Embed Size (px)
Citation preview

R E V I E W A R T I C L E
Candidaglabrata,Candida parapsilosis andCandida tropicalis:biology, epidemiology,pathogenicityandantifungal resistanceSonia Silva1, Melyssa Negri1, Mariana Henriques1, Rosario Oliveira1, David W. Williams2 &Joana Azeredo1
1Institute for Biotechnology and Bioengineering, Universidade do Minho, Campus de Gualtar, Braga, Portugal; and 2Tissue Engineering & Reparative
Dentistry, School of Dentistry, Heath Park, Cardiff, UK
Correspondence: Mariana Henriques,
Institute for Biotechnology and
Bioengineering, Universidade do Minho,
Campus de Gualtar 4710-057, Braga,
Portugal. Tel.: 1351 253 604 408; fax: 1351
253 604 429; e-mail: [email protected]
Received 29 December 2010; revised 31 March
2011; accepted 6 May 2011.
DOI:10.1111/j.1574-6976.2011.00278.x
Editor: Martin Kupiec
Keywords
Candida species; candidosis; epidemiology;
virulence factors; antifungal resistance.
Abstract
The incidence of infections caused by Candida species (candidosis) has increased
considerably over the past three decades, mainly due to the rise of the AIDS
epidemic, an increasingly aged population, higher numbers of immunocompro-
mised patients and the more widespread use of indwelling medical devices.
Candida albicans is the main cause of candidosis; however, non-C. albicans
Candida (NCAC) species such as Candida glabrata, Candida tropicalis and
Candida parapsilosis are now frequently identified as human pathogens. The
apparent increased emergence of these species as human pathogens can be
attributed to improved identification methods and also associated with the degree
of diseases of the patients, the interventions that they were subjected and the drugs
used. Candida pathogenicity is facilitated by a number of virulence factors, most
importantly adherence to host surfaces including medical devices, biofilm forma-
tion and secretion of hydrolytic enzymes (e.g. proteases, phospholipases and
haemolysins). Furthermore, despite extensive research to identify pathogenic
factors in fungi, particularly in C. albicans, relatively little is known about NCAC
species. This review provides information on the current state of knowledge on the
biology, identification, epidemiology, pathogenicity and antifungal resistance of
C. glabrata, C. parapsilosis and C. tropicalis.
Introduction
In last 30 years there has been a significant increase in the
incidence of fungal infections in humans (Lass-Florl, 2009).
Such infections may either be superficial, affecting the skin,
hair, nails and mucosal membranes, or systemic, involving
major body organs (Ruping et al., 2008). A number of
factors have been implicated in this increased occurrence of
fungal disease, but it is generally accepted that the increased
and widespread use of certain medical practices, such as
immunosuppressive therapies, invasive surgical procedures
and use of broad-spectrum antibiotics are significant (Sa-
maranayake et al., 2002; Hagerty et al., 2003; Kojic &
Darouiche, 2004).
Of the fungi regarded as human pathogens, the members
of the genus Candida are the most frequently recovered from
human fungal infection. The Candida genus contains over
150 heterogeneous species (Calderone, 2002), but only a
minority have been implicated in human candidosis. Addi-
tionally, it is known that approximately 65% of Candida
species are unable to grow at a temperature of 37 1C, which
precludes these species from being successful pathogens or
indeed commensals of humans (Calderone, 2002).
Of the Candida species isolated from humans, Candida
albicans is the most prevalent under both healthy and
disease (Calderone, 2002; Samaranayake et al., 2002) condi-
tions. However, while mycological studies have shown that
C. albicans represents over 80% of isolates from all forms of
human candidosis (Calderone, 2002) in the last two decades,
the number of infections due to non-C. albicans Candida
(NCAC) species has increased significantly (Kauffman et al.,
2000; Manzano-Gayosso et al., 2008; Ruan & Hsueh, 2009).
The apparent increased involvement of NCAC species in
human candidosis may partly be related to improvements in
diagnostic methods, such as the use of chromogenic media
with the ability to differentiate Candida species, as well as
c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
FEMS Microbiol Rev 36 (2012) 288–305
Final version published online 6 June 2011.
