Embed Size (px)
Citation preview

Plant Physiol. (1982) 70, 430-4360032-0889/82/70/0430/07/$00.50/0
Emission of Hydrogen Sulfide by Leaf Tissue in Response toL-Cysteine'
Received for publication April 6, 1981 and in revised form June 10, 1981
JIRO SEKIYA2, AHLERT SCHMIDT3, LLOYD G. WILSON, AND PHILIP FILNER4MSU-DOE Plant Research Laboratory, Michigan State University, East Lansing, Michigan 48824
ABSTRACT
Leaf discs and detached leaves exposed to L-cysteine emitted a volatilesulfur compound which was proven by gas chromatography to be H2S. Thisphenomenon was demonstrated in all nine species tested (Cucumis sativus,Cucurbita pepo, Nicotiana tabacum, Coleus bbumei, Beta vulgaris, Phaseolusvulgaris, Medicago sativa, Hordeum vulgare, and Gossypium hirsutum). Theemission of volatile sulfur by cucumber leaves occurred in the dark at asimilar rate to that in the light. The emission of leaf discs reached themaximal rate, more than 40 picomoles per minute per square centimeter,2 to 4 hours after starting exposure to L-cysteine; then it decreased. In thecase of detached leaves, the maximum occurred 5 to 10 h after startingexposure. The average emission rate of H2S during the first 4 hours fromleaf discs of cucurbits in response to 10 milimlar L-cysteine, was usuallymore than 40 picomoles per minute per square centimeter, i.e. 0.24 micro-moles per hour per square decimeter. Leaf discs exposed to 1 milHimolar L-cysteine emitted only 2% as much as did the discs exposed to 10 millmolarL-cysteine. The emission from leaf discs and from detached leaves lastedfor at least 5 and 15 hours, respectively. However, several hours after themaximal emission, injury ofthe leaves, manifested as chlorosis, was evdent.H2S emission was a specific consequence ofexposure to L-cysteine; neitherD-cysteine nor L-cystine elicited H2S emission. Aminooxyacetic acid, aninhibitor ofpyridoxal phosphate dependent enzymes, inhibited the emission.In a celi free system from cucumber leaves, H2S formation and its releaseoccurred in response to L-cysteine. Feeding experiments with 135SIL_CyS_teine showed that most of the sulfur in H2S was derived from sulfur in theL-cysteine supplied and that the H2S emitted for 9 hours accounted for 7to 10% of L-cysteine taken up. 35S-labeled S032- and S042- were found inthe tissue extract in addition to internal soluble S2-. These findigs suggestthe existence of a sulfur cycle which converts L-cysteine to S042- throughcysteine desulfhydration.
Illuminated green leaves emit H2S when plants are exposed tos042- (32, 35) or SO2 (6, 27). Plants have the potential forreduction of s042- to a bound form of sulfide, which is incorpo-rated into L-cysteine, by a light-driven assimilation pathway (1,26). Therefore, the conversion of bound sulfide to free sulfide andits release as H2S is one possible origin of the H2S emitted inresponse to s042- (35). Another possible route to H2S from SO2is via direct reduction of free s032- to H25, catalyzed by sulfite
' Supported by the United States Department of Energy under ContractDE-AC02-76ERO-1338.
2 Present address: Department of Agricultural Chemistry, YamaguchiUniversity, Yamaguchi 753, Japan.
3 Present address: Botanisches Institut der Universitat Munchen, Men-zinger Str. 67, D-8000 Munchen, FRG.
4 Present address: ARCO Plant Cell, Research Institute, Dublin, CA94566.
reductase (1, 26).Still another possibility is that L-cysteine could be a precursor
of H2S. L-Cysteine is a precursor ofmost organic sulfur compounds(9), and it regulates s042- uptake (13, 16, 29, 30), ATP sulfurylase(24, 33), adenosine-5'-phosphosulfate sulfotransferase (33, 36),thiosulfonate reductase (25), O-acetylserine sulfhiydrylase (20, 30),L-serine transacetylase (28), and nitrogen metabolism (33). InSalmonella, L-cysteine is degraded to pyruvate, NH4' and sulfideby L-cysteine desulfhydrase, which is induced by L-cysteine (4, 5,14, 15). L-cysteine desulfhydrase activity has also been reported toexist in the XD strain of cultured tobacco cells and to be inducedby L-cysteine in these cells (12). The H2S could also arise bycyanide-dependent desulfhydration of L-cysteine, which is cata-lyzed by f8-cyanoalanine synthase, an enzyme found in manyplant species (19). If leaves contain L-cysteine desulfhydratingactivity, it is conceivable that the L-cysteine level could be regu-lated by L-cysteine desulflhydration, which would prevent accu-mulation of L-cysteine to injurious levels.We report here evidence of sulfide formation and its emission
as H2S by cucurbit leaves exposed to L-cysteine.
