Embed Size (px)
Citation preview

INCIPIENT GENOME DIFFERENTIATION IN GOSSYPIUM. 111. COMPARISON OF CHROMOSOMES O F G. HZRSUTUM
AND ASIATIC DIPLOIDS USING HETEROZYGOUS TRANSLOCATIONS
MARGARET Y. MENZEL AND CLARE A. HASENKAMPF
Department of Biological Science, Florida State Uniuersity, Tallahassee, Florida 32306
AND
JAMES McD. STEWART
U . S. Department of Agriculture, Science and Education Administration, Agricultural Research, Department of Plant and Soil Science, University of Tennessee,
Knoxuille, Tennessee 37916
Manuscript received May 6, 1981 Revised copy accepted October 5, 1981
ABSTRACT
Hybrids between upland cotton (G. hirsutum, genome constitution 2A,Dh) and either A-genome or D-genome diploid species exhibit 26 paired and 13 unpaired chromosomes at metaphase I. The A , and D, genomes are therefore considered homoeologous with those of the respective diploids. Previous studies, nevertheless, revealed a low level of (“incipient”) differentiation between Dh and various diploid D genomes. The diploid A genomes have been regarded as more closely homologous to A, on the basis of low preferential pairing and autotetraploid segregation ratios in allohexap1oids.-The present study ad- dressed the following questions: Are the diploid A genomes differentiated from A, in meiotic homology? If so, is the differentiation manifested equally by all 13 chromosomes or is it localized in certain chromosomes?-Three diploid A-genome lines representing G. herbaceum and G. arboreum were hybridized by in ouulo culture of embryos (1) with a standard line of G. hirsutum, which differs from G. herbaceum by two and from G. arboreum by three naturally occurring reciprocal translocations involving chromosomes 1-5, and (2) with six lines homozygous for experimental translocations involving chromosomes 6, 7, IO, 11, 12 and 13. Chiasma frequencies in hybrids were compared with those in appropriate G. hirsutum controls. In every comparison overall chiasma frequencies were slightly lower i n the hybrids. Therefore A , appears to be differentiated from the diploid A genomes. No localized differentiation was de- tected in chromosomes marked by experimental translocations. The differentia- tion may be localized mainly in chromosomes 4 and 5.
T H E allotetraploid species of Gossypium, including G. hirsutum L. (upland cotton), all have the genome constitution ,?(AD). The 26 individual chromo-
somes of the allotetraploid genomes cannot at present be identified morphologi- cally at either mitosis or meiosis in normal material, and linkage maps are incom- pletely developed even for G. hirsutum (KOHEL 1978). However, chromosome
Genetics 100: 89-103 January, 1952.

90 M. Y. MENZEL, C . A. HASENKAMPF A N D J. M. STEWART
translocations available in G. hirsutum mark chromosomes 1-13 of the Ah and chromosomes 14-25 of the Dh genomes of G. hirsutum (BROWN 1980). At pres- ent no translocation is available for Dh chromosome 26. The translocated chromo- somes form the expected heterozygous configurations in interspecific hybrids with species carrying other A, D or (AD) genomes, and may be used to study chiasma frequencies in specific chromosome regions. The rationale for using data from such studies to investigate the initial stages in the evolution of meiotic chromosome nonhomology has been developed in some detail in earlier papers (MENZEL 1955; MENZEL, BROWN and NAQI 1978; HASENKAMPF et al. 1979; HASENKAMPF and MENZEL 1980).
Hybrids between allotetraploid species and the two diploid A-genome species, G. hmbaceum L. (2A,) and G. arboreum L. (2Ap), have been notoriously diffi- cult to obtain. However, recent technical improvements have made it possible to produce such triploid hybrids readily (STEWART 1979; STEWART and HSU 1977, 1978a, 1978b; LIANG et al. 1978). These advances made it practicable to produce hybrids between appropriate G. hirsutum translocation lines and the A-genome diploid species.
We report here a comparison of chiasma frequencies in triploid hybrids of genome constitution (AD)A and in control plants of G. hirsutum and G. ar- boreum. Six different translocations involving chromosomes 6, 7,10,11, 12 and 13 of the Ah genome were used to mark specific chromosome regions in hybrids and controls. The following questions will be addressed: (1) Has any divergence in meiotic homology occurred between the Ah genome and A, or A,? (2) If SO,
does it affect all chromosomes similarly (generalized genome differentiation) or to different extents in individual chromosomes and regions (localized differentia- tion) ?
Earlier studies used preferential pairing and genetic segregation ratios in ex- perimental allohexaploids to assess genome divergence (GERSTEL 1956; PHILLIPS 1962, 1964; PHILLIPS and GERSTEL 1959; SARVELLA 1958). Considerable evi- dence was found for divergence between Dh and various D-genome wild diploid species. However, no significant divergence was noted between A genomes from diploids and those from allotetraploids (G. hirsutum and G. barbadense L.) .
D-genome divergence was confirmed by study of chiasma frequencies in triploid hybrids of genome constitution (AD)D, (MENZEL, BROWN and NAQI 1978; HASENKAMPF et al. 1979). HASENKAMPF and MENZEL (1980) also found evidence of a low level of divergence, probably affecting both A and D genomes, between G. hirsutum and the wild allotetraploids G. mustelinum Miers ex Watt and G. tomentosum Nuttal ex Seeman.
MATERIALS A N D METHODS
The following translocations were used (MENZEL and BROWN 1978b; BROWN 1980; BROWN et al. 1981): T(H6L;HlOR) line Z9-9; T(H7R;HI IR) line 1052; T(H7L;HIZR) line 1043; T(HI0R;HZIR) line 2785; T(HIIR;H12L) line 6-5M; T ( H I l R ; H I 3 L ) line 10-5Kb. (For brevity, these will frequently be referred to hereinafter as T6;IO, T7;11, etc.) The inbred line TM-1, derived from ‘Deltapine 14’ by inbreeding (KOHEL, RICHMOND and LEWIS 1970), was used as the standard parent to produce all of the G. hirsutum heterozygous controls.
