Embed Size (px)
Citation preview

Shellfishing and the Colonization of Sahul:A multivariate model evaluating the dynamic effects of preyutility, transport considerations and life-history on foragingpatterns and midden composition
Brian F. Codding1,∗, James F. O’Connell 1 and Douglas W. Bird2
1. Department of Anthropology, University of Utah, Salt Lake City, UT, 84112, USA2. Department of Anthropology, Stanford University, Stanford, CA, 94305, USA∗ Corresponding author. E-mail: [email protected]
UNCORRECTED AUTHOR’S MANUSCRIPT
Citation: Codding, Brian F., James F. O’Connell and Douglas W. Bird (2014) Shellfishing and the Colonization ofSahul: A multivariate model evaluating the dynamic effects of prey utility, transport considerations and life historyon foraging patterns and midden composition. Journal of Island & Coastal Archaeology 9(2):238–252.
Abstract. Archaeological evidence of shellfishexploitation along the coast of Sahul (PleistoceneAustralia-New Guinea) points to an apparentparadox. While the continental record as a wholesuggests that human populations were very lowfrom initial colonization through early Holocene,coastal and peri-coastal sites dating to that timeare dominated by small, low-ranked littoral taxato the near-complete exclusion of large, higherranked sub-littoral species, precisely the oppo-site of theory-based expectations, if human pop-ulations and predation rates were indeed as lowas other data suggest. We present a model ofshellfish exploitation combining information onspecies utility, transport considerations and preylife-history that might account for this apparentmismatch, and then assess it with ethnographicand archaeological data. Findings suggest eitherthat high-ranked taxa were uncommon along thePleistocene coastlines of Sahul, or that abun-dant and commonly taken high-ranked prey areunder-represented in middens relative to theirrole in human diets largely as a function of hu-man processing and transport practices. If thelatter reading is correct, archaeological evidenceof early shellfishing may be mainly the product ofsubsistence activities by children and their moth-ers.
Keywords: Shellfish Exploitation | Coloniza-tion of Sahul (Australia/New Guinea) | Chil-dren’s Foraging | Division of Labor | Human Be-havioral Ecology
Introduction
Archaeological evidence of early shellfish collect-ing on Sahul (Pleistocene Australia-New Guinea)points to an apparent paradox. While theo-retical predictions from foraging theory lead tothe expectation that the earliest foragers onthe continent would have encountered abundantlarge high-ranking shellfish taxa offering rela-tively high average rates of nutrient return rela-tive to time spent collecting and processing, ar-chaeological data from coastal shell middens in-dicate that foraging emphasized smaller, muchlower ranked shellfish taxa. There are at leastthree plausible explanations for this pattern:
1. Rapidly changing Pleistocene sea levels keptthe abundance of large, high-ranked, slow-growing taxa low, leaving foragers no optionbut to pursue smaller, lower ranked prey(Beaton 1995).
2. Early human populations were much higherthan previously thought, placing sustained
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
collecting pressure on high ranked preywhich rapidly reduced their numbers, leav-ing only an ephemeral archaeological signa-ture.
3. Transport considerations favored the differ-ential discard of inedible components of rel-atively high ranked prey at or near the pointof acquisition, leaving little or no evidenceof their use in base camp middens. The ma-terial signature of lower-ranked taxa maythen be the result of foraging by children,who experience lower encounter rates withlarge prey due to their slower walking speed(Bird et al. 2004), and by their moth-ers, who may be more interested in low-riskthan high-energy resources for provisioningyoung (Codding et al. 2011).
Resolving this issue is crucial to understand-ing the process through which Sahul was colo-nized and the early foraging decisions of indi-viduals on a previously unoccupied landscape(O’Connell and Allen 2012). Here we addressthe problem with a novel model that examinesprey utility, transport considerations and life-history. Combined, this approach may betterpredict both resource exploitation patterns andtheir archaeological consequences. We assess themodel in light of ethnographic and ethnoarchae-logical data, then consider its implications forunderstanding the record of early shellfish ex-ploitation in Sahul.
The Model
Our model is based on well-established ideasabout prey choice, central place foraging andlife history. These allow us to make predic-tions about which resources will be more (orless) likely to be targeted, which will be more(or less) likely to be represented in archaeolog-ical deposits created by central place foragers,and which will be more (or less) resilient to pre-dation pressure or environmental change.
The prey choice model (PCM) allows thederivation of simple predictions about which
species are more likely to be pursued on en-counter (Bettinger 2009, Bird and O’Connell2006, Charnov and Orians 2006). When search-ing a patch, a forager will encounter a vari-ety of different resources. If foragers are tryingmaximize the rate of nutrient acquisition, thenthey should preferentially pursue resources withhigher post-encounter return rates (e/h), mea-sured as the expected nutrients (e) gained fromacquiring a resource (typically measured as en-ergy) over the handling costs (h) associated withpursuing and processing that resource (typicallymeasured in time). If higher ranking resourcesare sufficiently abundant, then foragers shouldonly take those resources on encounter, passingup all other lower ranking resources; however, asthe abundance of the highest ranking resourcesdecline, foragers should begin to take lower rank-ing resources in rank order. From a forager’sperspective, the question is: should I pursue thisresource, or pass it up and continue searchingfor other, higher-ranked resources? The answerdepends on whether the post-encounter returnrate (e/h) for that resource is expected to in-crease the overall return rate (E/T) which in-cludes all energy acquired in a patch (E) rel-ative to all in-patch search and handling time(T). These model dynamics allow the derivationof predictions about which taxa a forager shouldpreferentially pursue and a proxy for overall for-aging efficiency based on the presence or absenceof lower-ranking resources.