MATERIALS AND METHODS
Plant Materials. Cucumber plants (Cucumis sativus cv. Chipper,seeds multiplied after at least 5 cycles of sibling crosses) weregrown for 30 to 40 days with a half-strength Hoagland's solutionin a growth chamber programmed for 14 h of light (maximum 8mw cm-2) at 28°C followed by 10 h of dark at 16°C (35).Cucumber plants usually had six to nine leaves; the upper threeto five leaves were still expanding, and the lower three or fourleaves were fully expanded and matured. The second or third leaffrom an apex was used in the experiments presented, unless noted.Pumpkin plants (Cucurbita pepo cv. Small Sugar Pumpkin) weregrown for 25 to 30 days under the above conditions. Squash(Cucurbita pepo cv. Prolific Straight Neck Squash), another culti-var of cucumber (C. sativus cv. National Pickling), tobacco (Ni-cotiana tabacum cv. Burley 21), coleus (Coleus blumet), sugar beetBeta vulgaris cv. US H20), alfalfa (Medicago sativa cv. Pioneer531), barley (Hordeum vulgare cv. Kearney), and kidney bean(Phaseolus vulgaris cv. Scotia) were grown similarly. Cotton (Gos-sypium hirsutum cv. Acala SJ-1) was also grown in a growthchamber (17).Measurement of H2S Emission. Detached leaves with leaf area
of about 80 cm2 or leaf discs (2.65 cm2 disc-) were used tomeasure H2S emission. In the case of detached leaves, a singleleaf, the petiole of which was in 20 ml of treatment solution in asmall vial sealed with Parafilm, was placed in a 1-L flask. Theflask was sealed with a rubber stopper covered with Teflon TFEtape. The stopper had two ports: one port was an air inlet and theother was connected to the four-channel automatic sample selec-tor, which, in turn, was coupled to a sulfur analyzer with a flamephotometric detector (Model 8450, Monitor Labs). Air was pulledat 180 ml min-' through a leaf chamber by the pump of the sulfur
430
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

EMISSION OF HYDROGEN SULFIDE FROM CYSTEINE
analyzer or by another pump when that leaf chamber was notbeing monitored. When leaf discs were used, 10 discs punched outfrom one-half of a leaf divided at the midrib were floated on 10ml of reference solution in a 250-ml flask, and another 10 discsfrom the other one-half of the same leaf were floated on 10 ml ofsolution containing a chemical to be tested. The lower surface ofeach disc was exposed to the treatment solution, and the uppersurface was exposed to the air. Disc chambers (250-ml flasks) wereconnected to a sulfur analyzer through an automatic sampleselector in a similar way to that used for leaf chambers fordetached leaves. Detached leaves were illuminated at 1 mw cm-2by cool-white fluorescent lamps; leaf discs were illuminated at 8mw cm-2 by 400-w phosphor-coated metal halide lamps. Lightintensity was measured with a radiometer (Model YSI 65A,Yellow Springs Instruments). The temperature was 26 ± 1°Cduring all measurements. The pH of each treatment solution wasadjusted to 6. One g fresh weight corresponds to about 60 cm2 ofleaf area.Gas Chromatography Analysis. The volatile sulfur emitted was
analyzed with a gas chromatograph (Varian 3700) equipped witha flame photometric detector for sulfur, in which Chromosil 330packed in a Teflon FEP column (1.83 m x 0.32 cm, Supelco, Inc)was mounted. The column temperature was 55°C, and the flowrate of carrier gas (N2) was 15 ml min-'. Detector output, set atattenuation 2 and range 10-9 amp mv-1, was in the direct mode,so that response was proportional to the square of sulfur concen-tration in a sample. One-ml samples of air containing volatilesurfur were taken from each leaf chamber through a septum by a3-ml plastic syringe (Becton-Dickinson Co.) and injected into thegas chromatograph.Measurement of 135SIH2S Emitted by Leaves Treated with ["6Si
L-Cysteine. The petiole of a detached young leaf was put in 10mM L-cysteine solution containing [35S]L-cysteine (the Radiochem-ical Centre). The leaf mounted in the vial containing the treatmentsolution was placed in a 1-L flask leaf chamber, and air waspushed through the chamber at 180 ml min-' and then through atrap for H2S consisting of 10 ml of a mixture containing 100 mmzinc acetate and 40 mm sodium acetate in water (zinc acetate trap).Cool-white fluorescent lamps provided illumination at 1 mw cm-'.35S in aliquots of the zinc acetate trap or in ZnS precipitatescollected by centrifugation of trap solution at 12,000g for 10 minwere determined by liquid scintillation counting. The amount ofH2S in zinc acetate traps was determined colorimetrically.
Extraction and Fractionation of 3"S Compounds. To stabilizesulfhydryl compounds s032 and sulfide, NEM5 was used forextraction (7). After the petiole of a leaf incubated with [35S]L-cysteine was removed, the leaf was homogenized with 10 ml ofcold 80% (v/v) ethanol containing 50 mm NEM for 2 min by aTekmar homogenizer at full speed. The homogenate was centri-fuged at 12,000g for 10 min. Residue was reextracted with 10 ml80% ethanol containing 50 mm NEM. The supematants werecombined and vacuum-evaporated at 40°C. To the residue afterevaporation, CHC13:CH30H:H20 (55:15:30, v/v) was added. Thewhole mixture was transferred to a separatory funnel and shakenvigorously. CHC13 phase was removed, and 11 ml fresh CHC13was added to the aqueous phase and partitioned again. The pooledCHC13 fraction was washed with 6 ml H20. The aqueous washwas pooled with the aqueous fraction. The pooled aqueous frac-tion and CHC13 fraction were made to 50 ml with H20 and CHC13,respectively; these are referred to as ethanol-aqueous fraction andthe CHC13 fraction. The residue, after ethanol extraction, washomogenized with 10 ml 1% (w/v) TCA solution for 2 min atroom temperature to extract [35SISO42-, and the homogenate wascentrifuged at 12,000g for 10 min. The residue was reextractedwith 10 ml 1% TCA solution. The combined TCA supernatant
5 Abbreviations: NEM, N-ethylamaleimide; AOA, aminooxyacetic acid.
fraction was neutralized with 1 N NaOH and made to 25 ml; it isreferred to as TCA fraction. The residue after TCA extraction wastransferred onto cellulose filter paper. Radioactivities of 35S inaliquots of ethanol-aqueous, CHC13, and TCA fractions and ofthe whole residue on filter paper were measured by liquid scintil-lation counting using 10 ml scintillation fluid (4 g PPO, 0.1 gPOPOP, 330 ml Triton X-100, and 670 ml toluene). The rest ofthe ethanol-aqueous and TCA fractions were evaporated at 40°Cand dissolved separately in small amounts of H20. Both concen-trates were subjected to TLC and electrophoresis.
Colorimetric Determination of H2S. H2S was determined byformation of methylene blue from dimethyl-p-phenylenediaminein H2SO4, essentially according to the method by Gustafsson (10).An aliquot of the zinc acetate trap was put in a small test tube (12x 75 mm) and made up to 2.4 ml with H20. The test tube wassealed quickly with a rubber serum bottle cap with the insidesurface covered with Teflon TFE tape, after addition of 0.3 ml 5mM dimethyl-p-phenylenediamine dissolved in 3.5 M H2SO4. Then0.3 ml of 50 mm ferric ammonium sulfate in 100 mm H2SO4 wasinjected by means of a glass syringe through the serum bottle cap.The A at 667 nm was measured after leaving the mixture for 15min at room temperature. Blanks were prepared by the sameprocedures with unused zinc acetate solution. A calibration curvewas made with known concentrations of Na2S solution each timethe assay was done.
Preparation of Authentic NEM Adducts of Sulfur-ContainingCompounds. Unlabeled NEM adducts of L-cysteine and reducedglutathione were prepared by mixing L-cysteine or reduced glu-tathione with equivalent (molar) 1.1 mol ofNEM at pH 6.5 for 30min at 24°C. Formation of NEM adducts was followed by de-crease in A at 302 nm. After the reaction, NEM adducts of L-cysteine and glutathione were crystallized by addition of acetone(31). NEM adducts obtained are referred to as NEM-cysteine andNEM-glutathione, respectively. For preparation of[14C]NEM-cys-teine or ['4C]NEM-glutathione, 1ltCi of N-ethyl-[1_14C]maleimide(New England Nuclear) was mixed with L-cysteine or reducedglutathione and then 1.1 mol eq of unlabeled NEM were addedto each mixture. These [14C]NEM adducts were purified by TLCon cellulose plates (see below). Other NEM adducts of sulfhydrylcompounds were prepared in a similar manner. NEM-[35S]sulfitewas obtained by mixing [35S]Na2SO3, prepared from elemental[35S]sulfur (2), with 1.1 mol eq NEM and used without furtherpurification. NEM-[35S]sulfide was obtained by mixing NEM with[35S]H2S, prepared by reduction of [35SJNa2SO4 (New EnglandNuclear) (1 1). All reactions of sulfur compounds with NEM wereperformed at pH 6.5. The obtained NEM adducts were dissolvedin distilled H20 and used as authentic compounds for TLC andelectrophoresis.
