Embed Size (px)
Citation preview

OPEN 3 ACCESS Freely available online tlos oneReview
Global Diversity of Sponges (Porifera)Rob W. M. Van Soest1*, Nicole Boury-Esnault2, Jean Vacelet2, Martin Dohrmann3, Dirk Erpenbeck4, Nicole J. De V oogd1, Nadiezhda Santodom ingo5, Bart Vanhoorne6, Michelle Kelly7, John N. A. Hooper81 N etherlands C entre for Biodiversity Naturalis, Leiden, The N etherlands, 2 Aix-Marseille University, C en tre d 'O céano log ie d e Marseille, CNRS, DIMAR, UMR 6540, Marseille, France, 3 D epartm en t o f Inverteb rate Zoology, N ational M useum o f Natural History, Sm ithsonian Institution, W ashington , D.C., U nited S tates o f America, 4 D epartm en t o f Earth- and Environm ental Sciences & G eoBio-Center LMU, Ludwig-M aximilians-University M unich, Munich, G erm any, 5 Paleonto logy D epartm en t, Natural History M useum , London, United Kingdom, 6 Flanders M arine Institu te - VLIZ, Innovocean Site, O ostende, Belgium, 7 National C entre for A quatic Biodiversity & Biosecurity, National Institu te o f W ater and A tm ospheric Research Ltd, Auckland, New Zealand, 8 Q ueensland M useum , S outh Brisbane, Q ueensland , and Eskitis Institu te for Cell & M olecular Therapies, Griffiths University, Q ueensland, Australia
Abstract: With th e com pletion of a single unified classification, the Systema Porifera (SP) and subsequent developm ent of an online species database, the World Porifera Database (WPD), we are now equipped to provide a first com prehensive picture of the global biodiversity of the Porifera. An introductory overview of the four classes of the Porifera is followed by a description of the structure of our main source of data for this paper, the WPD. From this we extracted num bers of all 'know n' sponges to date: the num ber of valid Recent sponges is established at 8,553, with the vast majority, 83%, belonging to the class Demospongiae. We also m apped for th e first tim e the species richness of a com prehensive se t of marine ecoregions of the world, data also extracted from the WPD. Perhaps not surprisingly, these distributions appear to show a strong bias tow ards collection and taxonom y efforts. Only w hen species richness is accum ulated into large marine realms does a pattern em erge tha t is also recognized in many o ther marine animal groups: high num bers in tropical regions, lesser num bers in the colder parts of the world oceans. Preliminary similarity analysis of a matrix of species and marine ecoregions extracted from the WPD failed to yield a consistent hierarchical pattern of ecoregions into marine provinces. Global sponge diversity information is mostly generated in regional projects and resources: results obtained dem onstrate tha t regional approaches to analytical biogeography are at present more likely to achieve insights into the biogeographic history of sponges than a global perspective, which appears currently too ambitious. We also review information on invasive sponges tha t might well have som e influence on distribution patterns of the future.
IntroductionSponges, phylum Porifera, are the oldest m etazoan group still
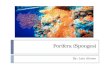
extant on our planet. T heir continued survival in vast num bers in R ecen t seas (and in freshw ater habitats) is closely linked to the apparen t adaptability o f their baup lan to dram atic changes in environm ental characteristics and com peting b io ta [1,2]. Sponges (Fig. 1A) are exclusively aquatic anim als, w hich are fixed on the substrate an d live by draw ing in w ater and filtering m icroscopic- size food particles from it. R ecen t research also indicates an ability to take up dissolved organic m atter [3]. Sponges have a simple level o f organization: there are specialized cells for a variety o f life functions, bu t these are no t organized into tissues o r organs. All sponges have a “ skin” o f T -shaped or flattened cells (called pinacocytes) w hich covers the outside o f the sponge) as well as its internal system o f canals, and m icroscopic cham bers (Fig. IB).
These cham bers have a lining of flagella-bearing cells (choanocytes, Fig. 1C) that generate the w ater currents necessary for the unique filtering activity characteristic to sponges. An exception to this is in the so-called carnivorous sponges, highly adap ted deep- sea forms, in w hich the aquiferous system is non-existent, bu t w hich have a sticky ou ter surface with w hich small prey anim als are cap tured [4], T h e space (Fig. IB) betw een canals and cham bers is filled w ith a collagenous m atrix, called the mesohyl, w hich harbors individual cells, supporting fibers, and inorganic structures o f the skeleton [5].
Sponges grow in distinct shapes (Fig. 1A) an d sizes due to the form of the internal m ineral a n d /o r organic skeletons secreted by specialized cells. T h e skeleton m ay also be supplem ented by exogenous m aterials, such as sand grains. Skeletons, w hen present, are constructed of discrete siliceous o r calcareous elem ents (spicules) a n d /o r organic collagenous fibers (spongia), and rarely skeletons m ay be aspicular massive lim estone constructions. D epending on the nature and density o f these building com ponents, sponge species m ay variously be soft, compressible, fragile o r rock h a rd in consistency. Sponges com e in various shapes and sizes, from flat cushions to elaborate b ranch ing or cup-shaped forms, from tiny crusts m easured in nini, to giant shapes in meters. Sponges have num erous m icroscopic openings (the incurren t pores) an d one or a few larger vents (the excurrent oscilles). T h e shapes o f sponges are variable am ong different species and genera, bu t also vary to some extent betw een individuals o f the same species in response to environm ental factors such as hydrodynam ics, light and turbidity.
A great diversity o f symbiotic organism s often thrive inside or on the body o f a sponge, from m icroscopic prokaryotes, e.g. [6,7] to m acroscopic organism s such as shrimps, polychaetes, hydrozoans and fishes, e.g. [8],
T h e simple body organization of sponges an d relative plasticity o f the cellular elem ents, coupled with a unique tolerance towards symbiotic m icroorganism s, allows for a great diversity o f
C ita tio n : Van S oest RWM, Boury-Esnault N, Vacelet J, D ohrm ann M, Erpenbeck D, e t al. (2012) Global Diversity o f Sponges (Porifera). PLoS ONE 7(4): e35105. dol:10.1371 /journal.pone.0035105
E d ito r: John Murray Roberts, H erlot-W att University, United Kingdom
P u b lish ed April 27, 2012
C o p y rig h t: © 2012 Van Soest e t al. This Is an open-access article d istributed under th e term s o f th e Creative C om m ons A ttribution License, w hich perm its unrestricted use, distribution , and reproduction In any m edium , provided th e original au th o r and source are credited .
F u n d in g : The au thors have no funding or su p p o rt to report.
C o m p e tin g In te rests : The au thors have declared th a t no com peting Interests exist.
* E-mail: rob.vansoest@ ncbnaturalls.nl
PLoS ONE I w ww.plosone.org 1 April 2012 I Volume 7 | Issue 4 | e35105
Global Diversity of Sponges
Figure 1. Porifera m orphology and internal structure. A. Callyspongia (Callyspongia) sam arensis (D em o sp o n g iae : H aplosclerida), T erna te , M aluku prov ince , Indonesia (p h o to N.J. d e V oogd); B. SEM im a g e o f c ross sec tio n o f m esohy l o f th e d e m o s p o n g e S copalina ruetz le ri o b ta in e d by freeze-frac tu ring te c h n iq u e (cou rte sy L. d e Vos); C. Detail o f c h o an o c y te c h a m b e r o f S copalina ruetz le ri (cou rtesy L. d e Vos). do i:10 .1371/jo u rn a l.pone.0035105 .g001
‘evolutionary solutions’ for environm ental challenges. Know ledge o f sponge biodiversity is still largely incom plete. T o date, about 11,000 species have been form ally described o f w hich approxim ately 8,500 are considered valid (see below), bu t as m any as twice that num ber are thought to exist. Sponges are currently divided am ong four distinct classes, 25 orders, 128 families and 680 genera [9,10], bu t m any o f these h igher taxa are under discussion due to new insights obtained from m olecular systematic m ethods and new considerations o f their m orphological characteristics. Fossil sponges com prise a similar additional diversity [11] T h ere are several hundred freshwater species.
D ue to the lim ited swim m ing capabilities o f m ost sponge larvae, and occasional asexual p ropagation , m ost sponges occur in regional o r local areas o f endem ism , unless spread globally or regionally in an inadvertent m anner by shipping traffic. Sponges m ay be found vertically from the eulittoral zone to hadai depths, horizontally from the tropics to the highest latitudes, locally from epifaunal rocky com m unities to m ud bottom s and ephem eral freshwater habitats. T h eir im portance for the global ecosystem is high b u t no t widely appreciated [12,13]. Sponges are efficient filter feeders, vital to the health an d econom ics o f all m arine systems by linking the nutrients o f the open w ater colum n w ith benthic
com m unities. Sym bionts o f sponges play a decisive role in the nitrogen cycle o f m any habitats and m ay contribute significantly to organic p roduction in oligotrophic habitats. Specialized sponges are im portan t bio-eroders in coral reefs, coralline bottom s and oyster beds and they m ay com pete successfully with o ther sessile organism s such as corals. Specific groups have an essential function in b inding unconsolidated substrate such as coral rubble and pebbles into stable surfaces. M any fossil sponges and a small group o f R ecen t sponges a re capable o f building extensive reef form ations that today, in some locations, shape the contours o f the benthos, an d now form uplifted terrestrial habitats. M egabenthic species m ay form high-density aggregations in m any shelf edge and seam ount regions playing a so far unexplored role in deep-sea ecosystems. These are only a few general features o f the ecosystem services provided by the global sponge com m unity [14-26].
A lthough sponges have been known to m ankind since the earliest civilizations (4000 YBP, see [27]) they w ere no t recognized as an independent m etazoan lineage until well into the 19th century, w hen R obert G ran t [28] first observed their unique m orphology and physiology and coined the nam e Porifera for them . Since then, spongology, the study o f all aspects o f the biology, ecology, taxonom y and chem istry o f sponges, has grown
PLoS ONE I w ww.plosone.org 2 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
into a discipline attracting a steadily increasing population of hundreds o f scientists worldwide, m any o f w hom devote a lifetime career to the study o f this group. Increasingly, sponges are studied as p a r t o f a b ro ad er enterprise a ttem pting to detail the T ree of Life. A part from nurtu ring academ ic interest, sponges play an im portan t role in hum an health as producers o f chem ical com pounds with useful pharm aceutical properties, including antitum or, anti-infective and anti-inflam m atory properties [29]. N atural sponges are still harvested for personal, industrial, and artistic use.
For the first tim e since the appearance o f the 2002 consensus classification, we review here the global diversity o f the R ecent Porifera, giving a sum m ary of the m ajor groups and their currently established taxon richness. W e also m ake a first a ttem pt to review distribution patterns o f species an d higher taxa over the global seas and oceans. O u r emphasis will be on the ‘know n’ species, b u t we will also briefly consider the ‘unknow n’ species.
MethodsBecause o f the review na ture o f this study, m ethods em ployed
are diverse. W e sum m arize here the m ajor m ethodological approaches, w hich are further explained in the various sections below. T axonom ic and distribution da ta were extracted from the online W orld Porifera D atabase [10] (accessed 2011 Sept 30), and supplem ented with a survey o f the literature on sponge diversity. Figures, tables and m aps are partially the result o f newly analyzed data. T h e type localities and additional confirm ed occurrences in neighboring areas o f a lm ost all ‘accepted’ species were en tered in the W PD in generalized areas (M arine Ecoregions o f the W orld, M E O W s, see [30]), b u t m any non-original distribution records are still to be evaluated and entered. M oreover, m any sponge taxa are recorded in the literature as ‘undeterm ined’ an d these a re not included in the W PD . Thus, the da ta an d m aps for species presented here are to be considered a conservative or ‘m inim al’ estim ate o f the actual distributional da ta and patterns. For the p roduction o f m aps and the tracing of species richness patterns, W PD da ta sets w ere com bined in geographic inform ation system (GIS) software (ESRI A rcG IS v9.3). A biodiversity analysis aim ed a t testing the aptness o f the M E O W hierarchical system o f M arine Provinces an d M arine Realm s for sponge richness data was carried ou t using the B ray-Curtis coefficient h ierarchical clustering o f W PD datasets perform ed w ith the P R IM E R -6 (PRIM ER-E) package. Presence/absence sponge species da ta were clustered at three levels distinguished in the M E O W [30] system: realm , province (> 50 records) and ecoregion (> 2 0 records) level. T he reduction in the num ber o f provinces an d ecoregions was determ ined empirically by repeated clustering attem pts with different m inim um record num bers in w hich level o f resolution o f the dendrogram was observed. T his reduction is justified by the lack o f sufficient exploration o f these geographic units, b u t precise levels (m inim um o f 50 an d 20 records) were chosen arbitrarily. A uthor contributions outlined below were solicited on the basis o f expert knowledge and skills.
Results
C urren tly r e c o g n iz e d h ig h e r taxa a n d n e w (molecular) d e v e lo p m e n t s
D em o sp o n g ia e . D em ospongiae is the largest and m ost diverse class o f the Porifera. It unites [9] sponges w ith siliceous spicules (Fig. 2G) (either m onaxonic o r tetraxonic, never triaxonic) a n d /o r with a skeleton o f organic fibers or fibrillar collagen. Like in H exactinellida (see below) siliceous spicules are divided into
megascleres, w hich strengthen the fram ew ork o f the sponges, and microscleres, w hich have various - possibly defensive, possibly supportive o f soft tissues, b u t generally unclear - functions. M icroscleres are frequently m ore com m on in the ou ter regions o f the sponges and often surround aquiferous canals. M em bers o f the class H om osclerom orpha also possess tetraxonic siliceous spicules, b u t they lack a subdivision in m ega- and microscleres. O ccasionally the skeleton is absent, a feature shared again with some H om osclerom orpha. R are forms with lim estone basal skeletons are living links to Paleozoic reef-building sponges. Larvae are usually o f the parenchym ella type (solid with overall ciliation), b u t in some groups hollow larvae occur [31,32]. T he m ost recent sum m ary o f the Porifera classification [9] recognized 15 ordinal groups, one o f w hich was recently transferred to the class H om osclerom orpha (see below). T h e m ajor groups include three orders possessing tetraxonic spicules (Spirophorida, A strophorida, and p a rt o f the “ Lithistida”), three orders lacking siliceous spicules th a t were historically called keratose o r horny sponges (Dictyoceratida, D endroceratida, and V erongida), a single large o rder based on the possession ‘chelae’ m icroscleres (order Poecilosclerida) an d a single large o rder based on the possession of skeletons built in a reticulate arrangem en t o f simple diactinal spicules called ‘oxeas’ and ‘strongyles’ (order Haplosclerida). F reshw ater sponges have so far been included in the latter o rder, b u t are p robably unrela ted (see below). T here are also several less firmly established orders th a t are based upon unique com binations o f non-exclusive skeletal or spicule characters (orders H adrom erida, H alichondrida), o r sm aller groups with unique skeletal o r spicule features (Agelasida, C hondrosida+H alisarcida). T h e integrity o f these groups is currently being investigated using m olecular techniques and proposals to rearrange all ordinal groups and their families is im m inent ([33]; see also below).
Dem osponges dem onstrate a trem endous diversity th a t can only be illustrated w ith a few iconic examples: T h e well-known b a th sponges (family Spongiidae, Fig. 2A) have excellent properties to appeal to hu m an use as a cleaning or scrubbing tool: a softly compressible consistency an d a silica-free resilient skeleton of horny fibres. T hey grow in w arm er waters worldwide an d have been exploited to near-extinction in m any areas. Nowadays, use o f b a th sponges is lim ited to specialized industries and as a curiosity for tourists [34], D eep-sea species o f the genus Thenea (Astrophorida, Fig. 2B), have strongly differentiated hairy stalked bodies specialized in living on bathyal an d abyssal m ud flats, using long laterally spreading spicules an d basal roots. Excavating (or boring) sponges (Fig. 2C) are able to penetrate and erode lim estone surfaces. T hey belong to families C lionaidae (order H adrom erida), T hoosidae (order Astrophorida) and genus A ka (family Phloeodictyidae). T he sponges use acid produced by special cells to etch small ‘chips’ o f calcium carbonate [35] from the substratum and th rough this activity recycle lim estone in e.g. coral reef ecosystems, coralline bottom s an d tem perate oysterbanks. R ock sponges, “ Lithistida” (Fig. 2D), are a polyphyletic group o f sponges with stone-hard silica skeletons com posed o f intim ately interlocking spicules. M any living species are found in deeper waters o f tropical and (warm -)tem perate regions an d are thought to be isolated survivors o f a m uch larger fossil sponge fauna, e.g. [36]. ‘G iant barre l sponges’, e.g. the haplosclerid Xestospongia muta, referred to by some as ‘Redw oods o f the R e e f [37,38], have been estim ated to reach ages o f 2000 years o r m ore in C aribbean seas. A coun terpart species in the Indo-Pacific (X. testudinaria, Fig. 2E) shows com parable sizes and m ay be similarly long-lived. T he A ustralian haplosclerid Amphimedon queenslandica (Fig. 2F) was the first, and thus far only sponge to have its entire genom e sequenced
PLoS ONE I w ww.plosone.org 3 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
Figure 2. Demospongiae morphology and spicule diversity. A. Bath sp o n g e , S pongia o ffic ina lis , G reece (p h o to c o u rte sy E. V oultsiadou); B. Bathyal m u d s p o n g e Thenea sch m id ti; C. Papillae o f excava ting s p o n g e Cliona ce lata p ro tru d in g from lim es to n e s u b s tra tu m (p h o to M.J. d e Kluijver); D. G ian t rock sp o n g e , Neophrissospongia, A zores (p h o to F.M. Porteiro /lm agD O P); E. G iant barre l s p o n g e Xestospongia testud inaria , Lesser S unda Islands, Indonesia (p h o to R. R oozendaal); F. A m ph im edon queenslandica (p h o to o f h o lo ty p e in aq u ariu m , p h o to S. Walker); G. SEM im ag e s o f a se lec tio n o f m icrosc leres a n d m eg asc le re s , n o t to scale, sizes vary b e tw e e n 0.01 an d 1 m m . do i:10 .1371/jo u rn a l.p o n e .0035105 .g002
[39,40]. It proved beyond reasonable doubt that sponges are at the very base o f the M etazoan T ree o f Life.
Carnivorous sponges. Som e sponges o f the order Poecilosclerida, class D em ospongiae, have a surprising carnivorous feeding regim e [4,41,42], instead o f being filter- feeders, as is typical o f sponges. These typically deep-sea sponges lack the aquiferous system and the choanocyte cells w hich are considered to be diagnostic for Porifera [1], M ost display a peculiar sym m etrical shape, generally with lateral appendages lined by hook-like microsclere spicules form ing a sticky ‘velcro’-like cover on w hich prey are trapped. An aquiferous system is m aintained only in the genus Chondrocladia, in which, however, it is apparently not used for w ater filtration bu t for the inflation of turgescent spheres lined by the same sticky cover o f hook-like spicules. T hey prey on a variety o f small invertebrates, mostly crustaceans, w ith setae o r bristles that ensnare on the spicule
cover. In the absence o f any gut o r digestive cavity, digestion is perform ed by cells m igrating tow ard the prey and acting individually to phagocytize an d digest its fragm ents intracellularly [43]. This system is unique in the M etazoa, bu t it parallels the behaviour o f individual sponge cells, w hich perform the various functions o f differentiated tissue, organs an d a nervous system, w hich sponges lack.
