol in acutely stressed beef-cows', Theriogenology, vol. 22, no. 3, pp. 305-11.
Edmonson, AJ, Lean, IJ, Weaver, LD, Farver, T & Webster, G 1989, 'A Body Condition Scoring Chart for Holstein Dairy-Cows', Journal of Dairy Science, vol. 72, no. 1, pp. 68-78.
Einspanier, R, Miyamoto, A, Schams, D, Müller, M & Brem, G 1990, 'Tissue concentration, mRNA expression and stimulation of IGF-I in luteal tissue during the oestrous cycle and pregnancy of cows', Journal of reproduction and fertility, vol. 90, no. 2, pp. 439-45.
El-Wishy, AB 2007, 'The postpartum buffalo II. Acyclicity and anestrus', Animal Reproduction Science, vol. 97, no. 3-4, pp. 216-36.
Elsasser, TH, Rumsey, TS & Hammond, AC 1989, 'Influence of diet on basal and growth hormone-stimulated plasma concentrations of IGF-I in beef cattle', Journal of animal science, vol. 67, no. 1, pp. 128-41.
Entwistle, K, Talib, C, Siregar, AR, Budiarti, S, Turner, W & Lidndsay, D 2001, 'Bali cattle performance: Current population dynamics and performance and some strategies for improvement (A preliminary report)', in Seminar Nasional Teknologi Peternakan dan Veteriner, Bogor, Indonesia.

125
Entwistle, KW 1983, Factors influencing reproduction in beef cattle in Australia, AMRC.
Everitt, GC 1978, 'Cattle production in Indonesia. Background to an overseas aid assignment', New Zealand Agricultural Science, vol. 12, no. 3, pp. 81-8.
Ezanno, P, Ickowicz, A & Bocquier, F 2003, 'Factors affecting the body condition score of N'Dama cows under extensive range management in Southern Senegal', Animal Research, vol. 52, no. 1, pp. 37-48.
Ezov, N, Maltz, E, Yarom, R, Lewis, GS, Schindler, D, Ron, M, Aizinbud, E & Lehrer, AR 1990, 'Cell-density, fluid volume and electrolyte content of bovine vulvar tissue during oestrus and dioestrus', Animal Reproduction Science, vol. 22, no. 4, pp. 281-8.
Fajersson, P, Barradas, HV, Romanponce, H & Cook, RM 1991, 'The effects of dietary protein on age and weight at the onset of puberty in Brown Swiss and Zebu heifers in the tropics', Theriogenology, vol. 35, no. 4, pp. 845-62.
Fallon, GR & Crofts, JM 1959, 'Some aspects of oestrus in cattle, with reference to fertility on artificial insemination. 2. Crystallisation patterns in cervico-vaginal mucus', Queensland Journal of Agricultural Science, vol. 16, pp. 431-7.
Fatet, A, Pellicer-Rubio, M-T & Leboeuf, B 2011, 'Reproductive cycle of goats', Animal Reproduction Science, vol. 124, no. 3-4, pp. 211-9.
Ferguson, JD, Blanchard, T, Galligan, DT, Hoshall, DC & Chalupa, W 1988, 'Infertility in dairy-cattle fed a high percentage of protein degradable in the rumen.', Journal of the American Veterinary Medical Association, vol. 192, no. 5, pp. 659-62.
Ferguson, JD, Galligan, DT, Blanchard, T & Reeves, M 1993, 'Serum urea nitrogen and conception rate - The usefulness of test information.', Journal of Dairy Science, vol. 76, no. 12, pp. 3742-6.

126
Fitko, R, Kucharski, J, Szlezyngier, B & Jana, B 1996, 'The concentration of GnRH in hypothalamus, LH and FSH in pituitary, LH, PRL and sex steroids in peripheral and ovarian venous plasma of hypo- and hyperthyroid, cysts-bearing gilts', Animal Reproduction Science, vol. 45, no. 1-2, pp. 123-38.
Fitzpatrick, LA 1994, 'Advances in the understanding of post-partum anoestrus in Bos indicus cows', Strengthening research on animal reproduction and disease diagnosis in Asia through the application of immunoassay techniques: Proceedings of the Final Research Co-ordination Meeting, Bangkok, Thailand, 1-5 February 1993., pp. 19-35.
Foote, R, Oltenacu, E, Mellinger, J, Scott, N & Marshall, R 1979, 'Pregnancy rate in dairy cows inseminated on the basis of electronic probe measurements', Journal of Dairy Science, vol. 62, no. 1, pp. 69-73.
Forde, N, Beltman, M, Lonergan, P, Diskin, M, Roche, J & Crowe, M 2011a, 'Oestrous cycles in Bos taurus cattle', Animal Reproduction Science, vol. 124, no. 3, pp. 163-9.
Forde, N, Beltman, ME, Lonergan, P, Diskin, M, Roche, JF & Crowe, MA 2011b, 'Oestrous cycles in Bos taurus cattle', Animal Reproduction Science, vol. 124, no. 3-4, pp. 163-9.
Fordyce, G, Fitzpatrick, LA, Cooper, NJ, Doogan, VJ, De Faveri, J & Holroyd, RG 2002, 'Bull selection and use in northern Australia. 5. Social behaviour and management', Animal Reproduction Science, vol. 71, no. 1-2, pp. 81-99.
Fordyce, G, Fitzpatrick, LA, Mullins, TJ, Cooper, NJ, Reid, DJ & Entwistle, KW 1997, 'Prepartum supplementation effects on growth and fertility in Bos indicus-cross cows', Australian Journal of Experimental Agriculture, vol. 37, no. 2, pp. 141-9.
Fordyce, G, Panjaitan, T, Julianto, T, Kurtz, E & Poppi, D 2010, 'Breeding Bos javanicus d’Alton cattle in Eastern Indonesia: Cattle reproduction', in The 5th International Seminar on Tropical Animal Production, Yogyakarta, pp. 19-22.
Fordyce, G, Panjaitan, TA, Poppi, M & Poppi, D 2003, 'Management to facilitate genetic improvement of Bali cattle in Eastern Indonesia', in K Entwistle & DR Lindsay

127
(eds), ACIAR PROCEEDINGS: Strategies to Improve Bali Cattle in Eastern Indonesia, pp. 23-8.
Fortune, JE 1993, 'Follicular dynamics during the bovine estrous-cycle - A limiting factor in improvement of fertility', Animal Reproduction Science, vol. 33, no. 1-4, pp. 111-25.
Fortune, JE 1994, 'Ovarian follicular-growth and development in mammals', Biology of Reproduction, vol. 50, no. 2, pp. 225-32.
Galina, CS, Calderon, A & McCloskey, M 1982, 'Detection of signs of estrus in the Charolais cow and its Brahman cross under continuous observation', Theriogenology, vol. 17, no. 5, pp. 485-98.
Galina, CS, Orihuela, A & Duchateau, A 1987, 'Reproductive physiology in Zebu cattle: unique reproductive aspects that affect their performance', Veterinary Clinics of North America: Food Animal Practice, vol. 3, no. 3, pp. 619-32.
Ganswindt, A, Brown, JL, Freeman, EW, Kouba, AJ, Penfold, LM, Santymire, RM, Vick, MM, Wielebnowski, N, Willis, EL & Milnes, MR 2012, 'International Society for Wildlife Endocrinology: the future of endocrine measures for reproductive science, animal welfare and conservation biology', Biology Letters, vol. 8, no. 5, pp. 695-7.
Garcia, M, Huanca, W & Echevarria, L 1990, 'Reproductive-performance of purebred and crossbred Zebu cattle under artificial-insemination in the Amazon tropics', Animal Production, vol. 50, pp. 41-9.
Garcia, MR, Amstalden, M, Williams, SW, Stanko, RL, Morrison, CD, Keisler, DH, Nizielski, SE & Williams, GL 2002, 'Serum leptin and its adipose gene expression during pubertal development, the estrous cycle, and different seasons in cattle', Journal of animal science, vol. 80, no. 8, pp. 2158-67.
Gartland, P, Schiavo, J, Hall, C, Foote, R & Scott, N 1976, 'Detection of estrus in dairy cows by electrical measurements of vaginal mucus and by milk progesterone', Journal of Dairy Science, vol. 59, no. 5, pp. 982-5.

128
Gearhart, MA, Curtis, CR, Erb, HN, Smith, RD, Sniffen, CJ, Chase, LE & Cooper, MD 1990, 'Relationship of Changes in Condition Score to Cow Health in Holsteins', Journal of Dairy Science, vol. 73, no. 11, pp. 3132-40.
Gilmore, HS, Young, FJ, Patterson, DC, Wylie, ARG, Law, RA, Kilpatrick, DJ, Elliott, CT & Mayne, CS 2011, 'An evaluation of the effect of altering nutrition and nutritional strategies in early lactation on reproductive performance and estrous behavior of high-yielding Holstein-Friesian dairy cows', Journal of Dairy Science, vol. 94, no. 7, pp.
3510-26.
Ginther, OJ, Knopf, L & Kastelic, JP 1989, 'Ovarian follicular dynamics in heifers during early pregnancy', Biology of Reproduction, vol. 41, no. 2, pp. 247-54.
Gombe, S & Hansel, W 1973, 'Plasma Luteinizing Hormone (LH) and Progesterone Levels in Heifers on Restricted Energy Intakes', Journal of animal science, vol. 37, no. 3, pp. 728-33.
Goodman, RL & Karsch, FJ 1980, 'Pulsatile secretion of luteinizing-hormone - differential suppression by ovarian-steroids', Endocrinology, vol. 107, no. 5, pp. 1286-90.
Goodman, RL, Legan, SJ, Ryan, KD, Foster, DL & Karsch, FJ 1981, 'Importance of variations in behavioral and feedback actions of estradiol to the control of seasonal breeding in the ewe', Journal of Endocrinology, vol. 89, no. 2, pp. 229-40.
Goto, Y, Noda, Y, Narimoto, K, Umaoka, Y & Mori, T 1992, 'Oxidative stress on mouse embryonic development in vitro.', Free Radical Biology and Medicine, vol. 13, no. 1, pp. 47-53.
Graham, L, Raeside, J, Goodrowe, K & Liptrap, R 1992, 'Measurements of faecal oestradiol and progesterone in non-pregnant and pregnant domestic and exotic cats', Journal of reproduction and fertility. Supplement, vol. 47, pp. 119-20.

