Embed Size (px)
Citation preview

[CANCER RESEARCH 53, 5589-5591, December 1, 1993]
Advances in Brief
APC Gene Messenger RNA: Novel Isoforms That Lack Exon 7
Masanobu Oshima, Hiroko Sugiyama, Kyoko Kitagawa, and Makoto Taketo ~ Genetic Animal Experiment Laboratory, Banyu Tsukuba Research Institute (Merck), 30kubo, Tsukuba, 300-33, Japan
Abstract
The APC gene at human chromosome 5q21 is responsible for familial adenomatous polyposis coli. Furthermore, sporadic cancers of not only colon but also other digestive organs often contain mutations in the APC gene. A dominant mouse mutation Min that was generated by chemical mutagenesis and causes polyposis in the digestive tract is in the mouse homologue of the human APC gene. The APC mRNA is generated from 15 exons. Two mRNA isoforms were reported which are produced by alter- native splicing in the 9th exon. Here, we report novel mRNA isoforms that lack the 7th exon in both mouse and human cells.
Introduction
The APC gene (for adenomatous polyposis coli) at human chro- mosome 5q21 was originally identified as one of the genes affected in patients of familial adenomatous polyposis coli. The gene appears responsible for the dominantly inherited disease characterized by mul- tiple benign colorectal tumors that often develop into malignant car- cinomas (1-4). In addition, many sporadic cancers of not only colon but also other organs of the digestive system such as stomach and esophagus turned out to contain mutations in the APC gene (5-8). At the same time, a dominant mouse mutation Min (for multiple intestinal neoplasia) that had been generated by chemical mutagenesis and causes polyposis in the digestive tract was located in the gene ho- mologous to the human APC gene (9, 10). The APC mRNA is gen- erated from 15 exons and encodes a protein of 2843 amino acids in humans and 2845 residues in mice. Exons 1 to 14 combined, occupy the NH2-terminal quarter, whereas exon 15 alone occupies three- fourths of the molecule at the COOH terminal (3, 4). The amino acid sequence of the APC protein deduced from the cDNA 2 sequence indicates a hydrophilic protein that contains heptad repeats, rat-2 homology, muscarinic acetylcholine receptor-G protein coupling ho- mology, and 20 amino acid repeats (1, 3). However, the physiological function(s) of this huge protein remains to be investigated. Although APC mRNA is detected in a wide variety of tissues (3), immunohis- tochemical analysis using antibodies against partial APC protein showed an intensive staining in the basolateral margins of the colonic epithelial cells, especially marked in the upper portions of crypts (11). Two mRNA isoforms were reported which are produced by alternative splicing in exon 9; one with the full length exon of 380 ntd and the other with a partial exon of 77 ntd (exon 9A) (3). Other polymor- phisms of APC mRNA were also reported that involve noncoding exons upstream of exon 1 and isoform mRNAs that are spliced into a downstream gene SRP19 (12). Here, we report novel mRNA isoforms that lack exon 7 sequence both in various mouse tissues and cell lines as well as in human cell lines.
Materials and Methods
Standard procedures (13, 14) were used unless otherwise stated.
Received 7/19/93; accepted 10/19/93. The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom requests for reprints should be addressed. 2 The abbreviations used are: cDNA, complementary DNA: ntd, nucleotide(s); PCR,
polymerase chain reaction; RT, reverse transcriptase.
Animals, Cell Lines, and RNA Preparation. Total RNA was isolated from various mouse organs/tissues of adult ICR mice (Charles River, Japan), and from mouse and human cell lines by the guanidine isothiocyanate method (15). Mouse embryonic stem cell line AB-1 (16), embryonal carcinoma cell line F9 (17), and human teratoma cell line NTera-2 (18) have been described previously, respectively. Other human cell lines, A431 epiderrnoid carcinoma (19), HT-1080 fibrosarcoma (20), and HLC-1 lung adenocarcinoma 3 cell lines were obtained from RIKEN Cell Bank, Tsukuba, Japan.