Central place foraging models (CPF) allowthe evaluation of the costs and benefits of fieldprocessing resources or transporting them wholefrom an acquisition locale to a central place (Bet-tinger 2009, Charnov and Orians 2006, Metcalfeand Barlow 1992, Orians and Pearson 1979).Once a forager has acquired a resource, theymust make decisions about how to best trans-port that resource to a central place. Assumingthat individuals are attempting to maximize therate at which resources are delivered to a centralplace, foragers may be better off culling low util-ity items (i.e., shell or bone) in the field and re-turning to their central place with only high util-
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
ity parts (i.e., meat) than making a greater num-ber of trips transporting both high and low util-ity parts. Assuming constant processing costsacross taxa, the net gain in utility as a functionof processing (the increase in high utility partsper load) can predict which items are more likelyto be field processed at a given distance froma central place. Given that individual shellfishhave an edible portion (meat) and a non-edibleportion (shell), meat:shell ratios provide a reli-able proxy of the potential benefit gained fromculling shell in the field, thereby increasing theutility of each load returned home. Those taxawith higher proportions of meat to shell are lesslikely to be field processed while those with lowmeat:shell ratios are more likely to be processedin the field. Because larger taxa generally havelower meat:shell ratios and require lower pro-cessing costs than smaller taxa (e.g., Bird et al.2002), these predictions likely hold with the in-clusion of processing costs.
Life history models (LFM) provide avenuesto investigate variability in the parameters thatgovern individual growth, maturation, reproduc-tion and mortality, which aggregate to producepopulation level effects. Because different taxahave different life-history characteristics, theyshould respond differently to human exploita-tion, which in turn, will alter a forager’s futureencounter rates. With invertebrate taxa, life-history parameters such as growth and matu-ration rates should provide a reliable proxy ofthe relative susceptibility of a taxon to overex-ploitation. Likely the result of natural selection(Gadgil and Bossert 1970), the rate of matura-tion covaries with a number of life-history pa-rameters and can be a predictor of populationlevel parameters (Peters 1983). As such, ratesof maturation for individual taxa should providea relative measure of the likelihood or speed atwhich a population could recover from exploita-tion (Whitaker 2008, Whitaker and Byrd 2012).Due to the effects of biological scaling and allom-etry, body size and post-encounter return rates(at least, for sessile resources, see Bird et al.2009) should co-vary positively while body size,
meat:shell ratios and maturation rates may co-vary negatively (Peters 1983). This could beproblematic as high ranking resources may bemore likely to be processed in the field, but ifparticular taxa exist as outliers in these rela-tionships (i.e., high ranking, slow-growing buthigh meat: shell ratios), they should provide keyproxies to monitor changes in foraging patternsthrough time.
Combined, these models allow us to make pre-dictions about which species should be prefer-entially pursued, which should be more or lesslikely to end up in archaeological sites and whichare more or less likely to be depleted by preda-tion. Following O’Connell (1995), these modelscan be used to derive specific deductive predic-tions that can be tested with ethnographic, eth-noarchaeological and archaeological data. Be-cause these predictions are derived from a gen-eral theory of behavior, they avoid direct ethno-graphic analogy.
Predictions
Imagine a parameter space represented by threevariables: shellfish utility (e/h, on the Figure1: y-axis), transport costs (operationalized bymeat:shell ratios, Figure 1: x-axis) and sus-ceptibility to overexploitation (measured herethrough the proxy of maturation rates). Howdifferent shellfish taxa fall out within this pa-rameter space should allow us to identify targettaxa that will allow us to investigate foragingbehavior, midden composition and how they areexpected to change over time.
Taxa with very low meat:shell ratios (Figure1: frame a) should rarely appear in middens dueto the low net utility of a load returned to acentral place with such a large amount of shellrelative to a limited amount of meat. If shellfishrank varies inversely with meat:shell ratios, thenevidence for the exploitation of high ranking taxais unlikely to be recovered from archaeologicaldeposits as their shells should be culled in thefield even over short transport distances. Thisis the case for many molluscan species from thisregion (Bird and Bliege Bird 2000).
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
Low High
Low
High
Meat:Shell
e/h
Dominate early middensAb
sent
from
mid
dens
Dominate late middens
(a) (b)
(c)
Figure 1: Figure 1. Schematic predictions of midden composition based on the combined effects ofeach taxon’s post-encounter return rate (e/h), and likelihood of field processing (meat:shell ratio).Taxa with low meat:shell ratios (frame a) are less likely to end up in middens as their shells shouldbe frequently culled in the field (unless if the shell itself provides some value). Early middensshould be dominated by higher ranking taxa with relatively high meat:shell ratios (frame b). Ifthese become less abundant in the environment due to overexploitation, then foragers should beginto take lower ranking resources which will come to dominate later middens (frame c). The rate oftransition in midden dominance from b→c depends on the life-history characteristics of the taxain question: fast growing species should be harder to deplete while slow growing species rapidlydecline in abundance. Note: these values are in dimensionless space.
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
High ranking taxa with high meat:shell ra-tios (Figure 1: frame b) should always be takenon encounter by foragers and are likely to al-ways be transported whole to the central place.As such, these taxa should dominate early mid-dens. Those with slow maturation rates shouldbe the best source to monitor the effects of re-source overexploitation through time. Thosethat mature early are likely to be fairly resilientto over-exploitation and may dominate middensthroughout the prehistoric record.
Lower-ranked taxa with high meat:shell ratios(Figure 1: frame c) should come to dominate lat-ter middens if higher ranking resources becomeless abundant due to overexploitation. Thosethat take longer to mature will be particularlyuseful to monitor human foraging and popula-tion pressure over time. Those that are rela-tively resilient to over-exploitation (rapid growthrates and short maturation rates) should domi-nate later shell middens (and shell mounds) fol-lowing declines of the highest ranking resources(e.g., Whitaker and Byrd, this issue).
