Embed Size (px)
Citation preview

Immunocytochemical Localization of Growth Factorsand Intermediate Filaments During the Establishmentof the Porcine PlacentaELISABETH PERSSON1* AND HERIBERTO RODRIGUEZ-MARTINEZ21Department of Anatomy and Histology, Faculty of Veterinary Medicine, Swedish University of Agricultural Sciences,S-750 07 Uppsala, Sweden2Department of Obstetrics and Gynaecology, Faculty of Veterinary Medicine, Swedish University of Agricultural Sciences,S-750 07 Uppsala, Sweden
KEY WORDS endometrium; fetal membranes; early pregnancy; immunohistochemistry; mor-phology; swine
ABSTRACT The immunocytochemical localization of insulin-like growth factor I (IGF-I) andplatelet-derived growth factorA (PDGF-A) chain ligands, PDGF-chain receptors, and the intermedi-ate filaments cytokeratin, desmin, and vimentin in the tissue reorganization and developmentduring the establishment of the porcine epitheliochorial placenta was studied at light and electronmicroscopic levels in sections of endometrium and/or placenta of gilts during early pregnancy up today 40 postmating. The endometrial epithelia (surface and glandular) as well as the trophoblastshowed PDGF-A and IGF-I labelling. The only temporal difference was seen regarding IGF-Ilabelling, this being more uniform in the glands during precontact and early contact stages (days7-12) compared to later stages. Cytokeratin labelling was conspicuous in all epithelia, including thetrophoblast. The endometrial stroma showed strong labelling for the PDGF receptors and theintermediate filament vimentin, staining being enhanced along with the increase in the vascularbed during the establishment of the placenta. The maternal endothelium immunoreacted to IGF-I,to both PDGF-A and PDGF receptors, to vimentin and to von Willebrand factor (Factor VIII).Desmin was also expressed in the capillary bed underneath the maternal surface epithelium. Inlarger blood vessels, desmin was detected in the smooth muscle layer but not in the endothelium.The vascular smooth muscle also reacted with IGF-I, PDGF-A, and PDGF-receptor antibodies.The results suggest the involvement of both growth factors, IGF-I and PDGF, in the early nutritionof the pig embryo and the tissue reorganization that encompasses placentation. IGF-I appearsto be related to the secretory tissue compartment (formation of histotrophe), whereas PDGFmight play a role in the reorganization of the stroma, particularly during placental angiogenesis.Desmin and von Willebrand factor immunolabelling appears to be useful to monitor the develop-ment of the subepithelial capillary bed in the porcine placenta.Microsc. Res. Tech. 38:165-175, 1997.r 1997 Wiley-Liss, Inc.
INTRODUCTIONPorcine embryos develop unattached in the uterine
lumen throughout a major part of the gastrulationprocess, experiencing a process of dramatic elongationof the trophoblast from days 10 to 12 postcoitum to theinitiation of the formation of the noninvasive epithelio-chorial placenta by day 15 (Dantzer, 1985). Two criticalevents are linked with the initiation of blastocystelongation, i.e., the secretion of oestrogens by theconceptuses (Perry et al., 1973) and the sequestering ofhistotroph produced by the stimulated, highly produc-ing endometrial glands (Geisert et al., 1982). Oestrogensecretion is related to the activation of major metabolicchanges in the endometrium, including the increase inglandular secretory activity and the mucosal foldingand development of the subepithelial vascular bed(Dantzer and Leiser, 1994; Geisert et al., 1990; Stro-band and Van der Lende, 1990).Although the initial survival of the embryos depends
on the availability of soluble nutrients in the lumen,
embryonic growth and development cannot be furthersustained solely by the histotrophe. Endometrial angio-genesis and vascularization of the fetal membranestogether with the intimate apposition of these tissuesfinally render the establishment of a functional pla-centa, which secures the nourishment of the rapidlydeveloping conceptuses (Dantzer, 1985, 1993).Direct and indirect control of conceptus growth is
modulated by many uterus-derived growth factors,which act as mitogens or modulate specific cell proteinsynthesis and secretion (Brigstock et al., 1989). Amongthe growth factors present in uterine tissues are theinsulin-like growth factors (IGFs), epidermal growth
Contract grant sponsor: Swedish Council for Forestry and Agricultural Re-search.*Correspondence to: Dr. Elisabeth Persson, Department of Anatomy and
Histology, Faculty of Veterinary Medicine, Swedish University of AgriculturalSciences, P.O. Box 7011, S-750 07 Uppsala, Sweden.Received 20 March 1995; Accepted 4 September 1995
MICROSCOPY RESEARCH AND TECHNIQUE 38:165–175 (1997)
r 1997 WILEY-LISS, INC.

factor (EGF), platelet-derived growth factor (PDGF),acidic and basic fibroblast growth factors (aFGF, bFGF),and transforming growth factors (TGF; Brigstock et al.,1989; Simmen and Simmen, 1991). These growth fac-tors act on their target cells via specific receptors, ofwhich the IGF type I receptor (Hofig et al., 1991), theEGF receptor (Zhang et al., 1992), and the PDGFreceptors (Rodriguez-Martinez et al., 1992; Ronnstrandet al., 1987; Terracio et al., 1988) have been shown to bepresent in the uterus of the pig.Vascular development follows two distinct mecha-
nisms, i.e., de novo, as in embryonic development(vasculogenesis), or by expansion from preexisting ves-sels, as is seen in the adult female genital organs(angiogenesis; Torry and Rongish, 1992). The establish-ment of the placenta seems to be accompanied by thesetwo mechanisms, the first in the fetal membranes andthe second in the endometrium (Torry and Rongish,1992). Several growth factors, including some of thosementioned above, appear to be involved as promoters ofangiogenesis (Folkman and Shing, 1992), a process thatcan be followed by the presence of cell type markerssuch as cytoskeletal proteins (Nehls et al., 1992; Pers-son et al., 1994) and the von Willebrand factor (Se-hested and Hou-Jensen, 1981), specific for endothelialcells.The present study attempted, therefore, to determine
the temporal localization of selected growth factors(ligands and receptors) and intermediate filamentsduring the establishment of the epitheliochorial pla-centa of the pig. Immunocytochemistry at light andelectron microscopic levels was used to localize reactionsites for IGF-I; PDGF-A; and PDGF-chain receptors;the intermediate filaments cytokeratin, desmin, andvimentin; and the von Willebrand factor (Factor VIII),on sections of endometrium and/or placenta from giltsduring early pregnancy up to day 40 postmating.
MATERIALS AND METHODSAnimals and Tissue Sampling
The management of the animals throughout theexperimental period was approved by the Ethical Com-mittee for Experimentation with Animals, Uppsala,Sweden. Twelve pregnant Landrace 3 Yorkshire gilts(Sus scrofa), 6-8 months of age and weighing 80-100 kg,were kept indoors in the Department of Obstetrics andGynaecology, Swedish University of Agricultural Sci-ences, and fed a commercial pig feed, according toSwedish standards. Oestrus detection was performedtwice daily in the presence of a boar. The gilts weremated with boars of proved fertility or were insemi-nated (with the boars’ undiluted semen) twice during anormal oestrus and then were slaughtered at days 7(precontact stage, n5 2), 12 (early contact stage, n5 2),15-17 (adhesion stage, n 5 3), or 24-40 (early placentalstage, n 5 5) of pregnancy. In all cases, the animalswere stunned and bled, and the internal genital organswere promptly removed and explored for confirmationof ovarian status and normality. The uterine hornswere longitudinally opened and samples of ad-mesome-trial endometrium or embryonic membranes/placentacollected. In early stages (up to 12 days postcoitum), thesamples were taken from the portion of the uterine
horn where the largest number of embryos had beenretrieved.
