Embed Size (px)
Citation preview

1097
Lymphatic Pumping in Response to Changesin Transmural Pressure Is Modulated by
Erythrolysate/HemoglobinRadu M. Elias, George Wandolo, Naren S. Ranadive,
John Eisenhoffer, and Miles G. Johnston
Red blood cells and lysate products (erythrolysate) are observed consistently in lymph drainingacute and chronic inflammatory reactions and from tissues subjected to trauma or surgicalprocedures. Using hemoglobin as a marker for erythrolysate, we have measured hemoglobin inlymph up to the 10-6M range in a number of pathophysiological states. Data demonstrate thaterythrolysate alters the pumping characteristics of lymphatic vessels. To test the effects oferythrolysate on lymphatic pumping, bovine lymphatics were suspended in an organ bathpreparation with the vessels cannulated at both inflow and outflow ends. By raising the heightsof the Krebs reservoir and the outflow catheters appropriately, a transmural pressure thatstimulated pumping activity could be applied to the vessels. With a fixed transmural pressure
of 6 cm H20 applied to the ducts, sheep erythrolysate depressed pumping activity between 40%oand 10099 with dilutions containing between 10`8 and 10` M hemoglobin. Although the activeprinciple in the red blood cells has not been characterized, evidence from precipitationpurification experiments suggests that hemoglobin is an important component. Once sup-pressed, pumping could be restored in many but not all vessels (often to control levels) byelevating the distending pressure above 6 cm H20. The relation between transmural pressureand fluid pumping is expressed as a bell-shaped curve, with pumping increasing up to a peakpressure (usually 8 cm H20) and declining at pressures above this level. By comparingpressure/flow curves, we were able to ascertain that hemoglobin shifted the lymphatic functioncurve to the right and, on average, reduced the maximum pumping capability of the vessels. Wespeculate that the presence of erythrolysate/hemoglobin in lymph may modulate the ability oflymphatic vessels to drain liquid and protein from the tissue spaces. (Circulation Research1990;67:1097-1106)
R ed blood cells (RBCs) are observed in lymphafter vascular damage associated with in-flammatory reactions, surgical procedures,
or traumatic injury; however, this has not beenconsidered to have any major significance in terms oflymphatic physiology. A number of recent observa-tions have encouraged us to look into this issue inmore detail. The ability of the lymphatics to contractand pump liquid and protein from the tissue spacesback to the bloodstream has been the major focus of
From the Departments of Pathology and Surgery, University ofToronto, and the Trauma Research Program, Sunnybrook MedicalCentre, Toronto, Canada.Supported by the Canadian Foundation for Ileitis and Colitis
Grant (to M.G.J.) and a grant from the Medical Research Councilof Canada (grant MA-7925 to M.G.J.).Address for correspondence: Dr. Miles G. Johnston, Depart-
ment of Pathology, Medical Sciences Building, 1 Kings CollegeCircle, University of Toronto, Toronto, Ontario, Canada M5S1A8.Received January 24, 1990; accepted July 2, 1990.
our studies over the past few years. Many factors arecapable of altering the contractile properties of thesevessels, including neurohumoral and other agents.1'2By isolating lymphatics in vivo from lymph input andapplying transmural pressures to the ducts, severalgroups have been able to study lymphatic pumpingwithout the complications of variable lymph inputs.3-6These studies suggest that the host may have theability to modulate pumping and, in this way, regu-late the drainage of liquid and protein from the tissuespaces. Any factor that alters pumping could conceiv-ably facilitate or interfere with this function. It is inthis regard that we first became interested in eryth-rolysate/hemoglobin.
In previous studies, we observed that systemicendotoxin administration to sheep inhibited lym-phatic pumping,5 and we obtained evidence thatsuggested a lymph-borne factor was at least partiallyresponsible for this effect.6 The unknown factor orfactors appeared to reduce the sensitivity of the
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

1098 Circulation Research Vol 67, No 5, November 1990
vessels to changes in transmural pressure. BecauseRBCs and hemoglobin often were observed in lymphcollected in these experiments, we began to investi-gate whether these elements could be contributing tothis effect. There are numerous reports in the liter-ature of the effects of erythrolysate on vascularsmooth muscle7-11 and cardiac muscle,12'3 and insome cases, it would appear that hemoglobin is theactive principle. Preliminary investigation revealedthat erythrolysate prepared from sheep RBCs had avery similar effect to that of the endotoxin-inducedunknown factor when tested on sheep vessels in vivoor on bovine vessels in an organ bath preparation.The purpose of this study was to examine in more
detail the effects of erythrolysate/hemoglobin onlymphatic pumping using the in vitro preparation.This included studying the dose-response effect ofhemoglobin and determining its impact on the rela-tion between transmural pressure and pumping ac-tivity. In addition, we have modified existing spectro-photometric techniques to permit assessment ofhemoglobin concentrations in lymph.
Materials and MethodsCollection of Bovine LymphaticsThe lymphatic vessels were obtained from bovine
mesentery 7-10 minutes after the cattle were slaugh-tered. The mesenteric lymph nodes were injectedwith Evans blue dye (0.1% in physiological phos-phate buffered saline) to facilitate vessel identifica-tion. The larger vessels with diameters of 2 mm ormore were carefully dissected and immersed in ster-ile saline solution (0.9% NaCl) at room temperature.After transport back to the laboratory, the vesselswere cannulated at both ends using polyethylenetubing (Adams PE 160, i.d. 1.14 mm, o.d. 1.57 mm;Intermedic Clay Adams, Parsippany, N.J.). As a wayof providing some standardization, lengths of vesselswere cut that contained a minimum of four valves.After cannulation, the vessels were infusion testedfor leaks with a 0.01% Evans blue solution, distend-ing them with a transmural pressure of 12 cm H2O.Vessels found to leak were discarded.
All cannulated vessels then were transferred to acustom-made water-jacketed organ bath14 and im-mersed in Krebs solution (millimoles per liter: NaCl120.0, KCl 5.9, MgSO4 1.2, NaH2P04 1.2, CaCl2 2.5,NaHCO3 15.0, glucose, 11.0). The Krebs was circu-lated continuously through the inner bath by a dual-channel peristaltic pump (MasterFlex Pump, Cole-Parmer Instrument Co., Chicago), maintaining a flowof 6.7 1/hr. The pH was maintained at 7.38±0.02 bycontinuous aeration with a mixture of 95% oxygenand 5% CO2 (Accumet model 810, pH meter, FisherScientific Co., Pittsburgh). The solution temperaturewas kept constant at 38+0.20 C using a circulatingwater bath (12/TE- 8D, Techne Inc., Princeton, N.J.).The inflow catheter to the lymphatic was connectedto a valve that in turn was connected to a commonfluid reservoir filled with Krebs solution (10 ml
volume). A four-channel peristaltic pump (BuchlerInstruments, Fort Lee, N.J.) was used to refresh theKrebs solution and to maintain a constant level offluid in the supply reservoir. The flow rate was set at10 ml/min.The reservoir and the outflow catheters were
mounted on a common adjustable stand. As soon asthe vessels were immersed in the Krebs solution, asmall hydrostatic gradient of 2-3 cm H2O was ap-plied for 15-20 minutes while the vessels equili-brated. After this period, the hydrostatic pressuregradient was removed. By raising the height of boththe reservoir and the outflow catheter relative to thecannulated vessel, but keeping the heights of bothequal to each other (i.e., no net driving pressure), atransmural pressure was applied to the ducts. Thesurface of the Krebs in the organ bath was made thezero reference level. All vessels were allowed toequilibrate for another 1-hour period with a trans-mural pressure of 6-8 cm H2O applied to the vessels.In the majority of cases, this procedure elicitedspontaneous pumping activity. Those vessels that didnot demonstrate spontaneous contractile activitywere discarded.
