Embed Size (px)
Citation preview

(CANCER RESEARCH 48, 6555-6559, November 15, 1988]
Intervention of T-Cells in Transportation of Mouse Mammary Tumor Virus(Milk Factor) to Mammary Gland Cells in Vivo1
Airo Tsubura,2 Muneo Inaba, Shunsuke Imai, Akira Murakami, Naoki Oyaizu, Ryoji Yasunii/u, Yoko Ohnishi,
Harutaka Tanaka, Sotokichi Morii, and Susumu IkeharaDepartment of Pathology, Kansai Medical University, Moriguchi, Osaka 570 [A. T., M. I., N. O., R. Y., Y. O., S. M., Su. l.¡;Department of Pathology, Nara MedicalCollege, Kashihara, Nara 634 fSh. I.J; and Institute for Virus Research, Kyoto University, Kyoto, Kyoto 606 [A. M., H. TJ; Japan
ABSTRACT
Using BALB/c nu/nu, BALB/c nu/nutOI I (BALB/c nu/nu mice raisedby C3H/HeN foster mother), BALB/c thymus-engrafted BALB/c nuln«fC3H,BALB/c nu/+, and BALB/c n«/+fC3Hmice, we examined whatkinds of cells are carriers of blood-borne mouse mammary tumor virus(B-MMTV). A radioimmunoassay and an immunoperoxidase assay revealed the presence of MMTV-gp52 antigen in the mammary glands ofall BALB/c n«/+fC3H and BALB/c thymus-engrafted BALB/c nulnufC3H mice (more than 60 days old) but only of some BALB/c nulnufC3H mice (more than 120 days old): those that possessed a significantnumber of functional T-cells. BALB/c nu/+ mice did not show the antigenexpression at any age. Transfer experiments of cells or plasma fromyoung (<12 weeks) BALB/c nu/nirfC3H to BALB/c +/+ virgins revealedthat cells besides T-cells can also become carriers of B-MMTV. Thiswas confirmed by Southern blotting analyses; exogenous provirus DNAsequences were found in B-cells as well as T-cells of BALB/c nu/+fC3Hmice. However, when young BALB/c nu/nu mice were inoculated withBALB/c i)u/nufC3H blood, they did not show the MMTV-gp52 antigenexpression. Transfer experiments of purified T-cells, B-cells, naturalkiller cells, and macrophages from BALB/c IX311 mice to BALB/c nu/nu mice revealed that only T-cells have the ability to transfer viralactivity to the mammary glands.
These results suggest that B-MMTV is carried from the gastrointestinal tract to the mammary glands by lymphoid cells such as T-cells andB-cells, then transferred to the mammary gland cells by the T-cells.
INTRODUCTIONThe in vivo life cycle of exogenous MMTV3 is still uncertain.
MMTV is produced in the mammary gland of infected femalesand transferred to suckling pups as a milk factor. Intact MMTVB-particles can be observed on the surface of the intestinalepithelium but not in the cytoplasm (1). Destruction of thevirus structure, therefore, seems to occur when the virus entersthe cell, and MMTV "footprints" can be traced through viral
antigens. Immunohistochemically, viral antigens are taken upin the intestinal epithelial cells by endocytosis in coated pitsand channeled toward larger vacuoles by fusion with lysosomes(2). Vacuolar contents are released at the basolateral membrane.However, the antigen is not detected in the intercellular spacesof intestinal absorptive cells or in the lymph spaces below thebasolateral membrane (2, 3).
Received 9/10/87; revised 2/22/88, 6/29/88; accepted 8/2/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported in part by Grants-in-Aid for Cancer Research,
61010037 and 62010025, from the Ministry of Education, Science, and Culture,Japan (to A. T.). This work was also supported by the Japanese Ministry ofHealth and Welfare, a grant from the Naito Foundation, a grant from theMitsubishi Foundation, a Grant-in-Aid from the Mochida Memorial Foundationfor Medical and Pharmaceutical Research, National Institutes of Health, theScience Research Promotion Fund of the Japan Private School PromotionFoundation ( 1987), and a Grant-in-Aid for Cancer Research, 62015088, from theMinistry of Education, Science and Culture (to Su. I.).