ResultsResource-Specific Attributes
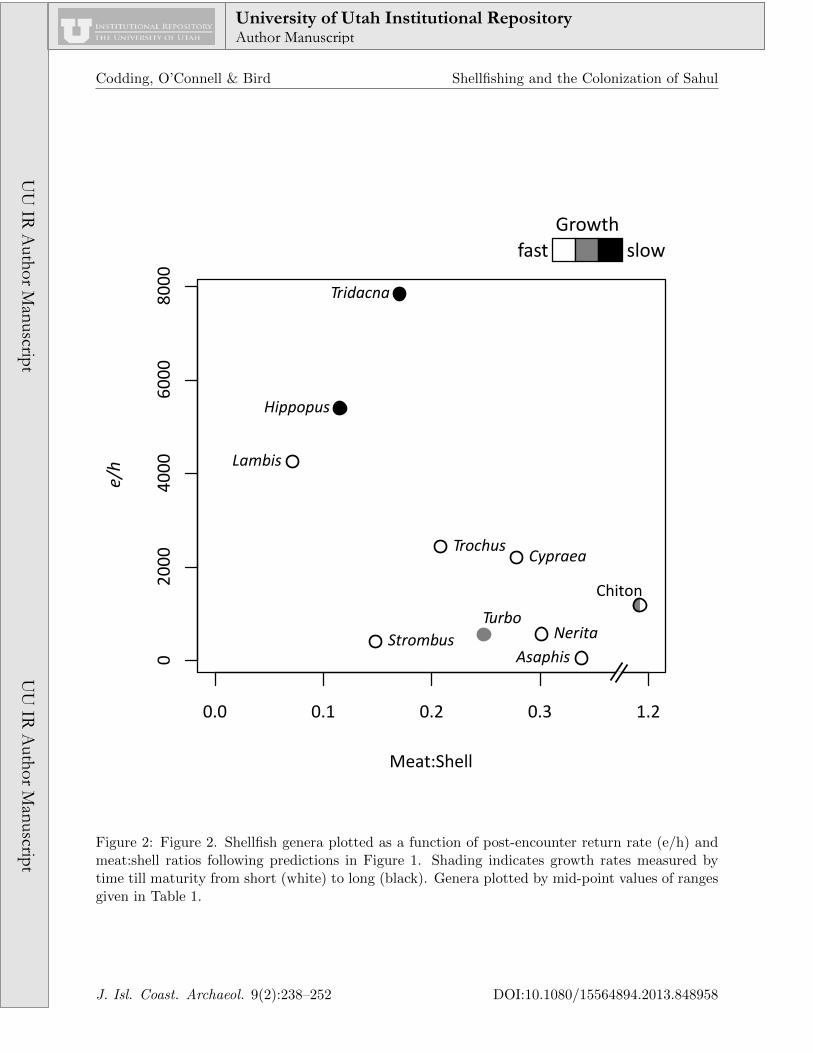
Table 1 provides a summary of available dataon shellfish genera post-encounter return rates(e/h), meat:shell ratios and age at maturity fortaxa potentially exploited by the earliest inhabi-tants of Sahul. Mean values are also representedin Figure 2 following the layout of predictions inFigure 1.
The two large bivalves, Tridacna and Hip-popus, would be excellent candidates to moni-tor foraging pressure given their high rank andslow growth rates. However, given their lowmeat:shell ratios, they are unlikely to be trans-ported to midden deposits proportionally to thefrequency with which they are taken. This ex-pectation is supported by ethnoarchaeologicalstudies indicating that the material record ofTridacna and Hippopus underrepresent the fre-quency with which they are actually acquired(see below, Bird and Bliege Bird 2000, Bird etal. 2002).
Table 1: Proxy data for shellfish utility (e/h),transport cost (meat:shell) and susceptibility tooverexploitation (age at maturity), ordered bymid-point return rate (e/h).
Taxon e/h Meat:Shell Age at(kcal/hr) Ratio Mat. (yr)
Tridacna 2,622-13,064 0.170 4-5Hippopus 1,680-9,120 0.115 6-7Lambis 3,412-5,106 0.092 2Trochus 977-3,904 0.208 1-2Cypraea 2,214 0.278 1Chiton 446-2,228 1.159 1-2+?Nerita 42-1,106 0.301 1.6Turbo 520-606 0.248 3-4Strombus 294-543 0.148 1-2Asaphis 42-78 0.338 1-2
Data on shellfish returns from Bird and Bliege Bird(2000), Bird et al. (2004), Kennedy (2005) and Thomas(2007); on meat:shell ratios from Bird and Bliege Bird(1997), Erlandson (1988); on age at maturity fromBeesley et al. (1998), Munro (1993), Nash (1993) andYamaguchi (1993).
Unfortunately, within the suite of resourcesavailable to the earliest colonists of Sahul (andfor which data are currently available), there areno species that can be considered high-rankingwith high meat:shell ratios (i.e., falling withinparameter space b in Figure 1). Evidence fromNorth America suggests that Abalone (Haliotisspp.) likely falls within this region of the param-eter space (Kennedy 2005), but conclusive dataon Austral species are lacking. The absence ofspecies within this parameter space suggests thatefficient tidal foraging in Sahul required field pro-cessing. As a result, we are unlikely to acquirean unbiased picture of the prehistoric exploita-tion of these higher ranking taxa.
With relatively low returns but high meat:shellratios, small gastropods such as Nerita and smallbivalves like Asaphis also have rapid matura-tion rates, suggesting they are resilient to overex-ploitation. As we would predict, these taxa dom-inate modern middens within permanent villagesin the Torres Strait Islands, Australia and West-ern Kiribati, Micronesia where sustained forag-ing pressure does not seem to diminish their pop-
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript
Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
0.0 0.1 0.2 0.3 1.2
020
0040
0060
0080
00
Meat:Shell
e/h
NeritaAsaphis
CypraeaTrochus
Turbo
Hippopus
Strombus
Lambis
Tridacna
Chiton
Growthfast slow
Figure 2: Figure 2. Shellfish genera plotted as a function of post-encounter return rate (e/h) andmeat:shell ratios following predictions in Figure 1. Shading indicates growth rates measured bytime till maturity from short (white) to long (black). Genera plotted by mid-point values of rangesgiven in Table 1.
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
ulations (Bird et al. 2002, Thomas 2007). Asthese taxa should be resilient to overexploitation,their abundance is unlikely to vary with humanforaging and population pressure over time.