Collection of Sheep Blood and Preparation ofRedBlood Cell and Platelet Concentrate
For the preparation of the RBC suspension, ovineblood was collected into acid citrate dextrose (triso-dium citrate, 25 g; citric acid, 15 g; dextrose, 20 g,dissolved in 1 1 distilled water; 6:1, vol/vol).'5 Theblood was centrifuged at 200g for 30 minutes, and theplatelet-rich plasma and the buffy coat were aspi-rated. The remaining erythrocyte concentrate wasdiluted with phosphate buffered saline and recentri-fuged at 200g for another 30 minutes. Again, theplatelet-rich plasma and buffy coat were aspirated.The cells were diluted with phosphate buffered salineto 50 ml, spun a third time at 1,000g for 10 minutes,and the supernatant and buffy coat were aspirated toremove most of the remaining leukocytes. One finalphosphate buffered saline wash and spin was used toremove any plasma still remaining in the RBC con-centrate. These procedures removed approximately50% of the platelets. To remove the remainingplatelets, the cell suspension was passed through acotton wool column (Leukopak Filter, Fenwal Labo-ratories, Morton Grove, Ill.). Cotton wool (0.5 g) wascompressed in the barrel of a 10 ml syringe. The filtercolumn was rinsed with 100 ml of saline, and theerythrocyte concentrate was filtered through the col-umn. After this filtration step, 95-99.5% of theplatelets in the original collection had been removed.
Platelets were removed from the pooled platelet-rich plasma obtained during the preparation of theRBCs. The pooled platelet-rich plasma was spun andwashed three times at 200g for 30 minutes to removeRBCs and white blood cell contamination. The RBC-and white blood cell-free platelet-rich plasma thenwas spun for 20 minutes at 1,000g.
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

Elias et al Erythrolysate and Lymphatic Pumping 1099
Preparation of Sheep ErythrolysateRBCs were lysed by freezing in dry ice and thawing
in a 370 C water bath. After centrifugation at 1,000gfor 30 minutes, the supernatant was assessed forhemoglobin content using the cyanmethemoglobinassay (Sigma Chemical Co., St. Louis). In someexperiments, the supernatant was dialyzed in a dia-lyzing membrane with a cutoff of 10,000 MW againsta phospate buffered solution (2.8 M P04, pH 6.8).When the volume of the dialysis bag was reduced toone third, the bag was cut and the contents werecollected in a flask. The same phosphate bufferedsolution (2.8 M) was added slowly until precipitationoccurred.'6 The hemoglobin precipitate was spuntwice at 10,000g for 30 minutes. The samples wereassessed for hemoglobin content and an absorptionspectrum obtained between 400 and 650 nm.
It was important to determine if any plateletproducts (from potential platelet contamination ofthe RBC preparation) could be contributing to theeffects on lymphatics. To achieve this, we treated theplatelets the same way as we treated the RBCs; thatis, they were frozen and thawed, the lysate wascentrifuged at 1,000g for 30 minutes, and the lysatewas tested for effects in the pumping preparation.
Quantitation ofHemoglobin in Lymph SamplesThe assay is based on spectrophotometric methods
in current use for measuring hemoglobin concentra-tions in blood, plasma, urine, and cerebrospinal fluid(see References 17 and 18). The major problem inmeasuring hemoglobin in lymph is that it is essentialto consider the other proteins (mainly albumin), asthese absorb to some extent at the wavelength we useto measure hemoglobin. Also, in some lymph sam-ples, for example intestinal lymph, a special problemoccurs in that lipids cause dispersion of light. Toreduce this latter effect, lymph was delipidated withthe addition of butanol/diisopropyl ether (40:60,vol/vol).19 Twenty milliliters of the solvent mixturewas added to 10 ml of lymph. The mixture wasshaken and centrifuged at 2,500g for 16 minutes toseparate the aqueous and organic phases while leav-ing the lipids layered between the phases. The aque-ous phase was removed and filtered through What-man filter paper number 1 (Whatman Inc., Clifton,N.J.). Absorbance due to proteins was eliminatedusing a 4 g% albumin solution as a blank for theassay. This value represents the average proteinconcentration in sheep intestinal lymph. We used awavelength of 410 nm, as this was the intersection ofthe absorbance of oxyhemoglobin and methemoglo-bin, both of which could be present in our lymphcollections.20 Even after delipidation and deprotein-ation with 10% trichloroacetic acid, which also wouldremove any hemoglobin present, the sample still hada small absorbance (in the vicinity of 0.01). In allassays, this value was determined and subtractedfrom the overall absorbance. To prepare the stan-dard curve, hemoglobin was isolated from sheep
blood as described above, and its concentration wasdetermined by the Drabkins method using the Sigmakit. The hemoglobin was added to saline containing 4g% fat-free albumin, and the absorbance values wereread at 410 nm against the 4% albumin blank. Thisprocess was repeated four times, and the average ofall absorbances was used to prepare the standardcurve. The lowest limit of detection was 1x 10 ` M.
Collection ofLymph Samples forHemoglobin Determninations
Female sheep weighing 25-35 kg were used. Plas-tic catheters were inserted into the efferent lymphat-ics 10-15 cm downstream from the terminal mesen-teric lymph node as described previously.5,6 Thecatheter was externalized, and lymph was collectedinto sterile plastic bottles attached to the animal'sflank. Lymph was collected continuously after sur-gery, and in some animals, endotoxin (33.0 ,ug/kg;055:B5, List Biological Laboratories, Inc., Camp-bell, Calif.) was infused intravenously 4-5 days aftercatheter placement.
Experimental ProtocolsFixed transmural pressure experiments. After equili-
bration of the bovine lymphatics, transmural pres-sures were reduced to zero. The transmural pressureand fluid pumping relation was obtained by elevatingthe pressure in increments of 2 cm H20 from 0 to 14cm H20. The vessels were allowed to pump for 10minutes at each pressure with an equilibration timeof 2 minutes between each pressure level. Pumpingactivity was assessed by measuring the weight of theliquid collected at the end of each 10-minute inter-val. After the maximum distending value wasreached, the transmural pressure was decreasedslowly over 10-15 minutes to a distending pressurethat induced 75% of maximum pumping activity. Theaverage distending pressure was in the range of 6 cmH20. In these experiments, transmural pressureswere fixed at 6 cm H20, and the vessels were allowedto pump for 1 hour. At this point, the input reservoirwas switched to that containing erythrolysate orplatelet lysate, and the pumping was monitored foranother 2 hours. To study the dose-response effect ofthe erythrolysate (in terms of hemoglobin concentra-tions), the same protocol was used except that theinput reservoir was switched to a higher concentra-tion at 1-hour intervals. All flows were averaged over10-minute intervals and were expressed as a percent-age of the mean pumping activity generated in thecontrol period. Mean±SEM of the flows was plottedagainst time.