2To whom requests for reprints should be addressed.3The abbreviations used are: MMTV, mouse mammary tumor virus; nu/nu,
congenitally athymic nude; gp52, glycoprotein with a molecular weight of 52,000;N'K, natural killer; IPA, immunoperoxidase assay; HAN, hyperplastic alveolar
nodule; RIA, radioimmunoassay; B-MMTV, blood-borne mouse mammary tumorvirus; Con A, concanavalin A.
A new method should be employed to determine the route ofthe virus from the gastrointestinal tract to the mammary glandcells. A number of investigators have reported that blood elements propagate MMTV from the digestive tract to the mammary gland (4). However, electron microscopic studies havefailed to demonstrate B-particles in blood cells, indicating thatMMTV is in a subviral state (5). Virus activity related to bloodcells has been designated blood-borne MMTV (B-MMTV), asdistinct from B-particle-associated mammary tissue-borneMMTV. The MMTV activity in the blood is found in cellularcomponents but not in the plasma (6). This ability to infect wasfirst thought to be associated with erythrocytes and later withnucleated hemic cells, i.e., leukocytes and/or hematopoieticstem cells (4). Lymphocytes have also been assumed to be thecarriers (7). Recently, exogenous MMTV DNA sequences inlymphoid cells of BALB/cfC3H mice have been found (8),suggesting that lymphoid cells are the carriers of B-MMTVactivity. However, it is not known which lymphoid cells are theactual carriers. Since B-MMTV is integral in the life cycle ofMMTV, it is of great interest to determine the cells responsiblefor carrying the B-MMTV activity. The present study focuseson lymphoid cells and has been designed to determine usingBALB/c «n//i«fC3Hmice which type of lymphocytes (T or B)are the carrier of B-MMTV.
MATERIALS AND METHODS
Mice. BALB/c +/+, BALB/c nu/+, BALB/c nu/nu, and C3H/HeNmice were purchased from CLEA Japan Inc., Osaka, Japan. TheBALB/c nu/+fC3H and BALB/c nu//iufC3H mice were raised in ouranimal facility. In brief, a BALB/c nu/+ female was mated with aBALB/c nu/nu male and was kept on an elevated wire mesh platformto allow her babies to drop into a cage in which a C3H foster motherwas kept. Subsequent generations were nursed by their natural mothers.All of these mice were kept under specific pathogen-free conditions.
Neonatal Thymus Graft. Whole or single thymus lobes of neonatalBALB/c mice were implanted under the shoulder skin of BALB/c nu/nufC3H mice (9).
Immunoperoxidase Assay. The ABC technique using ami MMTVgpS2 serum was applied to paraffin-embedded sections that had beenfixed in a 10% neutral buffered formalin solution. The method and thecharacterization of the antibody have been described in detail elsewhere(10). Mammary glands from mice were hormonally stimulated in orderto more easily detect the presence of antigen. Stimulation was inducedby mating with BALB/c male mice or pituitary transplantation in theinguinal mammary fat pad. This allowed detailed observations of thedistribution of viral antigens within the histológica! framework of theentire cell population. Samples considered positive consisted of clustersof gp52-positive alveoli surrounded by antigen negative alveoli. Samplesconsidered negative showed no viral antigen throughout the wholemammary gland.
Radioimmunoassay. Purified MMTV-gp52 was labeled with I25Ibythe chloramine-T method. The techniques used in the preparation oftissue extracts from the mammary glands, spleen, thymus, and bonemarrow have been described elsewhere (11). Measurements are definedin PÃŽÃŽ//J1of tissue extract.
Blood Cell Transfer Assay. Donors were 2- to 3-month-old BALB/c
6555
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

TRANSPORTATION OF EXOGENOUS MMTV IN VIVO
Table 1 Expression ofMMTV-gp52 antigen in various organs
MouseBALB/c
nu/+BALB/cm//+fC3HBALB/c
nu/nufC3H + thymusgraft
BALB/c n«/n«fC3HAge
(days)60-250
60-12060-12060-120
121-250Thy-1.2+
Spleencells(%)29
±430±620
±64±4
3±2Spleen0/5*
0/121/70/31/3(2)Thymus0/52/12(5.2-19.5)cRIA"Bone
marrow0/5
3/12(2-19.5)
0/70/30/3Mammary
gland0/5
11/12(26-3 x 10')
6/7(2-3.7 x IO9)
0/33/3
(2-1 x 10')IPAMammary
gland0/5
12/127/70/3
3/3
°Radioimmunoassay was performed, as described in "Materials and Methods" (MMTV-gp52 over 2 pg/^l was considered positive).