129
Graham, LH 2004, 'Non-Invasive Monitoring of Reproduction in Zoo and Wildlife Species', ARBS Annual Review of Biomedical Sciences, vol. 6, pp. 91-8.
Grings, EE, Staigmiller, RB, Short, RE, Bellows, RA & MacNeil, MD 1999, 'Effects of stair-step nutrition and trace mineral supplementation on attainment of puberty in beef heifers of three sire breeds', Journal of animal science, vol. 77, no. 4, pp. 810-5.
Gupta, KA & Purohit, GN 2001, 'Use of vaginal electrical resistance (VER) to predict estrus and ovarian activity, its relationship with plasma progesterone and its use for insemination in buffaloes', Theriogenology, vol. 56, no. 2, pp. 235-45.
Hadi, PU, Ilham, N, Thahar, A, Winarso, B, Vincent, D & Quirke, D 2002, 'Improving Indonesia's beef industry. ACIAR Monograph. Canberra: Australian Center for International Agriculture Research (ACIAR).'.
Halaas, JL, Gajiwala, KS, Maffei, M, Cohen, SL, Chait, BT, Rabinowitz, D, Lallone, RL, Burley, SK & Friedman, JM 1995, 'Weight-reducing effects of the plasma-protein encoded by the obese gene', Science, vol. 269, no. 5223, pp. 543-6.
Handiwirawan, E & Subandriyo 2004, 'Potensi dan Keragaman Sumberdaya Genetik Sapi Bali (Potency and genetic diversity of Bali cattle)', Wartazoa, vol. 14, no. 3, pp. 107-15.
Harrison, JH, Hancock, DD & Conrad, HR 1984, 'Vitamin-E and selenium for reproduction of the dairy-cow.', Journal of Dairy Science, vol. 67, no. 1, pp. 123-32.
Hattab, SA, Kadoom, AK, Palme, R & Bamberg, E 2000, 'Effect of crestar ™ on estrus synchronization and the relationship between fecal and plasma concentrations of progestagens in buffalo cows', Theriogenology, vol. 54, no. 7, pp. 1007-17.
Heckman, G, Katz, L, Foote, R, Oltenacu, E, Scott, N & Marshall, R 1979, 'Estrous cycle patterns in cattle monitored by electrical resistance and milk progesterone', Journal of Dairy Science, vol. 62, no. 1, pp. 64-8.

130
Heersche, G & Nebel, RL 1994, 'Measuring Efficiency and Accuracy of Detection of Estrus', Journal of Dairy Science, vol. 77, no. 9, pp. 2754-61.
Heistermann, M, Agil, M, Buthe, A & Hodges, JK 1998, 'Metabolism and excretion of oestradiol-17 beta and progesterone in the Sumatran rhinoceros (Dicerorhinus sumatrensis)', Animal Reproduction Science, vol. 53, no. 1-4, pp. 157-72.
Hemler, ME & Lands, WEM 1980, 'Evidence of peroxide-initiated free radical mechanism of prostaglandin biosynthesis.', Journal of Biological Chemistry, vol. 255,
no. 13, pp. 6253-61.
Henricks, DM, Dickey, JF & Niswender, GD 1970, 'Serum Luteinizing Hormone and Plasma Progesterone Levels During the Estrous Cycle and Early Pregnancy in Cows', Biology of Reproduction, vol. 2, no. 3, pp. 346-51.
Herbison, AE & Pape, JR 2001, 'New evidence for estrogen receptors in gonadotropin-releasing hormone neurons', Frontiers in Neuroendocrinology, vol. 22, no. 4, pp. 292-308.
Heuer, C, Schukken, YH & Dobbelaar, P 1999, 'Postpartum body condition score and results from the first test day milk as predictors of disease, fertility, yield, and culling in commercial dairy herds', Journal of Dairy Science, vol. 82, no. 2, pp. 295-304.
Hidiroglou, M 1979, 'Trace-element deficiencies and fertility in ruminants - Review.', Journal of Dairy Science, vol. 62, no. 8, pp. 1195-206.
Ho, KKY & Hoffman, DM 1993, 'Aging and growth-hormone', Hormone Research, vol. 40, no. 1-3, pp. 80-6.
Hockett, ME, Hopkins, FM, Lewis, MJ, Saxton, AM, Dowlen, HH, Oliver, SP & Schrick, FN 2000, 'Endocrine profiles of dairy cows following experimentally induced clinical mastitis during early lactation', Animal Reproduction Science, vol. 58, no. 3-4, pp. 241-51.

131
Hockey, CD, Norman, ST, Morton, JM, Boothby, D, Phillips, NJ & McGowan, MR 2010, 'Use of Vaginal Electrical Resistance to Diagnose Oestrus, Dioestrus and Early Pregnancy in Synchronized Tropically Adapted Beef Heifers', Reproduction in Domestic Animals, vol. 45, no. 4, pp. 629-36.
Hodges, J 1997, 'World watch list for domestic animal diversity. (1995) Second edition. Separate editions in English, French and Spanish. Ed.: Beate D. Scherf. Published by FAO and UNEP. FAO, Rome, Italy. ISBN 92-5-103729-9. 769 pp', Livestock Production Science, vol. 49, no. 1, p. 88.
Hoedemaker, M, Prange, D & Gundelach, Y 2008, 'Relationship between BCS pattern antepartum and metabolic parameters, fertility and health in dairy cows', Tieraerztliche Umschau, vol. 63, no. 6, pp. 299-+.
Hoedemaker, M, Prange, D & Gundelach, Y 2009, 'Body Condition Change Ante- and Postpartum, Health and Reproductive Performance in German Holstein Cows', Reproduction in Domestic Animals, vol. 44, no. 2, pp. 167-73.
Hopper, HW, Silcox, RW, Byerley, DJ & Kiser, TE 1993, 'Follicular development in prepubertal heifers', Animal Reproduction Science, vol. 31, no. 1-2, pp. 7-12.
Hoshino, S, Wakita, M, Kobayashi, Y, Sakauchi, R, Nishiguchi, Y, Ozawa, A, Hodate, K, Hamaguchi, I & Yotani, Y 1991, 'Variations in serum levels of insulin-like growth factor-1, growth hormone and thyroid hormones during lactation in dairy cows', Comparative Biochemistry and Physiology -- Part A: Physiology, vol. 99, no. 1, pp. 61-4.
Houghton, PL, Lemenager, RP, Horstman, LA, Hendrix, KS & Moss, GE 1990, 'Effects of body-composition, prepartum and postpartum energy-level and early weaning on reproductive-performance of beef-cows and preweaning calf gain', Journal of animal science, vol. 68, no. 5, pp. 1438-46.
Houseknecht, KL, Baile, CA, Matteri, RL & Spurlock, ME 1998, 'The biology of leptin: A review', Journal of animal science, vol. 76, no. 5, pp. 1405-20.

132
Houseknecht, KL & Portocarrero, CP 1998, 'Leptin and its receptors: Regulators of whole-body energy homeostasis', Domestic animal endocrinology, vol. 15, no. 6, pp. 457-75.
Houseknecht, KL, Portocarrero, CP, Ji, S, Lemenager, R & Spurlock, ME 2000, 'Growth hormone regulates leptin gene expression in bovine adipose tissue: correlation with adipose IGF-1 expression', Journal of Endocrinology, vol. 164, no. 1, pp. 51-7.
Hunter, GL, Belonje, PC & Van Niekerk, CH 1971, 'Synchronized mating and lambing in spring bred merino sheep the use of progestogen impregnated intra vaginal sponges and teaser rams', Agroanimalia, vol. 3, no. 4, pp. 133-40.
Hurley, WL & Doane, RM 1989, 'Recent Developments in the Roles of Vitamins and Minerals in Reproduction', Journal of Dairy Science, vol. 72, no. 3, pp. 784-804.
Hurnik, JF, King, GJ & Robertson, HA 1975, 'Estrous and related behaviour in postpartum Holstein cows', Applied Animal Ethology, vol. 2, no. 1, pp. 55-68.
Isle, BR, Kirsch, JD, Paasch, S, Odde, KG, Funston, RN, Lardy, GP & Vonnahme, KA 2007, 'Growth and attainment of puberty in heifers from cows supplemented with linseed meal during early lactation', Journal of animal science, vol. 85, pp. 169-.
Isobe, N, Akita, M, Nakao, I, Yamashiro, H & Kubota, H 2005, 'Pregnancy diagnosis based on the fecal progesterone concentration in beef and dairy heifers and beef cows', Animal Reproduction Science, vol. 90, no. 3-4, pp. 211-8.
Jelantik, IGN, Mullik, ML, Leo-Penu, C & Copland, R 2010, 'Factors affecting the response of Bali cattle (Bos sondaicus) calves to supplementation prior to weaning', Animal Production Science, vol. 50, no. 6, pp. 493-6.
Jelantik, IGN, Mullik, ML, Leo-Penu, C, Jeremias, J & Copland, R 2008, 'Improving calf survival and performance by supplementation in Bali cattle', Australian Journal of Experimental Agriculture, vol. 48, no. 7, pp. 954-6.

133
Jeong, S-W, Yoon, S-S, Hwang, W-S, Jean, Y-H, Joo, Y-S, Moon, O-K, Kim, J-H, Lee, B-C, Chang, C-H, Jung, S-C, Jang, H, Choi, S-H & Rhee, J-C 1996, 'Investigation on the incidence status of bovine reproductive disorders in Korea', RDA Journal of Agricultural Science Veterinary, vol. 38, no. 1, pp. 825-9.
Jilek, F, Pytloun, P, Kubesova, M, Stipkova, M, Bouska, J, Volek, J, Frelich, J & Rajmon, R 2008, 'Relationships among body condition score, milk yield and reproduction in Czech Fleckvieh cows', Czech Journal of Animal Science, vol. 53, no. 9, pp. 357-67.
Johnston, DJ, Barwick, SA, Fordyce, G, Holroyd, RG, Williams, PJ, Corbet, NJ & Grant, T 2014, 'Genetics of early and lifetime annual reproductive performance in cows of two tropical beef genotypes in northern Australia', Animal Production Science, vol. 54, no. 1, pp. 1-15.
Jolly, PD, McDougall, S, Fitzpatrick, LA, Macmillan, KL & Entwistle, KW 1995, 'Physiological effects of undernutrition on postpartum anoestrus in cows', Journal of reproduction and fertility. Supplement, vol. 49, pp. 477-92.
Jolly, PD, McSweeney, CS, Schlink, AC, Houston, EM & Entwistle, KW 1996, 'Reducing post-partum anoestrous interval in first-calf Bos indicus crossbred beef heifers .3. Effect of nutrition on responses to weaning and associated variation in metabolic hormone levels', Australian Journal of Agricultural Research, vol. 47, no. 6, pp. 927-42.
Jordan, ER & Swanson, LV 1979, 'Effect of crude protein on reproductive efficiency, serum protein, and albumin in the high-producing dairy cow', Journal of Dairy Science,
vol. 62, no. 1, pp. 58-63.
Joubert, DM 1954, 'The influence of winter nutritional depressions on the growth, reproduction and production of cattle', Journal of Agricultural Science, vol. 44, no. 1, pp. 5-66.
Kadokawa, H, Matsui, M, Hayashi, K, Matsunaga, N, Kawashima, C, Shimizu, T, Kida, K & Miyamoto, A 2008, 'Peripheral administration of kisspeptin-10 increases plasma concentrations of GH as well as LH in prepubertal Holstein heifers', Journal of Endocrinology, vol. 196, no. 2, pp. 331-4.