The moderately sized gastropod Turbo pro-vides a welcome exception: with relatively longtimes to maturity and high meat:shell ratios, weshould expect that Turbo will be exploited whenthe abundance of higher ranking resources arelow, and should be one of the first taxa withinthis lower-ranked set to exhibit overexploitation.As such, diachronic trends in the exploitationof Turbo should provide a key proxy to under-stand foraging pressure on shellfish species, andas such, a proxy for human population densi-ties. With by far the most extreme meat:shellratio, chiton (class Polyplacophora) may also fallwithin this category. Larger species of chitonare likely to have higher return rates and slowermaturation rates, while still maintaining rela-tively high meat:shell ratios; unfortunately, pre-cise data are not available for Austral species.Based on observations of Meriam shellfishers byone of the authors (DWB), limited data on chi-ton exploitation by contemporary foragers mayresult from low encounter rates with larger chi-ton coupled with consistent decisions to pass oversmaller chiton in search of more profitable taxa.
An Ethnographic Test
Previous ethnoarchaeological investigation withMeriam Islanders in the Torres Strait exploredthe archaeological consequences of trade-offsthat shellfishers face in acquiring, handling, andtransporting a number of the taxa that Pleis-tocene foragers would have likely encounteredon colonizing Sahul (Bird 1996, Bird and BliegeBird 1997, 2000; Bird et al. 2002, 2004). Con-sistent with expectations framed by the preychoice model (PCM), Meriam restrict the suiteof taxa they handle on encounter to those shell-fish that will increase overall foraging efficiency(E/T) while collecting on the reef flat or harvest-ing within the rocky shore. For example, whilereef flat collecting, foragers routinely select ex-posed tridacnid clams (Tridacna spp. and Hip-
popus hippopus), Lambis lambis conch, and largeTrochus niloticus specimens, all of which signif-icantly increase overall foraging return rates ifhandled on-encounter. Conversely adult shell-fishers almost always pass over specimens thatoffer returns for time handling that will reduceforaging efficiency: small Strombus (Conomurex)luhuanus conch, small Trochus, and small bi-valves embedded in the reef. Meriam also payclose attention to changes in patch return rateswhile intertidal foraging, and switch from reefflat collecting to rocky shore harvesting (forAsaphis violascens clams and Nerita spp. snails)when an incoming tide depresses the overall re-turn rate from collecting in the mid-littoral be-low that expected from harvesting in the nearshore.
While observed time allocation and selectiv-ity are consistent with predictions generated bythe PCM and patch residence models, they arein sharp contrast to the frequency of differenttaxa of shellfish found in either contemporary orprehistoric shellmiddens on the Meriam Islands.The most important resources for contemporarycollectors (especially large tridacnids and Lam-bis lambis) are very rare in the archaeologicaldeposits, which are dominated by small gastro-pod shells (especially nerites, small strombids,and small Trochus niloticus; Bird et al. 2002).We can account for these differences by consid-ering the effects of differential field processingand age-linked differences in foraging returns.While reef flat collecting, children walk slowerand encounter high ranked resources at a lowerrate than adults, resulting in lower overall re-turn rates and a predictably broader range ofselectivity that includes the smaller gastropods.So while both adults and children collect thehigher ranked shellfish on-encounter, only chil-dren regularly collect the abundant small strom-bids and small specimens of Trochus niloticusthat are common in the shell middens. But whyare tridacnid and Lambis shell, which make upthe bulk of shellfish harvested, so rare in the de-posits? If we consider how time spent field pro-cessing can increase the ratio of high to low util-
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
ity parts (meat:shell), the central place foragingmodel discussed above generates precise predic-tions about which types of molluscs should befield processed if a forager’s goal is to increasethe rate at which shellfish meat can be trans-ported home. The model well anticipates thefact that Meriam foragers almost always cull theshells of large tridacnid clams and Lambis conchwhile on the reef. These shellfish offer high post-encounter return rates and have low meat:shellratios, and thus field processing quickly frees upmore space for more meat, and more time formore foraging and transport. As such, Meriamadults and children commonly forage well be-yond the predicted thresholds at which in-bulktransportation (without culling the valves) ofthese large shellfish will increase the home de-livery rate of meat. Conversely, the smaller reefflat shellfish that children often collect, alongwith rocky shore taxa such as Asaphis and Ner-ita that both adults and children harvest, arealways transported in-bulk, and foragers veryrarely cross the predicted field processing thresh-olds while collecting these prey. We suspect thatPleistocene foragers would have faced similartrade-offs when shellfishing in intertidal zones.
Early Middens in Sahul
Data are available for at least six Pleistocenemiddens (coastal and peri-coastal sites yield-ing marine shell) with dates older than 30kya.Buang Merabak (Leavesley and Allen 1998,Rosenfeld 1997) and Matenkupkum (Gosden andRobertson 1991) are located on New Ireland,Kilu (Wickler 2001) on the northern end of theSolomon Islands and Noala Cave (Veth et al.2007), Mandu Mandu Creek (Morse 1988), andDevil’s Lair (Dortch 2004) are located alongthe coast of modern day Western Australia.1
For more detailed summaries on the dating ofeach site, see Allen and O’Connell (2003, 2008),O’Connell and Allen (2004, 2007, 2012) and
1Early middens (pre-30kya) for which data were un-obtainable include other sites along the Cape Range ofnorthwestern Australia (Przywolnik 2008, Morse 1988,Morse 1993).
O’Connell et al (2010). Evidence for shellfishexploitation at many of these sites is extremelysparse; nonetheless, the available data allow usto run an initial test of these predictions.
Higher ranking species with low meat:shellratios are rare in these early middens. AtBuang Merabak, chiton, Turbo and Nerita dom-inate the Pleistocene assemblage (Leavesley andAllen 1998). At Kilu, Nerita dominates assem-blages dating before the Late Holocene (Wickler2001). At Matenkupkum, Gosden and Robert-son (1991) report extremely large Turbo shells,indicating the exploitation of lower ranked taxa,but assuming that human predation will drivedown shellfish size (e.g., Klein and Steele 2013)only limited predation pressure on these taxa.While Tridacna is absent from the faunal assem-blages recovered from these early levels, Leaves-ley and Allen (1998) report that large Tridacnashells and Turbo opercula, originally recordedas white chert, were used to manufacture toolsat Buang Merabak. This suggest that thesetaxa were taken on encounter, likely for theirfood and shell, but only transported and de-posited within midden contexts for purposes re-lating to the latter. This also suggests thatsuch high-value shell might be secondarily trans-ported away from middens for subsequent use, apattern that may further bias such a taxon’s rep-resentation in the archaeological record.