Number of positive mice/number of mice examined.
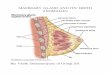
Fig. 1. MMTV-gp52 antigen-positive alveoli cluster surrounded by antigen-negative alveoli, in the mammary glands of lactating BALB/c nu/+fC3H mice(x 100).
nu/n«fC3H or BALB/c n«/+fC3H mice. The sampling of peripheralblood and plasma was according to the method of Nandi (12, 13). Toprepare the blood cell fraction, blood was pooled in a heparinized tubeand diluted with two volumes of saline. The mice in one group werethen injected with 0.3 ml of this solution. For plasma preparation,pooled blood was centrifuged at 150 x g for 15 min. The plasma wasseparated and diluted with two volumes of saline, and the mice in theother group injected with 0.3 ml of this solution. As in the method ofspleen cell preparation described by Bentvelzen (14), spleen cell suspensions were washed in Hanks' balanced salt solution, and the cell countwas adjusted to IO6 cells/0.3 ml. Six- to 7-month-old BALB/c nul+fC3H mice were used for the separation of T-cells, B-cells, NK-cells,and macrophages. B-cells were prepared as follows: spleen cells weretreated with monoclonal anti-Thy 1.2 antibody (Clone F7D5; Olac,Bicester, Oxon, UK) plus rabbit complement, and then passed throughSephadex G-10 columns (Pharmacia Fine Chemicals, Uppsala, Sweden). The resultant cell population, referred to as B-cells, containedmore than 95% surface immunoglobulin-positive cells; T-cells andmacrophages were not detected when checked using a cytofluorometer(FACStar, Becton and Dickinson, USA). T-cells were prepared bypassing spleen cells through a nylon wool-column (Wako pure Chemicals, Osaka, Japan). The purity of T-cells was more than 95% when
checked using a cytofluorometer. NK-cells were prepared as follows:spleen cells from MMTV-infected mice were passed through nylonwool columns twice, and the nylon wool-nonadherent cells were furthertreated with appropriate dilutions of anti-Thy 1.2 antibody plus complement to deplete mature T-lymphocytes. The resultant population,which consists of more than 80% asialo Gmi-positive cells, was referredto as NK-cell-enriched population. In addition, we have confirmed thatthis population shows killing activity to YAC-1 cells. Macrophageswere obtained from the plastic dish-adherent peritoneal exúdatecells.The purity of macrophages was more than 90%, when analyzed by acytofluorometer using anti-Mac 1 (Ml/70) antibodies. Each cell countwas adjusted to IO6 cells/0.3 ml. These cell suspensions were i.p.injected to 3- to 5-week-old BALB/c +/+, BALB/c nu/+, or BALB/cnu/nu females. Adult BALB/c mice served as hypophyseal donors forhormonal stimulation of the host mammary gland. One pituitary glandwas grafted to the right inguinal mammary fat pad. All mice wereallowed either 5 or 8 weeks of hormone stimulation. Right inguinalmammary glands were removed together with the grafted pituitarygland and were processed for IPA. All mice with hormone stimulationfor 8 weeks were checked using HAN assays 5 weeks after pituitarygraft removal in wholemount preparations. Hormonal stimulation wasalso achieved by mating with BALB/c male mice, and IPA was performed during lactation.
Cytotoxicity Text. The cytotoxicity test was carried out, as previouslydescribed (15).
Mitogen Assay. The mitogenic reactivity was determined by measuring the incorporation of [3H]thymidine into DNA as previously
reported (16).Restriction Endonuclease Digestion and Southern Blotting Analyses.
T-cells and B-cells were separated from the spleen of mammary tumor-bearing multiparous BALB/c nu/+fC3H mice (9- to 13-month old) andmultiparous BALB/c nu/+ females (16-month-old). MuMT73, an epithelial cell line which was derived from spontaneous mammary tumorsof BALB/CÕC3Hmice (17), was used for a positive control while BALB/c liver was used as a negative control. Each cellular DNA (10 /ig) wasisolated, digested with Pstl, subjected to electrophoresis on a 0.8%agarose gel, and transferred to nitrocellulose (18). The nitrocellulosefilter was then hybridized with a 32P-labeled probe specific for the gag-
poi fragment of MMTV(C3H) (8).Mammary Tumor Incidence and Latency. Multiparous BALB/c nu/
n«fC3H,BALB/c n«/+fC3H, and BALB/c nu/+ which survived morethan 200 days were sacrificed when carrying visible mammary tumors.