Electrophoresis and TLC. Thin layer electrophoresis was per-formed on 20-cm-long thin-layer cellulose plates (Baker-flex cel-lulose plate, J. T. Baker) with 100 mm sodium acetate buffer, pH4.0. A sample was spotted at 7 cm from the cathodal end and runat 600 v for 20 min at 4°C. For TLC, samples spotted at 3 cmfrom the lower edge on 20-cm-long thin-layer cellulose plates weredeveloped with tert-butyl alcohol:88% formic acid:H20 (14:3:3, v/v, pH 1.5). For two dimensional development, the sample waselectrophoresed as a first dimensional development and thenchromatograyhed as a second dimensional development. Extractscontaining 3 S compounds were usually coelectrophoresed or co-chromatographed with unlabeled authentic compounds: L-methi-onine, L-cysteic acid, NEM-cysteine, and NEM-glutathione. Afterdevelopment of the sample, 0.2% (w/v) ninhydrin dissolved inabsolute ethanol was sprayed to locate amino acids. 35S com-pounds were located by radioautography, radioactive spots werescraped, and 35S was determined by liquid scintillation counting.In the case of one dimensional development, 35S was measured byliquid scintillation counting after cutting cellulose plates into 0.5-
431
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 70, 1982
or 1-cm segments. When NEM-cysteine was exposed to alkalineconditions, including a developing solvent at alkaline pH, more
than 50% of NEM-cysteine degraded into a ninhydrin-negativecompound (RF relative to methionine, 1. I1), probably a thiazanederivative (31). Therefore, acidic conditions were used for electro-phoresis and TLC in the experiments presented here. Under theconditions used, each authentic compound tested, except for L-
methionine and NEM-sulfite, showed a single ninhydrin-positivespot and/or a single radioactive spot. L-Methionine containedmethionine sulfoxide, and NEM-sulfite contained So42
RESULTS
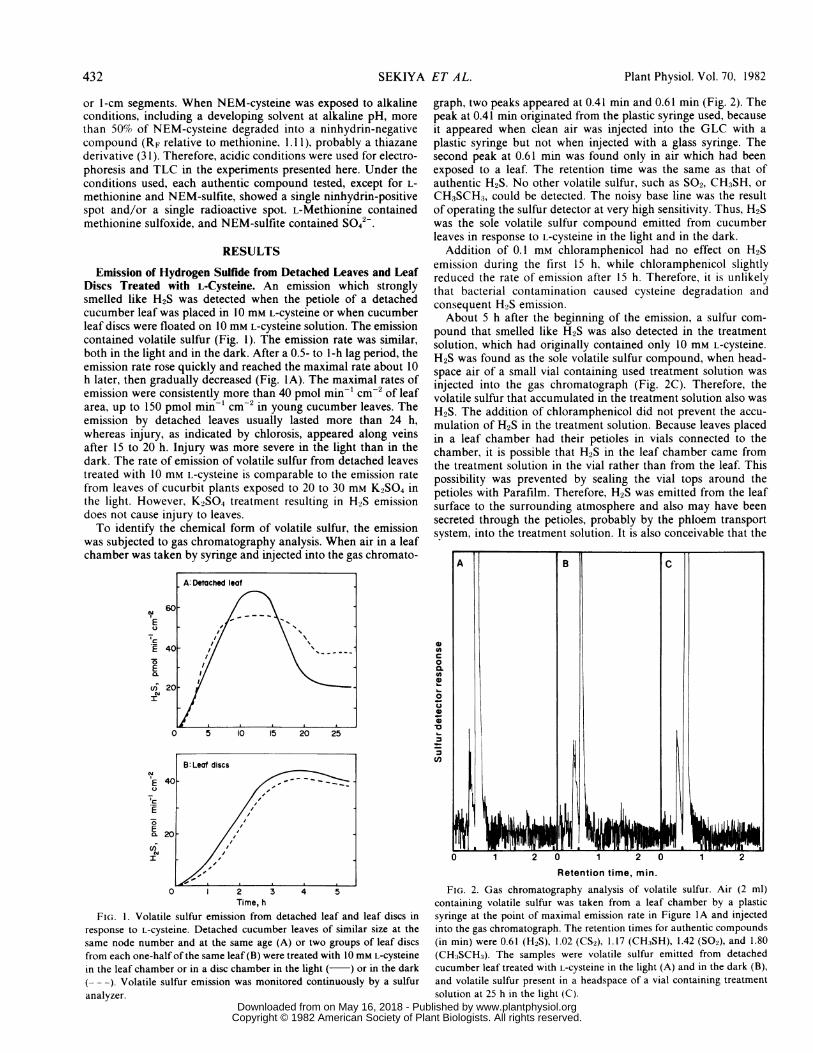
Emission of Hydrogen Sulfide from Detached Leaves and LeafDiscs Treated with L-Cysteine. An emission which stronglysmelled like H2S was detected when the petiole of a detachedcucumber leaf was placed in 10 mM L-cysteine or when cucumberleaf discs were floated on 10 mM L-cysteine solution. The emissioncontained volatile sulfur (Fig. 1). The emission rate was similar,both in the light and in the dark. After a 0.5- to 1-h lag period, theemission rate rose quickly and reached the maximal rate about 10h later, then gradually decreased (Fig. IA). The maximal rates ofemission were consistently more than 40 pmol min-' cm-2 of leafarea, up to 150 pmol min-' cm-2 in young cucumber leaves. Theemission by detached leaves usually lasted more than 24 h,whereas injury, as indicated by chlorosis, appeared along veinsafter 15 to 20 h. Injury was more severe in the light than in thedark. The rate of emission of volatile sulfur from detached leavestreated with 10 mM L-cysteine is comparable to the emission ratefrom leaves of cucurbit plants exposed to 20 to 30 mm K2SO4 inthe light. However, K2SO4 treatment resulting in H2S emission
does not cause injury to leaves.To identify the chemical form of volatile sulfur, the emission
was subjected to gas chromatography analysis. When air in a leafchamber was taken by syringe and injected into the gas chromato-
A: Detached leaf
60-
IT
0 5 10 15 20 25
B:Leafdiscs|
40.
