Embed Size (px)
Citation preview

Molecular Cell, Vol. 10, 1083–1096, November, 2002, Copyright 2002 by Cell Press
Evidence for Distinct CD4 Silencer Functionsat Different Stages of Thymocyte Differentiation
polypeptide and TCR�. After several rounds of prolifera-tion, the selected cells initiate expression of both CD4and CD8 and rearrangement of their TCR� genes. These
Ichiro Taniuchi,1,3,4 Mary Jean Sunshine,1
Richard Festenstein,2 and Dan R. Littman1,3
1Howard Hughes Medical InstituteCD4�8� (double-positive or DP) cells, which make up theMolecular Pathogenesis Programmajority of thymocytes, then undergo a second round ofSkirball Institute of Biomolecular Medicineselection for export to the periphery if they expressNew York University School of Medicineheterodimeric TCR�� that can bind with appropriate540 First Avenueaffinity to major histocompatibility complex (MHC) mole-New York, New York 10016cules (von Boehmer, 1994). This process, termed posi-2 Department of Medicinetive selection, is followed by loss of expression of one ofGene Control Mechanisms and Diseasethe coreceptors, as thymocytes differentiate into eitherImperial College School of MedicineCD4�8� T cells with helper function or CD4�8� cells withMedical Research Councilcytotoxic function. The commitment toward functionallyClinical Sciences Centredistinct lineages is thus marked by the transcriptionalHammersmith Hospital, Londondownregulation of either CD4 or CD8. It has been pro-United Kingdomposed that signals that govern the expression of thesecoreceptors are also involved in the commitment of thy-mocytes to either of the two lineages (reviewed in Ell-Summarymeier et al., 1999).
Whereas expression of the closely linked CD8� andAn intronic silencer within the CD4 gene is the criticalCD8� genes appears to be regulated by distinct en-cis regulatory element for T cell subset-specific ex-hancers that function at different stages of development,pression of CD4. We have combined transfection stud-transcription of the CD4 gene is regulated by both a Ties with gene targeting in mice to identify several keycell-specific enhancer and a developmentally regulatedsequences within the silencer core that are requiredsilencer (reviewed in Ellmeier et al., 1999). The CD4 en-for gene silencing during thymocyte development. Inhancer, located 13 kb upstream from the transcriptionalmice, mutations in individual sites resulted in varie-initiation site, is active in immature and mature T cellsgated, but heritable, derepression of CD4 in matureof all lineages (Sawada and Littman, 1991). In contrast,CD8� T lymphocytes, whereas compound mutationsthe silencer, located in the first intron of the CD4 gene,resulted in full derepression. These results indicatefunctions to restrict expression of CD4 to the appro-that there is partial redundancy in recruiting a chroma-priate subsets of thymocytes and T cells (Sawada ettin remodeling machinery that results in epigenetical., 1994; Siu et al., 1994). The silencer was previouslysilencing. Mutations in single sites also resulted inmapped to a 434 bp sequence that repressed expres-
partial derepression of CD4 in immature double-nega-sion of a reporter transgene specifically in immature DN
tive thymocytes, but there was no apparent variega-and in CD4�CD8� single-positive (SP) thymocytes and
tion. These findings suggest two distinct modes of cytotoxic T lymphocytes in a position- and orientation-CD4 silencer function at different developmental independent manner (Sawada et al., 1994). By gene tar-stages: active repression in CD4�CD8� thymocytes, geting in mice, the silencer was shown to be requiredin which silencing must be reversible, and epigenetic for the repression of CD4 expression in both DN andgene silencing upon differentiation to the CD8� cyto- CD8 SP thymocytes (Leung et al., 2001; Zou et al., 2001).toxic T cell lineage. A silencer has also been identified in the human CD4
locus (Donda et al., 1996). Several putative functionalIntroduction sites or factor binding sites within the CD4 silencer have
been reported (Donda et al., 1996; Duncan et al., 1996),In the course of their differentiation within the thymus, along with putative corresponding binding factors, in-T lymphocytes display precisely choreographed regula- cluding HES-1, SAF (silencer associated factor), andtion of expression of the coreceptor molecules, CD4 c-Myb (Allen et al., 2001; Kim and Siu, 1998, 1999). How-and CD8. Cell surface expression of these glycoproteins ever, little is known about the physiological function ofis coupled to T cell antigen receptor (TCR)-mediated any of these sites or of the proposed binding factors inselection events required for thymocytes to undergo CD4 gene regulation. Moreover, the mechanism of ac-two distinct developmental transitions. The first, named tion of the CD4 silencer has not been explored in depth� selection, occurs after CD4�8� (double-negative or in vivo.
Several mechanisms of transcriptional repressionDN) thymocytes undergo in-frame VDJ rearrangementsthrough negative regulatory elements have been pro-of their TCR� genes, which results in cell surface expres-posed. In one such mechanism, known as “active repres-sion of a pre-TCR composed of the invariant pre-T�sion,” repressor complexes recruited to cis regulatorysequences inhibit the basal transcriptional machinery3 Correspondence: [email protected] (D.R.L.), taniuchi@(Cowell, 1994; Hanna-Rose and Hansen, 1996; Johnson,bioreg.kyushu-u.ac.jp (I.T.)1995). Another mechanism, chromatin structure-medi-4 Present address: Department of Molecular Genetics, Medical Insti-
tute of Bioregulation, Kyushu University, Fukuoka, 812-8582, Japan. ated transcriptional repression, has been shown to be
Molecular Cell1084
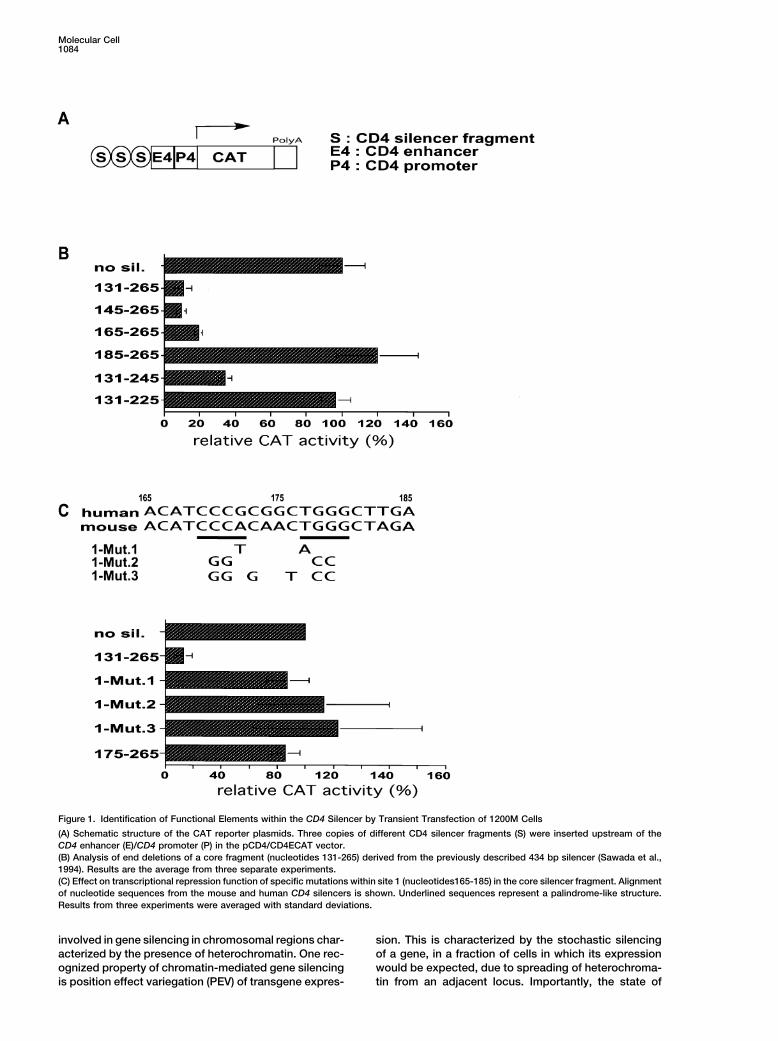
Figure 1. Identification of Functional Elements within the CD4 Silencer by Transient Transfection of 1200M Cells
(A) Schematic structure of the CAT reporter plasmids. Three copies of different CD4 silencer fragments (S) were inserted upstream of theCD4 enhancer (E)/CD4 promoter (P) in the pCD4/CD4ECAT vector.(B) Analysis of end deletions of a core fragment (nucleotides 131-265) derived from the previously described 434 bp silencer (Sawada et al.,1994). Results are the average from three separate experiments.(C) Effect on transcriptional repression function of specific mutations within site 1 (nucleotides165-185) in the core silencer fragment. Alignmentof nucleotide sequences from the mouse and human CD4 silencers is shown. Underlined sequences represent a palindrome-like structure.Results from three experiments were averaged with standard deviations.