MIC
ROBI
OLO
GY
REV
IEW
S

the introduction of molecular techniques in the routine
diagnosis of fungemia (Liguori et al., 2009). However, the
high prevalence of NCAC species in disease could also be a
reflection of their inherently higher level of resistance to
certain antifungal drugs (Gonzalez et al., 2008) compared
with C. albicans, as this would promote their persistence,
possibly to the detriment of C. albicans, in mixed species
infections treated with traditional antifungal agents.
Unfortunately, compared with C. albicans there are
relatively few studies examining the virulence factors of
NCAC species. This review therefore provides information
on the current state of knowledge on the biology, identifica-
tion, epidemiology, pathogenicity and antifungal resistance
of Candida glabrata, Candida parapsilosis and Candida
tropicalis, three of the most frequent causes of candidosis
after C. albicans.
Biology of NCAC species
Candida comprises an extremely heterogeneous group of
fungal organisms that can all grow as yeast morphology.
Macroscopically, colonies of Candida, on the routinely used
Sabouraud dextrose agar (SDA), are cream to yellow in
colour. Depending on the species, colony texture may be
smooth, glistening or dry, or wrinkled and dull. Under
standard conditions with optimal nutrients, yeast grow in
log phase as budding cells (blastoconidia), which are sphe-
rical to oval in shape and are approximately 2–5� 3–7mm in
size (Fig. 1) (Larone, 2002). Moreover, certain species, such
as C. albicans and Candida dubliniensis, can produce a
filamentous type of growth, such as true hyphae (Fig. 1) or
more frequently, pseudohyphae (Fig. 1).
The distinction between hyphae and pseudohyphae is
related to the way in which they are formed. Pseudohyphae
are formed from yeast cells or hyphae by budding (Fig. 1),
but the new growth remains attached to the parent cell and
elongates, resulting in filaments with constrictions at the
cell–cell junctions. There are no internal cross walls (septa)
associated with pseudohyphae (Fig. 1). In comparison, true
hyphae are formed from yeast cells or even as branches of
existing hyphae. The development of true hyphae is initiated
by a ‘germ tube’ projection (Fig. 1), which elongates and
then branches with defined septa that divide the hyphae into
separate fungal units (Fig. 1).
Candida albicans and C. dubliniensis are truly poly-
morphic, due to their ability to form hyphae and/or
pseudohyphae, and these species are also referred to germ
tube positive, a diagnostic feature (Table 1) (Calderone,
2002). In contrast, C. glabrata is not polymorphic, growing
only as blastoconidia (yeast) (Table 1; Fig. 2). Historically,
this species was originally classified in the genus Torulopsis
due to its lack of pseudohyphal formation. However, in
1978, it was determined that the ability to form pseudohy-
phae was not a reliable distinguishing factor for members of
the genus Candida and it was proposed that Torulopsis
glabrata could be classified in the genus Candida, due to its
association with human infection (Fidel et al., 1999). With
regard to C. parapsilosis, this species does not produce true
hyphae, but can generate pseudohyphae that are character-
istically large and curved, and often referred to as ‘giant cells’
Fig. 1. Epifluorescence photocomposition of the different morphological growth forms of Candida albicans stained with calcofluor white: (A)
blastoconidia; (B1) reproduction by budding; (B2) germ tube formation; (C1) pseudohyphae formation; (C2) yeast form; (C3) hyphae formation.�Internal cross walls (septa).
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 289

(Fig. 2) (Larone, 2002; Trofa et al., 2008). In contrast, on
corn meal Tween 80 agar and at 25 1C after 72 h, C. tropicalis
produces oval blastospores, pseudohyphae depending on
some reports, true hyphae (Fig. 2) (Calderone, 2002; Larone,
2002; Yoshio & Kouji, 2006).
It should also be highlighted that C. glabrata cells
(1–4 mm) are noticeably smaller than the blastoconidia of
C. albicans (4–6 mm), C. tropicalis (4–8mm) and C. para-
psilosis (2.5–4mm) (Larone, 2002) (Table 1). On SDA (Fig.