£f 20 /
0 2 3 4 5
Time, h
FIG. 1. Volatile sulfur emission from detached leaf and leaf discs inresponse to L-cysteine. Detached cucumber leaves of similar size at thesame node number and at the same age (A) or two groups of leaf discsfrom each one-half of the same leaf (B) were treated with 10 mM L-cysteinein the leaf chamber or in a disc chamber in the light ( ) or in the dark(--). Volatile sulfur emission was monitored continuously by a sulfur
analyzer.
graph, two peaks appeared at 0.41 min and 0.61 min (Fig. 2). Thepeak at 0.41 min originated from the plastic syringe used, becauseit appeared when clean air was injected into the GLC with aplastic syringe but not when injected with a glass syringe. Thesecond peak at 0.61 min was found only in air which had beenexposed to a leaf. The retention time was the same as that ofauthentic H2S. No other volatile sulfur, such as SO2, CH3SH, orCH3SCH3, could be detected. The noisy base line was the resultof operating the sulfur detector at very high sensitivity. Thus, H2Swas the sole volatile sulfur compound emitted from cucumberleaves in response to L-cysteine in the light and in the dark.
Addition of 0.1 mm chloramphenicol had no effect on H2Semission during the first 15 h, while chloramphenicol slightlyreduced the rate of emission after 15 h. Therefore, it is unlikelythat bacterial contamination caused cysteine degradation andconsequent H2S emission.About 5 h after the beginning of the emission, a sulfur com-
pound that smelled like H2S was also detected in the treatmentsolution, which had originally contained only 10 mM L-cysteine.H2S was found as the sole volatile sulfur compound, when head-space air of a small vial containing used treatment solution wasinjected into the gas chromatograph (Fig. 2C). Therefore, thevolatile sulfur that accumulated in the treatment solution also wasH2S. The addition of chloramphenicol did not prevent the accu-mulation of H2S in the treatment solution. Because leaves placedin a leaf chamber had their petioles in vials connected to thechamber, it is possible that H2S in the leaf chamber came fromthe treatment solution in the vial rather than from the leaf. Thispossibility was prevented by sealing the vial tops around thepetioles with Parafilm. Therefore, H2S was emitted from the leafsurface to the surrounding atmosphere and also may have beensecreted through the petioles, probably by the phloem transportsystem, into the treatment solution. It is also conceivable that the
A B C
CD0
0.
CD,
0 1 2 0 1 2 0 1 2
Retention time, min.
FIG. 2. Gas chromatography analysis of volatile sulfur. Air (2 ml)containing volatile sulfur was taken from a leaf chamber by a plasticsyringe at the point of maximal emission rate in Figure IA and injectedinto the gas chromatograph. The retention times for authentic compounds(in min) were 0.61 (H2S), 1.02 (CS2), 1.17 (CH:3SH), 1.42 (SO2), and 1.80(CH3SCH3). The samples were volatile sulfur emitted from detachedcucumber leaf treated with L-cysteine in the light (A) and in the dark (B),and volatile sulfur present in a headspace of a vial containing treatment
solution at 25 h in the light (C).
432 SEKIYA ET A L.
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

EMISSION OF HYDROGEN SULFIDE FROM CYSTEINE
H2S may have been produced by the petioles themselves.There was a difficulty in experiments with detached whole
leaves because of variability of leaves. Each detached leaf releasedvolatile sulfur at a different emission rate, even when leaves ofsimilar size at the same node number and at the same age wereused. Since the analytical equipment limited us to monitoring onlya small number of leaves at a time, the variability made it difficultor impossible to detect subtle differences in H2S output in responseto various treatments. However, use of leaf discs overcame thevariation problem. When two groups of leaf discs were made, onefrom the right one-half and one from the left one-half of the sameleaf, and the two groups of leaf discs were floated on 10 mM L-cysteine in separate flasks, they emitted volatile sulfur at the samerate. Since both groups of leaf discs have virtually identicalactivities for H2S emission, it is possible to use one as a controlwhen testing the effect of a treatment on the H2S emission of theother.
Floating such paired groups of leaf discs on 10 mM L-cysteineshowed that the discs initially emitted H2S at a similar rate in thelight and in the dark (Fig. IB). Consequently, H2S emission withL-cysteine is light-independent, unlike H2S emission caused bySo42- Leaf discs responded more quickly to chemicals and envi-ronmental changes that did the detached leaves above, but thepattern of changes in H2S emission with time were similar to thoseof detached leaves. H2S rose after a short lag period (about 15min). Then the emission rate increased quickly, reached themaximal rate 2 to 4 h later, and began to decrease gradually.Emission lasted more than 10 h. H2S emission from leaf discstreated with L-cysteine increased as the concentrations of L-Cys-teine increased (Fig. 3). Leaf discs with 1 mM L-cysteine emittedonly 2% of H2S emitted at 10 mM.A volatile sulfur emission, which smelled like H25, was observed
to be produced by all plant species tested but at different emissionrates, when leaf discs of each species were exposed to L-cysteine(Table I). Volatile sulfur from pumpkin leaves in response to L-cysteine also behaved like H2S when subjected to gas chromatog-raphy analysis.
Substrate Specificity of H2S Emission. Analogs of L-cysteinewere used to assess the substrate specificity of the H2S-formingprocess. D-Cysteine, cysteamine, 3-mercaptopropionic acid, L-ho-mocysteine, L-methionine, S-methyl-L-cysteine, glutathione at 10mM, and L-cystine at 2 mm caused no emission of H2S or othervolatile sulfur by leaves of cucumber or pumpkin. The enzyme(s)
150_0N
C~
0
0 100 _
E
~50-
1510 5 10 15
L-cysteine, mMFIG. 3. Dependence of H2S emission on L-cysteine concentration. Ten
cucumber leaf discs from one-half of a leaf were floated on 10 mM L-
cysteine solution as reference and another 10 leaf discs from the other one-
halfof the same leafwere floated on L-cysteine at a different concentration.The H2S emissions in two such experiments at a time were monitored bya sulfur analyzer coupled to an automatic four-channel sample selector.The emission is presented as percentage of emission caused by 10 mM L-
cysteine during the first 4 h in the light.
Table I. Emission of Volatile Sulfur by Leaf Discs in Response to L-Cysteine
Ten leaf discs (26.5 cm2), cut from young and actively growing leavesbetween first and third leaves from the apex, were floated on 10 ml of 10mM L-cystemne solution in a 250-ml-flask leaf chamber. Volatile sulfuremission rate in the light (8 mw cm-2) was continuously monitored by asulfur analyzer. Values for total sulfur emitted were obtained from inte-gration of peak area for first 4 h.
Volatile S Emission
Plant Plant Maximal emission Total SAge Total___
Rate Time emitted
week pmol min-' -2wek -2 h nmolcm2cmCucumber (Chipper) 5 127.0 2.4 18.2Cucumber (National Pickling) 5 87.9 2.5 13.8Pumpkin 5 93.6 3.4 14.1Squash 5 71.2 2.8 9.5Tobacco 8 7.1 2.8 1.3Coleus 24 18.3 5.5 1.6Sugar beet 4 72.5 2.3 10.8Cotton 7 23.0 1.4 4.0Kidney bean 3 26.1 2.7 3.6Alfalfa 4 36.3 5.5 3.6Barley 3 50.8 6.4 2.1
Table II. H2S Emission by Developmentally Different Leaves in Responseto L-Cysteine
Ten leaf discs from each leaf of a single cucumber plant with sevenleaves were floated on 10 mM L-cysteine in a 250-ml flask in the light. H2Semission was monitored by a sulfur analyzer coupled to a four-channeltimer-actuated automatic selector. H2S emitted during the first 4 h wasintegrated, and the average emission rate was calculated.