By the end o f the tw entieth century, 90 carnivorous sponges were classified in the family C ladorhizidae, w ithin three genera, Cladorhiza, Asbestopluma an d Chondrocladia. T hey were all found in the deep sea, including the dep th record for sponges, w ith a species know n from 8840 m. Increased interest in these sponges, due to the discovery that they are carnivorous, an d due to the developm ent o f m anned submersibles an d R O V s, has shown that this diversity was largely underestim ated. T o date, 119 species are
PLoS ONE I w ww.plosone.org 4 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
known, classified in three families an d eight genera, and several new species and a new genus are in the course o f description.
T h e m orphology o f carnivorous sponges is always erect, bu t is highly diverse (Fig. 3) an d often poorly know n because they are fragile and easily broken during collection in dredges. T heir stalk m ay be a ttached to h a rd substrate by an enlarged base o r rooted in the sediment. Som e are feather-shaped, others are pedunculate with a disc-shaped body bearing radiating filaments, while others have a fan-shaped m orphology w hich m ay be confused w ith that o f hydroids o r gorgonians. Som e Chondrocladia spp. are stalked, with lateral processes ending in translucent inflated spheres.
T h e evidence that this special m orphology is related to a carnivorous habit has been first obtained in an Asbestopluma species living in a cool-w ater littoral cave [4], T he latter species was able to thrive in laboratory conditions, offering excellent study conditions [43]. A lthough a carnivorous regim e is difficult to prove conclusively in the deep sea, it appears likely since several deep-sea sponges sharing this m orphology have shown partially digested crustaceans included in their body, see [44,45].
T h e spicule skeleton, on w hich the classification is based, includes m onaxonic m egascleres that usually include a special type o f tylostyle, a mycalostyle, that builds the axes o f the body and of the appendages, an d a large variety o f microscleres, generally chelae and derivatives, to w hich m ay be added sigmas, sigmancistras, microstyles and forceps. Interestingly, the chelae m icroscleres have no know n function in o ther poecilosclerids, bu t in carnivorous species seem to be used to trap the prey, by lining the surface o f body and appendages w ith the larger hook outw ardly directed.
T h e diversity o f the m icroscleres is rem arkable, especially the apparen t derivatives o f chelae [46-50], in w hich several new types are know n (Fig. 4). T hese microscleres, although diagnostic o f
Poecilosclerida, are not in agreem ent w ith the sub-ordinal classification o f poecilosclerid sponges. T h e family C ladorhizidae lacks a clear synapom orphy [51], an d some sponges with an undoubted carnivorous regim e are classified in the families Esperiopsidae or G uitarridae. T his could m ean either that the classification o f Poecilosclerida needs to be revised, or that carnivory appeared before the separation o f the evolutionary lineages o f Poecilosclerida. M olecular phylogenetic analyses in progress a re a ttem pting to resolve this problem .
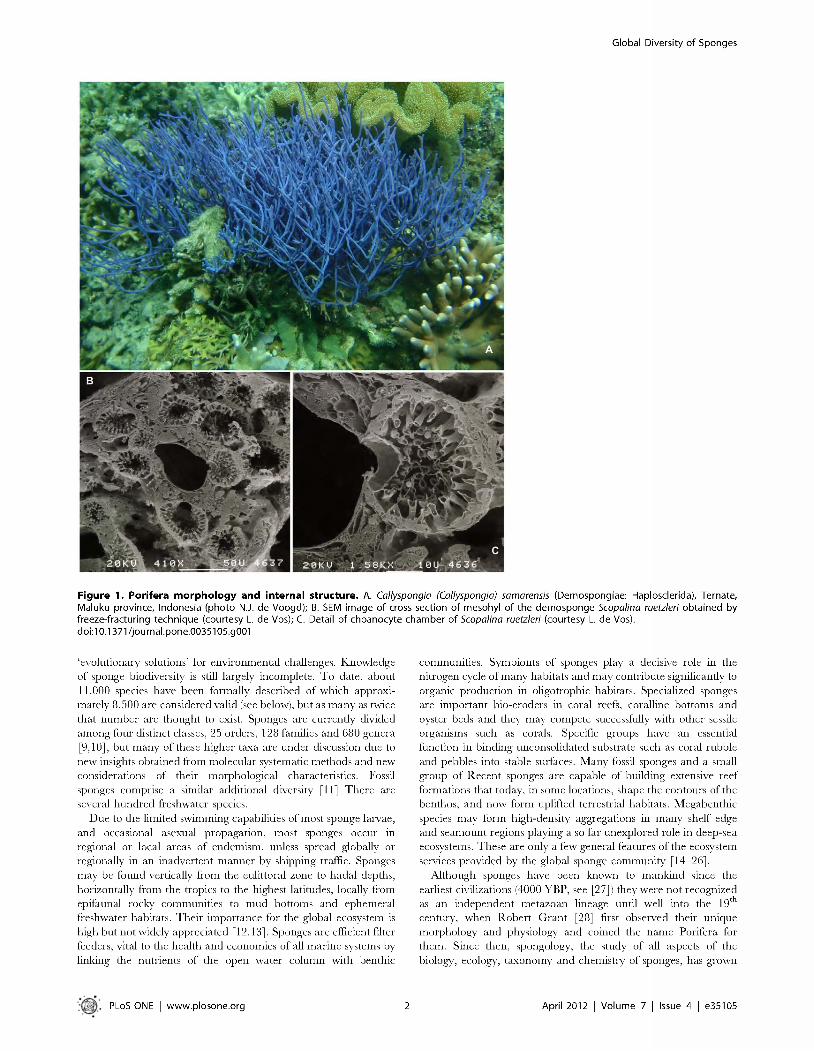
Hexactinellida. H exactinellida, o r glass sponges, are exclusively m arine and m ainly restricted to h a rd an d soft substrates in deeper waters (200 to > 6 0 0 0 m), a lthough they occasionally occur in shallower, scuba-accessible, w ater, such as subm arine caves in the M ed iterranean [52,53], o r off the coast o f British C olum bia w here they form massive structures analogous to M esozoic sponge reefs, e.g. [54—56], T h ey are mostly inconspicuously coloured and highly variable in body shape (e.g. sac-, vase-, blade-shaped, com posed o f b ranch ing tubes etc.; bu t never incrusting). H exactinellids are clearly distinct from other sponges in th a t their soft tissues are largely syncytial and their siliceous spicules have a triaxonic symmetry; they are viviparous and produce distinctive trichim ella larvae (see [57] for a com prehensive review o f glass sponge biology). T h e unusual properties o f their spicules have recently a ttracted the a ttention of m aterials scientists, e.g. [58,59]. Iconic hexactinellids include the venus flower basket (Euplectella aspergillum), w hich often encloses a pa ir o f shrim ps inside its body and was used as a bridal gift in ancient Jap a n , an d Monorhaphis chuni, w hich anchors its body in the soft deep-sea floor w ith a single giant (up to 3 m long) spicule. T o date there are ca. 600 described extant species, w hich is certainly an underestim ate o f their actual diversity, given their rem ote
Figure 3. Carnivorous sponge diversity. A. C ladorh iza abyssicola (from Fig. 17 in [172], scale ap p ro x im ate ); B. C ladorh iza sp., u n d e sc rib e d sp ec ies from W est Norfolk R idge (N ew Z ea land EEZ), 757 m (NIWA 25834); C. Abyssocladia sp., u n d e sc rib e d sp ec ies from B rothers S e a m o u n t (New Z ealand EEZ), 1336 m (NIWA 21378); D. Abyssoclad ia sp., u n d e sc rib e d sp ec ie s from C h ath am Rise (N ew Z ealand EEZ), 1000 m (NIWA 21337); E. Abyssocladia sp., u n d e sc rib e d sp ec ie s from S e a m o u n t 7, M acquarie R idge (A ustralian EEZ), 770 m (NIWA 40540); F. Asbestoplum a (Asbestoplum a) desm ophora, h o lo ty p e QM G 331844, from M acquarie R idge (A ustralian EEZ), 790 m (from Fig. 5A in [47]); G. Abyssocladia sp., u n d e sc rib e d sp ec ie s from S e a m o u n t 8, M acquarie R idge (A ustralian EEZ), 501 m (NIWA 52670); FI. Asbestoplum a hypogea from [41]; I. C hondroc lad ia lam pad ig lobus (from Fig. 17A in [48]). do i:10.1371 /jo u rn a l, p o n e .0035105 .g003
PLoS ONE I w ww.plosone.org April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
Figure 4. Examples of chelae and sigmancistras in carnivorous sponges. A. A rcuate iso ch e lae from Abyssocladia sp., an u n d e sc rib ed sp ec ies from M orgue S e a m o u n t, C h ath am Rise (N ew Z ealand EEZ), 1000 m (NIWA 21337); B. A byssochela from Abyssoclad ia sp., an u n d e sc rib e d sp ec ie s from M orgue S ea m o u n t, C h ath am Rise (New Z ealand EEZ), 1000 m (NIWA 21337); C. A byssochela from Abyssoclad ia carcharias, h o lo ty p e NIWA 62124, from M onow ai S e a m o u n t, K erm adec Volcanic Arc (N ew Z ealand EEZ, [47]), 1071 m; D. A byssochela from Abyssoclad ia sp., an u n d e sc rib e d sp ec ies from S e a m o u n t 8, M acquarie Ridge (A ustralian EEZ), 501 m (NIWA 52670); E. P a lm ate isochelae from Abyssoclad ia sp . (cf.), an u n d e sc rib e d sp ec ies from S e a m o u n t 7, M acquarie R idge (A ustralian EEZ), 770 m (NIWA 40486); F. A n ch o ra te u n g u ife ra te an iso ch e la from C ladorh iza sp., an u n d e sc rib e d sp ec ie s from W est N orfolk R idge (N ew Z ealand EEZ), 757 m (NIWA 25834); G. A n ch o ra te isoche lae from C hondroclad ia (M eliiderm a) tu rb ifo rm is, h o lo ty p e NIWA 21357, from Pyre S e a m o u n t, C h ath am Rise, 1075 m (from Fig. 2D righ t, in [46]); H-l. Palm ate an iso ch e la e from Asbestoplum a sp., an u n d e sc rib e d sp ec ie s from H ikurangi C hannel, o ff G isborne , e a s te rn N orth Island o f New Z ea land , 1119 m (NIWA 32053); J. A nisochela from Asbestoplum a sp., an u n d e sc rib e d sp ec ie s from G houl S e a m o u n t, C h ath am Rise, 922 m (NIWA 21343); K. Pa lm ate an iso c h e la from Abyssoclad ia sp., an u n d e sc rib e d sp ec ie s from S e a m o u n t 7, M acquarie R idge (A ustralian EEZ), 770 m (NIWA 40486); L. P lacochela from Euchelip lum a p ris tina ; M. A nisop lacochela from Asbestoplum a (A sbestoplum a ) anisoplacochela, h o lo ty p e 25835, from T hree Kings Ridge, n o rth e rn N ew Z ea lan d , 1690 m [47]; N. C ercichela from Cercicladia austra lis, h o lo ty p e NIWA 39599, from S e a m o u n t 1, M acquarie Ridge, 1060 m , (New Z ealand EEZ, [49]) (from Fig. 2H, u p p e r left in [49]); O. A n ch o ra te isoche la from Lo llipoc lad ia tib u ro n i (from Fig. 3E left, in [50]); P. S igm ancistra from Asbestoplum a sp., an u n d e sc rib e d sp ec ies from H ikurangi C hannel, o ff G isborne , e a s te rn N orth Island o f New Z ealand , 1119 m (NIWA 32053). do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 0 4
habitats and very small num ber o f taxonom ic experts for the group [60].
H exactinellida is divided into two subclasses, the A m phidiscophora , w hich have am phidisc microscleres, and the H exasterophora, w hich have hexaster m icroscleres (Fig. 5A). A m phidiscoph o ra (Fig. 5B) contains a single extant o rder with three families; am phidiscophoran species exclusively possess skeletons o f unfused spicules. In contrast, H exasterophora is divided into one order with m ostly unfused spicules (Lyssacinosida Fig. 5C with three families) and three orders characterized by fused (dictyonal) m ain skeletons (Hexactinosida [Fig. 5D] w ith 9 families, and Aulocalycoida and Lychniscosida w ith two small families each). Especially the H exasterophora display an astonishing diversity o f spicule forms and skeletal arrangem ents, and this (for sponges) unusual richness o f characters greatly facilitates the delineation of natural taxa. M olecular phylogenetic studies strongly support m onophyly o f H exactinellida and its two subclasses, as well as m ost families and genera sam pled so far [61-64], In contrast, order-level phylogeny and classification w ithin H exasterophora are still poorly resolved since there is strong evidence for paraphyly of H exactinosida w ith respect to Lyssacinosida [61] and DN A sequence da ta for Aulocalycoida, Lychniscosida an d m any families o f H exactinosida are still missing.
Homoscleromorpha. T he H om osclerom orpha com prise a small group o f m arine Porifera with unique features: flagellated opinacocytes and a basem ent m em brane lining bo th choanoderm and pinacoderm , oval to spherical choanocyte cham bers with large choanocytes, and a viviparous cinctoblastula larva. T he skeleton, if present, is com posed o f tetraxonic siliceous spicules with four equal rays (called calthrops) and derivatives showing reduced (diods, triods) or proliferated rays (lophocalthrops). T here is no differentiation betw een megascleres and microscleres, and the spicules are usually small (100 pm or less), no t localized in any particular region [65].
M ost o f the species are encrusting or cushion shaped and the colour varies from creani to blue, violet, green, yellow, deep brow n, orange or red (see Figure 6). T hey are often found in dark or sem i-dark ecosystems (caves, overhangs, coralligenous substratum). H om osclerom orpha are generally located in shallow waters, bu t some species have been found below 100 m [66], T hey have been perhaps overlooked in deep-sea ecosystems due to their encrusting shape.
T h e m onophyly of H om osclerom orpha has been accepted for m any years now [67-69], an d it was assigned to the rank of a subclass o f D em ospongiae [69,1], How ever, m olecular studies have shown that H om osclerom orpha are not a pa rt o f the
PLoS ONE I w ww.plosone.org April 2012 I Volume 7 | Issue 4 | e35105
Global Diversity of Sponges
Figure 5. Hexactinellida diversity. A. Scann ing e le c tro n m ic ro g rap h s o f m icrosc leres (co u rtesy o f H.M. Reiswigi, left: a h ex as te r, th e d ia g n o stic sp icu le ty p e o f su b c lass H ex as te ro p h o ra (scale b a r = 1 0 pm ), right: an am p h id isc , th e d ia g n o s tic sp icu le ty p e o f su b c lass A m p h id isco p h o ra (scale b a r= 100 pm ); B. H yalonem a sp ., an a m p h id isco p h o ran (A m phidiscosida: H yalonem atidae), B aham as; C. A tla n tise lla sp ., a lyssacine h e x as te ro p h o ra n (Lyssacinosida: Euplectellidae), G a lap ag o s Islands; D. Lefroyella decora, a d ic tyona l h e x as te ro p h o ra n ("H exactinosida": S cep tru lo p h o ra : Euretidae), B aham as. B-D co u rte sy o f H arbor B ranch O c ean o g rap h ic In s titu te (Ft. P ierce, Florida, U S A), im ag es tak en from m a n n e d su b m ersib le Jo h n so n -S ea - Link II.do i:10 .1371/jo u rn a l.p o n e .0035105 .g005
D em ospongiae [64,70-73], and recently, H om osclerom orpha was form ally proposed as the fourth class o f Porifera [65]. A m olecular phylogenetic study based on the internal relationships within H om osclerom orpha has shown that aspiculate an d spiculate genera belong to two distinct clades an d the families Plakinidae and Oscarellidae, w hich h ad been m erged in the past have now been restored [74],
H om osclerom orpha is the smallest class o f Porifera w ith two families, 7 genera and 87 species described so far: 16 species o f Oscarella (Fig. 6A-B), and one Pseudocorticium w ithin the family Oscarellidae; 6 species o f Corticium (Fig. 6E-F), 6 o f Placinolopha, 28 o f Plakina (Fig. 6C), 11 o f Plakinastrella, and 19 o f Plakortis (Fig. 6B) w ithin the family Plakinidae. A ltogether, 40 species have been described in the last 20 years, representing an increase o f 42% of the num ber o f H om osclerom orpha. T his clade has thus the highest ra te o f descriptions o f new species [66,75-76], 25% o f the species have been described from the M ed iterranean Sea 10 of w hich since 1992. T his high level o f biodiversity in the M ed iterranean Sea is a reflection o f special efforts undertaken by a M ed iterranean team to find new tools to discrim inate betw een cryptic species. It is predictable that a high diversity o f hom osclerom orph sponges is present in o ther regions such as the C aribbean an d the Indo-W est Pacific.
T h e H om osclerom orpha are considered too difficult to differentiate at the species level due to lack o f diagnostic characters, especially in genera w ithout skeleton (Oscarella), resulting in the perception that m any species are cosm opolitan. T h e high rate o f descriptions o f new species is linked to genetic studies, w hich show that m orphological variability betw een sym patric populations is linked to low levels o f genetic identity betw een them [77,78], All possible m orphological datasets (external features, spicule shapes w hen present [Fig. 6 D -E ], anatom y, cytology, m icrosymbionts) as well as m olecular and chem ical m arkers are used as diagnostic characters to discrim inate betw een these species [74,76,79-83]. T h e cytological dataset o f H om osclerom orpha facilitates discrim ination betw een cryptic aspiculate species o f Oscarella [76-77 ,84 -86], as well as spiculate species o f Plakina [80], M uricy [75] em phasized the benefit o f inclusion o f histological and cytological characters in the taxonom y of o ther spiculate hom osclerom orphs such as Plakortis, Plakinastrella, Placinolopha, and Corticium.
Calcarea. Calcareous sponges have a m ineral skeleton com posed entirely o f calcium carbonate, consisting o f free, rarely linked or cem ented, diactine, triactine, tetractine a n d /o r polyactinal spicules, to w hich can be added a solid basal calcitic skeleton. T h e aquiferous system ranges in complexity from the m ost simple (asconoid and syconoid) to a m ore com plex
PLoS ONE I w ww.plosone.org 7 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
Figure 6. Homosderom opha diversity. A. Oscarella lobu laris (O scarellidae): tw o co lo r m o rp h s from NW M ed ite rran ean Sea (p h o to s c o u rte sy o f Jea n V acelet & Thierry Pérez); B. Plakortis s im p lex (Plakinidae) sp ec im en h a n g in g from th e ceiling o f th e 3PPs cave (NW M ed ite rran ean Sea), a p a rad ise fo r H o m o scle ro m o rp h a sp ec ies (at least 8 sp ec ies b e lo n g in g to 4 d iffe ren t g e n e ra a re p resen t) ; red a rro w in d ica tes th e p re sen c e o f Oscarella m icro lo ba ta a n d a g re e n a rro w P lakina ja n i (p h o to co u rte sy Thierry Pérez); C. Plakina ja n i (Plakinidae) de ta il o f th e lobes, 3PPs cave (NW M ed ite rran ean Sea) (p h o to c o u rte sy Jean V acelet); D. S p icu les o f Plakinidae: triods, d io d s an d lo p h o se c a lth ro p s; E. Spicules o f C ortic ium cande labrum (Plakinidae): c a lth ro p s an d ca n d e la b ru m (h e te ro lo p h o se calth rops); F. C ortic ium cande labrum NW M ed ite rran ean Sea (p h o to s co u rte sy o f Jean Vacelet).do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 0 6
arrangem ent (leuconoid). T he m ode o f reproduction is viviparous and the larvae are always hollow (blástula) [87].
C alcarea are also called Calcispongiae in the o lder literature, and m ore recently in m olecular studies. Som e authors [88-90] propose to use the nam e Calcispongiae for the R ecent representatives to distinguish them from the exclusively fossil H eteractin ida (with polyactine spicules).