2) C. glabrata forms glistening, smooth, and cream-co-
loured colonies, which are largely indistinguishable from
those of other Candida species except for their relative size,
which can be quite small (Fig. 2). Furthermore, C. para-
psilosis, when grown on SDA, yields white, creamy, shiny and
smooth/wrinkled colonies (Fig. 2). On the same medium, C.
tropicalis forms colonies that are cream-coloured with a
slightly mycelial border (Fig. 2) (Calderone, 2002).
Concerning the biochemistry of Candida species, C.
glabrata ferments and assimilates only glucose and trehalose,
which contrasts with C. albicans, which ferments and/or
assimilates a number of sugars with the notable exception of
sucrose (Odds, 1988). Additionally, C. tropicalis has the
ability to ferment and assimilate sucrose and maltose
(Martin, 1979). Interestingly, C. parapsilosis was firstly
classified as a species of Monilia, due to its inability to
ferment maltose (Odds, 1988; Trofa et al., 2008).
A main distinguishing genetic characteristic of C. glabrata
is that it has a haploid genome, in contrast to the diploid
genome of C. albicans and several other NCAC species (Fidel
et al., 1999). Genetically, C. tropicalis has the highest
similarity to C. albicans, and C. glabrata the least (Butler
et al., 2009). It is through the advent of molecular genetics
that new identification methods for Candida have been
developed, leading to the identification of new species
together with their increased recognition in human infec-
tion. Before 2005, C. parapsilosis was separated into three
groups (I–III), but further studies revealing genomic differ-
ences that have led to the separation of these groups into
closely related, but distinct species, namely, C. parapsilosis,
Candida orthopsilosis and Candida metapsilosis (Tavanti
et al., 2005; Gacser et al., 2007a, b).
Laboratory identification of NCAC species
The laboratory diagnosis of candidoses continues to be
problematic. Microbiological confirmation can be difficult
Fig. 2. Candida species macroscopic colonies on cornmeal Tween 80 and microscopy structure on SDA. Microscopic structures: (a) Candida glabrata;
(b) Candida parapsilosis; (c) Candida tropicalis; macroscopic colonies: (d) C. glabrata; (e) C. parapsilosis; (f) C. tropicalis.
Table 1. Morphological characteristics of Candida albicans, Candida tropicalis, Candida parapsilosis and Candida glabrata species
Species Germ tube production Hyphae/pseudohyphae Yeasts size (mm) CHROM-agar colony colour
C. albicans 1 1/1 4–6�6–10 Blue-green
C. tropicalis � � /1 4–8�5–11 Dark blue
C. parapsilosis � � /1 2.5–4�2.5–9 White
C. glabrata � � /� 1–4 White, Pink-purple
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
290 S. Silva et al.

as blood cultures can be negative in up to 50% of autopsy-
proven cases of deep-seated candidoses, or may only become
positive late in the infection (Ellepola & Morrison, 2005).
Positive cultures from urine or mucosal surfaces do not
necessarily indicate invasive disease although may occur
during systemic infection (Ellepola & Morrison, 2005).
Furthermore, differences in virulence between Candida
species as well as in their susceptibility to antifungal drugs
make identification important for clinical management.
Laboratory diagnosis has improved with the advent of
new methods for Candida isolation and identification.
Technologies such as species-specific FISH (Alexander
et al., 2006), antibody and antigen detection (Pfaller, 1992;
Ellepola & Morrison, 2005) and molecular approaches for
typing and detection of fungal pathogens (Ellepola &
Morrison, 2005) have all been used successfully. However,
many of these approaches have not yet been standardized or
validated in large clinical trials and therefore are not widely
used in clinical laboratory settings (Ellepola & Morrison,
2005). Laboratory surveillance of ‘at-risk’ patients could
result in earlier initiation of antifungal therapy if sensitive
and specific diagnostic tests, which are also cost effective,
become widely available.
The clinical symptoms of fungemia are not indicative of
particular Candida species and may be induced by other
microorganisms. The laboratory identification of Candida is
therefore essential for establishing a definitive diagnosis. A
standard approach to the laboratory diagnosis generally
involves nonmolecular methods, although PCR is increas-
ingly being used.
