Embed Size (px)
Citation preview

Int J Clin Exp Pathol 2016;9(5):4943-4952www.ijcep.com /ISSN:1936-2625/IJCEP0024016
Original Article MicroRNA 21 and PLAP-1 regulate periodontal ligament remodeling during tooth movement of rats
Hong Hong1*, Jinxuan Zheng1*, Lu Liu2, Lixiang Mai1, Xi Wei2, Liping Wu1
1Department of Orthodontics, Guanghua School of Stomatology, Affiliated Stomatological Hospital, Guangdong Province Key Laboratory of Stomatology, Sun Yat-sen University, Guangzhou, P. R. China; 2Department of Operative Dentistry and Endodontics, Guanghua School of Stomatology, Affiliated Stomatological Hospital, Guangdong Province Key Laboratory of Stomatology, Sun Yat-sen University, Guangzhou, P. R. China. *Equal con-tributors.
Received January 14, 2016; Accepted March 24, 2016; Epub May 1, 2016; Published May 15, 2016
Abstract: Periodontal ligament associated protein-1 (PLAP-1), a newly discovered extracellular matrix protein in peri-odontal ligament (PDL), plays a unique biological role in periodontal ligament mineralization. MicroRNA 21 (miR-21) is one of the endogenous small noncoding RNAs which is involved in osteogenic differentiation of PDLCs by targeting PLAP-1 in vitro. Our study was to investigate the specific expression and function of PLAP-1 and miR-21 in PDL dur-ing orthodontic tooth movement and attempt to study on whether miR-21 modulated PDL remodeling by targeting PLAP-1 in vivo. We detected the expression of PLAP-1 and miR-21 in rat PDL during experimental tooth movement by in situ hybridization and immunohistochemical staining. The results showed that both PLAP-1 and miR-21 specifi-cally expressed in PDL. The expression of PLAP-1 on the pressure side increased to the peak in 7 days (7d-) group and decreased in 14d- and 21d- groups, while its expression on tension side decreased in 3d- and 5d- groups and increased later. MiR-21 expression on both pressure side and tension side exhibited a trend of increasing firstly and decreasing later. But the peak of expression appeared at different time on the two sides (7 days for pressure side and 3 days for tension side). Our study suggests that both miR-21 and PLAP-1 were involved in the reconstruction of periodontal tissue under the orthodontic force in rats. MiR-21 probably targeted PLAP-1 to implicate in the peri-odontal bone remodeling in the late period of tooth movement on tension side.
Keywords: PLAP-1, miR-21, tooth movement, periodontal ligament, orthodontics
Introduction
The periodontal ligament (PDL) is a group of connective tissue fibers that attaches the tooth roots to the inner wall of the alveolar bone. It can provide support, protection, nutrition and sensory input to the tooth. In addition, it also contributes to the homeostasis, remodeling and repair of periodontal tissue [1]. The PDL is always exposed to mechanical forces under physiological environment, such as speech, mastication, physiologic drifting and eruption of tooth. Since PDL can transmit the mechani-cal forces to alveolar bone, it can adapt to a rapidly changing applied force level and keep its own homeostasis [2, 3]. With the ability of self-repair and self-reconstruction, PDL plays a critical role in the bone remodeling of periodon-tal tissues under the orthodontic force [4].
Periodontal ligament associated protein-1 (PLAP-1), also named asporin, is a novel mem-ber of small leucine-rich repeat proteoglycan (SLRP) which expressed specifically and pre-dominantly in PDL [5]. Recent studies about PLAP-1 were focused on its biological function in osteoarticular diseases and considered as a susceptible gene of osteoarthritis [6-8]. Few researches were about the expression and function of PLAP-1 in PDL. PLAP-1 is specifically expressed in PDL, it primitively expresses in the dental follicle then enhances during the course of the cytodifferentiation of the PDL cells into mineralized tissue-forming cells such as osteo-blasts and cementoblasts [5]. Results from in vitro studies suggested that PLAP-1 plays a specific role in the PDL as a negative regulator of cytodifferentiation and mineralization to maintain the potential of the tissue structure and prevent the PDL from developing non-phys-

MicroRNA 21 and PLAP-1 in tooth movement
4944 Int J Clin Exp Pathol 2016;9(5):4943-4952
iological mineralization such as ankylosis [9]. However, its possible function during tooth movement remains unclear.
MicroRNAs are endogenous single-stranded noncoding RNAs, which are found in various organs from plants to humans. Some of them play important roles in cancers, heart diseases and cell differentiation [10]. MicroRNA 21 (miR-21), one member of microRNAs, has been proved to be involved in embryonic stem cells (ESCs) differentiation as an important compo-nent of mediating differentiation and stem cell fate determination [11]. The probable mecha-nism is that miR-21 can regulate the expres-sion and function of mRNAs through extracel-lular signaling by combining to their promoters and transcription factors [12]. Previous studies have concentrated in the regulating effect of miR-21 during adipogenesis and osteogenesis of mesenchymal stem cells (MSCs) and miR-21 was reported to be up-regulated consistent with differentiated potential of MSCs [13-15]. The latest research has found that miR-21 could modulate the osteogenic differentia- tion of periodontal ligament cells by targeting PLAP-1 in vitro by using bioinformation analy- sis [16]. However, the expression pattern and relationship of miR-21 and PLAP-1 in PDL dur-ing orthodontic tooth movement still remain unknown.