Leaf Position from Base H2S Emitted.-1 2~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
pmol min-' cm-21 55.22 57.63 67.94 68.75 67.06 84.27 100.8
responsible for H2S formation appears to be remarkably specificfor L-cysteine.
D-Cysteine and S-methyl-L-cysteine at 5 mm stimulated H2Semission in response to L-cysteine to a level twice as great as thatwith L-cysteine alone, while L-methionine, L-cysteic acid, taurine,and L-serine neither stimulated nor inhibited H2S emission causedby L-cysteine.H2S Emission in Response to L-Cysteine by Leaves in Different
Developmental Stages. Cucumber plants growing for 30 to 40days have 6 to 9 leaves which are progressively less mature,developmentally, as one proceeds upward from the base of theplant. The rate of H2S emission in response to L-cysteine wasslightly different between young and mature leaves, when meas-urements were performed on leaf discs from each leaf of a singleplant (Table II). The top leaf, which was still actively expanding,showed the highest rate of H2S emission (100 pmol minm1 cm ),and the bottom leaf had the lowest rate (55 pmol min-1 cm-2).However, leaves other than the upper two had similar rates of H2Semission (55 to 70 pmol min-1 cm-2), though H2S emissiondecreased gradually with leaf age.
Emission of 13551H2S from Leaves Treated with 135SIL-Cysteine.
433
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 70, 1982
When young detached leaves of cucumber were treated with[35SJL-cysteine, volatile 3S sulfur was emitted. It was caught by azinc acetate trap, as described in "Materials and Methods." Incor-poration of 35S into volatile sulfur increased with time (Table III).H2S, which can be determined chemically, was also emitted, andit increased with time (Table III). The volatile 35S trapped by zincacetate for 3, 6, and 9 h was collected by centrifugation as ZnSprecipitate. 35S in the precipitate was determined by liquid scintil-lation counting, and sulfide in the precipitate was determinedcolorimetrically. The specific radioactivities of sulfide in ZnSprecipitates, based on radioactivity and sulfide content of theprecipitates, were relatively constant: 0.20 to 0.29 nCi nmol-1,compared to 0.398 nCi nmol-' for the L-cysteine added initially(Table III). The specific radioactivity of the total 35S in the traps,which was calculated by dividing the total radioactivity in thetraps by the chemically determined amount of sulfide in the traps,was similar to the specific radioactivity of the isolated sulfideprecipitate. If an appreciable fraction of the trapped radioactivitywere not in sulfide, then the specific radioactivity of total 35Swould have been correspondingly greater than that of the sulfideprecipitate. Therefore, most of the volatile 35S had to be [15S]H2S.The small difference between the specific radioactivities of theZnS precipitate and the total zinc acetate trap may arise frompartial oxidation of sulfide in the zinc acetate trap; oxidizedproducts of sulfide do not react with Zn2+ ion to form a precipitate.However, the oxidation rate seems to be slow.The sulfur in H2S, emitted in response to L-cysteine, was mostly
derived from the sulfur in L-cysteine, since specific radioactivitiesof [35SJH2S emitted were 60 to 90%o of the specific radioactivity ofthe [35SJL-cysteine applied. A comparison of specific radioactivitiesalso indicates that the initial pool size of L-cysteine and interme-diate(s), if any, between L-cysteine and H2S are small comparedto that of exogenous L-cysteine taken up.The extents of conversion of [35SJL-cysteine taken up to [tSJH2S
emitted were 2.1, 3.3, and 6.5% during 3, 6, and 9 h in the light,respectively, and 9.9% during 9 h in the dark (Table IV). H2S was,therefore, a major metabolite of the [3S]L-cysteine taken up bythe leaves.
Inhibition of H2S Emission from L-Cysteine. L-Cysteine analogsdid not inhibit H2S emission by leaf discs exposed to L-cysteine.These L-cysteine analogs also have been reported not to inhibitL-cysteine desulfhydrase isolated from Salmonella typhimurium (4,
Table III. [35SJH2S Emission from Detached Young Cucumber LeavesTreated with [5SJL-Cysteine
Each detached cucumber leaf was treated in the leaf chamber with 10ml 10 mM L-cysteine containing 30.8 AiCi of [35SJL-cysteine, ie. 0.308 ,uCi,umol', in the light (I mw cm-2) or dark. H2S emitted was caught in a zincacetate trap as ZnS. Radioactivity and amount of sulfide in aliquots of thetrap solution were determined as volatile S trapped at the indicated timesby liquid scintillation counting and the colorimetric method for sulfide.ZnS precipitate was collected by centrifugation of the trap solution.Radioactivity and amount of sulfide in ZnS precipitate were determinedto calculate specific activity of [`*S]H2S emitted.
Time Volatile S Trapped Specific ActivityCollected H2S(A) 35S(B) B/A of H2S Emitted
h nmol (g nCfes nCi nimth fresh wt)- t(tfresh nmol-' nCi nmol-
Light 0 to 3 67 19.6 0.293 0.2020 to 6 379 77.3 0.204 0.2330 to 9 769 205 0.267 0.283
Dark 0 to 9 787 213 0.271 0.275
Table IV. Distribution of3Sfrom [35SJL-CysteineDetached cucumber leaves treated with ['SlL-cysteine in Table III were
used for extraction, fractionation, and determination of 'S compounds.Detailed procedures were described in the text.
35S X 10-3
Compound Light Dark
3h 6h 9h (9h)
cpm (gfresh wt)-'Cysteine and cystine 686 1,809 2,147 1,605H2S + S2- 195 568 970 875H2S emitted (43)- (170) (451) (469)s2- (152) (398) (519) (406)
S032- 23 121 167 127SO42- 114 239 374 299Met 87 173 200 282GSH 158 308 360 274Lipids 50 86 70 100Others (soluble) 691 1,798 2,618 1,176Precipitate 10 18 26 20
Total 2,015 5,120 6,930 4,756
H2S/total 2.1 3.3 6.5 9.9a Numbers in parentheses are subfractions of the H2S + S2- fraction.