Tissue ProcessingFor routine morphological examinations, tissue
samples were immersion fixed in a 3% solution ofglutaraldehyde in sodium cacodylate buffer (pH 7.2,500 mOsm) and stored at 4°C until further processing.Corresponding samples were collected for immunocyto-chemistry and frozen by direct plunging in liquidnitrogen and stored at -70°C alternatively fixed for30-60 minutes in a mixture of paraformaldehyde (4% or8%) and glutaraldehyde (0.1% or 0.5%) in phosphatebuffer (PFG-4 or PFG-8, respectively). The fixed sampleswere subjected to cryoprotection through saturation in2.3 M sucrose at least overnight, before being trimmed,placed on metal chucks, and stored at -70°C.
MorphologyThe glutaraldehyde-fixed tissues were trimmed,
washed in buffer, and either dehydrated and embeddedin water-soluble methylmetacrylate (Historesin; LKB,Bromma, Sweden) or postfixed in osmium tetroxide,dehydrated by exposure to graded concentrations ofethanol and propylene oxide, and embedded in Agar100 plastic resin (AgarAids, Essex, England). Semithinsections (1-2 µm) for light microscopy were obtainedwith a Historange (LKB) microtome (methylmetacry-late) or an LKBUltratome and stainedwith haematoxy-lin-eosin (HE) or buffered toluidine blue. The semithinsections were photographed with a Nikon microphot-FXA photomicroscope. Ultrathin sections for transmis-sion electron microscopy (TEM) were cut with a dia-mond knife from selected areas of the resin-embeddedmaterial. The ultrathin sections were picked up ontouncoated copper grids, counterstained with uranyl ac-etate and lead citrate, and examined in a Philips EM201 electron microscope at 60 or 80 kV. Samples fromall stages of pregnancy were examined by light micros-copy, and selected sections of glands and the interhemalbarrier of the placenta were further studied by TEM.
ImmunocytochemistryThe immunoreactivity of IGF-I; PDGF-A; and PDGF
receptors (a and b); the intermediate filaments cytokera-tin, desmin, and vimentin; and the endothelial cellmarker von Willebrand factor (Factor VIII) in thetissues sampled was studied at the light microscopiclevel using a peroxidase-antiperoxidase (PAP) tech-nique as described by Rodriguez-Martinez et al. (1992)as well as with modifications as follows. In brief,directly frozen tissue blocks were mounted on precooledchucks and cut in a cryostat (Starlet 2212R; Bright,Huntingdon, England) into 5-µm-thick sections andmounted on poly-L-lysine- orA-saline-coated glass slides(Sigma Chemical Co., St. Louis, MO). The sections wereleft in the cryostat for 1 hour and then stored at -70°C.Prior to incubation, the sections were fixed in concen-trated acetone for 5 minutes and air dried for 20minutes. The sections were thereafter briefly washed inTris buffer, pH 7.6, and then incubated with 0.3% H2O2for 15 minutes to block the endogenous peroxidasepresent. Pretreatments with 2% bovine serum albumin(BSA; Sigma) in the buffer were carried out to minimize
166 E. PERSSON AND H. RODRIGUEZ-MARTINEZ

nonspecific binding to immunoglobulins before a panelof primary antibodies was applied to the sections.The antibodies used were 1) against IGF-I: a rabbit
polyclonal against human IGF-I (courtesy of Prof. K.Hall, Dept. Endocrinology, Karolinska Hospital, Stock-holm, Sweden) and a mouse monoclonal against humanIGF-I (clone 82-9A, GF 21; Oncogene Science, Union-dale, NY); 2) against PDGF (chain A) and PDGFR-1(made against purified porcine uterus PDGF receptor,recognizing both a- and b-type PDGF receptors; cour-tesy of Prof. C.-H. Heldin, Ludwig Institute, Uppsala,Sweden); and 3) against the intermediate filamentscytokeratin, desmin, and vimentin (mouse monoclonalsantihuman [M 821], antiswine [M 724], and antiswine[M 725]; Dakopatts, Glostrup, Denmark); in addition,4) the von Willebrand factor (polyclonal rabbit antihu-man [A082]; Dakopatts) was included for comparison ofthe labelling with the desmin antibody.The antibodies were diluted in Tris with 1%BSA. The
sections were incubated with the antibodies for 30-60minutes at room temperature, then rinsed in PBS andincubated with an excess of the respective secondaryantibody (goat or pig antirabbit IgG [Z 421 and Z 196]and rabbit antimouse IgG [Z 259]; Dakopatts) andfinally with the PAP complex (rabbit [Z 113] and mouse[B 650]; Dakopatts). The developer DAB (3,3-diamino-benzidine; Sigma, or Saveen Biotech, Malmo, Sweden)was then added to visualize the antibody-antigen com-plex. Counterstaining with haematoxylin was carriedout on sections with clear and distinct labelling. Se-lected sections were photographed with a Nikon micro-phot-FXAphotomicroscope with and without Nomarskiinterference contrast optics.The PFG-4- or PFG-8-fixed and sucrose-saturated
samples were sectioned with glass knives on a Reichert(Reichert-Jung, Vienna, Austria) cryoultramicrotomeat -80°C. Semithin sections were placed on glass slidesand subjected to immunohistochemical labelling by thePAP method as described above, except for the fixationin acetone. Ultrathin cryosections were placed on goldgrids and immunolabelled using a Tokuyasu’s colloidalgold technique combinedwith silver enhancement (AuroProbe EMGAR 5 nm or GAM IgG 5 nm and IntenSEM;Amersham, Buckinghamshire, England). The sectionswere studied and photographed in a Philips EM 420electron microscope at 20 kV.For both methods, controls were run by omission of
the primary antibody and replacement of the primaryantibody with an irrelevant antibody raised in therabbit species (rabbit antirat [Z 494]; Dakopatts) respec-tive preimmune serum (mouse immunoglobulins-nega-tive control [X 931]; Dakopatts). Furthermore, theimmunogold labellings were also controlled by omissionof silver enhancement.All antibodies and stages of pregnancy were sub-
jected to PAP labelling of cryosections from directlyfrozen specimens. PAP labelling of PFG-4-fixed andsucrose-saturated specimens was carried out with theintermediate filament antibodies on endometrium ofday 17. The semithin sucrose sections showed consider-ably better morphology compared to the directly frozencryosections. However, immunoreactivity as seenagainst desmin in directly frozen sections was notmaintained after PFG fixation as performed in thepresent study. Only placentae from days 24 and 30 were
subject to immunolabelling with colloidal gold andsilver enhancement for electron microscopy, using thePDGF-R1 and intermediate filament antibodies.