Most of studies have used real time polymerase chain reaction (RT-PCR) or miRNAs microarray technology to evaluate miRNAs expression. Although they’re very sensitive for miRNAs detection, RT-PCR and microarray technology are unable to locate gene expression positions. It’s necessary to understand where the miRNAs and target protein locate and how specific they are. In situ hybridization has only been used to evaluate miRNA expression in solid tumor [17] and some organ [18]. There’s no research about miR-21 detected in situ hybridization to illustrate the relationship between miR-21 and dentoalveolar tissues. In this study, we used in situ hybridization and immunohistochemical staining to locate the positions of miR-21 and PLAP-1 in PDL of rats treated with orthodontic appliances, and investigate their expression in PDL during orthodontic tooth movement. This study attempted to explore the function and location of them in PDL remodeling and investi-gate whether miR-21 modulated PDL remodel-ing by targeting PLAP-1 in vivo.
Materials and methods
Animal experiment
Thirty male Sprague-Dawley rats (6-8 weeks, 250-300 g) were obtained from the Sun Yat- Sen University’s experimental animal center (Guangzhou, China). The animals were random-ly divided into 6 groups with 5 rats of each. During the experiment, the rats were kept in cages in a room maintained at 21°C-24°C with a 12 h/12 h light/dark cycle and fed a soft diet with water. All animals were treated under the ethical regulations for animal experiments, defined by the Institutional Ethics Committee. Each animal’s weight was recorded throughout the experimental period, and there was no sig-nificant loss of weight.
Establishment of tooth movement model
An orthodontic elastic Ni-Ti closed-coil spring (3M Unitek, USA) was fixed between the maxil-lary right first molar and the incisors with ultra-violet curable resin (3M Unitek, USA) under anesthesia by intraperitoneal injection of 3% sodium pentobarbital (40 mg/kg). The spring was activated immediately upon insertion with a force of 50 g and the maxillary first molars were moved medially. No reactivation was per-formed during the experimental periods. The
Figure 1. The model of experimental tooth move-ment in rat. Left side (L) is loading orthodontic force and right side (R) is remaining as control.

MicroRNA 21 and PLAP-1 in tooth movement
4945 Int J Clin Exp Pathol 2016;9(5):4943-4952
left side of maxilla (with a non-activated spring) was used as control (Figure 1). The fit of rats was checked daily.
Tissue preparation
At 0, 1, 3, 7, 14 and 21 d after springs activa-tion, the rats were anesthetized with 3% sodi-um pentobarbital (40 mg/kg intraperitoneally) and fixed by cardiac perfusion with 4% freshly prepared paraformaldehyde in 100 mL Phos- phate-buffered saline (PBS, pH 7.2-7.4). The maxillae were dissected and fixed with the same fixative for 72 h at 4°C. The samples were then decalcified with 10% EDTA in 100 mL PBS for 6-8 weeks and embedded in paraffin. Tissue blocks were cut into mesio-distal serial sec-tions (4 mm thick) and mounted on Poly-L-Lysine coated glass slides (Shitai, Shanghai, China). Selected sections were stained with hematoxylin and eosin (MP Biomedicals, USA).
Immunohistochemical staining
Tissue sections were placed in a Tris-hydroxy methyl aminomethane buffered saline solution (TBS) at pH 7.4 for 10 minutes. Thereafter, endogenous peroxidase activity was blocked in hydrogen peroxide for 10 minutes in the dark.
and then counterstained with Mayer's hema-toxylin, dehydrated, and cover-slipped for light microscopic analysis. To prove the specificity of the immunoreactions, negative controls were carried out by omitting the primary antibody or using nonimmune immunoglobulin G instead, omitting both the primary and secondary anti-body, and using TBS/BSA instead.
In situ hybridization
In situ hybridization was performed according to a previous study [19]. The cRNA probe of PLAP-1 was obtained from Land Biosciences Inc. (Guangzhou, China). The cRNA probe of miR-21 was purchased from Exigon Company (Netherlands). In the presence of digoxigenin-labeled 11-dUTP (Roche, Germany), the anti-sense and sense cRNA probes were prepared by in vitro transcription using SP6 and T7 RNA polymerases, respectively. Digoxigenin (DIG)-11-UTP-labeled single-stranded cRNA probes were prepared with a DIG RNA labeling kit (Roche, Germany) according to the manufac-turer’s instructions.
The sections were deparaffinized in xylene, hydrated, postfixed in 4% paraformaldehyde, and sequentially treated with 10 mg/ml pro-
Figure 2. Results of hematoxylin-eosin staining of mesial roots of upper first molar during experimental tooth movement of day 0 (A), day 7 (B), day 21 (C). In the experimental groups, the collagen fibers in the periodontal ligament of the pressure side became lining up tightly with the extension of time, bone resorption pits with osteoclasts were seen on the surface of alveolar bone (D, arrows). On the tension side, collagen fibers became loose, cell compo-nents increased, osteoblasts were seen in the periodontal ligament closing to the surface of alveolar bone (E). Until 21 days, bone resorption pits on pressure side decreased (F). (M mesial side of periodontal ligament, D distal side of periodontal ligament, P pulp, R root, A alveolar, A-C×200, D-F×400).