EIC
ECII~
Time, h
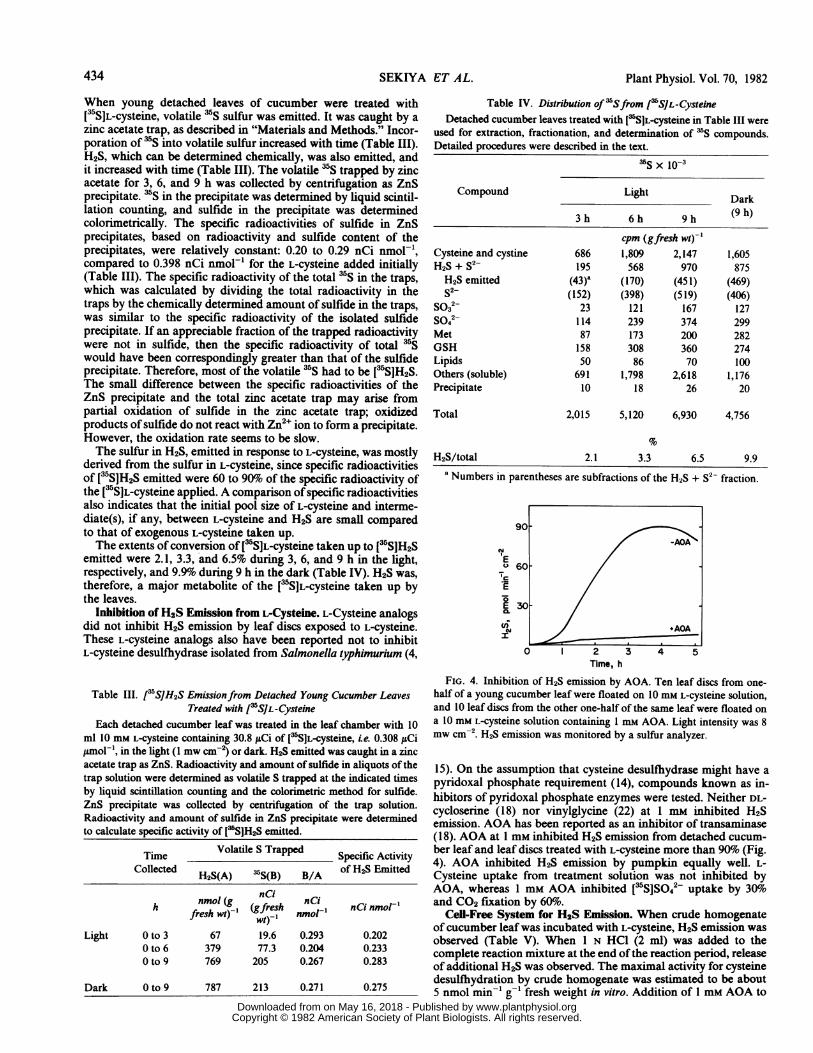
FIG. 4. Inhibition of H2S emission by AOA. Ten leaf discs from one-half of a young cucumber leaf were floated on 10 mM L-cysteine solution,and 10 leaf discs from the other one-half of the same leaf were floated ona 10 mM L-cysteine solution containing I mm AOA. Light intensity was 8mw cm-'. H2S emission was monitored by a sulfur analyzer.
15). On the assumption that cysteine desulfhydrase might have apyridoxal phosphate requirement (14), compounds known as in-hibitors of pyridoxal phosphate enzymes were tested. Neither DL-cycloserine (18) nor vinylglycine (22) at 1 mm inhibited H2Semission. AOA has been reported as an inhibitor of transaminase(18). AOA at 1 mm inhibited H2S emission from detached cucum-ber leaf and leaf discs treated with L-cysteine more than 90% (Fig.4). AOA inhibited H2S emission by pumpkin equally well. L-Cysteine uptake from treatment solution was not inhibited byAOA, whereas 1 mm AOA inhibited [5SJS042 uptake by 30%oand CO2 fixation by 60%o.
Cell-Free System for H2S Emission. When crude homogenateof cucumber leaf was incubated with L-cysteine, H2S emission wasobserved (Table V). When 1 N HCI (2 ml) was added to thecomplete reaction mixture at the end of the reaction period, releaseof additional H2S was observed. The maximal activity for cysteinedesulfhydration by crude homogenate was estimated to be about5 nmol min-' g9' fresh weight in vitro. Addition of 1 mm AOA to
434 SEKIYA ET AL.
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

EMISSION OF HYDROGEN SULFIDE FROM CYSTEINE
Table V. H2S Emission in Response to L-Cysteine in Cell-Free SystemYoung cucumber leaf was homogenized with 10 volumes of 100 mm
Tris-HCl buffer (pH 7.5), containing 1% (w/v) ascorbic acid. The homog-enate was filtered through cheesecloth. Complete reaction mixture con-sisted of 8 ml crude homogenate and 2 ml 100 mM L-cysteine. Incubationlasted 2 min, and H2S released to the atmosphere was monitored by asulfur analyzer. The activity presented is an average emission rate for first2 min. The data presented represent about 4% of the sulfide formed in thereaction mixture. The rest remains ionized and dissolved at pH 7.5 butcan be released by acidification of the mixture.
Reaction Mixture H2S Emission
nmol min-' g- 'fresh wtComplete 0.215 (l00)O+AOA, I mm 0.013 (6)-Enzyme 0.022 (10)-L-Cysteine 0.018 (8)
a Numbers in parentheses are rates as percentage of rate with a completereaction mixture.
the reaction mixture caused complete inhibition of H2S emission.The enzymic activity for H2S emission was found in the superna-tant after centrifugation of the crude homogenate of cucumberand pumpkin leaves at 15,000g for 10 mi.
Distribution of 35S from 135S L-Cysteine. After volatile sulfurwas trapped for 3, 6, and 9 h, 3 S compounds in each leaf wereextracted, fractionated, and determined. The uptake of [35SJL-cysteine increased with the time (Table IV). Thirty to 35% of MStaken up was found as cysteine and cystine in the light or in thedark (Table IV). More than 10%Yo of S taken up was found asinternal sulfide (HS-, S2-) and volatile H2S during 3, 6, and 9 hin the light. The 5S in H2S, as the percentage of 35S in total sulfide(H2S and S2-), increased from 22% at 3 h to 46% at 9 h in thelight, in addition to an increase in 35S in internal sulfide. Thisindicates accumulation of soluble sulfide prior to emission of H2S.At 9 h, H2S emitted was a major metabolite of [ssS]L-cysteine, aswas internal sulfide. It was also notable that appreciable amountsof S032- and S042- were formed. These are presumed to beproducts of sequential oxidation of sulfide. Thirty to 40o of total5S was incorporated into unidentified compounds in the ethanol-aqueous fractions.
In the dark, the [35S]H2S emitted during 9 h accounted for 10%oof the total taken up, and total sulfide accounted for 18%, com-pared to 14% in the light. The relative distribution of 35S in othercompounds in the dark was similar to that in the light at 9 h,whereas uptake of [35]L-cysteine was reduced by 30%1o in the dark.At least one-half of the sulfide formed in the dark is released asH2S.
In conclusion, sulfide, found as internal sulfide and emittedH2S, is a major metabolite of [35S]L-cysteine in green leaves in thelight and in the dark.