RESULTSMorphology of the Porcine Endometrium
and Early PlacentaThe morphology of the early pregnant endometrium
and placenta of the females studied here did not deviatefrom the expected histology for the species. During theprecontact stage, the endometrium was conspicuouslyfolded, including primary and secondary folds of theepithelial-lamina propria layers, and blood vessels werecommonly seen in the lamina propria. The endometrialglands (Fig. 1A,B) showed accumulations of clear, elec-tron-lucent secretory vesicles in the supranuclear cyto-plasm. The early contact and adhesion stages (days12-17) were characterized by decreasing amounts ofglandular secretory vesicles and changes in surfacefolding, vascular development, and contact betweenendometrium and trophoblast as known for the species.By days 24-40 of pregnancy (early placental stage), thetrophoblast had developed a close contact with thematernal surface epithelium,with interdigitatingmicro-villi from each cell layer (Fig. 2A), except for the areolarareas (Fig. 2B). The number of subepithelial capillarieshad now increased markedly, forming a continuouslayer underneath the maternal epithelium (Fig. 2A)very close to its basal lamina (Fig. 2B). The allantocho-rial capillaries were readily seen but were only occasion-ally found close to the trophoblast (Fig. 2A). Thesecretory activity of the endometrial glands decreasedfurther, and the secretory vesicles changed character,becoming dark and electron-dense by days 30-40.At theend of the period studied, the porcine epitheliochorialplacenta type was well established, with a rich vascularlayer developed especially under the endometrial laminaepithelialis but also in the allantochorion.
ImmunocytochemistryThe immunocytochemical localization of the various
growth factors, growth factor receptors, intermediatecytoskeletal filaments, and vonWillebrand factor in theporcine endometrium and placenta is shown in Figures3-8. The control immunostainings were consistentlynegative (Fig. 9A-E), although the use of polyclonalrabbit antibodies gave some unspecific backgroundstaining. As can be seen in Figure 9, the endogenousperoxidase activity found in cells such as mast cells andeosinophils of the endometrium could not be quenchedwith the treatment used. Therefore, cells in the stromawith very strong immunolabelling could not be consid-ered as either positive or negative.IGF-I. Positive immunoreactivity to the polyclonal
rabbit and monoclonal mouse antibodies assayedshowed a similar localization, although the rabbitantibody gave a stronger staining intensity. Immuno-staining in the endometrium was present on the sur-face and glandular epithelium, the endothelium, andthe vascular smooth muscle cells of the endometrialblood vessels (Fig. 3A-C). The vascular smooth muscleshowed strong reactivity (Fig. 3B,C). The immunolabel-ling intensity and localization varied at the glandular
167IMMUNOCYTOCHEMISTRY OF THE PORCINE PLACENTA
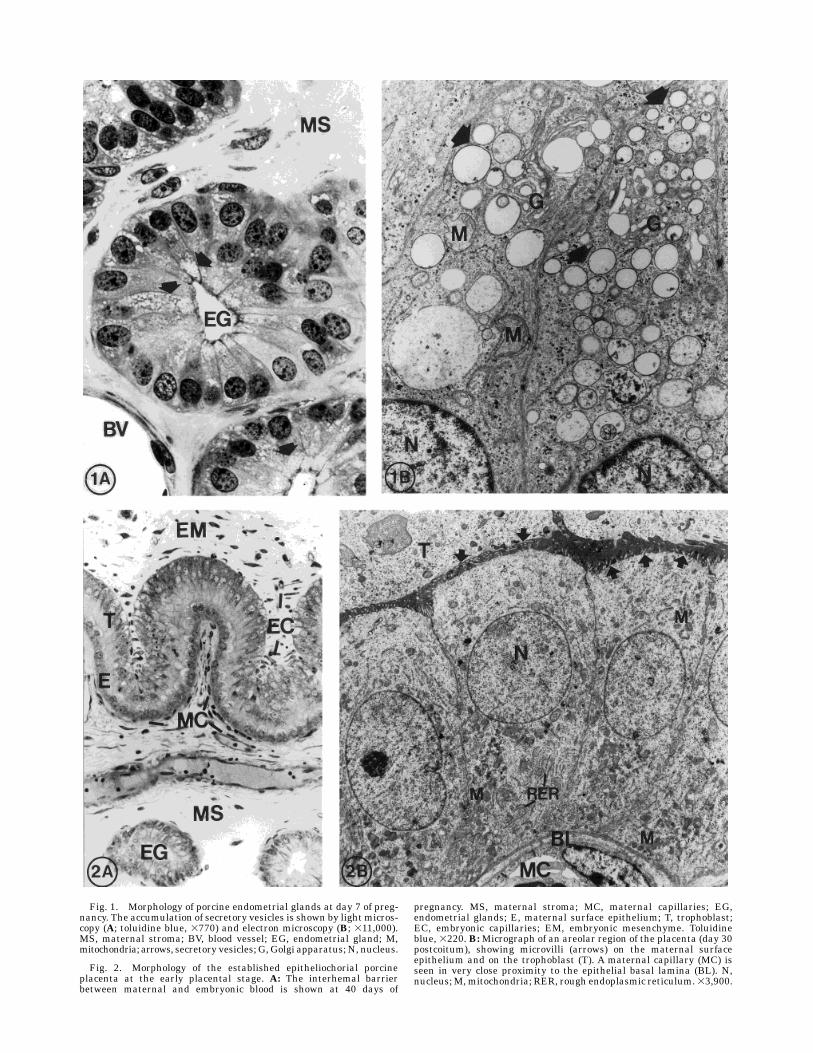
Fig. 1. Morphology of porcine endometrial glands at day 7 of preg-nancy. The accumulation of secretory vesicles is shown by light micros-copy (A; toluidine blue, 3770) and electron microscopy (B; 311,000).MS, maternal stroma; BV, blood vessel; EG, endometrial gland; M,mitochondria; arrows, secretory vesicles; G, Golgi apparatus; N, nucleus.
Fig. 2. Morphology of the established epitheliochorial porcineplacenta at the early placental stage. A: The interhemal barrierbetween maternal and embryonic blood is shown at 40 days of
pregnancy. MS, maternal stroma; MC, maternal capillaries; EG,endometrial glands; E, maternal surface epithelium; T, trophoblast;EC, embryonic capillaries; EM, embryonic mesenchyme. Toluidineblue, 3220. B:Micrograph of an areolar region of the placenta (day 30postcoitum), showing microvilli (arrows) on the maternal surfaceepithelium and on the trophoblast (T). A maternal capillary (MC) isseen in very close proximity to the epithelial basal lamina (BL). N,nucleus;M,mitochondria; RER, rough endoplasmic reticulum.33,900.