Three types of A-genome parents were used to obtain the interspecific hybrids: G. herbaceum cv. ‘Jayahdar’ (line AIJ), and G. arboreum CY. ‘Nanking’ (line ASN), both from station stocks

COTTON A GENOMES 91
at Knoxville; and a strain purporting to be the wild form of G. herbaceum srp. africanum (Watt) Mauer, obtained as 1977 greenhouse open pollinated seeds from the species collection at Stone- d e , Mississippi (line AIA). The intent was to compare wild and cultivated G. herbaceum and cultivated G. arboreum (which is unknown in the wild) with regard to the similarity of their genomes to the A, genome of G. hirsutum.
Each experiment consisted of hybrids (“Tt”) between a homozygous translocation line (‘‘TI?’) and TM-1, AIA, AIJ and A2N (“tt”). Twenty-four pedigrees were represented among the various experiments. However, hybrids of T11;12 x AIJ did not survive transfer to Tallahas- see.
The standard A , and A , genomes differ by two and the A, and A , genomes by three naturally occurring reciprocal translocations (BEASLEY 1942; GERSTEL 1953; GERSTEL and SARVELLA 1956; MENZEL and BROWN 1954; ENDRIZZI and BROWN 1962). In analyses of the hybrids, the multi- valents due to the spontaneous translocations were confounded with those due to the transloca- tion introduced via the TT parent. Therefore, it was necessary to include as controls hybrids between the A-genome diploids and G. hirsutum with the standard end arrangements. ‘Deltapine 14.‘ (hereinafter referred to as DPL) rather than TM-1 was used as the G. hirsutum parent of these control triploids.
Hybrids were obtained by in ouulo culture of embryos. Ovules were removed from the ovaries on the second or third day after controlled pollination and cultured in the dark at 30-32” on a modified Beasley and Ting medium (STEWART and Hsu 1978a, 1978b; STEWART 1979). After germination seedlings were transferred to a low-salt rooting medium until large enough to be planted in small pots.
The hybrids were produced at Knoxville and transposed to Tallahassee as potted plants with three or four leaves. They were transplanted first to 6-inch pots in the greenhouse and later to rows in the cotton garden at the Mission Road Research Facility at appropriate seasons. Pre- liminary cytological examination in the 1978 fall garden confirmed that they were hybrids. The pots were brought into the green house to overwinter and returned to the experimental garden in spring 1979. Most of the data reported here were taken from buds collected in October and November 1979. However, controls for the T11;13 translocation were not available until Spring 1980. Hence hybrids involving T11;13 were again overwintered in the greenhouse. Buds for this experiment were collected from the experimental garden in May and June 1980. A second set of control data from TM-1 and the DPL x AIA and DPL x A2N hybrids was recorded from the same season.
Slides were prepared by a standard acetocarmine anther squash technique (MENZEL and BROWN 1978a). Slides from each of two plants from each pedigree were scanned systematically until 50 pollen mother cells (PMC‘s) at metaphase I (MI) had been recorded or the material was exhausted. In a few pedigrees, only one plant produced analyzable cells. In these cases two samples were taken from the same plant on different days.
For each cell, the number of univalents, the number of one- and two-chiasma bivalents, the number of multivalents of each type, and the number and location of chiasmate regions in each multivalent were recorded, and the presence of the expected number of chromosomes was verified. Chromosome configurations found in each cell were recorded on file cards and later entered into a computer. From the computerized data the various statistics were generated. The analyses of variance used to compare samples were carried out using the SPSS statistical packages available through the Florida State University Computing Center.
Two types of statistical comparisons were made. First, the total number of chiasmate arms per pair of A genomes (hereinafter referred to as “total chiasma frequency” for brevity) was com- pared. Then a more detailed comparison was made for specific chromosomes and chromosome regions marked by the translocation.
RESULTS A N D DISCUSSION
Chromosome configurations in parents and hybrids: Four of the chromosomes of A , and Ah, corresponding to chromosomes H2, H3, H4 and H5 of the standard A?* genome, differ in their end arrangements so as to form 2 IV’s (one involving
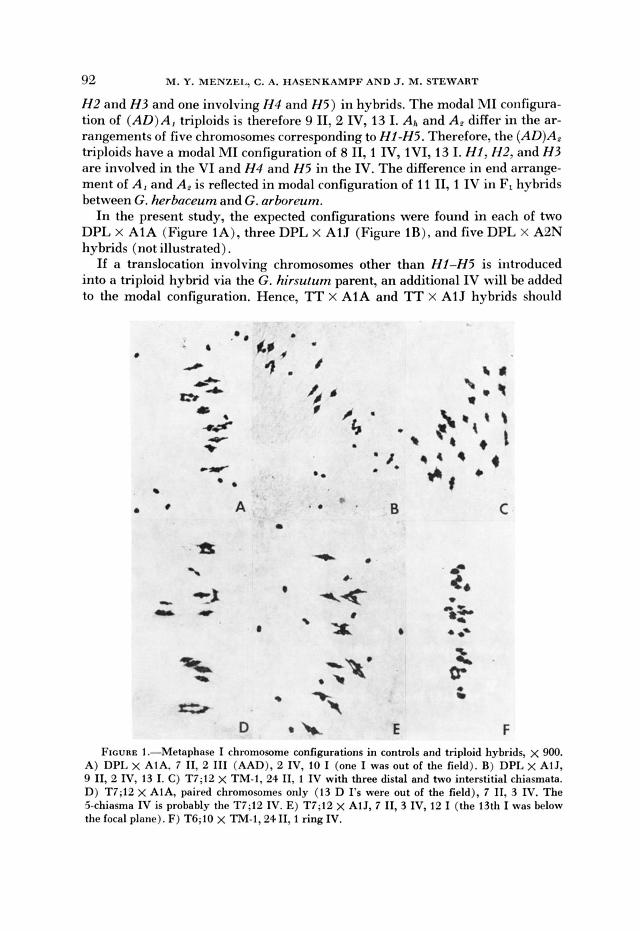
92 M. Y. MENZEI, C. A. HASENKAMPF AND J. M. STEWART
H2 and H3 and one involving I14 and H5) in hybrids. The modal MI configura- tion of (AD)A, triploids is therefore 9 11, 2 IV, 13 I. Ai and A, differ in the ar- rangements of five chromosomes corresponding to HI-H5. Therefore, the (AD)A, triploids have a modal MI configuration of 8 11, 1 IV, IVI, 13 I. HI, H2, and H3 are involved in the VI and H 4 and H5 in the IV. The difference in end arrange- ment of A, and A, is reflected in modal configuration of 11 11,l IV in F, hybrids between G. herbaceum and G. arboreum.