134
Kanitz, W 2003, 'Follicular dynamic and ovulation in cattle - a review', Archiv Fur Tierzucht-Archives of Animal Breeding, vol. 46, no. 2, pp. 187-98.
Kanitz, W & Becker, F 2005, 'Are methods of oestrous synchronisation and synchronisation of oestrus and ovulation practicable and successful alternatives to the conventional heat detection and AI management?', Zuchtungskunde, vol. 77, no. 2-3, pp. 128-39.
Karsch, FJ, Dahl, GE, Evans, NP, Manning, JM, Mayfield, KP, Moenter, SM & Foster, DL 1993, 'Seasonal-changes in gonadotropin-releasing-hormone secretion in the ewe - Alteration in response to the negative feedback action of estradiol', Biology of Reproduction, vol. 49, no. 6, pp. 1377-83.
Karsch, FJ, Legan, SJ, Ryan, KD & Foster, DL 1980, 'Importance of estradiol and progesterone in regulating LH-secretion and estrous behavior during the sheep estrous-cycle', Biology of Reproduction, vol. 23, no. 2, pp. 404-13.
Kementan-BPS 2011, Rilis Hasil Akhir PSPK 2011, Kementerian Pertanian (Ministry of Agriculture of Republic of Indonesia, Badan Pusat Statistik (Beurau of Statistics of Republic of Indonesia), Jakarta.
Kempster, AJ 1981, 'Fat partition and distribution in the carcasses of cattle, sheep and pigs: A review', Meat Science, vol. 5, no. 2, pp. 83-98.
Kesler, DJ & Garverick, HA 1982, 'Ovarian cysts in dairy-cattle - A review', Journal of animal science, vol. 55, no. 5, pp. 1147-59.
Kikkawa, Y, Amano, T & Suzuki, H 1995, 'Analysis of genetic diversity of domestic cattle in east and Southeast Asia in terms of variations in restriction sites and sequences of mitochondrial DNA', Biochemical genetics, vol. 33, no. 1-2, pp. 51-60.
King, JOL 1971, 'Nutrition and fertility in dairy cows', Veterinary Record, vol. 89, no. 12, pp. 320-&.

135
Kinoshita, K, Inada, S, Seki, K, Sasaki, A, Hama, N & Kusunoki, H 2011, 'Long-Term Monitoring of Fecal Steroid Hormones in Female Snow Leopards (Panthera uncia) during Pregnancy or Pseudopregnancy', Plos One, vol. 6, no. 5.
Kirby, GWM 1979, 'Bali cattle in Australia', World Animal Review, no. 31, pp. 24-9.
Klopfenstein, T 1996, 'Need for escape protein by grazing cattle', Animal Feed Science and Technology, vol. 60, no. 3-4, pp. 191-9.
Kornmatitsuk, B, Thitaram, C & Kornmatitsuk, S 2007, 'Measurement of Faecal
Progesterone Metabolites and its Application for Early Screening of Open Cows Post‐insemination', Reproduction in Domestic Animals, vol. 42, no. 3, pp. 238-42.
Krisnamurthi, B 2011, 'Bedah Masalah dan Rancang Bangun Kebijakan Supply Demand Daging Sapi/Kerbau Indonesia, Ministry of Agriculture of the Republic of Indonesia.'.
Kugelmeier, T, do Valle, RdR, de Barros Vaz Guimaraes, MA, Pereira Carneiro Muniz, JA, Barros Monteiro, FO & de Oliveira, CA 2011, 'Tracking the Ovarian Cycle in Black-and-Gold Howlers (Alouatta caraya) by Measuring Fecal Steroids and Observing Vaginal Bleeding', International Journal of Primatology, vol. 32, no. 3, pp. 605-15.
Labhsetwar, AP 1970, 'Role of estrogens in ovulation - A study using estrogen-antagonist, ICI 46,474', Endocrinology, vol. 87, no. 3, pp. 542-&.
Laflamme, LF & Connor, ML 1992, 'Effect of postpartum nutrition and cow body condition at parturition on subsequent performance of beef-cattle', Canadian Journal of Animal Science, vol. 72, no. 4, pp. 843-51.
Lall, D, Dixit, VB, Arora, U, Kumar, B & Chauhan, TR 2000, 'Effect of mineral supplementation on reproductive performance of anestrous buffaloes under field conditions', Indian Journal of Animal Nutrition, vol. 17, no. 1, pp. 34-9.

136
Lamb, GC 1999, 'Influence of nutrition on reproduction in the beef cow herd', Univ. of Minnesota, Dept. of Animal Science, Extension Service, vol. 8.
Lammoglia, MA, Willard, ST, Hallford, DM & Randel, RD 1997, 'Effects of dietary fat on follicular development and circulating concentrations of lipids, insulin, progesterone, estradiol-17 beta, 13,14-dihydro-15-keto-prostaglandin F-2 alpha and growth hormone in estrous cyclic Brahman cows', Journal of animal science, vol. 75, no. 6, pp. 1591-600.
Larson, LL & Ball, PJH 1992, 'Regulation of estrous cycles in dairy cattle - A review', Theriogenology, vol. 38, no. 2, pp. 255-67.
Layek, SS, Mohanty, TK, Kumaresan, A, Behera, K & Chand, S 2011, 'Behavioural signs of estrus and their relationship to time of ovulation in Zebu (Sahiwal) cattle', Animal Reproduction Science, vol. 129, no. 3, pp. 140-5.
Le Thi Thanh, H, Herold, P & Zarate, AV 2010, 'Farm types for beef production and their economic success in a mountainous province of northern Vietnam', Agricultural Systems, vol. 103, no. 3, pp. 137-45.
Ledger, HP 1959, 'Possible Explanation for Part of the Difference in Heat Tolerance Exhibited by Bos-taurus, and Bos-indicus Beef Cattle', Nature, vol. 184, no. 4696, pp. 1405-6.
Legan, SJ, Karsch, FJ & Foster, DL 1977, 'Endocrine control of seasonal reproductive function in ewe - Marked change in response to negative feedback action of estradiol on luteinizing-hormone secretion', Endocrinology, vol. 101, no. 3, pp. 818-24.
Leidl, W & Stolla, R 1976, 'Measurement of electric resistance of the vaginal mucus as an aid for heat detection', Theriogenology, vol. 6, no. 2-3, pp. 237-49.
Liefers, SC, Veerkamp, RF, Pas, M, Chilliard, Y & Van der Lende, T 2005, 'Genetics and physiology of leptin in periparturient dairy cows', Domestic animal endocrinology, vol. 29, no. 1, pp. 227-38.

137
Liefers, SC, Veerkamp, RF, Te Pas, MFW, Delavaud, C, Chilliard, Y & Van der Lende, T 2003, 'Leptin concentrations in relation to energy balance, milk yield, intake, live weight, and estrus in dairy cows', Journal of Dairy Science, vol. 86, no. 3, pp. 799-807.
Lincoln, DT, Sinowatz, F, Elhifnawi, E, Hughes, RL & Waters, M 1995, 'Evidence of a direct role for growth-hormone (GH) in mammary-gland proliferation and lactation', Anatomia Histologia Embryologia-Journal of Veterinary Medicine Series C-Zentralblatt Fur Veterinarmedizin Reihe C, vol. 24, no. 2, pp. 107-15.
Lisson, S, MacLeod, N, McDonald, C, Corfield, J, Pengelly, B, Wirajaswadi, L, Rahman, R, Bahar, S, Padjung, R, Razak, N, Puspadi, K, Dahlanuddin, Sutaryono, Y, Saenong, S, Panjaitan, T, Hadiawati, L, Ash, A & Brennan, L 2010, 'A participatory, farming systems approach to improving Bali cattle production in the smallholder crop-livestock systems of Eastern Indonesia', Agricultural Systems, vol. 103, no. 7, pp. 486-97.
Lisson, S, MacLeod, N, McDonald, C, Corfield, J, Rachman, R & Wirajaswadi, L 2011, Crop-livestock farming systems in eastern Indonesia, Beef production in crop-livestock systems: simple approaches for complex problems.
Llewelyn, CA, Munro, CD, Luckins, AG, Jordt, T, Murray, M & Lorenzini, E 1987, 'Behavioural and ovarian changes during the oestrous cycle in the boran (Bos indicus)', British Veterinary Journal, vol. 143, no. 1, pp. 75-82.
Lobo, RB, Fernandes, MB, Maffei, WE, Bezerra, LAF, Lobo, MP, Gavio, D & Ribeiro, LB 2001, 'Preliminary study of ovarian perimeter in Nellore heifers of the PMGRN', Revista Brasileira de Reproducao Animal, vol. 25, no. 2, pp. 127-8.
Lopez-Gatius, F, Mirzaei, A, Santolaria, P, Bech-Sabat, G, Nogareda, C, Garcia-Ispierto, I, Hanzen, C & Yaniz, JL 2008, 'Factors affecting the response to the specific treatment of several forms of clinical anestrus in high producing dairy cows', Theriogenology, vol. 69, no. 9, pp. 1095-103.
López-Gatius, F, Yániz, J & Madriles-Helm, D 2003, 'Effects of body condition score and score change on the reproductive performance of dairy cows: a meta-analysis', Theriogenology, vol. 59, no. 3, pp. 801-12.