level, with the epithelium showing moderate and uni-form labelling during the precontact (Fig. 3A) and earlycontact stages to gradually change, showing stronglabelling in only some of the glandular cells by the timeof apposition (Fig. 3B) and early placental stages (Fig.3C). In the embryonic component of the placenta, thetrophoblast (Fig. 3C) and themesenchymal cells showedmoderate labelling.PDGF (chain A). Moderate immunolabelling was
present in the smoothmuscle of the blood vessels and inthe endometrial glands (Fig. 4A). The trophoblast andthe surface epithelium of the endometrium showedweaker labelling (Fig. 4A). No temporal variations werenoticed in the immunostaining.PDGF receptors. Strong immunoreactivity to the
PDGF-R1 antibody (against receptors of a and b type)was present in all endometrial samples explored, beingprincipally located in the subepithelial stroma, endothe-lium and vascular smooth muscle as well as in bloodvessels on the embryonic side of the placenta (Fig. 4B).Weaker immunoreactivity was seen inmaternal epithe-lia and trophoblast. At the EM level, distinct labellingwas seen along the membranes of the vascular smoothmuscle cells (Fig. 4C), whereas it was less apparent inother cell types.Intermediate filaments. Cytokeratin immunolabel-
ling was seen in the trophoblast and in the surface andglandular epithelia of the endometrium, as shown byPAP labelling of directly frozen cryosections (Fig. 5A)and semithin, sucrose-saturated cryosections (Fig. 8A)for LM and ultrathin cryosections for EM (Fig. 6A). Thesame localization was found throughout the differentstages of placentation. Vimentin immunostaining wasclearly present in the connective tissue of the endome-trial lamina propria and in the embryonic mesenchyme(Fig. 5B). Also, the vascular smooth muscle and theendothelia of both embryonic and maternal sides werestained. In the semithin, sucrose-saturated cryosec-tions as well as at the ultrastructural level, fibroblasts(Figs. 6B, 8B) and endothelial cells (Figs. 6C, 8B) wereseen with distinct cytoplasmic labelling. Immunostain-ing against desmin was present solely in the directlyfrozen tissues (Figs. 5C, 7A), labelling vascular smoothmuscle cells as well as the subepithelial blood vessels,probably pericytes (or possibly endothelial cells) of thecapillaries. This observation could not be confirmed;labelling of the tissue samples fixed and prepared forultrastructural studies was negative.Von Willebrand factor (Factor VIII). Labelling of
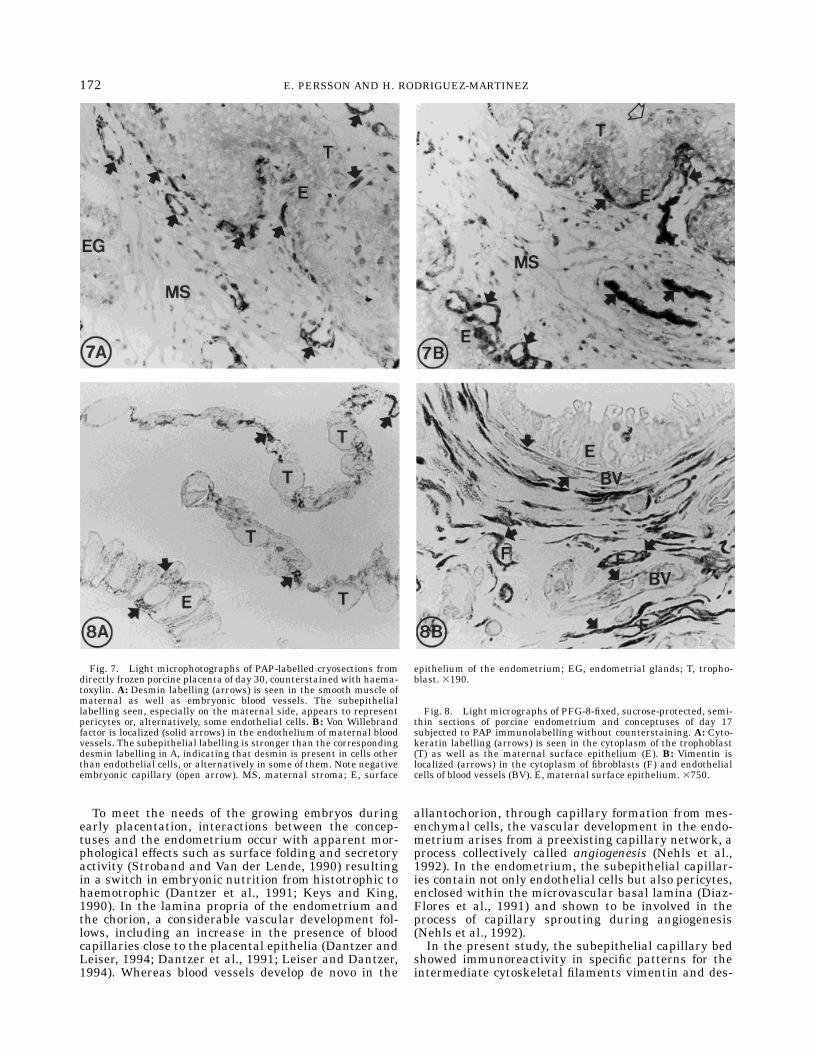
von Willebrand factor was included in order to revealwhether it labelled the same cells underneath theendometrial surface epithelium as the desmin antibodydid. A very strong immunolabelling was found in theendothelium of all vessels larger than capillaries. Thesubepithelial capillaries, however, did not stain asstrongly, but the growth of the capillary bed along thesurface epithelium could clearly be followed with thehelp of the immunoreactivity (Fig. 7B) throughout thestages studied. The embryonic endothelium showedsome reactivity in blood vessels larger than capillaries.The subepithelial labelling of von Willebrand factor onthe maternal side of the placenta was stronger andmore uniform compared to the desmin labelling of theday-30 porcine placenta (Fig. 7A,B).
DISCUSSIONThe present study, immunocytochemically estab-
lished by light and electron microscopy, shows that theIGF-I and PDGF ligands; the PDGF-chain receptors;the intermediate filaments cytokeratin, desmin, andvimentin; and the von Willebrand factor are present inspecific patterns during the tissue reorganization anddevelopment involved in the establishment of the epithe-liochorial placenta of the pig. Early pregnant gilts wereused up to day 40 of gestation.The porcine conceptuses migrate along the uterine
horns after their entrance into the uterine lumen,undergoing a dramatic elongation, finally to establishcontact with the endometrium to develop a noninva-sive, epitheliochorial placenta. The endometrium corre-spondingly increases its secretory activity during thefirst 2 weeks following fertilization, i.e., during theperiod when the embryos are free and are nurturedthrough absorption of histotrophe (secretion by theendometrial glands mainly; Geisert at el., 1982; Stro-band et al., 1984; Stroband and Van der Lende, 1990).The changes in endometrial secretory activity seem
to be modulated not only by the maternal ovariansteroid hormones but also by the oestrogens producedin varying amounts by the elongating blastocysts,rendering the signal for maternal recognition of preg-nancy by acting upon the uterus (Bazer et al., 1991).The effects of oestradiol appear to be mediated with thehelp of autocrine or paracrine polypeptide growth fac-tors and/or their receptors (Eriksson, 1993). Oestradiolup-regulates expression of IGF-I in the porcine uterus(Sahlin et al., 1990; Simmen et al., 1990), and IGF-I hasbeen shown to mediate growth in steroid-responsiveorgans (Sahlin et al., 1994); hormonal influence hasalso been seen through a relationship between tissuesteroid receptor levels and expression and IGF-I mRNAlevels in the porcine uterus (Persson et al., in press).Both pig blastocysts and pig uterine tissues possess
specific receptors for IGF-I (Corps et al., 1990; Hofig etal., 1991), thus relating IGF-I to embryonic develop-ment and uterine function. Porcine embryos have alsobeen shown to contain IGF-I at maximal levels in thebeginning of the placentation process on day 12, withmRNA levels constant and significantly lower than inthe corresponding endometrium (Letcher et al., 1989),implying an absorption from the uterine secretions.Peak levels of endometrial IGF-I mRNAwere also seenat day 12 in both cyclic and pregnant gilts but wereclearly higher during pregnancy (Simmen et al., 1992),coinciding with peak levels of IGF-I in the uterineluminal fluid (Letcher et al., 1989) and a rise insecretory activity of the endometrial glands as well aswith the onset of embryonic elongation (Geisert et al.,1982) and oestrogen secretion from the porcine blasto-cysts (Perry et al., 1973). Preliminary results (Perssonand Rodriguez-Martinez, 1993; Persson et al., 1992) aswell as the present study localized IGF-I immunocyto-chemically in the trophoblast of porcine embryos and inthe epithelia of the early pregnant endometrium. Thetemporal relationship between morphological evidenceof secretory accumulation and release and a change inIGF-I immunolabelling in the endometrial glands indi-cates a common regulator, possibly conceptus-derivedoestrogens.