In the present study, the expected configurations were found in each of two DPL X A1A (Figure lA), three DPL x A1J (Figure lR), and five DPL X A2N hybrids (not illustrated).
If a translocation involving chromosomes other than HI-H5 is introduced into a triploid hybrid via the G. hirsutum parent, an additional IV will be added to the modal configuration. Hence, TT x AlA and TT x AlJ hybrids should
d - 0
It=r D F
FIGURE 1 .-Metaphase I chromosome configurations in controls and triploid hybrids, x 900. A) DPL x AIA, 7 11, 2 111 (AAD), 2 IV, 10 I (one I was out of the field). B) DPL x AlJ, 9 11, 2 IV, 13 I. C) T7;13 x TM-1, 24 11, 1 IV with three distal and two interstitial chiasmata. D) T7;12 x AIA, paired chromosomes only (13 D 1's were out of the field), 7 11, 3 IV. The 5-chiasma IV is probably the T7;12 IV. E) T7;12 x AtJ, 7 11.3 IV, 12 I (the 13th I was below the focal plane). F) T6;10 x TNI-1,2411,1 ring IV.
COTTON A GENOMES 93
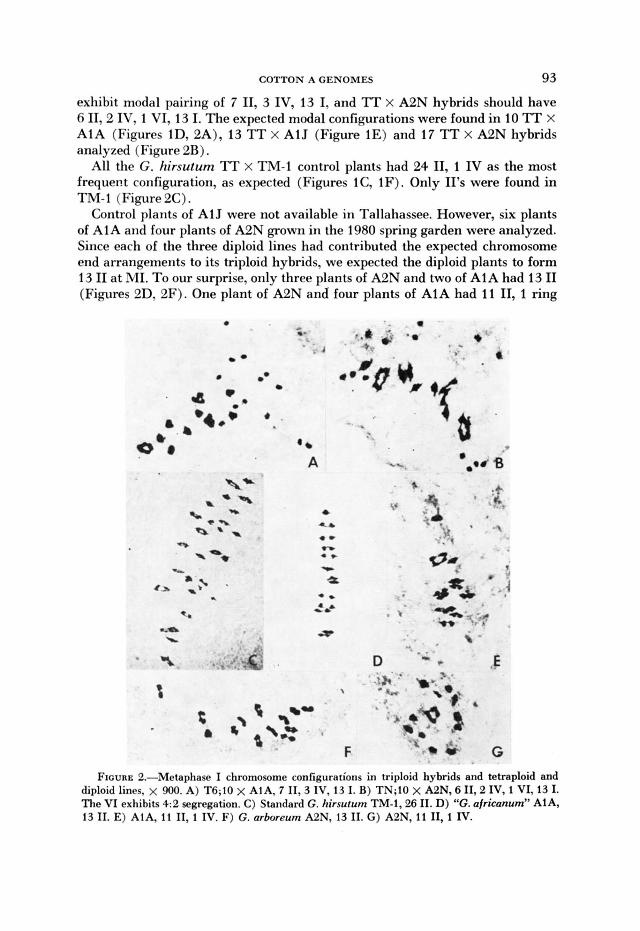
exhibit modal pairing of 7 11, 3 IV, 13 I, and ‘IT x A2N hybrids should have 6 IT, 2 IV, 1 VI, 13 I. The expected modal configurations were found in 10 TT X A1A (Figures lD, 2A), 13 TT x A1J (Figure 1E) and 17 TT X A2N hybrids analyzed (Figure 2B).
All the G. hirsutum ‘IT x TM-1 control plants had 24 11, 1 IV as the most frequem configuration, as expected (Figures lC, 1F). Only 11’s were found in TNI-1 (Figure 2C).
Control plants of A1 J were not available in Tallahassee. However, six plants of AlA and four plants of A2N grown in the 1980 spring garden were analyzed. Since each of the three diploid lines had contributed the expected chromosome end arrangements to its triploid hybrids, we expected the diploid plants to form 13 I1 at M I . To our surprise, only three plants of A2N and two of AlA had 13 I1 (Figures 2D, 2F). One plant of A2N and four plants of A1A had 11 IT, 1 ring
FIGURE 2.-Metaphase I chromosome configurations in triploid hybrids and tetraploid and diploid lines, x 900. A) T6;lO x AlA, 7 II ,3 IV, 13 I. B) TN;lO x A2N, 6 II ,2 IV, 1 VI, 13 I. The VI exhibits 4:2 segregation. C) Standard G. hirsutum TM-1,26 11. D) “G. africunum” AIA, 13 11. E) AlA, 11 11, 1 IV. F) G. arboreum A2N, 13 11. G) A2N, 11 11, I IV.

94 M. Y. MENZEI,, C. A. HASENKAMPF AND J. M. STEWART
IV in nearly every cell (Figures 2E. 2G). The appearance of heterozygous trans- locations in the A1A and A2N diploids is puzzling since other plants from the same strains apparently transmitted only the standard end arrangements to the hybrids.