Non-PCR based methods of Candidaidentification
CHROMagars Candida (CHROMagars, Paris, France), is a
relatively new differential agar medium for Candida species
identification and has been particularly useful in the pre-
sumptive identification of C. albicans, C. tropicalis and
Candida krusei upon primary culture of clinical specimens.
On CHROMagars Candida, C. glabrata colonies appear
white, pink or purple in contrast to C. albicans colonies,
which are blue-green, while C. parapsilosis colonies are white
and C. tropicalis dark blue (Table 1). Moreover, it is possible
to detect coinfection with different Candida species on
primary culture plates and this can have importance in
infection management strategies (Ellepola & Morrison,
2005; Furlaneto-Maia et al., 2007).
After Candida isolation, species can also be identified by
carbohydrate assimilation and fermentation tests as well as
morphological characteristics such as germ tube and chla-
mydospore development (Fig. 2). In addition, more rapid
and less laborious phenotypic identification methods have
become available. Perhaps the most widely used methods for
Candida species identification are those based on the format
of carbohydrate assimilation and/or enzyme detection with-
in plastic wells of commercially available kits. Examples of
such biochemical tests include the API 20C AUX (API
Candida) Auxacolor (Bio-Rad) and the Uni-Yeast-Tek kit
(Ellepola & Morrison, 2005). These tests generate reliable
identification for the most common species of Candida,
while identification of other Candida species may not be so
accurate. For example, the differentiation of C. dubliniensis
from C. albicans often requires the use of supplemental
biochemical or morphological tests for definitive identifica-
tion (Verweij et al., 1999; Ellepola & Morrison, 2005).
Additional methods for Candida species identification
include tests that allow the detection of an isolate in 1 day,
such as the RapID Yeast Plus System (Innovative Diagnostic
Systems, Norcross), the Fongiscreen test (Sanofi Diagnostics
Pasteur, France) and the automated Rapid Yeast Identifica-
tion Panel (Dade Microscan). However, as mentioned
above, most of these tests tend to be most accurate for the
identification of the more frequently encountered yeast
pathogens (Ellepola & Morrison, 2005).
The diagnosis of invasive candidosis should include a
collection of adequate volumes of blood and an agar-based
blood culture method for optimal detection of candidemia
(Pfaller, 1992). Several advances in blood culturing techni-
ques have been developed, which appear to have improved
the sensitivity and/or reduced the time required to obtain a
positive blood culture. Two automated methods for mon-
itoring of blood culture bottles, based on colour (BacT/
ALERT 3D, Organon Teknika Corp., Durham, NC) and
fluorescence (BACTEC 9240, Becton Dickinson), have been
developed recently (Ellepola & Morrison, 2005).
The identification of typical blastospores and pseudohy-
phae of Candida species on microscopic examination of
tissue remains the unequivocal standard for the diagnosis of
invasive or disseminated candidoses. Unfortunately, the
usefulness of this approach is frequently limited by sampling
problems (isolation source and sample size) (Pfaller, 1992).
The use of fluorescent antibody, acridine orange or calco-
fluor-white staining (Pfaller, 1992; Ellepola & Morrison,
2005) may enhance the sensitivity of microscopic examina-
tion. However, the production of fluorescent antibodies
specific for the identification of individual Candida species
has proved to be extremely difficult.
A relatively recent laboratory method based on PNA FISH
targeting the 26S rRNA gene allows the reliable detection of C.
albicans from NCAC species, within 2.5 h of yeast growth
detection in blood culture, with a sensitivity of 99% and
specificity of 100% (Rigby et al., 2002). According to recent
studies PNA FISH also results in substantial cost savings for
hospitals, making the method both an effective and affordable
one for the laboratory diagnosis of candidoses (Rigby et al.,
2002; Ellepola & Morrison, 2005; Alexander et al., 2006).
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 291

PCR-based methods of Candida identification
The molecular-based technology that has undoubtedly had
the greatest impact in the clinical diagnosis of Candida
infections is PCR. This technique can detect highly limited
quantities of microbial nucleic acid from blood, tissue
specimens as well as cultured microorganisms. Over the last
decade, numerous studies have been performed to investi-
gate the effectiveness of PCR in diagnosis of systemic
infection caused by Candida (Williams et al., 1995, 2001;
Chen et al., 2000; Carvalho et al., 2007; Orazio et al., 2009).