138
Lopez, H, Satter, LD & Wiltbank, MC 2004, 'Relationship between level of milk production and estrous behavior of lactating dairy cows', Animal Reproduction Science, vol. 81, no. 3-4, pp. 209-23.
Lowman, BG, Scott, N & Somerville, S 1973, 'Condition scoring of cattle', Bulletin, East of Scotland College of Agriculture, no. 6, pp. iv + 28pp.-iv + pp.
Lucy, MC 2008, 'Functional differences in the growth hormone and insulin-like growth factor axis in cattle and pigs: Implications for post-partum nutrition and reproduction', Reproduction in Domestic Animals, vol. 43, pp. 31-9.
Lucy, MC, Billings, HJ, Buttler, WR, Ehnis, LR, Fields, MJ, Kesler, DJ, Kinder, JE, Mattos, RC, Short, RE, Thatcher, WW, Wettemann, RP, Yelich, JV & Hafs, HD 2001a, 'Efficacy of an intravaginal progesterone insert and an injection of PGF2α for synchronizing estrus and shortening the interval to pregnancy in postpartum beef cows, peripubertal beef heifers, and dairy heifers', Journal of Animal Science, vol. 79, pp. 982-95.
Lucy, MC, Jiang, H & Kobayashi, Y 2001b, 'Changes in the Somatotrophic Axis Associated with the Initiation of Lactation', Journal of Dairy Science, vol. 84, pp. E113-E9.
Lucy, MC, Savio, JD, Badinga, L, Delasota, RL & Thatcher, WW 1992, 'Factors that affect ovarian follicular dynamics in cattle', Journal of animal science, vol. 70, no. 11, pp. 3615-26.
Lyimo, ZC, Nielen, M, Ouweltjes, W, Kruip, TAM & van Eerdenburg, F 2000, 'Relationship among estradiol, cortisol and intensity of estrous behavior in dairy cattle', Theriogenology, vol. 53, no. 9, pp. 1783-95.
Macdonald, KA, Penno, JW, Bryant, AM & Roche, JR 2005, 'Effect of feeding level pre- and post-puberty and body weight at first calving on growth, milk production, and fertility in grazing dairy cows', Journal of Dairy Science, vol. 88, no. 9, pp. 3363-75.

139
Major, S, Randhawa, SS & Sushma, C 2007, 'Manganese deficiency in dairy animals in relation to its status in soil-plant system', Indian Journal of Veterinary Medicine, vol. 27, no. 2, pp. 145-6.
Mancio, AB, Viana, JDC, Deazevedo, NA, Rehfeld, OAM, Ruas, JRM & Amaral, R 1982, 'Effects of urea and soybean supplementation on the potential reproductive-performance of zebu heifers', Arquivos Da Escola De Veterinaria Da Universidade Federal De Minas Gerais, vol. 34, no. 3, pp. 573-85.
Mann, GE, Keatinge, R, Hunter, M, Hedley, BA & Lamming, GE 2005, 'The use of milk progesterone to monitor reproductive function in beef suckler cows', Animal Reproduction Science, vol. 88, no. 3-4, pp. 169-77.
Maree, C, Jurriaanse, A & Venter, HAW 1974, 'The occurrence of postpartum anoestrus in Bonsmara cows on supplemented sourveld grazing', South African Journal of Animal Science, vol. 4, pp. 181-3.
Martin, GB, Boukhliq, R, Tjondronegoro, S, Hotzel, MJ & Fisher, JS 1992a, 'The effect of nutrition on reproductive endocrinology', Proceedings of the Nutrition Society of Australia, vol. 17, pp. 177-85.
Martin, GB, Scaramuzzi, RJ, Oldham, CM & Lindsay, DR 1983, 'Effects of Progesterone on the Responses of Merino Ewes to the Introduction of Rams during Anoestrus', Australian Journal of Biological Sciences, vol. 36, no. 4, pp. 369-78.
Martin, LC, Brinks, JS, Bourdon, RM & Cundiff, LV 1992b, 'Genetic effects on beef heifer puberty and subsequent reproduction', Journal of animal science, vol. 70, no. 12, pp. 4006-17.
Martinez, N, Lopez, S & Combellas, J 1984, 'Effect of post-partum nutrition on blood level of progesterone in dairy cows [concentrate feeds]', Colloques de l'INRA (France).
Martojo, H 2012, 'Indigenous Bali Cattle is Most Suitable for Sustainable Small Farming in Indonesia', Reproduction in Domestic Animals, vol. 47, pp. 10-4.

140
Masunda, B, Mutisi, C, Hamudikuwanda, H & Agumbah, JGO 1999, 'The concentration of faecal progestins during the oestrous cycle in Nkone cows and the effect of duration of storage of faecal samples at room temperature on faecal progestin levels', Tropical animal health and production, vol. 31, no. 6, pp. 373-81.
Masunda, B, Mutisi, C, Hamudikuwanda, H & Agumbah, JGO 2002, 'The Use of Faecal Progestin Measurements to Monitor Reproductive Activity in Mashona Cows in a Smallholder Farming Area of Zimbabwe', Tropical animal health and production, vol. 34, no. 4, pp. 309-18.
Mattoni, M, Mukasa-Mugerwa, E, Cecchini, G & Sovani, S 1988a, 'The reproductive performance of East African (Bos indicus) Zebu cattle in Ethiopia. 1. Estrous cycle length, duration, behavior and ovulation time', The reproductive performance of East African (Bos indicus) Zebu cattle in Ethiopia. 1. Estrous cycle length, duration, behavior and ovulation time, no. 5, pp. 961-71.
Mattoni, M, Mukasamugerwa, E, Cecchini, G & Sovani, S 1988b, 'The reproductive-performance of east-African (Bos-Indicus) Zebu cattle in Ethiopia. 1. Estrous cycle length, duration, behavior and ovulation time', Theriogenology, vol. 30, no. 5, pp. 961-71.
Mayberry, D, Cowley, F, Cramb, R, Poppi, D, Quigley, S, McCosker, K & Priyanti, A 2016, 'Project Improving the reproductive performance of cows and performance of fattening cattle in low input systems of Indonesia and northern Australia'.
McBride, BW, Burton, JL & Burton, JH 1988, 'The influence of bovine growth-hormone (somatotropin) on animals and their products', Research and Development in Agriculture, vol. 5, no. 1, pp. 1-21.
McCaughey, W & Patterson, A 1981, 'Vaginal electrical resistance in cows: 1. Measurements in isolated reproductive tracts', Veterinary Research Communications, vol. 5, no. 1, pp. 73-6.
McCool, C 1992, 'Buffalo and Bali cattle--exploiting their reproductive behaviour and physiology', Tropical animal health and production, vol. 24, no. 3, pp. 165-72.

141
McCormick, ME, French, DD, Brown, TF, Cuomo, GJ, Chapa, AM, Fernandez, JM, Beatty, JF & Blouin, DC 1999, 'Crude protein and rumen undegradable protein effects on reproduction and lactation performance of Holstein cows', Journal of Dairy Science, vol. 82, no. 12, pp. 2697-708.
McDougall, S, Macmillan, KL & Williamson, NB 1998, 'Factors associated with a prolonged period of postpartum anoestrum in pasture-fed dairy cattle', in Proc. World Assoc., Buiatrics, vol. 22, pp. 657-62.
McGowan, MR, McCosker, KD, Fordyce, G, Smith, DR, O'Rourke, P, Perkins, N, Barnes, T, Marquart, L, Morton, J & Newsome, T 2014, Northern Australian beef fertility project: CashCow, Final Report, Project B.NBP.0382, Meat & Livestock Australia, Sydney, Australia.
McGowan, MR, McCosker, KD, Fordyce, G, Smith, DR, Perkins, NR, O'Rourke, PK, Barnes, T, Marquart, L, Menzies, D & Newsome, T 2013, 'Factors affecting performance of beef breeding herds: findings from the Cash Cow project', in Proceedings of the Australian Cattle Veterinarians Conferences, pp. 112-6.
McKenzie, RC, Rafferty, TS & Beckett, GJ 1998, 'Selenium: An essential element for immune function', Immunology Today, vol. 19, no. 8, pp. 342-5.
McNeilly, AS 1987, 'Prolactin and the control of gonadotropin-secretion', Journal of Endocrinology, vol. 115, no. 1, pp. 1-5.
Meikle, A, Kulcsar, M, Chilliard, Y, Febel, H, Delavaud, C, Cavestany, D & Chilibroste, P 2004, 'Effects of parity and body condition at parturition on endocrine and reproductive parameters of the cow', Reproduction, vol. 127, no. 6, pp. 727-37.
Messager, S, Chatzidaki, EE, Ma, D, Hendrick, AG, Zahn, D, Dixon, J, Thresher, RR, Malinge, I, Lomet, D & Carlton, MB 2005, 'Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54', Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no. 5, pp. 1761-6.

142
Mihm, M, Baguisi, A, Boland, MP & Roche, JF 1994, 'Association between the duration of dominance of the ovulatory follicle and pregnancy rate in beef heifers', Journal of reproduction and fertility, vol. 102, no. 1, pp. 123-30.
Miller, C & Holtan, D 1996, 'Progestin analysis by gas chromatography/mass spectrometry in plasma and feces of horses, cattle, and sheep', Thesis, Oregon State University, Corvallis, OR.
Mohamad, K, Colenbrander, B, Lenstra, JA, Olsson, M, van Tol, HTA, Mikko, S, Vlamings, BH, Andersson, G, Rodríguez-Martínez, H, Purwantara, B & Paling, RW 2009, 'On the origin of Indonesian cattle', Plos One, vol. 4, no. 5, p. e5490.
Mohamad, K, Olsson, M, Andersson, G, Purwantara, B, van Tol, HTA, Rodriguez-Martinez, H, Colenbrander, B & Lenstra, JA 2012, 'The Origin of Indonesian Cattle and Conservation Genetics of the Bali Cattle Breed', Reproduction in Domestic Animals, vol. 47, pp. 18-20.
Mohamed, AA, Babiker, IA & Eltayeb, AE 2010, 'Effect of concentrate supplementation on growth and sexual development of dairy heifers', Journal of Applied Sciences Research, no. March, pp. 212-7.
Mohammed, OB, Greene, DI & Holt, WV 2011, 'Fecal progesterone metabolites and ovarian activity in cycling and pregnant mountain gazelles (Gazella gazella)', Theriogenology, vol. 75, no. 3, pp. 542-8.
Montiel, F & Ahuja, C 2005, 'Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: a review', Animal Reproduction Science, vol. 85, no. 1, pp. 1-26.
Morris, ST, Morel, PCH & Kenyon, PR 2006, 'The effect of individual liveweight and condition of beef cows on their reproductive performance and birth and weaning weights of calves', New Zealand veterinary journal, vol. 54, no. 2, pp. 96-100.
Morrow, DA 1980, The role of nutrition in dairy cattle reproduction, Current therapy in theriogenology: diagnosis, treatment and prevention of reproductive diseases in animals.