Herbarium specimens (deposited at FSU) from each of the analyzed A1A and A2N plants were examined by J. E . ENDRIZZI, L. L. PHILLIPS and P. A. FRYXELL. ENDRIZZI also examined the live plants in the garden in September 1980. They concluded that line A1A is not typical G. h. africcmum. The plants appear to be segregating to some extent for leaf shape, indument, habit and bract traits in a manner suggesting introgression of genes from G. arboreum. The most probable, though not the only, explanation for the presence of a IV in some plants is introgression of the A, end arrangement via an earlier inadvertent hy- bridization which must have occurred before we received the stock.
The plants of A2N were more constant in phenotype and were judged morpho- logically acceptable as G. arboreum. If the IV in some A2N plants were the result of introgression of the A , end arrangement, the history of the line requires that it represent a separate hybridization event from that involving A1 A. Avail- able information concerning the history of these lines does not shed any further light on the question. It cannot be ruled out that spontaneous translocations in- volving other chromosomes have arisen in the diploid species. Further study of the identity of the chromosomes involved in the IV’s is continuing. However, in each case only the standard A , or A, end arrangement expected was transmitted to each of the triploid hybrids included in the study.
Since the purity and species identity of two of the diploid lines is called in question, our original objective of comparing the degree of divergence in meiotic homology of Ah from wild and cultivated G. herbaceum and cultivated G. ar- boreum was somewhat compromised. However, it is still feasible to determine whether the genomes of the three diploid lines, viewed as representatives of the A genomes as a group, show any divergence from A?,.
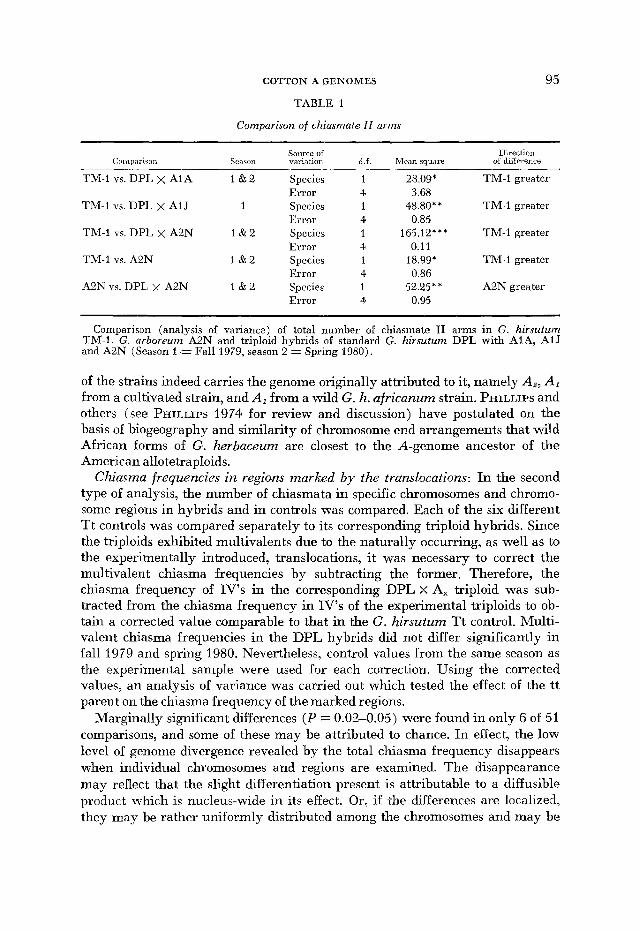
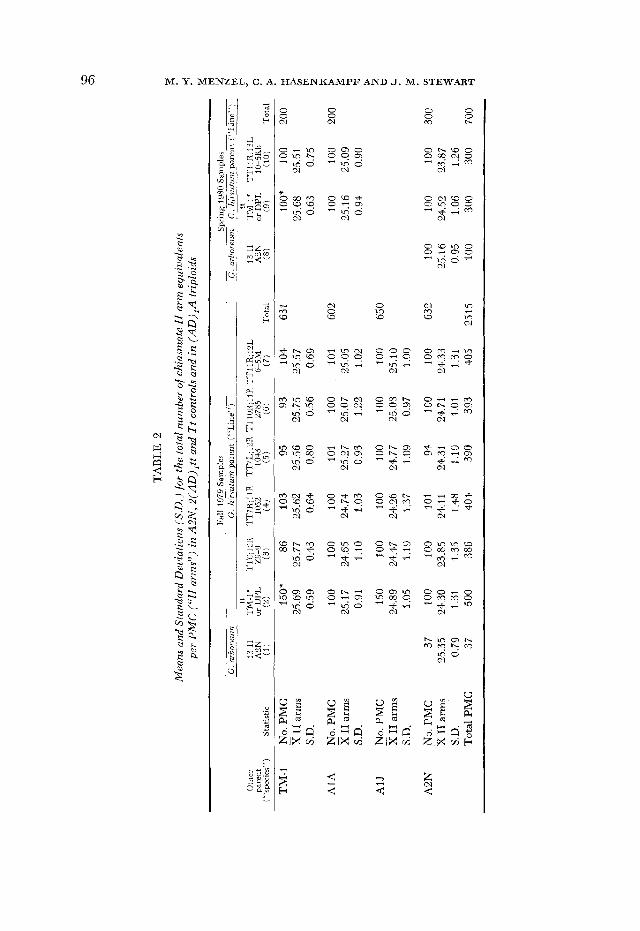
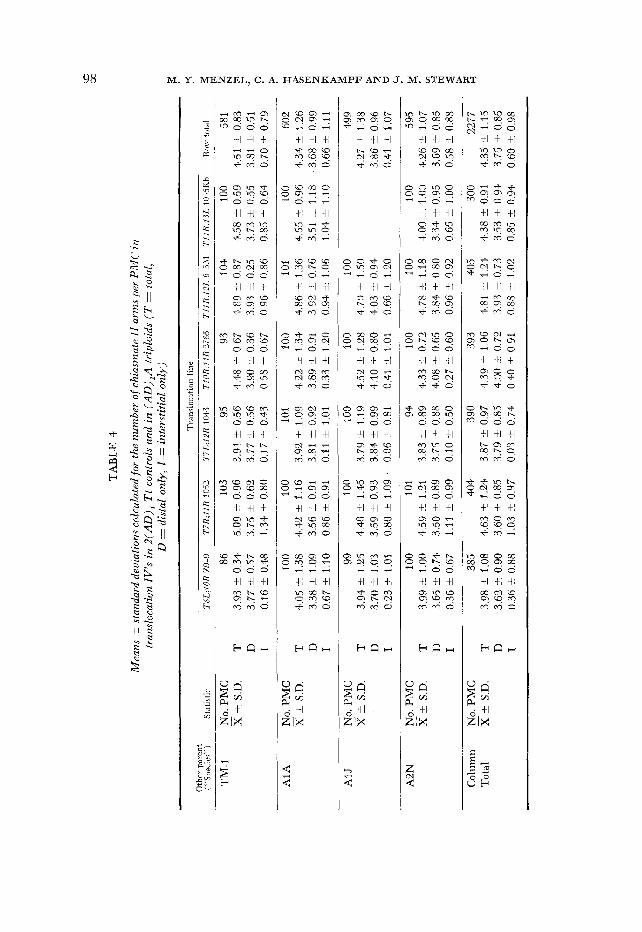
Total chiasma frequency: In the first type of analysis, the total estimated A-genome chasma frequency in TM-1 controls was compared to the total ob- served in each of the DPL x A1 A, x A1J and x A2N triploids (Table 1 ) . Each comparison consisted of an analysis of variance in which the effect of the species used as the pollen parent was assessed. Where data for two seasons were avail- able, b3th cets of data were used and the effect of season was also assessed. Then each set of six Tt tripbid hybrids having the same pollen parent was compared with the set of six Tt G. hirsutum controls, and the effects of species, TT line and the interaction between the two were assessed (Table 2, 3 ) .