169IMMUNOCYTOCHEMISTRY OF THE PORCINE PLACENTA
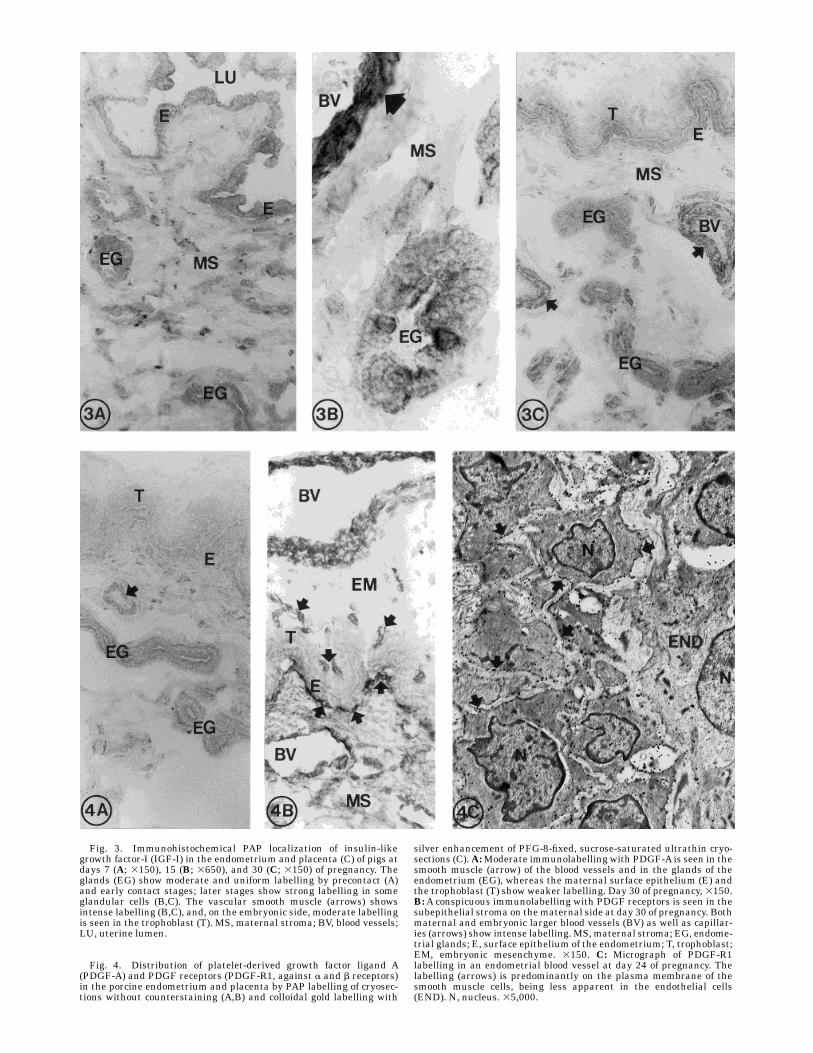
Fig. 3. Immunohistochemical PAP localization of insulin-likegrowth factor-I (IGF-I) in the endometrium and placenta (C) of pigs atdays 7 (A; 3150), 15 (B; 3650), and 30 (C; 3150) of pregnancy. Theglands (EG) show moderate and uniform labelling by precontact (A)and early contact stages; later stages show strong labelling in someglandular cells (B,C). The vascular smooth muscle (arrows) showsintense labelling (B,C), and, on the embryonic side, moderate labellingis seen in the trophoblast (T). MS, maternal stroma; BV, blood vessels;LU, uterine lumen.
Fig. 4. Distribution of platelet-derived growth factor ligand A(PDGF-A) and PDGF receptors (PDGF-R1, against a and b receptors)in the porcine endometrium and placenta by PAP labelling of cryosec-tions without counterstaining (A,B) and colloidal gold labelling with
silver enhancement of PFG-8-fixed, sucrose-saturated ultrathin cryo-sections (C).A:Moderate immunolabelling with PDGF-A is seen in thesmooth muscle (arrow) of the blood vessels and in the glands of theendometrium (EG), whereas the maternal surface epithelium (E) andthe trophoblast (T) show weaker labelling. Day 30 of pregnancy, 3150.B:A conspicuous immunolabelling with PDGF receptors is seen in thesubepithelial stroma on thematernal side at day 30 of pregnancy. Bothmaternal and embryonic larger blood vessels (BV) as well as capillar-ies (arrows) show intense labelling.MS,maternal stroma; EG, endome-trial glands; E, surface epithelium of the endometrium; T, trophoblast;EM, embryonic mesenchyme. 3150. C: Micrograph of PDGF-R1labelling in an endometrial blood vessel at day 24 of pregnancy. Thelabelling (arrows) is predominantly on the plasma membrane of thesmooth muscle cells, being less apparent in the endothelial cells(END). N, nucleus. 35,000.
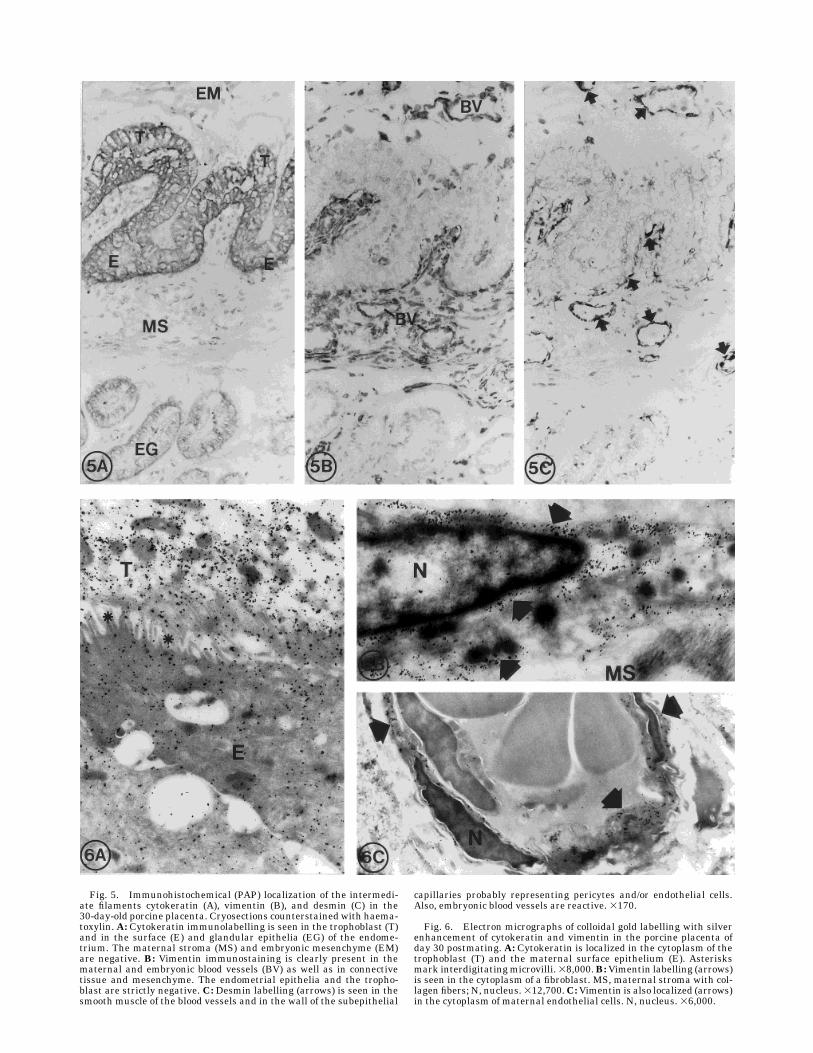
Fig. 5. Immunohistochemical (PAP) localization of the intermedi-ate filaments cytokeratin (A), vimentin (B), and desmin (C) in the30-day-old porcine placenta. Cryosections counterstained with haema-toxylin. A: Cytokeratin immunolabelling is seen in the trophoblast (T)and in the surface (E) and glandular epithelia (EG) of the endome-trium. The maternal stroma (MS) and embryonic mesenchyme (EM)are negative. B: Vimentin immunostaining is clearly present in thematernal and embryonic blood vessels (BV) as well as in connectivetissue and mesenchyme. The endometrial epithelia and the tropho-blast are strictly negative. C: Desmin labelling (arrows) is seen in thesmooth muscle of the blood vessels and in the wall of the subepithelial
capillaries probably representing pericytes and/or endothelial cells.Also, embryonic blood vessels are reactive. 3170.