Subsequently, sections were rinsed and then pre-incubat-ed with TBS containing 4% goat’s serum albumin (TBS/BSA) for 20 minutes to pre-vent unspecific background staining. Then, sections we- re incubated with a primary rabbit anti-asporin polyclonal antibody (Sigma, USA) in a 1:200 working solution of TBS/BSA at 4°C overnight in a humidified chamber. The slides were rinsed again and incubated for 30 minutes with a rabbit anti-goat immuno-globulin diluted 1:100 in TBS/BSA (ZSGB-BIO, Beijing, Ch- ina) as secondary antibody. Incubation was stopped in TBS before the peroxidase-antiperoxidase complex was administered for 30 minutes. After another rinse, the tissue sections were stained in a 3, 3’-diaminobenzidine solution for about 5 minutes, rinsed,
MicroRNA 21 and PLAP-1 in tooth movement
4946 Int J Clin Exp Pathol 2016;9(5):4943-4952
teinase K at 37°C for 30 min and with 0.1 M triethanolamine containing 0.25% acetic anhy-dride for 10 min. Hybridization was done at 50°C under high stringency conditions using the denatured probes at concentrations of about 1 ng/ml in a freshly prepared hybridiza-tion mixture. All samples were hybridized over-night at 50°C. Post-hybridization treatment included incubation with RNase A at 37°C for 30 min followed by thorough washes. The washed slides were incubated with anti-digoxi-genin monoclonal antibody (Roche, Germany) overnight at 4°C. After the application of bioti-nylated rabbit anti-rat IgG antibody (Dako, Glostrup, Denmark), the sections were incub- ated with alkaline phosphatase-conjugated streptavidin (Dako) and then with nitro blue tetrazolium/5-bromo-4-chloro-3-indolyl phos-phate solution (Roche, Germany) for 3-5 h to
ators (Hong Hong, Jinxuan Zheng) and any dis-agreement was resolved through discussion or assessment by a third investigator (Liping Wu). Both operators were blinded to the treatment allocation.
Statistical analysis
The data were processed with SPSS 22.0 soft-ware (Chicago, USA). The results were expressed as means and standard deviations. For in situ hybridization and immunohistochemical stain-ing data, the results for each group were the means and standard deviations of the 5 mea-surements, each as an average of the 3 sec-tions from a specimen. The data between groups were evaluated by independent T test analysis of variance. The significance level was set at P<0.05.
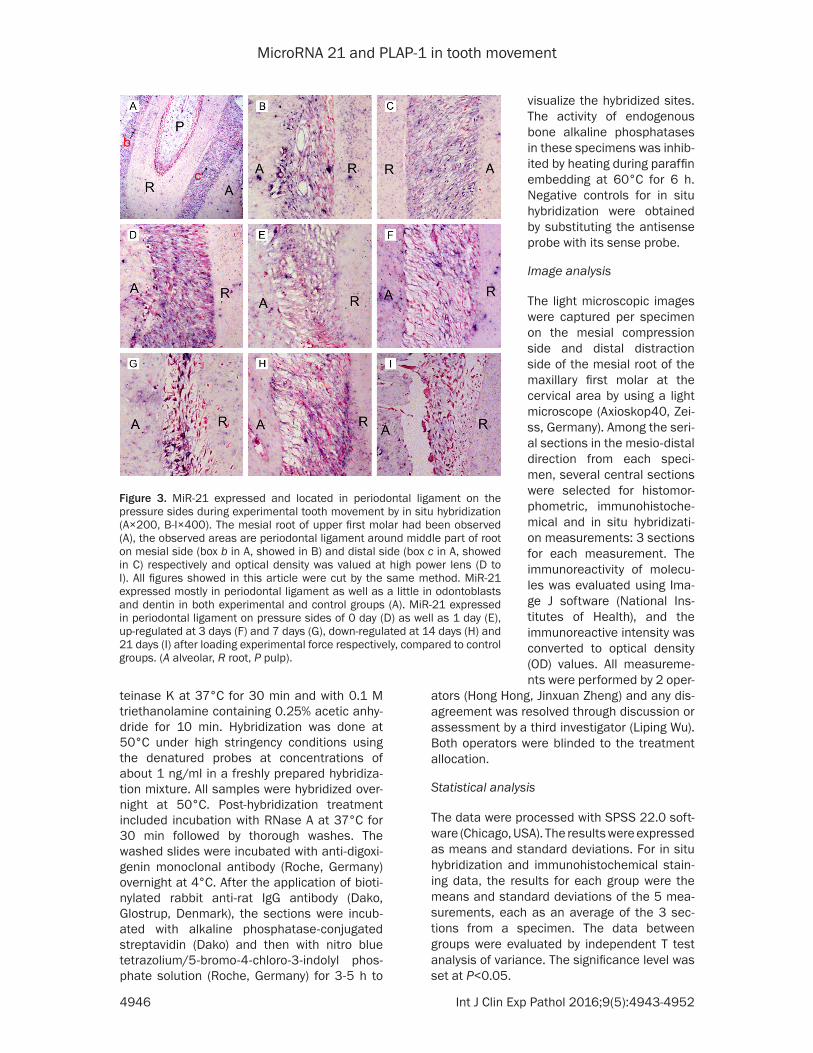
Figure 3. MiR-21 expressed and located in periodontal ligament on the pressure sides during experimental tooth movement by in situ hybridization (A×200, B-I×400). The mesial root of upper first molar had been observed (A), the observed areas are periodontal ligament around middle part of root on mesial side (box b in A, showed in B) and distal side (box c in A, showed in C) respectively and optical density was valued at high power lens (D to I). All figures showed in this article were cut by the same method. MiR-21 expressed mostly in periodontal ligament as well as a little in odontoblasts and dentin in both experimental and control groups (A). MiR-21 expressed in periodontal ligament on pressure sides of 0 day (D) as well as 1 day (E), up-regulated at 3 days (F) and 7 days (G), down-regulated at 14 days (H) and 21 days (I) after loading experimental force respectively, compared to control groups. (A alveolar, R root, P pulp).
visualize the hybridized sites. The activity of endogenous bone alkaline phosphatases in these specimens was inhib-ited by heating during paraffin embedding at 60°C for 6 h. Negative controls for in situ hybridization were obtained by substituting the antisense probe with its sense probe.