involved in gene silencing in chromosomal regions char- sion. This is characterized by the stochastic silencingof a gene, in a fraction of cells in which its expressionacterized by the presence of heterochromatin. One rec-
ognized property of chromatin-mediated gene silencing would be expected, due to spreading of heterochroma-tin from an adjacent locus. Importantly, the state ofis position effect variegation (PEV) of transgene expres-
Mechanisms of CD4 Silencing1085
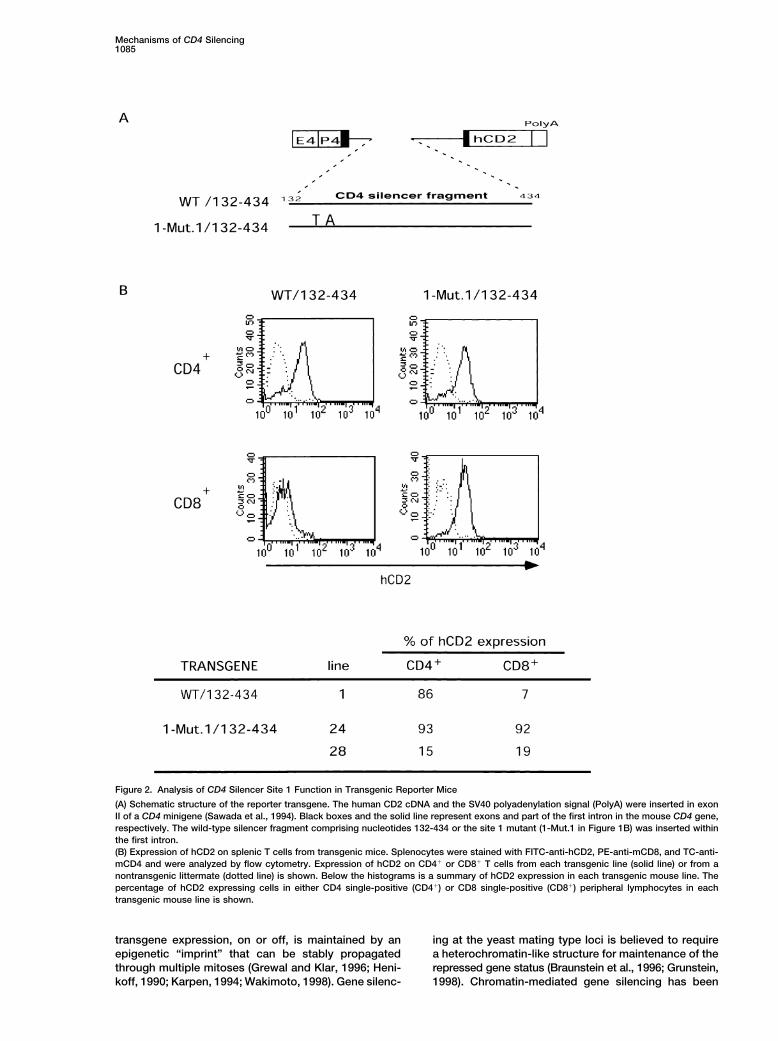
Figure 2. Analysis of CD4 Silencer Site 1 Function in Transgenic Reporter Mice
(A) Schematic structure of the reporter transgene. The human CD2 cDNA and the SV40 polyadenylation signal (PolyA) were inserted in exonII of a CD4 minigene (Sawada et al., 1994). Black boxes and the solid line represent exons and part of the first intron in the mouse CD4 gene,respectively. The wild-type silencer fragment comprising nucleotides 132-434 or the site 1 mutant (1-Mut.1 in Figure 1B) was inserted withinthe first intron.(B) Expression of hCD2 on splenic T cells from transgenic mice. Splenocytes were stained with FITC-anti-hCD2, PE-anti-mCD8, and TC-anti-mCD4 and were analyzed by flow cytometry. Expression of hCD2 on CD4� or CD8� T cells from each transgenic line (solid line) or from anontransgenic littermate (dotted line) is shown. Below the histograms is a summary of hCD2 expression in each transgenic mouse line. Thepercentage of hCD2 expressing cells in either CD4 single-positive (CD4�) or CD8 single-positive (CD8�) peripheral lymphocytes in eachtransgenic mouse line is shown.
transgene expression, on or off, is maintained by an ing at the yeast mating type loci is believed to requirea heterochromatin-like structure for maintenance of theepigenetic “imprint” that can be stably propagated
through multiple mitoses (Grewal and Klar, 1996; Heni- repressed gene status (Braunstein et al., 1996; Grunstein,1998). Chromatin-mediated gene silencing has beenkoff, 1990; Karpen, 1994; Wakimoto, 1998). Gene silenc-
Molecular Cell1086

Mechanisms of CD4 Silencing1087
shown to be important during several early develop- CAT plasmid (Figure 1A). These reporter plasmids weretransfected into 1200M, and reporter gene activity wasmental processes in multicellular organisms. These in-
clude genomic imprinting (Bartolomei and Tilghman, compared to that of cells transfected with a reporterplasmid lacking the CD4 silencer fragment. Suppression1997; Jaenisch et al., 1998; Tilghman, 1999) and X chro-
mosome inactivation in mammals (Brockdorff, 1998; of CAT activity was observed with a construct containinga fragment lying at the center of the CD4 silencer (nucle-Jaenisch et al., 1998) and homeotic gene regulation by
the polycomb group (PcG) proteins in Drosophila (Or- otides 131-265, Figure 1B), but not with constructs con-taining only the 5� (1-132) or 3� (265-434) fragments.lando et al., 1998; Paro et al., 1998). Importantly, these
processes of gene silencing are often maintained and Paradoxically, no silencing of the reporter gene wasobserved in transgenic mice when fragment 132-279inherited by epigenetic mechanisms (Cavalli and Paro,
1998; Csankovszki et al., 1999; Lewin, 1998; Panning from the CD4 silencer was used, whereas lineage-spe-cific silencing of the reporter gene was achieved usingand Jaenisch, 1998; Paro et al., 1998). As cells acquire
their phenotypic identity during developmental pro- additional flanking sequences (1-279 or 132-434) (Ell-meier et al., 1999).cesses by establishing and maintaining cell type-spe-
cific gene expression, chromatin-mediated gene silenc- To further narrow down the critical regions within thecentral fragment, the effects of sequential deletions froming may play an essential role in lineage commitment.
However, little is known about how developmentally reg- either end and of internal deletions were determined. A101 bp region of the CD4 silencer (165-265) retainedulated “facultative” heterochromatin is established.
The CD4 locus presents a unique opportunity to study repressive activity that was nearly equivalent to thatof 131-265 (Figure 1B), but further deletion of the 5�a developmentally regulated vertebrate gene silencer.