Living calcareous sponges are often delicate with th in coalescent tubes (Fig. 7A , C) or m ay be urn-shaped (Fig. 7G). Som e cave- dwelling species are stony (Fig. 7D). M ost o f the species a re white or creani, bu t some species m ay be also red, yellow or pink (Fig. 7A). Calcareous sponges are relatively small, m easured in nini o r a few cm, however in especially rich tem perate estuaries Sycon ciliatum can reach m ore th an 50 cm in length an d 3 cm in d iam eter [91]. Pacific coral reefs m ay also harb o r several larger
PLoS ONE I w ww.plosone.org 8 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
species such as Leucetta avocado and Pericharax heteroraphis, w hich m ay reach 20 cm in height. In m ost textbooks calcareous sponges are regarded as exclusively shallow-water organisms. However, calcareous sponges are repeatedly collected from bathyal and abyssal zones in the N orth A dantic as well as in the Southern O cean [92]. K now ledge o f living calcareous sponges is fragm entary: the total num ber o f described species (ca. 680) represents only abou t 8% o f all described extant sponges. This is partially due to a bias in taxonom ic effort an d the com m on perception that calcareous sponges are difficult to identify. M ore recently, efforts have been m ade to be tte r understand C alcarea diversity in several poorly studied biogeographical areas, e.g. [93-95], and in deep- sea ecosystems [92]. As an exam ple, 67 species o f Clathrina are now known, w ith 22 species described since 2000 [94,96].
T h e m onophyletic origin o f calcareous sponges, w ith their unique m orphological feature o f m onocrystalline calcareous spicules, has never been seriously doubted; m olecular phylogenies using the full 18S an d partial 28S rD N A sequences confirm with high support the m onophyly o f the R ecent C alcarea [97-100].
C urrently , the accepted classification is th a t proposed by B idder [101] following observations by M inchin [102], an d w hich is based on the position o f the nucleus w ithin the choanocytes, the shape of the spicules, the type o f larva and the first type o f spicule to appear during ontogeny. B idder’s classification [101], based on several independent datasets and recognized by several subsequent authors [103-104], was only adopted a t the end o f the 20th century and validated by the first m olecular results [98-100]. T he two clades recognized within R ecen t C alcarea are the Calcinea and the C alcaronea. Calcinea has equiangular triactine spicules (Fig. 7B), a basal nucleus in the choanocytes, a flagellum arising independently from the nucleus, a coeloblastula larva, and triactines as the first spicules to appear during ontogenesis. C alcaronea possess inequiangular triactines (Fig. 7E), an apical nucleus in the choanocytes, a flagellum arising from the nucleus, a stom oblastula larva w hich after eversión (turning inside out) becom es an am phiblastula, and diactines as the first spicules to appear during ontogenesis.
W ith in Calcinea, 166 species have been allocated to two orders (C lathrinida and M urrayonida). W ith in C alcaronea, 515 species have been allocated to three orders (Leucosolenida, L ithonida and Baerida). T h e family G ran tiidae (Calcaronea) has the highest biodiversity w ith 206 species, 138 of w hich w ithin the genus Leucandra.
C ongruence betw een the m olecular results and the curren t classification [87] is no t ap p aren t a t lower taxonom ic levels [88,96,98-100], necessitating a thorough revision th rough an integrative approach.
R ecen t d ev e lo p m en ts fro m m o lecu la r p h y lo g en etic s tu d ie s . Phylogenomics has recently suggested solutions for decades o f differing class-level hypotheses on poriferan phylogeny by showing th a t sponges are m onophyletic, an d th a t classes D em ospongiae and H exactinellida form a sister group to classes C alcarea an d H om osclerom orpha [72]. R ecen t m olecular data also shed new light on the classification and phylogenetic relationships w ithin C alcarea, Hexactinellida and H om osclerom orpha, as briefly m entioned in their respective contributions, bu t D em ospongiae systematics appears to be dem anding m ajor changes.
In D em ospongiae (Fig. 8), the usage o f m olecular systematic techniques revealed weaknesses and inconsistencies o f the m orphology-based classification (for reviews see e.g. [33,105— 106]) and dem onstrated th a t m orphological characters are o f lim ited use, especially a t h igher taxonom ic levels. O f particular interest a re recent studies using m itochondrial and nuclear
ribosom al m arkers, w hich independently [70,107] suggest a deep split betw een (mostly) spiculose, an d (mostly) spicule-lacking dem osponges. T h e latter comprises the K eratosa form ed by the orders D ictyoceratida [including Verticillitida, see [108]) and D endroceratida, and the ‘M yxospongiae’ form ed by Halisarci- da+C hondrosida (which do no t fall in distinct orders, see [105]) as sister group to the o rder V erongida. This implies th a t sponge orders with predom inantly spongin skeletons are no t as closely related as previously assum ed and the aster-type spicules o f chondrosids are no t hom ologous to their hadrom erid or tetractinellid counterparts.
In the clade o f (mostly) spiculose demosponges, the m arine haplosclerid taxa (suborders H aplosclerina and Petrosina) split first. T h e th ird haplosclerid suborder, Spongillina (freshwater sponges), forms a clade in a m ore derived position, leaving H aplosclerida non-m onophyletic. T h e orders H adrom erida, H alichondrida, and Poecilosclerida canno t be recovered m onophyletic either, see details in [105] an d are subsequently proposed to undergo a re-classification based on m olecular results [109]. M olecular data revealed th a t Raspailiidae and Desm acellidae, w hich are poecilosclerid families w ithout the characteristic chelae- microscleres, are unrela ted to the chelae-bearing Poecilosclerida sensu stricto (see details in [105]). Likewise, H alichondrida an d its families have repeatedly shown to be non-m onophyletic in m olecular analyses, w ith some genera closely related e.g. to Raspailiidae o r to the o rder Agelasida. H alichondrid taxa are also found in new taxon compositions (e.g., the re-defined Dictyonellidae an d Axinellidae) or in newly erected families o f yet unclear relationships to o ther taxa (e.g., Scopalinidae) [109]. T h e nom inal family H alichondriidae forms a clade w ith the hadrom erid family Suberitidae. T h e m onophyly of the rem aining hadrom erid families also canno t be dem onstrated. M olecular da ta suggests a h adrom erid clade consisting o f T ethyidae, Hem iasterellidae, T im eidae, an d T rachycladidae, b u t a m onophyletic relationship to o ther hadrom erid families such as Polym astiidae or the closely related C lionaidae and Spirastrellidae still awaits further support [109]. M ore distantly, m olecular da ta indicate a potential close relationship o f some hadrom erid genera and some halichondrid taxa, resulting in a proposed re-erection o f the family Stelligeridae [109]. T h e orders A strophorida and Spirophorida form a m onophyletic group for w hich the previously em ployed taxon T etractinellida can be revived. After inclusion of several ‘lithistid’ families, the m onophyly o f this group and the apom orphic nature o f triaene m egascleres is supported by m olecular data.
T he Sponge B arcod in g P roject. T h e paucity o f com plex m orphological characters in sponges in com bination with a high degree o f plasticity, increased chances o f hom oplasy an d cryptic spéciation m ake species identification difficult even for the expert. M olecular tools have recently been em ployed to a ttem pt to surm ount such shortcom ings o f m orphological taxonom y by the usage o f D N A signature sequences (DNA-Barcoding) [110]. T he Sponge B arcoding Project (ww w.spongebarcoding.org) [111] has been the first barcoding project for a non-bilaterian m etazoan taxon and aims to provide D N A -based identification tools for every poriferan species. C urrently the Sponge B arcoding Project builds up a reference database from type m aterial an d curated collections from various m useums, particularly the Q ueensland M useum , Brisbane.
T he World Porifera D a ta b a seT h e W orld Porifera D atabase (WPD) [10] is an online
searchable catalogue o f all nam es of R ecen t Porifera erected since 1759. T h e catalogue is p a r t o f the W orld R egister o f M arine Species (W oRM S [112], available: h ttp ://w w w .m arinespecies.o rg)
PLoS ONE I w ww.plosone.org 9 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
Figure 7. Calcarea diversity. A. C la th rina rubra (Calcinea, C lathrinida), NW M ed ite rran ean Sea (p h o to co u rte sy Jea n V acelet); B. C alcinean sp icu les: eq u ia n g u la r an d e q u ira d ia te triac tin e s (p h o to c o u rte sy Jea n V acelet); C. G uancha lacunosa (Calcinea, C lathrinida), NW M ed ite rran ean Sea; D. Petrob iona m assiliana (C alcaronea, L ithonida), tw o sp ec im e n s from caves, NW M ed ite rran ean Sea. Spicule c o m p le m e n t o f P. m assiliana: from left to r igh t p u g io le , sag itta l triac tines, m icrod iac tine (p h o to s co u rte sy J e a n V acelet); E. C alcaronean sp icu les: sag itta l (inequ iangu lar) triac tin es an d d iac tines; F. S yconoid aq u ife ro u s system from Sycon c ilia tu m (SEM p h o to , co u rte sy Louis De Vos, ULB); G. Sycon c ilia tu m (C alcaronea, L eucoso len ida), sp ec im e n a b o u t 10 cm , from th e English C hannel. do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 0 7
hosted by the Flanders M arine Institute (VLIZ), O ostende, Belgium. It is an aim o f the W PD to be the w orld standard for sponge nam es and the w orld portal for in ternet access to inform ation on Porifera. W ith its expert team of editors, the W PD acts to stabilize an d regulate the use o f sponge nam es in science and society. It serves as a tool for taxonom y by facilitating inventories o f taxa, literature references, distributional data, and knowledge gaps. A great advantage over traditionally published inventories is the continuous updating that takes place with each
new item o f taxonom ic inform ation that becom es available in the literature. C urrently, the W PD contains approx. 20,000 taxon nam es o f w hich approx. 8,500 are considered valid (see T able 1).
Basic data. T able 1 lists the fields and their contents for a standard entry in the W orld Porifera D atabase. M ost fields are linked to further entries an d subsidiary databases. E ach entry page contains navigation buttons to various sections o f the database (Introduction, Species, D istribution, Checklist, Sources) and contact buttons for editors an d database m anagers.
PLoS ONE I w ww.plosone.org 10 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
("G4”)
“m arine hap losclerids”
•'M yxospongiae''
I Raspailiidae I Desm acellidae I Poecilosclerida,I Suberitidae+H alichondriidaeI Tethyidae+H em rasterellidae+Tim eidae+Trachycladidae I C lionaidae+Spirastrellidae I Polym astiidae I Axinellidae I Dictyonellidae I Scopalinidae I Stelligeridae I Agelasida I Tetractinellida I Spongillina
I H ap losc le rina + P e tro s in a
C h o n d ro s id a (incl. H a lisa rc id ae )
V erong ida
“K eratosa" D en d ro cera tid a
D ictyoceratida
Figure 8. Phylogenetic relationships of higher demosponge taxa as evident from various molecular phytogenies. Sources e.g., [70,107,109], The ap p ro x im a te co m p o sitio n o f th e "G 4" su b tax a is know n , b u t th e p h y lo g e n e tic re la tio n sh ip s o f th e se a re still to b e a sse ssed . do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 0 8
Geographic entries. T he W oR M S database architecture provides various geographic resources w hich can be linked to the taxon entries. Editors can choose betw een three com peting global
geographic classification systems: terrestrial, oceanic or‘alternative’. T h e first two classifications are nation-oriented (for the oceans the basic system is the Exclusive Econom ic Z one (EEZ)
Table 1. Standard record of the World Porifera Database (available: w w w .m arm especles.org/ponfera, accessed 2011 Aug 31) with field names (left column) and con ten t of each field (right column).
Field C o n te n t
ta x o n n am e e.g. genus and species combination with authorship and year, or ditto for family and higher taxon names, including unique database number
C lass ifica tion hierarchical, collapsible higher taxa names to which the taxon belongs
S tatus accepted or unaccepted, checked or unchecked by taxonomic editor
R ank species, genus etc.
p a re n t ta x o n first higher taxon
s y n o n y m ize d taxa each linked to its own entry
ch ild ta x o n nam es each linked to its own entry
source re fe ren ce e.g. source of original description, basic source of current classification, additional sources
E n v iro n m e n t marine, brackish, freshwater or terrestrial
fossil ran g e Recent only, fossil+Recent, fossil only, unknown
d is trib u tio n linked to pages containing source references and additional data, including a summary map
specim en link to pages containing type specimen information and source references, additional data on individual specimens
Links buttons linking to other internet resources e.g. Encyclopedia of Life, PESI, Genbank etc.
N otes any additional information or explanations of entries
Im ages thumbnails linking to photos and o ther illustrations
Lsid unique species nam e reference number
e d it h is to ry who created or changed the entry when
T re e lin k and G o o g le lin k links to Taxonomic Tree, Google, Google Scholar, Google Images
C ita tio n requested way of citing the entry
doi:10.1371 /journal.pone .0035105. t001
p | ) . PLoS ONE I w w w .p lo so n e .o rg 11 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
o f countries). A m ong the alternative classifications are FA O Fishing Areas, Longhurst Provinces an d M arine Realm s (also know n as the M arine Ecoregions o f the W orld (M EOW s), see [30], available: h t tp : / /w w w .w orldw ild life .org/science/ecoreg ions/m arin e /item 1266.html). For the W orld Porifera D atabase em phasis is based on the M E O W system because it is constructed from anim al distribution patterns an d is also the m ost refined, and the only h ierarchical system o f the existing alternative classifications. From a scientific poin t o f view, this appears to provide a good opportunity to explore distribution patterns o f sponges (see below), a lthough dep th occurrence cannot be properly docum ented. Proposals for im plem entation o f G lobal O pen O ceans and D eep Sea-habitats (G O O D S) bioregional classification (h ttp :// w w w .ias.u n u .ed u /re so u rce_ cen tre /o cean % 20bioregionalisation. pdf), w hich accom m odates open-ocean a n d deep-sea d istribu tions, have no t yet b een honored , a n d this is anxiously aw aited. T h e W PD editors are also in the process o f en tering the EEZ occurrences as this m ay facilitate retrieval o f inform ation dem an d ed by n a tion states.
Completeness. L iterature on the taxonom y o f sponges is scattered over thousands of journals and dozens of books spanning a 250-year period, so any claim o f com pleteness is b o u n d to be false. Nevertheless, thanks to inform al card systems and early electronic nam e lists, a basic catalogue was entered relatively quickly into the W oR M S systems. T h e T axonom ic T ree a t the heart o f the W PD was provided by the editorial team o f the System a Porifera [9], so we can rely on this resource for completeness o f all taxa dow n to the level o f genus and subgenus. O nline sources such as the Biodiversity Fleritage L ibrary (available: h ttp ://w w w .b iod iversity lib rary .o rg /), N om enclátor Zoologicus (available: h t tp :/ /u io .m b l.e d u /N om enclatorZoologicus/), T h e Zoological R ecord O nline (h ttp ://w w w .o v id .c o m /site /cata lo g /D a taB ase/2 0 0 .jsp ), an d o ther similar resources allowed quick retrieval o f (older) literature records.
All in all, we believe that nam es of all h igher sponge taxa and species nam es for all extant sponges are virtually completely present in the W PD . This does not imply th a t all com binations o f species nam es an d genus nam es are incorporated, bu t original com binations and accepted com binations have been entered to the best o f our ability. If a com bination cannot be found in the W PD it usually m eans that it is neither an original nor an accepted com bination.
Accepted and unaccepted names. O riginal com binations can be declared unaccepted for two reasons: (1) a published statem ent o f synonym y by one or m ore taxonom ists underbuilt by argum ents, (2) an im plied synonym y based on the Systema
Porifera [9]. An exam ple for the latter reason w ould be that when a particu lar genus is considered a ju n io r synonym o f ano ther older genus by one o f the authors o f the Systema Porifera then all species described in the ju n io r genus are autom atically transferred to the older genus even though in m ost cases there is no published statem ent. T h e System a Porifera usually only discusses the type species o f genera, leaving the status o f the rem aining species to subsequent reviewers o f the genera. I f these species were left in their original com bination, the structure o f the T axonom ic T ree o f the W PD w ould have been com prom ised. For largely the same reason, the W PD can only accom m odate taxon nam es following the In ternational C ode of Zoological N om enclature, as rivaling codes are incom patible. Species com binations that do not fall under reasons 1 o r 2 are considered accepted for the tim e being, unless they are know n insufficiently to assess their genus affinity, in w hich case they are declared ‘species inqu irenda’.
S p o n g e diversityNumbers of taxa tabulated. Based on the above
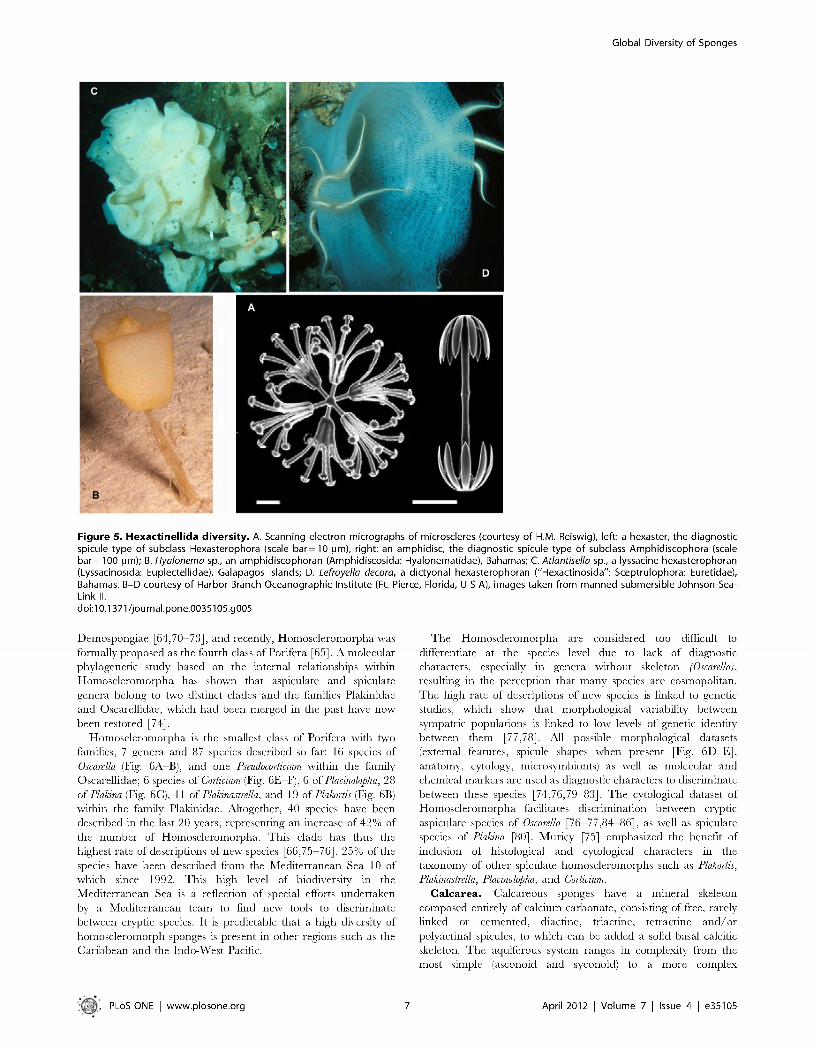
considerations, T ab le 2 lists the num bers o f W PD entries o f species and lower-level com binations (varieties o r subspecies) o f the four recognized classes. So far (2011 August 31) the num ber o f accepted species o f Porifera is 8,553, the vast m ajority o f which (83%) are D em ospongiae (Fig. 9). T h e num ber o f ju n io r synonyms is currently approxim ately 28% o f the num ber o f accepted species. A striking difference in num bers o f ju n io r synonyms is observed in the three small classes (respectively 5% , 9% and 3%) as opposed to the D em ospongiae (32%). This reflects an overall low scientific effort in the study of these smaller classes: H exactinellida have a ttracted few taxonom ists over prolonged periods o f time, p robably due to the perceived difficulty o f identification and a lack o f taxonom ic resource m aterial caused by predom inantly deep-sea occurrence. Despite ubiquitous occurrence in m any habitats, C alcarea have been neglected as well, possibly because of their small size and ap paren t uniform ity o f characters. H om osclerom orpha were only recently separated from D em ospongiae [65] and like C alcarea show few classical differentiating features.