143
Mukasa-Mugerwa, E 1989, 'A review of reproductive performance of female Bos indicus (zebu) cattle', ILCA Monograph, no. 6, pp. x + 134pp.-x + pp.
Mukasa-Mugerwa, E & Tegegne, A 1989, 'Peripheral plasma progesterone concentration in Zebu (Bos-indicus) cows during pregnancy', Reproduction Nutrition Development, vol. 29, no. 3, pp. 303-8.
Munro, C & Stabenfeldt, G 1984, 'Development of a microtitre plate enzyme immunoassay for the determination of progesterone', Journal of Endocrinology, vol. 101, no. 1, pp. 41-9.
Murphy, MG, Boland, MP & Roche, JF 1990, 'Pattern of follicular-growth and resumption of ovarian activity in postpartum beef suckler cows', Journal of reproduction and fertility, vol. 90, no. 2, pp. 523-33.
Murphy, MG, Enright, WJ, Crowe, MA, McConnell, K, Spicer, LJ, Boland, MP & Roche, JF 1991, 'Effect of dietary-intake on pattern of growth of dominant follicles during the estrous-cycle in beef heifers', Journal of reproduction and fertility, vol. 92, no. 2, pp. 333-8.
Mwaanga, ES & Janowski, T 2000, 'Anoestrus in dairy cows: Causes, prevalence and clinical forms', Reproduction in Domestic Animals, vol. 35, no. 5, pp. 193-200.
Nagatani, S, Zeng, YH, Keisler, DH, Foster, DL & Jaffe, CA 2000, 'Leptin regulates pulsatile luteinizing hormone and growth hormone secretion in the sheep', Endocrinology, vol. 141, no. 11, pp. 3965-75.
Naor, Z & Catt, KJ 1981, 'Mechanism of action of gonadotropin-releasing hormone - Involvement of phospholipid turnover in luteinizing-hormone release', Journal of Biological Chemistry, vol. 256, no. 5, pp. 2226-9.
Ndlovu, T, Chimonyo, M, Okoh, AI, Muchenje, V, Dzama, K & Raats, JG 2007, 'Assessing the nutritional status of beef cattle: current practices and future prospects', African Journal of Biotechnology, vol. 6, no. 24, pp. 2727-34.

144
Niswender, G, Juengel, J, McGuire, W, Belfiore, C & Wiltbank, M 1994, 'Luteal function: the estrous cycle and early pregnancy', Biology of Reproduction, vol. 50, no. 2, pp. 239-47.
Niswender, GD, Juengel, JL, Silva, PJ, Rollyson, MK & McIntush, EW 2000, 'Mechanisms controlling the function and life span of the corpus luteum', Physiological Reviews, vol. 80, no. 1, pp. 1-29.
Nogueira, GP 2004, 'Puberty in South American Bos indicus (Zebu) cattle', Animal Reproduction Science, vol. 82, pp. 361-72.
Obese, FY, Stirling, TE, Stockdale, RC, Macmillan, KL, Egan, AR & Humphrys, S 2011, 'Effect of concentrate supplementation during early lactation on plasma IGF-I concentrations and reproductive performance of dairy cows', South African Journal of Animal Science, vol. 41, no. 4, pp. 360-7.
Oldham, CM, Cognie, Y, Poindron, P & Gayerie, P 1980, The influence of progesterone or FGA priming on the ovarian function of seasonally anovular ewes induced to ovulate by their reintroduction to teaser rams, 9th International Congress on Animal Reproduction and Artificial Insemination, 16th-20th June 1980. III. Symposia.
Olson, JD 1996, 'The role of selenium and vitamin E in mastitis and reproduction of dairy cattle', Irish Veterinary Journal, vol. 49, no. 6, pp. 362-4.
Orihuela, A 2000, 'Some factors affecting the behavioural manifestation of oestrus in cattle: a review', Applied Animal Behaviour Science, vol. 70, no. 1, pp. 1-16.
Oyedipe, EO, Osori, DIK, Akerejola, O & Saror, D 1982, 'Effect of level of nutrition on onset of puberty and conception rates of zebu heifers', Theriogenology, vol. 18, no. 5, pp. 525-39.
Palme, R 2005, 'Measuring fecal steroids - Guidelines for practical application', in U Bauchinger, W Goymann & S JenniEiermann (eds), Bird Hormones and Bird

145
Migrations: Analyzing Hormones in Droppings and Egg Yolks and Assessing Adaptations in Long-Distance Migration, vol. 1046, pp. 75-80.
Palme, R, Fischer, P, Schildorfer, H & Ismail, MN 1996, 'Excretion of infused 14C-steroid hormones via faeces and urine in domestic livestock', Animal Reproduction Science, vol. 43, no. 1, pp. 43-63.
Palme, R, Mostl, E, Brem, G, Schellander, K & Bamberg, E 1997, 'Faecal metabolites of infused C-14-progesterone in domestic livestock', Reproduction in Domestic Animals, vol. 32, no. 4, pp. 199-206.
Panjaitan, T, Fordyce, G & Poppi, D 2003, 'Bali Cattle Performance in the Dry Tropics of Sumbawa', Jurnal Ilmu Ternak dan Veteriner (JITV), vol. 8, no. 3, pp. 1-6.
Panjaitan, T, Fordyce, G & Poppi, D 2014, 'Bali Cattle Performance in the Dry Tropics of Sumbawa', JITV, vol. 19, no. 3.
Paschal, JC, Sanders, JO & Kerr, JL 1991, 'Calving and weaning characteristics of Angus-sired, Gray Brahman-sired, Gir-sired, Indu-Brazil-sired, Nellore-sired, and Red Brahman-sired F1 calves', Journal of animal science, vol. 69, no. 6, pp. 2395-402.
Patterson, DJ, Corah, LR, Brethour, JR, Spire, MF, Higgins, JJ, Kiracofe, GH, Stevenson, JS & Simms, DD 1991, 'Evaluation of reproductive traits in Bos-taurus and Bos-indicus crossbred heifers - effects of postweaning energy manipulation', Journal of animal science, vol. 69, no. 6, pp. 2349-61.
Patterson, DJ, Perry, RC, Kiracofe, GH, Bellows, RA, Staigmiller, RB & Corah, LR 1992, 'Management considerations in heifer development and puberty', Journal of animal science, vol. 70, no. 12, pp. 4018-35.
Peralta, OA, Pearson, RE & Nebel, RL 2005, 'Comparison of three estrus detection systems during summer in a large commercial dairy herd', Animal Reproduction Science, vol. 87, no. 1, pp. 59-72.

146
Perry, GA, Dalton, JC & Geary, TW 2011, 'Management factors influencing fertility in beef cattle breeding programs', in Applied Reproductive Strategies in Beef Cattle-Northwest, Northwest, Boise, ID, USA.
Perry, RC, Corah, LR, Cochran, RC, Beal, WE, Stevenson, JS, Minton, JE, Simms, DD & Brethour, JR 1991, 'Influence of dietary energy on follicular development, serum gonadotropins, and 1st postpartum ovulation in suckled beef-cows', Journal of animal science, vol. 69, no. 9, pp. 3762-73.
Peter, AT, Simon, JE, Luker, CW & Bosu, WTK 1990, 'Site of action for endotoxin-induced cortisol release in the suppression of preovulatory luteinizing-hormone surges', Theriogenology, vol. 33, no. 3, pp. 637-43.
Peters, A & Lamming, G 1986, 'Regulation of ovarian function in the post partum cow: an endocrine model', The Veterinary record, vol. 118, no. 9, pp. 236-9.
Pielecka-Fortuna, J, Chu, Z & Moenter, S 2008, 'Kisspeptin Acts Directly and Indirectly to Increase Gonadotropin-Releasing Hormone Neuron Activity and Its Effects Are Modulated by Estradiol', Endocrinology, vol. 149, no. 4, pp. 1979-86.
Pinheiro, O, Barros, C, Figueiredo, R, Do Valle, E, Encarnação, R & Padovani, C 1998a, 'Estrous behavior and the estrus-to-ovulation interval in nelore cattle (Bosindicus with natural estrus or estrus induced with prostaglandin F2α or norgestomet and estradiol valerate', Theriogenology, vol. 49, no. 3, pp. 667-81.
Pinheiro, OL 1998, 'Estrous behavior and the estrus-to-ovulation interval in nelore cattle (View the MathML source with natural estrus or estrus induced with prostaglandin F2α or norgestomet and estradiol valerate', Theriogenology, vol. 49, pp. 667-81.
Pinheiro, OL, Barros, CM, Figueiredo, RA, do Valle, ER, Encarnacao, RO & Padovani, CR 1998b, 'Estrous behavior and the estrus-to-ovulation interval in Nelore cattle (Bos indicus) with natural estrus or estrus induced with prostaglandin F2 alpha or norgestomet and estradiol valerate', Theriogenology, vol. 49, no. 3, pp. 667-81.

147
Plasse, D, Koger, M & Warnick, AC 1969, 'Reproductive behaviour of Bos indicus females in a subtropical environment. 3. Calving intervals from first exposure to conception and intervals from parturition to conception', Journal of animal science, vol. 27, pp. 105-12.
Plasse, D, Warnick, AC & Koger, M 1965, 'Age at maturity and seasonal influence on rate of ovulation in Brahman and crossbred heifers in Florida. Engl. summ.', Z Tierzuecht Zuechtungs Biol, vol. 81, no. (3), pp. 231-40.
Plasse, D, Warnick, AC & Koger, M 1970, 'Reproductive behavior of Bos-indicus females in a subtropical environment .4. Length of estrous cycle, duration of estrus, time of ovulation, fertilization and embryo survival in grade Brahman heifers', Journal of animal science, vol. 30, no. 1, pp. 63-&.
Pohan, A & Talib, C 2014, 'Aplikasi Hormone Progesterone dan Estrogen pada Betina Induk Sapi Bali Anestrus Post-Partum yang Digembalakan di Timor Barat, Nusa Tenggara Timur', JITV, vol. 19, no. 3.
Poppi, D 2009, 'Improving reproductive performance of cows and performance of fattening cattle in low input systems of Indonesia and northern Australia', ACIAR Publications.
Poppi, D, Fordyce, G, Panjaitan, T, Dahlanuddin & Quigley, S (eds) 2011, Developing an Integrated Production System for Bali Cattle in the Eastern Islands of Indonesia, vol. 145, Beef Production in Crop–Livestock Systems; Simple Approaches for Complex Problems ACIAR.
Post, TB & Reich, MM 1980, 'Puberty in tropical breeds of heifers as monitored by plasma progesterone', in Proceedings of the Australian Society of Animal Production, Australia, vol. 13, pp. 61-2.
Priyanti, A, Cramb, RA, Hanifah, VW & I.G.A.P, M 2013, 'Small-Scale Cattle Raising in East Java: A Pathway out of Poverty?', paper presented to Asia Pacific Viewpoint.
Pugh, DG, Elmore, RG & Hembree, TR 1985, 'A review of the relationship between mineral nutrition and reproduction in cattle', Bovine Practitioner, no. 20, pp. 10-3.