Image analysis
The light microscopic images were captured per specimen on the mesial compression side and distal distraction side of the mesial root of the maxillary first molar at the cervical area by using a light microscope (Axioskop40, Zei- ss, Germany). Among the seri-al sections in the mesio-distal direction from each speci-men, several central sections were selected for histomor-phometric, immunohistoche- mical and in situ hybridizati-on measurements: 3 sections for each measurement. The immunoreactivity of molecu- les was evaluated using Ima- ge J software (National Ins- titutes of Health), and the immunoreactive intensity was converted to optical density (OD) values. All measureme- nts were performed by 2 oper-
MicroRNA 21 and PLAP-1 in tooth movement
4947 Int J Clin Exp Pathol 2016;9(5):4943-4952
Results
HE staining
Representative histology of the mesial root of the upper first molars of control specimens was shown in Figure 2. The area observed in this study contained the dentin and pulp of the mesial root, PDL and the adjacent alveolar bone. In the experimental groups, the collagen
expression of miR-21 was significantly higher in 3d- group than control group (P<0.05). The opti-cal density results were shown in Figure 5.
Expression of PLAP-1 by in situ hybridization and immunohistochemical staining
In our study, both in situ hybridization and immunohistochemical staining results showed that the PLAP-1 was specifically expressed in
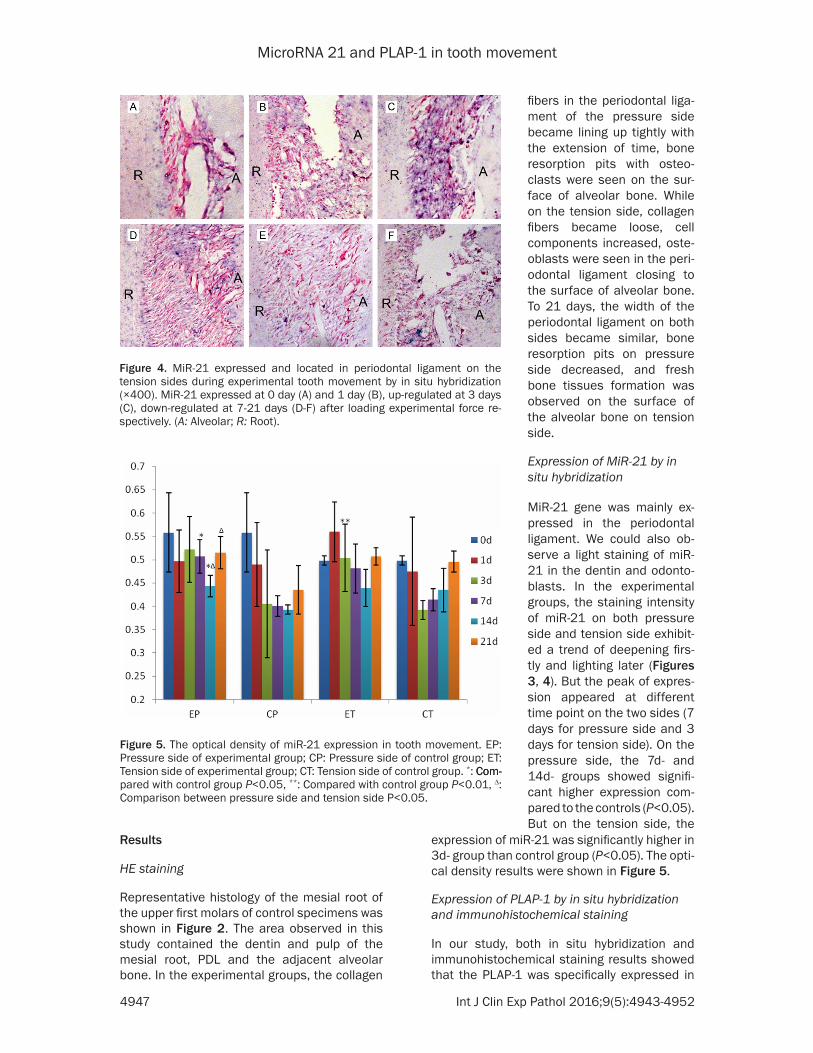
Figure 4. MiR-21 expressed and located in periodontal ligament on the tension sides during experimental tooth movement by in situ hybridization (×400). MiR-21 expressed at 0 day (A) and 1 day (B), up-regulated at 3 days (C), down-regulated at 7-21 days (D-F) after loading experimental force re-spectively. (A: Alveolar; R: Root).
fibers in the periodontal liga-ment of the pressure side became lining up tightly with the extension of time, bone resorption pits with osteo-clasts were seen on the sur-face of alveolar bone. While on the tension side, collagen fibers became loose, cell components increased, oste- oblasts were seen in the peri-odontal ligament closing to the surface of alveolar bone. To 21 days, the width of the periodontal ligament on both sides became similar, bone resorption pits on pressure side decreased, and fresh bone tissues formation was observed on the surface of the alveolar bone on tension side.
Expression of MiR-21 by in situ hybridization
MiR-21 gene was mainly ex- pressed in the periodontal ligament. We could also ob- serve a light staining of miR-21 in the dentin and odonto-blasts. In the experimental groups, the staining intensity of miR-21 on both pressure side and tension side exhibit-ed a trend of deepening firs- tly and lighting later (Figures 3, 4). But the peak of expres-sion appeared at different time point on the two sides (7 days for pressure side and 3 days for tension side). On the pressure side, the 7d- and 14d- groups showed signifi-cant higher expression com-pared to the controls (P<0.05). But on the tension side, the
Figure 5. The optical density of miR-21 expression in tooth movement. EP: Pressure side of experimental group; CP: Pressure side of control group; ET: Tension side of experimental group; CT: Tension side of control group. *: Com-Com-pared with control group P<0.05, **: Compared with control group P<0.01, ∆: Comparison between pressure side and tension side P<0.05.