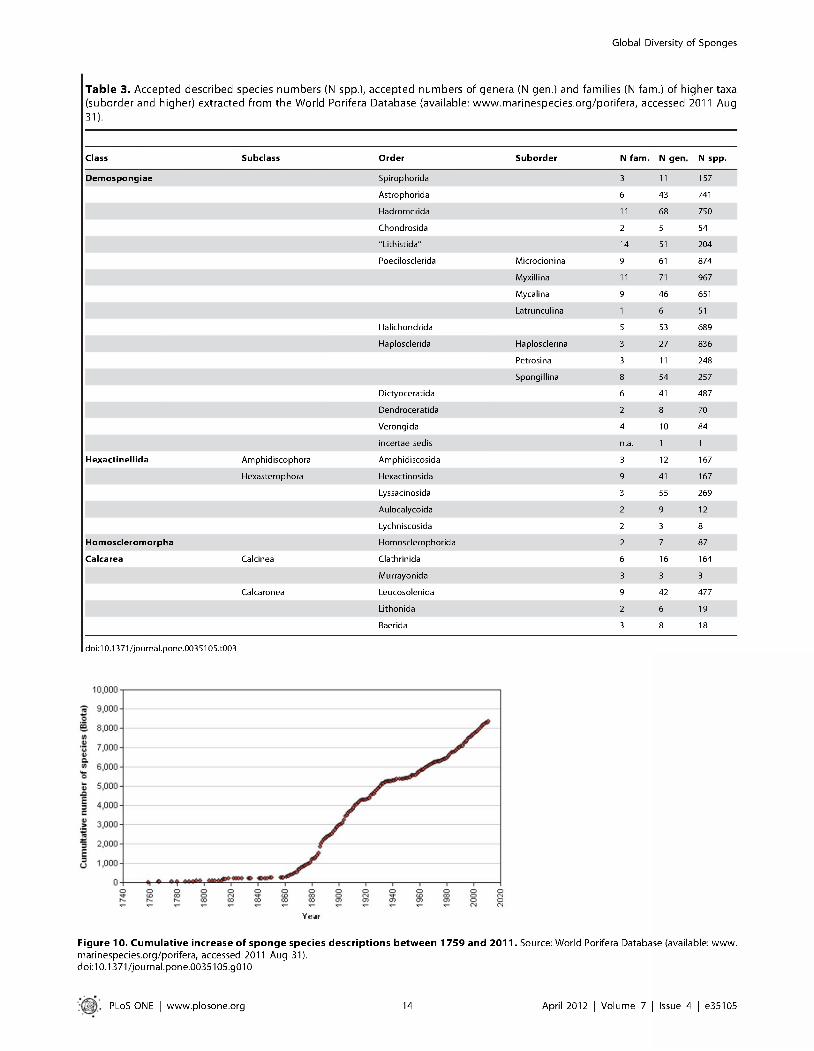
T h e num ber o f sponge taxa increases steadily a t a ra te o f 35-87 each year, w ith lim ited variations over the years, bu t a striking difference in the num ber o f ‘au thors’ for a single new species is apparen t over the last century, with an overall single au tho r for each nam e before the 1980s and a growing num ber o f authors after that. Apparently, species recognition is nowadays a team effort necessitating inclusive authorship.
T ax a equal to or above the (sub-)genus level entered in the W PD num ber 2004 (see T able 3) overall, approxim ately ha lf o f
Table 2. Described species num bers of the four Porifera classes and total num ber of Porifera species extracted from the World Porifera Database (available: w ww.m arm especles.org/porlfera, accessed 2011 Aug 31).
T axon A c cep ted co m b in a tio n U n accep ted co m b in a tio n U n accep ted c o m b in a tio n T o ta l Entries
ju n io r syn onym gen us tra n s fe r
D e m o s p o n g ia e 7164 2314 6552 16030
H e x a c tin e llid a 623 33 427 1083
H o m o s c le ro m o rp h a 87 3 108 198
C alcarea 681 64 588 1333
T o ta l species 8553 2414 7675 18644
A ccepted com bination: valid species com binations according to th e WPD. U naccep ted described species num bers divided in colum ns 'junior synonym ' an d 'g enus transfer' com binations o f e ith e r accep ted nam es o r synonym s. doi:10.1371 /journal.pone.0035105.t002
PLoS ONE I w w w .plosone.org 12 April 2012 I Volume 7 | Issue 4 | e35105
Global Diversity of Sponges
Accepted species by Class
Hc**ctincllk}#8%
C ito re* Momowierofncrph*8% »
- Hexactinellida
- Caicatr*
H om osclerurnorpha
- D em ospongiae
OemotpongiM83%
Figure 9. Percentual species diversity of the four classes of sponges. S o u rc e : W orld P o r ife ra D a ta b a s e (a v a ila b le : w w w . m arin e sp ec ie s.o rg /p o rife ra , acce sse d 2011 Aug 31). do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 0 9
w hich (1026) is currently considered accepted, mostly based on conclusions derived from the System a Porifera. As explained above, h igher taxa are under enhanced investigation using m olecular sequence data . R earrangem ents a t all levels are anticipated in the n ear future.
Numbers of taxa collected but not yet described. T here is a large num ber o f ‘unknow n’ species: h idden in the m any collections worldwide are num erous sponge species aw aiting description. Unlike m ost o ther m arine taxa sponges show dram atic post-collection preservation changes in habit and color, m aking com parison w ith living m aterial difficult unless the species has a uniquely recognizable form. Unless good images o f living m aterial are available, discovery o f new taxa is alm ost invariably a m atter o f com paring preserved samples w ith type m aterial o f previously described species. It is no t uncom m on to discover undescribed sponges in collections that have been preserved for a hundred years or m ore. T h e building of a database o f in situ images in com bination w ith classical im aging an d taxonom ic descriptions should alleviate the curren t im pedim ent o f post-collection species discovery (see also below).
Numbers of taxa expected to be extant. T h e cum ulative num ber o f described species is increasing a t a steady rate (Fig. 10) and there is no indication that it is asym ptotic. R egional species accum ulation curves m ay differ as is the case for Australia, where effectively sponge discovery halted after the 1920s and was taken up again only in the last few decades. This caused a dram atically stepped discovery curve w ith a m uch steeper-angled increase in recent decades. Following the global curve, it is likely th a t at the end o f the present century the num ber o f know n Porifera species will have risen to at least 12,000. New techniques and increased efforts m ay well accelerate species discovery beyond that. An extra boost in the num ber o f described species m ay be expected w hen a posteriori m orphological studies o f previously recognized ‘cryptic’ species, i.e. sponges showing genetic distinctness in the absence of m orphological differentiation, are launched in earnest, similar to pioneering studies o f e.g. [77,113-116], A persistent problem , preventing the form al recognition of such cryptic species, is the
lack of m orphological evidence o f such differentiation at the genetic level, e.g. [117]. This is the cause of a w idespread reluctance to describe and nam e these potentially thousands of putatively new species.
Numbers of freshwater sponges. Freshw ater sponges are united in the suborder Spongillina (Class Dem ospongiae), num bering approxim ately 200-250 accepted species. A W PD search p roduced 257 accepted species, w hereas only 219 are acknowledged in [118], the m ost recent overview o f the freshwater sponges. Spongillina are distributed over all continents except A ntarctica, an d show high endem ism w ith the exception o f a few w idespread species such as Spongilla lacustris and Ephydatia fluviatilis. T h e suborder is divided into six families (and an incertae sedis com plem ent), the largest o f which, Spongillidae, contains m ore th an ha lf the num ber o f species. T h ere has been some debate over the likelihood of m ultiple invasions o f the freshwater hab ita t by sponges, so prudency dictates that the issue rem ains unsolved [118], Flowever, curren t knowledge o f phylogeny and distribution favours a single Palaeozoic invasion linked evolutionarily to the developm ent o f specialized resting stages (called gemmules) found in m ost freshwater sponges all over the globe.
S p o n g e d is tr ibu t ion sGlobal distributions. C om prehensive analyses o f
distribution patterns have been m ade previously only for the classes D em ospongiae [119] and hexactine llida [120], based on global distributions o f all taxa o f these classes. T h e m ethod o f these studies was tracing distributions over large pre-conceived areas o f endem ism . M any m ore such studies were done in m ore lim ited geographic areas, such as M editerranean-A tlantic [121-123], and A ntarctica [124-125]. M ore sophisticated attem pts at analytical biogeography were invariably m ore lim ited in their scope regarding a rea a n d /o r taxon coverage, e.g. those using biogeographic indices and com plicated statistical treatm ent (M editerranean- A tlantic areas [126], A ustralia [127] (see also below), and South Africa [128], o r areacladistic analyses (four unrela ted genera [129]; suborder M icrocionina [130]; genus Mycale [131]; 20 selected genera [132]. Panbiogeographic analysis w ith selected dem osponge genera and families was a ttem pted by [133-134], Recently, phylogeographic studies em ploying various genes a t the infraspecific o r supraspecific levels were perform ed w ith several species complexes in lim ited geographic areas o f the N ortheast A tlantic (Cliona celata [115], Phorbas fictitius [135], deep-w ater Hexadella [116] an d Plocamionida. [136]) an d the Indo-W est Pacific (Leucetta chagosensis [137]). All these studies w ere diverse in m ethodology and taxon content, an d it is no t possible to arrive at a com prehensive sum m ary at this m om ent in time.
H e re we will large ly confine ourselves to revisit th e b ro a d e r com p reh en siv e ap p ro ach es m ad e ea rlie r by sim ply m ap p in g th e d is tr ib u tio n a l d a ta from th e W o rld P orife ra D a tab ase in to a n u m b er o f g lobal m aps b ased on the schem e [30] o f the M arin e E coreg ions o f th e W o rld (M EO W s), availab le: h t t p : / / w w w .w orldw ild life .o rg / sc ie n c e /e c o re g io n s /m a r in e / ite m 1266. h tm l. D a ta sets w ere co m b in ed in a g eo graph ic in fo rm atio n system (GIS) softw are (E SR I A rcG IS v9.3 [138]), thus n u m b ers o f species, gen era , a n d fam ilies w ere p lo tte d in to m arin e R ealm s, m arin e P rovinces, a n d M E O W s. F ro m all the m aps th a t we have g e n e ra te d for this study (see the links to in d iv idua l m aps) a c lear collection bias is ev iden t. T h is is d e m o n s tra ted in Fig. 11, w h ich p ic tu res th e species c o n te n t o f all M E O W s. In v ariab ly , th e m ost diverse a reas a p p e a r to be in th e N o rth e as t A tlan tic , a n d in m ore ideta il th e M e d ite rran e a n - A tlan tic areas, w hereas the tro p ica l co ral re e f regions, rep u ted ly th e m ost rich a reas, com e ou t w ith low er diversities.
PLoS ONE I w ww.plosone.org 13 April 2012 I Volume 7 | Issue 4 | e35105
Global Diversity of Sponges
Table 3. A c c e p te d d e s c r ib e d s p e c i e s n u m b e r s (N s p p .) , a c c e p t e d n u m b e r s o f g e n e r a (N g e n .) a n d fa m il ie s (N fa m .) o f h ig h e r ta x a ( s u b o r d e r a n d h ig h e r ) e x t r a c t e d f r o m t h e W o rld P o r ife ra D a ta b a s e (a v a ila b le : w w w .m a r in e s p e c ie s .o r g /p o r i f e r a , a c c e s s e d 2011 A u g 31).
Class Subclass O rd e r S u b o rd e r N fa m . N gen N spp.
D e m o s p o n g ia e Spirophorida 3 11 157
Astrophorida 6 43 741
Hadromerida 11 68 750
Chondrosida 2 5 54
"Lithistida" 14 51 204
Poecilosclerida Microcionina 9 61 874
Myxillina 11 71 967
Mycalina 9 46 651
Latrunculina 1 6 51
Halichondrida 5 53 689
Haplosclerida Haplosclerina 3 27 836
Petrosina 3 11 248
Spongillina 8 54 257
Dictyoceratida 6 41 487
Dendroceratida 2 8 70
Verongida 4 10 84
incertae sed is n.a. 1 1
H e x a c tin e llid a Amphidiscophora Amphidiscosida 3 12 167
Hexasterophora Hexactinosida 9 41 167
Lyssacinosida 3 55 269
Aulocalycoida 2 9 12
Lychniscosida 2 3 8
H o m o s c le ro m o rp h a Homosclerophorida 2 7 87
C alcarea Calcinea Clathrinida 6 16 164
Murrayonida 3 3 3
Calcaronea Leucosolenida 9 42 477
Lithonida 2 6 19
Baerida 3 8 18
doi:10.1371 /journal.pone.0035105.t003
10,000
9,000-
E 8,000-
7,000-
“ 6 ,0 0 0 -
5,000-
§ 4,000-
2 ,0 0 0 -
1 ,0 0 0 -
o'd- O■X-
o oo o o~r OX '
o oo o■ J
oT
o■X o oQ o
Y ear
Figure 10. Cumulative increase of sponge species descriptions between 1759 and 2011 . Source: World Porifera Database (available: www.marinespecies.org/porifera, accessed 2011 Aug 31).doi:10.1371/journal.pone.0035105.g010
PLoS ONE I w w w .plosone.org 14 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
SpeciesH o 1 - 100 101 -2 0 0 201 -4S1
Figure 11. Global diversity of the Porifera. N um bers o f s p o n g e sp ec ie s re co rd ed in e a c h o f 232 m arine eco reg io n s o f th e w orld [30] e x trac ted from th e W orld Porifera D a tab ase (available: w w w .m arin e sp ec ie s .o rg /p o rife ra , acce sse d 2011 Aug 31). The ty p e localities an d ad d itio n a l con firm ed o c cu rre n ces in n e ig h b o rin g a re as o f a lm o s t all 'a c c e p te d sp ec ie s ' w e re e n te re d in o n e o r m o re o f th e M arine E coregions o f th e W orld, b u t m any n o n orig inal d is trib u tio n re co rd s in th e lite ra tu re a re still to b e ev a lu a te d an d e n te re d . M oreover, m any s p o n g e taxa a re re co rd ed in th e lite ra tu re u n d e te rm in e d an d th e se a re n o t inc lu d ed in th e WPD. Thus, th e d a ta p re s e n te d he re a re to b e co n sid e re d a co n se rv a tiv e o r 'm in im al' e s tim a te o f th e ac tu a l d is trib u tio n a l d a ta . do i:10 .1371/jo u rn a l.pone.0035105 .g011
W e are fo rced to co n clu d e th a t c u rre n t know ledge as la id dow n in th e W PD is likely defic ien t in show ing less th a n the ac tu a l d iversity p a tte rn s o f sponges. M an y m ore a reas rem a in to be ex p lo red a n d m an y re co rd e d u n d e te rm in e d tax a rem a in to be n am ed .
A dditionally, we present a prelim inary biogeographic analysis o f the aptness o f the M E O W scheme as a tool for representing sponge distributions. In the next section, we will present a clustering o f B ray-Curtis indices obtained from com parisons o f the sponge contents o f all Realm s, Provinces an d M E O W s. Finally, we will discuss the advantages o f a regional approach and briefly review w hat is know n about alien sponge invaders.
Marine Realms. At the R ealm level species num bers (File SI p a rt A), disregarding the high num ber in the N orth Atlantic for reasons explained above, do reflect partially a p a tte rn that is found in m any o ther m arine groups: highest diversity in the C entral Indo-Pacific, som ewhat lower in the W estern Indo-Pacific, lower again in the T ropical A tlantic and lowest in the E astern Indo- Pacific an d T ropical E astern Pacific. T h e latter two realm s appear severely understudied. T em pera te Southern realm s probably correctly show highest diversity in T em pera te Australasia with its m ore extended habitats and island groups. T h e Southern O cean appears to h a rbour m ore species th an the Arctic for the same reasons. G enus (File S I p a rt B) and family (File SI pa rt C) num bers show similar results.
Marine Provinces. Species patterns (File S2 p a rt A) are som ewhat surprising, w ith the T ropical W estern Atlantic province as the m ost diverse province closely followed by the N ortheastern A tlantic provinces and the Indo-W est Pacific provinces at some distance. G enus (File S2 pa rt B) an d family (File S2 pa rt C)
patterns are similar, although the differences betw een the family diversity o f circum global tropical areas and the M editerranean and L usitanian provinces are minim al.
Marine Ecoregions. Species patterns (Fig. 11) are com plicated and difficult to sum m arize. M E O W s with high species num bers m ay be adjacent to very poor ones, often explained by hab ita t differences (e.g. the 295 species recorded for the Celtic Sea are contrasted by 140 species o f the N o rth Sea for reasons o f lack o f h a rd substratum in the latter region), bu t very often also because exploration has been differently intense (e.g. in the South A ustralian M EO W s). Genus (File S3 p a rt A) and family distributions (File S3 p a rt B) are less extrem ely different in m any M E O W s and probably reflect a m ore realistic diversity o f sponges over the M E O W s m ore closely th an the species distributions.
Selected higher taxa patterns: Classes. W e provide m aps o f the species num bers a t the R ealm (File S4) and the M E O W level (File S5). T h e dem osponge distributions (File S4 pa rt A and File S5 p a rt A) are closely similar to those corresponding to all sponges (see above). H exactinellida distributions (File S4 pa rt B for Realm s an d File S5 pa rt B for M EO W s) look surprisingly com m onplace, with highest num bers in the W est Pacific, bu t the m aps are deceitful by not revealing the predom inantly bathyal and abyssal occurrence o f these sponges. C alcarea patterns (File S4 pa rt D for Realm s an d File S5 p a rt D for M EOW s) are obviously biased, with highest num bers in South A ustralia and Ja p a n and very low num bers in the tropics, reflecting an alarm ingly low exploration and description status. Please note that this is the first tim e a com prehensive m ap of global C alcarea distributions has been published. H om osclerom orpha is a small group with m uch of the effort concen tra ted in the M editerranean, bu t the distribution
PLoS ONE I w ww.plosone.org 15 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
at the R ealm and M E O W level (File S4 pa rt Cl and File S5 p a rt Cl, respectively) appears to be largely confined to w arm er waters.
Selected higher taxa patterns: Genera. W e provide some exam ples o f distinct generic distribution patterns, w hich were already observed in [119] an d later studies. Com m only, genera occur circum globally in b ro ad er o r narrow er latitudinal zones. Increasingly, patterns th a t appeared disjunct or restricted at the tim e have since been found to be m uch m ore continuous. Exam ples o f such patterns are: virtually cosm opolitan, e.g. Tedania (File S6 pa rt A), w arm -tem perate, e.g. Spongia (Fig. 12), circum tropical, bu t lacking in the tropical East Pacific an d W est Africa as found in Agelas (File S6 p a rt Cl), and b ip o la r /antitropical,e.g. Iophon (File S6 pa rt B). V ariations on these com m on distributions are e.g. cosm opolitan with a cold-w ater bias as in M yxilla (S6 p a rt D), b ipo lar and cosm opolitan deep-sea as in Asbestopluma (File S6 p a rt F), and restricted tropical as in Carteriospongia (File S6 p a rt E), w hich is not found outside the Indo-W est Pacific. Interestingly, such distribution patterns are not predicted by the h ierarchical system [30] o f M EO W s, Provinces and Realm s: there are no cosm opolitan, bipolar, o r circum tropical units distinguished. Obviously, the m arine ecoregion subdivision scheme is based on species distributions as they are observed today, lacking biogeographic history. It needs similarity studies to explore such disjunct patterns.
