Embed Size (px)
Citation preview

Heredity 80 (1998) 732–741 Received 10 June 1997
Experimental evolution in Chlamydomonas.IV. Selection in environments that vary
through time at different scales
REES KASSEN* & GRAHAM BELLDepartment of Biology and Redpath Museum, McGill University, 1205 avenue Dr Penfield, Montreal, Quebec,
Canada H3A 1B1
Specialists and two kinds of generalist were selected for in a genetically heterogeneous basepopulation of the unicellular chlorophyte Chlamydomonas reinhardtii. The selection environ-ments consisted of alternating periods of light and dark. When the environment remainsconstant (light or dark), specialists are expected to evolve; when the environment variesthrough time, generalists are expected to evolve. The kind of generalist that evolves dependson the period of environmental variation: versatile generalists capable of reversible responsesto growth conditions are expected to evolve when the environment is fine-grained, whereasplastic generalists that respond irreversibly to the conditions of growth are expected to evolvewhen the environment is coarse-grained. The results indicate that specialists evolve in constantenvironments and generalists evolve in variable environments, as expected, but no evidencewas found to support the idea that versatility and plasticity evolve in fine-grained and coarse-grained environments respectively. Moreover, the evolved generalists performed well in everyenvironment and were insensitive to environmental variation. These results are interpreted tomean: (1) selection in the variable environments acted on the mean performance in eachenvironment, rather than on the variance in performance across environments; (2) there waslittle cost to being a generalist.
Keywords: Chlamydomonas reinhardtii, environmental grain, generalist, plasticity, specialist,versatility.
Introduction
The terms ‘specialist’ and ‘generalist’ describe twoends of a spectrum of adaptation: at one extremeare specialists, which tolerate a narrow range ofenvironmental conditions, whereas at the other aregeneralists, which tolerate a much broader range. Inthis paper, we present the results of an experimentdesigned to investigate how such differences in thebreadth of adaptation evolve. More specifically, ourexperiment addresses the following questions: (1)under what environmental conditions do specialistsand generalists evolve?; (2) How does the temporalscale of environmental variation affect the evolutionof generalists?; (3) What prevents the evolution of auniversally superior generalist?
When do specialists and generalists evolve?
The simplest theory of the evolution of specialistsand generalists is that specialists should evolve whenthe environment remains constant through time,whereas generalists should evolve when the environ-ment varies through time (see the general review byFutuyma & Moreno, 1988). Specialists evolve in atemporally constant environment because selectionfavours types whose fitness is highest in thatenvironment. Generalists, on the other hand, evolvein environments that vary through time becauseevery type is compelled to grow in several environ-ments, each of which may differ in the direction inwhich selection is operating; the type that evolves isone that has the highest fitness over all environ-ments.
Note that this theory does not require that gener-alists be ‘jacks-of-all-trades and masters of none’;there need not be a trade-off between ecological*Correspondence. E-mail: [email protected]
©1998 The Genetical Society of Great Britain.732

breadth and mean fitness (although this possibilityhas been raised by some authors; reviewed inFutuyma & Moreno, 1988; Sultan, 1992; see alsoVan Tienderen, 1991). Nor does the theory specifythe kinds of adaptations that will be favoured ineither situation, although these have been dealt within some theoretical treatments (for example Lloyd,1984; Moran, 1992; Padilla & Adolph, 1996). Thecentral issue is how fitness is maximized when theenvironment is more or less variable through time(see, for example, Lynch & Gabriel, 1987).
Scales of temporal variation and the evolution ofversatility and plasticity
Environmental change through time can occur on atleast two different scales relative to the lifetime ofan individual. As an illustration, consider Fig. 1. Onthe left-hand side are three generations of a lineageof vegetatively reproducing unicells. On the right-
hand side are three environments that alternatebetween two states (‘white’ and ‘black’) at threedifferent scales: the environment may never change(Fig. 1a), it may change every few generations (Fig.1b) or it may change repeatedly within the lifetimeof an individual (Fig. 1c). In Fig. 1a, only the whitestate is experienced in every generation, so selectionacts consistently in the same direction and thelineage should evolve to become specialized to thatstate; Fig. 1(b,c) experience both white and blackstates with equal frequency, so selection repeatedlychanges direction and this should lead to the evolu-tion of generalism. What distinguishes Fig. 1b fromFig. 1c is the time-scale on which change occurs; Fig.1b is called coarse-grained variation and Fig. 1c fine-grained variation, terms first coined by Levins(1968). Under coarse-grained variation (Fig. 1b),each individual experiences a single environmentalstate, although individuals living at different timesmay experience different states — the environmentis ‘patchy’ in time. Fig. 1c depicts fine-grained varia-tion, a situation in which every individual experi-ences both states, and individuals in subsequentgenerations experience the same series of states.
The dynamics of selection in fine-grained andcoarse-grained environments depends on how theshort-term fitness of an individual within a genera-tion is related to the long-term fitness of the lineageas a whole (Levins, 1968; Lloyd, 1984; Gomulkiewicz& Kirkpatrick, 1992). Assuming that individualsacquire resources additively within a patch, thenwhen the environment is fine-grained, the overallfitness of an individual is the arithmetic average ofits fitness in all the patches it experiences. Becauseeach generation experiences the same set of patches,the fitness of a lineage in the long-term is the sameas its average fitness among patches in the short-term. This is not true in coarse-grained environ-ments in which each generation does not necessarilyexperience the same type of patch. Here, any vari-ance in fitness among patches reduces the long-termgeometric mean fitness below that of the short-termarithmetic average fitness (for detailed discussions ofthe relationship between arithmetic and geometricmean fitness, see Gillespie, 1973; Bulmer, 1994; Bell,1997).