RT-PCR. Reverse transcription reaction was performed using 10 /~g of total RNA and 100 pmol of a synthetic oligonucleotide primer (5'-GAA- AGCTTCATFAGAACACAC-3', complimentary to ntd 1344-1364 in exon 10) in 40 Izl of reverse transcription buffer at 42~ for 1 h with 200 units (1/,1) of Superscript RT (BRL) and diluted with 60/xl of H20. Standard PCR was then performed with 10 t~l of the reverse transcription product, 100 pmol of an exon 6 primer (5'-ATAGCCAGGATCCAGCAAAT-3', for ntd 646-665) and 0.2 mM deoxynucleotide triphosphates, 0.5 txl (2.5 units) of Taq polym- erase (Takara, Japan) in 100 p~l of PCR buffer (Perkin-Elmer/Cetus GeneAmp kit; 10 • = 500 mM KCI/100 mM Tris-HC1, pH 8.3/15 mM MgC12/0.1% gelatin). PCR was performed under the following condition: 95~ 1 min; 60~ 1 min; 35 cycles of 60~ 1 rain; 72~ 1 min; and 95~ 1 rain; and 72~ 10 rain. Ten ill of the PCR product were loaded onto each lane of 2% agarose gel and electrophoresed. Although the exon-intron structure has been published only for the human gene (3), it is reasonable to assume the same structure in the mouse because the cDNA sequences are almost identi- cal between the two species except Exon 7 and parts of Exon 15 (see "Discussion").
DNA Sequencing. The cDNA preparations amplified by PCR were di- gested with BamHI and HindIII and subcloned into pUC19 (Takara). The sequences were determined and compared with that of mouse APC cDNA (10) (downloaded from GenBank; accession no. M88127).
Amino Acid Sequence Analysis. Computer-assisted statistical analysis of the similarity between mouse and human APC amino acid sequences was performed by the Best Fit program of GCG package (Version 7.0) from the University of Wisconsin, Madison, WI, on a MicroVAX II computer (Digital Equipment Corporation, Boston, MA).
Results
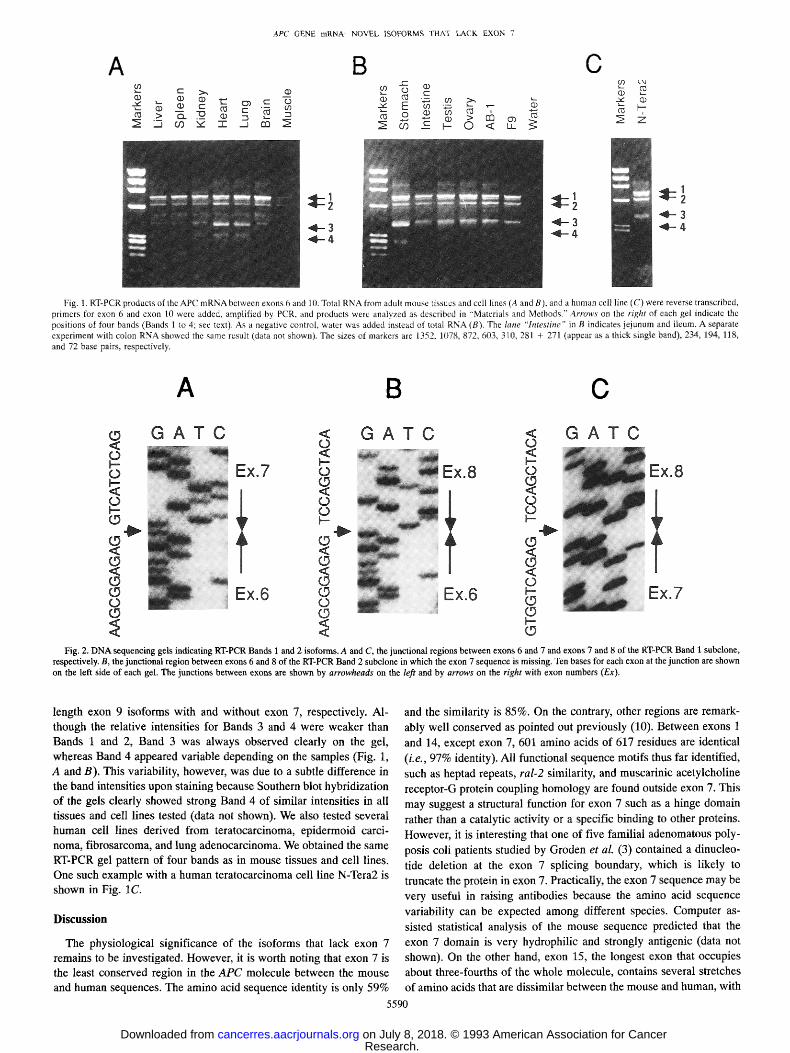
During the course of RT-PCR studies of the APC mRNA in mouse liver, we detected four strong bands instead of one, when PCR primers for exons 6 and 10 were used (Fig. 1,4, Bands 1-4). Because such multiple bands were not observed for other regions of the cDNA (i.e., between exons 1 and 6 or exons 10 and 15; data not shown), we cloned these bands in a plasmid vector and determined their sequences. Fig. 2 shows the DNA sequencing gels for cloned Bands 1 and 2. While Band 1 contained the exon 7 sequence (Fig. 2, A and C), the sequence of exon 6 was directly followed by that of exon 8 in the Band 2 clone (Fig. 2B). On the other hand, the RT-PCR Bands 3 and 4 contained exon 9A in place of exon 9. Like Bands 1 and 2, Bands 3 and 4 represented isoforms with and without exon 7 sequence, respectively (data not shown). These results are summarized diagrammatically in Fig. 3. To determine whether these isoforms with and without exon 7 are expressed differentially in a tissue-specific manner, we examined by RT-PCR various adult mouse tissues as well as embryonic cell lines. However, all samples tested showed essentially the same pat- tern. They all contained intense Bands 1 and 2, representing full-