Biodiversity analysis: hierarchical clustering of MEOW contents. W ithin P R IM E R -6 (PR IM ER -E package) p resen ce / absence sponge species da ta were used to perform a hierarchical cluster analysis at R ealm , Province (> 50 records) an d M E O W (> 2 0 records) level. In Fig. 13 the dendrogram is given at R ealm level an d four assem blage types were identified a t various degrees o f similarity. T h e 12 different Realm s contained records differing from the lowest num ber o f species present in the Arctic and T em pera te Southern Realm s (both 310 spp.) and the highest num ber o f sponge species present in the T em pera te N orthern A tlantic (1664 spp.) and the C entra l Indo-Pacific (1325 spp.). T he different types o f assem blage identified represent either the m ajor oceans o r a b ip o la r /antitropical distribution. For instance, the C entra l Indo-Pacific is m ost similar to the W estern Indo-Pacific together with T em pera te Australasia (including Shark Bay and H o u tm an Abrolhos); the T em pera te N orthern A tlantic is most similar to the T ropical A tlantic and the Arctic; the Southern O cean clusters together w ith T em pera te South A m erica and T em pera te Southern Africa. T h e Realm s w ith the lowest num ber o f records cluster together and have a low similarity (Eastern Indo- Pacific an d T ropical E astern Pacific), and these Realm s only
Figure 12. W arm -tem perate distribution of the genus S p o ng ia .All k now n sp ec ie s o f th e g e n u s reco rd e d w e re e n te re d in th e re levan t M arine E coreg ions o f th e W orld [30], y ie ld ing th e c ircum global w a rm er w a te r d is t r ib u t io n o f th is g e n u s . This ty p e o f d is t r ib u t io n is re p re sen ta tiv e fo r a la rge n u m b e r o f s p o n g e g en era . do i:10 .1371/jo u rn a l.p o n e .0035105 .g012
consist o f a few ecoregions o f w hich m any have no sponge records at all. T hey do no t only reflect a low exploration status bu t also the seclusion of their geographical position (e.g. Galapagos, Cllipperton, and Polynesia). T h e endem ism of some o f the m arine ecoregions becom es clearer in the dendogram at the M E O W level (File S7), bu t in general there are few assemblages at this level that conform to the Provinces distinguished in [30],
T h e dendogram given a t Province level (File S8) also does not clearly represent a nested system w here the Provinces are clustered w ithin their R ealm . M ost Provinces are clustered together adjacent to their closest geographical Province. For instance the Provinces nested w ithin the W estern Indo-Pacific R ealm are found together with m ost Provinces o f the C entra l Indo-Pacific R ealm (with exception o f the C entral Ind ian O cean Islands and the South C hina Sea). T h e similarity o f the different identified clusters is very low, reflecting again the low exploration status in m any o f the Provinces. T h e Provinces with a high num ber o f records clearly follow the position of the oceans bette r than provinces w ith a low num ber o f records.
Freshwater sponge distributions. As these w ere the subject o f a recent contribution to the G lobal Diversity o f Freshw ater habitats series [118] we will confine ourselves to cite several o f the conclusions from th a t study. D istributions were tabu lated in seven classical terrestrial regions (Palaearctic, Nearctic, N eotropical, Afrotropical, O riental, Australasian and Pacific Oceanic). T h e m ost diverse region is the N eotropical region with m ore th an 65 species, closely followed by the Palaearctic regions with a round 60 species. Smallest num bers are found on Pacific O ceanic Islands (5 species) and this is also the case for the C aribbean.
A t the family an d genus level there are some interesting m ore restricted distribution patterns. T he ancient lakes each have distinct endem ic species and genera, an d the family Lubom irskiidae is restricted to Lake Baikal, the family M etschnikow iidae to the C aspian Sea, and the family M alaw ispongiidae to the Rift lakes. T h e family M etaniidae appears restricted to the tropical rainforest belt o f all continents, w hich m ay be in terpreted as a typical G ondw ana distribution. This is possibly also the case for the family Potam olepidae, bu t m em bers o f this family are so far no t found in O rienta l and Australasian forests.
Regional data systems and online identification tools. Progress o f knowledge o f global sponge diversity is generated predom inantly in m any regional efforts, m ost p re em inently in the A ustralian region (Fig. 14). Sim ilar to, bu t a t that tim e independent o f the W o R M S /W P D global effort was an A ustralian regional inventory o f the “know n” sponge fauna from the A ustralian m arine territories (amongst the largest in the world, w ith 6,819,501 km~ o f seabed jurisdiction, an d also the largest in term s of the num ber o f described m arine species, 32,900 so far [138]). Since the sponge com ponent o f this fauna had largely been untouched since the early 20th century, it also requ ired an a ttem pt to significantly revise this know n fauna w ithin a contem porary systematica (ZCIA [139]). T h e initial hardcopy publication listed 1,385 valid species-group nam es an d 338 genus-group nam es. T he subsequent online version (the AFD [140]) currently contains 1,650 species an d subspecies in 330 genera an d 102 families.
Know ledge o f regional m arine sponge diversity has expanded considerably over the past two decades in particular, thanks to m any new biodiversity discovery initiatives. These range from m any small-scale local studies to m am m oth voyages over larger continental spatial scales. A few exam ples are the G reat B arrier R eef Seabed Biodiversity project in northeast Australia (GBRSBD[141]), and the N orthw est Shelf project in W estern A ustralia[142]. Elsewhere recent expeditions were held and regional guides
PLoS ONE I w ww.plosone.org 16 April 2012 I Volume 7 | Issue 4 | e35105
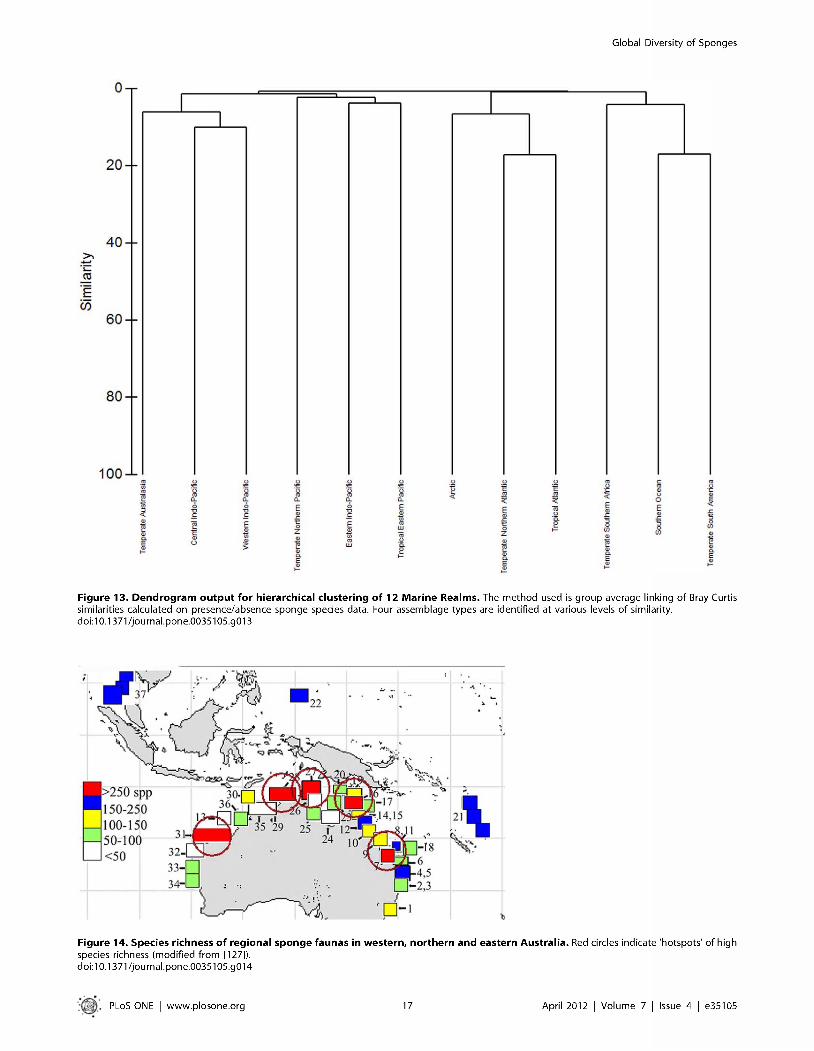
Global Diversity of Sponges
0 - r
20 —
4 0 -
ICO
6 0 —
8 0 —
100 -L .2 o u o o o o o cŒo(75 Ü o -j o
Figure 13. Dendrogram output for hierarchical clustering of 12 M arine Realms. The m e th o d u sed is g ro u p -av e ra g e linking o f Bray-Curtis sim ilarities c a lcu la ted on p re s e n c e /a b se n c e s p o n g e sp ec ie s d a ta . Four a sse m b la g e ty p e s a re iden tified a t v a rious levels o f sim ilarity. do i:10 .1371/jo u rn a l.p o n e .0035105 .g013
Figure 14. Species richness of regional sponge faunas in western, northern and eastern Australia. Red circles in d ica te 'h o ts p o ts ' o f h igh sp ec ie s richness (m odified from [127]). do i:10 .1371/jo u rn a l.pone .00 3 5 1 0 5 .g 0 1 4
PLoS ONE I w ww.plosone.org 17 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
to sponges were developed o f the British Isles [143], N orth East A dantic [144], C aribbean [145], M edite rranean [146], New C aledonia [147], Indonesia [148], South Africa [149-150], M ariana Islands [151], N ew Z ealand [152-153], N o rth East Pacific [154], California [155], G ulf o f M exico [156], Florida [157], an d B ahia Brazil [158], to nam e ju st a very few. T he products from these various initiatives include basic species inventories, bu t often also extensive databases, websites, CDs and interactive keys. These were derived from an escalated collecting effort over the past two decades driven m ainly by a relatively small num ber o f factors. O n e of these m ajor factors has been the need to know m ore abou t the regional inventories o f sponge faunas based on the econom ic potential o f sponges for their bioactive com pounds as new pharm aceutical products. Biodiscovery for sponges th roughout the w estern Pacific in general, e.g. [159-160], an d in A ustralia in particular, e.g [161-162], has p roduced a surge o f new sponge collections in the m agnitude o f several hundreds o f thousands o f specimens. A nother im portan t factor th a t has accelerated sponge collections is the increasing responsibilities o f governm ents under various agreem ents within the CBD (h ttp ://w w w .cb d .in t/co n v en tio n ) to curb environm ental degradation, p ro tect native genetic resources, an d im prove food security such th a t m arine jurisdictions are increasingly “ ground tru th ing” their seabed for bioregional planning, hab ita t assessment and conservation purposes. A n exam ple o f sponge species distributions used in these regional environm ental assessments is the rezoning o f the G reat B arrier R eef M arine Park [163] that included approxim ately 1,200 sponge O T U ’s o f w hich even two of the five m ost prevalent species were new to science, e.g. [164],
T o cope with the vast num bers o f collected ‘unknow ns’ online tools are now available to help fast-track the description and illustration o f know n an d potential new species, including au tom ated taxonom ic keys and o ther initiatives to im prove diagnostic capabilities across a range o f biota. These tools include E D IT ’s Scratchpads (w w w.e-taxonom y.eu), E O L ’s LifeDesks (www.lifedesks.org), an d the A L A -E O L -C B IT (Centre for Biological Inform ation T echnology [ww w .cbit.uq.edu.au] partnership o f the IdentifyLife.initiative [www.identifylife.org], am ongst others). T h e Porifera LifeDesks project (porifera.lifedesks.org), currendy contains only a round 200 species, m ostly C aribbean, b u t is a working com ponent o f the Porifera T ree o f Life (PorToL, www. portol.org) initiative under developm ent, and is contributing to the global Assembling the T ree o f Life project (US N SF funded). In the A ustralian context the Atlas o f Living A ustralia (ALA, www. ala.org.au) is the m ost recent developm ent as biodiversity eR esearch infrastructure. A further tool in developm ent focused on collaborative work on raw sponge taxonom ic da ta is SpongeM aps wiki (w ik i.trin .o rg .au /b in /v iew au th /M arine /S pong- es), a new tool th a t has been developed from the T R IN wiki (wiki.trin.org.au).
In vasive sp e c ie s . As in m any o ther m arine groups, there are several cases o f sponge species know n or suspected to have crossed oceanic o r terrestrial barriers an d showing disjunct distributions. From the 1950s onw ard, E uropean Halichondria species, especially H . bowerbanki, have been reported as in troduced species in the San Francisco Bay area (available: h ttp ://re search arch iv e.ca lacad em y . o rg /resea rch /izg /S F B ay 2 K /H alich o n d ria % 2 Obowerbanki.htm). D ue to the variability o f these sponges an d the paucity o f distinctive m orphological m arkers the assertion o f these being alien species rem ains inconclusive. An extensively studied case is the Indonesian sponge Mycale (Mycale) ‘armata’ (identification questioned), w hich was identified as a potential th rea t to coral reefs o f H aw aii (available: h ttp ://hbs .b ish o p m u seu m .o rg /in v asiv es/reports/m ycale.h tm l), following a 1996 invasion o f Pearl H arbor.
Four o ther species w ere identified as ‘unintentionally in troduced’ in H aw aiian waters (h ttp ://w w w 2 .b ish o p m u seu m .o rg /H B S / invertg u id e /sponges.htm), the W est Ind ian species Haliclona caerulea an d Suberites zeteki, Philippine Gelliodes fibrosa, and Indo- M alayan Mycale parishi. T hese species are no t very well know n nor do they seem to be reliably identified so we reserve judgem en t on the source origin. Nevertheless, the m onitoring da ta indicate their recent range extensions. M ore spectacular is the case o f Celtodoryx ciocalyptoides, originally described from the Sea o f Jap a n . T he species was discovered on the west coast o f France from 1996 onw ards [165] and described as a new genus an d species, Celtodoryx girardae, w ith unknow n origin. V ery shordy afterwards the sponge was also discovered in the O osterschelde estuary in the SW p a rt o f the N etherlands [166], w here it is now one of the m ore com m on and conspicuous sponges. Both studies expressed a likely connection w ith shellfish culture bu t w ere unable to provide evidence for this o ther than th a t the species was previously unknow n from their areas. H enkel & Janussen [167] discovered the likely source populations in the northw est Pacific, and provided convincing p ro o f o f the conspecificity o f the Asian and E uropean populations. D utch waters contain several o ther species no t know n from elsewhere in adjacent regions and suspected to have been in troduced by shellfish transports: Mycale (Carmia) micracanthoxea, Haliclona (Soestella) xena, an d Sycon scaldiense [166]. A possible recent introduction from Brazil to the M edite rranean of a calcareous sponge, Paraleucilla magna, was reported in [168].
Discussion
Global diversity patterns o f ‘know n’ m arine sponges very probably reflect sam pling bias similar to th a t w hich is shown for Ascidiacea [169]. T his m ay be partly explained (a) by our focus on the ‘know n’ sponges, i.e. fully described ‘accepted’ species, and the ‘know n’ distributions, i.e. vouchered records o f ‘know n’ species. T h e scientific literature contains m any regional o r local species lists with unsubstantiated records o f ‘know n’ species and undeterm ined species, and na tura l history m useum collections contain m any identified bu t unpublished specimens th a t are partly accessible th rough GBIF and O B IS (iobis.org /m apper, data.gbi-f.org, 2011-11-05). A lthough partly to be considered ‘know n’ we decided against using these da ta in view o f the m ixture o f reliable and unreliable identifications inevitably adhering to them . A further explanation for the assum ed bias is (b) the lack o f reliable identifications o f sponges from several o f the w orld’s m arine habitats, notably all sciophilous an d deep-sea habitats, and from several m arine regions such as the South East Pacific, the Indian subcontinent, the A rabian and Persian Gulf, tropical W est Africa, South East Asia and the Pacific islands. Deep-sea sponge biogeography is still anecdotal. Also, the neglect o r lack o f effort o f the study of m ajor taxa such as the C alcarea an d the m arine Haplosclerida, respectively 8 an d 12% o f the total num ber o f species, m ay have contributed to biased results. Clearly, there is a significant sponge diversity im pedim ent to overcom e.
For the next decades, a large am oun t o f sponge specimens and da ta aw ait treatm ent. M any of these sponges are already collected and m any m ore are p lanned to be collected in various regions for biodiscovery an d conservation purposes. W e have the tools available (e.g. the System a Porifera classification, the W orld Porifera D atabase catalogue, C IS tools, and rap id sequencing) to process these specimens and data, b u t there is a very significant lag betw een docum enting the specimens, defining these w ithin the L innaean systematics, an d m aking their distributions widely accessible - the differential being a gap betw een the “ adequately know n” , the “poorly know n” an d the “ unknow n” in the o rder o f
PLoS ONE I w ww.plosone.org 18 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
one or m ore m agnitudes. For exam ple, it is estim ated th a t o f the > 3 ,0 0 0 sponge species collected from N orth East Australia alone, a round 70% are thought to be new to science [170], o r canno t be reconciled with any “know n” , m ostíy ancient species concepts as no ted above. This also ignores the extra dim ension o f the quantities o f cryptic sibling species h iding am ongst alleged w idespread m orphospecies, e.g. [137], an d the tiny, encrusting, parasitic sponge com m unities th a t have barely been sam pled, and therefore contribute to a potentially even bigger “unknow n” . T o resolve this using global datasets a t the level o f realm s is a t present p robably unhelpful, in view o f the assum ed collection bias, especially w hen the presendy “ unknow n” (but collected) species are excluded, and w ithout corrections for factors like differential collecting effort and sponge taxonom ic research effort betw een the various regions. N ear-fu ture efforts m ight m ore productively focus on sm aller m ore m anageable regional case studies, w hereas the ultim ate goal o f a global sponge richness assessment is o f necessity a d istant perspective.
N otw ithstanding this, there is great optim ism th a t m olecular tools will bette r define the identities o f m any o f the “know n” taxa, and therefore also fast-track the assignm ent o f these vast “ unknow n” collections to a new or know n taxon, e.g. [171]), bu t m uch w ork rem ains (see Sponge B arcoding Project rem arks above).
T h e M E O W scheme o f ecoregions, provinces and realms clearly accom m odates only the R ecent species distributions an d is indeed essentially an ecological instrum ent. It should be com plem ented by a higher-taxa scheme o f regions, notably for groups o f species belonging to the same phylogenetic clade or for genera with unchallenged synapom orphies. Exam ples we genera ted here show circum tropical, b ipolar, and antitropical distribution patterns, w hich provide insights in the biogeographic history o f taxa and will docum ent faunal changes.
Sponges were initially collected during the halcyon days o f curiosity-driven around-the-w orld expeditions in the 1800s, and in the 1980s they becam e the focus o f the new drive to understand coral reef and tem perate m arine ecology and invertebrate interactions. Sponges have since escalated in prom inence due to their potential value as new sources o f pharm aceutical products, transform ing our perspective on, an d understand ing o f the biology and biodiversity o f these allegedly simple basal m etazoans. In conclusion, to ou r constant am azem ent, sponges have sustained a high diversity and variety o f forms over the entire Phanerozoic Eon, and we continue to find new unpreceden ted species. W e can only im agine the limits o f this intriguing group of invertebrates.