It is particularly interesting because the same intronic sequence to nucleotide 185 resulted in complete lossof repression. Deletions of 20 and 40 bp from the 3� endsilencer is required for downregulation of CD4 at two
distinct stages during a linear progression of develop- of the silencer resulted in partial and complete loss ofsilencing, respectively (Figure 1B). Thus, nucleotidesment. In this report, we have used targeted mutagenesis
of the CD4 locus to demonstrate the physiological im- 165-265 constitute the minimal core fragment that con-fers full silencing activity in the in vitro transient transfec-portance of the CD4 silencer and of two novel functional
sites within the silencer. We have observed distinct pat- tion assay.Further characterization of sequence 165-185, whichterns of CD4 derepression at two different stages of T
cell development in mice harboring mutations within the is required for repression in the transfection assay, indi-cated that a centrally placed palindromic sequence issilencer: uniform derepression in immature DN thymo-
cytes and variegated derepression in mature CD8� T essential in the context of the 131-265 CD4 silencerfragment (Figure 1C). As substitution of only two nucleo-lymphocytes. The mutations did not affect maintenance
of silencing in mature T cells in which CD4 had been shut tides (1-Mut.1) was sufficient to abrogate repression, itis likely that a transcription factor participates in CD4off. Together, these findings suggest that chromatin-
mediated gene silencing is involved in shutting off CD4 silencing by binding specifically to this region, whichwe refer to as silencer site 1. Another sequence that isin mature CD8� T lymphocytes. Based on these results,
we propose that the CD4 silencer adopts different func- essential for repression in this assay is 5�-232GACCACA238-3�, a site for binding of runt domain transcrip-tions in the course of T lymphocyte development: it
directs active repression in DN thymocytes and epige- tion factors, which we refer to as site 2 (Taniuchi et al.,2002). Other deletions and substitutions in nucleotidesnetic gene silencing in mature CD8 lineage T lympho-
cytes. 206-225 also resulted in partial loss of repressor activity(data not shown). There are therefore at least three siteswithin the core CD4 silencer that are required for repres-Resultssion of the reporter gene in 1200M cells: site 1 (165-185), site 2 (232-238), and site 3 (206-225).Mapping of Sequences Required for CD4 Silencer
Function in Transient Transfection AssaysTo identify important sequences within the 434 bp CD4 Analysis of CD4 Silencer Function in Transgenic
Reporter Micesilencer, we used the mouse CD8�CD4� thymoma line1200M, in which CD4 silencer-dependent repression of To determine whether point mutations that lead to a
loss of CD4 silencer function in 1200M cells also affectreporter gene activity can be demonstrated using invitro transient transfection assays (Sawada et al., 1994). silencer activity in vivo, we analyzed transgenic mice
prepared with a reporter gene construct containing wild-Three copies of various silencer fragments were insertedupstream of the minimal CD4 enhancer/CD4 promoter- type or mutant silencer fragments. The CD4 silencer
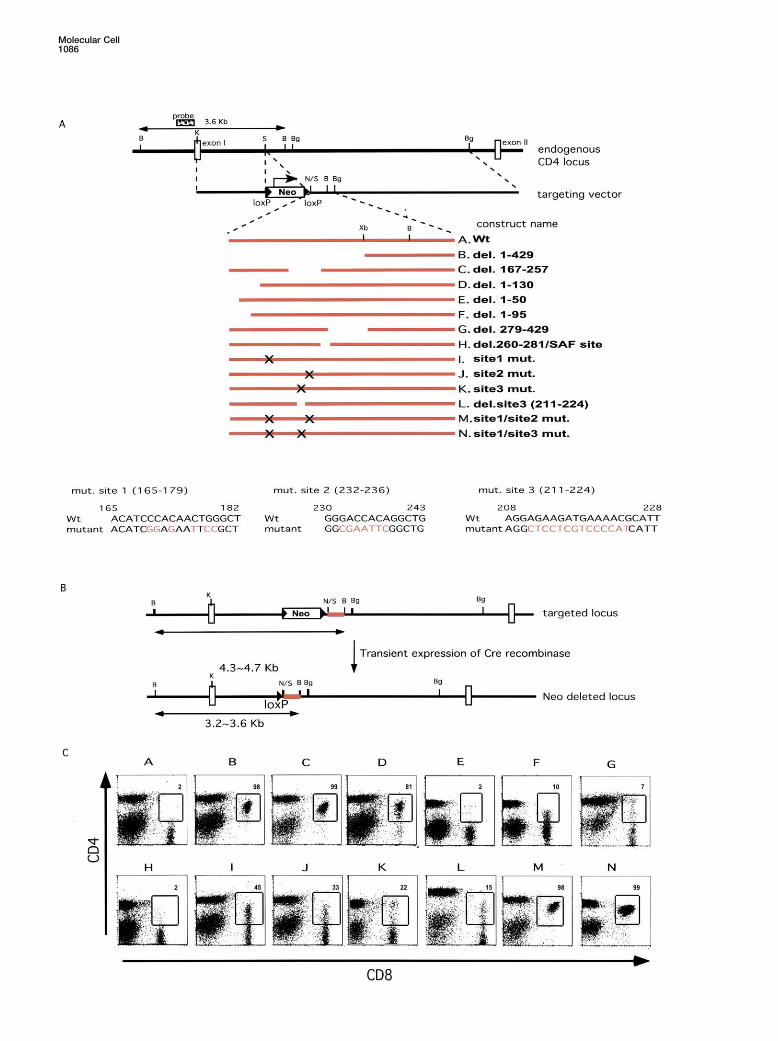
Figure 3. CD4 Expression in Mice with Targeted Mutations in the Silencer
(A) Strategy for introducing deletions or mutations into the silencer region within the CD4 locus. The targeting vectors consist of a 1.8 kb 5�-homology region, a Neomycin resistance gene (Neo) flanked with loxP sites (arrow heads), a segment containing wild-type or mutant silencer,and 5 kb of 3�-homology sequence. Mutant sequences at sites 1-3 in constructs I, J, and K are shown in red at the bottom. The restrictionsites shown are BamHI (B), KpnI (K), SacI (S), BglII (Bg), NotI (N), and XbaI (Xb).(B) Strategy for Cre-mediated removal of the Neo gene in ES clones with targeted silencer mutations.(C) Flow cytometric analysis and CD4 and CD8 expression on ES cell-derived peripheral blood lymphocytes from chimeric mice. ES cell-derived lymphocytes were identified by gating for surface expression of Ly9.1.

Molecular Cell1088
fragments were inserted between exons I and II in a gion, we also tested the effect of shorter deletions fromthe 5� end of the silencer. Deletion of 50 bp had no effectmouse CD4 minigene, in which human CD2 expression
was under the control of the minimal CD4 enhancer (construct E), but further deletion to position 95 resultedin variegated expression of CD4 in a sizeable proportionand promoter (Sawada et al., 1994) (Figure 2A). Upon
insertion of a functional CD4 silencer fragment con- of CD8� T cells (construct F). The mean fluorescenceintensity of derepressed CD4 in cells from these micetaining the core sequence and the 3� flanking sequence
(132-434), hCD2 expression was observed in CD4� T was notably lower than in mice with the 1-130 mutation.These results indicate that there are key factor bindinglymphocytes, but not in CD8� T lymphocytes (Figure
2B). In contrast, insertion of the same fragment con- sites between nucleotides 50-130 in the CD4 silencerand suggest that there may be independent sites intaining the 2 bp substitution in site 1 (1-Mut.1), which
had no activity in 1200M cells, resulted in equal percent- regions 50-95 and 95-130. Deletion of the 3� flankingsequence, 279-429 (construct G), resulted in derepres-ages of CD8� and CD4� T lymphocytes expressing
hCD2 (Figure 2B). Similar results were obtained with sion of CD4 in a small population of CD8� T lymphocytes(�10%). Because SAF (silencing associated factor) wasmultiple transgenic founders prepared with a construct
containing a deletion of sequence 205-225 (site 3) within reported to bind to the CD4 silencer at nucleotides 260-281 (Kim and Siu, 1999), we also deleted this region ofthe silencer (data not shown). These results demonstrate
that the same mutations can abrogate CD4 silencer the gene to analyze the potential role of SAF in silencing(construct H). Elimination of the SAF binding region hadfunction in both the transient transfection assays and
in transgenic reporter assays in vivo. no effect on CD4 silencing.We then analyzed the consequence of mutating sites
that we had mapped within the core sequence of theEffects on Silencing of Mutations withinsilencer. Mutations of each of these sites resulted inthe Endogenous CD4 Silencer Locusdifferent degrees of variegated derepression of CD4 inTo confirm the in vivo importance of individual sitesmature CD8 lineage T cells (Figure 3, constructs I, J, K,within the CD4 silencer, we targeted mutations withinL). These results demonstrate that sites 1, 2, and 3 arethe endogenous locus by homologous recombination ineach required for full silencer activity but that there isES cells. As both CD4 alleles are expressed during Tsome level of redundant function. This was reinforcedlymphocyte development, loss of silencer function onby the finding that combinations of mutations at siteseither allele is expected to result in derepression of CD41 and 2 (construct M) and sites 1 and 3 (construct N)on peripheral CD8� T lymphocytes. A 770 bp SacI-BglIIresulted in complete loss of CD4 silencing in maturefragment containing the wild-type CD4 silencer was re-CD8� T cells (Figure 3). Together, these results clearlyplaced with a series of mutant CD4 silencer fragments,demonstrate that the three sites perform partially redun-as shown in Figure 3A and described in Experimentaldant functions.Procedures. A neomycin resistance gene (Neo) flanked
