Embed Size (px)
Citation preview

Lineages and Stem Cells: B-cell development

The Circulatory System
The circulatory system consists of the heart (to pump blood),the blood vessels (to transport blood) and the blood itself. All of these must develop concurrently in the early embryo to allow a functioning circulatory system

What is blood?
Mammalian blood is 45% red blood cells, 1% whiteblood cells and the remainder is plasma (the liquid portion containing various dissolved proteins).

Why do we need blood cells?
1.) Red blood cells carry oxygen to tissues and carbon dioxide to lungs (or to the placenta prior to birth)
2.) Platelets are crucial for blood clotting, which prevents excess bleeding if there is a breach in the integrity of a blood vessel
3.) White blood cells fight infection.
The first two functions need to be peformed at an early stage of embryonic development (as soon as blood vessels form), but the last one is only needed mainly after birth. So not all blood cell types are produced at the same time in the embryo

Early human (before 16 days gestation) and mouse embryos(before 7 days gestation) do not have blood. Instead the embryo obtains all the required nutrients and oxygen from simple diffusion. Waste products also diffuse out of the embryo into the mother’s circulation.
Once the embryo reaches a large enough size, simple diffusion is not sufficient to bring nutrients or remove waste productsnecessitating the development of a circulatory system and blood.
When is blood first made in an embryo?
12 days7 days 8 days5 days 6 days

HematopoiesisHematopoiesis means forming blood
Hematopoiesis has discrete stages in embryonicdevelopment
In mammals, blood cell development takes place in four main organs - yolk sac, placenta, fetal liver and bone marrow and two discrete stages - primitive and definitive

Places of Hematopoiesis
Note: The AGM is part of the early embryonic body, described later
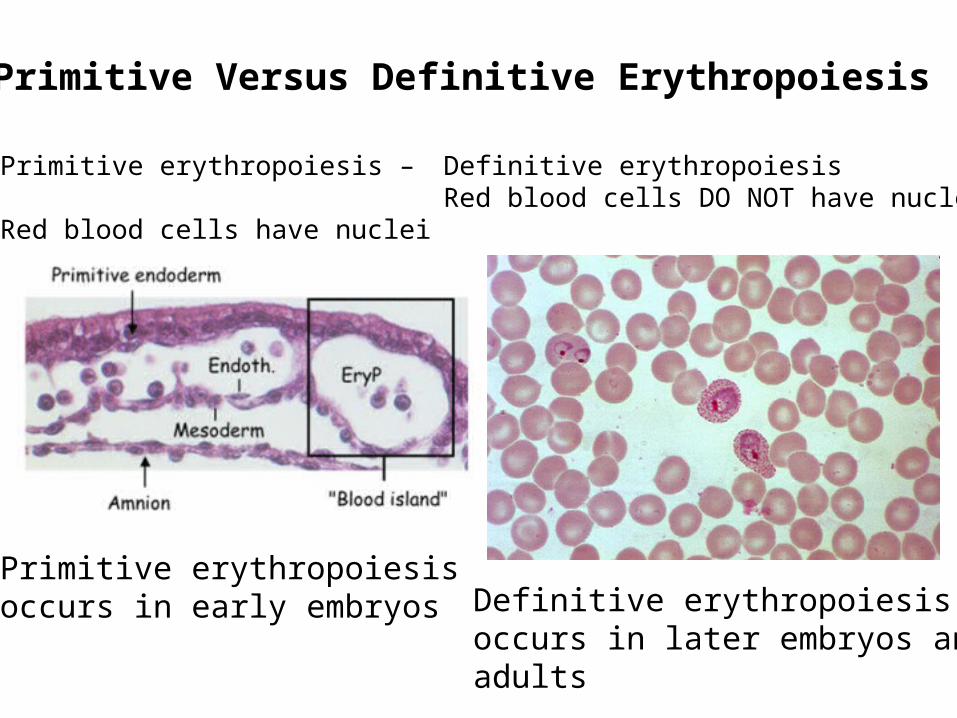
Primitive Versus Definitive Erythropoiesis
Primitive erythropoiesis –
Red blood cells have nuclei
Definitive erythropoiesisRed blood cells DO NOT have nuclei
Primitive erythropoiesisoccurs in early embryos Definitive erythropoiesis
occurs in later embryos andadults
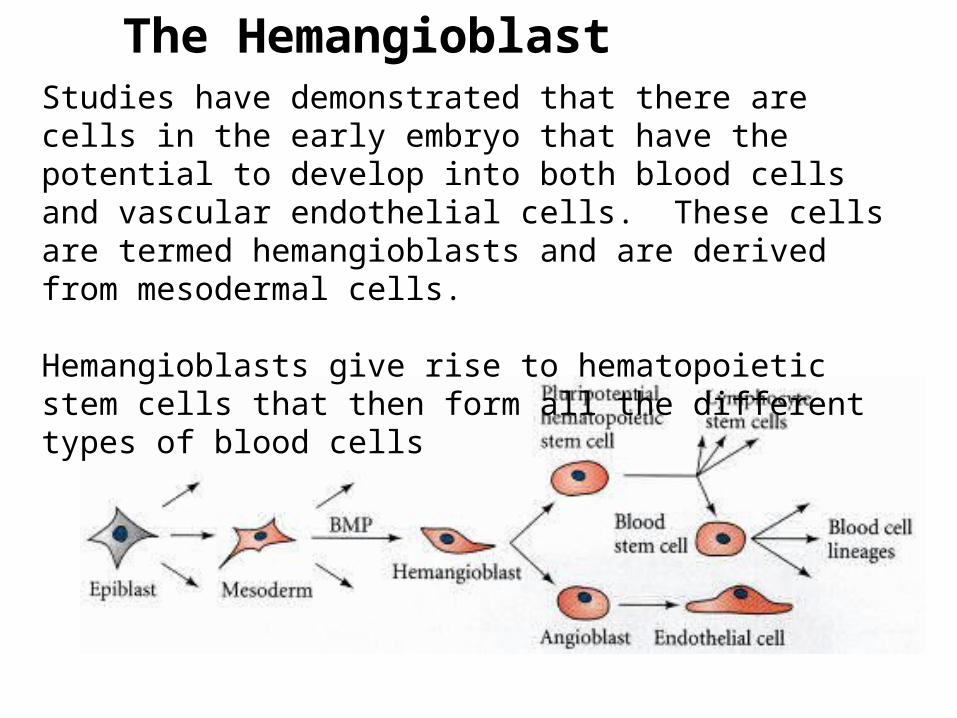
The HemangioblastStudies have demonstrated that there are cells in the early embryo that have the potential to develop into both blood cells and vascular endothelial cells. These cells are termed hemangioblasts and are derived from mesodermal cells.
Hemangioblasts give rise to hematopoietic stem cells that then form all the different types of blood cells