These different scales of variation can also influ-ence the kinds of adaptations that are favoured. Infine-grained environments, each generation experi-ences the same set of patches as every other genera-tion, so the fittest type is one that is versatile; itsphenotype should change appropriately as condi-tions change during its lifetime. Examples of versa-tile characters (also called labile characters by
Fig. 1 Environmental variation at different time-scalesrelative to the lifetime of an individual. The solid whitebar (a) represents an environment with one patch thatremains constant through time; (b) represents a coarse-grained environment in which the environment consists ofa series of patches that alternate once every few genera-tions; (c) represents an environment that is fine-grained,the patches alternating within the lifetime of an indivi-dual. (a) favours the evolution of specialists; (b) and (c)favour the evolution of versatile and plastic generalists,respectively.
SELECTION IN TEMPORALLY VARYING ENVIRONMENTS 733
© The Genetical Society of Great Britain, Heredity, 80, 732–741.

Scheiner, 1993) are inducible enzyme systems (e.g.Nguyen et al., 1989) and animal behaviour (Komers,1996). In a coarse-grained environment, the fittesttype is plastic; its phenotype is influenced by thestate of the environment but, once determined, doesnot change afterwards. Examples of plastic charac-ters are gender in species with environmental sexdetermination (Charnov, 1982) or alternativemorphologies in species with heteroeocious lifecycles (Moran, 1988).
The outcome of selection in variable environ-ments is therefore expected to be the evolution ofdifferent kinds of generalist, as a consequence of thescale at which the environment changes throughtime. This amounts to saying that the variance of theenvironment through time is itself an environmentalfactor independently of the states of the patchesconsidered separately; lineages may adapt toparticular patterns of environmental change ratherthan, or in addition to, adaptation to particularenvironmental states. There is no experimentalsupport for this conclusion as yet, although at leasttwo experiments have shown that generalism evolveswhen temporal change is coarse-grained (bacteria,Bennett et al., 1992; Chlamydomonas, Reboud &Bell, 1997).
Costs of adaptation prevent the evolution ofsuperior generalists
If versatility and plasticity are specific adaptations todifferent scales of environmental variation, they canbe interpreted in the same way as any other adapta-tion. This means that it should be possible to detecta cost of adaptation not only in specialists adaptingto particular patches in an environment, but also ingeneralists adapting to different patterns of change.
There are two sources of a cost of adaptation:antagonistic pleiotropy and mutation accumulation.Both lead to negative genetic correlations in fitnessacross environments, but for different reasons.Antagonistic pleiotropy occurs when genes thatimprove fitness in one environment are detrimentalin another. Mutation accumulation, on the otherhand, occurs because mutations that are neutral inone environment are likely to be detrimental inanother. Over time, selection will erode the antago-nistic effects of pleiotropic gene action, although thiswill be balanced by the continuing accumulation ofconditionally deleterious mutations. Which compo-nent makes the greater contribution to the overallcost of adaptation will depend on how geneticeffects are correlated and on how long selectionacts. Reboud & Bell (1997) have discussed which
components are responsible for the cost of adapta-tion in specialists and generalists. For now, it issufficient to note that the existence of a cost ofadaptation guarantees that a universally superiorspecialist or generalist should never evolve; adapta-tion will always be environment-specific, regardlessof whether the environment is constant or varyingthrough time.
Materials and methods
The general biology of Chlamydomonas reinhardtiiand descriptions of the laboratory methods used inthis experiment can be found in The Chlamydomo-nas Sourcebook (Harris, 1989) and in previouspapers describing experimental evolution in C. rein-hardtii (for example Reboud & Bell, 1997, and refer-ences therein).
Constructing the base population
The base population was a genetically hetero-geneous mixture consisting of the parents, F1, back-cross (F1Åparents) and F2 progeny of matingsbetween three mating type+ strains and threemating typeµ strains (CC1010+, CC1871+,F94-161+, CC410µ, CC1009µ and CC1952µ). Allare standard laboratory strains that have been inculture for many years except for F94-161, which wasisolated from a cornfield in Farnham, Quebec in1994.
Selection environments
The two different environmental states or patches inour experiment were light and dark. Chlamydomo-nas reinhardtii normally grows vegetatively as aphotoautotroph in the light but can also grow as aheterotroph in the dark when given acetate as acarbon source (Harris, 1989). A strong negativegenetic correlation between these two modes ofgrowth has been shown to evolve in clonal popula-tions after approximately 275 generations of selec-tion in one or the other environment (Bell &Reboud, 1997), which suggests that autotrophy andheterotrophy require different physiological compe-tences. We selected for specialists by maintainingpopulations in either a constant light environment(light specialists) or a constant dark environment(dark specialists). Versatile generalists were selectedin a fine-grained environment, which was con-structed by alternating light and dark every hour(about five alternations within every generation).