MicroRNA 21 and PLAP-1 in tooth movement
4948 Int J Clin Exp Pathol 2016;9(5):4943-4952
the periodontal ligament without any expres-sion in the alveolar bone, cementum or gums. Under the high power microscope, PLAP-1 was mainly located in the osteoblasts, fibroblasts and cementoblasts adjacent to the surface of the alveolar bone and cementum, but there’s no expression of PLAP-1 in mineralized alveolar bone and cementum. For in situ hybridization experiments, the expression of PLAP-1 on the
mineralizated state, which plays a key role in the self-repair of periodontal tissue during tooth movement [2, 3, 20].
As a kind of newfound extracellular matrix pro-tein, PLAP-1 has the function to inhibit mineral-ization. According to previous studies, PLAP-1 inhibits cartilage formation [21] and suppre- ssed the mineralization of PDL [22]. Over-
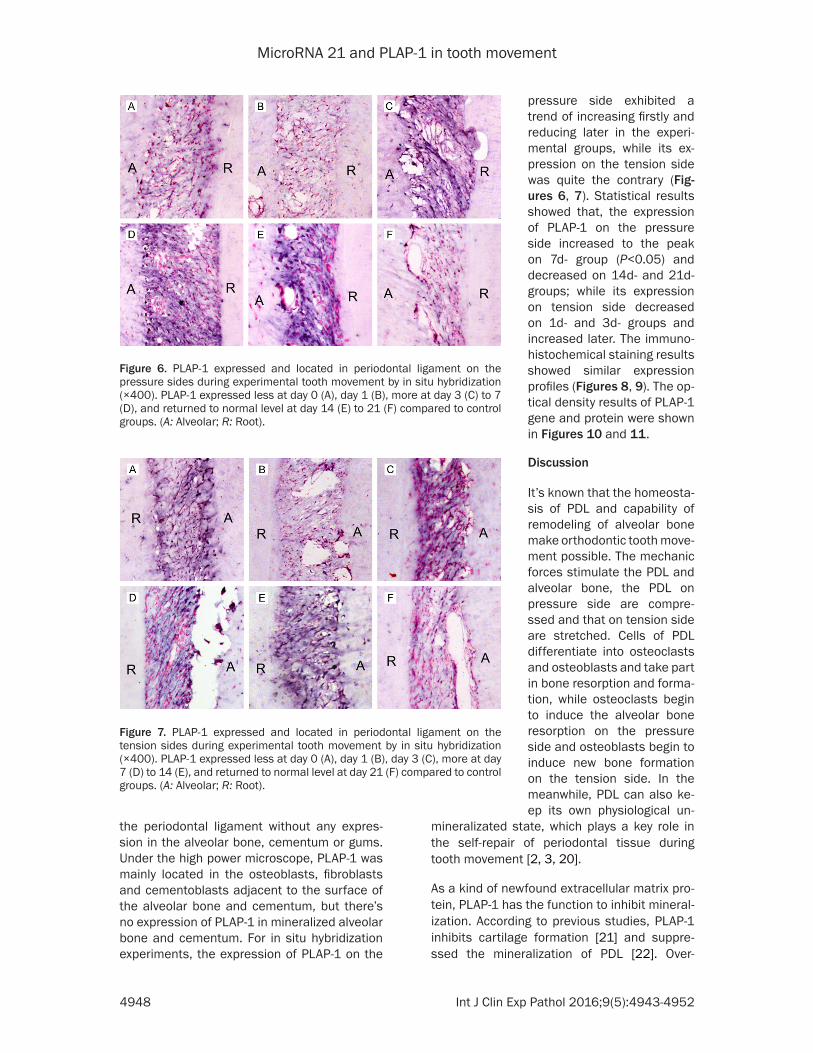
Figure 6. PLAP-1 expressed and located in periodontal ligament on the pressure sides during experimental tooth movement by in situ hybridization (×400). PLAP-1 expressed less at day 0 (A), day 1 (B), more at day 3 (C) to 7 (D), and returned to normal level at day 14 (E) to 21 (F) compared to control groups. (A: Alveolar; R: Root).
Figure 7. PLAP-1 expressed and located in periodontal ligament on the tension sides during experimental tooth movement by in situ hybridization (×400). PLAP-1 expressed less at day 0 (A), day 1 (B), day 3 (C), more at day 7 (D) to 14 (E), and returned to normal level at day 21 (F) compared to control groups. (A: Alveolar; R: Root).
pressure side exhibited a trend of increasing firstly and reducing later in the experi-mental groups, while its ex- pression on the tension side was quite the contrary (Fig- ures 6, 7). Statistical results showed that, the expression of PLAP-1 on the pressure side increased to the peak on 7d- group (P<0.05) and decreased on 14d- and 21d- groups; while its expression on tension side decreased on 1d- and 3d- groups and increased later. The immuno-histochemical staining results showed similar expression profiles (Figures 8, 9). The op- tical density results of PLAP-1 gene and protein were shown in Figures 10 and 11.
Discussion
It’s known that the homeosta-sis of PDL and capability of remodeling of alveolar bone make orthodontic tooth move-ment possible. The mechanic forces stimulate the PDL and alveolar bone, the PDL on pressure side are compre- ssed and that on tension side are stretched. Cells of PDL differentiate into osteoclasts and osteoblasts and take part in bone resorption and forma-tion, while osteoclasts begin to induce the alveolar bone resorption on the pressure side and osteoblasts begin to induce new bone formation on the tension side. In the meanwhile, PDL can also ke- ep its own physiological un-
MicroRNA 21 and PLAP-1 in tooth movement
4949 Int J Clin Exp Pathol 2016;9(5):4943-4952
expression of PLAP-1 plays a negative role in osteogenic differentiation of mesenchynal stem cells [23]. These studies suggested that PLAP-1 is associated with alveolar bone remod-eling and maintain of homeostasis of PDL dur-ing orthodontic tooth movement. In our study, the data of both in situ hybridization and immu-
PLAP-1 and investigate whether miR-21 modu-lated tooth movement remodeling by targeting PLAP-1.