3 T. Okabe, personal communication.
5589
Research. on July 8, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
A C
l/) ~ "r" ..j
APC GENE mRNA: NOVEL ISOFORMS THA'I LACK EXON 7
B , , , - g o
�9 to to -= "- ...
m ~ ~ ~ _ 0 < w
C o 'r O k,. I---
, 9 t -3 . ~ - - 4
4 - 3 . 4 - 4 .
1 l=z 4 - 3 .,,111-4.
Fig. 1. RT-PCR products of the APC mRNA between exons 6 and 10. Total RNA from adult mouse tissues and ccll lines (A and B), and a human cell line (C) were reverse transcribed, primers for exon 6 and exon 10 were added, amplified by PCR, and products were analyzed as described in "Materials and Methods." Arrows on the right of each gel indicate the positions of four bands (Bands 1 to 4; see text). As a negative control water was added instead of total RNA (B). The lane "Intestine" in B indicates jejunum and ileum. A separate experiment wilh colon RNA showed the same result (data not shown). The sizes of markers are 1352, 1078, 872, 603, 310, 281 + 271 (appear as a thick single band), 234, 194, 118, and 72 base pairs, respectively.
A B C
o G A T C
o
o
O
O
41-
G A T C < G A T C
E x . 7 o E x . 8
t t o
<
E x . 6 E x . 6
<
O
O <
(.9
E x . 8
t E x . 7
Fig. 2. DNA sequencing gels indicating RT-PCR Bands 1 and 2 isoforms. A and C, the junctional regions between exons 6 and 7 and exons 7 and 8 of the RT-PCR Band 1 subclone, respectively. B, the junctional region between exons 6 and 8 of the RT-PCR Band 2 subclone in which the exon 7 sequence is missing. Ten bases for each exon at the junction are shown on the left side of each gel The junctions between exons are shown by arrowheads on the left and by arrows on the right with exon numbers (Ex).
length exon 9 isoforms with and without exon 7, respectively. Al- though the relative intensities for Bands 3 and 4 were weaker than Bands 1 and 2, Band 3 was always observed clearly on the gel, whereas Band 4 appeared variable depending on the samples (Fig. 1, A and B). This variability, however, was due to a subtle difference in the band intensities upon staining because Southern blot hybridization of the gels clearly showed strong Band 4 of similar intensities in all tissues and cell lines tested (data not shown). We also tested several human cell lines derived from teratocarcinoma, epidermoid carci- noma, fibrosarcoma, and lung adenocarcinoma. We obtained the same RT-PCR gel pattern of four bands as in mouse tissues and cell lines. One such example with a human teratocarcinoma cell line N-Tera2 is shown in Fig. 1C.
Discuss ion
The physiological significance of the isoforms that lack exon 7 remains to be investigated. However, it is worth noting that exon 7 is the least conserved region in the APC molecule between the mouse and human sequences. The amino acid sequence identity is only 59%
and the similarity is 85%. On the contrary, other regions are remark- ably well conserved as pointed out previously (10). Between exons 1 and 14, except exon 7, 601 amino acids of 617 residues are identical
(i .e. , 97% identity). All functional sequence motifs thus far identified, such as heptad repeats, ra l -2 similarity, and muscarinic acetylcholine receptor-G protein coupling homology are found outside exon 7. This may suggest a structural function for exon 7 such as a hinge domain rather than a catalytic activity or a specific binding to other proteins.