In every case the hybrids had significantly fewer chiasmata regardless of the season of the sample. The difference in overall chiasma frequency between con- trols and triploid hybrids was small (2-6%) but highly significant (Table 3 ) . Since the overall chiasma frequency is consistently higher in controls than in hybrids, it may be concluded that a low level of divergence does exist between the Ah genome and the genomes carried by AIA, A1J and A2N. The order of divergence, A2N > A1J > AIA, corresponds to what might be expected if each
COTTON A GENOMES 95
TABLE 1
Comparison of chiasmate II arms
TM-1 VS. DPL X AIA 1 & 2 Species
TM-I vs. DPL x AIJ 1 Species
TM-1 VS. DPL X A2N 1 & 2 Species
TM-I vs. A2N 1 & 2 Species
AQN vs. DPL, x A2N 1 & 2 Species
Error
Error
Error
Error
Error
1 4 1 4 1 4 1 4 1 4
28.09* TM-1 greater
48.80** TM-1 greater
165.12*** TM-1 greater
18.99* TM-1 greater
52.25** A2N greater
3.68
0.85
0.11
0.86
0.95
Comparison (analysis of variance) of total number of chiasmate I1 arms in G. hirsutum TM-1, G. arboreum A2N and triploid hybrids of standard G. hirsutum DPL with AIA, AIJ and A2N (Season 1 = Fall 1979, season 2 = Spring 1980).
of the strains indeed carries the genome originally attributed to it, namely A,, AI from a cultivated strain, and A, from a wild G. h. africanum strain. PHILLIPS and others (see PHILLIPS 1974 for review and discussion) have postulated on the basis Qf biogeography and similarity of chromosome end arrangements that wild African forms of G. herbaceum are closest to the A-genome ancestor of the American allotetraploids.
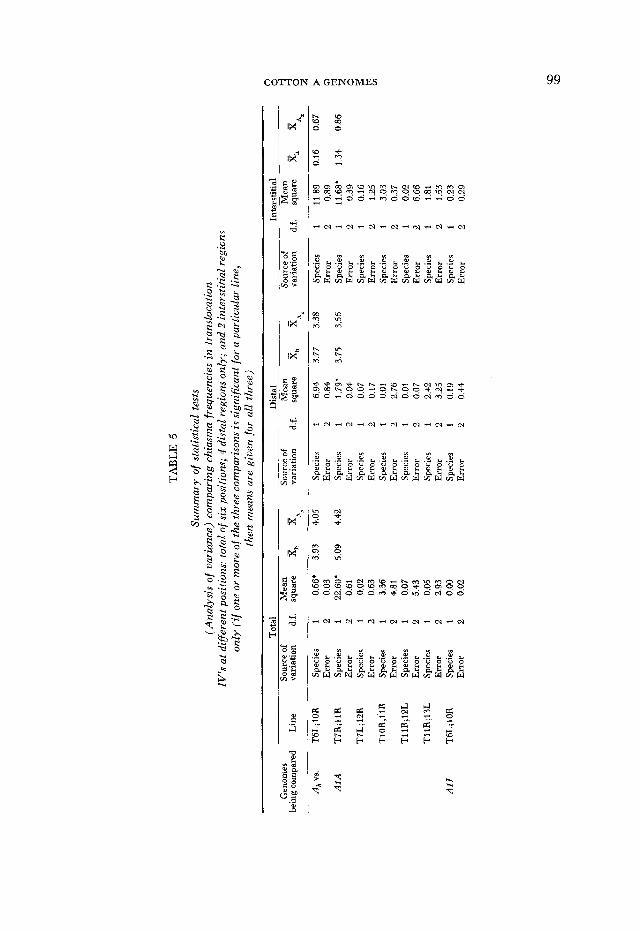
Chiasma frequencies in regions marked by the trdnslocat.ions: In the second type of analysis, the number of chiasmata in specific chromosomes and chromo- some regions in hybrids and in controls was compared. Each of the six different Tt controls was compared separately to its corresponding triploid hybrids. Since the triploids exhibited multivalents due to the naturally occurring, as well as to the experimentally introduced, translocations, it was necessary to correct the multivalent chiasma frequencies by subtracting the former. Therefore, the chiasma frequency of IV’s in the corresponding DPL x A, triploid was sub- tracted from the chiasma frequency in IV’s of the experimental triploids to ob- tain a corrected value comparable to that in the G. hirsutum Tt control. Multi- valent chiasma frequencies in the DPL hybrids did not differ significantly in fall 1979 and spring 1980. Nevertheless, control values from the same season as the experimental sample were used for each correction. Using the corrected values, an analysis of variance was carried out which tested the effect of the t t parent on the chiasma frequency of the marked regions.