734 R. KASSEN & G. BELL
© The Genetical Society of Great Britain, Heredity, 80, 732–741.

Plastic generalists were selected in a coarse-grainedenvironment in which the light and dark conditionsalternated every 24 h (about once every five genera-tions). Each environment was located in a separategrowth chamber in the McGill University Phytotron,except for the two specialist lines, which shared thesame growth chamber. Dark conditions were createdby maintaining the cultures in an opaque box.
Maintaining the selection lines
The lines were maintained on agar plates containingBold’s medium with 1.2 g Lµ1 sodium acetate as anadditional carbon source. Every 3 or 4 days (every 7days in the case of the dark lines, which grow muchmore slowly than the others) approximately 1.5Å105
cells were transferred by washing the cells off thesurface of the agar and transferring an aliquot(250 mL) by pipette to a fresh plate. The experimentlasted for 10 weeks, representing roughly 400 gener-ations in constant light, 150–200 generations in thefine-grained and coarse-grained environments and80 generations in constant dark.
Three replicates of the base population were keptin the laboratory under dim illumination on platescontaining identical medium and wrapped in paraf-fin wax. The stored base population was transferredonce and underwent no more than eight generationsduring the course of the experiment.
Three replicate lines within each treatment (selec-tion environment) were arranged as a randomizedblock design within each environment chamber.Block effects were significant (F2,45 = 9.10,P = 0.0005), so all the analyses and data presentedin the paper have been adjusted for these effects.
Measuring the effects of selection
Assay procedure We grew samples from each selec-tion line and the base population in every environ-ment. The assay plates received an inoculum thesame size as those used in routine transfers. Theassay thus comprised (four selection lines+basepopulation)Å(four environments)Å(three repli-cates) = 60 plates. These were laid out in the growthchambers in the same manner as the selection lines.
Fitness We used the initial growth rate of coloniesas our measure of fitness, which we estimated bycalculating the mean number of divisions of 100colonies on a plate 24 h after it was inoculated. Thisprovides an estimate of rmax, the intrinsic rate ofgrowth in pure culture, which should be a good esti-
mate of fitness in competition as long as nutrientsremain unlimiting (see Bell, 1997, for a moredetailed discussion). Our selection lines were trans-ferred often enough to ensure that most lines weremaintained near their maximum rate of increasethroughout the experiment.
This procedure differed slightly for assaysconducted in the coarse-grained environment, inwhich one cycle of growth (five generations in thelight, five generations in the dark) comprised 24 h inthe light and then 24 h in the dark. In order toensure that separate colonies could be distinguished,lines assayed in the coarse-grained environmentnecessarily experienced less than one full growthcycle. As a result, not all lines assayed in the coarse-grained environment experienced the same photo-period, although all experienced at least one fullgeneration in either the light or the dark. Toaccount for these differences in photoperiod, weadjusted by regression the growth rates of linesassayed in the coarse-grained environment to a 12 hlight/12 h dark photoperiod.
Adaptation The difference in fitness between aselection line and the base population was calculatedas a measure of adaptation. A positive value indi-cates that a selection line grew faster than the basepopulation in a particular environment; a negativevalue indicates that it grew more slowly.
Results
The general response to selection
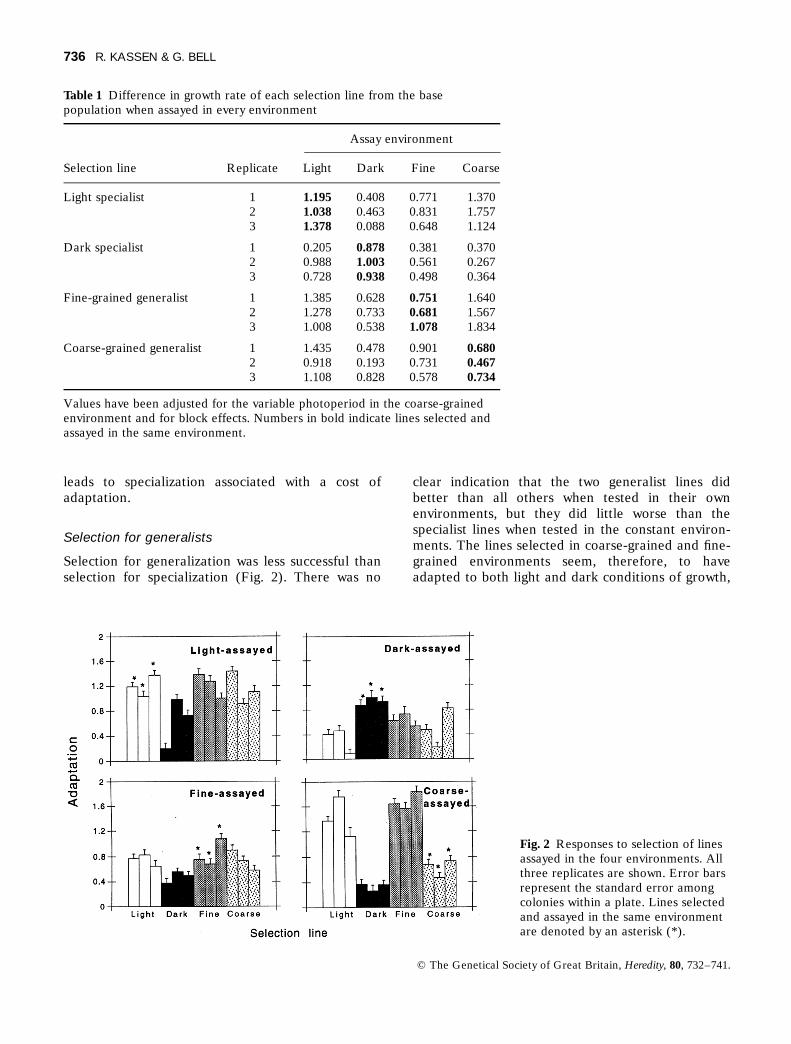
The results of selection are given in Table 1 andshown graphically in Fig. 2. The values represent thedifference in growth rate from the base population.All selection lines grew faster than the base popula-tion. There were significant differences in theresponse to selection among selection lines, amongassay environments and an interaction of the two,indicating that the lines diverged substantiallybecause of selection in different environments(Table 2). The synclinal response to selection in alllines suggests that part of the response observed wascaused by a general adaptation to the conditions ofthe growth chambers.