At peri-coastal sites along the shore of modernday Western Australia, shellfish taxa are dom-inated by different genera specific to the localenvironment; but even so, the taxa exploited atthese sites share characteristics with those ex-ploited in the Bismarks. At Noala Cave on theMonte Bello Islands, foragers were exploiting avariety of whelks (Terebralia, Veth et al. 2007),which are expected to be low ranked. Perhapsat odds with findings elsewhere, the earliest lay-ers at Mandu Mandu Creek reveal chiton andbaler (Melo) (Morse 1988). While precise dataon these taxa are not yet available, what we knowsuggests that they may be relatively high rankedwith high meat:shell ratios (Figure 1, frame b).If these were large chiton and Melo, this evi-
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
Buang MerabakMatenkupkum
Kilu
Devil’s Lair
Noala Cave
Mandu Mandu
60 m contour120 m contour
W A L L A C E A
S U N D A
S A H U L
P a c i f i c O c e a n
I n d i a n O c e a n
B i s m a r k s
S o l o m o n s
Figure 3: Map of coastal and peri-coastal archaeological sites with evidence of marine shell ex-ploitation prior to 30kya.
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
dence for the exploitation of high-ranking taxaduring this interval supports the idea that large,high-ranking taxa were present, pursued on en-counter, but only transported to central placeswhen meat:shell ratios were high or when theshells has some added value. This might alsobe true in the southwestern corner of the conti-nent, where small fragments of abalone (Halio-tis) are some of the only taxa represented in theearly levels at Devil’s Lair (and at Tunnel Cave;Dortch 2004; J. Dortch, personal communication2013). Abalone too likely falls within the param-eter space shown in Figure 1, frame b represent-ing high-rank and high meat:shell ratios, leadingus to expect that it will often be transportedwhole. The additional value of abalone shell forritual, ornamental or other purposes beyond sub-sistence likely increase the probability that itwill often be transported whole. This is consis-tent with finds at Carpenter’s Gap, where earlylevels contain pearl shell (O’Connor 1999).
Diachronic Trends at Buang Merabak
Temporal trends from the well reported finds atBuang Merabak can further our understandingof these patterns. As predicted from our model,while we should not expect high-ranking, lowmeat:shell taxa to be deposited at central places,we should be able to monitor the abundance ofTurbo and chiton through time as an indicatorof foraging pressure.
As evident in Figure 4, the temporal trends atBuang Merabak show a significant decline in theproportion of Turbo (τ = 17.6, p = 0.0361) and amarginally significant decline in the proportionof chiton (τ = 4.2, p = 0.1498) through the Pleis-tocene. These are mirrored by a marginally sig-nificant increase in Nerita (τ = 7.0, p = 0.0905).2 Such patterning has also been reported at Kilu(Wickler 2001). While significant, declines inthe relative abundance of Turbo and chiton oc-cur over a long span of time, suggesting that it
2Pearson’s correlation analyses run in R (R Develop-ment Core Team, 2013). Figure 4 produced using thebattleship.plot function in the plotrix library.
Other
Nerita
Chiton
Turbo
1
2
3
4
5
6P
leis
toce
ne
Hol
ocen
e
Ana
lytic
al U
nit
Figure 4: Battleship plot showing the propor-tion of total weight per analytical unit for im-portant Pleistocene shellfish taxa at Buang Mer-abak. Data from Leavesley and Allen (1998),shown here in Table 2. Based on radiocarbondates form Rosenfeld (1997), analytical unit 6(2-sigma range 38,090-34,523 cal BP), 5 (24,141-22,505 cal BP) and 4 (24,782-23,356 cal BP) dateto the Pleistocene, while analytical units 3, 2 and1 all date to the Holocene (2-sigma ranges cali-brated in OxCal 4.2.2, Bronk Ramsey 2009, us-ing a marine curve from Reimer et al. 2009 andlocal correction by Petchey et al. 2004). The twomost abundant taxa in the earliest Pleistocenedeposits (Turbo and chiton) are also relativelyslow growing. Their overexploitation throughthe Pleistocene is mirrored in the rising dom-inance of faster-growing Nerita. Discontinuitybetween the Pleistocene-Holocene record is likelydue to sea level stabilization at distances closerto the site (see Lambeck and Chappell 2001).
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
Table 2: Shell weight (g) by taxa across chronological analytical units at Buang Merabak.
Taxa 1 2 3 4 5 6
Cypraea 90.6 223.0 16.6 0.0 0.0 13.6Strombus 34.2 62.8 1.2 0.6 0.0 0.0Isognomon 15.3 66.1 4.7 0.0 0.0 0.0Trochus 41.4 153.0 83.8 23.9 12.5 9.3Barnacle 50.0 136.9 797.9 8.9 0.9 2.0Limpet 7.8 26.5 33.3 24.2 0.5 0.0Nerita 52.7 226.0 258.0 1398.4 122.0 265.8Tectarius 2.5 3.1 11.3 79.2 11.3 49.9Chiton 22.6 61.0 53.9 267.3 99.7 509.4Turbo 13.1 71.1 7.9 165.5 58.9 408.9
From Balean (1989) reported in Leavesley and Allen (1998), see also Rosenfeld (1997). Given their high meat:shellratio, chiton likely contributed more to the early diets than is represented by shell weight alone (Leavesley 2004).
took most of the Pleistocene record before hu-man populations were high enough to place con-sistent pressure on these taxa. This interpre-tation is supported by the radiocarbon proba-bilities from Buang Merabak which suggest in-termittent occupations (Figure 5a) and occu-pational estimates from neighboring Australia,which suggest very low populations levels dur-ing this time (Figure 5b; Williams 2013). Thesefindings suggest that Pleistocene occupations atBuang Merabak were intermittent, perhaps co-varying negatively with the distance to the coast(cf., Figure 5a and 5c), and were always at a lowdensities.