DISCUSSION
It is clear from the results presented that a common property ofplants is that their leaves emit volatile sulfur when exposed to L-cysteine. Based on our detailed studies of the phenomenon incucurbits, the emitted sulfur is entirely in the form of H2S.Furthermore, the sulfur of the H2S is derived from the sulfur ofthe added L-cysteine with very little dilution by sulfur from othercompounds in the plants.The process occurs in the dark. In young leaves of cucumber or
pumpkin, the initial rates of H2S emission in dark or light arevirtually identical, but, in mature leaves, the initial rate in light ismore than twice that in the dark (H. Rennenberg, unpublisheddata).
L-Cysteine desulihydrase, a pyridoxal phosphate enzyme, has
been purified from Salmonella typhimurium (14). A similar enzymeactivity was detected in extracts of the XD strain of culturedtobacco cells by Harrington and Smith (12). In both Salmonellaand tobacco cells, the enzyme was apparently induced by growthof the cells on L-cysteine. We have found the same kind of activityin extracts of cucumber and pumpkin leaves. The reaction can beinhibited by AOA, a well known inhibitor ofpyridoxal phosphate-dependent enzyme reactions (18).The activity found in leaf extracts was in the range of 5 nmol
min-' g-1 fresh weight. The maximal induced activity reported byHarrington and Smith (12) was about 50 nmol min-' g-' freshweight, assuming 5 mg protein g-' fresh weight in their prepara-tions, while the basal activity was less than 10 nmol min-' g-'fresh weight. The desulfhydrating activity level in leaf extracts iscomparable to that of ATP sulfurylase extractable from leaves ofvarious plant species (23) and to the rate at which actively growingplant material assimilates sulfur into protein. It remains to bedetermined if exposure to L-cysteine causes an increase in cysteinedesulflydrase activity, as it does in Salmonella and tobacco XDcells. Since 1 g fresh weight corresponds to about 60 cm2 of leaf,the extractable cysteine desulfhydrase activity is equivalent to 100pmol minm cm2, which is similar to the rates of H2S emissionmeasured in vitro.AOA inhibits H2S production of leaf discs and detached leaves
in response to L-cysteine by 80 to 90%, which is consistent withthe idea that the H2S formed in vivo arises by direct desulfhydra-tion of L-cysteine. The strict substrate specificity of the H2S-forming process, i.e. the lack of production of H2S in response tocompounds as closely related to L-cysteine as L-cystine or D-cysteine, or in response to structural analogs such as L-homocys-teine, or to derivatives such as S-methyl-L-cysteine and cyste-amine, also points to a direct enzymic attack on L-cysteine in vivo.The stimulation of H2S emission in response to L-cysteine by D-cysteine or S-methyl-L-cysteine suggests that the enzyme(s) re-sponsible for H2S production may be regulated, ie. induced oractivated by sulfur-containing compounds other than L-cysteine.Another possibility is that the enzyme is less specific than itappears to be, and it may be induced/activated by S-methyl-L-cysteine and may catalyze release of CH3SH from S-methyl-L-cysteine or SH- from L-cysteine; however, the CH3SH may not beemitted. These possibilities should be considered, because S-methyl-cysteine induces cysteine desulfhydrase activity in Chlo-rella (Schmidt, unpublished observations).We have shown previously that young cucumber leaves have a
much greater potential to produce H2S than do mature leaves inresponse to SO2 (27) or SO4 - (unpublshed data). It was, therefore,of interest to determine the L-cysteine-dependent rate of emissionof H2S as a function of leaf age, as part of an evaluation of thehypothesis that H2S production from s042- or SO2 was via L-cysteine desulfhydration. We found that the rates of cysteine-dependent H2S emission did, indeed, decrease with leaf age butby less than a factor of two from the youngest leaf to the oldestleaf. This difference is insufficient to account for the 10- to 100-fold difference in H2S emission rates which we have observedbetween young and mature leaves exposed to SO2 or s042.
It became evident, from the experiments in which ['SJL-cysteinewas used, that some of the [35S]sulfide released by desulfhydrationwas oxidized to [35S132- and [5SISO42- . Because plants normallyreduce So42- and synthesize L-cysteine from the reduced sulfur,it appears that there exists a seemingly futile sulfur cycle in leaves,as in the case of the XD strain of cultured tobacco cells (12). Theflux of sulfur through this cycle under normal conditions, ie. inthe absence of a high concentration of exogenous L-cysteine,cannot be estimated from the data available so far. However, it isnoteworthy that there is sufficient cysteine desulfhydrase activityin leaves to divert about as much sulfur through the proposedfutile sulfur cycle as there is ATP sulfurylase to initiate the
435
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 70, 1982
assimilation of sulfate into protein. It is conceivable, therefore,that, under certain circumstances (eg. a low rate of activation ofL-cysteine to L-cysteinyl tRNACYS, so that L-cysteine concentrationrises), most of the sulfur reduced in a leaf would be recycled ratherthan incorporated into protein. However, one would expect thissituation to be short-lived, because it has been shown that H2Scauses a rapid decrease in APS sulfotransferase, the second en-zymic step of sulfate reduction, in Lemna (3, 34) and Phaseolus(36). Also, L-cysteine, itself, inhibits or represses sulfate assimila-tion into L-cysteine (13, 20, 24, 25, 33).
Exposure of leaf discs or petioles of detached leaves to mmconcentrations of L-cysteine is a somewhat artificial situation, soone must consider the possibility that cysteine-stimulated H2Semission is entirely a man-made artifact. Since tissue exposed to10 mMt [a5S]L-cysteine contains about 1 ,umol [a5S]L-cysteine g'fresh weight, we can estimate that the internal concentration ofL-cysteine is about 10o ofthe external concentration. The emissioncan be detected when the leaf tissue is exposed to as little as 1 mmL-cysteine. Therefore, we detected H2S emission when the internalL-cysteine concentration was raised to about 100 JIM. L-Cysteinepools are in the range of 10 to 20 ,UM in Lemna tissues (9), 30 to 70ZlM in cultured tobacco XD cells (29), and 40 liM in potato tuber(16). Therefore, it is not unreasonable to propose that H2S mightbe produced spontaneously and emitted at a low rate undercircumstances in which the internal L-cysteine level increasesperhaps 2- to 3-fold (8, 21). At lower concentrations of L-cysteine,it is conceivable that desulflydration could occur but with suchefficient recycling of S2- that H2S emission would not be detect-able.
Acknowledgments-We thank H. Rennenberg for suggesting the use of leaf discsfrom halves of a single leaf to get around variability among leaves. We also thank N.Amrhein for a useful discussion on AOA. L. LeCureux and T. Shimei providedexcellent technical assistance.