Supporting InformationFile SI M a p s h o w in g n u m b e r s o f s p o n g e s p e c ie s a n d h ig h e r ta x a fo u n d in e a c h o f 12 M a r in e R e a lm s [30],extracted from the W orld Porifera D atabase (available: www. m arinespecies.org/porifera, accessed 2011 A ug 31). A. Species num bers, B. G enus num bers, C. Fam ily num bers.(TIF)
File S2 M a p s h o w in g n u m b e r s o f s p o n g e s p e c ie s a n d h ig h e r ta x a fo u n d in e a c h o f 62 M a r in e P r o v in c e s [30],extracted from the W orld Porifera D atabase (available: www. m arinespecies.org/porifera, accessed 2011 A ug 31). A. Species num bers, B. G enus num bers, C. Fam ily num bers.(TIF)
File S3 M a p s h o w in g n u m b e r s o f s p o n g e s p e c ie s a n d h ig h e r ta x a fo u n d in e a c h o f 232 M a r in e E c o r e g io n s [30],extracted from the W orld Porifera D atabase (available: www.
m arinespecies.org/porifera, accessed 2011 Aug 31). A. G enus num bers, B. Fam ily num bers (for Species num bers see Figure 11). (TIF)
File S4 M a p s h o w in g n u m b e r s o f s p e c ie s o f th e fo u r s p o n g e c la s s e s fo u n d in e a c h o f 12 M a r in e R e a lm s [30],extracted from the W orld Porifera D atabase (available: www. m arinespecies.org/porifera, accessed 2011 A ug 31). A. D em ospongiae, B. H exactinellida, C. H om osclerom orpha, D. C alcarea. (TIF)
File S5 M a p s h o w in g n u m b e r s o f s p e c ie s o f th e fo u r s p o n g e c la s s e s fo u n d in e a c h o f 232 M a r in e E c o r e g io n s[30] , ex tracted from the W orld Porifera D atabase (available: www. m arinespecies.org/porifera, accessed 2011 A ug 31). A. D em ospongiae, B. H exactinellida, C. H om osclerom orpha, D. C alcarea. (TIF)
File S6 D is tr ib u t io n p a t te r n s o f r e p r e se n ta t iv e g e n e r a r e c o r d e d in 232 M a r in e E c o r e g io n s [30], extracted from the W orld Porifera D atabase (available: w w w .m arinespecies.org/ porifera, accessed 2011 A ug 31). A. C osm opolitan distribution of Tedania; B. Bipolar distribution o f Iophon; C. C ircum tropic distribution o f Agelas; D. A ntitropical distribution o f Myxilla; E. R estricted tropical Indo-W est Pacific distribution o f Carteriospongia; F. D eep-sea distribution o f Asbestopluma (for an exam ple o f w arm - tem perate distribution see Fig. 12 showing the distribution o f the genus Spongia).(TIF)
File S7 D e n d r o g r a m o u tp u t fo r h ie r a r c h ic a l c lu s te r in g o f M a r in e E c o r e g io n s [30], using group-average linking of Bray-Curtis similarities calculated on p resence/absence sponge species data. O f the 232 provinces recognized by [30], those with less than 20 species recorded were om itted, resulting in 132 ecoregions analyzed.(TIF)
File S8 D e n d r o g r a m o u tp u t fo r h ie r a r c h ic a l c lu s te r in g o f M a r in e P r o v in c e s [30], using group-average linking o f Bray- Curtis similarities calculated on presence absence sponge species data. O f the 62 provinces recognized by [30], those with less than 50 species recorded were om itted, resulting in 44 provinces analyzed.(TIF)
AcknowledgmentsT hanks are due to: Elly B eglinger (N C B /Z M A ), L. de V os (ULB, Belgium), E. V oultsiadou (Thessaloniki, G reece), Filipe Porteiro (A zores/Im agD O P ), T . P é rez (M arseille, F rance), M .J. de K lu ijv e r (N C B /Z M A ), R . R oozendaa l (N etherlands), S.A. P om pon i (H arbo r B ranch O ceanograph ic Institute, Ft. P ierce, F lorida, U S A), H .M . R eisw ig (Royal British C o lum bia M useum ), a n d S. W alker (Q ueensland M useum ) fo r perm ission to use im ages. Special thanks are m ade for perm ission to use im ages o f carnivorous sponges: several specim ens illustrated w ere supplied by the N IW A Inverteb ra te C ollection (NIC), W ellington. T hese w ere collected as p a r t o f the N IW A research p rog ram m e Seamounts, their importance to fisheries and marine ecosystems. Several specim ens illustrated w ere collected from w ith in A ustra lian E E Z w aters a ro u n d M acquarie Island on the T A N 0803 voyage o f R V Tangaroa. A ustra lian E E Z specim ens curren tly w ith N IW A registra tion num bers will be accessioned b y the Q ueensland M useum once described. T h e M acR idge 2 voyage (TAN0803) rep resen ted a co llaboration betw een N ew Z ealand a n d A ustra lian scientists from N IW A and G eological N uc lea r Sciences Institu te (GNS) in N ew Z ealand , an d C S IR O D ivision o f M arine an d A tm ospheric R esearch , the A ntarctic C lim ate and Ecosystem s C ooperative R esearch C en tre (A C EC R C), an d the R esearch School o f E a rth Sciences, A ustra lian N a tio n a l U niversity (ANU) in A ustralia.
PLoS ONE I w ww.plosone.org 19 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
Author ContributionsA nalyzed the data: N d V N S BV. W ro te the paper: R vS N B E J V M D D EN d V N S BV M K J H . C onceived, con tribu ted in troduction , D em ospongiae, W P D , diversity, distributions, invasive species, conclusions: RvS.C o n trib u ted carn ivorous sponges: J V M K . C o n trib u ted H exactinellida:
References1. B ergqu is t P R (1978) Sponges. L o n d o n : H u tch in so n . 268 p.2. M ü lle r W E G (2003) S ponges (Porifera). Berlin: S pringer. 258 p.3. D e G oeij J M , V a n d e n B erg H , V a n O o s tv een M M , E p p in g E H G , V a n
D u y l F C (2008) M a jo r b u lk disso lved o rg an ic c a rb o n (D O C ) rem o v a l byen c ru s tin g c o ra l re e f cavity sponges. M a r E co l P ro g S er 357: 139—151.
4. V a ce le t J , B o u ry -E sn au lt N (1995) C a rn iv o ro u s sponges. N a tu re 373(6512): 3 3 3 -3 3 5 .
5. D e V o s L, R ü tz le r K , B o u ry -E sn au lt N , D o n a d e y G , V a c e le tJ (1991) A d as o f sponge m o rp h o lo g y . A d as d e m o rp h o lo g ie des éponges. W a sh in g to n & L on d o n : S m ith so n ian In stitu tio n Press. 117 p.
6. H e n tsch e l U , F ieseler L, W e h ri M , G e rn e r t G , S te in e ri M , e t al. (2003) M ic ro b ia l d iversity o f m arin e sponges. P ro g r M o l Subcell B iol 37: 59—88.
7. T a y lo r M W , R a d a x R , S teger D , W a g n er M (2007) S p onge-assoc ia ted m icroorgan ism s: evo lu tion , ecology a n d b io tech n o lo g ica l p o ten d a l. M icro b io l M o l Biol R ev 71: 2 9 5 -3 4 7 .
8. W estinga E , H o e tje s P (1981) T h e in tra sp o n g e fau n a o f Spheciospongia vesparia (Porifera , D em o sp o n g iae) a t C u ra ç ao a n d B onaire . M a r B iol 62: 139—150.
9. H o o p e r J N A , V a n Soest R W M , eds. System a P orifera: a gu ide to the c lassification o f Sponges. 2 volum es. N ew Y ork: K lu w er A c a d e m ic /P le n u m P ublishers. 1718 p.
10. V a n Soest R W M , B o u ry -E sn au lt N , H o o p e r J N A , R ü tz le r K , d e V o o g d N J, e t al. (2011) W o rld P o rife ra d a tab ase . A vailab le: h t tp : / /w w w .m a rin e sp ec ie s .o rg / p o rife ra . A ccessed 2011 A u g u st 31.
11. R ig b y J K , K aes le r R L (2003) T re a tise o n In v e r teb ra te Paleon to logy , P a r t E, P o rife ra (Revised) vol. 2: In tro d u c tio n to th e P orifera . L aw ren ce , K ansas: G eo log ical Society o f A m erica , xxvii+ 349 p.
12. S a rà M , V a c e le t J (1973) Écologie des D em o sp o n g es. In: PP . G rassé, ed. T ra ité d e Z oo log ie vol. 3. Spongiaires. Paris: M asso n e t C ie. p p 462—576.
13. Bell I I (2008) T h e fu n c tio n a l ro les o f m a r in e sponges. E s tu ar C o a s t S h e lf Sei 79(3): 3 4 1 -3 5 3 .
14. R eisw ig H M (1974) W a te r tra n sp o rt, resp ira tio n , a n d energetics o f th ree tro p ica l m arin e sponges. J E xp M a r B iol a n d E co l 14: 231—249.
15. R ü tz le r K (1975) T h e ro le o f b u rro w in g sponges in b io ero sio n . O eco lo g ia 19:2 0 3 -2 1 9 .
16. W ilk inson C R , F ay P (1979) N itro g e n fix a tio n in co ra l re e f sponges w ith sym bio tic c y an o b ac te ria . N a tu re 279: 52 7 —529.
17. W u lf f JL , Buss L W (1979) D o sponges h e lp h o ld co ra l reefs together? N a tu re 281: 4 7 4 M 7 5 .
18. W u lf f J L (1984) S p o n g e-m ed ia ted c o ra l-re e f g ro w th a n d reju v en a tio n . C o ra l R eefs 3: 15 7 -1 6 3 .
19. W ilk inson C R (1987) In te ro ce a n differences in size a n d n u tritio n o f co ra l ree f sponge pop u latio n s. Science 236: 1654—1657.
20. A erts L A M , V a n S o es t R W M (1997) Q u a n t if ic a t io n o f sp o n g e -c o ra l in te rac tio n s in a physically stressed re e f c o m m u n ity , N E C o lo m b ia . M a r E col P ro g r S er 148: 12 5 -1 3 4 .
21. H ill M S (1998) S p o n g iv o ry o n C a r ib b e a n reefs re leases co ra ls f ro m c o m p e titio n w ith sponges. O eco lo g ia 117: 143—150.
22. C o n w ay K W , K ra u tte r M , B arrie J V , N eu w eiler M (2001) H ex actin e llid sponge reefs o n th e C a n a d ia n co n tin e n ta l shelf: A u n iq u e “ L iv ing Fossil” . G eosci C a n a d a 28(2): 7 1 -7 8 .
23. D iaz M C , R ü tz le r K (2001) Sponges: a n essentia l c o m p o n e n t o f C a r ib b e a n co ra l reefs. Bull M a r Sei 69(2): 5 3 5 -5 4 6 .
24. V a n Soest R W M (2007) S ponge bid iversity . J M a r B iol Assoc U K 87: 13 4 5 -1 3 4 8 .
25. H o g g M M , T e n d a l O S , C o n w ay K W , P o m p o n i SA, V a n S oest R W M , e t al. (2010) D eep -sea sponge g rounds: reservo irs o f b iodiversity . U N E P -W C M C : C am b rid g e , U K . A vailable: h t tp : //w w w .u n e p -w c m c .o rg /m e d ia l ib ra ry /2 0 1 0 / 0 9 /0 7 /7 f0 3 c 5 ef /S p o n g e s_ B S 3 2 -R S 1 8 9 .p d f. A ccessed 2011 A u g u st 31.
26. P a w lik JR (2011) T h e ch em ical ecology o f sponges o n C a r ib b e a n reefs: n a tu ra l p ro d u cts shape n a tu ra l system s. Bio Sei 61(11): 88 8 —898.
27. V o u ltsiad o u E , D ailian is T , A n to n iad o u C , V afid is D , D o u n as, e t al. (2011) A eg ean b a th sponges: h isto rica l d a ta a n d c u rre n t status. R e v Fish Sei 19(1): 3 4 -5 1 .
28. G ra n t R E (1836) A n im al K in g d o m . In: T o d d R B , ed. T h e cy c lo p aed ia o f a n a to m y a n d physiology. V o lu m e 1. L on d o n : S h e rw o o d ,G ilb e rt, a n d P iper, p p 107-118 .
29. P o m p o n i SA (2006) Biology o f th e P orifera: cell cu ltu re . C a n J Z o o l 84: 167-174 .
30. S p a ld in g M , F o x H , A llen G , D a v id so n N , F e rd a ñ a Z , e t al. (2007) M arin e ecoreg ions o f th e w orld : A b io reg io n aliza tio n o f coasta l a n d sh e lf A reas. Bio Sei57: 5 7 3 -5 8 3 .
31. M a ld o n a d o M , B erg q u is t P R (2002) C h a p te r II: P h y lu m P orife ra . In: Y o u n g C M , ed. A das o f M a rin e In v e r teb ra te L arvae . L o n d o n : A cad em ic Press, p p 2 1 -5 0 .
M D . C o n trib u ted H om osclerom orpha an d C alcarea: N B E. C o n trib u ted new d evelopm en ts : D E . C o n tr ib u te d d is trib u tio n , m aps: N S BV. Perfo rm ed biodiversity analysis: N dV . C o n trib u ted reg ional a n d online resources, conclusions a n d perspectives: J H .
32. Ereskovsky A V (2010) T h e c o m p a ra tiv e em bryo logy o f sponges. L o n d o n , N ew Y ork , H eid e lb erg : S pringer. 329 p.
33. B o u ry -E sn au lt N (2006) S y stem a tic s a n d e v o lu tio n o f D e m o sp o n g ia e . C a n J Z o o l 84: 2 0 5 -2 2 4 .
34. V o u ltsiad o u E , V afid is D , A n to n iad o u C (2008) Sponges o f e co n o m ical in te res t in the E a s te rn M e d ite rra n ea n : a n assessm ent o f d iversity a n d p o p u la tio n density . J N a t H is t 42(5): 5 2 9 -5 4 3 .
35. P o m p o n i SA (1980) C y to log ica l m ech an ism s o f ca lc iu m c a rb o n a te ex cav atio n b y b o r in g sponges. I n t R e v C y to l 65: 301—319.
36. K elly M (2007) T h e m arin e fau n a o f N ew Z ealan d : Porife ra : lith istid D em o sp o n g iae (R ock Sponges). N IW A B iodiv M em 121: 1 -1 0 0 .
37. M c M u rra y SE, B lu m J E , P a w lik J R (2008) R ed w o o d o f the reef: g ro w th a n dage o f th e g ian t b a rre l spo n g e Xestospongia muta in th e F lo rid a K eys. M a r Biol155: 15 9 -1 7 1 .
38. P a w lik J R (2008) R ed w o o d s o f th e reef. S c ience 321: 19.39. H o o p e r J N A , V a n S oest R W M (2006) A n ew species o f Amphimedon (Porifera,
D em o sp o n g iae , H ap lo sc le rid a , N ip h a tid a e ) f ro m th e C ap ric o rn -B u n k w e r G ro u p o f Islands, G re a t B a rrie r R eef, A ustralia : ta rg e t species fo r th e ‘sponge g en o m e p ro je c t’. Z o o ta x a 1314: 31—39.
40. S rivastava M , S im akov O , C h a p m a n J , F ah ey B, G a u th ie r M E A , e t al. (2010) T h e Amphimedon queenslandica g en o m e a n d th e evo lu tio n o f a n im a l com plexity . N a tu re 466: 7 2 0 -7 2 7 .
41. V a c e le t J , B o u ry -E s n a u lt N , F ia la -M é d io n i A , F ish e r C R (1995) A m e th a n o tro p h ic c a rn iv o ro u s sponge. N a tu re 377(6547): 296.
42. V a c e le t J (2007) D iversity a n d evo lu tio n o f deep -sea carn iv o ro u s sponges. In: C u sto d io M R , L ô b o -H a jd u G , H a jd u E , M u ricy G , eds. P o rife ra research : b iodiversity , in n o v atio n a n d susta inability . S érie L ivros 28. R io de Ja n e iro : M u seu N ac io n al, p p 1 0 7 -1 1 5 .
43. V a ce le t J , D u p o r t É (2004) P rey c a p tu re a n d d igestion in th e carn iv o ro u s sponge Asbestopluma hypogea (Porifera: D em ospong iae). Z o o m o rp h o lo g y 123: 17 9 -1 9 0 .
44. R e isw ig H M , L e e W L (2007) A n e w species o f Cladorhiza (P orifera : C lad o rh iz id ae ) f ro m S C a lifo rn ia (U S A). In: C u sto d io M R , L ô b o -H a jd u G, H a jd u E , M u ric y G , eds. P o r ife ra re se a rch : b io d iv e rs ity , in n o v a tio n , susta inability . S érie L ivros 28. R io d e ja n e i ro : M u seu N ac io n al, p p 517—523.
45. W a d in g L (2007) P re d a tio n o n cop ep o d s b y a n A laskan c lad o rh iz id sponge. J M a r Biol Ass U K 87: 1 7 2 1 -1 7 2 6 .
46. V a ce le t J , K elly M , S ch lach e r-H o en lin g e r M (2009) T w o n ew species o f Chondrocladia (D em ospong iae: C lad o rh iz id ae ) w ith a n ew sp icule type fro m the deep so u th Pacific, a n d a d iscussion o f th e genus Meliiderma. Z o o ta x a 2073:5 7 -6 8 .
47. K elly M , V a c e le t J (2011) T h re e n e w rem a rk a b le c a rn iv o ro u s sponges (Porifera , C lad o rh iz id ae ) f ro m d eep N ew Z e a la n d a n d A u s tra lian (M acq u arie Island) w aters. Z o o ta x a 2976: 5 5 -6 8 .
48. V a c e le t J (2006) N e w carn iv o ro u s sponges (P orifera , P oecilosclerida) co llected f ro m m a n n e d subm ersib les in the d eep Pacific. Z o o lJ L in n Soc 148: 55 3 —584.
49. R ios P , K elly M , V a c e le tJ (2012) Cercicladia australis, a n ew c a rn iv o ro u s sponge w ith novel ch e lae f ro m th e T a s m a n B asin a n d the A d a n tic P a tag o n ia n M arg in (Porifera , C lad o rh iz id ae). Z o o ta x a 3131: 5 2 -6 2 .
50. V a c e le tJ (2008) A n ew genus o f carn iv o ro u s sponges (Porifera: Poecilosclerida , C lad o rh iz id ae ) f ro m th e d eep N -E Pacific, a n d rem ark s o n th e genus Neocladia. Z o o ta x a 1752: 5 7 -6 5 .
51. H a jd u E , V a c e le t J (2002) F am ily C lad o rh iz id ae . In : H o o p e r J N A , v an Soest R W M , eds. System a Porifera: A gu ide to th e c lassification o f sponges. N ew Y ork: K lu w er A c a d e m ic /P le n u m P ublishers, p p 6 3 6 —641.
52. V a c e le tJ , B o u ry -E sn au lt N , H a rm e l in J G (1994) H e x a c d n e llid cave, a u n iq u e deep -sea h a b ita t in the scuba zone. D eep -S ea R es (I) 41: 965—973.