Discussion
This paper began with a conundrum: the ar-chaeological record suggests that the earliest col-onizing populations of Sahul disproportionatelyexploited low-ranked littoral shellfish species,despite theoretical predictions to the contrary.While a number of hypotheses could be proposedto explain this conundrum, we suggest that thesecan be assessed by incorporating models of preyefficiency, transport utility and life-history topredict the targets of foraging, the resulting mid-den composition and the potential impact of hu-man foraging on shellfish species. In light ofour model, the conundrum actually makes sense:
high-ranking shellfish taxa available to the earlyforagers of Sahul had low meat:shell ratios (Fig-ure 1: frame b). As it would not make economicsense to transport shell from these taxa (unlessif the shell itself had some added value; e.g.,Leavesley and Allen 1998), we should not expectmiddens to contain shell from these taxa. Butwhat then accounts for the deposition of lowerranking littoral shellfish species?
Given the extremely low estimated populationlevels and intermittent occupation of sites likeBuang Merabak, it seems unlikely that the ac-quisition and deposition of low-ranked taxa wasdue to overexploitation of higher ranking taxa.If the abundance of these larger high-rankedspecies declined rapidly, then we might have ev-idence for a larger colonizing population placingmore pressure on resources than traditionally as-sumed. However, given the long period of timethat passed before human populations reacheddensities required to overexploit fairly slow grow-ing taxa like Turbo, we suggest that human pop-ulation densities were quite low, their mobilityquite high, and as such, they placed minimalpredation pressure on the littoral environment.Combined with evidence for the rapid dispersalof peoples along the coastlines of Sahul, this im-plies that it takes very little impact to make adja-cent, unoccupied patches more appealing to for-aging populations (O’Connell and Allen 2012).
It still could be that these taxa occurred in low
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
50 45 40 35 30 25
−120
−80
−40
0
Years cal BP
Sea
Lev
el (m
bel
ow p
rese
nt)
Relative Sea Level(Huon Peninsula, PNG)
40k
20k
0 Estimated Total Population of Australia
Pop
Buang Merabak(a)
(b)
(c)
Figure 5: Occupational history of Buang Merabak (a, Leavesley 2004, Leavesley and Allen 1998,summed calibrated radiocarbon proababilites from OxCal 4.2.2, Bronk Ramsey 2009, using a marinecurve from Reimer et al. 2009 and a local correction by Petchey et al. 2004) relative to the estimatedtotal population of Australia (b, assuming a founding population of 5,000 people, Williams 2013)and estimated fluctuations in sea level (c, Lambeck and Chappell 2001).
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
abundance due to the natural effects of a fluctu-ating coastline (Beaton 1995). While alternativelines of evidence (such as paleontological finds)will be required to test this further, we suspectthat habitats (e.g. broad fringing reefs) sup-porting healthy populations of some of the high-est ranked shellfish (such as Tridacna) wouldhave been commonly encountered when Sahulwas first colonized (Pope and Terrell 2008). Butagain, that such taxa are high-ranked and of-ten associated with low meat:shell ratios leads usto suspect that they would have been importantprey options whose archaeological presence washeavily filtered by field processing. This agreeswith ethnographic observations on tropical reefflat foraging, where highly ranked Tridacna arealways exploited on-encounter, but the valves arerarely transported back to a central place (Birdet al. 1997, 2002, 2004; Thomas 2007).
If our interpretations of these results are cor-rect, it seems that the most reasonable expla-nation for the high abundance of low-rankingtaxa at early sites may result from the differ-ential foraging constraints faced by children andtheir mothers. Because children generally walkmore slowly than adults and because they mayhave less strength and skill in processing, theyare likely to have lower encounter rates with highranked prey and are less able to field process taxaeven when it might be desirable to do so (Birdand Bliege Bird 2000, Bird et al. 2004, Meehan1982). Further, given the limited availability ofalternative caregivers expected with smaller pop-ulations (Codding et al. 2011), mothers withyoung children will generally experience greatertrade-offs between childcare and foraging. Inter-tidal shellfish foraging provides an activity withlower costs of childcare than other alternatives(e.g., sub-tidal foraging, pelagic fishing, etc.)and despite being low return, shellfish (espe-cially small taxa) are characterized by low acqui-sition variance, making them ideal resources forprovisioning dependents (Codding et al. 2011).Indeed, cooperative parent-juvenile units maydo better overall by working together in lower-return activities than by working apart (Hawkes
et al. 1995, Kramer 2011).
While this seems like the most plausibleexplanation for the Pleistocene record of shell-fishing on Sahul given the available evidence,more work is required to truly understand thesepatterns. Continued archaeological and pale-oenvironmental work is necessary, but researchmight benefit most from continued ethnoarchae-ologial and simple actualistic experiments. Thisincludes simple measures of utility, processingcosts and meat:shell ratios for key species(e.g., Halitois, Melo and chiton) and additionalstudies of well-known taxa (e.g., Turbo, Neritaand Asaphis) to further our understandingof variability within species. Additional workalong these lines should greatly improve ourinterpretations of the archaeological record andallow us to solve such conundrums as the recordof early shellfishing by the colonists of Sahul.
Acknowledgments. Thanks to Richard Kleinfor suggesting the idea for this symposium andcollection of papers. We remain indebted to theMeriam Community for hosting DWB’s ethno-graphic work over many years, which was sup-ported by the National Science Foundation andthe LSB Leakey Foundation. This paper hasbenefited from comments, suggestions and workby Jim Allen, Rebecca Bliege Bird, ShannonArnold-Boomgarden, Joe Dortch, James HollandJones, Peter Veth, Adie Whitaker, and DavidZeanah. Three anonymous reviewers provideddetailed comments that greatly improved the fi-nal result.