Selection for specialists
The line selected in the light grew better whentested in the light than in the dark; the reverse wastrue for the dark-selected line (Fig. 2). This supportsthe idea that selection in constant environments
SELECTION IN TEMPORALLY VARYING ENVIRONMENTS 735
© The Genetical Society of Great Britain, Heredity, 80, 732–741.
leads to specialization associated with a cost ofadaptation.
Selection for generalists
Selection for generalization was less successful thanselection for specialization (Fig. 2). There was no
clear indication that the two generalist lines didbetter than all others when tested in their ownenvironments, but they did little worse than thespecialist lines when tested in the constant environ-ments. The lines selected in coarse-grained and fine-grained environments seem, therefore, to haveadapted to both light and dark conditions of growth,
Table 1 Difference in growth rate of each selection line from the basepopulation when assayed in every environment
Assay environment
Selection line Replicate Light Dark Fine Coarse
Light specialist 1 1.195 0.408 0.771 1.3702 1.038 0.463 0.831 1.7573 1.378 0.088 0.648 1.124
Dark specialist 1 0.205 0.878 0.381 0.3702 0.988 1.003 0.561 0.2673 0.728 0.938 0.498 0.364
Fine-grained generalist 1 1.385 0.628 0.751 1.6402 1.278 0.733 0.681 1.5673 1.008 0.538 1.078 1.834
Coarse-grained generalist 1 1.435 0.478 0.901 0.6802 0.918 0.193 0.731 0.4673 1.108 0.828 0.578 0.734
Values have been adjusted for the variable photoperiod in the coarse-grainedenvironment and for block effects. Numbers in bold indicate lines selected andassayed in the same environment.
Fig. 2 Responses to selection of linesassayed in the four environments. Allthree replicates are shown. Error barsrepresent the standard error amongcolonies within a plate. Lines selectedand assayed in the same environmentare denoted by an asterisk (*).
736 R. KASSEN & G. BELL
© The Genetical Society of Great Britain, Heredity, 80, 732–741.
without evolving plasticity or versatility as a responseto specific patterns of environmental change.
The response to selection expressed as progress inthe light and dark
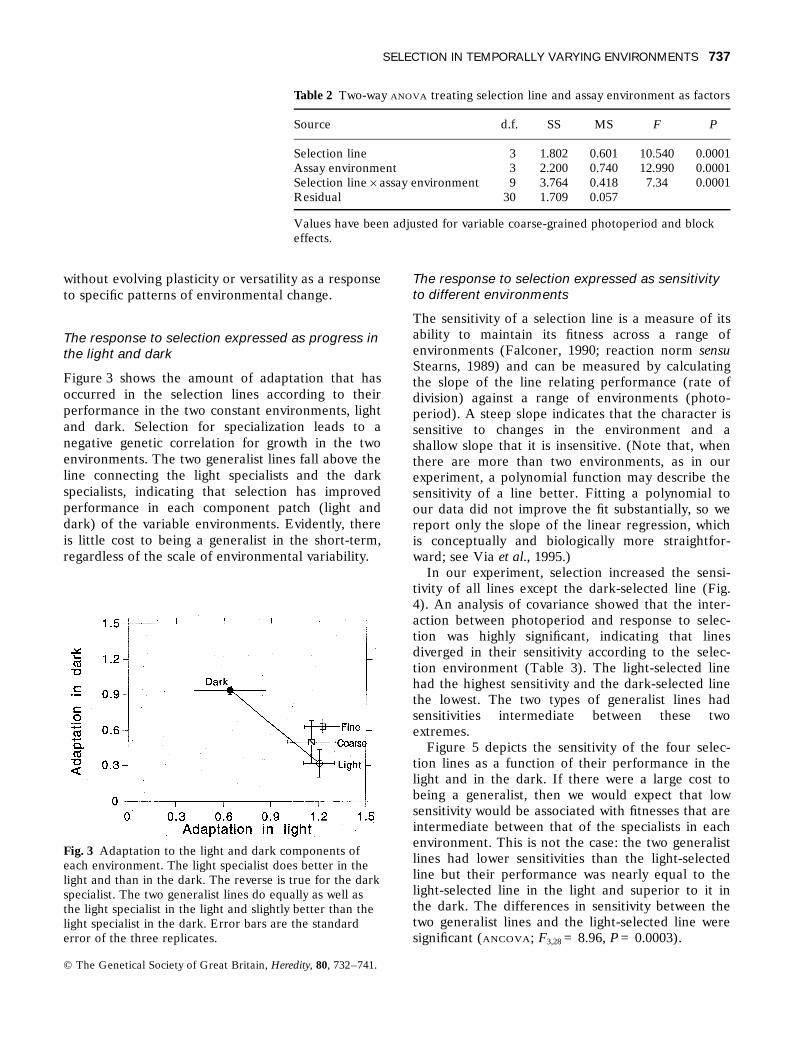
Figure 3 shows the amount of adaptation that hasoccurred in the selection lines according to theirperformance in the two constant environments, lightand dark. Selection for specialization leads to anegative genetic correlation for growth in the twoenvironments. The two generalist lines fall above theline connecting the light specialists and the darkspecialists, indicating that selection has improvedperformance in each component patch (light anddark) of the variable environments. Evidently, thereis little cost to being a generalist in the short-term,regardless of the scale of environmental variability.