Marginally significant differences (P = 0.02-0.05) were found in only 6 of 51 comparisons, and some of these may be attributed to chance. In effect, the low level of genome divergence revealed by the total chiasma frequency disappears when individual chromosomes and regions are examined. The disappearance may reflect that the slight differentiation present is attributable to a diffusible product which is nucleus-wide in its effect. Or, if the differences are localized, they may be rather uniformly distributed among the chromosomes and may be
96 M. Y. MENZEL, C. 4. HASENKAMPF AND J. M. STEWART
0 0 N
0 0 N
* 0" o w *
m o m o N N
z:cr\rq z . - s
N 4 1
u?
1 0 N
- 0 N VI (0
U) W
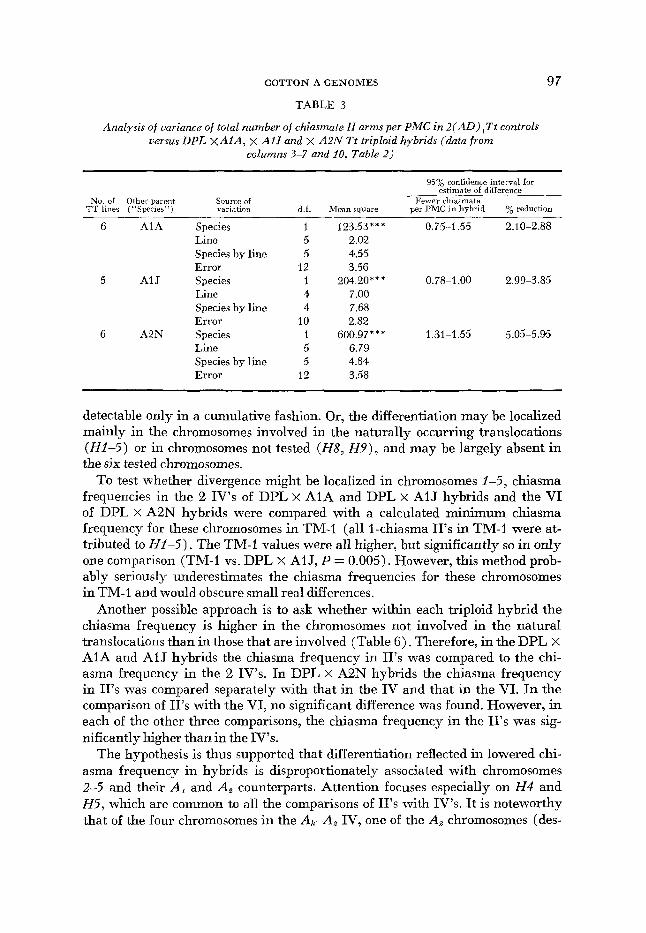
COTTON A GENOMES 97 TABLE 3
Analysis of variance of total number of chiasmate II arms per PMC in 2(AD),Tt controls versus DPL X A I A , x AlJ and X A2N Tt triploid hybrids (data f rom
columns 3-7 and 10, Table 2)
95% confidence interval for estimate of difference
No. of Olhei parent Source of Fewer chiarmata ’IT lines (“Species”) variation d.f. Mean square per PMC in hybrid % reduction
6 AIA Species 1 Line 5 Species by line 5 Error 12
5 AIJ Species 1 Line 4 Species by line 4 Error 10
6 A2N Species I Line 5 Species by line 5 Error 12
123.53** * 0.75-1.55 2.10-2.88 2.02 4a.55 3.56
7.00 7.68 2.82
6.79 4.84 3.58
204.20*** 0.78-1.00 2.99-3.85
600.97** * 1.31-1.55 5.05-5.96
detectable only in a cumulative fashion. Or, the differentiation may be localized mainly in the chromosomes involved in the naturally occurring translocations (HI-5) or in chromosomes not tested (Ha , H 9 ) , and may be largely absent in the six tested chromosomes.
To test whether divergence might be localized in chromosomes 1-5, chiasma frequencies in the 2 IV’s of DFL x A1A and DPL x A1J hybrids and the VI of DPL x A2N hybrids were compared with a calculated minimum chiasma frequency for these chromosomes in TM-1 (all l-chiasma 11’s in TM-1 were at- tributed to HI-5) . The TM-1 values were all higher, but significantly so in only one comparison (TM-1 vs. DPL X A1 J, P = 0.005). However, this method prob- ably seriously underestimates the chiasma frequencies for these chromosomes in TM-1 and would obscure small real differences.