However, it is interesting that one of five familial adenomatous poly- posis coli patients studied by Groden et al. (3) contained a dinucleo- tide deletion at the exon 7 splicing boundary, which is likely to truncate the protein in exon 7. Practically, the exon 7 sequence may be very useful in raising antibodies because the amino acid sequence variability can be expected among different species. Computer as- sisted statistical analysis of the mouse sequence predicted that the exon 7 domain is very hydrophilic and strongly antigenic (data not shown). On the other hand, exon 15, the longest exon that occupies about three-fourths of the whole molecule, contains several stretches of amino acids that are dissimilar between the mouse and human, with
5590
Research. on July 8, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

APC GENE mRNA: NOVEL 1SOFORMS THAT LACK EXON 7
RT-PCR
Band 1
Band 2
Exon6 Exon7 Exon8 Exon9(gA)
0 ExonlO
Band 3
Band 4
Fig. 3. Schematic diagram showingAPC mRNA isoforms in the region between Exons 6 and 10. Exon lengths are drawn to scale whereas the introns are not. Arrows above exon 6 and beneath exon 10 indicate the positions of the PCR primers.
the rest o f the s e q u e n c e b e i n g a lmos t ident ica l . Fu r the r inves t iga t ion
such as gene k n o c k o u t e x p e r i m e n t s i n v o l v i n g e x o n 7 shou ld he lp
e luc ida te its ro le in p h y s i o l o g y and po lypos i s .
Acknowledgments
We thank Drs. T. Nagata and Y. Saga for total RNA preparations from various tissues and cell lines of the mouse and human, respectively. We also
thank Dr. S. Nishimura for discussion and critical reading of the manuscript.
References 1. Kinzler, K. W., Nilbert, M. C., Su, L-K., Vogelstein, B., Bryan, T. M., Levy, D. B.,
Smith, K. J., Preisinger, A. C., Hedge, P, McKechnie, D., Finniear, R., Markham, A., Groffen, J., Boguski, M. S., Altschut, S. E, Horii, A., Ando, H., Miyoshi, Y., Miki, Y., Nishisho, I., and Nakamura, Y. Identification of FAP locus genes from chromosome 5q21. Science (Washington DC), 253: 661--665, 1991.
2. Nishisho, I., Nakamura, Y., Miyoshi, Y., Mild, Y., Ando, H., Horii, A., Koyama, K., Utsunomiya, J., Baha, S., Hedge, P., Markham, A., Krush, A. J., Petersen, G., Hamil- ton, S. R., Nilbert, M. C., Levy, D. B., Bryan, T. M., Preisinger, A. C., Smith, K. J., Su, L-K., Kinzler, K. W., and Vogelstein, B. Mutations of chromosome 5q21 genes in FAP and colorectal cancer patients. Science (Washington DC), 253: 665-669, 1991.
3. Groden, J., Thliveris, A., Samowitz, W., Carlson, M., Gelbert, L., Albertsen, H., Joslyn, G., Stevens, J., Spirio, L., Robertson, M., Sergeant, L., Krapcho, K., Wolff, E., Burt, R., Hughes, J. P., Warrington, J., McPherson, J., and White, R. Identification and characterization of the familial adenomatous polyposis coli gene. Cell, 66: 589--600, 1991.
4. Joslyn, G., Carlson, M., Thtiveris, A., Albertson, H., Getbert, L., Samowitz, W., Groden, J., Stevens, J., Spirio, L., Robertson, M., Sergeant, L., Krapcho, K., Wolff, E., Burt, R., Hughes, J. E, Warrington, J., McPhearson, J., Wasmuth, J., Paslier, D. L, Abderrahim, H., Cohen, D., Leppert, M., and White, R. Identification of deletion mutations and three new genes at the familial polyposis locus. Cell, 66: 601-613, 1991.
5. Horii, A., Nakatsuru, S., Miyoshi, Y., Ichii, S., Nagase, H., Kato, Y., Yanagisawa, A., and Nakamura, Y. Multiple forms of the APC gene transcripts and their tissue-specific expression. Cancer Res., 52: 3231-3233, 1992.
6. Powell, S. M., Zilz, N., Beazer-Barclay, Y., Bryan, T. M., Hamilton, S. R., Thibodeau, S. N., Vogelstein, B., and Kinzler, K. W. APC mutation occur early during colorectal tumorigenesis. Nature (Lond.), 359: 235-237, 1992.
7. Boynton, R. E, Bloant, E L., Yin, J., Brown, V. L., Huang, Y., Tong, Y., McDaniel, T., Newkirk, C., Resau, J. H., Raskind, W. H., Haggitt, R. C., Reid, B. J., and Mettzer, S. J. Loss of heterozygosity involving the APC and MCC genetic loci occurs in the majority of human esophageal cancers. Proc. Natl. Acad. Sci. USA, 89: 3385-3388, 1992.