The response to selection expressed as sensitivityto different environments
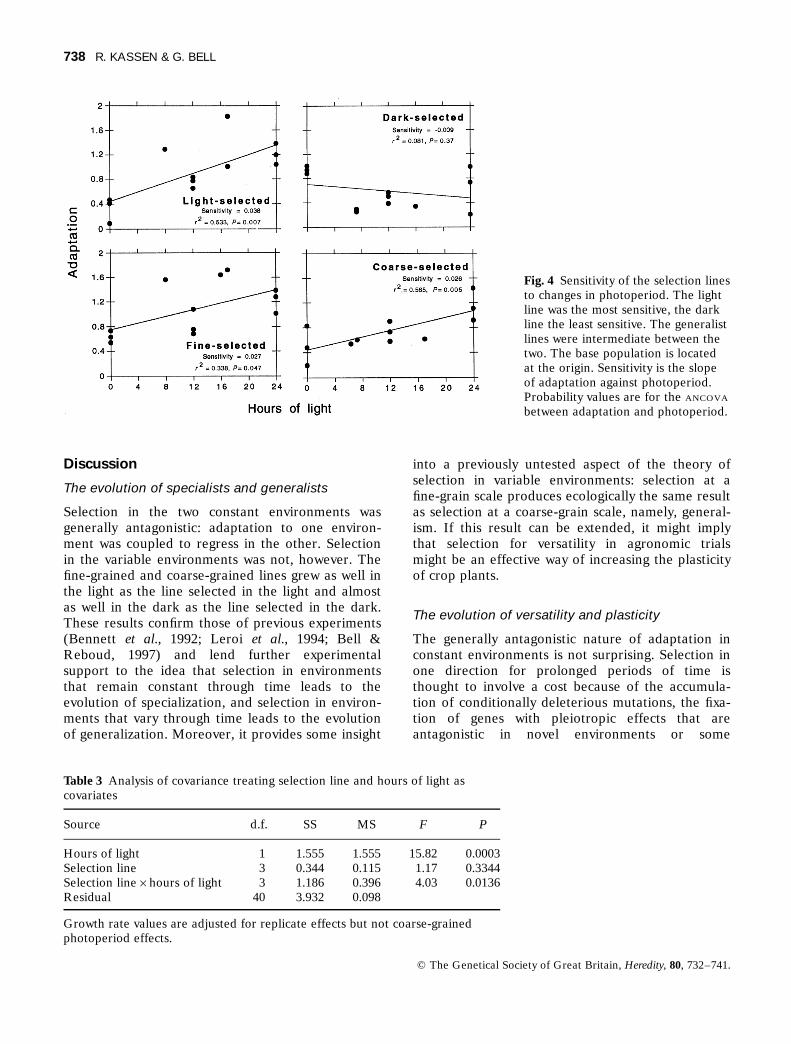
The sensitivity of a selection line is a measure of itsability to maintain its fitness across a range ofenvironments (Falconer, 1990; reaction norm sensuStearns, 1989) and can be measured by calculatingthe slope of the line relating performance (rate ofdivision) against a range of environments (photo-period). A steep slope indicates that the character issensitive to changes in the environment and ashallow slope that it is insensitive. (Note that, whenthere are more than two environments, as in ourexperiment, a polynomial function may describe thesensitivity of a line better. Fitting a polynomial toour data did not improve the fit substantially, so wereport only the slope of the linear regression, whichis conceptually and biologically more straightfor-ward; see Via et al., 1995.)
In our experiment, selection increased the sensi-tivity of all lines except the dark-selected line (Fig.4). An analysis of covariance showed that the inter-action between photoperiod and response to selec-tion was highly significant, indicating that linesdiverged in their sensitivity according to the selec-tion environment (Table 3). The light-selected linehad the highest sensitivity and the dark-selected linethe lowest. The two types of generalist lines hadsensitivities intermediate between these twoextremes.