148
Purwantara, B, Noor, RR, Andersson, G & Rodriguez-Martinez, H 2012, 'Banteng and Bali Cattle in Indonesia: Status and Forecasts', Reproduction in Domestic Animals, vol. 47, pp. 2-6.
Rabiee, AR, Macmillan, KL & Schwarzenberger, F 2001, 'The effect of level of feed intake on progesterone clearance rate by measuring faecal progesterone metabolites in grazing dairy cows', Animal Reproduction Science, vol. 67, no. 3, pp. 205-14.
Rahman, M, Khan, M, Islam, M, Haque, M & Sarker, M 2001, 'Genetic differences in the performance of Local, Pure and Crossbred cows', Bang. J. Anim. Sci, vol. 30, no. 1-2, pp. 155-64.
Randel, RD 1990, 'Nutrition and postpartum rebreeding in cattle', Journal of animal science, vol. 68, no. 3, pp. 853-62.
Randel, RD & Welsh, TH, Jr. 2013, 'Joint Alpharma-Beef Species Symposium: Interactions of feed efficiency with beef heifer reproductive development', Journal of animal science, vol. 91, no. 3, pp. 1323-8.
Rawlings, NC 1987, 'Reproductive activity in Finnish Landrace × Rambouillet ewes exposed to various daylength cycles applied over periods of six months', Theriogenology, vol. 28, no. 1, pp. 33-42.
Reid, JT 1960, 'Effect of energy intake upon reproduction in farm animals. In: The effect of germ cell damage on animal reproduction', Jour Dairy Sci, vol. 43, no. (Suppl.), pp. 103-22.
Reynolds, WL, Urick, JJ & Knapp, BW 1990, 'Biological type effects on gestation length, calving traits and calf growth rate', Biological type effects on gestation length, calving traits and calf growth rate, vol. 68, no. 3, pp. 630-9.
Richards, MW, Spitzer, JC & Warner, MB 1986, 'Effect of varying levels of postpartum nutrition and body condition at calving on subsequent reproductive performance of beef cattle', Journal of animal science, vol. 62, no. 2, pp. 300-6.

149
Ricoy, EF, Acua, C, Rincon, RM, Cortes, DF, Bauelos-Valenzuela, R & Arechiga, CF 2001, 'True estrus determination through evaluation of serum-progesterone levels at the time of insemination of dairy cows from semi-intensive dairies in north-central Mexico', Journal of Dairy Science, vol. 84, no. Supplement 1, pp. 227-.
Roberson, MS, Wolfe, MW, Stumpf, TT, Kittok, RJ & Kinder, JE 1989, 'Luteinizing-hormone secretion and corpus-luteum function in cows receiving 2 levels of progesterone', Biology of Reproduction, vol. 41, no. 6, pp. 997-1003.
Roberts, AJ, Nugent, RA, Klindt, J & Jenkins, TG 1997, 'Circulating insulin-like growth factor I, insulin-like growth factor binding proteins, growth hormone, and resumption of estrus in postpartum cows subjected to dietary energy restriction', Journal of animal science, vol. 75, no. 7, pp. 1909-17.
Roberts, SJ 1956, 'Veterinary obstetrics and genital diseases', Veterinary obstetrics and genital diseases.
Robinson, JE & Short, RV 1977, 'Changes in breast sensitivity at puberty, during menstrual-cycle, and at parturition', British Medical Journal, vol. 1, no. 6070, pp. 1188-91.
Robinson, JJ 1990, 'Nutrition in the reproduction of farm animals', Nutrition Research Reviews, vol. 3, pp. 253-76.
Roche, JF 2006, 'The effect of nutritional management of the dairy cow on reproductive efficiency', Animal Reproduction Science, vol. 96, no. 3-4, pp. 282-96.
Roche, JF, Crowe, MA & Boland, MP 1992, 'Postpartum anestrus in dairy and beef-cows', Animal Reproduction Science, vol. 28, no. 1-4, pp. 371-8.
Roche, JF, Mackey, D & Diskin, MD 2000, 'Reproductive management of postpartum cows', Animal Reproduction Science, vol. 60, no. 1-3, pp. 703-12.

150
Ronge, H & Blum, J 1989, 'Insulin-like growth factor I responses to recombinant bovine growth hormone during feed restriction in heifers', Acta Endocrinologica, vol. 120, no. 6, pp. 735-44.
Rorie, RW, Bilby, TR & Lester, TD 2002, 'Application of electronic estrus detection technologies to reproductive management of cattle', Theriogenology, vol. 57, no. 1, pp. 137-48.
Rosnina, N, Wahid, Y, Bukar, H, Malik, MM, Yap, A, Fahmi, KC, Ganesamurthi, M, Saharee, P & Yimer, AA 2012, 'Fecal progestin extraction and analysis for non-invasive monitoring of ovarian cycle in beef cows', Pakistan Veterinary Journal, vol. 32, no. 4, pp. 584-8.
Ruegg, PL, Goodger, WJ, Holmberg, CA, Weaver, LD & Huffman, EM 1992, 'Relation among body condition score, serum urea nitrogen and cholesterol concentrations, and reproductive-performance in high-producing
Holstein dairy-cows in early lactation', American Journal of Veterinary Research, vol. 53, no. 1, pp. 10-4.
Ruegg, PL & Milton, RL 1995, 'Body Condition Scores of Holstein Cows on Prince-Edward-Island, Canada - Relationships with Yield, Reproductive-Performance, and Disease', Journal of Dairy Science, vol. 78, no. 3, pp. 552-64.
Rutter, LM & Randel, RD 1984, 'Postpartum nutrient intake and body condition - Effect on pituitary-function and onset of estrus in beef-cattle', Journal of animal science, vol. 58, no. 2, pp. 265-74.
Saleh, N, Mahmud, E & Waded, E 2011, 'Interactions between insulin like growth factor 1, thyroid hormones and blood energy metabolites in cattle with postpartum inactive ovaries', Nat. Sci, vol. 9, no. 5, pp. 56-63.
Sariubang, M, Siregar, AR, Talib, C, Purba, M & Priadi, I 2000, Perbandingan produksi susu induk sapi bali dengan survival rate tinggi dan rendah (Comparison of milk production of Bali cows with high and low survival rate). Instalasi Penelitan dan Pengkajian Teknologi Pertanian Gowa-Makassar, Sulawesi Selatan.
Balai Penelitian Ternak, P.O. Box 221, Bogor 16002, Indonesia.

151
Pusat Penelitian dan Pengembangan Peternakan, Jalan Raya Pajajaran Kav. E59, Bogor, Indonesia.
Sauerwein, H, Miyamoto, A, Günther, J, Meyer, H & Schams, D 1992, 'Binding and action of insulin-like growth factors and insulin in bovine luteal tissue during the oestrous cycle', Journal of reproduction and fertility, vol. 96, no. 1, pp. 103-15.
Savio, JD, Thatcher, WW, Morris, GR, Entwistle, K, Drost, M & Mattiacci, MR 1993, 'Effects of induction of low plasma progesterone concentrations with a progesterone-releasing intravaginal device on follicular turnover and fertility in cattle', Journal of reproduction and fertility, vol. 98, no. 1, pp. 77-84.
Scaramuzzi, RJ, Campbell, BK, Downing, JA, Kendall, NR, Khalid, M, Munoz-Gutierrez, M & Somchit, A 2006, 'A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate', Reproduction Nutrition Development, vol. 46, no. 4, pp. 339-54.
Scaramuzzi, RJ, Tillson, SA, Thorneyc.Ih & Caldwell, BV 1971, 'Action of exogenous progesterone and estrogen on behavioral estrus and luteinizing hormone levels in ovariectomized ewe', Endocrinology, vol. 88, no. 5, pp. 1184-&.
Schams, D, Schallenberger, E, Hoffmann, B & Karg, H 1977, 'The oestrous cycle of the cow: hormonal parameters and time relationships concerning oestrus, ovulation, and electrical resistance of the vaginal mucus', Acta Endocrinologica, vol. 86, no. 1, pp. 180-92.
Schams, D, Schallenberger, E, Menzer, C, Stangl, J, Zottmeier, K, Hoffmann, B & Karg, H 1978, 'Profiles of LH, FSH and progesterone in postpartum dairy cows and their relationship to the commencement of cyclic functions', Theriogenology, vol. 10, no. 6, pp. 453-68.
Schatz, TJ & Hearnden, MN 2008, 'Heifer fertility on commercial cattle properties in the Northern Territory', vol. 48, pp. 940-4.

152
Schottler, JH & Williams, WT 1975, 'The effect of early weaning of Brahman cross calves on calf growth and reproductive performance of the dam', Australian Journal of Experimental Agriculture and Animal Husbandry, vol. 15, no. 75, pp. 456-9.
Schwarzenberger, F 2007, 'Non-invasive endocrine monitoring using fecal steroid analysis: opportunities and challenges', Revista Brasileira de Zootecnia, vol. 36, pp. 87-8.
Schwarzenberger, F, Mostl, E, Palme, R & Bamberg, E 1996, 'Faecal steroid analysis for non-invasive monitoring of reproductive status in farm, wild and zoo animals', Animal Reproduction Science, vol. 42, no. 1-4, pp. 515-26.
Segerson, EC & Libby, DW 1982, 'Ova fertilisation and sperm number per fertilised ovum for selenium and vitamin-E treated Charolais cattle', Theriogenology, vol. 17, no. 3, pp. 333-41.
Seminara, SB, Messager, S, Chatzidaki, EE, Thresher, RR, Acierno, JS, Shagoury, JK, Bo-Abbas, Y, Kuohung, W, Schwinof, KM, Hendrick, AG, Zahn, D, Dixon, J, Kaiser, UB, Slaugenhaupt, SA, Gusella, JF, O'Rahilly, S, Carlton, MBL, Crowley, WF, Aparicio, SAJR & Colledge, WH 2003, 'The GPR54 Gene as a Regulator of Puberty', The New England Journal of Medicine, vol. 349, no. 17, pp. 1614-27.
Senger, PL 2003, Pathways to pregnancy and parturition, Current Conceptions, Pullman, WA.
Sheldon, M 2004, 'The postpartum uterus', Veterinary Clinics of North America-Food Animal Practice, vol. 20, no. 3, pp. 569-91.
Short, RE & Adams, DC 1988, 'Nutritional and hormonal interrelationships in beef-cattle reproduction', Canadian Journal of Animal Science, vol. 68, no. 1, pp. 29-39.
Short, RE & Bellows, RA 1971, 'Relationships amond weight gains, age at puberty and reproductive performance in heifers', Journal of animal science, vol. 32, no. 1, pp. 127-&.