Hematopoietic Stem CellsAll blood cells are derived from hemato-poietic stem cells (HSC). Stem cell can prolif-erate to replace them-selves and to give rise to more committed progeny.
The HSC
Lots of redblood cells and platelets are needed. Many fewer of the other cell types are required

Blood and a circulatory system need to start formingbeginning around day 18-20 of human development and around day 7-8 of mouse development
At this stage, the embryos are still very tiny, so there is not much room to generate blood
Therefore, the first blood cells to form are made outside the embryo in a membrane called the yolk sac
Yolk Sac Hematopoiesis
Yolk SacYolk Sac EmbryoEmbryo

In the yolk sac, the first place of hematopoiesis, undifferentiated mesenchymal cells differentiate to clusters of hemangioblast cells. The hemangioblasts further differentiate into endothelial cells and primitive blood cells. This is the first “blood vessel” like structure in the embryo.
Yolk Sac Hematopoiesis (blood islands)

Fetal Liver Hematopoiesis
When the embryo is bigger and more developed, hematopoiesis switches from the yolk sac to the fetal liver. This occurs around day 30-40 of human development and around day 10-11 of mouse develop-ment.

The AGM regionDerived from mesoderm that is associated with the internal organs of the body. This mesoderm lies close to and contributes to the development of the aorta, the early kidney (mesonephros) and the gonad.