In PCR, a pair of synthetic oligonucleotides homologous
to specific sequences serves to prime the amplification of
target DNA. The most important feature of any PCR
primers used directly on clinical samples is that they are
specific and do not amplify host DNA or that of other
microorganisms.
To improve the sensitivity of PCR, many investigators
have designed primers that amplify regions of DNA that are
repeated in the fungal genome. The most commonly used
target for yeast diagnostic PCR primers is the rRNA gene
operon, encoding the 18S, 5.8S, and 28S rRNA gene
subunits, namely internal transcribed spacer 1 (ITS1), ITS2
and ITS4 (Fell et al., 1992; Sullivan et al., 1995; Williams
et al., 1995, 2001; Haynes & Westerneng, 1996; Chen et al.,
2000). More recently, multiplex targets, coupled to real-time
PCR, have been used successfully (Sampaio et al., 2005;
Carvalho et al., 2007; Orazio et al., 2009) for Candida species
identification.
Despite the increased development of new molecular
approaches, the great majority of clinical diagnosis of
candidosis are based on nonmolecular methodologies due
the reduced amount of PCR equipment in hospital labora-
tories, the problems with sample preparation and environ-
mental contamination and the lack of standardized
protocols for PCR methodologies.
Epidemiology and risk factors in NCACspecies infection
The mortality rates associated with different microorgan-
isms have declined with the early administration of empiri-
cal antibiotics and antifungal agents. However, despite this,
systemic fungal infections are increasingly recognized as
important causes of morbidity and mortality. Candida
species are among the most frequently recovered fungi from
blood cultures of hospitalized patients (Pfaller et al., 1998,
2010). In fact, an increasing incidence of fungal infections
with Candida species has been noted in immunocompro-
mised patients, including those in intensive care, postsurgi-
cal units and suffering from cancer (Kiehn et al., 1980;
Samaranayake et al., 2002; Hagerty et al., 2003). Candida
species are most frequently isolated from the oral cavity, and
vulvovaginal and urinary tracts and are detected in approxi-
mately 31–55% of healthy individuals. Historically, C.
albicans has accounted for 70–80% of clinical isolates, with
other NCAC species only rarely encountered (Odds, 1988;
Calderone, 2002; Samaranayake et al., 2002). Nevertheless,
over the last 10–30 years NCAC species have emerged as
important opportunistic pathogens of humans and the
reasons for this might be related to improved diagnostic
methods or altered medical practices, as mentioned above.
Regardless of the basis of this change, recent epidemiological
data reveal a mycological shift, and while C. albicans remains
the most common causative agent, its relative incidence in
infection is declining with the increasing prevalence of other
species such as C. glabrata, C. tropicalis and C. parapsilosis
(Chandra et al., 2001; Colombo et al., 2003; Bassetti et al.,
2006).
In a study on the epidemiology of invasive candidosis,
Pfaller & Diekema (2007) observed that C. albicans, C.
glabrata, C. tropicalis and C. parapsilosis collectively ac-
counted for about 95% of identifiable Candida infections.
Moreover, in the 1980s, according to Kiehn et al. (1980), C.
albicans constituted 68% of Candida isolates from sites
other than blood in cancer patients, while C. tropicalis, C.