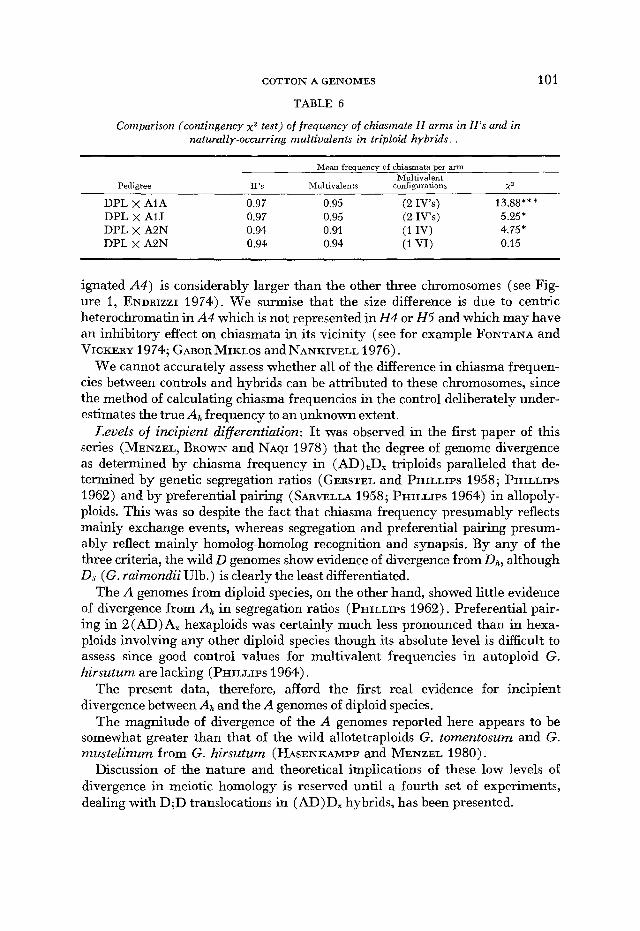
Another possible approach is to ask whether within each triploid hybrid the chiasma frequency is higher in the chromosomes not involved in the natural translocations than in those that are involved (Table 6). Therefore, in the DPL x A1A and A1J hybrids the chiasma frequency in 11’s was compared to the chi- asma frequency in the 2 IV’s. In DPL X A2N hybrids the chiasma frequency in II’s was compared separately with that in the IV and that in the VI. In the comparison of II’s with the VI, no significant difference was found. However, in each of the other three comparisons, the chiasma frequency in the 11’s was sig- nificantly higher than in the IV’s.
The hypothesis is thus supported that differentiation reflected in lowered chi- asma frequency in hybrids is disproportionately associated with chromosomes 2-5 and their A , and A, counterparts. Attention focuses especially on H4 and H5, which are common to all the comparisons of II’s with IV’s. It is noteworthy that of the four chromosomes in the Ah-A, IV, one of the A, chromosomes (des-
98 M. Y. MENZEL, C. A. HASENKAMPF AND J. M. STEWART
c . Y
COTTON A GENOMES 99

100 M. Y. MENZEL, C. A. HASENKAMPF A N D J. M. STEWART
I I IX”
4- IX
IX“
8 d
01 n 9 ? on
n
2
0
* h. 8 a‘
COTTON A GENOMES 101
TABLE 6
Comparison (contingency x g test) of frequency of chiasmate I1 arms in Il’s and in natumlly-occurring multivalents in triploid hybrids
Mean frequency of chiasmata per arm - ~ _ _ _ Multivalent
Pedigree 11’s Multi1 dents configurations x? DPL x A1A 0.97 0.95 (2 Iv’s) 13.88*** DPL x AIJ 0.97 0.95 (2 IVS) 5.25’
DPL x A2N 0.94 0.94 (1 VI) 0.15 DPI, x A2N 0.94 0.91 (1 IV) 4.75 *
ignated A4) is considerably larger than the other three chromosomes (see Fig- ure 1, ENDRIZZI 1974). We surmise that the size difference is due to centric heterochromatin in A4 which is not represented in H 4 or H5 and which may have an inhibitory effect on chiasmata in its vicinity (see for example FONTANA and VICKERY 1974; GABOR MIKLOS and NANKIVELL 1976).
We cannot accurately assess whether all of the difference in chiasma frequen- cies between controls and hybrids can be attributed to these chromosomes, since the method of calculating chiasma frequencies in the control deliberately under- estimates the true Ah frequency to an unknown extent.
Levels of incipient differentiation: I t was observed in the first paper of this series (MENZEL, BROWN and NAQI 1978) that the degree of genome divergence as determined by chiasma frequency in (AD),D, triploids paralleled that de- termined by genetic segregation ratios (GERSTEL and PHILLIPS 1958; PHILLIPS 1962) and by preferential pairing (SARVELLA 1958; PHILLIPS 1964) in allopoly- ploids. This was so despite the fact that chiasma frequency presumably reflects mainly exchange events, whereas segregation and preferential pairing presum- ably reflect mainly homolog-homolog recognition and synapsis. By any of the three criteria, the wild D genomes show evidence of divergence from Dh, although D,5 (G. raimondii Ulb.) is clearly the least differentiated.
The A genomes from diploid species, on the other hand, showed little evidence of divergence from Ah in segregation ratios (PHILLIPS 1962). Preferential pair- ing in 2 (AD) A, hexaploids was certainly much less pronounced than in hexa- ploids involving any other diploid species though its absolute level is difficult to assess since good control values for multivalent frequencies in autoploid G. hirsutum are lacking (PHILLIPS 1964).
The present data, therefore, afford the first real evidence for incipient divergence between Ah and the A genomes of diploid species.
The magnitude of divergence of the A genomes reported here appears to be somewhat greater than that of the wild allotetraploids G. tomentosum and G. mustelinum from G. hirsutum (HASENKAMPF and MENZEL 1980).
Discussion of the nature and theoretical implications of these low levels of divergence in meiotic homology is reserved until a fourth set of experiments, dealing with D;D translocations in (AD)D, hybrids, has been presented.

102 M. Y. M E N Z E L , C. A. H A S E N K A M P F A N D J. NI. S T E W A R T
Supported in part by Grants No. DEB 77-24476 from the National Science Foundation and 5901-0410-8-0116-0 and 59-2123-11 -6360 from the Competitive Research Grants Office, Science and Education Administration, U.S.D.A. We thank META S. BROWN and others at the Texas Agri- cultural Experiment Station for supplying the cotton translocation stocks; the late VESTA L. MEYER of the Delta Branch Experiment State, Stoneville, Mississippi for supplying one of the diploid strains used; CECELIA Hsu, LESLIE B. CAMPBELL, KAREN GRAGFIUS, ANNE THISTLE, SUSAN WOODYARD, JAMES COX, DANA BRYAN and K. L. RICHMOND for technical assistance, and JOE B.ARRETT~ of the F. S. U. Statistical Consulting Center for help with statistical analysis.