Figure 5 depicts the sensitivity of the four selec-tion lines as a function of their performance in thelight and in the dark. If there were a large cost tobeing a generalist, then we would expect that lowsensitivity would be associated with fitnesses that areintermediate between that of the specialists in eachenvironment. This is not the case: the two generalistlines had lower sensitivities than the light-selectedline but their performance was nearly equal to thelight-selected line in the light and superior to it inthe dark. The differences in sensitivity between thetwo generalist lines and the light-selected line weresignificant (ANCOVA; F3,28 = 8.96, P = 0.0003).
Table 2 Two-way ANOVA treating selection line and assay environment as factors
Source d.f. SS MS F P
Selection line 3 1.802 0.601 10.540 0.0001Assay environment 3 2.200 0.740 12.990 0.0001Selection lineÅassay environment 9 3.764 0.418 7.34 0.0001Residual 30 1.709 0.057
Values have been adjusted for variable coarse-grained photoperiod and blockeffects.
Fig. 3 Adaptation to the light and dark components ofeach environment. The light specialist does better in thelight and than in the dark. The reverse is true for the darkspecialist. The two generalist lines do equally as well asthe light specialist in the light and slightly better than thelight specialist in the dark. Error bars are the standarderror of the three replicates.
SELECTION IN TEMPORALLY VARYING ENVIRONMENTS 737
© The Genetical Society of Great Britain, Heredity, 80, 732–741.
Discussion
The evolution of specialists and generalists
Selection in the two constant environments wasgenerally antagonistic: adaptation to one environ-ment was coupled to regress in the other. Selectionin the variable environments was not, however. Thefine-grained and coarse-grained lines grew as well inthe light as the line selected in the light and almostas well in the dark as the line selected in the dark.These results confirm those of previous experiments(Bennett et al., 1992; Leroi et al., 1994; Bell &Reboud, 1997) and lend further experimentalsupport to the idea that selection in environmentsthat remain constant through time leads to theevolution of specialization, and selection in environ-ments that vary through time leads to the evolutionof generalization. Moreover, it provides some insight
into a previously untested aspect of the theory ofselection in variable environments: selection at afine-grain scale produces ecologically the same resultas selection at a coarse-grain scale, namely, general-ism. If this result can be extended, it might implythat selection for versatility in agronomic trialsmight be an effective way of increasing the plasticityof crop plants.
The evolution of versatility and plasticity
The generally antagonistic nature of adaptation inconstant environments is not surprising. Selection inone direction for prolonged periods of time isthought to involve a cost because of the accumula-tion of conditionally deleterious mutations, the fixa-tion of genes with pleiotropic effects that areantagonistic in novel environments or some
Table 3 Analysis of covariance treating selection line and hours of light ascovariates
Source d.f. SS MS F P
Hours of light 1 1.555 1.555 15.82 0.0003Selection line 3 0.344 0.115 1.17 0.3344Selection lineÅhours of light 3 1.186 0.396 4.03 0.0136Residual 40 3.932 0.098
Growth rate values are adjusted for replicate effects but not coarse-grainedphotoperiod effects.
Fig. 4 Sensitivity of the selection linesto changes in photoperiod. The lightline was the most sensitive, the darkline the least sensitive. The generalistlines were intermediate between thetwo. The base population is locatedat the origin. Sensitivity is the slopeof adaptation against photoperiod.Probability values are for the ANCOVA
between adaptation and photoperiod.
738 R. KASSEN & G. BELL
© The Genetical Society of Great Britain, Heredity, 80, 732–741.
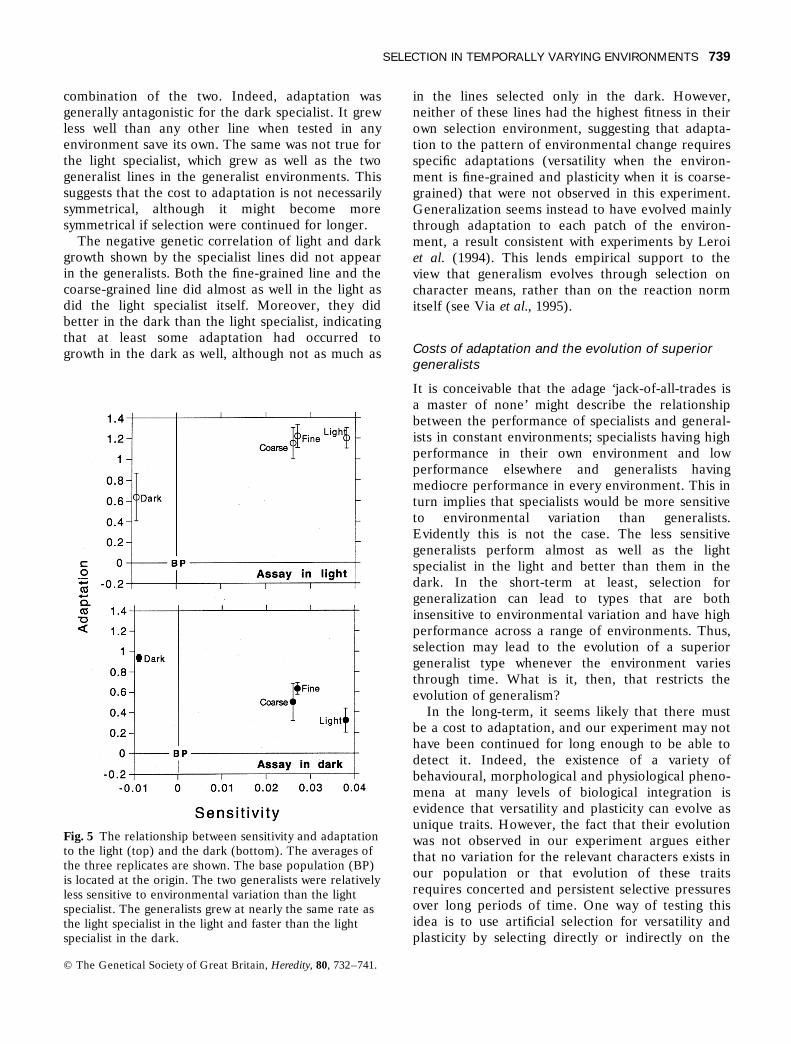
combination of the two. Indeed, adaptation wasgenerally antagonistic for the dark specialist. It grewless well than any other line when tested in anyenvironment save its own. The same was not true forthe light specialist, which grew as well as the twogeneralist lines in the generalist environments. Thissuggests that the cost to adaptation is not necessarilysymmetrical, although it might become moresymmetrical if selection were continued for longer.