parapsilosis and C. glabrata accounted only for 12%, 10%
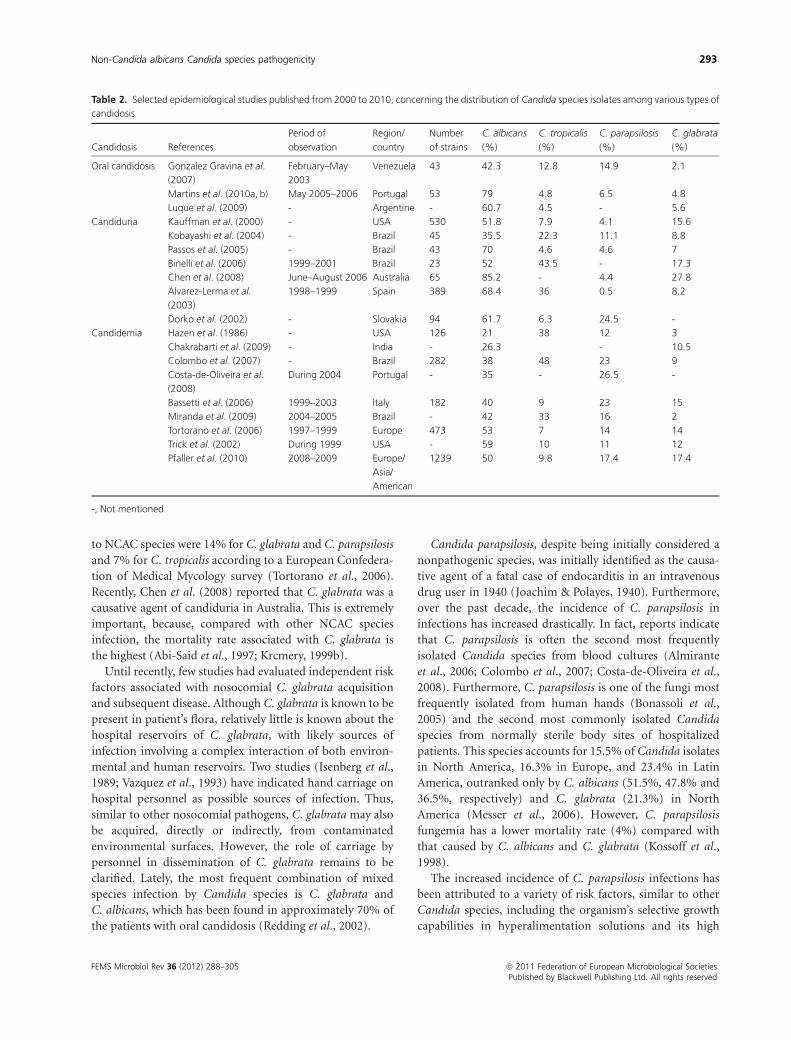
and 3.0% of the isolates, respectively. Table 2 presents
epidemiologic studies published between 2000 and 2010,
concerning oral candidosis, candiduria and candidemia. In
more recent studies, most cases of fungemia have been
significantly associated with NCAC species (Bassetti et al.,
2006; Colombo et al., 2007; Chakrabarti et al., 2009; Pfaller
et al., 2010). However, it is important to emphasize that
there are significant variations in Candida species isolation
depending on the geographical region and patient group,
with some NCAC species being more prevalent, even
compared with C. albicans, in certain countries (Colombo
et al., 2007).
The incidence of C. glabrata is higher in adults than in
children, and lower in neonates (Krcmery & Barnes, 2002).
In contrast, C. parapsilosis appears to be a significant
problem in neonates, transplant recipients and patients
receiving parenteral nutrition (Trofa et al., 2008). Further-
more, C. tropicalis is commonly associated with patients
with neutropenia and malignancy (Colombo et al., 2007).
For many years C. glabrata was considered a relatively
nonpathogenic saprophyte of the normal flora of healthy
individuals and certainly not readily associated with serious
infection in humans. However, following the widespread
and the increased use of immunosuppressive therapies
together with broad-spectrum antibiotic treatment, the
frequency of mucosal and systemic infections caused by
C. glabrata has increased significantly (Hajjeh et al., 2004).
Although the mortality rate associated with Candida infec-
tions varies with the type of patient and with the causative
agent, the incidence rates of candidosis infections attributed
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
292 S. Silva et al.
to NCAC species were 14% for C. glabrata and C. parapsilosis
and 7% for C. tropicalis according to a European Confedera-
tion of Medical Mycology survey (Tortorano et al., 2006).
Recently, Chen et al. (2008) reported that C. glabrata was a
causative agent of candiduria in Australia. This is extremely
important, because, compared with other NCAC species
infection, the mortality rate associated with C. glabrata is
the highest (Abi-Said et al., 1997; Krcmery, 1999b).