with loxP sequences was used for selection and wasremoved, by transient transfection with a vector encod- Variegation in CD4 Expression Is Due to Stochastic
Loss of CD4 Silencinging Cre recombinase, from cells in which homologousrecombination had been confirmed (Figure 3B). The ES The observation that single mutations in the CD4 si-
lencer result in either loss or maintenance of silencingcells were used to generate chimeric mice, and periph-eral T cells expressing the ES-derived isotype marker in CD8� T cells can be explained by two models. There
may be specific subpopulations of CD8 lineage cells inLy9.1 were analyzed for CD4 and CD8 expression byflow cytometry (Figure 3C). There was no derepression which individual silencer binding sites are not essential
for CD4 silencing. In this case, the percentage of CD8�of CD4 in CD8� T lymphocytes from chimeric mice gen-erated with the wild-type construct A, indicating that T lymphocytes in which CD4 is derepressed would be
expected to be the same in heterozygous and homozy-introduction of a single loxP sequence upstream of theCD4 silencer had no effect (Figure 3C). gous mutant mice. Alternatively, CD8� T lymphocytes
in which silencing fails or succeeds may be generatedAll peripheral CD8� T lymphocytes with deletions ofeither the full silencer (1-429, construct B) or of the core stochastically in mutant mice. If this occurs, the percent-
age of CD8� T lymphocytes derepressing CD4 in homo-sequence (165-257, construct C) expressed CD4. Theseresults demonstrate that the core sequence is essential zygous mutant mice should be higher than in heterozy-
gous mutant mice, since CD4 would often be expressedfor silencing CD4 expression in mature cytotoxic T cells.We also deleted each of the sequences flanking the from both alleles. We compared CD4 expression on
CD8� T lymphocytes from mice heterozygous or homo-core, since we had observed that either the 5� or 3�sequence is required for silencing reporter transgenes zygous for the germline transmitted site 1 mutation (Fig-
ure 4A). CD4 was expressed on 36% (n � 4, 36.05 �in mice. Deletion of the 5� flanking sequence, 1-130,resulted in variegated derepression of CD4 in CD8� T 7.2) and 64% (n � 3, 64.3 � 6.4) of CD8� T lymphocytes
in heterozygous and homozygous mutant mice, respec-lymphocytes (construct D, Figure 3C). Although CD4expression was observed in approximately 80% of the tively. This result corresponds to the expected fre-
quency of CD4 derepression if loss of silencing occursCD8� T cells, there remained a clearly defined popula-tion in which CD4 expression was shut off. Interestingly, stochastically on each allele in the homozygous site 1
mutant mice.in the transgenic reporter expression system, this se-quence was dispensable as long as the 3� flanking se- Loss of CD4 silencing can be observed in thymocytes
from mice with a nonfunctional silencer (Leung et al.,quence was included (Figure 2; data not shown). Tofurther map functional sequences in the 5� flanking re- 2001; Zou et al., 2001). Deletion of the entire silencer
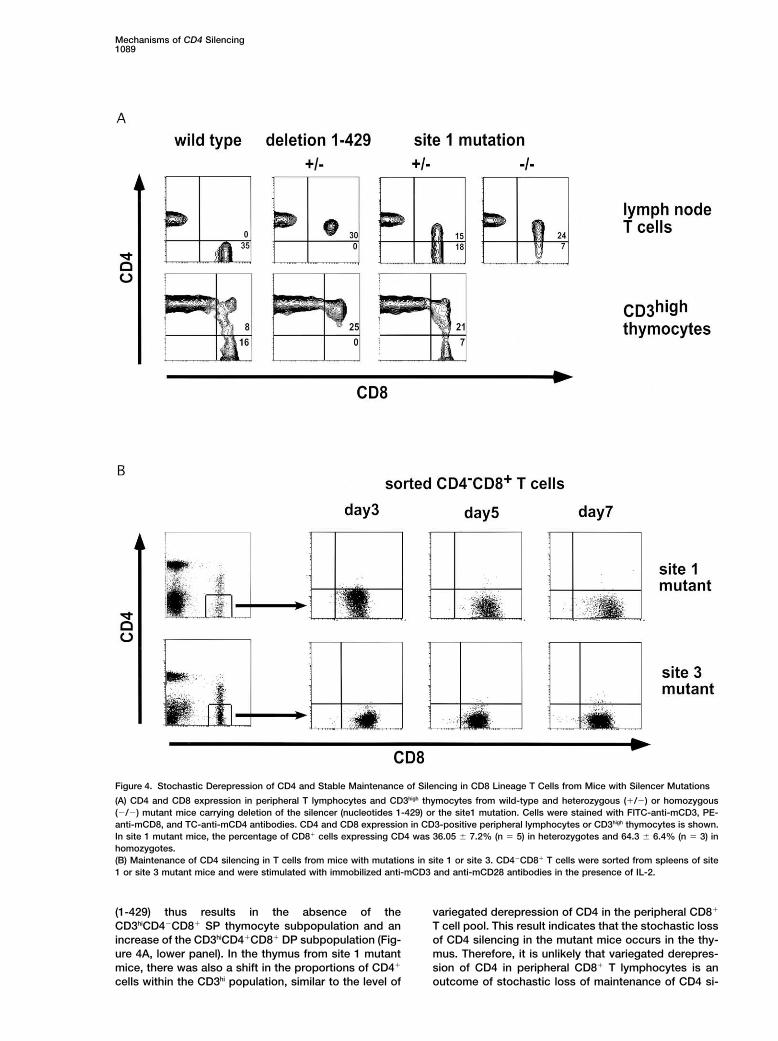
Mechanisms of CD4 Silencing1089
Figure 4. Stochastic Derepression of CD4 and Stable Maintenance of Silencing in CD8 Lineage T Cells from Mice with Silencer Mutations
(A) CD4 and CD8 expression in peripheral T lymphocytes and CD3high thymocytes from wild-type and heterozygous (�/�) or homozygous(�/�) mutant mice carrying deletion of the silencer (nucleotides 1-429) or the site1 mutation. Cells were stained with FITC-anti-mCD3, PE-anti-mCD8, and TC-anti-mCD4 antibodies. CD4 and CD8 expression in CD3-positive peripheral lymphocytes or CD3high thymocytes is shown.In site 1 mutant mice, the percentage of CD8� cells expressing CD4 was 36.05 � 7.2% (n � 5) in heterozygotes and 64.3 � 6.4% (n � 3) inhomozygotes.(B) Maintenance of CD4 silencing in T cells from mice with mutations in site 1 or site 3. CD4�CD8� T cells were sorted from spleens of site1 or site 3 mutant mice and were stimulated with immobilized anti-mCD3 and anti-mCD28 antibodies in the presence of IL-2.
(1-429) thus results in the absence of the variegated derepression of CD4 in the peripheral CD8�
T cell pool. This result indicates that the stochastic lossCD3hiCD4�CD8� SP thymocyte subpopulation and anincrease of the CD3hiCD4�CD8� DP subpopulation (Fig- of CD4 silencing in the mutant mice occurs in the thy-
mus. Therefore, it is unlikely that variegated derepres-ure 4A, lower panel). In the thymus from site 1 mutantmice, there was also a shift in the proportions of CD4� sion of CD4 in peripheral CD8� T lymphocytes is an
outcome of stochastic loss of maintenance of CD4 si-cells within the CD3hi population, similar to the level of

Molecular Cell1090
lencing in these cells. This conclusion is reinforced by establish, but not maintain, an epigenetically stablestate of gene inactivation. Several recent studies havethe finding that silencing is stably maintained for up to
7 days in sorted CD4�CD8� T cells from site 1 and shown that heterochromatin protein 1 (HP-1) has a rolein formation of inactive regions of chromatin observedsite 3 mutant mice subjected to multiple rounds of cell
division in the presence of anti-CD3, anti-CD28, and IL-2 in position effect variegation in Drosophila and in mouse(Eissenberg and Elgin, 2000; Festenstein et al., 1999).(Figure 4B). Intracellular staining showed that there was
no CD4 expression in these cells, indicating that ab- To determine if HP-1 may be involved in the stochasticestablishment of silencing in mice with mutations in thesence of CD4 from the surface is not due to its internal-
ization after T cell activation (data not shown). The find- CD4 silencer, we bred the site 3 mutant mice to micethat express an HP-1� transgene regulated by the CD2ing that binding sites within the CD4 silencer are not
required for maintenance of silencing is consistent with locus control region in thymocytes (Festenstein et al.,1999). Whereas CD4 was expressed on approximatelyour recent finding that the entire silencer, contained
within a 1.6 kb fragment, can be excised from mature 35% of CD8� peripheral T cells in the site 3 mutantmice, the proportion of cells with derepressed CD4 wasCD8 lineage cells without derepression of the CD4 gene