MI.B/C nu/tfC,K(17/T7)
Fig. 2. Age-dependent increase in MMTV-gp52 antigen expression in the mammaryglands of BALB/c nu/nufC3H mice as estimated by immunoperoxidase assay. •¿�,antigenpositive; O, antigen negative.
tALI/c nu/nufCjH•¿�•ft
U)ii ort(15/1
IALt/c iMi/iwfCjH(13/35)
Oo0 0oo o ooooooooo0000•oO*
»00••¿�•¿�••¿�••• «o» o
1(0
•¿�Positiv« O K«.t K.
6556
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

TRANSPORTATION OF EXOGENOUS MMTV IN VIVO
Table 2 Correlation between MMTV-gp52 antigen expression in the mammary glands and mitogen responsiveness in spleen cells ofBALB/c nu/+fC3H,BALB/c nu/nu with neonatal thymus grafts, and BALB/c nu/nufC3H mice
MouseBALB/c
n«/+fC3HBALB/c
nw/nufC3H+thymusgraftBALB/c
nu/nufC3HAge
(days)162170222231243206229229313346717882157162238310Thy-
1.2*
spleencells(%)303737402520223224340304137cpmBackground5.6
±0.06.4±1.91.8
±0.61.4±0.61.2±0.38.6
±0.36.7±1.16.0±0.76.3±0.71.1
±0.30.4
±0.20.6±0.10.7
±0.47.6±0.44.8±0.28.2±2.76.7±0.7x
10~3(meanPHA96.1
±9.435.5±3.144.8
±1.530.2±2.215.8+5.519.6
±2.47.2±0.114.7
±1.820.7±4.38.1±1.91.7
±1.21.4±0.41.6
+0.66.1±1.34.2±0.39.9±1.311.4±0.0±SD)Con
A220.8
±13.2190.0+3.042.0±3.438.1
±6.425.9±2.0162.6
±9.956.2±2.7155.0
±7.2145.2+3.067.0+
1.80.8
±0.00.7+0.10.7+0.131.6+0.634.8±2.082.9+5.170.7±5.2PHA17.05.524.420.913.22.31.12.43.36.83.72.32.30.80.91.21.7SI"Con
A39.129.522.926.421.518.88.425.722.957.61.81.21.04.27.210.110.9IPAMMTV-gp52++•f+++++++_——++++
1Stimulation index.
Table 3 IPA and HAN assay for MMTV-gp52 expression in the mammaryglands of BALB/c +/+ virgins inoculated i.p. with peripheral blood cells, spleen
cells, or plasma of BALB/c nu/+fC3H or BALB/c nu/nufC3H miceBALB/c nu/+fC3H and BALB/c nu/nufC3H mice (2-3 months old) were
used as donors.
7IH/+InoculationPeripheral
blood"Spleen cell'Plasma'Control'IPA7/10*
7/100/50/5HAN3/10
1/100/5
0/5nu/nuIPA9/10
8/100/80/5HAN2/10
1/100/80/5
" Triple-diluted peripheral blood (0.3 ml) inoculation.* Numbers indicate positive mice/mice examined.c Spleen cell (1 x 10' in 0.3 ml) inoculation.'' Triple-diluted plasma (0.3 ml) inoculation.'' Saline (0.3 ml) inoculation.
Table 4 Immunoperoxidase assay for MMTV-gp52 antigen expression in thelactating mammary glands of female BALB/c nu/nu or BALB/c nu/+ inoculatedi.p. with peripheral blood of 2 to 3-month-old BALB/c nu/+ or nu/nufC3H mice
Donor
Recipient" nu/nu
BALB/c nu/+BALB/c nu/nu
14/16*
13/1512/130/12
" All recipients were below 100 days of age when examined.* Number of positive mice/number of mice examined.
Table 5 Immunoperoxidase assay for MMTV-gp52 antigen expression in themammary glands of BALB/c nu/nu virgins inoculated i.p. with splenic
T-cells, B-cells, or peritoneal macrophages of 6- to /'-month-old
BALB/c nu/+fC3H miceInoculation"
Recipient* T-cell B-cell NK cell Macrophage Control
BALB/c nu/nu 7/1 Oc 0/10 0/10 0/5 0/5•¿�1 x 10" cells in 0.3 ml RPMI 1640 or 0.3 ml RPMI 1640 alone.* 5-week-old mice with 1 pituitary transplant were used.' Number of positive mice/number of mice examined.