Until recently, few studies had evaluated independent risk
factors associated with nosocomial C. glabrata acquisition
and subsequent disease. Although C. glabrata is known to be
present in patient’s flora, relatively little is known about the
hospital reservoirs of C. glabrata, with likely sources of
infection involving a complex interaction of both environ-
mental and human reservoirs. Two studies (Isenberg et al.,
1989; Vazquez et al., 1993) have indicated hand carriage on
hospital personnel as possible sources of infection. Thus,
similar to other nosocomial pathogens, C. glabrata may also
be acquired, directly or indirectly, from contaminated
environmental surfaces. However, the role of carriage by
personnel in dissemination of C. glabrata remains to be
clarified. Lately, the most frequent combination of mixed
species infection by Candida species is C. glabrata and
C. albicans, which has been found in approximately 70% of
the patients with oral candidosis (Redding et al., 2002).
Candida parapsilosis, despite being initially considered a
nonpathogenic species, was initially identified as the causa-
tive agent of a fatal case of endocarditis in an intravenous
drug user in 1940 (Joachim & Polayes, 1940). Furthermore,
over the past decade, the incidence of C. parapsilosis in
infections has increased drastically. In fact, reports indicate
that C. parapsilosis is often the second most frequently
isolated Candida species from blood cultures (Almirante
et al., 2006; Colombo et al., 2007; Costa-de-Oliveira et al.,
2008). Furthermore, C. parapsilosis is one of the fungi most
frequently isolated from human hands (Bonassoli et al.,
2005) and the second most commonly isolated Candida
species from normally sterile body sites of hospitalized
patients. This species accounts for 15.5% of Candida isolates
in North America, 16.3% in Europe, and 23.4% in Latin
America, outranked only by C. albicans (51.5%, 47.8% and
36.5%, respectively) and C. glabrata (21.3%) in North
America (Messer et al., 2006). However, C. parapsilosis
fungemia has a lower mortality rate (4%) compared with
that caused by C. albicans and C. glabrata (Kossoff et al.,
1998).
The increased incidence of C. parapsilosis infections has
been attributed to a variety of risk factors, similar to other
Candida species, including the organism’s selective growth
capabilities in hyperalimentation solutions and its high
Table 2. Selected epidemiological studies published from 2000 to 2010, concerning the distribution of Candida species isolates among various types of
candidosis
Candidosis References
Period of
observation
Region/
country
Number
of strains
C. albicans
(%)
C. tropicalis
(%)
C. parapsilosis
(%)
C. glabrata
(%)
Oral candidosis Gonzalez Gravina et al.
(2007)
February–May
2003
Venezuela 43 42.3 12.8 14.9 2.1
Martins et al. (2010a, b) May 2005–2006 Portugal 53 79 4.8 6.5 4.8
Luque et al. (2009) - Argentine - 60.7 4.5 - 5.6
Candiduria Kauffman et al. (2000) - USA 530 51.8 7.9 4.1 15.6
Kobayashi et al. (2004) - Brazil 45 35.5 22.3 11.1 8.8
Passos et al. (2005) - Brazil 43 70 4.6 4.6 7
Binelli et al. (2006) 1999–2001 Brazil 23 52 43.5 - 17.3
Chen et al. (2008) June–August 2006 Australia 65 85.2 - 4.4 27.8
Alvarez-Lerma et al.
(2003)
1998–1999 Spain 389 68.4 36 0.5 8.2
Dorko et al. (2002) - Slovakia 94 61.7 6.3 24.5 -
Candidemia Hazen et al. (1986) - USA 126 21 38 12 3
Chakrabarti et al. (2009) - India - 26.3 - 10.5
Colombo et al. (2007) - Brazil 282 38 48 23 9
Costa-de-Oliveira et al.
(2008)
During 2004 Portugal - 35 - 26.5 -
Bassetti et al. (2006) 1999–2003 Italy 182 40 9 23 15
Miranda et al. (2009) 2004–2005 Brazil - 42 33 16 2
Tortorano et al. (2006) 1997–1999 Europe 473 53 7 14 14
Trick et al. (2002) During 1999 USA - 59 10 11 12
Pfaller et al. (2010) 2008–2009 Europe/
Asia/
American
1239 50 9.8 17.4 17.4
-, Not mentioned.