(Zou et al., 2001). significantly reduced to 27% in animals that also carriedthe HP-1� transgene (Figure 6). In contrast, the level ofCD4 expression in immature DN thymocytes was notCD4 Derepression in DN Thymocytes from Silenceraltered in the same transgenic mice (Figure 6A). TheMutant Miceenhancement of CD4 silencing in CD8� T cells was notThe CD4 silencer was previously shown to repress re-the result of a nonspecific inhibitory effect of the HP-porter transgene expression in immature DN thymo-1� transgene on transcription, as CD4 expression oncytes, and targeted deletion of the silencer in mice re-mature CD4�CD8� T cells was not affected in thesesulted in derepression of CD4 in these cells (Leung et al.,mice (data not shown). These results support a role for2001; Sawada et al., 1994; Zou et al., 2001). To determinethe heterochromatin proteins in the establishment ofwhether the same cis acting sites are involved in silenc-epigenetic silencing of CD4 in mature CD8 lineage thy-ing in DN and CD8 SP thymocytes, we examined expres-mocytes and T cells.sion of CD4 in thymocytes derived from mice with germ-
line targeted mutations in the silencer. For this analysis,we gated on CD8�CD3�TCR�Thy1� thymocytes, Discussionwhich correspond to DN thymocytes in wild-type mice(Figure 5). In thymi from mice with deletion of the entire Gene silencing during development of multicellular or-silencer (1-429), CD4 expression was detected in the ganisms is likely to have a critical role in the regulationentire population of immature DN thymocytes. The ex- of cell and tissue fates and in the stable inheritancepression level of derepressed CD4 in CD8�CD3�� of gene expression programs in differentiated tissues.TCR�Thy1� cells was lower than in peripheral CD8� T While genetic studies in Drosophila and yeast have iden-lymphocytes but was similar to that observed previously tified important components involved in silencing of indi-upon deletion of a 1.6 kb segment encompassing the vidual genes, there is relatively little known about mech-silencer (Zou et al., 2001). In mice with a germline muta- anisms and factors involved in gene silencing duringtion in site 1, there was uniform derepression of CD4 in vertebrate development. We have focused our attentionthe CD8�CD3��TCR�Thy1� thymocytes, in contrast on the cis elements required for silencing of the CD4to the variegated derepression observed in peripheral gene because of the importance of the gene product inCD8� T lymphocytes (Figure 5). Although derepression T cell differentiation and function and, also, because ofwas observed in all DN thymocytes, the level of CD4 the likelihood that studies of this locus will contributeexpression was lower than in mice with deletion of the to our general understanding of how individual genes areentire silencer. Similarly, there was expression of CD4 regulated epigenetically during vertebrate development.in the entire population of immature thymocytes from By using conditional excision of the intronic silencer,mice with a mutation in site 3, although expression was we previously showed that it is required for shutting offeven lower than in mice with the mutation of site 1. expression of CD4 in both immature double-negativeSites 1 and 3 in the core region of the CD4 silencer are and mature CD8 lineage thymocytes, but that it is dis-therefore required for efficient silencing in both DN and pensable for stably maintaining a silenced state in ma-CD8 SP thymocytes. However, CD4 derepression in ture cytotoxic T cells (Zou et al., 2001). DNA methylation,mice with mutations at these sites differs significantly which has a key role in X chromosome inactivation andin its pattern according to the stage of development: genomic imprinting, does not appear to be involved inthere is uniform partial derepression in CD4�CD8� DN maintenance of CD4 silencing (Zou et al., 2001). Otherthymocytes versus variegated derepression in mature mechanisms that could be involved in the heritability ofCD8� T lymphocytes, suggesting that different mecha- silencing at this locus have not yet been explored.nisms of gene silencing are operating in the different T By generating mutations in individual elements withincell subsets. the CD4 silencer, we have been able to show that it
shares several properties with silencers in yeast andDrosophila. Like the HML and HMR mating type locusHP-1 Enhances CD4 Silencing in Mice with a Site 3
Silencer Mutation silencers in the budding yeast Saccharomyces cerevis-iae (Pillus and Rine, 1989), there are several cis actingThe observation that CD4 expression is derepressed in
a variegated pattern in CD8 lineage T cells in mutant sites with partially redundant function, such that muta-tion of any single site results in only partial loss of silenc-mice is consistent with a requirement for the silencer to
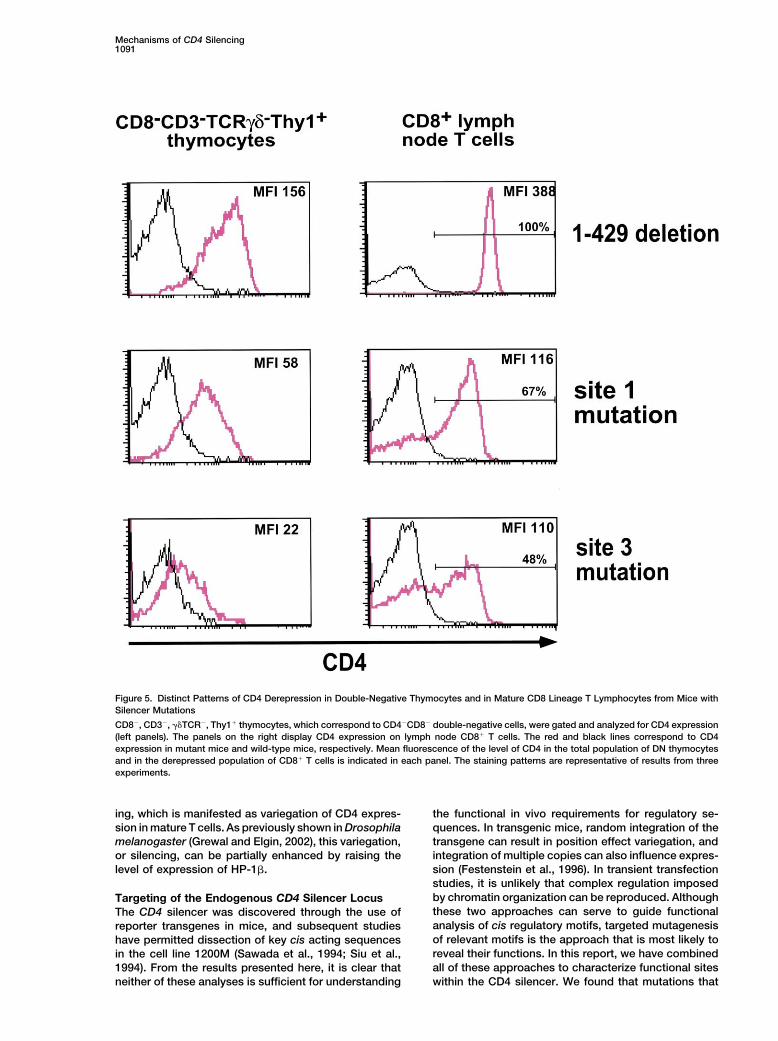
Mechanisms of CD4 Silencing1091
Figure 5. Distinct Patterns of CD4 Derepression in Double-Negative Thymocytes and in Mature CD8 Lineage T Lymphocytes from Mice withSilencer Mutations
CD8�, CD3�, TCR�, Thy1� thymocytes, which correspond to CD4�CD8� double-negative cells, were gated and analyzed for CD4 expression(left panels). The panels on the right display CD4 expression on lymph node CD8� T cells. The red and black lines correspond to CD4expression in mutant mice and wild-type mice, respectively. Mean fluorescence of the level of CD4 in the total population of DN thymocytesand in the derepressed population of CD8� T cells is indicated in each panel. The staining patterns are representative of results from threeexperiments.
ing, which is manifested as variegation of CD4 expres- the functional in vivo requirements for regulatory se-quences. In transgenic mice, random integration of thesion in mature T cells. As previously shown in Drosophila
melanogaster (Grewal and Elgin, 2002), this variegation, transgene can result in position effect variegation, andintegration of multiple copies can also influence expres-or silencing, can be partially enhanced by raising the
level of expression of HP-1�. sion (Festenstein et al., 1996). In transient transfectionstudies, it is unlikely that complex regulation imposedby chromatin organization can be reproduced. AlthoughTargeting of the Endogenous CD4 Silencer Locusthese two approaches can serve to guide functionalThe CD4 silencer was discovered through the use ofanalysis of cis regulatory motifs, targeted mutagenesisreporter transgenes in mice, and subsequent studiesof relevant motifs is the approach that is most likely tohave permitted dissection of key cis acting sequencesreveal their functions. In this report, we have combinedin the cell line 1200M (Sawada et al., 1994; Siu et al.,all of these approaches to characterize functional sites1994). From the results presented here, it is clear that
neither of these analyses is sufficient for understanding within the CD4 silencer. We found that mutations that
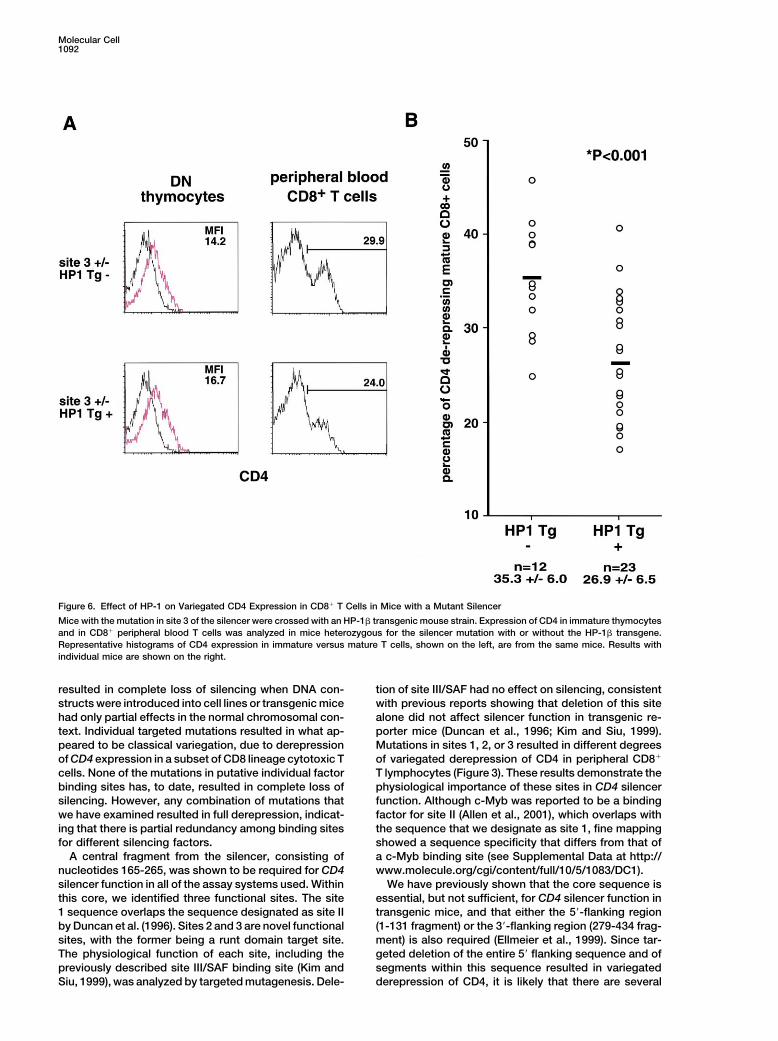
Molecular Cell1092
Figure 6. Effect of HP-1 on Variegated CD4 Expression in CD8� T Cells in Mice with a Mutant Silencer
Mice with the mutation in site 3 of the silencer were crossed with an HP-1� transgenic mouse strain. Expression of CD4 in immature thymocytesand in CD8� peripheral blood T cells was analyzed in mice heterozygous for the silencer mutation with or without the HP-1� transgene.Representative histograms of CD4 expression in immature versus mature T cells, shown on the left, are from the same mice. Results withindividual mice are shown on the right.