Histológica! examinations including wholemount preparation and immunoperoxidase staining for MMTV-gp52 were performed.
RESULTS
Age-dependent Increase in Expression of MMTV-gp52 Antigen in BALB/c nu/nufC3H. We examined the expression ofMMTV-gp52 antigen in various organs using RIA and IPA.As shown in Table 1, BALB/c nu/+ mice did not show the
9.4-
7.4-6.6-5.8-5.7~4.9-
3.5-
2.3-
2.0-
123456
Fig. 3. Detection of exogenous MMTV proviral sequences. The sizes offragments are indicated in kilobase pairs. Lanes: I, BALB/c liver; 2, BALB/c nul+ T-cells; 3, BALB/c nu/+ B-cells; 4, BALB/c n«/+fC3H T-cells; 5, BALB/cn«/+fC3H B-cells; 6, BALB/cfC3H mammary tumor cell line MuMT 73.
antigen in any organs, whereas BALB/c /»//+fC3H miceshowed the antigen in the thymus, bone marrow, and mammaryglands, and BALB/c thymus-engrafted BALB/c nu/nufC3Hmice exhibited the antigen in the spleen and mammary glands.The positive rate of the MMTV-gp52 antigen in RIA was highin the mammary glands but low in the spleen, bone marrow,and thymus. The level of MMTV-gp52 in the mammary glands,which was measured by RIA, varied considerably from mouseto mouse (26 to 3 x IO9 pg/til). In contrast, IPA was more
sensitive and the results more consistent than those of RIA,although IPA does not allow quantitation of viral antigens. In
6557
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

TRANSPORTATION OF EXOGENOUS MMTV IN VIVO
agreement with the results of St. George et al. (19), lactatingBALB/c HM/+ÕC3Hmice showed MMTV-gp52-positive alveoliclusters surrounded by antigen-negative alveoli in the mammaryglands (Fig. 1). This uneven distribution of viral antigen mightreflect the wide range in the RIA data or false negative resultsin RIA. BALB/c nu/nufC3H mice (60-120 days) did not showthe antigen in RIA, whereas BALB/c nu/nufC3H mice (121-250 days) did, although the percentage of Thy 1.2+ cells waslow. Fig. 2 more clearly shows an age-dependent increase inMMTV-gp52 antigen expression of mammary glands as estimated by IPA; the viral antigen was detected in the mammaryglands of BALB/c nu/nufC3H mice from about 120 days ofage.
Relationship between Viral Antigen Expression in MammaryGlands and T-Cell Functions. The next step was to determinewhether or not there is a correlation between the viral antigenexpression and T-cell functions. As shown in Table 2, allBALB/c /iH/+fC3H mice and BALB/c nu/nutC3H with thymusgrafts (5 mice, respectively) showed MMTV-gp52 antigenexpression and high responsiveness to T-cell mitogens (phyto-hemagglutinin and Con A). Young BALB/c nu/nuïC3VLmice(3 mice; <120 days) exhibited neither MMTV-gp52 antigenexpression nor response to T-cell mitogens, whereas old BALB/c nu/nufC3H mice (4 mice; >120 days) showed MMTV-gp52antigen expression and significant responsiveness to Con A,although the percentage of Thy 1.2* cells was low. We thereforeconcluded that there is a correlation between MMTV-gp52antigen expression and T-cell functions (but not percentage ofThy 1.2-positive cells).
Transfer of Viral Activity. We examined whether viral activitycan be transferred by cells or plasma. As shown in Table 3,inoculation of peripheral blood or spleen cells from BALB/cnu/nufC3H as well as BALB/c w«/+fC3H mice (2-3 monthsold) caused the MMTV-gp52 antigen expression in the mammary glands of BALB/c +/+ virgins, although the incidence inHAN assays was relatively low compared with IPA. Plasmainoculation however did not cause any viral activity. To excludethe influence of host T-cells on the expression of the antigen,lactating mammary glands of young BALB/c nu/nu recipientwere used. As shown in Table 4, the BALB/c nu/nu miceshowed MMTV-gp52 antigen expression in mammary glandswhen inoculated with BALB/c /iw/+fC3H blood (but notBALB/c nu/nufC3H blood). Furthermore, as shown in Table5, only young BALB/c nu/nu mice which had been inoculatedwith exogenous M MTV-infected T-cells (but not B-cells, NKcells, or macrophages) exhibited the MMTV-gp52 antigenexpression in the mammary glands. These results indicate thatT-cells are essential for MMTV-gp52 antigen expression, although cells other than T-cells can also be carriers of B-MMTV.