Variable transmural pressure experiments. To deter-mine the ability of the lymphatics to respond totransmural pressure changes in the presence oferythrolysate, two transmural pressure and fluidpumping curves were generated for each vessel, onewith Krebs and one with Krebs plus erythrolysate/hemoglobin. In some experiments, two curves weregenerated with Krebs to be certain that the genera-
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
1100 Circulation Research Vol 67, No 5, November 1990
tion of a second transmural pressure/flow curve froma given vessel had no deleterious effect on thepumping mechanism. All flows were averaged over10-minute intervals and were expressed as a percent-age of the maximum pumping activity generated inthe first control curve. Mean+SEM of the flows ateach pressure was plotted against the distendingpressure values.
Analysis of DataThe graphs illustrate results from a minimum of
three experiments conducted on separate days withvessels obtained from six or more animals. Data wereassessed using analysis of variance and the Duncan'smultiple range test. The analysis of variance that weuse is based on the Statistical Analysis System,General Linear Model procedure, in which effects ofgroups are tested against the variability of time ortransmural pressure within the group. We inter-preted p<0.05 as significant.
ResultsResponse of Lymphatic Vessels toTransmural PressureThe relation between transmural pressure and
pumping activity in this model system has beendescribed in our earlier publications.14'2' Once atransmural pressure was applied to the vessels, themajority of ducts (approximately 80% in this study)pumped fluid spontaneously. The average maximumpumping activity occurred at a distending pressure of8 cm H2O, and as pressures were increased beyondthis point, pumping declined. Maximum flow ratesranged from 0.35 to 1.4 ml/min. To study the effectsof erythrolysate on steady-state pumping, the trans-mural pressure was fixed at 6 cm H20 and kept at thislevel for the duration of the experiment. This pres-sure level represented approximately 75% of thepressure that induced maximum pumping.
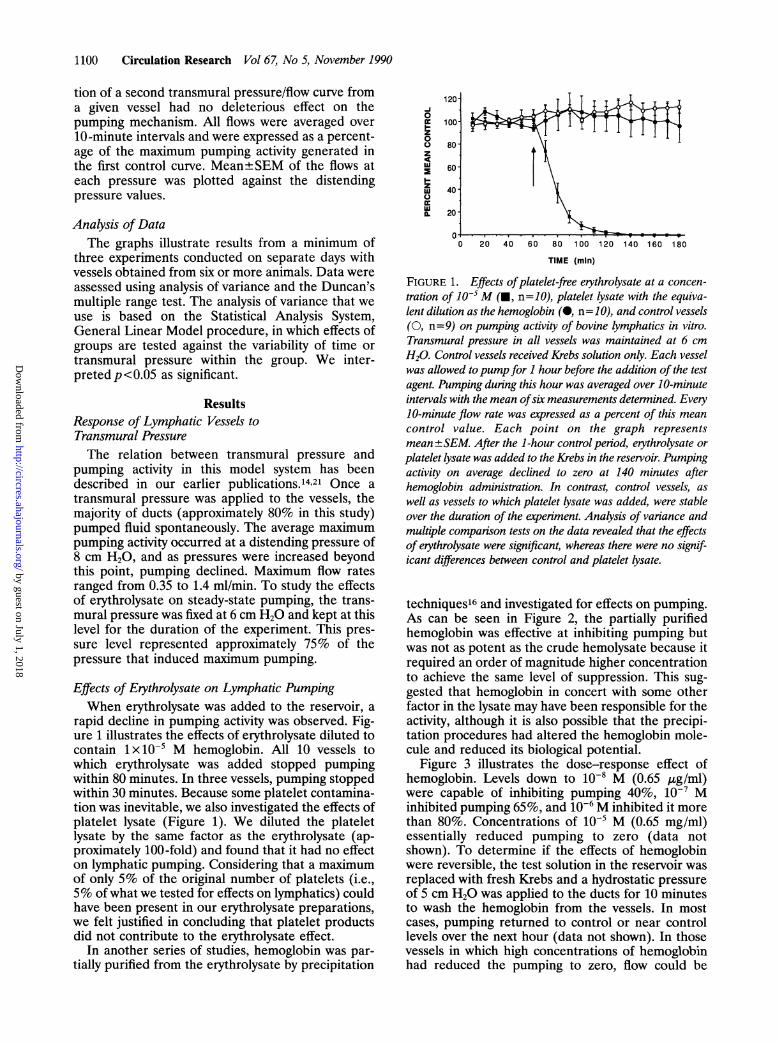
Effects of Erythrolysate on Lymphatic PumpingWhen erythrolysate was added to the reservoir, a
rapid decline in pumping activity was observed. Fig-ure 1 illustrates the effects of erythrolysate diluted tocontain lx 10-5 M hemoglobin. All 10 vessels towhich erythrolysate was added stopped pumpingwithin 80 minutes. In three vessels, pumping stoppedwithin 30 minutes. Because some platelet contamina-tion was inevitable, we also investigated the effects ofplatelet lysate (Figure 1). We diluted the plateletlysate by the same factor as the erythrolysate (ap-proximately 100-fold) and found that it had no effecton lymphatic pumping. Considering that a maximumof only 5% of the original number of platelets (i.e.,5% of what we tested for effects on lymphatics) couldhave been present in our erythrolysate preparations,we felt justified in concluding that platelet productsdid not contribute to the erythrolysate effect.
In another series of studies, hemoglobin was par-tially purified from the erythrolysate by precipitation
0x 100z0c) 80-zW 60
,, 4040
CL 20
0-0 20 40 60 80 100 120 140 160 180
TIME (min)
FIGURE 1. Effects ofplatelet-free erythrolysate at a concen-tration of 10-5M (U, n=10), platelet lysate with the equiva-lent dilution as the hemoglobin (@, n=10), and control vessels(0, n=9) on pumping activity of bovine lymphatics in vitro.Transmural pressure in all vessels was maintained at 6 cmH20. Control vessels received Krebs solution only. Each vesselwas allowed topump for 1 hour before the addition of the testagent. Pumping during this hour was averaged over 10-minuteintervals with the mean ofsix measurements deternined. Every10-minute flow rate was expressed as a percent of this meancontrol value. Each point on the graph representsmean ±SEM. After the 1-hour control period, erythrolysate orplatelet lysate was added to the Krebs in the reservoir. Pumpingactivity on average declined to zero at 140 minutes afterhemoglobin administration. In contrast, control vessels, aswell as vessels to which platelet lysate was added, were stableover the duration of the experiment. Analysis of variance andmultiple comparison tests on the data revealed that the effectsof erythrolysate were significant, whereas there were no signif-icant differences between control and platelet lysate.
techniques16 and investigated for effects on pumping.As can be seen in Figure 2, the partially purifiedhemoglobin was effective at inhibiting pumping butwas not as potent as the crude hemolysate because itrequired an order of magnitude higher concentrationto achieve the same level of suppression. This sug-gested that hemoglobin in concert with some otherfactor in the lysate may have been responsible for theactivity, although it is also possible that the precipi-tation procedures had altered the hemoglobin mole-cule and reduced its biological potential.