53. B ak ran -P etric io li T , V a c e le tJ , Z ib row ius H , Petric io li D , C h e v a ld o n n é P , e t al. (2007) N e w d a ta o n th e d istr ib u tio n o f th e ‘d eep -sea ’ sponges Asbestopluma hypogea a n d Oopsacas minuta in th e M e d ite rra n e a n Sea. M a r E co l 28: 10—23.
54. C o n w ay K W , B arrie J V , A u sd n W C , L u te rn a u e r J L (1991) H o lo c e n e sponge b io h erm s o n th e w estern C a n a d ia n co n tin e n ta l shelf. C o n t S h e lf R es 11: 7 7 1 -7 9 0 .
55. K ra u tte r M , C o n w ay K W , B arrie J V , N eu w eiler M (2001) D iscovery o f a ‘liv ing d in o sa u r’: globally u n iq u e m o d e rn hex actin ellid sponge reefs o ff B ritish C o lu m b ia , C a n a d a . F acies 44: 2 6 5 -2 8 2 .
56. C o o k SE, C o n w ay K W , B u rd B (2008) S tatus o f th e glass sponge reefs in the G eo rg ia B asin. M a r E n v iro n m R es 66: S 8 0 -S 8 6 .
57. Leys SP, M ack ie G O , R eisw ig H M (2007) T h e b io logy o f glass sponges. A dv M a r B iol 52: 1 -1 4 5 .
58. S u n d a r V C , Y ab lo n A D , G ra z u l J L , B an M , A izen b erg J (2003) F ib re-o p tica l fea tu re s o f a glass sponge. N a tu re 424: 8 9 9 —900.
PLoS ONE I w ww.plosone.org 20 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
59. A iz e n b e rg J , W eav er J C , T h a n a w a la M S , S u n d a r V C , M o rse D E , e t al. (2005) Skeleton o f Euplectella sp.: s tru c tu ra l h ie ra rc h y fro m th e n an o sca le to the m acrosca le . Science 309: 275—278.
60. R eisw ig H M (2002) C lass H ex ac tin e llid a S ch m id t, 1870. In : H o o p e r JN A , V a n Soest R W M , eds. System a P orifera . A gu ide to th e c lassification o f sponges. N ew Y ork: P lenu . p p 1 2 0 1 -1 2 0 2 .
61. D o h rm a n n M , C ollins A G , W ö rh e id e G (2009) N e w insights in to the phy lo g en y o f glass sponges {Porifera, H exactinellida): m o n o p h y ly o f Lyssacin o sid a a n d E u p lecte llin ae , a n d th e ph y lo g en etic p o sitio n o f E u re tid ae . M ol P hy l E vol 52: 2 5 7 -2 6 2 .
62. D o h rm a n n M , G öcke C , J a n u ss e n D , R e itn e r J , L ü te r C , e t al. (2011) S ystem atics a n d sp icule ev o lu tio n in d ic ty o n a l sponges {H exactinellida: S cep tru lo p h o ra ) w ith d escrip tio n o f tw o n ew species. Z o o l J L in n Soc 163: 10 0 3 -1 0 2 5 .
63. D o h rm a n n M , H a e n K M , L av ro v D V , W ö rh e id e G {2012) M o lecu lar phy lo g en y o f glass sponges {Porifera, H exactinellida): in creased ta x o n sam pling a n d in clusion o f th e m ito c h o n d ria l p ro te in -c o d in g gene, cy to ch ro m e oxidase su b u n it I. H y d ro b io l d o i.1 0 .1 0 0 7 /s l0 7 5 0 -0 1 1 -7 2 7 -z .
64. D o h rm a n n M , Ja n u ss e n D , R e itn e r J , C ollins A G , W ö rh e id e G {2008) P hy logeny a n d evo lu tion o f glass sponges {Porifera, H exac tine llida). Syst Biol 57: 3 8 8 -4 0 5 .
65. G azav e E , L a p éb ie P, Ereskovsky A V , V a c e le tJ , R e n a rd E , e t al. {2012) N o lo n g er D em ospong iae : H o m o sc le ro m o rp h a fo rm a i n o m in a tio n as a fo u rth class o f P orifera . H y d ro b io l D O I 1 0 .1 0 0 7 /s l0 7 5 0 -0 1 1 -0 8 4 2 -x .
66. Ereskovsky A V , Ivanisevic J , P é rez T {2009) O v erv iew o n th e H o m o sc le ro m o rp h a sponges d iversity in th e M e d ite rra n ea n . In: P ro ceed in g s o f th e 1st S ym posium o n th e co ra lligenous a n d o th e r ca lcareo u s b io -co n c re tio n s o f the M e d ite rra n e a n Sea, T a b a rk a , 1 5 -1 6 J a n u a r y 2009. T un is : C A R /A S P . p p 8 9 -9 5 .
67. Lévi C {1956) E tu d e des Halisarca de Roscoff. E m b ry o lo g ie e t systém atique des dém osponges. A rc h Z o o l exp gén 93: 1—184.
68. Lévi C {1957) O n to g e n y a n d system atics in sponges. Syst Z o o l 6: 174—183.69. Lévi C {1973) S y stém atiq u e d e la classe des D em o sp o n g ia ria {Dém osponges).
In: G rassé P P , ed. T ra ité de Z oologie. III. Spong iaires. Paris: M asso n & C ie . p p 5 7 7 -6 3 2 .
70. B orchiellin i C , C h o m b a rd C , M a n u e l M , A livon E , V a c e le t J , e t al. {2004) M o lecu lar ph y lo g en y o f D em o sp o n g iae : im plica tions fo r c lassification a n d scenarios o f c h a ra c te r evo lu tion . M o l P hy l E vol 32: 8 2 3 —837.
71. S p erlin g E A , P e te rso n K L , P isan i D {2009) P hy logenetic-signa l d issection o f n u c lea r h o u sek eep in g genes su p p o rts th e p a ra p h y ly o f sponges a n d the m o n o p h y ly o f E u m etazo a . M o l B iol E vol 26: 2 2 6 1 -2 2 7 4 .
72. P h ilippe H , D ere lle R , L o p ez P , P ick K , B orchiellin i C , B o u ry -E sn au lt N , e t al. {2009) P hy logenom ics revives tra d itio n a l view s o n d eep a n im a l rela tionsh ips. C u r r Biol 19: 1-7 .
73. S p erlin g EA , R o b in so n J M , P isan i D , P e te rso n K J {2010) W h e re ’s the glass? B iom arkers, m o lecu la r clocks, a n d m ic ro R N A s suggest a 2 0 0 -M y r m issing P re ca m b ria n fossil rec o rd o f siliceous sponge spicules. G eob io l 8: 24—36.
74. G azav e E , L ap éb ie P, R e n a rd E , V a c e le tJ , R o c h e r C , e t al. {2010) M o lecu lar phy lo g en y resto res the su p ra -g en e ric subd iv ision o f h o m o sc le ro m o rp h sponges {Porifera, H o m o sc le ro m o rp h a ). PL oS O N E 5: e l4 2 9 0 .
75. M u ricy G {2011) D ivers ity o f In d o -A u s tra lian Plakortis {D em ospongiae:Plakin idae), w ith d escrip tio n o f fo u r n ew species. T M a r B iol Ass U K 91:3 0 3 -3 1 9 .
76. P é rez T , Ivanisevic J , D u b o is M , T h o m a s O P , T o k in a D , e t al. {2011) Oscarella balibaloi, a n ew sponge species { H om osclerom orpha: P lak in idae) f ro m the W este rn M e d ite rra n e a n Sea: cyto logical d escrip tio n , rep ro d u c tiv e cycle a n d ecology. M a r E co l 32: 174—187.
77. B o u ry -E sn au lt N , So lé-C av a A M , T h o rp e J P {1992) G en e tic a n d cytological d ivergence b e tw een c o lo u r m o rp h s o f th e M e d ite rra n e a n sponge Oscarella lobularis S ch m id t {Porifera, D e m o sp o n g iae , O scarellidae). J N a t H is t 26: 2 7 1 -2 8 4 .
78. M u ricy G , S o lé-C ava A M , T h o rp e J P , B o u ry -E sn au lt N {1996) G en eticev idence fo r ex tensive c ryp tic sp écia tio n in th e su b tid a l spo n g e Plakina trilopha{Porifera: D em o sp o n g iae : H o m o sc le ro m o rp h a). M a r E co l P ro g r Ser 138: 181-187 .
79. B o u ry -E sn au lt N , M u ricy G , G allissian M F , V a c e le tJ {1995) S ponges w ith o u t skeleton: a n e w M e d ite rra n e a n g en u s o f H o m o sc le ro m o rp h a {Porifera, D em ospong iae). O p h e lia 43: 25—43.
80. M u ricy G , B o u ry -E sn au lt N , B ézac C , V a c e le tJ {1998) A tax o n o m ic rev ision o f th e M e d ite rra n e a n Plakina S chulze {Porifera, D e m o sp o n g iae , H o m o sc le ro m o rpha). Z oo l J L in n Soc 124: 169-203 .
81. B e rg q u is t P R , K e lly M {2004) T a x o n o m y o f so m e H a l is a rc id a a n d H o m o s c le ro p h o r id a { P o rife ra : D e m o s p o n g ia e ) f ro m th e In d o -P a c if ic . N Z J M a r F resh w R es 38: 5 1 -6 6 .
82. V ishnyakov A E , Ereskovsky A V {2009) B ac teria l sym bion ts as a n ad d itio n a l cyto logical m ark e r fo r iden tificatio n o f sponges w ith o u t a skeleton. M a r Biol 156: 1 6 2 5 -1 6 3 2 .
83. Ivanisevic J , T h o m a s O P , L ejeusne C , C h e v a ld o n n é P, P é rez T {2011) M etab o lic f in g e rp rin tin g as a n in d ic a to r o f biodiversity : to w ard s u n d e rs ta n d in g in ter-specific re la tio n sh ip s a m o n g H o m o sc le ro m o rp h a sponges. M etab o lo m ics7: 2 8 9 -3 0 4 .
84. M u ricy G , B o u ry -E sn au lt N , B ézac C , V a c e le t J {1996b) C y to log ica l ev idence for c ryp tic sp écia tio n in M e d ite rra n e a n Oscarella species {Porifera, H o m o sc le ro m o rp h a). G a n J Z oo l 74: 8 8 1 -8 9 6 .
. PLoS ONE I w w w .plosone.org 21
85. M u ricy G , P earse J S {2004) A n ew species o f Oscarella {D em ospongiae: Plak in idae) fro m C alifo rn ia . P ro c C a lif A c a d Sei 55: 5 9 8 -6 1 2 .
86. Ereskovsky A V {2010) A n ew species o f Oscarella {D em ospongiae: P lakin idae) f ro m th e W e ste rn Sea o f ja p a n . Z o o ta x a 1376: 37—51.
87. M a n u e l M , B orojevic R , B o u ry -E sn au lt N , V a c e le t J {2002) C lass C a lcarea B o w erb an k , 1864. In : H o o p e r J N A , V a n So est R W M , eds. S y stem a P o rife ra : a gu ide to th e c lassification o f sponges. N e w Y ork: K lu w e r A c a d e m ic /P le n u m P ublishers, p p 1103—1110.
88. M a n u e l M {2006) P hy logeny a n d ev o lu tion o f c a lcareo u s sponges. C a n J Z ool 84: 2 2 5 -2 4 1 .
89. V a c e le t J {2012) C lass C a lc isp o n g iae B lainville , 1830. In : T re a tise o f P a leon to logy o n line. K ansas: T h e U n iversity o f K an sas , P a leon to log ica l Institu te .
90. C á rd e n a s P , P é rez T , B o u ry -E sn au lt N {2012) Sponge system atics fac in g new challenges. A dv M a r B iol 61: 7 9 -2 0 9 .
91. K o ech lin N {1977) In sta lla tio n d ’u n e ép ifau n e à Spirographis spallanzani V iv ian i, Sycon ciliatum F ab ric iu s e t Ciona intestinalis {L.) d an s le p o r t d e p la isan ce de L é z a rd rieu x {Côtes d u N ord). C a h B iol M a r 18: 325—337.
92. R a p p H T , Ja n u ss e n D , T e n d a l O S {2011) C a lcareo u s sponges f ro m abyssal a n d b a th y a l d ep th s in th e W ed d ell Sea, A n ta rc tica . D eep -S ea R es {II) 58:5 8 -6 7 .
93. W ö rh e id e G , H o o p e r J N A {1999) C a lc a re a f ro m th e G re a t B a rrie r Reef. 1: C ry p tic C a lc in ea f ro m H e ro n Islan d a n d W is ta ri re e f { C aprico rne-B unker group). M em Q u e e n s l M u s 43: 8 5 9 -8 9 1 .
94. K la u ta u M , V a le n tin e C {2003) R ev is io n o f th e genus Clathrina {Porifera, C alcarea). Z o o l J L in n Soc 139: 1—62.
95. R a p p H T {2006) C a lc are o u s sponges o f th e g e n e ra Clathrina a n d Guancha {C alcinea, C a lc a re a , P orife ra ) o f N o rw ay {north -east A d an tic ) w ith the d escrip tio n o f five n ew species. Z o o l J L in n Soc 147: 331—365.
96. R ossi A L , R usso C A M , S o lé-C av a A M , R a p p H T , K la u ta u M {2011) P hy logenetic signal in th e ev o lu tion o f b o d y co lo u r a n d sp icule skele ton in ca lcareo u s sponges. Z o o l J L in n Soc 163: 1026—1034.
97. A d am s C L , M c ln e rn e y J O , K elly M {1999) In d ica tio n s o f re la tionsh ips b e tw een p o r ife ra n classes using fu ll-leng th 18S rR N A gene sequences. M em Q u e e n s l M u s 44: 33^44.
98. M a n u e l M , B orch iellin i C , A livon E , L e P a rco Y, V a c e le t J , e t al. {2003) Phy logeny a n d ev o lu tion o f ca lcareo u s sponges: M o n o p h y ly o f C a lc in ea a n d C a lc a ro n e a , H ig h level o f m o rp h o lo g ica l hom o p lasy , a n d th e p rim itiv e n a tu re o f ax ial sym m etry . Syst Biol 52: 311—333.
99. M a n u e l M , B orchiellin i C , A livon E , B o u ry -E sn au lt N {2004) M o lecu lar ph y lo g en y o f ca lcareo u s sponges using 18S rR N A a n d 28S rR N A sequences. Boli M u s 1st B iol U n iv G e n o v a 68: 4 4 9 -4 6 1 .
100. D o h rm a n n M , V o ig t O , E rp e n b ec k D , W ö rh e id e G {2006) N o n -m o n o p h y ly o f m o st supraspecific tax a o f c a lcareo u s sponges {Porifera, C a lcarea ) rev ea led by in creased tax o n sam p lin g a n d p a rtitio n e d B ayesian analysis o f rib o so m al D N A . M o l P hy l E vol 40: 8 3 0 -8 4 3 .
101. B id d er G P {1898) T h e skele ton a n d classification o f ca lcareo u s sponge. P ro c R o y Soc 64: 6 1 -7 6 .
102. M in c h in E A {1896) Suggestions fo r a n a tu ra l classification o f th e A scon idae. A n n M a g N a t H is t 18: 3 4 9 -3 6 2 .
103. H a r tm a n W D {1958) A re -ex a m in a tio n o f B id d e r’s c lassification o f the C a lcarea . Syst Z o o l 7: 5 5 -1 0 9 .
104. B orojevic R {1979) E vo lu tio n des éponges C a lcarea . In: L évi C , B oury- E sn a u lt N , eds. B iologie des Spong iaires. Paris: E d itions C N R S , p p 527—530.
105. E rp e n b ec k D , W ö rh e id e G {2007) O n th e m o lecu la r ph y lo g en y o f sponges {Porifera). Z o o tax a . p p 107-126 .
106. R e d m o n d N E , R a le ig h J , v a n Soest R W M , K elly M , T ra v e rs SAA, e t al. {2011) P hy logenetic rela tio n sh ip s o f m arin e H a p lo sc le rid a {Phylum P orifera) em p lo y ing rib o so m al {28S rR N A ) a n d m ito c h o n d ria l (coxi y nadl) g en e seq u en ce d a ta . P L oS O N E 6 (9) e24344: 1-10 .
107. L avrov D , W a n g X , K elly M {2008) R eco n s tru c tin g o rd in a l rela tio n sh ip s in the D em o sp o n g iae using m ito c h o n d ria l g en o m ic d a ta . M o l P hy l E vol 49: 111—124.
108. W ö rh e id e G {2008) A hyperca lcified sp onge w ith soft relatives: Vaceletia is a ke ra to se dem o sp o n g e . M o l P hy l E vol 47: 4 33^438 .
109. M o rro w C C , P ic to n B E , E rp e n b ec k D , B o u ry -E sn au lt N , M aggs C A , e t al. {2012) C o n g ru e n c e b e tw een n u c lea r a n d m ito c h o n d ria l genes in D e m o sp o n giae: a n e w hypo thesis for re la tio n sh ip s w ith in th e G 4 c lade {Porifera: D em ospong iae). M o l P hy l E vol 62: 1 7 4 -1 9 0 .
110. H e b e r t P D N , C y w in sk a A , B a ll SL , d e W a a r d J R {2003) B io lo g ica l id en tifications th ro u g h D N A b a rco d es . P ro c R o y Soc L o n d o n {B) 270: 3 1 3 -3 3 2 .
111. W ö rh e id e G , E rp e n b ec k D {2007) D N A tax o n o m y o f sponges - p ro g ress a n d perspectives. J M a r Biol Ass U K 87: 1629-1 6 3 3 .
112. A p p e ltan s W , B o u ch e t P , Boxshall G A , F a u c h a ld K , G o rd o n D P , e t al. {2011) W o rld R eg is te r o f M a rin e Species. A vailable: h t tp : //w w w .m a rin e sp e c ie s .o rg . A ccessed 2011 A u g u st 31.
113. K la u ta u M , R usso C A M , Lazoski C , B o u ry -E sn au lt N , T h o rp e J P , Solé- C a v a A M {1999) D o es c o sm o p o lita n ism re su lt f ro m o v e rc o n se rv a tiv e system atics? A case study usin g th e m arin e sponge Chondrilla nucula. E vo lu tio n 53{5): 1414—1422.
114. B lan q u er A, U riz M J {2008) ‘A posteriori ’ search in g fo r p h e n o ty p ic c h a ra c te rs to describe n ew cryp tic species o f sponges rev ea led b y m o lecu la r m ark ers {D ictyonellidae : Scopalina). In v e r t Syst 22: 4 8 9 -5 0 2 .
April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
115. X a v ie r J R , R a c h e llo -D o lm e n P G , P a r ra -V e la n d ia F , S c h ö n b e rg C H L , B reeuw er J A J , e t al. (2010a) M o lecu lar ev idence o f c ryp tic sp écia tio n in the “co sm o p o litan ” ex cav a tin g sponge Cliona celata (P orifera , C liona idae). M o l P hyl E vol 56: 13 -2 0 .
116. R e v e illa u d J , R e m e rie T , V a n Soest R , E rp e n b ec k D , C á rd e n as P, e t al. (2010) Species b o u n d a rie s a n d p h y lo g en e tic re la tio n sh ip s b e tw een A d an to -M ed ite r- ra n e a n shallow -w ater a n d deep -sea co ra l a ssoc ia ted Hexadella species (Porifera , Ian thellidae). M o l P hy l E vol 56: 10 4 -1 1 4 .
117. A lvarez B, B ergqu is t P R , B attersh ill C N (2002) T a x o n o m ic rev ision o f the genus Latrunculia D u B ocage (Porifera: D em o sp o n g iae : L a tru n cu liid ae) in N ew Z ea lan d . N Z J M a r F resh w Res.