The negative genetic correlation of light and darkgrowth shown by the specialist lines did not appearin the generalists. Both the fine-grained line and thecoarse-grained line did almost as well in the light asdid the light specialist itself. Moreover, they didbetter in the dark than the light specialist, indicatingthat at least some adaptation had occurred togrowth in the dark as well, although not as much as
in the lines selected only in the dark. However,neither of these lines had the highest fitness in theirown selection environment, suggesting that adapta-tion to the pattern of environmental change requiresspecific adaptations (versatility when the environ-ment is fine-grained and plasticity when it is coarse-grained) that were not observed in this experiment.Generalization seems instead to have evolved mainlythrough adaptation to each patch of the environ-ment, a result consistent with experiments by Leroiet al. (1994). This lends empirical support to theview that generalism evolves through selection oncharacter means, rather than on the reaction normitself (see Via et al., 1995).
Costs of adaptation and the evolution of superiorgeneralists
It is conceivable that the adage ‘jack-of-all-trades isa master of none’ might describe the relationshipbetween the performance of specialists and general-ists in constant environments; specialists having highperformance in their own environment and lowperformance elsewhere and generalists havingmediocre performance in every environment. This inturn implies that specialists would be more sensitiveto environmental variation than generalists.Evidently this is not the case. The less sensitivegeneralists perform almost as well as the lightspecialist in the light and better than them in thedark. In the short-term at least, selection forgeneralization can lead to types that are bothinsensitive to environmental variation and have highperformance across a range of environments. Thus,selection may lead to the evolution of a superiorgeneralist type whenever the environment variesthrough time. What is it, then, that restricts theevolution of generalism?
In the long-term, it seems likely that there mustbe a cost to adaptation, and our experiment may nothave been continued for long enough to be able todetect it. Indeed, the existence of a variety ofbehavioural, morphological and physiological pheno-mena at many levels of biological integration isevidence that versatility and plasticity can evolve asunique traits. However, the fact that their evolutionwas not observed in our experiment argues eitherthat no variation for the relevant characters exists inour population or that evolution of these traitsrequires concerted and persistent selective pressuresover long periods of time. One way of testing thisidea is to use artificial selection for versatility andplasticity by selecting directly or indirectly on the
Fig. 5 The relationship between sensitivity and adaptationto the light (top) and the dark (bottom). The averages ofthe three replicates are shown. The base population (BP)is located at the origin. The two generalists were relativelyless sensitive to environmental variation than the lightspecialist. The generalists grew at nearly the same rate asthe light specialist in the light and faster than the lightspecialist in the dark.
SELECTION IN TEMPORALLY VARYING ENVIRONMENTS 739
© The Genetical Society of Great Britain, Heredity, 80, 732–741.

environmental variance. We intend to set up experi-ments of this kind.
An alternative idea, put forward by Whitlock(1996), is that the appropriate trade-off is betweenthe rates at which specialists and generalists evolve.When the environment is constant, beneficial allelesgo to fixation faster and deleterious ones are lostfaster than when the environment is variable. Thus,specialists may evolve faster than generalists becausetheir environment is persistent, whereas the environ-ment of a generalist is not. Our results provide somecircumstantial support for this idea. The darkspecialist was superior to all other lines in the darkafter just 80 generations of selection, despite the factthat selection had occurred for almost twice as longin the two generalist lines.
These two explanations need not be mutuallyexclusive. Indeed, a trade-off between the rates atwhich specialists and generalists evolve seems likely,at least in the short-term, considering that geneticcorrelations can prevent a population from achievingthe optimal phenotype in any given environment(Via & Lande, 1985; Gomulkiewicz & Kirkpatrick,1992). In the long-term, however, these correlationswould be broken down by mutation, so a cost ofadaptation must be ultimately responsible forpreventing the evolution of universally superiorgeneralists.
Acknowledgements
We are grateful to M. Romer and C. Cooney forassistance with the growth chambers in the Phyto-tron, Y. Ducharme and S. Goho for technical assist-ance, D. Kramer and G. Stirling for discussion. Thiswork was supported by a Natural Sciences andEngineering Research Council (NSERC) of CanadaResearch Grant to G.B. and an NSERC Postgrad-uate Fellowship to R.K.