resulted in complete loss of silencing when DNA con- tion of site III/SAF had no effect on silencing, consistentwith previous reports showing that deletion of this sitestructs were introduced into cell lines or transgenic mice
had only partial effects in the normal chromosomal con- alone did not affect silencer function in transgenic re-porter mice (Duncan et al., 1996; Kim and Siu, 1999).text. Individual targeted mutations resulted in what ap-
peared to be classical variegation, due to derepression Mutations in sites 1, 2, or 3 resulted in different degreesof variegated derepression of CD4 in peripheral CD8�of CD4 expression in a subset of CD8 lineage cytotoxic T
cells. None of the mutations in putative individual factor T lymphocytes (Figure 3). These results demonstrate thephysiological importance of these sites in CD4 silencerbinding sites has, to date, resulted in complete loss of
silencing. However, any combination of mutations that function. Although c-Myb was reported to be a bindingfactor for site II (Allen et al., 2001), which overlaps withwe have examined resulted in full derepression, indicat-
ing that there is partial redundancy among binding sites the sequence that we designate as site 1, fine mappingshowed a sequence specificity that differs from that offor different silencing factors.
A central fragment from the silencer, consisting of a c-Myb binding site (see Supplemental Data at http://www.molecule.org/cgi/content/full/10/5/1083/DC1).nucleotides 165-265, was shown to be required for CD4
silencer function in all of the assay systems used. Within We have previously shown that the core sequence isessential, but not sufficient, for CD4 silencer function inthis core, we identified three functional sites. The site
1 sequence overlaps the sequence designated as site II transgenic mice, and that either the 5�-flanking region(1-131 fragment) or the 3�-flanking region (279-434 frag-by Duncan et al. (1996). Sites 2 and 3 are novel functional
sites, with the former being a runt domain target site. ment) is also required (Ellmeier et al., 1999). Since tar-geted deletion of the entire 5� flanking sequence and ofThe physiological function of each site, including the
previously described site III/SAF binding site (Kim and segments within this sequence resulted in variegatedderepression of CD4, it is likely that there are severalSiu, 1999), was analyzed by targeted mutagenesis. Dele-

Mechanisms of CD4 Silencing1093
functional sites that are important in silencing within the In mice with targeted mutations of the CD4 silencer,the level of derepressed CD4 expression in the DN cellscontext of the chromosome, but not in repression in
transient tranfection reporter analysis. correlated with the proportion of mature CD8� T cellsin which CD4 was expressed. Thus, in DN thymocytes,the level of CD4 expression was greater in site 1 mutantDifferent Modes of Silencer Function in DNmice than in site 3 mutant mice, and expression of bothversus CD8 SP Thymocyteswas considerably lower than that observed when theDuring the last decade, many studies have shown thatentire silencer was deleted; and the proportion of CD8heterochromatin has an inhibitory effect on gene ex-lineage cells in which CD4 was expressed was corre-pression. A well-characterized effect of heterochromatinspondingly greater in mice harboring a mutation in siteis position effect variegation of transgene expression1 than in those with the site 3 mutation. This may reflect(Festenstein et al., 1996; Milot et al., 1996). Propagationthe relative abilities of specific factors to bind to theseof heterochromatin structure toward a locus in whichtwo sites or to interact in ternary complexes at boththe transgene is integrated is believed to inhibit its ex-stages of development. Thus, in the absence of bindingpression in a proportion of cells (Wakimoto, 1998; Wall-of a site 3 factor, there may still be relatively strongrath, 1998). In this study, we have observed variegatedbinding of factors to other sites, ensuring that a largederepression of CD4 following introduction of individualproportion of cells can initiate silencing. Conversely, inmutations into the CD4 silencer locus. In this context,the absence of binding to site 1, binding to the otherit is unlikely that the variegation is a result of propagatedsites may be less frequent, and the silencing machineryheterochromatin from another locus. Since variegatedwould have a lower probability of being recruited at theCD4 derepression was observed upon mutation of indi-appropriate time in development. We also noted that,vidual motifs, but not upon deletion of the entire CD4even in CD8� mature T cells, CD4 derepression wassilencer, it probably results from stochastic loss of si-below the full level observed in the absence of the entirelencing due to reduced recruitment of the silencing ma-silencer or when double mutations were present (Figurechinery to the variant CD4 locus. Like variegated trans-5). This may reflect the sustained ability of silencer-gene expression, CD4 derepression was suppressed byassociated factors to bind to the region even if epige-expression of HP-1�, consistent with a role for hetero-netic chromatin remodeling fails to occur (see model inchromatin binding proteins in the stochastic process ofFigure 7). Such binding would interfere with silencing inestablishing a silenced state at the CD4 locus. Anothera manner analogous to that observed in the DN thymo-important characteristic of chromatin-mediated gene si-cytes, in which an active form of transcriptional repres-lencing is the epigenetic maintenance of the repressedsion may be operating.status of a locus (Lewin, 1998; Paro et al., 1998; Wallrath,
Based on these observations, we propose that the1998). We showed that, once established, silencing isCD4 gene is repressed by distinct mechanisms at differ-maintained in mature CD8� T lymphocytes from miceent stages of differentiation, even though the same ciswith mutations of the silencer. This is consistent withacting sequences appear to be required at both stagesour previous finding that there was no derepression of(Figure 7). Our model is further supported by the findingCD4 in mature CD8� T lymphocytes even after deletion
of the entire silencer (Zou et al., 2001). Together, these of distinct requirements for trans acting factors for CD4silencing at different stages of differentiation: Runx1observations demonstrate that silencing of the CD4 lo-
cus is maintained by epigenetic mechanisms in mature for DN thymocytes and Runx3 for mature CD8� T cells(Taniuchi et al., 2002). Moreover, mutation of both con-CD8 lineage cells. Since variegated derepression of CD4
was already observed in CD8�CD3high thymocytes, epi- sensus motifs for runt domain transcription factor bind-ing (sites 2 and 2�, see Figure 7) resulted in completegenetic CD4 silencing appears to be established in the
thymus during maturation of the CD4loCD8� thymocytes loss of silencing in mature CD8� T cells but only partialderepression in DN thymocytes (Taniuchi et al., 2002).(Zou et al., 2001). It has been reported that the inactive
CD4 locus is localized close to heterochromatic struc- In addition, compromised function of the BAF chromatinremodeling complex enhanced derepression of CD4 intures in nuclei from B lymphocytes, although this has
not been demonstrated in CD8 lineage T cells (Brown DN thymocytes, but not in mature CD8 lineage cells, inmice harboring a mutation in site 1 of the silencer (Chi etet al., 1997).
In contrast to the variegation observed in mature T al., 2002). In contrast, overexpression of HP-1 enhancedCD4 silencing in mature CD8 lineage T cells, but not incells, immature thymocytes from mice with mutations
of individual sites in the CD4 silencer exhibited uniform, DN thymocytes (Figure 6). In DN thymocytes, silencerbinding factors may recruit corepressors that activelypartial derepression of CD4. Since the pattern of dere-
pression in CD4�CD8� DN thymocytes was different inhibit the basal transcriptional machinery through pro-tein-protein interactions. Corepressors may recruit his-from that in mature CD8 lineage cells, it is likely that
repression operates by different mechanisms at these tone deacetylases, resulting in a transient reversible al-teration in local chromatin structure. This inhibitorytwo stages of development. This is perhaps not surpris-
ing, since the repressed state of CD4 expression is re- process may require remodeling of chromatin by theBAF complex. In CD8 SP thymocytes and mature T cells,versed after � selection, whereas it appears to be stably
inherited in mature cytotoxic T cells. Therefore, it is silencer binding factors likely recruit not only histonedeacetylases, but also factors that can alter the stateunlikely that an irreversible chromatin modification at
the CD4 locus is responsible for repression in of the chromatin, resulting in its condensation into het-erochromatin, thus rendering the inactive state perma-CD4�CD8� DN thymocytes. Further experiments will be
necessary to distinguish the chromatin state of the CD4 nent. The initial silencing mechanism in these cells maybe similar to that operating in DN thymocytes, resultinglocus at the different stages of T cell development.