Fig. 6. Electron micrographs of colloidal gold labelling with silverenhancement of cytokeratin and vimentin in the porcine placenta ofday 30 postmating. A: Cytokeratin is localized in the cytoplasm of thetrophoblast (T) and the maternal surface epithelium (E). Asterisksmark interdigitatingmicrovilli.38,000.B:Vimentin labelling (arrows)is seen in the cytoplasm of a fibroblast. MS, maternal stroma with col-lagen fibers; N, nucleus.312,700.C:Vimentin is also localized (arrows)in the cytoplasm of maternal endothelial cells. N, nucleus. 36,000.
To meet the needs of the growing embryos duringearly placentation, interactions between the concep-tuses and the endometrium occur with apparent mor-phological effects such as surface folding and secretoryactivity (Stroband and Van der Lende, 1990) resultingin a switch in embryonic nutrition from histotrophic tohaemotrophic (Dantzer et al., 1991; Keys and King,1990). In the lamina propria of the endometrium andthe chorion, a considerable vascular development fol-lows, including an increase in the presence of bloodcapillaries close to the placental epithelia (Dantzer andLeiser, 1994; Dantzer et al., 1991; Leiser and Dantzer,1994). Whereas blood vessels develop de novo in the
allantochorion, through capillary formation from mes-enchymal cells, the vascular development in the endo-metrium arises from a preexisting capillary network, aprocess collectively called angiogenesis (Nehls et al.,1992). In the endometrium, the subepithelial capillar-ies contain not only endothelial cells but also pericytes,enclosed within the microvascular basal lamina (Diaz-Flores et al., 1991) and shown to be involved in theprocess of capillary sprouting during angiogenesis(Nehls et al., 1992).In the present study, the subepithelial capillary bed
showed immunoreactivity in specific patterns for theintermediate cytoskeletal filaments vimentin and des-
Fig. 7. Light microphotographs of PAP-labelled cryosections fromdirectly frozen porcine placenta of day 30, counterstained with haema-toxylin. A: Desmin labelling (arrows) is seen in the smooth muscle ofmaternal as well as embryonic blood vessels. The subepitheliallabelling seen, especially on the maternal side, appears to representpericytes or, alternatively, some endothelial cells. B: Von Willebrandfactor is localized (solid arrows) in the endothelium of maternal bloodvessels. The subepithelial labelling is stronger than the correspondingdesmin labelling in A, indicating that desmin is present in cells otherthan endothelial cells, or alternatively in some of them. Note negativeembryonic capillary (open arrow). MS, maternal stroma; E, surface
epithelium of the endometrium; EG, endometrial glands; T, tropho-blast. 3190.
Fig. 8. Light micrographs of PFG-8-fixed, sucrose-protected, semi-thin sections of porcine endometrium and conceptuses of day 17subjected to PAP immunolabelling without counterstaining. A: Cyto-keratin labelling (arrows) is seen in the cytoplasm of the trophoblast(T) as well as the maternal surface epithelium (E). B: Vimentin islocalized (arrows) in the cytoplasm of fibroblasts (F) and endothelialcells of blood vessels (BV). E, maternal surface epithelium. 3750.
172 E. PERSSON AND H. RODRIGUEZ-MARTINEZ
min as well as for von Willebrand factor (Factor VIII).Vimentin expression is characteristic for cells of mesen-chymal origin (Franke et al., 1979), including endothe-lia as well as pericytes. Von Willebrand factor is foundin endothelial cells (Sehestad and Hou-Jensen, 1981)whereas desmin, usually present inmuscle cells but notin endothelium, also appeared in pericytes (Nehls et al.,1992), indicating that the subepithelial desmin label-ling as described above could be confined to pericytes.The difference in immunoreactivity between desminand von Willebrand factor suggests that pericytes areinvolved in the angiogenesis occurring during porcineplacentation. Desmin labelling was readily seen insmall as well as large embryonic vessels, although vonWillebrand factor was clearly seen only in larger ones.The difference between maternal and embryonic capil-
laries indicates different regulatorymechanisms regard-ing the embryonic vasculogenesis (Torry and Rongish,1992), which is considered to be a developmental pro-cess independent of angiogenic factors (compared tomaternal vascular growth by Reynolds et al., 1992). Inrecent studies, Reynolds et al. have pointed out that theendometrial, but not the embryonic, part of the ovineplacenta produce endothelial mitogens during earlypregnancy (days 12-40).Many polypeptides possess angiogenic activities in
vitro and in vivo (Folkman and Shing, 1992; Risau,1990). One of these, the vascular endothelial growthfactor (VEGF/VPF), is structurally related to PDGF(Keck et al., 1989), and its function is specifically on theendothelial cells, increasing vascular permeability. En-hanced vascular permeability occurs during the period
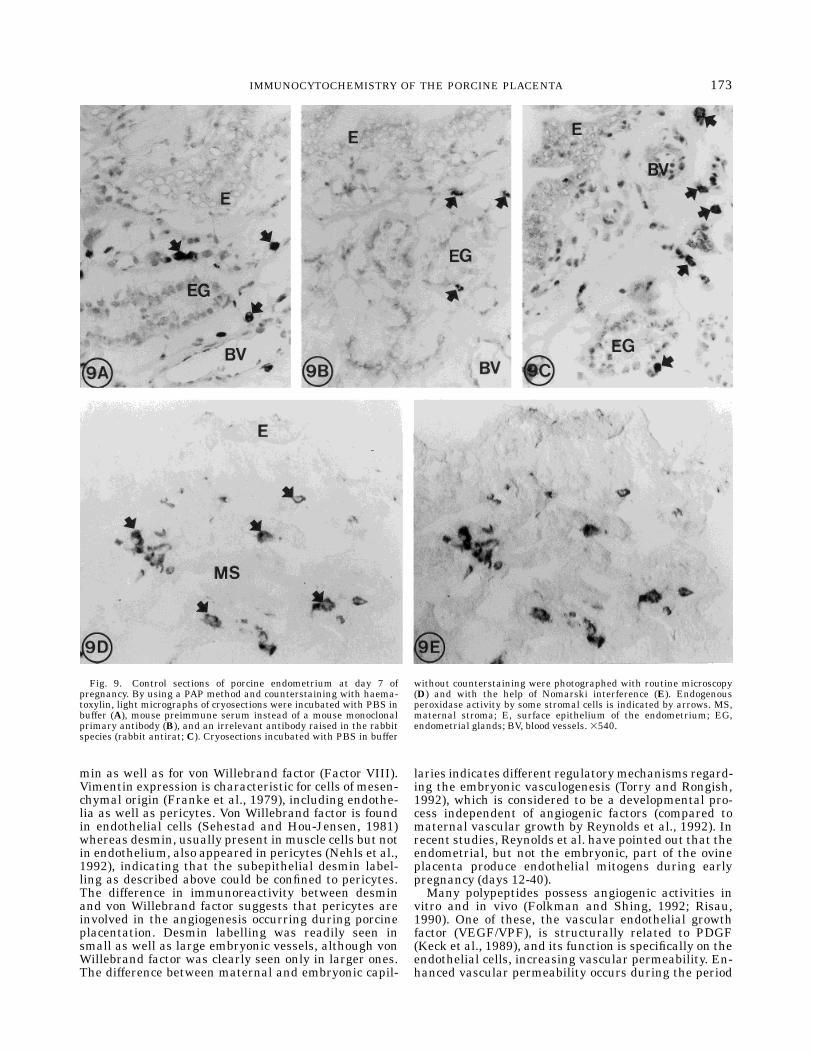
Fig. 9. Control sections of porcine endometrium at day 7 ofpregnancy. By using a PAP method and counterstaining with haema-toxylin, light micrographs of cryosections were incubated with PBS inbuffer (A), mouse preimmune serum instead of a mouse monoclonalprimary antibody (B), and an irrelevant antibody raised in the rabbitspecies (rabbit antirat; C). Cryosections incubated with PBS in buffer
without counterstaining were photographed with routine microscopy(D) and with the help of Nomarski interference (E). Endogenousperoxidase activity by some stromal cells is indicated by arrows. MS,maternal stroma; E, surface epithelium of the endometrium; EG,endometrial glands; BV, blood vessels. 3540.