References
Allen, J. and J.F. O’Connell. 2003. The longand the short of it: Archaeological approachesto determining when humans first colonisedAustralia and New Guinea. Australian Archae-ology 57:5–19.
Allen, J. and J. F. O’Connell. 2008. Gettingfrom Sunda to Sahul. In Islands of Inquiry:colonization, seafaring and the archaeology
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
of maritime landscapes (G. Clark, F. Leachand S. O’Connor, eds): 31–46. ANU E Press,Canberra: Australian National University.
Balean, C. E. 1989. Caves as refuge sites: ananalysis of shell material from Buang Merabak.BA Honours Thesis, Deptartment of Prehistoryand Anthropology, Canberra: Australian Na-tional University.
Beaton, J. M. 1995. The transition on thecoastal fringe of greater Australia. Antiquity69:798–806.
Beesley, P. L., G. J. B. Ross and A.Wells(eds). 1998. Mollusca: The Southern Synthesis.Australian Biological Resources Study. CSIROPublishing. Parts 1 & 2.
Bettinger, R. L. 2009. Hunter-Gatherer For-aging: Five Simple Models. New York: EliotWerner Publications.
Bird, D. W. 1996. Intertidal Foraging Strate-gies Among the Meriam of the Torres StraitIslands, Australia: An Evolutionary EcologicalApproach to the Ethnoarchaeology of TropicalMarine Subsistence. Ph.D. dissertation, Univer-sity of California, Davis.
Bird, D. W. and R. Bliege Bird. 1997. Contem-porary shellfish gathering strategies among theMeriam of the Torres Strait Islands, Australia:Testing predictions of a central place foragingmodel. Journal of Archaeological Science24(1):39–63.
Bird, D. W. and R. Bliege Bird. 2000. The eth-noarchaeology of juvenile foragers: Shellfishingstrategies among Meriam children. Journal ofAnthropological Archaeology 19(4):461–476.
Bird, D. W. and J.F. O’Connell. 2006. Be-havioral ecology and archaeology. Journal ofArchaeological Research 14:143–188.
Bird, D. W., R. Bliege Bird, and B.F. Codding.2009. In pursuit of mobile prey: Martu huntingstrategies and archaeofaunal interpretation.American Antiquity 74(1):3–29.
Bird, D. W., R. Bliege Bird and Richardson, J.L. 2004. Meriam ethnoarchaeology: Shellfishingand shellmiddens. Memoirs of the QueenslandMuseum, Cultural Heritage Series 3(1):183–197.
Bird, D. W., J.L. Richardson, P.M. Veth, andA.J. Barham. 2002. Explaining shellfish vari-ability in middens on the Meriam Islands, TorresStrait, Australia. Journal of ArchaeologicalScience 29(5):457–469.
Blurton Jones, N. 1986. Bushman Birth Spac-ing: A Test for Optimal Interbirth Intervals.Ethnology and Sociobiology 7:91–105.
Bronk Ramsey, C. 2009. Bayesian analysis ofradiocarbon dates. Radiocarbon 51: 337–360.
Charnov, E. L. and G.H. Orians. 2006. Op-timal foraging: Some theoretical explorations.Mimeo: Department of Biology, Universityof Utah, Salt Lake City, UT. Digital ver-sion of 1973 edition. Available online athttp://repository.unm.edu/handle/1928/1649.
Codding, B.F., R. Bliege Bird and D.W.Bird. 2011. Provisioning offspring and others:risk-energy trade-offs and gender differencesin hunter -gatherer foraging strategies. Pro-ceedings of the Royal Society, B. 278: 2502–2509.
Dortch, J. 2004. Palaeo-environmental changeand the persistence of human occupation insouth-western Australian forests. British Ar-chaeological Reports. Oxford: Archaeopress.
Erlandson, J. 1988. Of Millingstones andMollusks: Cultural Ecology of Early HoloceneHunter-Gatherers on the California Coast. PhDDissertation. Santa Barbara: University ofCalifornia.
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
Gadgil, M. and W. H. Bossert. 1970. Lifehistorical consequences of natural selection. TheAmerican Naturalist, 104(935): 1–24.
Gosden, C. and N. Robertson. 1991. Models forMatenkupkum: interpreting a late Pleistocenesite from southern New Ireland, paupa newguinea. In Report of the Lapita HomelandProject, number 20 in Occasional Papers, pages20–45. Department of Prehistory, ResearchSchool of Pacific Studies, Canberra: AustralianNational University.
Hawkes, K. J. F. O’Connell and N. G. BlurtonJones. 1995. Hadza Children’s Foraging:Juvenile Dependency, Social Arrangements, andMobility among Hunter-Gatherers. CurrentAnthropology 36(4):688–700.
Kennedy, M. 2005. An Investigation ofHunter-Gatherer Shellfish Foraging Practices:Archaeological and Geochemical Evidence fromBodega Bay, California. PhD Dissertation.Davis: University of California.
Klein, R.G. and T. E. Steele. 2013. Archaeo-logical shellfish size and later human evolutionin Africa. Proceedings of the National Academyof Sciences, published ahead of print June 17,2013, doi:10.1073/pnas.1304750110.
Kramer, K. 2011. The evolution of humanparental care and recruitment of juvenile help.Trends in Ecology and Evolution 26(10):533–540.
Leavesley, M. G. 2004. Trees to the Sky:Prehistoric Hunting in New Ireland, PaupaNew Guinea. PhD Dissertation. Canberra:Australian National University.
Leavesley, M. G. and J. Allen. 1998. Dates,disturbance and artefact distributions: Anotheranalysis of Buang Merabak, a pleistocene site onNew Ireland, Papua New Guinea. Archaeologyin Oceania 33:63–82.
Meehan, B. 1982. Shell Bed to Shell Midden.Canberra: Australian Institute of AboriginalStudies.
Metcalfe, D. and K. R. Barlow. 1992. A Modelfor Exploring the Optimal Trade-off betweenField Processing and Transport. AmericanAnthropologist 94:340–356.