Southern Blotting Analyses. As shown in Fig. 3, both lane 4(extract from T-cells from BALB/c nw/+fC3H mice) and lane5 (extract from B-cells of BALB/c nu/+K3H mice) displayednot only 5.0- and 5.4-kilobase endogenous fragments but alsothe 4.0-kilobase («—)genomic band (exogenous MMTV band)as seen in lane 6 (extract from MuMT73, a cell line originatingfrom spontaneous mammary tumors of BALB/cfC3H mice).In contrast, lane 1 (extract from BALB/c liver), lane 2 (T-cellsof BALB/c nu/+ mice), and lane 3 (B-cells of BALB/c nu/+mice) lacked the 4.0-kilobase band, which is specific for exogenous MMTV.
Mammary Tumor Incidence and Latency in BALB/c ««/+,BALB/c iw/+fC3H and BALB/c nu/nuÕC3H Mice. All micewere normally bred females (maximum three pregnancies). Theincidence and mean latency of mammary tumors after intro
ducing milk factor to BALB/c nu/+ or BALB/c nu/nu micewere 31/51 (60.8%) and 284 ±66 days in BALB/c nu/+ mice,and 5/13 (38.5%) and 338 ±50 days in BALB/c nu/nu mice.
Histological examination revealed that all were mammaryadenocarcinomas, which expressed MMTV-gp52 antigen. Although two mammary tumors were found in BALB/c nu/+mice (2/13 = 15.4% and 379 ±30 days), they were MMTV-gp52 antigen-negative mammary sarcomas.
DISCUSSION
Nude mice have been thought to exhibit no T-cell functionsdespite the presence of some Thy 1.2-positive cells (20). However, it has recently been demonstrated that they possess functional T-cells, the number of which increases with age (15, 16,21). In the present study, we examined the expression ofMMTV-gp52 antigen in the mammary glands of BALB/c nu/nu mice in relation to T-cell functions. As shown in Table 1and Fig. 1, an age-dependent increase in the expression of theantigen in BALB/c nu/nufC3H mice was observed. In addition,we have found that there is a correlation between the antigenexpression and T-cell functions (but not percentage of Thy 1.2-positive cells), as shown in Table 2. These results stronglysuggest that functional T-cells are essential to the expressionof MMTV-gp52 antigen in the mammary glands.
To examine whether T-cells are the only carriers of B-MMTV, transfer experiments were carried out. As shown inTable 3, inoculation of peripheral blood and spleen cells (butnot plasma) from young BALB/c nu/nufC3H mice caused theMMTV-gp52 antigen expression in the mammary glands ofBALB/c +/+ virgins. However, if there are no T-cells in vivo,no viral transmission occurred (Table 4). Table 5 shows thatonly MMTV-infected T-cells (but not B-cells, NK cells, ormacrophages) can induce MMTV-gp52 antigen expression inthe mammary gland of BALB/c nu/nu mice. These resultsindicate that cells other than T-cells can also be carriers of B-MMTV, but that T-cells are essential for transmission of B-MMTV to the mammary gland cells in vivo.
MMTV DNA sequences in lymphoid cells have been documented; several B-lymphocyte cell lines of BALB/c origin haveendogenous MMTV DNA sequences (22), and splenic B-cellsexpress MMTV-related antigen(s) (23-26). It has been demonstrated that T-cell lymphomas of BALB/c origin amplifyendogenous MMTV sequences (27). BALB/c mice have beenshown to possess one subgenomic-size and two genomic-sizeMMTV proviruses, but exogenous MMTV has not been foundin these strains (28-30). A method to discriminate between theexogenous and endogenous MMTV DNA sequences is therefore necessary. Recently, Liegler and Blair have demonstratedthe exogenous MMTV DNA sequences in lymphoid cells ofBALB/cfC3H mice using the DNA hybridization method (8).Using this same method, we have clearly demonstrated theexogenous MMTV DNA sequences in both T-cells and B-cellsof BALB/c nti/+fC3H mice.