L I T E R A T U R E C I T E D
BEASLEY, J. O., 1942
BROWN, M. S., 1980
BROWN, M. S., M. Y. MENZEL, C. A. HASENKAMPF and S. NAQI, 1981
Meiotic chromosome behavior in species, species hybrids, haploids, and induced polyploids in Gossypium. Genetics 27: 25-54.
Identification of the chromosomes of Gossypium hirsutum L. by means of translocations. J. Heredity 71 : 266-274.
Chromosome configura- tions and orientations in fifty-eight heterozygous translocations in Gossypium hirsutum. J. Hered. 72: 161-171.
ENDRIZZI, J. E., 1974 Alternate1 and alternate-2 disjunctions in heterozygous reciprocal trans- locations. Genetics 77: 55-60.
ENDRIZZI, J. E. and M. S. BROWN, 1962 Identification of a ring of four and two chains of three chromosomes from the Gossypium arboreum-hirsutum ring of six. Can. J. Genet. Cytol. 4: 458-468.
Heterochromatin content and chi,asma distribution in the megameric chromosomes of Stethophymn gracilis and S. linearum (Orthoptera: Acrididae). Chromosoma (Berl.) 46: 375-395.
GABOR MIKLOS, G. L., and R. N. NANKIVELL, 1976 Telomeric satellite DNA functions in regu- lating recombination. Chromosoma (Berl.) 56: 143-167.
GERSTEL, D. U,, 1953 Chromosomal translocations in interspecific hybrids of the genus Gos- sypium. Evolution 7: 234-244. -, 1956 Segregation in new allopolyploids of Gossypium I. The R, locus in certain New World-Wild American hexaploids. Genetics 41 : 31-M.
Segregation of synthetic amphiploids in Gossypium
Additional observations on chromosomal translocations
FONTANA, P. G., and V. R. VICKERY, 1974
GERSTEL, D. U,,, and L. L. PHILLIPS, 1958
GERSTEL. D. U. and P. A. SARVELLA, 1956 and Nicotiana. Cold Spring Harbor Symp. Quant. Biol. 23: 225-237.
in cotton hybrids. Evolution 10: 408-414.
HASENKAMPF, C. A., and M. Y. MENZEL, 1980 Incipient genome differentiation in Gossypium 11. Comparison of 12 chromosomes in G. hirsutum, G. mustelinum and G. tomentosum using heterozygous translocations. Genetics 95: 971-983.
Progress in detecting incipient genome differentiation in Gossypium by means of the Rrown chromosome trans- locations. Proc. 1979 Beltwide Cotton Production Research Conferences (Phoenix) : 79-80.
Linkage tests in upland cotton. 111. Crop Sci. 18: 844.-847.
HASENKAMPF, C. A., M. Y. MENZEL, M. S. BROWN and S. NAQI, 1979
KOHEL, R. J., 19'78
KOHEL, R. J., T. R. RICHMOND and C. F. LEWIS, 1970
LIANG, C. L., C. W. SUN, T. L. LIU and J. C. CHIANG, 1978
MENZEL, M. Y., 1955
Texas Marker-I. Description of a genetic
Studies on interspecific hybridiza-
A cytological method for genome analysis in Gossypium. Genetics 40:
standard for Gossypium hirsutum L. Crop Sci. 10: 670-671.
tion in cotton. Scientia Sinica 21: 545-555.
214233.

COTTON A GENOMES 103
MLNZEL, M. Y., and M. S. BROWN, 1954 The significance of multivalent formation in three- species Gossypium hybrids. Genetics 39 : 546-557. - , 1978a Genetic lengths and breakpoints in twelve chromosomes of Gossypium hirsutum involved in ten reciprocal trans- locations. Genetics 88: 541-558. - , 1978b Reciprocal chromosome translocations in Gossypium hirsutum: Arm location of breakpoints and recovery of duplication-deficiencies. 5. Hered. 69 : 383-390.
Incipient genome differentiation in Gossypium. I. Chromosomes 14, 15, 16,19, and 20 assessed in G. hirsutum, G . raimcndii and G. Eobatum with seven A-D translocations. Genetics 90: 133-14Q.
PHILLIPS, L. L., 1962 Segregation in new allopolyploids of Gossypium. IV. Segregation in New World x Asiatic and New World x American hexaploids. Am. J. Bot. 49: 51-57. -, 1964 Segregation in New World allopolyploids of Gossypium. V. Multivalent formation in New World x Asiatic and New World x wild American hexaploids. Am. J. Bot. 51: 324-329. -, 1974 Cotton (Gossypium). pp. 111-133. In: Handbook of Ge- netics 2: Plants, Plant Viruses, and Protists. Edited by R. C. KING. Plenum Press, New York.
PHILLIPS, L. L., and D. U. GERSTEL, 1959 Segregation in new allopolyploids of Gossypium. 111. Leaf stape segregation in hexaploid hybrids of New World cottons. J. Hered. 50: 103-108.
SARVELLA, P., 1958 Multivalent formation and genetic segregation in some allopolyploid Gossypium hybrids. Genetics 43: 601-619.
STEWART, J. McD., 1979 Use of ovule cultures to obtain interspecific hybrids of Gossypium. Symp. Southern Sect. Am. Soc. Plant Physiol., New Orleans. Feb. 8, 1979: 44-56.
STEWART, J. McD., and C. L. HSU, 1977 In-ouulo embryo cultures and seedling development of cotton (Gossypium hirsutum L.). Planta 137: 113-117. - , 1978a Interspecific Gos- sypium hybrids through ovule culture. Proc. 1978 Beltwide Cotton Production Research Conferences (Dallas) : 68. - , 1978b Hybridization of diploid and tetraploid cottons through in-ouulo embryo cultures. J. Hered. 69 : 4.04408.
Corresponding editor: R. L. PHILLIPS
MENZEL, M. Y., M. S. BROWN and S . NAQI, 1978





