173IMMUNOCYTOCHEMISTRY OF THE PORCINE PLACENTA

studied here in the porcine uterus during early preg-nancy (Keys et al., 1986).PDGF is a platelet-contained serum mitogen that
exerts major pleiotropic effects (growth, motility) onmesenchyme-derived cells (Ross et al., 1986). PDGF is acationic protein (30 kD) consisting of two polypeptidechains, denominated A and B (Heldin et al., 1985).Recently, PDGF (especially the BB homodimer) hasbeen demonstrated to have angiogenic actions on embry-onic endothelial cells in vivo (Risau et al., 1992). Thetwo polypeptide chains, when combined as homo- orheterodimers, have different functional activities, result-ing from different binding specificities to two distinctPDGF receptor classes, denoted a and b, present on thecell surface of responsive cells (Heldin et al., 1988). ThePDGF receptors (of a and b type) are present in theendometrium and myometrium of cycling and pregnantpigs, and their expression seems to be accentuatedunder oestrogenic influence (Rodriguez-Martinez et al.,1992). In the present study, both the PDGF-A ligandand the PDGF receptors were immunolocalized invascular smooth muscle cells, indicating an autocrinecommunication, which has also been suggested forvascular smooth muscle in in vitro studies of the rat(Sjolund et al., 1988). Whereas the endothelial/perivas-cular areas of the subepithelial layer expressed thereceptors strongly, especially during the latest stageexamined (24-40 days of pregnancy), the PDGF-A li-gand was expressed in epithelial cell layers, indicatingparacrine targets for the factor as well as a role for thisgrowth factor in the angiogenesis of the porcine pla-centa. The weak to moderate labelling of the PDGF-Aligand as seen in the present study may be explained bythe lower angiogenic effect resulting from the ho-modimer PDGF-AA compared to PDGF-BB (Risau etal., 1992). It is tempting to speculate that the coincidinglocalization and increase in PDGF-receptor labellingwith vascular development are modulated by steroids,as has been suggested regarding angiogenesis by Ash-worth et al. (1990) and Reynolds et al. (1992).The results of the present work indicate that both
IGF-I and PDGF are involved in the early nutrition ofpig embryos and in porcine placentation but thatdifferent roles may be ascribed to the factors. WhereasIGF-I appears to be related more to the secretoryactivities of the uterine glands and the histotrophicnurturing of the elongating embryos, PDGF might playan angiogenic role in the establishment of the develop-ing placenta.
ACKNOWLEDGMENTSThe authors thank H. Ekwall, M. Ekwall, and Å.
Jansson, for excellent technical assistance. We grate-fully acknowledge the kind supply of material by Dr. V.Dantzer, Copenhagen.
REFERENCESAshworth, C.J., Fliss, M.F.V., and Bazer, F.W. (1990) Evidence forsteroid control of a putative angiogenic factor in the porcine uterus.J. Endocrinol., 125:15-19.
Bazer, F.W., Simmen, R.C.M., and Simmen, F.A. (1991) Comparativeaspects of conceptus signals for maternal recognition of pregnancy.Ann. N.Y. Acad. Sci., 622:202-211.
Brigstock, D.R., Heap, R.B., and Brown, K.D. (1989) Polypeptidegrowth factors in uterine tissues and secretions. J. Reprod. Fertil.,85:747-758.
Corps, A.N., Brigstock, D.R., Littlewood, C.J., and Brown, K.D. (1990)Receptors for epidermal growth factor and insulin-like growth
factor-I on preimplantation trophectoderm of the pig. Development,110:221-227.
Dantzer, V. (1985) Electronmicroscopy of the initial stages of placenta-tion in the pig. Anat. Embryol., 172:281-293.
Dantzer, V. (1993) Initial placentation in farm animals. Morphologicalevents where growth factors must be implicated. Reprod. DomesticAnim., 28:199-201.
Dantzer, V., and Leiser, R. (1994) Initial vascularisation in the pigplacenta: I. Demonstration of nonglandular areas by histology andcorrosion casts. Anat. Rec., 238:177-190.
Dantzer, V., Svendsen, A.-M., and Leiser, R. (1991) Correlation be-tween morphological events during the initial stages of placentationin the pig. In: Placenta: Basic Research for Clinical Application. Int.Conf. Placenta, Tokyo, 1990. H. Soma, ed. Karger, Basel, pp.188-199.
Diaz-Flores, L., Gutierrez, R., Varela, H., Rancel, N., and Valladares,F. (1991) Microvascular pericytes: A review of their morphologicaland functional characteristics. Histol. Histopathol., 6:269-286.
Eriksson, H. (1993) Regulation of growth factor expression via estro-gens. Reprod. Domestic Anim., 28:195-198.
Folkman, J., and Shing, Y. (1992) Angiogenesis. J. Biol. Chem.,267:10931-10934.
Franke, W.W., Schmid, E., Osborn, M., and Weber, K. (1979) Interme-diate-sized filaments of human endothelial cells. J. Cell Biol.,81:570-580.
Geisert, R.D., Renegar, R.H., Thatcher, W.W., Roberts, R.M., andBazer, F.W. (1982) Establishment of pregnancy in the pig: I.Interrelationships between preimplantation development of the pigblastocyst and uterine endometrial secretions. Biol. Reprod., 27:925-939.
Geisert, R.D., Zavy, M.T., Moffatt, R.J., Blair, R.M., and Yeltin, T.(1990) Embryonic steroids and the activation of pregnancy in pigs. J.Reprod. Fertil., 40:293-305.
Heldin, C.-H., Wasteson, Å., and Westermark, B. (1985) Platelet-derived growth factor. Mol. Cell. Endocrinol., 39:169-187.