In conclusion, both T-cells and B-cells are B-MMTV carriers,but T-cells are essential for the transfer of B-MMTV activityto mammary gland cells. There are two possibilities: (a) T-cellscarry B-MMTV the whole way from the gastrointestinal tractto the mammary gland cells, and (b) B-cells first carry B-MMTV in the gastrointestinal tract and then transfer it to T-cells. Since MMTV B-particles are not produced in blood cells(including T-cells), unencapsulated RNA and DNA may betransferred by cell-to-cell contact. However, the possibility thatadditional factors such as cytokines are involved in the transfer
6558
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

TRANSPORTATION OF EXOGENOUS MMTV IN VIVO
of B-MMTV activity to mammary glands still remains. We arenow attempting to elucidate the exact transfer mechanism ofMMTV activity from the gastrointestinal tract to the mammarygland cells.
REFERENCES
1. Tanaka, II.. and Tsujimura, D. An electron microscopic study on the fate ofingested mammary tumor virus in the alimentary canal of suckling mice.Annual Report of the Institute of Virus Research, Kyoto University, 10:94-100, 1967.
2. Tsubura, A., Ueda, S., Nakao, I., and Morii, S. Electron-immunocytochem-ical observations on MTV envelope glycoprotein (gpS2) in mammary glandsand intestines of GR mice. Proceedings of the Breast Cancer ResearchConference, p. 55. Denver, CO, 1983.
3. Hainaut, P., Francois, C, Calberg-Bacq, C. M., Vaira, D., and Osterrieth,P. M. Peroral infection of suckling mice with milk-borne mouse mammarytumor virus: uptake of the main viral antigens by the gut. J. Gen. Virol., 64:2535-2548, 1983.
4. Nandi, S., and McGrath, C. M. Mammary neoplasia in mice. Adv. CancerRes., / 7: 353-414, 1973.
5. Daams, J. II.. Calafat, J., Lasfargues, E. Y.. Kramarsky, B., and Bentvelzen,P. Mammary tumor virus-associated antigens on the membrane of infectedmouse spleen cells. Virology, 41: 184-186, 1970.
6. Nandi, S., Haslam, S., and Helmich, C. Cell-associated mammary tumorvirus in blood of BALB/cfC3H mice. J. Nati. Cancer Inst., 48: 1085-1088,1972.
7. Hageman, Ph. C., Calafat, J., and Daams, J. H. The mouse mammary tumorvirus. In: P. Emmelot and P. Bentvelzen (eds.), RNA Virus and Host Genomein Oncogenesis, pp. 283-300. Amsterdam: North-Holland, 1972.
8. Liegler, T. J., and Blair, P. B. Direct detection of exogenous mouse mammarytumor virus sequences in lymphoid cells of BALB/cfC3H female mice. J.Virol., 59: 159-162, 1986.
9. Loor, F., and Kindred, B. Differentiation of It-ell precursors in nude micedemonstrated by immunofluorescence of T-cell membrane markers. J. Exp.Med., 138: 1044-1055, 1973.
10. Tsubura, A., Ueda, S., Shikata, V, Morii, S., and Tanaka, H. Immunohis-tochemical expression of mammary tumor virus antigens in mammary glandsof virgin mice, in relation to Aftv genes. Acta Pathol. J pn., 36: 15-22, 1986.
11. Imai, S., Morimoto, J., Tsubura, Y., Iwai, Y., Okumoto, M., Takamori, Y.,Tsubura, A., and Hilgers, J. Tissue and organ distribution of mammarytumor virus antigens in low and high mammary cancer strain mice. Eur. J.Cancer Clin. Oncol., 19: 1011-1019, 1983.
12. Nandi, S., DeOme, K. B., and Handin, M. Mammary tumor virus activity inblood and mammary tissues of C3H and BALB/cfC3H strains of mice. J.Nati. Cancer Inst., 35: 309-318, 1965.
13. Ritter, R. I., and Nandi, S. Time of appearance of viral activity in red bloodcells of mice infected naturally or artificially with mammary tumor virus. J.Nati. Cancer Inst.. 40: 1313-1317, 1968.
14. Bentvelzen, P., and Brinkhof, J. Organ distribution of exogenous murinemammary tumor virus as determined by bioassay. Eur. J. Cancer, 13: 241-245, 1977.