Stem cells in the AGMHematopoietic stem cells from the yolk sac or the aorta-gonad-mesonephros (AGM) region of the embryo migrate to the fetal liver to give rise to fetal liver hematopoiesis
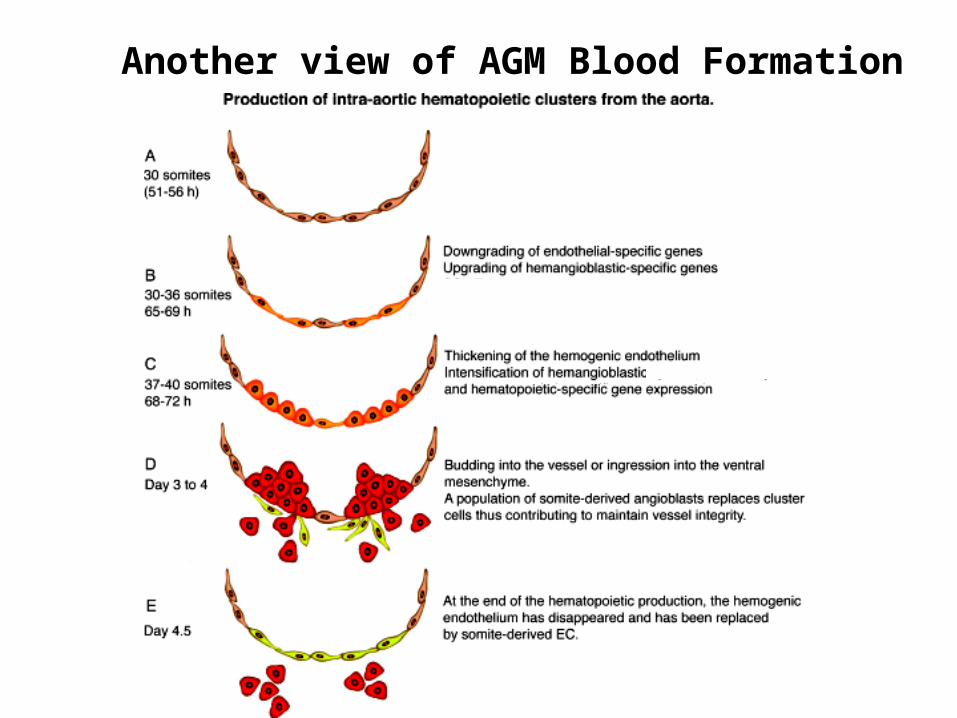
Another view of AGM Blood Formation

Fetal Liver Hematopoiesis
Colonized by definitive hematopoietic stem cellsderived from the AGM and possibly also from the yolk sac blood islands.
All blood cell types (except T cells) can differentiatein the fetal liver.
Fetal liver cellsNeutrophil (n)

Placental hematopoiesisThe discovery of the placenta as a major source of embryonic blood cells came later than the other tissues and is less well-understood than yolk sac, fetal liver or bone marrow hematopoiesis.