Figure 3 illustrates the dose-response effect ofhemoglobin. Levels down to 10-8 M (0.65 ,g/ml)were capable of inhibiting pumping 40%, 10` Minhibited pumping 65%, and 10-6M inhibited it morethan 80%. Concentrations of 10-5 M (0.65 mg/ml)essentially reduced pumping to zero (data notshown). To determine if the effects of hemoglobinwere reversible, the test solution in the reservoir wasreplaced with fresh Krebs and a hydrostatic pressureof 5 cm H2O was applied to the ducts for 10 minutesto wash the hemoglobin from the vessels. In mostcases, pumping returned to control or near controllevels over the next hour (data not shown). In thosevessels in which high concentrations of hemoglobinhad reduced the pumping to zero, flow could be
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

Elias et al Erythrolysate and Lymphatic Pumping 1101
0
z
I-
zW
C.)
120-
100
80
60
40
20
0- . . .
|CONTROL
..
0 20 40 60 80 100 120 140 160 180
TIME (m;n)
FIGURE 2. Effects of erythrolysate (hemoglobin content,10-6 M) (U, n=15) or hemoglobin precipitate 10-5 M (Z,n=9) on pumping activity of bovine lymphatics in vitro.Transmural pressure in all vessels was maintained at 6 cmH20. After a 1-hour control period, erythrolysate or hemoglo-bin precipitate was added to the Krebs in the reservoir. For thecontrol period, pumping was averaged over 10-minute inter-vals, with the mean of six measurements determined. Every10-minute flow rate was expressed as a percent of the meancontrol value. Each point on the graph represents mean +
SEM. After the addition ofthe hemoglobin precipitate solutionor the erythrolysate, pumping declined to less than 20% ofcontrol (>80% inhibition) by approximately 140 minutes.Effects with erythrolysate or the precipitation purified hemo-globin were significant when compared with pumping in theabsence of these agents. Ten times more of the partiallypunified hemoglobin was required to elicit the same level ofdepression caused by the crude erythrolysate.
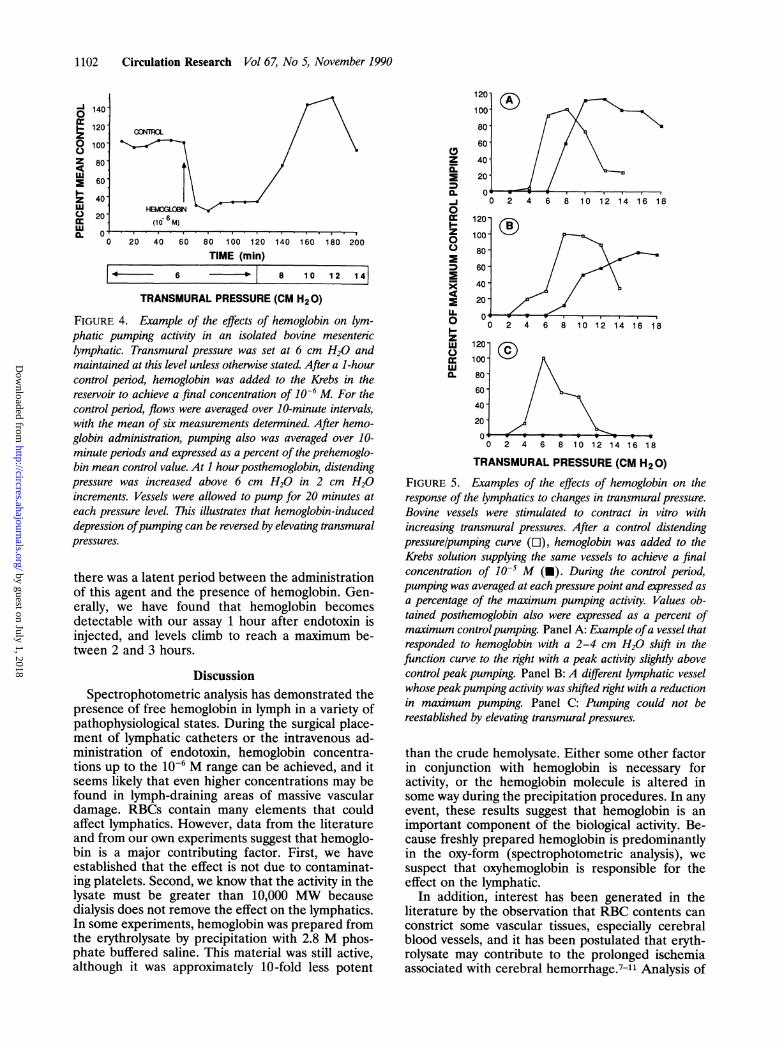
reestablished in only a few preparations. Becausethese experiments were carried out with a fixedtransmural pressure of 6 cm H20, we wanted todetermine if, after suppression by hemoglobin,pumping could be restored or elevated by increasingthe pressure above this level. As can be seen in theexample illustrated in Figure 4, increasing the trans-mural pressure in increments of 2 cm H20 increasedpumping in the presence of hemoglobin. This sug-gested that the transmural pressure and fluid pump-ing curve can be shifted to the right by hemoglobin.To study this possibility further, lymphatic functioncurves were generated in the presence and absenceof various concentrations of hemoglobin.
Effects ofHemoglobin on the Lymphatic VesselResponse to Transmural Pressure Changes
For each vessel, a transmural pressure and fluidpumping curve was obtained with Krebs in the res-ervoir and then repeated with the appropriate con-centration of hemoglobin added to the Krebs. Wepreviously determined (and it was confirmed in thisstudy) that the generation of two transmural pressureand fluid pumping curves in the same vessel had nodeleterious effect on the ducts.'4 Figure 5 demon-strates three representative examples of the effects ofhemoglobin. In all of these examples, 10- M hemo-globin was used. In panels A and B, it is clear that thehemoglobin shifted the function curve to the right. Insome cases, the maximum pumping activity was
_30
00zcc
zMLLc)MlW.