15. Ikehara, S., Pahwa, R. N., Fernandes, G., Hansen, C. T.. and Good, R. A.Functional T cells in athymic nude mice. Proc. Nati. Acad. Sci. USA, 81:886-888, 1984.
16. Ikehara, S., Shimizu, G., Yasumizu, K . Nakamura, T., Inaba. M., Inoue, S.,Oyaizu, V. Sugiura, K.. Maung Maung Oo, Hamashima, Y., and Good, R.A. Thymic rudiments are responsible for induction of functional T cells innu/nu mice. Thymus, 10: 193-205, 1987.
17. Sarkar, N. H., Pomenti, A. A., and Dion, A. S. Replication of mousemammary tumor virus in tissue culture. I. Establishment of a mouse mammary tumor cell line, virus characterization, and quantitation of virus production. Virology, 77: 12-30, 1977.
18. Southern, E. M. Detection of specific sequences among DNA fragmentsseparated by gel electrophoresis. J. Mol. Biol., 38: 503-517, 1975.
19. St. George, J. A., Cardiff, R. D., Young, L. J. T., and Faulkin, L. J.Immunohistochemical distribution of mouse mammary tumor virus antigensin BALB/cfC3H mammary epithelium. J. Nati. Cancer Inst., 63: 813-820,1979.
20. Raff, M. C. 9-bearing lymphocytes in nude mice. Nature (Lond.), 246: 350-351, 1973.
21. MacDonald, H. R., Lees, R. K., Sordat, B., Zaech, P., Maryanski, J. L., andBron, C. Age-associated increase in expression of the T cell surface markersThy-1, Lyt-1 and Lyt-2 in congenially athymic (nu/nu) mice: analysis byflow microfluorometry. J. Immunol., 126: 865-870, 1981.
22. Pauley, R. J., Parks, W. P., and Popko, B. J. Expression and demethylationof germinally-transmitted BALB/c mouse mammary tumor virus DNA inAbelson MuLV B-lymphoid cell lines. Virus Res., /: 381-400, 1984.
23. Gillette, R. W., Robertson, S., Brown, R., and Blackman, K. E. Expressionof mammary tumor virus antigen on the membranes of lymphoid cells. J.Nati. Cancer Inst., 53:499-505, 1974.
24. Charyulu, V., Sigel, M. M., Durden, D. L., and Lopez, D. M. Mousemammary tumor virus (MMTV) antigen(s) are present on B lymphocytes ofBALB/c mice. Int. J. Cancer, 24:813-818, 1979.
25. Tax, A., Ewert, D., and Manson, L. A. An antigen cross-reactive with gp52of mammary tumor virus is expressed on a B cell subpopulation of mice. J.Immunol., 130: 2368-2371, 1983.
26. Lopez, D. M., Charyulu, V., and Paul, R. D. B cell subsets in spleens ofBALB/c mice: identification and isolation of MMTV expressing andMMTV-responding subpopulations. J. Immunol.. ¡34:603-607, 1985.
27. Dudley, J., and Risser, R. Amplification and novel locations of endogenousmouse mammary tumor virus genomes in mouse T-cell lymphomas. J. Virol.,49:92-101, 1984.
28. Lopez, D. M., Pauley, R. J., and Lozzio, B.B. Interaction of the heterozygousnude gene with the asplenia trait in mammary tumorigenesis. J. Exp. Med.,161:629-634, 1985.
29. Cohen, J. C., Majors, J. E., and Varmus, H. E. Organization of mousemammary tumor virus-specific DNA endogenous to BALB/c mice. J. Virol.,32:483-496, 1979.
30. Knepper, J. E., Medina, D., and Butel, J. S. Differential expression ofendogenous mouse mammary tumor virus genes during development of theBALB/c mammary gland. J. Virol., 51: 518-521, 1986.
6559
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

1988;48:6555-6559. Cancer Res Airo Tsubura, Muneo Inaba, Shunsuke Imai, et al.
in VivoTumor Virus (Milk Factor) to Mammary Gland Cells Intervention of T-Cells in Transportation of Mouse Mammary
Updated version
http://cancerres.aacrjournals.org/content/48/22/6555
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/48/22/6555To request permission to re-use all or part of this article, use this link
Research. on February 15, 2021. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from