It’s generally accepted that pressure induces osteoclast-mediated bone resorption during orthodontic tooth movement in rats [25]. Pre-
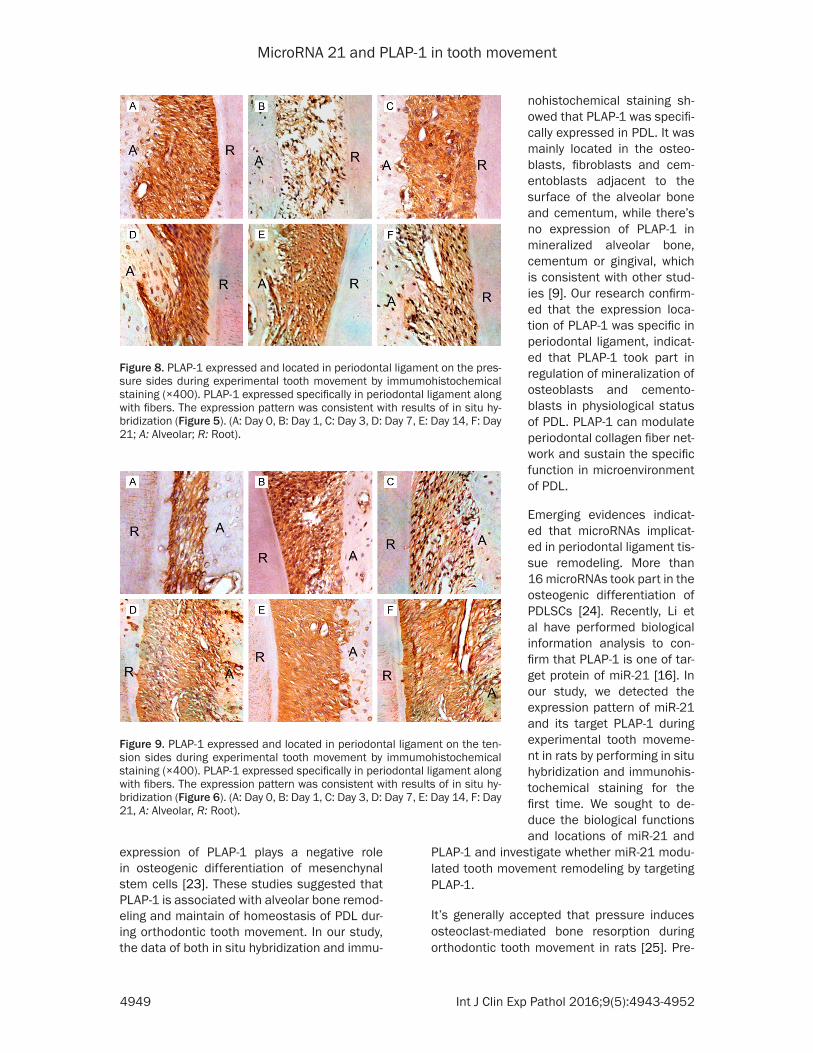
Figure 8. PLAP-1 expressed and located in periodontal ligament on the pres-sure sides during experimental tooth movement by immumohistochemical staining (×400). PLAP-1 expressed specifically in periodontal ligament along with fibers. The expression pattern was consistent with results of in situ hy-bridization (Figure 5). (A: Day 0, B: Day 1, C: Day 3, D: Day 7, E: Day 14, F: Day 21; A: Alveolar; R: Root).
Figure 9. PLAP-1 expressed and located in periodontal ligament on the ten-sion sides during experimental tooth movement by immumohistochemical staining (×400). PLAP-1 expressed specifically in periodontal ligament along with fibers. The expression pattern was consistent with results of in situ hy-bridization (Figure 6). (A: Day 0, B: Day 1, C: Day 3, D: Day 7, E: Day 14, F: Day 21, A: Alveolar, R: Root).
nohistochemical staining sh- owed that PLAP-1 was specifi-cally expressed in PDL. It was mainly located in the osteo-blasts, fibroblasts and cem- entoblasts adjacent to the surface of the alveolar bone and cementum, while there’s no expression of PLAP-1 in mineralized alveolar bone, cementum or gingival, which is consistent with other stud-ies [9]. Our research confirm- ed that the expression loca-tion of PLAP-1 was specific in periodontal ligament, indicat-ed that PLAP-1 took part in regulation of mineralization of osteoblasts and cemento-blasts in physiological status of PDL. PLAP-1 can modulate periodontal collagen fiber net-work and sustain the specific function in microenvironment of PDL.