1 U -
100-
80
60
40
20
v
0
CONTROL HB= 10 M |HB =107M |HB = 10M |
4 6 80 1001201401601820 04. ..0 _
20 40 60 80 100 120 140 160 180 200 220240
TIME (min)
FIGURE 3. Effects of hemoglobin (HB) at 10-8 M, 10` M,and 10'6M on lymphatic pumping activity in vitro. Transmu-ralpressure was set at 6 cm H20 and maintained at this level.After a 1-hour control period, hemoglobin was added to theKrebs in the reservoir in increasing concentrations. During60-120 minutes, the final hemoglobin concentration was 10 8.between 120 and 180 minutes, 10' M; and between 180 and240 minutes, 10-6M During the control period, pumping wasaveraged over 10-minute intervals, with the mean of sixmeasurements determined. Every 10-minute flow rate (controland hemoglobin) was expressed as a percent of the meancontrol value. Each point on the graph represents mean +SEM(data from 12 vessels). In each of the 12 vessels in whichhemoglobin was added, pumping declined in direct relation tothe hemoglobin concentration. Hemoglobin at 10-8M causeda 40% decrease in pumping activity; at 10`" M, a 65%decrease in pumping; and at 10`6 M, greater than 80%inhibition. Analysis of variance revealed significant effects ofhemoglobin on pumping activity. Analysis with the multiplerange test indicated that pumping with each concentration ofhemoglobin was significantly different from the control (pre-hemoglobinpumping level) and significantly differentfrom theother doses of hemoglobin.
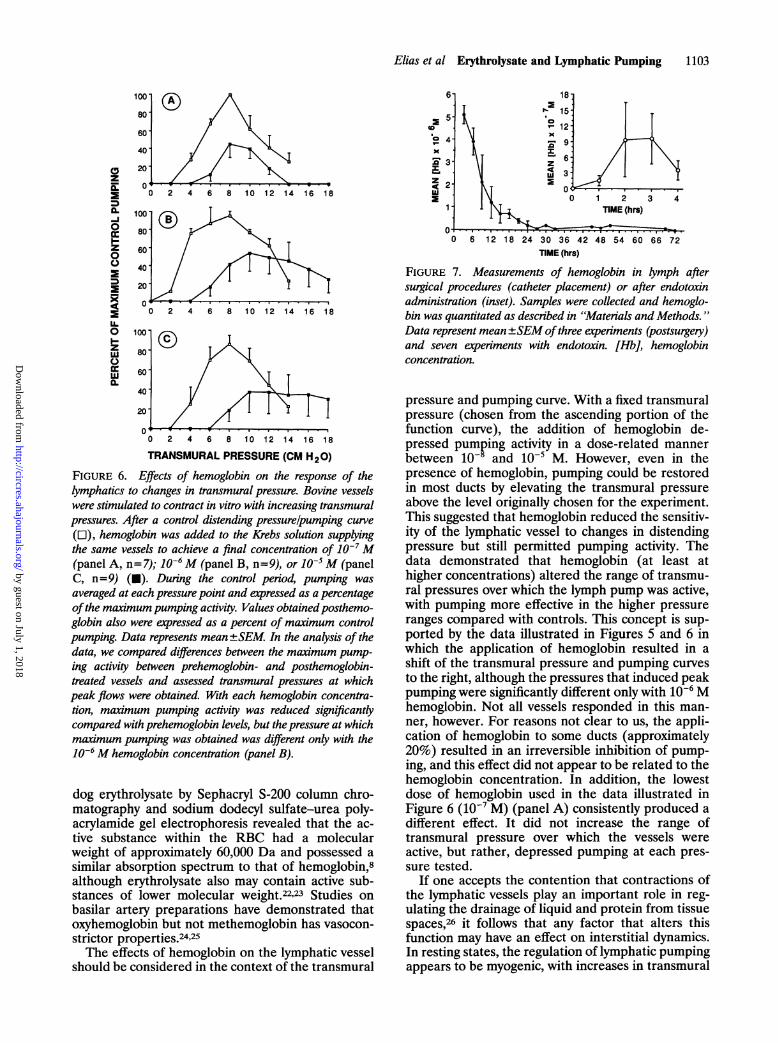
slightly more than that in the control curve (panel A),and in others, maximum pumping was somewhatless (panel B). Some vessels could not be induced topump when pressures were elevated (panel C).Taking all the results together (Figure 6), it is clearthat hemoglobin at 10', 10-6, and 10` M shifted thefunction curves to the right and reduced the maxi-mum pumping available to the vessels. With thetwo higher concentrations (10` and 10-6 M), thepumping tended to plateau beyond 10 cm H20, withflows being maintained even with high levels ofdistending pressure.
Hemoglobin Levels in LymphWe have begun to estimate hemoglobin levels in
lymph collections routinely. Figure 7 illustrates thehemoglobin levels in sheep lymph samples collectedafter the surgical placement of catheters in mesen-teric lymphatics or after endotoxin administration.High hemoglobin levels are consistently present in
lymph after surgery (into the 10-6 M range). In theanimals that received endotoxin (Figure 7, inset),
1-IL
n . . -.U r
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
1102 Circulation Research Vol 67, No 5, November 1990
-J0c-z0c)zLUWt
a.
140
120
100
80
60
40-
20
CONlROL
HEMOGLON- 6 M
0 20 40 60 80 100 120 140 160 180 200
TIME (min)- 6 - 7 8 10 12 14j
TRANSMURAL PRESSURE (CM H20)
FIGURE 4. Example of the effects of hemoglobin on lym-phatic pumping activity in an isolated bovine mesentericlymphatic. Transmural pressure was set at 6 cm H20 andmaintained at this level unless otherwise stated. After a 1-hourcontrol period, hemoglobin was added to the Krebs in thereservoir to achieve a final concentration of 10-6 M For thecontrol period, flows were averaged over 10-minute intervals,with the mean of six measurements determined. After hemo-globin administration, pumping also was averaged over 10-minute periods and expressed as a percent of the prehemoglo-bin mean control value. At 1 hourposthemoglobin, distendingpressure was increased above 6 cm H20 in 2 cm H20increments. Vessels were allowed to pump for 20 minutes ateach pressure level. This illustrates that hemoglobin-induceddepression ofpumping can be reversed by elevating transmuralpressures.
there was a latent period between the administrationof this agent and the presence of hemoglobin. Gen-erally, we have found that hemoglobin becomesdetectable with our assay 1 hour after endotoxin isinjected, and levels climb to reach a maximum be-tween 2 and 3 hours.
DiscussionSpectrophotometric analysis has demonstrated the
presence of free hemoglobin in lymph in a variety ofpathophysiological states. During the surgical place-ment of lymphatic catheters or the intravenous ad-ministration of endotoxin, hemoglobin concentra-tions up to the 10-6 M range can be achieved, and itseems likely that even higher concentrations may befound in lymph-draining areas of massive vasculardamage. RBCs contain many elements that couldaffect lymphatics. However, data from the literatureand from our own experiments suggest that hemoglo-bin is a major contributing factor. First, we haveestablished that the effect is not due to contaminat-ing platelets. Second, we know that the activity in thelysate must be greater than 10,000 MW becausedialysis does not remove the effect on the lymphatics.In some experiments, hemoglobin was prepared fromthe erythrolysate by precipitation with 2.8 M phos-phate buffered saline. This material was still active,although it was approximately 10-fold less potent
0z
E-J0mC-
00
0
zm5
X
LU0U.0
z
120
100
80
60
40
20
120
100'
80
60
40
20
2 4 6 8 10 12 14 16 18
0 2 4 6 8 10 12 14 16 18
TRANSMURAL PRESSURE (CM H20)
FIGURE 5. Examples of the effects of hemoglobin on theresponse of the lymphatics to changes in transmural pressure.Bovine vessels were stimulated to contract in vitro withincreasing transmural pressures. After a control distendingpressure/pumping curve (L), hemoglobin was added to theKrebs solution supplying the same vessels to achieve a finalconcentration of 10`5 M (M). During the control period,pumping was averaged at each pressure point and expressed asa percentage of the maximum pumping activity. Values ob-tained posthemoglobin also were expressed as a percent ofmaximum controlpumping. Panel A: Example ofa vessel thatresponded to hemoglobin with a 2-4 cm H20 shift in thefunction curve to the right with a peak activity slightly abovecontrol peak pumping. Panel B: A different lymphatic vesselwhosepeakpumping activity was shifted right with a reductionin maximum pumping. Panel C: Pumping could not bereestablished by elevating transmural pressures.