Hematopoiesis in the placenta overlaps with yolk sac and fetal liver hematopoiesis.
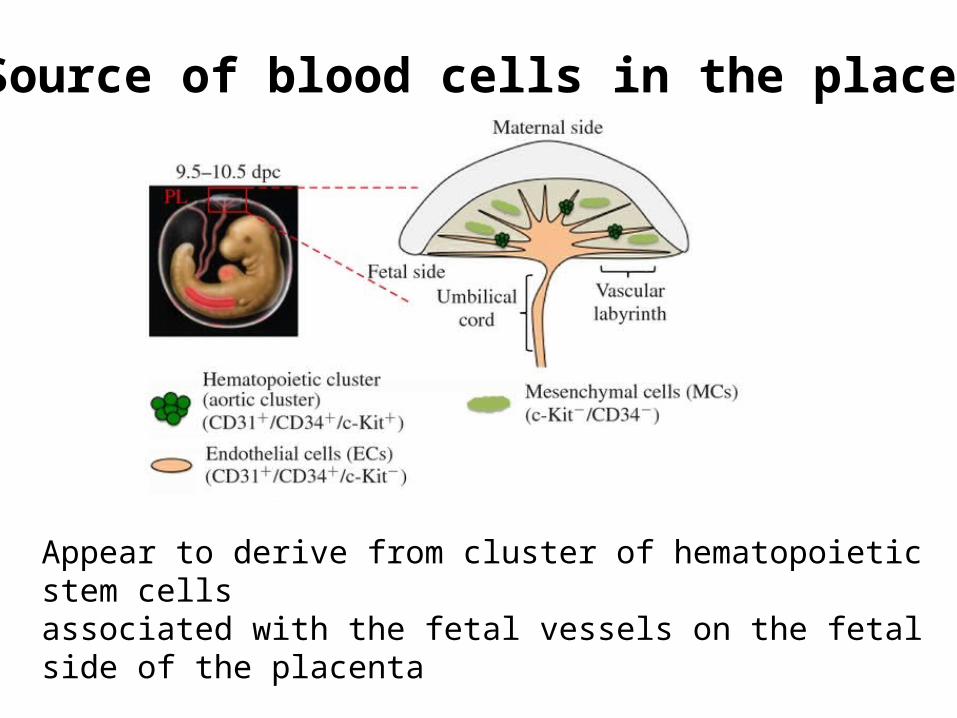
Source of blood cells in the placenta
Appear to derive from cluster of hematopoietic stem cellsassociated with the fetal vessels on the fetal side of the placenta
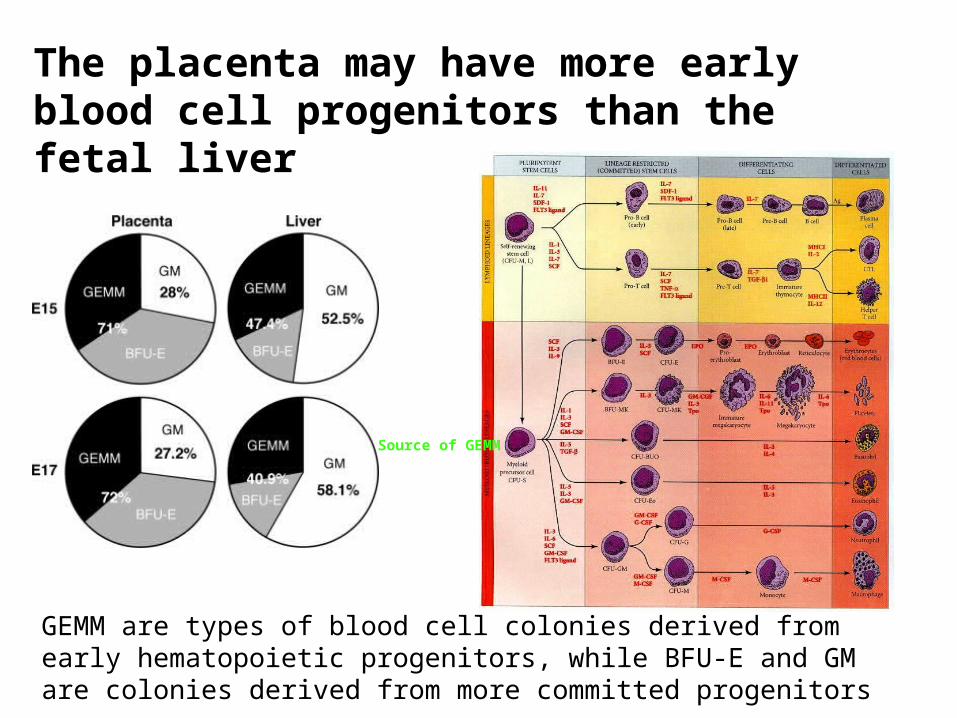
The placenta may have more early blood cell progenitors than the fetal liver
GEMM are types of blood cell colonies derived from early hematopoietic progenitors, while BFU-E and GM are colonies derived from more committed progenitors
Source of GEMM