118. M a n co n i R , P ro n z a to R (2008) G lo b a l d iv ers ity o f sp onges (Porifera: Spongillina) in fresh w ater. H y d ro b io l 595: 27—33.
119. V a n S o est R W M (1994) D e m o sp o n g e d is t r ib u tio n p a tte rn s . In : V a n Soest R W M , V a n K e m p e n T M G , B ra e k m a n J C , eds. S ponges in T im e a n d Space. R o tte rd a m : B alkem a. p p 213—223.
120. T a b a c h n ic k K R (1994) D is tr ib u tio n o f R e c e n t H ex ac tin e llid a . In : V a n Soest R W M , V a n K e m p e n T M G , B ra e k m a n J C , eds. S ponges in T im e a n d Space. R o tte rd a m : B alkem a. p p 2 2 5 -2 3 2 .
121. M ald o n a d o M , U riz M J (1995) B iotic affinities in a tran s itio n a l zo n e b e tw een th e A d a n tic a n d the M ed ite rra n ea n : a b io g eo g rap h ica l a p p ro a c h b a sed o n sponges. J B iogeogr 22(1): 8 9 -1 1 0 .
122. C a rb a llo J L , N a ra n jo S, G a rc ia -G o m e z J C (1997) W h e re d o e s th e M e d ite rra n e a n Sea beg in? Z o o g eo g rap h ica l affinities o f th e litto ra l sponges o f th e S traits o f G ib ra lta r . J B iogeogr 24(2): 2 2 3 -2 3 2 .
123. B o u ry -E sn au lt N , P an sin i M , U riz M J (1994) S pong ia ires b a th y a u x d e la m er d ’A lb o ran e t d u golfe ib éro -m a ro ca in . M é m M u s n a tio n H is t n a t 160: 1—174.
124. S a rà M , B a ld u z z i A , B a rb ie r i M , B av estre llo G , B u rlan d o B (1992) B iogeograph ic tra its a n d checklist o f A n ta rc tic d em osponges. P o l B iol 12(6— 7): 5 5 9 -5 8 5 .
125. M cC lin to ck J B , A m sler C D A , B aker BJ, V a n Soest R W M (2005) E cology o f A n ta rc tic m ar in e sponges: a n overview . In te g r C o m p Biol 45: 359—368.
126. X a v ie r J R , V a n Soest R W M (2012) D iversity p a tte rn s a n d z o o g eo g rap h y o f the N o rth e a s t A d a n tic a n d M e d ite rra n e a n sha llow -w ater sp onge fau n a . H y d ro b io l doi: 1 0 .1 0 0 7 /s l0 7 5 0 -0 1 1-0880-4.
127. H o o p e r J N A , K e n n e d y J , Q u in n B J (2002) B iodiversity ‘h o tsp o ts ’, p a tte rn s o f richness a n d e n d em ism , a n d tax o n o m ic affinities o f tro p ica l A u s tra lian sponges (Porifera). B iodiv C o n s 11: 8 5 1 -8 8 5 .
128. S am aa i T , G ib b o n s M J (2005) D em o sp o n g iae tax o n o m y a n d b io d iversity o f th e B enguela reg io n o n th e w est co ast o f S o u th A frica. A fr N a t H is t 1: 1—96.
129. V a n Soest (1993) Affinities o f th e D e m o sp o n g e fau n a o f th e C a p e V erd e Islands a n d tro p ica l W est A frica. C o u r S enckenb F o rsch In st 159: 205—219.
130. H o o p e r JN A , Lévi C (1994) B iog eo g rap h y o f In d o -w est Pacific sponges: M ic ro c io n id a e , R a s p a il i id a e , A x in e llid ae . In : V a n S o es t R W M , V a n K e m p e n T M G , B raek m an J C , eds. S ponges in tim e a n d space. R o tte rd a m : B alkem a. p p 1 9 1 -2 1 2 .
131. H a jd u E (1995) M a cro ev o lu tio n ary p a tte rn s w ith in th e D em o sp o n g e o rd e r Poecilosclerida . P hy logeny o f th e m arin e co sm o p o litan genus Mycale, a n d a n in te g ra te d a p p ro a c h to b io g eo g rap h y o f the seas. P h D T hesis U n iversity o f A m ste rd am . 173 p.
132. V a n S oest R W M , H a jd u E (1997) M a rin e a re a rela tio n sh ip s f ro m tw en ty sponge phylogenies. A c o m p ariso n o f m eth o d s a n d co d in g strateg ies. C ladistics 13: 1-20 .
133. H a jd u E (1998) T o w a rd a p a n b io g e o g ra p h y o f th e seas: spo n g e phylogen ies a n d g en e ra l tracks. In : W a ta n a b e Y, F u se tan i N , eds. S p o n g e sciences. M ultid isc ip lin ary perspectives. T okyo : S p rin g e r V erlag , p p 95—108.
134. H a jd u E , D esq u ey ro u x -F aú n d ez R (2008) A reassessm en t o f th e ph y lo g en y a n d b io g eo g rap h y o f R h a b d e re m ia T o p se n t, 1890 (R h a b d e re m iid a e , Poecilosclerid a, D em ospong iae). R ev Suisse Z oo l 115(2): 3 7 7 -3 9 5 .
135. X a v ie r J R , V a n S oest R W M , B reeuw er JA J , M a rtin s A M F , M e n k en SBJ (2011b) P hy lo g eo g rap h y , gene tic diversity a n d s tru c tu re o f the poecilosclerid sponge Phorbas fictitius a t o cean ic islands. C o n tr ib Z o o l 79(3): 119—129.
136. R e v e i l la u d J , V a n Soest R , D erycke S, P ic to n B, R ig a u x A , e t al. (2011) P hy logenetic rela tio n sh ip s a m o n g N E A tlan tic Plocamionida T o p se n t (1927) (Porifera , Poecilosclerida): U n d e r-es tim a te d diversity in re e f ecosystem s. PLoS O N E 6(2): e l6 5 3 3 p p l - 1 0 .
137. W ö rh e id e G , E p p L, M acis L (2008) D e e p genetic d ivergences a m o n g In d o - Pacific p o p u la tio n s o f th e co ra l re e f sponge Leucetta chagosensis (L eucettidae): fo u n d e r effects, v ica rian ce , o r b o th ? B M C E vol Biol 8: 1—4-6.
138. B u tle r A J, R ees T , Beesley P , B ax N J (2010) M a rin e B iodiversity in the A u s tra lia n R eg io n . P L oS O N E 5(8): e l 1831. d o i:1 0 .1 3 7 1 /jo u rn a l .p o n e . 0011831.
139. H o o p e r J N A , W ie d e n m ay e r F (1994) P o rife ra . In : W ells A , ed. Z oological C a ta lo g u e o f A u stra lia . V ol. 12. M e lb o u rn e : C S IR O A u stra lia . 621 p. (u p d a ted o n line v ersion 200 4 availab le: h t tp : //w w w .e n v iro n m e n t.g o v .a u / b io d iv e rs i ty /a b rs /o n lin e -re s o u rc e s /fa u n a /a fd /g ro u p s . A ccessed 2012 A u g 31.
140. A B R S (2011) A u s tra lian F a u n a l D irec to ry . P h y lu m P o rife ra G ra n t, 1836. A u stra lian B io logical R eso u rces S tudy , C a n b e r ra . A vailable: h t tp : / /w w w . e n v i r o n m e n t .g o v .a u /b io d iv e r s i ty /a b r s / o n l i n e - r e s o u r c e s / f a u n a /a f d / t a x a / P orifera . A ccessed 2011 N o v 4.
141. P itch e r G R , D o h e r ty P, A rn o ld P , H o o p e r J , G rib b le N , e t al. (2007) S eab ed b iod iversity o n th e co n tin e n ta l sh e lf o f th e G re a t B a rrie r R e e f W o rld H eritag e A rea. A I M S /C S I R O /Q M /Q D P I C R C R e e f R e se a rc h T ask F in a l R e p o rt. IS B N 9 7 8 1 9 2 1 2 3 2 8 7 9 (pbk.) IS B N 9 7 8 1 9 2 1 2 3 2 8 8 6 (web) H o b a r t, C leveland :
C S IR O M a rin e & A tm o sp h eric R esea rch , p p 1—315. A vailab le: h t tp : / /w w w . r e e f .c r c .o r g .a u / r e s p r o g r a m /p r o g r a m C /s e a b e d / fm a l-re p o r t .h tm . A ccessed 2012 A ug 31.
142. F ro m o n t J , H ass C , M a rsh L , M o o re G , S alo tti M , e t al. (2006) B iodiversity o f m arin e fau n a o n th e C e n tra l W est C oast. F in a l M ilestone R e p o r t to the S tra teg ic R e se a rc h F u n d for th e M a rin e E n v iro n m e n t (S R F M E ). P erth : W e ste rn A u s tra lian M u seu m . 85 p.
143. P ic to n B E , H o w so n C M , eds. T h e species d irec to ry o f th e m arin e fau n a a n d flora o f th e B ritish Isles a n d su rro u n d in g seas. Belfast a n d R oss-on-W ye: U ls te r M u seu m a n d T h e M a rin e C o n se rv a tio n Society. C D -R O M E dition .
144. V a n S oest R W M , P ic to n B E , M o rro w GG (2000) S ponges o f th e N o r th E ast A d an tic . A m ste rd am : E T I, S p rin g e r-V erlag . IS B N 3 -5 4 0-14774-8 (W indow s C D -R O M ). A vailable: h ttp : / /s p e c ie s -id e n tf f ic a tio n .o rg /species.php?species_ g ro u p —sp o n g es& m en u en try —inleid ing . A ccessed 2012 A u g 31.
145. Z e a S, H en k e l T P , P a w lik J R (2009) T h e S p o n g e G uide: a p ic tu re gu ide to C a r ib b e a n sponges. A vailable: w w w .sp o n g eg u id e .o rg . A ccessed 2011 N o v 4.
146. P an sin i M , L ongo C (2003) A rev iew o f th e M e d ite rra n e a n Sea sponge b io g eo g rap h y w ith , in a p p e n d ix , a list o f th e dem o sp o n g es h ith e rto rec o rd e d f ro m this sea. B iogeogr 24: 59—90.
147. Lévi C , L ab o u te P, B a rg ib a n t G , M e n o u JL , B atte rsh ill C , e t al. (1998) Sponges o f the N ew C a le d o n ia n L agoon . E d itions de 1’O rs to m . C o llec tio n F a u n e e t flore trop icales P aris 33: 1—214.
148. V a n Soest R W M (1989) T h e In d o n es ia n spo n g e fau n a: A sta tus rep o rt. N e t h J Sea R es 23(2): 2 2 3 -2 3 0 .
149. S am aa i T (2006) B iodiversity “ h o tsp o ts” , p a tte rn s o f richness a n d en d em ism , a n d d istr ib u tio n o f m arin e sponges in S o u th A frica b a sed o n ac tu a l a n d in te rp o la tio n da ta : A c o m p a ra tiv e a p p ro a c h . Z o o ta x a 1358: 1—37.
150. S am aa i T , G ib b o n s M J (2006) D em o sp o n g iae tax o n o m y a n d b io d iversity o f the B enguela reg io n o n th e w est co ast o f S o u th A frica. A fr N a t H is t 1: 1—96.
151. K elly M , H o o p e r JN A , P a u l V , P au lay G , V a n Soest R W M , e t al. (2003) T a x o n o m ic in v en to ry o f th e sponges (Porifera) o f th e M a ria n a Islands. M ic ro n e s ic a 35—36: 100—120. A v a ilab le : h t t p : / / u n iv e r s i ty .u o g .e d u /u p / m ic ro n es ic a /a b s tra c ts_ 3 5 -3 6 /p d fs_ 3 5 3 6 /5 -sp o n g e s .p d f. A ccessed 2012 A ug31.
152. D aw so n E W (1993) T h e m arin e fau n a o f N ew Z ea lan d . In d e x to th e F au n a : 2. Porifera . N Z O c e a n o g r In st M e m 100: 1-98 .
153. K elly M , E d w ard s A R , W ilk inson M R , A lvarez B, D e C o o k SC , e t al. (2009) P h y lu m P o rife ra sponges. In: G o rd o n D P , ed. N ew Z e a la n d in v en to ry o f b io d iv e rsity 1. R a d ia ta , L o p h o tro c h o z o a , D e u te ro s to m ia . C h ris tc h u rc h : C a n te rb u ry U n iversity Press, p p 1—46.
154. A u stin B, O t t B, M c D a n ie l N , R o m a g o sa P (2007) S ponges o f th e co ld te m p e ra te N E Pacific. A vailab le: h t tp : / /w w w .m a re c o .o rg /K M L /P ro je c ts / N E sp o n g es_ co n ten t.a sp . A ccessed 2012 A u g u st 31.
155. L ee W L , E lv in D W , R eisw ig H M (2007) T h e sponges o f C alifo rn ia . A guide a n d key to th e m arin e sponges o f C alifo rn ia . M o n terey : M o n te re y Bay S a n c tu a ry F o u n d a tio n , C A , U S A. x+ 265 p .
156. R ü tz le r K , V a n S oest R W M , P ia n to n i G (2009) S ponges (Porifera) o f th e G u lf o f M exico . In: F e ld er D L , C a m p D K , eds. G u lf o f M exico — O rig in s , w aters, a n d b io ta . 1. B iodiversity. C o llege S ta tio n , T exas: T e x a s A & M Press, p p 2 8 5 -3 1 3 .
157. M essing G G , D iaz M G , K o h le r K , T a y lo r K H , R e e d J K , e t al. (2009) So u th F lo rid a sp o n g es. A g u id e to id e n tif ic a tio n . D a n ia B e a c h , FL : N o v a S o u th e a s te rn U n iv e rs i ty . A v a ila b le : h t t p : / / w w w .n o v a .e d u /n c r i / s o f l a _ sp o n g e _ g u id e /. A ccessed 2012 A u g u st 31.
158. H a jd u E , P e ix in h o S, F e rn a n d e z J C C (2011) E spon jas m arin h as d a B ahia: gu ia de c a m p o e lab o ra to r io . M u s N a c R io J a n Sér L ivr 45: 1—276.
159. C o lin P L (1998) M a rin e p h a rm a c eu tica ls f ro m th e reef: A v iew fro m th e field. In : C o ra l Reefs: challenges a n d o p p o rtu n itie s for susta in ab le m an a g e m en t, P ro c Assoc E v en t 5 th W o rld B ank C o n f E n v iro n m Social S ustain D evel. p p 7 6 -7 8 .
160. D eb itu s C (2005) D e v e lo p m e n t o f m arin e active sub tances - C o llec tio n a n d screen ing , co ra l re e f in itia tives for th e Pacific (C R ISP). C o m p o n e n t 2C: B io p ro sp e c tio n a n d m a r in e ac tiv e su sb stan ces . A v ailab le : h t tp : / /w w w . re e fb a se .o rg /p a c ff ic /d a tab ase . asp x ?sea rch d a ta= 2 & p ro jec tid = e0 0 0 0 0 0 0 0 3 0 .
161. C a p o n R (2001) M a rin e B io p ro sp ec tin g — T ra w lin g fo r tre a su re a n d p leasu re . E u r J O rg G h e m 2001(4): 6 3 3 -6 4 5 . doi: 10 .10 0 2 /1 0 9 9 -0 6 9 0 (2 0 0 1 0 2 )2 0 0 1 : 4 < 6 3 3 : :A ID -E J O C 6 3 3 > 3 .0 .C O ;2 -Q
162. Q u in n R J, D e A lm eid a L e o n e P , G u y m er G , H o o p e r J N A (2002) A u stra lian b iod iversity v ia its p lan ts a n d m arin e o rganism s. A h ig h -th ro u g h p u t sc reen ing a p p ro a c h to d ru g discovery. P u re A p p l C h e m 74(4): 51 9 —526.
163. G B R M P A (2011) G re a t B a r r ie r R e e f R e p re se n ta tiv e A reas P ro g ra m . A vailab le: h t t p : / /w w w .g b rm p a .g o v .a u /z o n in g -p e rm its -a n d -p la n s /ra p /d o c s / rep resen ta tiv e -areas-p ro g ram -p u b lica tio n s . A ccessed 2012 A u g 31.
164. Sutcliffe P R , H o o p e r JN A , P itc h e r G R (2010) T h e m o st co m m o n sponges o n the G re a t B a rrie r R e e f seab ed , A u stra lia , include species n ew to science (P hy lum Porifera). Z o o tax a 2616: 1—30. A vailable: h t tp : / /w w w .m a p re s s .c o m / z o o ta x a /2 0 1 0 /f /z t0 2 6 1 6 p 0 3 0 .p d f . A ccessed 2012 A u g u st 31.
165. P e rez T , P e rrin B, C a r te ro n S, V a c e le t J , B o u ry -E sn au lt N (2006) Celtodoryx girardae gen .nov . sp. nov ., a n ew sponge species (Poecilosclerida: D e m o sp o n giae) in v ad in g th e G u lf o f M o rb ih a n (N o rth E a s t A tlan tic , F rance). C a h Biol M a r 47: 2 0 5 -2 1 4 .
166. V a n S oest R W M , D e K lu ijver M J, V a n B rag t P H , F aasse M , N ijla n d R , e t al. (2007) S p o n g e in v ad ers in D u tc h coasta l w aters. J M a r Biol Ass U K 87(6): 1 7 3 3 -1 7 4 8 .
PLoS ONE I w ww.plosone.org 22 April 2012 I Volume 7 | Issue 4 | e35105

Global Diversity of Sponges
167. H en k e l D , J a n u ss e n D (2011) R e d esc rip tio n a n d n e w reco rd s o f Celtodoryx 170.ciocalyptoides {D em ospongiae: Poecilosclerida) - a sponge in v ad e r in the n o r theast A d a n d c O c e a n o f A sian o rig in ? J M a r B iol Ass U K 91(2): 3 4 7 -3 5 5 . 171.
168. L ongo C . M a stro to ta ro F , C o rr ie ro G (2007) O c c u rre n c e o f Paraleucilla magna(Porifera: C a lcarea) in th e M e d ite rra n e a n Sea. J M a r B iol Ass U K 87: 1 7 4 9 -1 7 5 5 . 172.
169. S h en k a r N , Sw alla BJ (2011) G lo b a l diversity o f A scid iacea. PL oS O N E 6(6): e20657 . doi: 10 .1 3 7 1 / jo u rn a l.p o n e .0020657 .
Q u in n B J, C a m p D (2007) B iodiscovery for n a tu ra l p ro d u c t p h a rm aceu tica ls : A n A u stra lian ex p erien ce . J B io law Business, A u s tra lia n E d itio n 2007: 39—43. P o p p e J , Sutcliffe P, H o o p e r J N A , W ö rh e id e G , E rp e n b ec k D (2009) G O I b a rc o d in g reveals n ew clades a n d rad ia tio n p a tte rn s o f Indo-P ac ific sponges o f the fam ily I rc in iid ae (D em ospong iae: D ic tyoceratida). PL oS O N E 5: e9950. Sars G O (1872) Spong iae. In :, , K o n g elig e N o rsk eU n iv e rs ite t (ed.) (1872) O n som e rem a rk a b le fo rm s o f a n im a l life f ro m th e g rea t d ep th s o ff th e N o rw eg ian coast. I. P a rd y fro m p o s th u m o u s m an u sc r ip tso f th e la te p ro fesso r D r. M ich ae l Sars. C h ris tia n ia , N o rw ay : B rogger & C h risd e . p p 62—82.
PLoS ONE I w w w .plosone.org 23 April 2012 | Volume 7 | Issue 4 | e35105