than the crude hemolysate. Either some other factorin conjunction with hemoglobin is necessary foractivity, or the hemoglobin molecule is altered insome way during the precipitation procedures. In anyevent, these results suggest that hemoglobin is animportant component of the biological activity. Be-cause freshly prepared hemoglobin is predominantlyin the oxy-form (spectrophotometric analysis), wesuspect that oxyhemoglobin is responsible for theeffect on the lymphatic.
In addition, interest has been generated in theliterature by the observation that RBC contents canconstrict some vascular tissues, especially cerebralblood vessels, and it has been postulated that eryth-rolysate may contribute to the prolonged ischemiaassociated with cerebral hemorrhage.7-1' Analysis of
n ...
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
Elias et al Erythrolysate and Lymphatic Pumping 1103
AD~~~~~~~~~~" aCD
x
zw0. . T -
2 4 6 8 lo 12 14 16 18
0 2 4 6 8 10 12 14 16 18
TRANSMURAL PRESSURE (CM H20)
FIGURE 6. Effects of hemoglobin on the response of thelymphatics to changes in transmural pressure. Bovine vesselswere stimulated to contract in vitro with increasing transmuralpressures. After a control distending pressure/pumping curve(Ol), hemoglobin was added to the Krebs solution supplyingthe same vessels to achieve a final concentration of i0`' M(panel A, n= 7); 10-6M (panel B, n=9), or 1i-O M (panelC, n=9) (M). During the control period, pumping was
averaged at each pressure point and expressed as a percentageof the maximum pumping activity. Values obtainedposthemo-globin also were exressed as a percent of maximum controlpumping. Data represents mean ±SEM. In the analysis of thedata, we compared differences between the maximum pump-
ing activity between prehemoglobin- and posthemoglobin-treated vessels and assessed transmural pressures at whichpeak flows were obtained. With each hemoglobin concentra-tion, maximum pumping activity was reduced significantlycompared with prehemoglobin levels, but the pressure at whichmaximum pumping was obtained was different only with the10-6M hemoglobin concentration (panel B).
dog erythrolysate by Sephacryl S-200 column chro-matography and sodium dodecyl sulfate-urea poly-acrylamide gel electrophoresis revealed that the ac-tive substance within the RBC had a molecularweight of approximately 60,000 Da and possessed asimilar absorption spectrum to that of hemoglobin,8although erythrolysate also may contain active sub-stances of lower molecular weight.2223 Studies onbasilar artery preparations have demonstrated thatoxyhemoglobin but not methemoglobin has vasocon-strictor properties.24,25The effects of hemoglobin on the lymphatic vessel
should be considered in the context of the transmural
18-
' 15-0_ 12-
.i' 9:
z.9
W 3-
o0 1 2 3 4
TIME (hrs)
6 12 18 24 30 36 42 48 54 60 66 72
TIME (hrs)
FIGURE 7. Measurements of hemoglobin in lymph aftersurgical procedures (catheter placement) or after endotoxinadministration (inset). Samples were collected and hemoglo-bin was quantitated as described in "Materials and Methods."Data represent mean+SEM ofthree experiments (postsurgery)and seven experiments with endotoxin. [Hb], hemoglobinconcentration.
pressure and pumping curve. With a fixed transmuralpressure (chosen from the ascending portion of thefunction curve), the addition of hemoglobin de-pressed pumping activity in a dose-related mannerbetween 10-8 and 10`- M. However, even in thepresence of hemoglobin, pumping could be restoredin most ducts by elevating the transmural pressureabove the level originally chosen for the experiment.This suggested that hemoglobin reduced the sensitiv-ity of the lymphatic vessel to changes in distendingpressure but still permitted pumping activity. Thedata demonstrated that hemoglobin (at least athigher concentrations) altered the range of transmu-ral pressures over which the lymph pump was active,with pumping more effective in the higher pressureranges compared with controls. This concept is sup-ported by the data illustrated in Figures 5 and 6 inwhich the application of hemoglobin resulted in ashift of the transmural pressure and pumping curvesto the right, although the pressures that induced peakpumping were significantly different only with 10-6 Mhemoglobin. Not all vessels responded in this man-ner, however. For reasons not clear to us, the appli-cation of hemoglobin to some ducts (approximately20%) resulted in an irreversible inhibition of pump-ing, and this effect did not appear to be related to thehemoglobin concentration. In addition, the lowestdose of hemoglobin used in the data illustrated inFigure 6 (10` M) (panel A) consistently produced adifferent effect. It did not increase the range oftransmural pressure over which the vessels were
active, but rather, depressed pumping at each pres-sure tested.
If one accepts the contention that contractions ofthe lymphatic vessels play an important role in reg-ulating the drainage of liquid and protein from tissuespaces,26 it follows that any factor that alters thisfunction may have an effect on interstitial dynamics.In resting states, the regulation of lymphatic pumpingappears to be myogenic, with increases in transmural
100i80
60
40
20C,zE
Jm0c-z0C.)U
m:
RIL0zwLC)wrW.