LITERATURE CITED
1. ANDERSON JW 1980 Assimilation of inorganic sulfate into cysteine. In BJ Miflin,ed, The Biochemistry of Plants, Vol 5. Academic Press, New York, pp 203-223
2. BRESSAN RA, L LECUREUX, LG WILSON, P FILNER 1979 Emission of ethyleneand ethane by leaf tissue exposed to injurious concentration of sulfur dioxideor bisulfite ion. Plant Physiol 63: 924-930
3. BRUNOLD C, A SCHMIDT 1976 Regulation of adenosine-5'-phosphosulfate sulfo-transferase activity by H2S in Lemna minor L. Planta 133: 85-88
4. COLLINS JM, KJ MONTY 1973 The cysteine desulfhydrase of Salmonella typhi-murium. Kinetic and catalytic properties. J Biol Chem 248: 5943-5949
5. COLLINS JM, A WALLENSTEIN, KJ MONTY 1973 Regulatory features of thecysteine desulfhydrase of Salmonella typhimurium. Biochim Biophys Acta 313:156-162
6. DE CoRMIs L 1968 Degagement d'hydrogene sulfure par des plantes soumises aune atmosphere contenant de lanhydride sulfureux. C R Acad Sci Ser D 266:683-685
7. ELLIS RJ 1966 Sulfur metabolism: the usefulness of N-ethylmaleimide. Nature(Lond) 211: 1266-1268
8. ESTERBAUER H, D GRILL 1978 Seasonal variation of glutathione and glutathionereductase in needles of Picea abies. Plant Physiol. 61: 119-121
9. GIOVANELLI J, SH MUDD, AH DATKO 1980 Sulfur amino acids in plants. In BJMiflin, ed, The Biochemistry of Plants, Vol 5. Academic Press, New York, pp
453-50510. GUSTAFSSON L 1960 Determination of ultramicro amounts of sulphate as meth-
ylene blue. 1. The colour reaction. Talanta 4: 227-23511. GUSTAFSSON L 1960 Determination of ultramicro amounts of sulphate as meth-
ylene blue. II. The reduction. Talanta 4: 236-24312. HARRINGTON HM, IK SMITH 1980 Cysteine metabolism in cultured tobacco cells.
Plant Physiol 65: 151-15513. HART JW, P FILNER 1969 Regulation ofsulfate uptake by amino acids in cultured
tobacco cells. Plant Physiol 44: 1253-125914. KREDICH NM, BS KEENAN, LJ FOOTE 1972 The purification and subunit
structure of cysteine desulfhydrase from Salmonella typhimurium. J Biol Chem247: 7157-7162
15. KREDICH NM, LJ FOOTE, BS KEENAN 1973 The stoichiometry and kinetics ofthe inducible cysteine desulfhydrase from Salmonella typhimurium. J BiolChem 248: 6187-6196
16. MAGGIONI A, F RENOSTO 1977 Cysteine and methionine regulation of sulfateuptake in potato tuber discs (Solanum tuberosum). Physiol Plant 39: 143-147
17. MEINERT MC, DP DELMER 1977 Changes in biochemical composition of the cellwall of the cotton fiber during development. Plant Physiol 59: 1088-1097
18. MEISTER A 1965 Biochemistry of the Amino Acids, Ed 2, Vol 1. Academic Press,New York, p 411
19. MILLER JM, EE CONN 1980 Metabolism of hydrogen cyanide by higher plants.Plant Physiol 65: 1199-1202
20. NG BH, JW ANDERSON 1978 Chloroplast cysteine synthases of Trifolium repensand Pisum sativum. Phytochemistry 17: 879-885
21. RAYBOLILD C, MH UNSWORrH, PJ GREGORY 1977 Sources of sulphur in raincollected below a wheat canopy. Nature (Lond) 267: 146-147
22. RANDO RR 1974 Irreversible inhibition of aspartate aminotransferase by 2-amino-3-butenoic acid. Biochemistry 13: 3859-3863
23. REUVENY Z, P FILNER 1976 A new assay for ATP sulfurylase based on differentialsolubility of the sodium salts of adenosine 5'-phosphosulfate and sulfate. AnalBiochem 75: 410-428
24. REUVENY Z, P FILNER 1977 Regulation of adenosine triphosphate sulfurylase incultured tobacco cells. Effect of sulfur and nitrogen sources on the formationand decay of the enzyme. J Biol Chem 252: 1858-1864
25. SCHMIDT A 1973 Sulfate reduction in a cell-free system of Chlorella. Theferredoxin dependent reduction of a protein-bound intermediate by a thiosul-fonate reductase. Arch Mikrobiol 93: 29-52
26. SCHMIDT A 1979 Photosynthetic assimilation of sulfur compounds. In M Gibbs,E Latzko, eds, Encyclopedia of Plant Physiology, Vol 6. Springer-Verlag, NewYork, pp 481-496
27. SEKIYA J, LG WILSON, P FILNER 1980 Positive correlation between H2S emissionand SO2 resistance in cucumber. Plant Physiol 65: S-74
28. SMITH IK, JF THOMPSON 1971 Purification and characterization of L-serinetransacetylase and O-acetyl-L-serine sulfhydrylase from kidney bean seedlings(Phaseolus vulgaris). Biochim Biophys Acta 227: 288-295
29. SMITH IK 1975 Sulfate transport in cultured tobacco cells. Plant Physiol 55:303-307
30. SMITH 1K 1980 Regulation of sulfate assimilation in tobacco cells. Effect ofnitrogen and sulfur nutrition on sulfate permease and o-acetylserine sulfhydry-lase. Plant Physiol 66: 877-883
31. SMYTH DG, A NAGAMATSU, JS FRUTON 1960 Some reactions of N-ethylmaleim-ide. J Am Chem Soc 82: 4600-4604
32. SPALENY J 1977 Sulfate transformation to hydrogen sulfide in spruce seedlings.Plant Soil 48: 557-563
33. TRINITY PM, C VON ARB, D RHODES, T YAMAYA, A SCHMIDT, P FILNER 1980Regulation of APS sulfotransferase and other enzymes of S and N metabolismin cultured tobacco cells. Plant Physiol 65: S-16
34. VON ARB C, C BRUNOLD 1980 Analysis of the regulation of adenosine-5'-phosphosulfate sulfotransferase activity in Lemna minor L. using '5N-densitylabeling. Planta 149: 355-360
35. WILSON LG, RA BRESSAN, P FILNER 1978 Light-dependent emission ofhydrogensulfide from plants. Plant Physiol 61: 184-189
36. WYss H-R, C BRUNOLD 1979 Regulation of adenosine 5'-phosphosulfate sulfo-transferase activity by H2S and cysteine in primary leaves of Phaseolus vulgarisL. Planta 147: 37-42
436 SEKIYA ET AL.
www.plantphysiol.orgon May 16, 2018 - Published by Downloaded from Copyright © 1982 American Society of Plant Biologists. All rights reserved.





