Bone Marrow Hematopoiesis
In adult humans and mice, hematopoiesis mainly takes place in the bone marrow. The figure above shows the main sites of hematopoiesis in adult humans (red bones are the ones with highest blood cell production)

Bone marrow hematopoiesis begins in late in gestation (day 17-18 until birth in mice, week 22-birth in humans), because the bones have not developed sufficiently before this time to support hematopoiesis
Bone marrow hematopoiesis continues throughout adult life, but hematopoiesis can also occur in the liver or spleen if the bone marrow is not producing enough blood cells
Timing of bone marrow hematopoiesis

The Bone Marrow Hematopoiesis
Colonized late in embryogenesis by definitive hemato-poietic stem cells derived from the fetal liver (and placenta?). All blood cell types (except T cells) can differentiate in the bone marrow.

Generalities of Hematopoiesis
No matter which organ (yolk sac, placenta, fetal liver orbone marrow, hematopoietic stem cells and their moredifferentiated progeny need a proper environment togrow and differentiate.
This includes proper cell-cell contacts, proper cytokines and growth factors and other additional requirements forparticular blood cell lineages
These specific requirements can explain, at least in part, why blood cannot form anywhere in an embryo, but is localized to very specific regions

Various types of cytokines and growth factors are also required for hemato-poiesis to occur
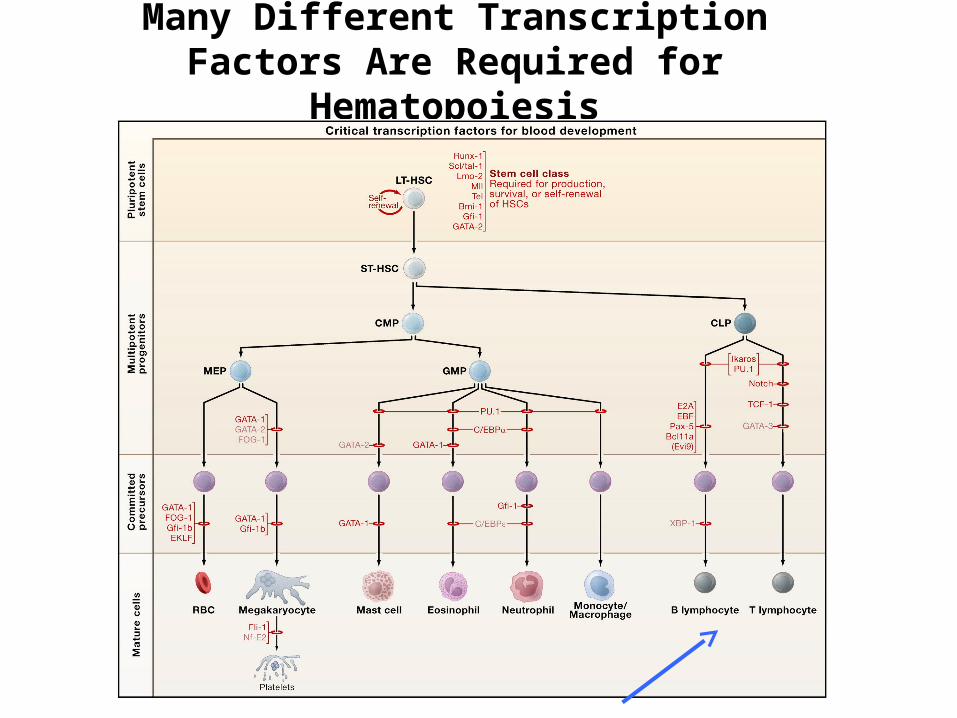
Many Different Transcription Factors Are Required for
Hematopoiesis

Differentiation of a specific blood cell type-the B lymphocyte
As an example of hematopoiesis, we will discuss the development of one type of white blood cell, the B cell, in the adult bone marrow.
To better understand their development, it is important to know what B cells do – they produce antibodies to fight infection
Antibody genes are not encoded in our DNA in an assemble form, but rather need to be generated by DNA rearrangements that must take place during B cell development

B Cell Development
Hematopoietic stem cells give rise to a lymphoidprogenitor cell, which is thought to be able to differentiate into both B cells and T cells as well as natural killer (NK) cells.
This lymphoid progenitor can migrate to the thymusand initiate T cell development or remain in the bonemarrow to initiate B cell development.
Bone marrow B cell development can be subdivided into various stages: pro-B, pre-B, immature B and mature B cells.

B Cell Development is Dependent on Several Factors
A.) The presence of bone marrow stromal cells (a kind of fibroblast) providing cell- mediated contacts and secreting the cytokine IL-7.
B.) The rearrangement of the immunoglobulin (antibody) genes. In the common lymphoid progenitor both the immunoglobulin heavy and light chains are in their germ-line configuration (the unrearranged state) and must be rearranged (so they can give rise to an in-frame antibody protein) for B cell development to progress.
C.) Expression of particular cytokine receptors and transcription factors by the developing B cell

Bone Marrow Stromal Cell-Cell Contacts
Bone marrow stromal cells provide at least two importantcontacts to B cell progenitors: an adhesive interaction through the integrin VLA-4 binding to VCAM-1 and asignaling one through surface bound SCF interacting with the Kit receptor on the lymphoid progenitor.
Bone marrow B lymphoidprogenitors adhering to bone marrow stromal cellsin culture.

Bone Marrow Stromal Cell Derived IL-7
IL-7 secreted by bone marrow stromal cells binds tothe IL-7 receptor expressed on pro- and pre-B cells and stimulates the survival and proliferation of the B cells.

Rearrangement of the Immunoglobulin Genes
B cells exist to produce antibodies (immunoglobulins)Thus, B cell progenitors in the bone marrow will only survive and proliferate if they are able to rearrange the immunoglobulin genes to generate functionalImmunoglobulins.

How can a developing B cell tell if it has a functionally-rearranged immunoglobulin gene?
The cells make a surface receptor out of the rearranged immunoglobulin and see if it signals!

pro-B pre-B immature Bmature Bactivated B
Bone Marrow Periphery
AntibodyProductionHeavy
ChainRearrangement
LightChain
Rearrangement
Surface Immunoglobulin Receptors on B Cell Progenitors
In pre-B cells, the rearranged immunoglobulinheavy chain pairs with two other proteins 5 and V-preB, the surrogate light chains. This formsthe pre-B cell receptor, which signals the cell toproliferate and further differentiate.

Summary of B Cell Development
Stromal Cell Dependence? Yes Yes Yes No No No NoIL-7 Dependence? No Yes Yes Yes Yes No No