Morse, K. 1988. Mandu mandu creek rockshel-ter: Pleistocene human coastal occupation ofnorth west cape, Western Australia. Archaeol-ogy in Oceania 23(3):81–88.
Morse, K. 1993. New radiocarbon dates fromthe north west cape, Western Australia: Apreliminary report. In Sahul in Review: Pleis-tocene Archaeology in Australia, New Guineaand Island Melanesia, (M.A. Smith, M. Spriggs,and B. Fankhauser, eds): 155–163. The Aus-tralian National University, Canberra.
Munro, J. L. 1993. Giant clams. In NearshoreMarine Resources of the South Pacific (A.Wrightand L. Hill, eds): 431–449. Institute for PacificStudies.
Nash, W. J. 1993. Trochus. In NearshoreMarine Resources of the South Pacific (A.Wright and L. Hill, eds): 451–495. Institute forPacific Studies.
O’Connell, J. F. 1995. Ethnoarchaeology Needsa General Theory of Behavior. Journal ofArchaeological Research 3(3):205–255.
O’Connell, J. F. and J. Allen. 2004. Dating thecolonization of Sahul (Pleistocene Australia-NewGuinea): A review of recent research. Journalof Archaeological Science 31(8):835–853.
O’Connell, J. F. and J. Allen. 2007. Pre-LGMSahul (Pleistocene Australia-New Guinea) andthe archaeology of early modern humans. InRethinking the Human Revolution (P. Mellars,K. Boyle, O. Bar-Yosef, and C. Stringer, eds):
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
395–410. McDonald Institute for ArchaeologicalResearch, Cambridge University, Cambridge.
O’Connell, J. F. and J. Allen. 2012. The restau-rant at the end of the universe: modeling thecolonization of Sahul. Australian Archaeology74:5–17.
O’Connell, J. F., J. Allen, and K. Hawkes. 2010.Pliestocene Sahul and the origins of seafaring.In The Global Origins and Development ofSeafaring (A. Anderson, J. Barrett, and K.Boyle, eds): 57–68. The McDonald Institute forArchaeological Research, Cambridge University,Cambridge.
O’Connor, S. 1999. 30,000 Years of Aboriginaloccupation: Kimberley, North West Australia.Number 14 in Terra Australis. ANH Pub-lications and The Centre for ArchaeologicalResearch, Canberra: The Australian NationalUniversity.
Orians, G. H. and N.E. Pearson. 1979. On thetheory of central place foraging. In Analysis ofEcological Systems (D.J. Horn, G. R. Stairs andR.T. Mitchell, editors): 155–177. Ohio StateUniversity Press.
Petchey, F., M. Phelan and J.P.White. 2004.New ∆R values for the southwest Pacific Ocean.Radiocarbon 46: 1005–1014.
Peters, R. H. 1983. The Ecological Implicationsof Body Size. Cambridge University Press.
Pope, K.O. and J.E. Terrell. 2008. Environ-mental setting of human migrations in thecircum-Pacific region. Journal of Biogeography35:1–21.
Przywolnik, K. 2008. Long-term transitionsin hunter-gatherers of coastal northwesternAustralia. In Desert Peoples: ArchaeologicalPerspectives (P. Veth, M. Smith and P. Hiscock,eds): 117–205. Oxford: Blackwell.
R Development Core Team. 2013. R: ALanguage and Environment for StatisticalComputing. R Foundation for Statistical Com-puting, Vienna, Austria.
Reimer, P. J., M. G. L. Baillie, E. Bard, A.Bayliss, J.W. Beck, P.G. Blackwell, C. BronkRamsey, C.E. Buck, G.S. Burr, R.L. Edwards,M. Friedrich, P.M. Grootes, T.P. Guilderson, I.Hajdas, T.J. Heaton, A.G. Hogg, K.A. Hughen,K.F. Kaiser, B. Kromer, F.G. McCormac, S.W.Manning, R.W. Reimer, D.A. Richards, J.R.Southon, S. Talamo, C.S.M. Turney, J. van derPlicht, and C.E. Weyhenmeyer. 2009. IntCal09and Marine09 radiocarbon age calibrationcurves, 0-50,000 years cal BP. Radiocarbon, 51:1111–1150.
Rosenfeld, A. 1997. Excavation at Buang Mer-abak, Central New Ireland. The Indo-PacificPrehistory Association Bulletin (The ChangMai Papers, Vol. 3) 16:213–224.
Thomas, F. R. 2007. The behavioral ecology ofshellfish gathering in Western Kiribati, Microne-sia 1: Prey choice. Human Ecology 35:179–194.
Veth, P., K. Alpin, L. Wallis, T. Manne, T.Pulsford, E. White, and A. Chappell. 2007. TheArchaeology of Montebello Islands, North-WestAustralia. BAR International Series 1668.British Archaeological Reports.
Whitaker, A. 2008. The role of human predationin structuring prey populations in NorthwesternCalifornia. PhD Dissertation. Davis: Universityof California.
Whitaker, A. and B.F. Byrd. 2012. Boat-basedforaging and discontinuous prehistoric redabalone exploitation. Journal of Anthropologi-cal Archaeology 31:196–214.
Wickler, S. 2001. The Prehistory of Buka: Step-ping Stone Island in the Northern Solomons.Terra Australis. Department of Archaeology
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript

Codding, O’Connell & Bird Shellfishing and the Colonization of Sahul
and Natural History, and Centre for Archaeo-logical Research, Australian National University.
Williams, A. N. 2013. A new population curvefor prehistoric Australia. Proceedings of theRoyal Society, B.doi: 10.1098/rspb.2013.0486.
Yamaguchi, M. 1993. Green snail. In NearshoreMarine Resources of the South Pacific (Wright,A. and L. Hill, eds): 488–511. Institute forPacific Studies.
J. Isl. Coast. Archaeol. 9(2):238–252 DOI:10.1080/15564894.2013.848958
UU
IR Author M
anuscript UU
IR Author M
anuscript
University of Utah Institutional Repository Author Manuscript





