n v v* v *qr . v.- - -
7@
Uv
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

1104 Circulation Research Vol 67, No 5, November 1990
pressure stimulating an appropriate response. If oneargues that the transmural pressure stimulation (orequivalent forces) remain constant, one might expectthat the presence of hemoglobin in the lymph woulddepress lymph flow rates. However, this seems un-likely in vivo. Many situations in which erythrolysate/hemoglobin enters the lymph occur in conjunctionwith increases in filtration and lymph formation (forexample, endotoxin-induced increases in vascularpermeability or damage to the microcirculation).Therefore, variable increases in filtration in concertwith a less sensitive lymph pump could produceincreases, decreases, or no change in lymph flowrates, depending on the magnitude of the filtrationchanges and the concentration of hemoglobin in thelymph. In vivo, it is likely that alterations in pumpingactivity represent part of a continuum, as the lym-phatic vessel responds to progressive changes inhemoglobin concentration and other factors andchanges in tissue and transmural pressures. There-fore, the curve shift concept does not imply thatlymph flow rates will decline. By altering the sensi-tivity of the lymph pump, lymph flow is still main-tained, but an increase in flow rates after an elevationin filtration may not be as large as that occurring inthe absence of erythrolysate. The most importantconsequence of this may be that tissue water mayincrease in such a circumstance, because the normalbalance between filtration and lymphatic drainagemay be subtly altered by the presence of hemoglobin.Of course, one might turn these arguments around
and take the position that, as liquid accumulates intissue spaces, transmural pressures may be applied tothe ducts at levels beyond that required to inducepeak pumping, that is, on the downside of thetransmural pressure and pumping relation. A curveshift to the right permits the lymphatic to pump moreeffectively in the higher pressure ranges. If transmu-ral pressure increases are not imparted to the initialvessels or collecting ducts, the lymphatics simply maybecome passive conduits, and the maintenance oflymph flow would have to be solely due to extrinsicforces or to pressure gradients from the interstitiumto the initial lymphatics.The mechanism of action of hemoglobin is un-
known. There are a number of clues from the liter-ature in terms of its effect on blood vessels, and someof these may be applicable to lymphatics. For exam-ple, hemoglobin is known to generate free radicalsand induce lipid peroxidation,27 29 to affect pros-taglandin synthesis,11'30 to inhibit endothelium-de-rived relaxing factor,31,32 and to suppress vasodilatornerve activity.10'33 We believe we can exclude thenerve effect for the following reasons. Some bloodvessels have nonadrenergic, noncholinergic vasodila-tor innervation. Electrical field stimulation in vitroactivates transmural nerves to produce dilation. RBChemolysate products (most likely oxyhemoglobin)inhibit this response.1033 Whereas lymphatics havenoradrenergic innervation and contain a- and fl-re-ceptors, tetrodotoxin (a nerve blocker) has no effect
on the pumping activity of isolated vessels,34,35 illus-trating that nerves do not play a role in the regulationof spontaneous contractile activity. Therefore, itseems unlikely that hemoglobin acts on the lymphaticthrough an effect on nerve function.
Pharmacological responses to some agents aremediated by the production of factors from theendothelium. As one example, the dilator responsesto acetylcholine are mediated by endothelium-de-rived relaxing factor.36 Hemoglobin has been demon-strated to inhibit dilation in endothelium-dependentreactions such as those induced with acetylcholine.Endothelium-derived relaxing factor binds to theheme group of guanylate cyclase to cause enzymeactivation and stimulation of cyclic GMP.3' Theendothelium-derived relaxing factor receptor maytherefore contain a heme group, and it has beenpostulated that hemoglobin competes with the hemereceptor. In our first attempts to study the effects ofhemoglobin on lymphatic contractions, we used aring preparation and observed that hemoglobin in-creased contractions in some preparations, de-pressed contractions in others, and in some cases,had no effect (unpublished observations, 1989). Partof the problem may have been that the endotheliumwas being damaged by the wires used to suspend thevessel segments. It is interesting to note that a similarvariation was observed in studies designed to test theeffects of acetylcholine on vascular preparations, andit turned out that the presence or absence of theendothelium was the key to this effect.36There is some experimental evidence that pros-
taglandin synthesis is involved in mediating the effectsof hemoglobin on some vascular smooth muscle prep-arations.30,37 This perhaps is not surprising becauseheme is a cofactor in the synthesis of prostaglandinsand is believed to be involved in the step in whichmolecular 02 is attached to unsaturated fatty acids.38In support of this, cyclooxygenase inhibitors such asaspirin and indomethacin can markedly alter theresponse of basilar artery preparations to hemoglo-bin.30'37 Further confirmation of this relation comesfrom superfusion studies that demonstrated that he-moglobin was capable of causing the release of variousprostaglandin types from vascular preparations.'1 Thepossibility that arachidonic acid metabolism may beinvolved with the hemoglobin effect seems to warrantconsideration in our model system, especially becausewe have obtained evidence in the past that the syn-thesis of cyclooxygenase and lipoxygenase productswithin the lymphatic vessel may be important inregulating the spontaneous contractions.39-42
It has been suggested that free radical reactionsmay be responsible in part for the effects of hemo-globin on cerebral vascular smooth muscle prepara-tions and contribute to the vasospasm and ischemiaafter subarachnoid hemorrhage. Whether this effectis due to free radical production24 or whether it issecondary to the generation of various lipid peroxidesby these reactions27,28 is unknown. In this regard, theoxidation of oxyhemoglobin to methemoglobin (if
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

Elias et al Erythrolysate and Lymphatic Pumping 1105
this occurs in vivo or in the organ bath preparation)could result in the generation of free radical reac-tions,43 and these products conceivably could alterlymphatic pumping.
AcknowledgmentsThe authors wish to thank Mr. M. Walker for his
technical assistance and Dr. Rollin Brant and Mr.Yong Hao (Department of Biostatistics, University ofToronto) for help in the statistical analysis of thedata. The authors are grateful to the managementpersonnel and veterinary inspectors at Grace MeatPackers (York) Ltd. for their cooperation and help inproviding the bovine tissues.
References1. Johnston MG: Involvement of lymphatic collecting ducts in
the physiology and pathophysiology of lymph flow, in JohnstonMG (ed): Experimental Biology of the Lymphatic Circulation.New York, Elsevier Science Publishers, 1985, pp 81-120
2. McHale NG: Innervation of the lymphatic circulation, inJohnston MG (ed): Experimental Biology of the LymphaticCirculation. New York, Elsevier Science Publishers, 1985,pp 121-140
3. McHale NG, Thornbury KD: A method for studying lymphaticpumping activity in conscious and anesthetized sheep. JPhysiol(Lond) 1986;378:109-118