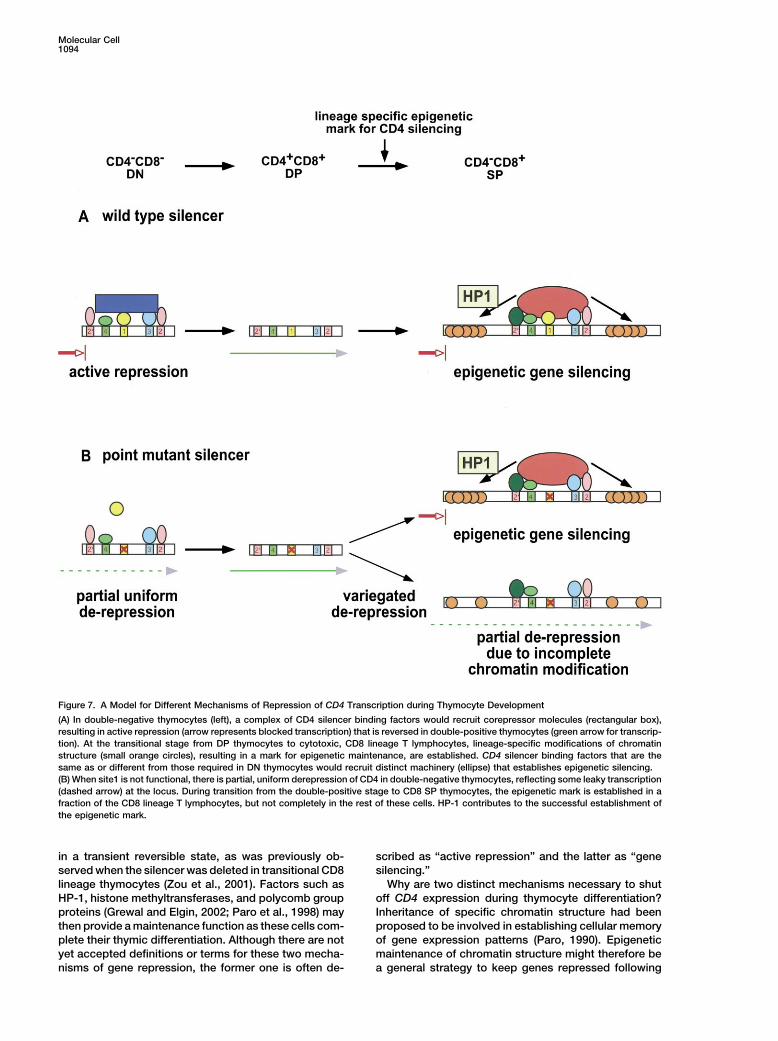
Molecular Cell1094
Figure 7. A Model for Different Mechanisms of Repression of CD4 Transcription during Thymocyte Development
(A) In double-negative thymocytes (left), a complex of CD4 silencer binding factors would recruit corepressor molecules (rectangular box),resulting in active repression (arrow represents blocked transcription) that is reversed in double-positive thymocytes (green arrow for transcrip-tion). At the transitional stage from DP thymocytes to cytotoxic, CD8 lineage T lymphocytes, lineage-specific modifications of chromatinstructure (small orange circles), resulting in a mark for epigenetic maintenance, are established. CD4 silencer binding factors that are thesame as or different from those required in DN thymocytes would recruit distinct machinery (ellipse) that establishes epigenetic silencing.(B) When site1 is not functional, there is partial, uniform derepression of CD4 in double-negative thymocytes, reflecting some leaky transcription(dashed arrow) at the locus. During transition from the double-positive stage to CD8 SP thymocytes, the epigenetic mark is established in afraction of the CD8 lineage T lymphocytes, but not completely in the rest of these cells. HP-1 contributes to the successful establishment ofthe epigenetic mark.
in a transient reversible state, as was previously ob- scribed as “active repression” and the latter as “genesilencing.”served when the silencer was deleted in transitional CD8
lineage thymocytes (Zou et al., 2001). Factors such as Why are two distinct mechanisms necessary to shutoff CD4 expression during thymocyte differentiation?HP-1, histone methyltransferases, and polycomb group
proteins (Grewal and Elgin, 2002; Paro et al., 1998) may Inheritance of specific chromatin structure had beenproposed to be involved in establishing cellular memorythen provide a maintenance function as these cells com-
plete their thymic differentiation. Although there are not of gene expression patterns (Paro, 1990). Epigeneticmaintenance of chromatin structure might therefore beyet accepted definitions or terms for these two mecha-
nisms of gene repression, the former one is often de- a general strategy to keep genes repressed following

Mechanisms of CD4 Silencing1095
PCR and was used to replace the wild-type NotI-BglII CD4 silencerterminal differentiation of diverse cell types. Therefore,fragment, to generate constructs B-N.it is reasonable that such mechanisms would keep the
30 �g of each targeting vector was excised by KpnI digestion andCD4 gene silenced after commitment to the cytotoxicwas transfected into the ES cell line E14, by electroporation of 5 �
T cell lineage, and that other reversible mechanisms 106 cells at 240 mV, 500 �F using a Gene Pulser. After selectingwould be involved in repression prior to this stage. cells with 350 �g/ml of G418, individual colonies generated with
each construct were subjected to Southern blotting to identify ho-mologous recombination events, using a 0.5 kb XbaI-KpnI fragmentExperimental Procedurescorresponding to the promoter region. ES clones that had under-gone homologous recombination were transfected with 30 �g ofIn Vitro Transient Transfection AssaypMC-Cre to excise the loxP-flanked Neo gene. Colonies wereFor CAT reporter plasmids, CD4 silencer fragments were generatedscreened for acquired sensitivity to G418, and this was confirmedby PCR with appropriate primers. The DNA sequence of each frag-by Southern blotting. ES clones were injected into blastocysts fromment was confirmed. Three copies of each CD4 silencer fragmentC57BL/6 mice to generate chimeric mice.were placed in the polylinker site at the 5� end of pCD4/CD4ECAT
(Sawada and Littman, 1991). 20 �g of CAT reporter plasmid wasLymphocyte Stimulation and Cell Culturetransfected into 5 � 106 to 1 � 107 1200M cells, together with 10Splenic T lymphocytes were prepared by purification on columns�g of pGL3-enhancer luciferase vector (Promega) as internal controlfrom R&D Systems and were stained with anti-CD4 and anti-CD8for transfection efficiency, by electroporation (260 mV, 975 �F) usingantibodies for subsequent cell sorting on a Coulter Elite sorter. 1-4 �a Bio-Rad Gene Pulser. All experiments were performed in duplicate.106 T lymphocytes were stimulated with 2 �g of plate-bound anti-Forty-eight hours after transfection, the CAT activity from cell ex-CD3 (Pharmingen) and 2 �g of anti-mCD28 antibodies (Pharmingen).tracts of each transfection was assayed as described previouslyTwo days after the first stimulation, T cells were cultured with 20(Sawada and Littman, 1991). In all experiments, values were normal-units/ml of mIL-2 (Boehringer Mannheim) and were restimulatedized relative to the activity of the plasmid pCD4/CD4ECAT (E4P4),with fresh mIL-2 every 2 days.which does not contain the CD4 silencer fragment.
AcknowledgmentsGeneration of Transgenic MiceThe CD4 silencer fragment, wild-type 132-434 and 1-Mut.1 132-434,
We thank Shinichiro Sawada and Sanjeev Mahanta for their contri-were generated by PCR with appropriate primers, and the sequencebutions to the characterization of the core silencer sequence. Weof this fragment was confirmed. Each of these CD4 silencer frag-are grateful to John Hirst for cell sorting, to Balthazar Cazac andments was inserted into the HindIII site of p4617 (transgene c, Sa-Takeshi Egawa for their critical reading of the manuscript, and towada et al., 1994) to produce the transgene constructs, wild-typemembers of the Littman laboratory for helpful discussions. D.R.L.132-434 and 1-Mut.1 132-434. B6/D2 F1 eggs were microinjectedis an investigator of the Howard Hughes Medical Institute.with these transgene constructs to generate transgenic mice.
Received: June 18, 2002Antibodies and Flow CytometryRevised: September 3, 2002PE-conjugated anti-mCD8, Tri-Color (TC)-conjugated anti-mCD4,
APC-conjugated anti-mThy-1.2, and FITC-conjugated anti-hCD2Referenceswere from Caltag. FITC-conjugated anti-Ly9.1, FITC-conjugated
anti-mCD3, FITC-conjugated anti-mCD4, PE-conjugated anti-Allen, R.D., 3rd, Kim, H.K., Sarafova, S.D., and Siu, G. (2001). Nega-mouse TCR, biotin-conjugated anti-mCD3, biotin-conjugatedtive regulation of CD4 gene expression by a HES-1-c-Myb complex.anti-mCD8, and APC-conjugated anti-mCD3 were from PharMingen.Mol. Cell. Biol. 21, 3071–3082.Biotin-conjugated antibodies were visualized by TC-conjugated
strepavidin from Caltag. For analysis of immature thymocytes, the Bartolomei, M.S., and Tilghman, S.M. (1997). Genomic imprinting incells were stained with FITC-anti-mCD4, PE-anti-mCD3, PE-anti- mammals. Annu. Rev. Genet. 31, 493–525.mCD8, PE-anti-TCR, and APC-anti-Thy1 antibodies. Peripheral
Braunstein, M., Sobel, R.E., Allis, C.D., Turner, B.M., and Broach,blood and lymph node cells were stained with FITC-anti-mCD4 and
J.R. (1996). Efficient transcriptional silencing in Saccharomyces cer-PE-anti-mCD8 antibodies. Cells stained with antibodies were ana-
evisiae requires a heterochromatin histone acetylation pattern. Mol.lyzed using Becton Dickinson FACScan or FACs Calibur flow cyto-
Cell. Biol. 16, 4349–4356.meters and CELLQuest software.