Emerging evidences indicat-ed that microRNAs implicat- ed in periodontal ligament tis-sue remodeling. More than 16 microRNAs took part in the osteogenic differentiation of PDLSCs [24]. Recently, Li et al have performed biological information analysis to con-firm that PLAP-1 is one of tar-get protein of miR-21 [16]. In our study, we detected the expression pattern of miR-21 and its target PLAP-1 during experimental tooth moveme- nt in rats by performing in situ hybridization and immunohis-tochemical staining for the first time. We sought to de- duce the biological functions and locations of miR-21 and

MicroRNA 21 and PLAP-1 in tooth movement
4950 Int J Clin Exp Pathol 2016;9(5):4943-4952
vious study has suggested that the upregula-tion of miR-21 decreases the apoptosis of osteoclasts and promotes osteoclastogenesis [26]. Our results showed that miR-21 was expressed in PDL and partly expressed in den-tin and odontoblasts, indicated that miR-21 was involved in PDL tissue reconstruction and
7d- groups while it happened earlier in 3d- groups on tension side. This indicated that miR-21 was implicated in orthodontic experimental tooth movement and was probably an impor-tant regulator of PDL tissue remodeling. Acc- ording to previous studies in vitro, down-regula-tion of miR-21 could induce the up-regulation of
Figure 10. The optical density of PLAP-1 gene expression in tooth move-ment by in situ hybridization. EP: Pressure side of experimental group; CP: Pressure side of control group; ET: Tension side of experimental group; CT: Tension side of control group. *: Compared with control group P<0.05, **: Compared with control group P<0.01, ∆: Comparison between pressure side and tension side P<0.05.
Figure 11. The optical density of PLAP-1 protein expression in tooth move-ment by immunohistochemical staining. EP: Pressure side of experimental group; CP: Pressure side of control group; ET: Tension side of experimen-tal group; CT: Tension side of control group. *: Compared with control group P<0.05, **: Compared with control group P<0.01, ∆: Comparison between pressure side and tension side P<0.05, ∆∆: Comparison between pressure side and tension side P<0.01.
dentinogenesis. On the pres-sure side of PDL, the expres-sion profile of miR-21 was increased firstly and decrea- sed late. The 7d- and 14d- groups showed significant hi- gher expression of miR-21 compared to the controls. The data showed that miR-21 was involved in bone resorption of PDL on the pressure side; the expression pattern was con-sistent with the study in vitro [26]. On the tension side of experimental groups, althou- gh the expression of miR-21 was in an increased-decrea- sed manner, it was signifi-cantly upregulated in 3d- groups compared with con-trols. After that, it decreased to the level of control groups (P<0.05). Takanori E, et al. found that miR-21 was high- ly expressed in 4 hours and down-expressed in 14 days after induction of mineraliza-tion of MSCs in vitro [27]. Combining to our results, the two studies showed that miR-21 might be the initiator of differentiation; it was highly expressed in the early stage of mineralization and decrea- sed in the late period of bone formation.
The expression pattern of miR-21 on both pressure si- des and tension sides of ex- perimental groups was char-acterized in an increased-decreased manner, but the turning point occurred at dif-ferent time point. As we sh- owed in figures, miR-21 upreg-ulated on the pressure side in

MicroRNA 21 and PLAP-1 in tooth movement
4951 Int J Clin Exp Pathol 2016;9(5):4943-4952
target protein PLAP-1 in the late induction of mineralization of PDLCs [16], however, when loaded stretch forces, upregulated miR-21 pro-moted osteogenic differentiation of PDLSCs [28]. Our study confirmed that miR-21 indeed overexpressed on both pressure and tension sides in experimental groups. The expression patterns of PLAP-1 and miR-21 were reversed on tension side of experimental groups. We can deduce that miR-21 probably targeted PLAP-1 to regulate bone remolding after loading orth-odontic forces on tension side. Since contain-ing a lot of different factors and complicated modulated networks, the in vivo environment is much more complicated than in vitro condi-tions, there’re other regulators of PDL tissue remodeling needing further study.
In conclusion, our study makes it clear that miR-21 and PLAP-1 located in PDL and were involved in the reconstruction of periodontal tissue during the experimental tooth movement in rats. The expression of miR-21 and PLAP-1 both upregulated at the early time point of tooth movement, then reversed in the later period. Considering the complication of in vivo environment, it can be concluded that miR-21 may probably modulated PDL remodeling by targeting PLAP-1 in the late period of tooth movement and maintained the potential of bone remodeling to improve tooth movement.
Acknowledgements
This study was supported by National Natural Science Foundation of China (No. 814030607) and Natural Science Foundation of Guangdong Province China (No. 10451008901005923).
Disclosure of conflict of interest
None.
Address correspondence to: Liping Wu, Department of Orthodontics, Guanghua School of Stomatology, Affiliated Stomatological Hospital, Guangdong Province Key Laboratory of Stomatology, Sun Yat-sen University, 56 Ling Yuan Xi Road, Guangzhou 510055, P. R. China. Tel: (86-20) 83802802; E-mail: [email protected]
References
[1] Beertsen W, McCulloch CA, Sodek J. The peri-odontal ligament: a unique, multifunctional connective tissue. Periodontol 2000 1997; 13: 20-40.
[2] Xu H, Han X, Meng Y, Gao L, Guo Y, Jing Y, Bai D. Favorable effect of myofibroblasts on colla-gen synthesis and osteocalcin production in the periodontal ligament. Am J Orthod Den- tofacial Orthop 2014; 145: 469-479.
[3] Krishnan V, Davidovitch Z. Cellular, molecular, and tissue-level reactions to orthodontic force. Am J Orthod Dentofacial Orthop 2006; 129: 461-469.
[4] Chamila Prageeth Pandula PK, Samaranayake LP, Jin LJ, Zhang C. Periodontal ligament stem cells: an update and perspectives. J Investig Clin Dent 2014; 5: 81-90.
[5] Yamada S, Murakami S, Matoba R, Ozawa Y, Yokokoji T, Nakahira Y, Ikezawa K, Takayama S, Matsubara K, Okada H. Expression profile of active genes in human periodontal ligament and isolation of PLAP-1, a novel SLRP family gene. Gene 2001; 275: 279-286.
[6] Xing D, Ma XL, Ma JX, Xu WG, Wang J, Yang Y, Chen Y, Ma BY, Zhu SW. Association between aspartic acid repeat polymorphism of the asporin gene and susceptibility to knee osteo-arthritis: a genetic meta-analysis. Osteoarthritis Cartilage 2013; 21: 1700-1706.