4. Hayashi A, Elias RM, Nelson W, Hamilton S, Johnston MG:Inhibition of intrinsic pumping activity in ovine mesentericlymphatics during endotoxin shock. Curr Surg 1987;45:131-134
5. Elias R, Johnston MG, Hayashi A, Nelson WR: Decreasedlymphatic pumping following intravenous endotoxin adminis-tration in sheep. Am J Physiol 1987;253:H1349-H1357
6. Elias RM, Johnston MG: Modulation of lymphatic pumpingby lymph-borne factors following endotoxin administration insheep. JAppl Physiol 1990;68:199-208
7. Osaka K: Prolonged vasospasm produced by the breakdownproducts of erythrocytes. JNeurosurg 1977;47:403-411
8. Okwuasaba F, Cook D, Weir B: Changes in vasoactive prop-erties of blood products with time and attempted identificationof the spasmogens. Stroke 1981;12:775-780
9. Fujiwara S, Kuriyama H: Hemolysate-induced contraction insmooth muscle cells of the guinea pig basilar artery. Stroke1984;15:503-510
10. Lee TJF, McIlhany MP, Sarwinski S: Erythrocyte extractsenhance neurogenic vasoconstriction of dog cerebral arteriesin vitro. J Cereb Blood Flow Metab 1984;4:474-476
11. Okamoto S, Handa H, Handa Y: Hemolysate-induced releaseof prostaglandinlike substances from the canine cerebralarteries. Surg Neurol 1985;23:421-424
12. Walter SV, Chang TMS: Chronotropic effect of stroma-freehemoglobin and polyhemoglobin on cultured myocardiocytesderived from newborn rats. Biomater Artif Cells Artif Organs1988;16:701-703
13. Kim HW, Feola M, Rowley BA, Roberts LA: Effects ofhemoglobin perfusion on contractile function of the isolatedventricular septa. Biomater Artif Cells Artif Organs 1988;16:331-345
14. Elias RM, Johnston MG: Modulation of fluid pumping inisolated bovine mesenteric lymphatics by thromboxane/endoperoxide analogue. Prostaglandins 1988;36:97-106
15. Clemmons RM, Bliss ER, Dorsey-Lee MR, Meyers KM:Platelet function, size and yield in whole blood and in platelet-rich plasma prepared using different centrifugation force andtime in domestic and food-producing animals. Thromb Hae-most 1983;50:838-843
16. De Venuto F, Zuck TF, Zegna AI, Moores WY: Characteris-tics of stroma-free hemoglobin prepared by crystallization. JLab Clin Med 1977;80:509-516
17. MacFate RP: Measurement of hemoglobin in blood, in Sun-derman FW, Sunderman FW Jr (eds): Hemoglobin, Its Precur-sors and Metabolites. Philadelphia, JB Lippincott Co, 1964,pp 31-39
18. Naumann HN: The measurement of hemoglobin in plasma, inSunderman FW, Sunderman FW Jr (eds): Hemoglobin, ItsPrecursors and Metabolites. Philadelphia, JB Lippincott Co,1964, pp 40-48
19. Cham BE, Knowles BR: A solvent system for delipidation ofplasma or serum without protein precipitation. J Lipid Res1976;17:176-181
20. Sugita Y, Negai M, Yoshimasa Y: Circular dichroism ofhemoglobin in relation to the structure surrounding the heme.JBiol Chem 1971;246:383-386
21. Hanley CA, Elias RM, Movat HZ, Johnston MG: Suppressionof fluid pumping in isolated bovine mesenteric lymphatics byInterleukin-1: Interaction with prostaglandin E2. MicrovascRes 1989;37:218-229
22. Cheung ST, Mcllhany MP, Lim R, Mullan S: Preliminarycharacterization of vasocontractile activities in erythrocytes. JNeurosurg 1980;53:37-43
23. Gilroy D, Shaw C, Parry E, Odling-Smee W: Detection of avasoconstrictor factor in stroma-free haemoglobin solutions. JTrauma 1988;28:1312-1316
24. Wellum GR, Irvine TW, Zervas NT: Cerebral vasoactivity ofheme proteins in vitro: Some mechanistic considerations. JNeurosurg 1982;56:777-783
25. Boullin DJ, Tagari P, Boulay DG, Aitken V, Hughes JT: Therole of hemoglobin in the etiology of cerebral vasospasm. JNeurosurg 1983;59:231-236
26. Adair TH, Guyton AC: Lymph formation and its modificationin the lymphatic system, in Johnston MG (ed): ExperimentalBiology of the Lymphatic Circulation. New York, ElsevierScience Publishers, 1985, pp 13-44
27. Sasaki T, Tanishima T, Asano T: Significance of lipid peroxi-dation in the genesis of chronic vasospasm following ruptureof an intracranial aneurysm. Acta Neurochir (Wien) 1979;suppl28:536-540
28. Sasaki T, Wakai S, Asano T: The effect of a lipid hydroper-oxide of arachidonic acid on the canine basilar artery: Anexperimental study on cerebral vasospasm. J Neurosurg 1981;54:357-365
29. Sasaki T, Asano T, Takakura K, Sano K, Kassell NF: Natureof the vasoactive substance in CSF from patients with sub-arachnoid hemorrhage. J Neurosurg 1984;60:1186-1191
30. Okamoto S, Handa H, Toda N: Role of intrinsic arachidonatemetabolites in the vascular action of erythrocyte breakdownproducts. Stroke 1984;15:60-64
31. Ignarro LJ: Endothelium-derived nitric oxide: Actions andproperties. FASEB J 1989;3:31-36
32. Kanamaru K, Waga S, Kojima T, Fujimoto K, Niwa S:Endothelium-dependent relaxation of canine basilar arteries:Part 2. Inhibition by hemoglobin and cerebrospinal fluid frompatients with aneurysmal subarachnoid hemorrhage. Stroke1987;18:938-943
33. Bowman A, Gillespie JS: Block of some non-adrenergic inhib-itory responses of smooth muscle by a substance from hemo-lysed erythrocytes. J Physiol (Lond) 1982;328:11-25
34. McHale NG, Roddie IC, Thornbury KD: Nervous modulationof spontaneous contractions in bovine mesenteric lymphatics.J Physiol (Lond) 1980;309:461-472
35. Allen JM, McHale NG, Rooney BM: Effect of norepinephrineon contractility of isolated mesenteric lymphatics. Am JPhysiol1983;244:H479-H486
36. Furchgott RF, Zawadzki JV: The obligatory role of endothe-lial cells in the relaxation of arterial smooth muscle byacetylcholine. Nature 1980;288:373-376
37. Brandt L, Ljunggren B, Andersson K-E, Hindfelt B, Uski T:Effect of indomethacin and prostacyclin on isolated humanpial arteries contracted by CSF from patients with aneurysmalSAH. J Neurosurg 1981;55:877-883
38. Yoshimoto A, Ito H, Tomita K: Cofactor requirements of theenzyme synthesizing prostaglandin in bovine seminal vesicles.J Biochem 1970;68:487-499
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

1106 Circulation Research Vol 67, No 5, November 1990
39. Johnston MG, Gordon JL: Regulation of lymphatic vesselcontractility by arachidonate metabolites. Nature 1981;293:294-297
40. Johnston MG, Kanalec A, Gordon JL: Effects of arachidonicacid and its cyclo-oxygenase and lipoxygenase products on
lymphatic vessel contractility in vitro. Prostaglandins 1983;25:85-98
41. Johnston MG, Feuer C: Suppression of lymphatic vesselcontractility with inhibitors of arachidonic acid metabolism. JPharmacol Exp Ther 1983;226:603-607
42. Allen JM, Burke EP, Johnston MG, McHale NG: The inhib-itory effect of aspirin on lymphatic contractility. Br J Phanna-col 1984;82:509-514
43. Misra HP, Fridovich I: The generation of superoxide radicalduring the autoxidation of hemoglobin. J Biol Chem 1972;247:6960-6962
KEY WORDS * lymphatic vessel * erythrolysate * hemoglobin* pumping activity
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

R M Elias, G Wandolo, N S Ranadive, J Eisenhoffer and M G Johnstonerythrolysate/hemoglobin.
Lymphatic pumping in response to changes in transmural pressure is modulated by
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1990 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.67.5.10971990;67:1097-1106Circ Res.
http://circres.ahajournals.org/content/67/5/1097World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from















![Inositol 1,4,5-Triphosphate-induced Granule Secretion ...dm5migu4zj3pb.cloudfront.net/manuscripts/112000/... · am (U co Co en.01.1 1 10 [U46619] FM Figure2. Secretion in responseto](https://img.pdfslide.us/doc/110x75/5f7b9628f18fdc1987195590/inositol-145-triphosphate-induced-granule-secretion-am-u-co-co-en011-1.jpg)