Brockdorff, N. (1998). The role of Xist in X-inactivation. Curr. Opin.Genet. Dev. 8, 328–333.
Construction of Target Vectors and Generation of Chimeric MiceBrown, K.E., Guest, S.S., Smale, S.T., Hahm, K., Merkenschlager,For the 5� region of homology in the CD4 gene, a 2.5 kb KpnI-BglIIM., and Fisher, A.G. (1997). Association of transcriptionally silentfragment spanning part of exon I and the first intron was insertedgenes with Ikaros complexes at centromeric heterochromatin. Cellinto the KpnI-BamHI sites in the pCR2.1-TOPO vector (Invitrogen).91, 845–854.This plasmid was then digested with SacI/BstXI to remove the 0.7Cavalli, G., and Paro, R. (1998). The Drosophila Fab-7 chromosomalkb SacI-BglII fragment that included the intronic silencer (Figure 3).element conveys epigenetic inheritance during mitosis and meiosis.The neomycin resistant (Neo) gene flanked with loxP sites from theCell 93, 505–518.pL2-Neo (2) plasmid (Gu et al., 1993) was digested with SalI/XhoI,
blunted, and inserted into the SacI/BstXI-digested plasmid. The 3� Chi, T.H., Wan, M., Zhao, K., Taniuchi, I., Chen, L., Littman, D.R.,region of homology, a 4.5 kb BglII fragment from the first intron, and Crabtree, G.R. (2002). Reciprocal regulation of CD4/CD8 ex-was subcloned into a separate pCR2.1-TOPO vector to allow exci- pression by SWI/SNF-like BAF complexes. Nature 418, 195–199.sion with NotI and KpnI. Inserts containing the 5� region of homology Cowell, I.G. (1994). Repression versus activation in the control ofplus the Neo gene and the 3� region of homology were excised from gene transcription. Trends Biochem. Sci. 19, 38–42.their respective plasmids as HindIII-NotI and NotI-Kpn fragments
Csankovszki, G., Panning, B., Bates, B., Pehrson, J.R., and Jaenisch,and were ligated into HindIII/KpnI-digested pUC19, yielding theR. (1999). Conditional deletion of Xist disrupts histone macroH2ApCassette vector. The 770 bp SacI-BglII fragment, which containslocalization but not maintenance of X inactivation. Nat. Genet. 22,the CD4 silencer region and an engineered NotI site at the 5�-end,323–324.was generated by PCR with an appropriate primer set and was
inserted into NotI/BglII-digested pCassette vector to generate wild- Donda, A., Schulz, M., Burki, K., De Libero, G., and Uematsu, Y.(1996). Identification and characterization of a human CD4 silencer.type construct A. Each mutant CD4 silencer fragment with a NotI
site at the 5�-end and BglII site at the 3�-end was generated by Eur. J. Immunol. 26, 493–500.

Molecular Cell1096
Duncan, D.D., Adlam, M., and Siu, G. (1996). Asymmetric redundancy Sawada, S., Scarborough, J.D., Killeen, N., and Littman, D.R. (1994).A lineage-specific transcriptional silencer regulates CD4 gene ex-in CD4 silencer function. Immunity 4, 301–311.pression during T lymphocyte development. Cell 77, 917–929.Eissenberg, J.C., and Elgin, S.C. (2000). The HP1 protein family:Siu, G., Wurster, A.L., Duncan, D.D., Soliman, T.M., and Hedrick,getting a grip on chromatin. Curr. Opin. Genet. Dev. 10, 204–210.S.M. (1994). A transcriptional silencer controls the developmentalEllmeier, W., Sawada, S., and Littman, D.R. (1999). The regulationexpression of the CD4 gene. EMBO J. 13, 3570–3579.of CD4 and CD8 coreceptor gene expression during T cell develop-Taniuchi, I., Osato, M., Egawa, T., Sunshine, M.J., Bae., S.-C., Ko-ment. Annu. Rev. Immunol. 17, 523–554.mori, T., Ito, Y., and Littman, D.R. (2002). Differential requirementsFestenstein, R., Tolaini, M., Corbella, P., Mamalaki, C., Parrington,for Runx proteins in CD4 repression and epigenetic silencing duringJ., Fox, M., Miliou, A., Jones, M., and Kioussis, D. (1996). LocusT lymphocyte development. Cell 111, in press.control region function and heterochromatin-induced position effectTilghman, S.M. (1999). The sins of the fathers and mothers: genomicvariegation. Science 271, 1123–1125.imprinting in mammalian development. Cell 96, 185–193.Festenstein, R., Sharghi-Namini, S., Fox, M., Roderick, K., Tolaini,von Boehmer, H. (1994). Positive selection of lymphocytes. Cell 76,M., Norton, T., Saveliev, A., Kioussis, D., and Singh, P. (1999). Het-219–228.erochromatin protein 1 modifies mammalian PEV in a dose- and
chromosomal-context-dependent manner. Nat. Genet. 23, 457–461. Wakimoto, B.T. (1998). Beyond the nucleosome: epigenetic aspectsof position-effect variegation in Drosophila. Cell 93, 321–324.Grewal, S.I., and Klar, A.J. (1996). Chromosomal inheritance of epi-
genetic states in fission yeast during mitosis and meiosis. Cell 86, Wallrath, L.L. (1998). Unfolding the mysteries of heterochromatin.95–101. Curr. Opin. Genet. Dev. 8, 147–153.Grewal, S.I., and Elgin, S.C. (2002). Heterochromatin: new possibili- Zou, Y.R., Sunshine, M.J., Taniuchi, I., Hatam, F., Killeen, N., andties for the inheritance of structure. Curr. Opin. Genet. Dev. 12, Littman, D.R. (2001). Epigenetic silencing of CD4 in T cells commit-178–187. ted to the cytotoxic lineage. Nat. Genet. 29, 332–336.Grunstein, M. (1998). Yeast heterochromatin: regulation of its as-sembly and inheritance by histones. Cell 93, 325–328.
Gu, H., Zou, Y.R., and Rajewsky, K. (1993). Independent control ofimmunoglobulin switch recombination at individual switch regionsevidenced through Cre-loxP-mediated gene targeting. Cell 73,1155–1164.
Hanna-Rose, W., and Hansen, U. (1996). Active repression mecha-nisms of eukaryotic transcription repressors. Trends Genet. 12,229–234.
Henikoff, S. (1990). Position-effect variegation after 60 years. TrendsGenet. 6, 422–426.
Jaenisch, R., Beard, C., Lee, J., Marahrens, Y., and Panning, B.(1998). Mammalian X chromosome inactivation. Novartis Found.Symp. 214, 200–209.
Johnson, A.D. (1995). The price of repression. Cell 81, 655–658.
Karpen, G.H. (1994). Position-effect variegation and the new biologyof heterochromatin. Curr. Opin. Genet. Dev. 4, 281–291.
Kim, H.K., and Siu, G. (1998). The notch pathway intermediate HES-1silences CD4 gene expression. Mol. Cell. Biol. 18, 7166–7175.
Kim, W.W., and Siu, G. (1999). Subclass-specific nuclear localizationof a novel CD4 silencer binding factor. J. Exp. Med. 190, 281–291.
Leung, R.K., Thomson, K., Gallimore, A., Jones, E., Van den Broek,M., Sierro, S., Alsheikhly, A.R., McMichael, A., and Rahemtulla, A.(2001). Deletion of the CD4 silencer element supports a stochasticmechanism of thymocyte lineage commitment. Nat. Immunol. 2,1167–1173.
Lewin, B. (1998). The mystique of epigenetics. Cell 93, 301–303.
Milot, E., Strouboulis, J., Trimborn, T., Wijgerde, M., de Boer, E.,Langeveld, A., Tan-Un, K., Vergeer, W., Yannoutsos, N., Grosveld,F., et al. (1996). Heterochromatin effects on the frequency and dura-tion of LCR-mediated gene transcription. Cell 87, 105–114.
Orlando, V., Jane, E.P., Chinwalla, V., Harte, P.J., and Paro, R. (1998).Binding of trithorax and Polycomb proteins to the bithorax complex:dynamic changes during early Drosophila embryogenesis. EMBOJ. 17, 5141–5150.
Panning, B., and Jaenisch, R. (1998). RNA and the epigenetic regula-tion of X chromosome inactivation. Cell 93, 305–308.
Paro, R. (1990). Imprinting a determined state into the chromatin ofDrosophila. Trends Genet. 6, 416–421.
Paro, R., Strutt, H., and Cavalli, G. (1998). Heritable chromatin statesinduced by the Polycomb and trithorax group genes. NovartisFound. Symp. 214, 51–61.
Pillus, L., and Rine, J. (1989). Epigenetic inheritance of transcrip-tional states in S. cerevisiae. Cell 59, 637–647.
Sawada, S., and Littman, D.R. (1991). Identification and character-ization of a T-cell-specific enhancer adjacent to the murine CD4gene. Mol. Cell. Biol. 11, 5506–5515.