153
Short, RE, Bellows, RA, Staigmiller, RB, Berardinelli, JG & Custer, EE 1990, 'Physiological-mechanisms controlling anestrus and infertility in postpartum beef-cattle', Journal of animal science, vol. 68, no. 3, pp. 799-816.
Short, RE, Randel, RD, Staigmiller, RB & Bellows, RA 1979, 'Factors affecting estrogen-induced LH-release in the cow', Biology of Reproduction, vol. 21, no. 3, pp. 683-9.
Siegmund-Schultze, M, Rischkowsky, B, da Veiga, JB & King, JM 2007, 'Cattle are cash generating assets farms in the Eastern for mixed smallholder Amazon', Agricultural Systems, vol. 94, no. 3, pp. 738-49.
Sirois, J & Fortune, JE 1988, 'Ovarian Follicular Dynamics during the Estrous-Cycle in Heifers Monitored by Real-Time U Itrasonography', Biology of Reproduction, vol. 39, no. 2, pp. 308-17.
Sirois, J & Fortune, JE 1990, 'Lengthening the bovine estrous-cycle with low-levels of exogenous progesterone - A model for studying ovarian follicular dominance', Endocrinology, vol. 127, no. 2, pp. 916-25.
Smith, IJ, Rao, IJ, Pereira, IJ, Clarke, IJ, Caraty, IJ & Millar, IJ 2008, 'Kisspeptin is present in ovine hypophysial portal blood but does not increase during the preovulatory luteinizing hormone surge: Evidence that gonadotropes are not direct targets of kisspeptin in vivo', Endocrinology, vol. 149, no. 4, pp. 1951-9.
Smith, J, Clay, C, Caraty, A & Clarke, I 2007, 'KiSS-1 Messenger Ribonucleic Acid Expression in the Hypothalamus of the Ewe Is Regulated by Sex Steroids and Season', Endocrinology, vol. 148, no. 3, pp. 1150-7.
Smith, JF 1988, 'Influence of nutrition on ovulation rate in the ewe', Australian Journal of Biological Sciences, vol. 41, no. 1, pp. 27-36.
Smith, OB & Akinbamijo, OO 2000, 'Micronutrients and reproduction in farm animals', Animal Reproduction Science, vol. 60, pp. 549-60.

154
Soares, FS & Dryden, GM 2011, 'A Body Condition Scoring System for Bali Cattle', Asian-Australasian journal of animal sciences, vol. 24, no. 11, pp. 1587-94.
Soca, P, Carriquiry, M, Claramunt, M, Gestido, V & Meikle, A 2013, 'Metabolic and endocrine profiles of primiparous beef cows grazing native grassland. 1. Relationships between body condition score at calving and metabolic profiles during the transition period', Animal Production Science.
Soca, P, Carriquiry, M, Claramunt, M, Ruprechter, G & Meikle, A 2014, 'Metabolic and endocrine profiles of primiparous beef cows grazing native grassland. 2. Effects of body condition score at calving, type of suckling restriction and flushing on plasmatic and productive parameters', Animal Production Science, vol. 54, no. 7, pp. 862-8.
Solano, J, Orihuela, A, Galina, CS, Montiel, F & Galindo, F 2005, 'Relationships between social behaviour and mounting activity of Zebu cattle (Bos indicus)', Applied Animal Behaviour Science, vol. 94, no. 3-4, pp. 197-203.
Somchit-Assavacheep, A 2011, 'Influence of Nutritional Management on Folliculogenesis in Ewes', Thai Journal of Veterinary Medicine, vol. 41, pp. 25-9.
Sorensen, AM, Hansel, W, Hough, WH, Armstrong, DT, McEntee, K & Bratton, RW 1959, 'Causes and prevention of reproductive failures in dairy cattle. 1. Influence of underfeeding and overfeeding on growth and development of Holstein heifers', Bull. Cornell Agric. Exp. Stn., vol. 936, pp. 51 pp.- pp.
Spicer, LJ 2001, 'Leptin: a possible metabolic signal affecting reproduction', Domestic animal endocrinology, vol. 21, no. 4, pp. 251-70.
Spicer, LJ, Tucker, WB & Adams, GD 1990, 'Insulin-like growth factor-I in dairy cows: relationships among energy balance, body condition, ovarian activity, and estrous behavior', Journal of Dairy Science, vol. 73, no. 4, pp. 929-37.
Staats, DA, Lohr, DP & Colby, HD 1988, 'Effects of tocopherol depletion on the regional differences in adrenal microsomal lipid-peroxidation and steroid-metabolism.', Endocrinology, vol. 123, no. 2, pp. 975-80.

155
Stagg, K, Spicer, LJ, Sreenan, JM, Roche, JF & Diskin, MG 1998, 'Effect of calf isolation on follicular wave dynamics, gonadotropin and metabolic hormone changes, and interval to first ovulation in beef cows fed either of two energy levels postpartum', Biology of Reproduction, vol. 59, no. 4, pp. 777-83.
Stahringer, RC, Byerley, DJ, Kiser, TE & Randel, RD 1994, 'Serum luteinizing-hormone, 13,14-dihydro-15-keto-prostaglandin F2-alpha and cortisol profiles during postpartum anestrus in Brahman and Angus cows', Theriogenology, vol. 41, no. 5, pp.
1069-80.
Staples, CR, Burke, JM & Thatcher, WW 1998, 'Influence of supplemental fats on reproductive tissues and performance of lactating cows', Journal of Dairy Science, vol. 81, no. 3, pp. 856-71.
Steffan, CA, Kress, DD, Doorbos, DE & Anderson, DC 1983, 'Beef heifers that differ in milk production. 1. Age and weight at puberty', in Proceedings of the Western Section of the American Animal Science Society, vol. 34, p. 78.
Stobart, S 1994, 'Aspects of reproductive physiology in Droughtmaster heifers from High and Low fertility lines', Master of Science thesis, James Cook.
Strauch, TA, Neuendorff, DA, Brown, CG, Wade, ML, Lewis, AW, Keisler, DH & Randel, RD 2003, 'Effects of lasalocid on circulating concentrations of leptin and insulin-like growth factor-I and reproductive performance of postpartum Brahman cows', Journal of animal science, vol. 81, no. 6, pp. 1363-70.
Stroebel, A, Swanepoel, FJC, Nthakheni, ND, Nesamvuni, AE & Taylor, G 2008, 'Benefits obtained from cattle by smallholder farmers: a case study of Limpopo Province, South Africa', Australian Journal of Experimental Agriculture, vol. 48, no. 6-7, pp. 825-8.
Sukanata, IW 2008, 'Dampak kebijakan kuota perdagangan terhadap penawaran dan populasi sapi serta kesejahteraan peternak di provinsi Bali (Impact of trade quota policy on the supply, beef cattle population and farmer’s surplus in Bali province)', Magister Sains thesis, Institut Pertanian Bogor (IPB) (Bogor Agricultural University).

156
Sumantran, VN, Tsai, ML & Schwartz, J 1992, 'Growth-hormone induces C-FOS and C-JUN expression in cells with varying requirements for differentiation', Endocrinology, vol. 130, no. 4, pp. 2016-24.
Sunderland, SJ, Crowe, MA, Boland, MP, Roche, JF & Ireland, JJ 1994, 'Selection, dominance and atresia of follicles during the estrous-cycle of heifers', Journal of reproduction and fertility, vol. 101, no. 3, pp. 547-55.
Syahniar, TM, Antari, R, Pamungkas, D, Marsetyo, Mayberry, DE & Poppi, DP 2012, 'The level of tree legumes required to meet the maintenance energy requirements of Ongole (Bos indicus) cows fed rice straw in Indonesia', Animal Production Science, vol. 52, no. 6-7, pp. 641-6.
Talib, C 1998, 'Effect of season on pasture production and their relationship to the cattle productivity'.
Talib, C 2002, 'Sapi Bali di daerah sumber bibit dan peluang pengembangannya (Bali cattle in the breeding stock areas and their future development)', Wartazoa, vol. 12, no. 3, pp. 100-7.
Talib, C, Bamualim, A & Pohan, A 1998, 'Problematika pengembangan sapi Bali dalam pemeliharaan di padang penggembalaan', in Pros. Seminar Nasional Peternakan dan Veteriner, Puslitbangnak, pp. 1-2.
Talib, C, Entwistle, K, Siregar, A, Budiarti-Turner, S & Lindsay, D 2003, Survey of population and production dynamics of Bali cattle and existing breeding programs in Indonesia, Strategies to Improve Bali Cattle in Eastern Indonesia.
Taylor, VJ, Beever, DE, Bryant, MJ & Wathes, DC 2003, 'Metabolic profiles and progesterone cycles in first lactation dairy cows', Theriogenology, vol. 59, no. 7, pp. 1661-77.
Teleni, E, Campbell, RSF & Hoffmann, D 1993, Draught animal systems and management : an Indonesian study / editors: E. Teleni, R.S.F. Campbell and D.