8. Horii, A., Haktsuru, S., Miyoshi, Y., Ichii, S., Nagase, H., Ando, H., Yanagisawa, A., Tsuchiya, E., Kato, Y., and Nakamura, Y. The APC gene, responsible for familial adenomatous polyposis, is mutated in human gastric cancer. Cancer Res., 52: 6696- 6698, 1992.
9. Moser, A. R., Dove, W. E, Rothe, K. A., and Gordon, J. I. The M/n (multiple intestinal neoplasia) mutation: its effect on gut epithelial cell differentiation and interaction with a modifier system. J. Cell Biol., 116: 1517-1526, 1992.
10. Su, L-K., Kinzler, K. W., Vogelstein, B., Preisinger, A. C., Moser, A. R., Luongo, C., Gould, K. A., and Dove, W. E Multiple intestinal neoplasia caused by a mutation in the murine homolog of the APC gene. Science (Washington DC), 256: 668--671, 1992.
11. Smith, K. J., Johnson, K. A., Bryan, T. M., Hill, D. E., Markowitz, S., Willson, J. IC V., Paraskeva, C., Petersen, G. M., Hamilton, S. R., Vogelstein, B., and Kinzler, K. W. The APC gene product in normal and tumor cells. Proc. Natl. Acad. Sci. USA, 90: 2846-2850, 1993.
12. Horii, A., Nakatsuru, S., Ichii, S., Nagase, H., and Nakamura, Y. Multiple forms of the APC gene transcripts and their tissue-specific expression. Hum. Mol. Genet., 2: 283- 287, 1993.
13. Ausubet, E M., Brent, R., Kingston, R. E., Moore, D. D., and Smith, J. A. Current Protocols in Molecular Biology. Wiley, New York, 1989.
14. Sambrook, J., Fritsch, E. E, and Maniatis, T. (eds.). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1989.
15. Chirgwin, J. M., Przybyla, A. E., MacDonald, R. J., and Rutter, W. J. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochem- istry, 18: 5294-5299, 1979.
16. McMahon, A. E, and Bradley, A. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell, 62: 1073-1085, 1990.
17. Taketo, M., Gilboa, E., and Sherman, M. I. Isolation of embryonal carcinoma cell lines that express integrated recombinant genes flanked by the Moloney murine leukemia virus long terminal repeat. Proc. Natl. Acad. Sci. USA, 82: 2422-2426, 1985.
18. Nagata, T., Segars, J. H., Levi, B-Z., and Ozato, K. Retinoic acid-dependent trans- activation of major histocompatibility complex class I promoters by the nuclear hormone receptor H-2RIIBP in undifferentiated embryonal carcinoma cells. Proc. Natl. Acad. Sci. USA., 89: 937-941, 1992.
19. Giard, D. J., Aaronson, S. A, Todaro, G. J., Kersey, J. H., Dosik, H., and Parks, W. P= In vitro cultivation of human tumors: establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 51: 1417-1423, 1973.
20. Rasheed, S., Nelson-Rees, W. A., Toth, E. M., Arnstein, P., and Gardner, M. B. Characterization of a newly derived human sarcoma cell line. Cancer 33: 1027-1033, 1974.
5591
Research. on July 8, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

1993;53:5589-5591. Cancer Res Masanobu Oshima, Hiroko Sugiyama, Kyoko Kitagawa, et al.
Gene Messenger RNA: Novel Isoforms That Lack Exon 7APC
Updated version
http://cancerres.aacrjournals.org/content/53/23/5589
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/53/23/5589To request permission to re-use all or part of this article, use this link
Research. on July 8, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from







![· Gefitinib Gefitinib 1. Non-small cell lung cancer EGFR DNA EGF-R exon 19 deletion, exon 21 [1.858R] substitution mutations, L861Q G719X EGFR exon 20](https://img.pdfslide.us/doc/110x75/5e51ddba1b664701f40175b0/gefitinib-gefitinib-1-non-small-cell-lung-cancer-egfr-dna-egf-r-exon-19-deletion.jpg)


![CRISPR/Cas9-mediated genome editing induces exon skipping ... · HeLa cells can cause skipping of exon 3, exon 4, or exons 3, 4, and 5 [18]. We also detected infrequent exon skipping](https://img.pdfslide.us/doc/110x75/60db8f117fb86d112c69c947/crisprcas9-mediated-genome-editing-induces-exon-skipping-hela-cells-can-cause.jpg)