[7] Duval E, Bigot N, Hervieu M, Kou I, Leclercq S, Galera P, Boumediene K, Bauge C. Asporin ex-pression is highly regulated in human chondro-cytes. Mol Med 2011; 17: 816-823.
[8] Juchtmans N, Dhollander AA, Coudenys J, Audenaert EA, Pattyn C, Lambrecht S, Elewaut D. Distinct dysregulation of the small leucine-rich repeat protein family in osteoarthritic ace-tabular labrum compared to articular carti-lage. Arthritis Rheumatol 2015; 67: 435-441.
[9] Yamada S, Tomoeda M, Ozawa Y, Yoneda S, Terashima Y, Ikezawa K, Ikegawa S, Saito M, Toyosawa S, Murakami S. PLAP-1/asporin, a novel negative regulator of periodontal liga-ment mineralization. J Biol Chem 2007; 282: 23070-23080.
[10] Kumarswamy R, Volkmann I, Thum T. Re- gulation and function of miRNA-21 in health and disease. RNA Biol 2011; 8: 706-713.
[11] Lakshmipathy U, Hart RP. Concise review: MicroRNA expression in multipotent mesen-chymal stromal cells. Stem Cells 2008; 26: 356-363.
[12] Singh SK, Kagalwala MN, Parker-Thornburg J, Adams H, Majumder S. REST maintains self-renewal and pluripotency of embryonic stem cells. Nature 2008; 453: 223-227.
[13] Kim YJ, Hwang SJ, Bae YC, Jung JS. MiR-21 regulates adipogenic differentiation through the modulation of TGF-beta signaling in mes-enchymal stem cells derived from human adi-pose tissue. Stem Cells 2009; 27: 3093-3102.
[14] Kim YJ, Hwang SH, Cho HH, Shin KK, Bae YC, Jung JS. MicroRNA 21 regulates the prolifera-

MicroRNA 21 and PLAP-1 in tooth movement
4952 Int J Clin Exp Pathol 2016;9(5):4943-4952
tion of human adipose tissue-derived mesen-chymal stem cells and high-fat diet-induced obesity alters microRNA 21 expression in white adipose tissues. J Cell Physiol 2012; 227: 183-193.
[15] Mei Y, Bian C, Li J, Du Z, Zhou H, Yang Z, Zhao RC. miR-21 modulates the ERK-MAPK signal-ing pathway by regulating SPRY2 expression during human mesenchymal stem cell differ-entiation. J Cell Biochem 2013; 114: 1374-84.
[16] Li C, Li C, Yue J, Huang X, Chen M, Gao J, Wu B. miR-21 and miR-101 regulate PLAP-1 expres-sion in periodontal ligament cells. Mol Med Rep 2012; 5: 1340-1346.
[17] Dillhoff M, Liu J, Frankel W, Croce C, Bloomston M. MicroRNA-21 is Overexpressed in Pan- creatic Cancer and a Potential Predictor of Survival. J Gastrointest Surg 2008; 12: 2171-2176.
[18] Hu J, Huang J, Zeng L, Wang G, Cao M, Lu H. Anti-Apoptotic Effect of MicroRNA-21 after Contusion Spinal Cord Injury in Rats. J Neurotrauma 2013; 30: 1349-1360.
[19] Toyosawa S, Shintani S, Fujiwara T, Ooshima T, Sato A, Ijuhin N, Komori T. Dentin matrix pro-tein 1 is predominantly expressed in chicken and rat osteocytes but not in osteoblasts. J Bone Miner Res 2001; 16: 2017-2026.
[20] Kajikawa T, Yamada S, Tauchi T, Awata T, Yamaba S, Fujihara C, Murakami S. Inhibitory effects of PLAP-1/asporin on periodontal liga-ment cells. J Dent Res 2014; 93: 400-405.
[21] Kou I, Nakajima M, Ikegawa S. Binding charac-teristics of the osteoarthritis-associated pro-tein asporin. J Bone Miner Metab 2010; 28: 395-402.
[22] Kajikawa T, Yamada S, Tauchi T, Awata T, Yamaba S, Fujihara C, Murakami S. Inhibitory effects of PLAP-1/asporin on periodontal liga-ment cells. J Dent Res 2014; 93: 400-405.
[23] Zhang PP, Li S, Yang PS, Sun J, Hou C. [Expression of periodontal ligament-associat-ed protein-1 in normal periodontal tissues and cells in rat]. Zhonghua Kou Qiang Yi Xue Za Zhi 2011; 46: 338-341.
[24] Ng TK, Carballosa CM, Pelaez D, Wong HK, Choy KW, Pang CP, Cheung HS. Nicotine alters MicroRNA expression and hinders human adult stem cell regenerative potential. Stem Cells Dev 2013; 22: 781-790.
[25] Skripitz R, Aspenberg P. Pressure-induced peri-prosthetic osteolysis: a rat model. J Orthop Res 2000; 18: 481-484.
[26] Sugatani T, Vacher J, Hruska KA. A microRNA expression signature of osteoclastogenesis. Blood 2011; 117: 3648-3657.
[27] Eguchi T, Watanabe K, Hara ES, Ono M, Kuboki T, Calderwood SK. OstemiR: A Novel Panel of MicroRNA Biomarkers in Osteoblastic and Osteocytic Differentiation from Mesencymal Stem Cells. PLoS One 2013; 8: e58796.
[28] Wei F, Liu D, Feng C, Zhang F, Yang S, Hu Y, Ding G, Wang S. microRNA-21 mediates stretch-induced osteogenic differentiation in human periodontal ligament stem cells. Stem Cells Dev 2015; 24: 312-319.