157
Hoffmann, vol. 19, Canberra : Australian Centre for International Agricultural Research (ACIAR), Canberra.
Thompson, WR, Theuninck, DH, Meiske, JC, Goodrich, RD, Rust, JR & Byers, FM 1983, 'Linear measurements and visual appraisal as estimators of percentage empty body-fat of beef-cows', Journal of animal science, vol. 56, no. 4, pp. 755-60.
Tran, TQ, Warnick, AC, Hammond, ME & Koger, M 1988, 'Reproduction in Brahman cows calving for the first time at two or three years of age', Theriogenology, vol. 29,
no. 3, pp. 751-6.
Trinder, N, Woodhous.Cd & Renton, CP 1969, 'Effect of vitamin E and selenium on the incidence of retained placentae in dairy cows.', Veterinary Record, vol. 85, no. 20, pp. 550-&.
Vailes, LD, Washburn, SP & Britt, JH 1992, 'Effects of various steroid milieus or physiological states on sexual-behavior of Holstein cows', Journal of animal science, vol. 70, no. 7, pp. 2094-103.
Valdez, KE, Cuneo, SP, Gorden, PJ & Turzillo, AM 2005, 'The role of thecal androgen production in the regulation of estradiol biosynthesis by dominant bovine follicles during the first follicular wave', Journal of animal science, vol. 83, no. 3, pp. 597-603.
VanEerdenburg, F, Loeffler, HSH & vanVliet, JH 1996, 'Detection of oestrus in dairy cows: A new approach to an old problem', Veterinary Quarterly, vol. 18, no. 2, pp. 52-
4.
Vargas, CA, Olson, TA, Chase, CC, Hammond, AC & Elzo, MA 1999, 'Influence of frame size and body condition score on performance of Brahman cattle', Journal of animal science, vol. 77, no. 12, pp. 3140-9.
Veerkamp, RF, Beerda, B & Lende, vdT 2003, 'Effects of genetic selection for milk yield on energy balance, levels of hormones, and metabolites in lactating cattle, and possible links to reduced fertility', Livestock Production Science, vol. 83, pp. 257-75.

158
Velazquez, M, Spicer, L & Wathes, D 2008, 'The role of endocrine insulin-like growth factor-I (IGF-I) in female bovine reproduction', Domestic animal endocrinology, vol. 35, no. 4, pp. 325-42.
Venkata Naidu, G & Rao, K 2006, 'Estrus pattern and conception rate in postpartum lactating Ongole (Zebu) cows', Indian J. Anim. Reprod, vol. 27, no. 1, pp. 14-7.
Vizcarra, JA, Wettemann, RP, Spitzer, JC & Morrison, DG 1998, 'Body condition at parturition and postpartum weight gain influence luteal activity and concentrations of glucose, insulin, and nonesterified fatty acids in plasma of primiparous beef cows', Journal of animal science, vol. 76, no. 4, pp. 927-36.
Walker, ML, Wilson, ME & Gordon, TP 1984, 'Endocrine control of the seasonal occurrence of ovulation in rhesus-monkeys housed outdoors', Endocrinology, vol. 114, no. 4, pp. 1074-81.
Walters, DL & Schallenberger, E 1984, 'Pulsatile secretion of gonadotropins, ovarian-steroids and ovarian oxytocin during the periovulatory phase of the estrous-cycle in the
cow', Journal of reproduction and fertility, vol. 71, no. 2, pp. 503-12.
Waltner, SS, McNamara, JP & Hillers, JK 1993, 'Relationships of Body Condition Score to Production Variables in High Producing Holstein Dairy-Cattle', Journal of Dairy Science, vol. 76, no. 11, pp. 3410-9.
Webb, R, Garnsworthy, PC, Gong, JG & Armstrong, DG 2004, 'Control of follicular growth: local interactions and nutritional influences', Journal of animal science, vol. 82 E-Suppl, pp. E63-74.
Werre, JF 1980, 'Relationship of age at puberty in heifers to subsequent fertility and productivity', M. S. thesis thesis, Colorado State University.
Wettemann, R, Lents, C, Ciccioli, N, White, F & Rubio, I 2003, 'Nutritional-and suckling-mediated anovulation in beef cows', Journal of animal science, vol. 81, no. 14_suppl_2, pp. E48-E59.

159
Wettemann, RP, Hafs, HD, Edgerton, LA & Swanson, LV 1972, 'Estradiol and progesterone in blood serum during the bovine estrous cycle', Journal of animal science, vol. 34, no. 6, pp. 1020-4.
Whisnant, C, Kiser, T, Thompson, F & Hall, J 1985, 'Effect of nutrition on the LH response to calf removal and GnRH', Theriogenology, vol. 24, no. 5, pp. 565-73.
Wichtel, JJ, Craigie, AL, Thompson, KG & Williamson, NB 1996, 'Effect of selenium and A-tocopherol supplementation on postpartum reproductive function of dairy heifers at pasture', Theriogenology, vol. 46, no. 3, pp. 491-502.
Wilde, D 2006, 'Influence of macro and micro minerals in the peri-parturient period on fertility in dairy cattle', Animal Reproduction Science, vol. 96, no. 3-4, pp. 240-9.
Williams, GL 1990, 'Suckling as a regulator of postpartum rebreeding in cattle - A review', Journal of animal science, vol. 68, no. 3, pp. 831-52.
Williams, GL & Amstalden, M 2010, 'Understanding postpartum anestrus and puberty in the beef female', Proceedings, Applied Reproductive Strategies in Beef Cattle, vol. 28, pp. 55-71.
Williams, GL, Amstalden, M, Garcia, MR, Stanko, RL, Nizielski, SE, Morrison, CD & Keisler, DH 2002, 'Leptin and its role in the central regulation of reproduction in cattle', Domestic animal endocrinology, vol. 23, no. 1-2, pp. 339-49.
Wiltbank, JN, Gregory, KE, Swiger, LA, Ingalls, JE, Rothlisb.Ja & Koch, RM 1966, 'Effects of heterosis on age and weight at puberty in beef heifers', Journal of animal science, vol. 25, no. 3, pp. 744-&.
Wiltbank, JN, Kasson, CW & Ingalls, JE 1969, 'Puberty in crossbred and straightbred beef heifers on 2 levels of feed', Journal of animal science, vol. 29, no. 4, pp. 602-&.

160
Winaya, A, Rahayu, ID, Amin, M & Herliantin 2011, 'The genetic variation of Bali cattle ( Bos javanicus) based on sex related Y chromosome gene', Journal of Animal Production, vol. 13, no. 3, pp. 150-5.
Winter, B 2011, Beef production in crop-livestock systems: simple approaches for complex problems, Beef production in crop-livestock systems: simple approaches for complex problems.
Wintermantel, TM, Campbell, RE, Porteous, R, Bock, D, Groene, H-J, Todman, MG, Korach, KS, Greiner, E, Perez, CA, Schuetz, G & Herbison, AE 2006, 'Definition of estrogen receptor pathway critical for estrogen positive feedback to gonadotropin-releasing hormone neurons and fertility', Neuron, vol. 52, no. 2, pp. 271-80.
Winugroho, M 2002, 'Strategi pemberian pakan tambahan untuk memperbaiki efisiensi reproduksi induk sapi', Jurnal litbang pertanian, vol. 21, no. 1, pp. 19-23.
Wirdahayati, RB & Bamualim, A 1990, 'Performance production and population structure of Bali cattle in Timor, East Nusa Tenggara (Penampilan produksi dan struktur ternak sapi Bali di Pulau Timor, Nusa Tenggara Timur)', in GNR Haryana, AAB Palguna, IB Djagra, KBGN Saka, IB Mantra, IM Mastika & DKH Putra (eds), Proceedings of Bali Cattle Meeting (Proceeding seminar nasional sapi Bali’).(Ed. GNR Haryana) pp. C1–C5.(Universitas Udayana: Denpasar, Indonesia), Bali, Indonesia., pp. C1-C5.
Wiryosuhanto, S 1997, 'Bali cattle--their economic importance in Indonesia', in Jembrana disease and the bovine lentiviruses: Proceedings of a workshop 10-13 June 1996 Bali, Indonesia.
Woller, M, Tessmer, S, Neff, D, Nguema, AA, Van Roo, B & Waechter-Brulla, D 2001, 'Leptin stimulates gonadotropin releasing hormone release from cultured intact hemihypothalami and enzymatically dispersed neurons', Experimental Biology and Medicine, vol. 226, no. 6, pp. 591-6.
Wright, IA, Rhind, SM, Russel, AJF, Whyte, TK & McBean, AJ 1987, 'Effects of body condition, food-intake and temporary calf separation on the duration of the post-partum anestrous period and associated LH, FSH and prolactin concentrations in beef-cows', Animal Production, vol. 45, pp. 395-402.

161
Wright, IA, Rhind, SM, Whyte, TK & Smith, AJ 2010, 'Effects of body condition at calving and feeding level after calving on LH profiles and the duration of the postpartum anoestrous period in beef cows', Animal Production, vol. 55, no. 01, pp. 41-6.
Wright, PJ & Malmo, J 1992, 'Pharmacological manipulation of fertility', Veterinary Clinics of North America-Food Animal Practice, vol. 8, no. 1, pp. 57-89.
Yimer, N, Rosnina, Y, Wahid, H, Bukar, MM, Malik, A, Yap, KC, Fahmi, M, Ganesamurthi, P & Saharee, AA 2012, 'Fecal Progestin Extraction and Analysis for Non-invasive Monitoring of Ovarian Cycle in Beef Cows', Pakistan Veterinary Journal, vol. 32, no. 4, pp. 584-8.
Yu, WH, Kimura, M, Walczewska, A, Karanth, S & McCann, SM 1997a, 'Role of leptin in hypothalamic-pituitary function (vol 94, pg 1023, 1997)', Proceedings of the National Academy of Sciences of the United States of America, vol. 94, no. 20, pp. 11108-.
Yu, WH, Walczewska, A, Karanth, S & McCann, SM 1997b, 'Nitric oxide mediates leptin-induced luteinizing hormone-releasing hormone (LHRH) and LHRH and leptin-induced LH release from the pituitary gland', Endocrinology, vol. 138, no. 11, pp. 5055-8.
Zakari, AY, Molokwu, ECI & Osori, DIK 1981, 'Effect of season on the estrous-cycle of cows (Bos-indicus) indigenous to northern Nigeria', Veterinary Record, vol. 109, no. 11, pp. 213-5.
Zieba, DA, Amstalden, M, Maciel, MN, Keisler, DH, Raver, N, Gertler, A & Williams, GL 2003, 'Divergent effects of leptin on luteinizing hormone and insulin secretion are dose dependent', Experimental Biology and Medicine, vol. 228, no. 3, pp. 325-30.
Zieba, DA, Amstalden, M & Williams, GL 2005, 'Regulatory roles of leptin in reproduction and metabolism: A comparative review', Domestic animal endocrinology, vol. 29, no. 1, pp. 166-85.