References
BELL, G. 1997. Selection: the Mechanism of Evolution.Chapman & Hall, Toronto.
BELL, G. AND REBOUD, X. 1997. Experimental evolution inChlamydomonas. II. Genetic variation in stronglycontrasted environments. Heredity, 78, 498–506.
BENNETT, A. F., LENSKI, R. E. AND MITTLER, J. E. 1992.Evolutionary adaptation to temperature. 1. Fitnessresponses of Escherichia coli to changes in its thermalenvironment. Evolution, 46, 16–30.
BULMER, M. 1994. Theoretical Evolutionary Ecology.Sinauer Associates, Sunderland, MA.
CHARNOV, E. L. 1982. The Theory of Sex Allocation.Princeton University Press, Princeton, NJ.
FALCONER, D. S. 1990. Selection in different environments:effects on environmental sensitivity (reaction norm) andon mean performance. Genet. Res., 56, 57–70.
FUTUYMA, D. J. AND MORENO, G. 1988. The evolution ofecological specialization. Ann. Rev. Ecol. Syst., 19,207–233.
GILLESPIE, J. H. 1973. Natural selection with varying selec-tion coefficients — a haploid model. Genet. Res., 21,115–120.
GOMULKIEWICZ, R. AND KIRKPATRICK, M. 1992. Quantita-tive genetics and the evolution of reaction norms.Evolution, 46, 390–411.
HARRIS, E. 1989. The Clamydomonas Sourcebook.Academic Press, New York.
KOMERS, P. 1996. Behavioural plasticity in variableenvironments. Can. J. Zool., 75, 161–169.
LEROI, A., LENSKI, R. E. AND BENNETT, A. F. 1994. Evolu-tionary adaptation to temperature. III. Adaptation ofEscherichia coli to a temporally varying environment.Evolution, 48, 1222–1229.
LEVINS, R. 1968. Evolution in Changing Environments.Princeton University Press, Princeton, NJ.
LLOYD, D. G. 1984. Variation strategies of plants in hetero-geneous environments. Biol. J. Linn. Soc., 21, 357–385.
LYNCH, M. AND GABRIEL, W. 1987. Environmental toler-ance. Am. Nat., 129, 283–303.
MORAN, N. A. 1988. The evolution of host-plant alternationin aphids: evidence for specialization as a dead end.Am. Nat., 132, 681–706.
MORAN, N. A. 1992. The evolutionary maintenance ofalternative phenotypes. Am. Nat., 139, 971–989.
NGUYEN, T. N. H., PHAN, Q. G., DUONG, L. P., BERTRAND, K.P. AND LENSKI, R. E. 1989. Effects of carriage andexpression of the Tn10 tetracycline-resistance operonon the fitness of Escherichia coli K12. Mol. Biol. Evol.,6, 213–225.
PADILLA, D. K. AND ADOLPH, S. C. 1996. Plastic induciblemorphologies are not always adaptive: the importanceof time delays in a stochastic environment. Evol. Ecol.,10, 105– 117.
REBOUD, X. AND BELL, G. 1997. Experimental evolution inChlamydomonas. III. Evolution of specialist and gener-alist types in environments that vary in space and time.Heredity, 78, 507–514.
SCHEINER, S. M. 1993. Genetics and evolution of pheno-typic plasticity. Ann. Rev. Ecol. Syst., 24, 35–68.
STEARNS, S. C. 1989. The evolutionary significance ofphenotypic plasticity. BioScience, 39, 436–445.
SULTAN, S. E. 1992. Phenotypic plasticity and the neo-Darwinian legacy. Evol. Trends Plants, 6, 61–71.
VAN TIENDEREN, P. H. 1991. Evolution of generalists andspecialists in spatially heterogeneous environments.Evolution, 45, 1317–1331.
VIA, S. AND LANDE, R. 1985. Genotype–environment inter-action and the evolution of phenotypic plasticity. Evolu-tion, 39, 505–522.
740 R. KASSEN & G. BELL
© The Genetical Society of Great Britain, Heredity, 80, 732–741.

VIA, S., GOMULKIEWICZ, R., DE JONG, G., SCHEINER, S. M.,SCHLICHTING, S. M., AND VAN TIENDEREN, P. H. 1995.Adaptive phenotypic plasticity: consensus and contro-versy. Trends Ecol. Evol., 10, 212–217.
WHITLOCK, M. C. 1996. The Red Queen beats the jack-of-all-trades: the limitations on the evolution of pheno-typic plasticity and niche breadth. Am. Nat., 148,S65–S77.
SELECTION IN TEMPORALLY VARYING ENVIRONMENTS 741
© The Genetical Society of Great Britain, Heredity, 80, 732–741.






![Remodeling of Chlamydomonas Metabolism Using Synthetic Inducers … · Remodeling of Chlamydomonas Metabolism Using Synthetic Inducers Results in Lipid Storage during Growth1[OPEN]](https://img.pdfslide.us/doc/110x75/5f0b74797e708231d43099e1/remodeling-of-chlamydomonas-metabolism-using-synthetic-inducers-remodeling-of-chlamydomonas.jpg)