Heldin, C.-H., Backstrom, G., Ostman,A. Hammacher,A., Ronnstrand,L., Rubin, K., Nister, M., and Westermark, B. (1988) Binding ofdifferent dimeric forms of PDGF to human fibroblasts: Evidence fortwo separate receptor types. EMBO J., 7:1387-1393.
Hofig, A., Michel, F.J., Simmen, F.A., and Simmen, R.C.M. (1991)Constitutive expression of uterine receptors for insulin-like growthfactor-I during the peri-implantation period in the pig. Biol. Reprod.,45:533-539.
Keck, P.J., Hauser, S.D., Krivi, G., Sanzo, K., Warren, T., Feder, J., andConnolly, D.T. (1989) Vascular-permeability factor, an endothelial-cell mitogen related to PDGF. Science, 246:1309-1312.
Keys, J.L., and King, G.J. (1990) Microscopic examination of porcineconceptus-maternal interface between days 10 and 19 of pregnancy.Am. J. Anat., 188:221-238.
Keys, J.L., King, G.J., and Kennedy, T.G. (1986) Increased uterinevascular permeability at the time of embryonic attachment in thepig. Biol. Reprod., 34:405-411.
Leiser, R., and Dantzer, V. (1994) Initial vascularisation in the pigplacenta: II. Demonstration of gland and areola-gland subunits byhistology and corrosion casts. Anat. Rec., 238:326-334.
Letcher, R., Simmen, R.C.M., Bazer, F.W., and Simmen, F.A. (1989)Insulin-like growth factor-I expression during early conceptus devel-opment in the pig. Biol. Reprod., 41:1143-1151.
Nehls, V., Denzer, K., and Drenckhahn, D. (1992) Pericyte involvementin capillary sprouting during angiogenesis in situ. Cell Tissue Res.,270:469-474.
Perry, J.S., Heap, R.B., and Amoroso, E.C. (1973) Steroid hormoneproduction by pig blastocysts. Nature, 245:45-47.
Persson, E., and Rodriguez-Martinez, H. (1993) Immunocytochemicalstudies of growth factors in porcine conceptus and endometriumduring early pregnancy. Proc. 4th Int. Conf. Pig Reprod., Columbia,Missouri, p. 70.
Persson, E., Rodriguez-Martinez, H., and Dantzer, V. (1992) Immuno-cytochemical localization of growth factors during porcine placenta-tion. Proc. 12th Int. Congr. Animal Reprod., The Hague, Vol. 3, pp.1267-1269.
Persson, E., Dantzer, V., and Rodriguez-Martinez, H. (1994) Cytoskel-etal proteins as cell-type markers in the porcine epithelio-chorialplacenta. Placenta, 15:A.55.
Persson, E., Sahlin, L., Masironi, B., Dantzer, V., Eriksson, H., andRodriguez-Martinez, H. (1997) Insulin-like growth factor-I in theporcine endometrium and placenta. Localization and concentrationin relation to steroid influence during early pregnancy. AnimalReproduction Science, in press.
Reynolds, L.P., Killilea, S.D., and Redmer, D.A. (1992) Angiogenesis inthe female reproductive system. FASEB J., 6:886-892.
174 E. PERSSON AND H. RODRIGUEZ-MARTINEZ

Risau, W. (1990) Angiogenic growth factors. Progr. Growth FactorRes., 2:71-79.
Risau, W., Drexler, H., Mironov, V., Siegbahn,A., Funa, K., and Heldin,C.-H. (1992) Platelet-derived growth factor is angiogenic in vivo.Growth Factors, 7:261-266.
Rodriguez-Martinez, H., Persson, E., Hurst, M., and Stanchev, P.(1992) Immunohistochemical localization of platelet-derived growthfactor receptors in the porcine uterus during the oestrous cycle andpregnancy. J. Vet. Med., A39:1-10.
Ross, R., Raines, E.W., and Bowen-Pope, D.F. (1986) Biology ofplatelet-derived growth factor. Cell, 46:155-169.
Ronnstrand, L., Beckmann, P.M., Faulders, B., Ostman,A., Er, B., andHeldin, C.-H. (1987) Purification of the receptor for platelet-derivedgrowth factor from porcine uterus. J. Biol. Chem., 262:2929-2932.
Sahlin, L., Rodriguez-Martinez, H., Stanchev, P., Dalin, A.-M., Nor-stedt, G., and Eriksson, H. (1990) Regulation of the uterine expres-sion of messenger ribonucleic acids encoding the oestrogen receptorand IGF-I peptides in the pig uterus. J. Vet. Med., A37:795-800.
Sahlin, L., Norstedt, G., and Eriksson, H. (1994) Estrogen regulationof the estrogen receptor and insulin-like growth factor-I in the ratuterus: a potential coupling between effects of estrogen and IGF-1.Steroids, 59:421–430.
Sehested, M., and Hou-Jensen, K. (1981) Factor VIII related antigenas an endothelial cell marker in benign and malignant diseases.VirchowsArch. Pathol. Anat., 391:217-225.
Simmen, F.A., and Simmen, R.C.M. (1991) Peptide growth factors andproto-oncogenes in mammalian conceptus development. Biol. Re-prod., 44:1-5.
Simmen, R.C.M., Simmen, F.A., Hofig, A., Farmer, S.J., and Bazer,
F.W. (1990) Hormonal regulation of insulin-like growth factor geneexpression in pig uterus. Endocrinology, 127:2166-2174.
Simmen, F.A., Simmen, R.C.M., Geisert, R.D., Martinat-Botte, F.,Bazer, F.W., and Terqui, M. (1992) Differential expression, duringthe estrous cycle and pre and postimplantation conceptus develop-ment, of messenger ribonucleic acids encoding components of the piguterine insulin-like growth factor system. Endocrinology, 130:1547-1556.
Sjolund, M., Hedin, U., Sejersen, T., Heldin, C.-H., and Thyberg, J.(1988) Arterial smooth muscle cells express platelet-derived growthfactor (PDGF) A chain mRNA, secrete a PDGF-like mitogen, andbind exogenous PDGF in a phenotype- and growth state-dependentmanner. J. Cell Biol., 106:403-413.
Stroband, H.W.J., and Van der Lende, T. (1990) Embryonic and uterinedevelopment during early pregnancy in pigs. J. Reprod. Fertil.,Suppl. 40:261-277.
Stroband, H.W.J., Taverne, N., and Bogaard, M. (1984) The pigblastocyst: Its ultrastructure and the uptake of protein macromol-ecules. Cell Tissue Res., 235:347-356.
Terracio, L., Ronnstrand, L., Tingstrom, A., Rubin, K., Claesson-Welsh, L., Funa, K., and C.-H. Heldin (1988) Induction of platelet-derived growth factor receptor expression in smooth muscle cellsand fibroblasts upon tissue culturing. J. Cell Biol., 107:1947-1957.
Torry, R.J., and Rongish, B.J. (1992) Angiogenesis in the uterus:Potential regulation and relation to tumor angiogenesis. Am. J.Reprod. Immunol., 27:171-179.
Zhang, Z., Krause, M., and Davis, D.L. (1992) Epidermal growth factorreceptors in porcine endometrium: Binding characteristics and theregulation of prostaglandin E and F2. Biol. Reprod., 46:932-936.
175IMMUNOCYTOCHEMISTRY OF THE PORCINE PLACENTA





