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 293

ability to colonize intravascular devices and prosthetic
materials. Additionally, patients requiring prolonged used
of a central venous catheter or indwelling devices, such as
cancer patients, are at increased risk of infection with
C. parapsilosis. A recent Spanish study of 72 patients with
invasive C. parapsilosis identified vascular catheterization
(97%), prior antibiotic therapy (91%), parenteral nutrition
(54%), prior surgery (46%), prior immunosuppressive
therapy (38%), malignancy (27%), transplant receipt
(16%), neutropenia (12%) and prior colonization (11%),
as risk factors for infection (Almirante et al., 2006). In a
report of 64 episodes (between 2002 and 2003) of
C. parapsilosis candidemia in Brazilian hospitals, the pri-
mary risk factors were neutropenia, the use of central venous
catheters and cancer chemotherapy (Brito et al., 2006). The
population at greatest risk for nosocomial infection with
C. parapsilosis is that of extremely low-birth-weight neonates
(Solomon et al., 1984; Voss et al., 1994). In fact, colonization
of the skin or gastrointestinal tract is frequently the first step in
the pathogenesis of invasive candidosis, and neonates are
especially prone to such infections given their compromised
skin integrity, susceptibility to gastrointestinal tract infection,
long-term need for central venous or umbilical catheters and
prolonged endotracheal intubation (Benjamin et al., 2000).
Furthermore, C. parapsilosis has been isolated from approxi-
mately one-third of neonates with gastrointestinal coloniza-
tion by Candida species (Saiman et al., 2001) and from
oropharynges of 23% of healthy neonates (Contreras et al.,
1994). Furthermore, in contrast to other NCAC species, the
rates of mortality in low-birth-weight neonates caused by
C. parapsilosis are drastically higher and sometimes equivalent
to those associated with C. albicans (Trofa et al., 2008).
Candida tropicalis is one of the three most commonly
isolated NCAC species (Alvarez-Lerma et al., 2003; Binelli
et al., 2006; Colombo et al., 2007; Hasan et al., 2009).
Usually, C. tropicalis is considered the third most frequently
isolated NCAC species from blood and urine cultures (Table
2) (Kauffman et al., 2000; Alvarez-Lerma et al., 2003).
Moreover, in a recent epidemiology study conducted in 12
Brazilian medical centres, C. tropicalis was the second most
frequently recovered Candida species, accounting for
33–48% of all candidemia cases (Colombo et al., 2007;
Miranda et al., 2009). Additionally, C. tropicalis is often
found in patients admitted to intensive care units, especially
in patients requiring prolonged catheterization, receiving
broad-spectrum antibiotics or with cancer (Kauffman et al.,
2000; Rho et al., 2004; Colombo et al., 2007; Nucci &
Colombo, 2007). Furthermore, C. tropicalis appears to dis-
play a higher potential for dissemination in neutropenic
individuals compared with C. albicans and other NCAC
species (Colombo et al., 2007).
According to Kontoyiannis et al. (2001), there are distinct
differences in the presentation and risk factors of C. tropica-
lis and C. albicans fungemia, with the former more persis-
tent and leading to longer intensive care unit stays during
the course of infection. This may imply a higher virulence
and greater resistance to commonly used antifungals by
C. tropicalis when compared with C. albicans. In fact, some
epidemiologic studies (Krcmery, 1999a; Kontoyiannis et al.,
2001; Eggimann et al., 2003; Colombo et al., 2007)
documented that C. tropicalis was associated with higher
mortality than other NCAC species and C. albicans.
This propensity of C. tropicalis for dissemination and the
associated high mortality may be related to the virulence
factors exhibited by this species such as biofilm formation,
proteinases secretion and dimorphism (Krcmery, 1999b;
Negri et al., 2010a).
Pathogenicity and virulence factors ofNCAC species
There remains a debate over what actually constitutes a
virulence factor. It can be argued that all the traits required
for establishing disease are virulence factors; however,
strictly speaking, virulence factors are those that interact
directly with host cells causing damage (Haynes, 2001). The
pathogenicity of Candida species is mediated by a number of
virulence factors, including adherence and biofilm forma-
tion on host tissue as well as medical devices, the ability to
evade host defences and the production of tissue-damaging
hydrolytic enzymes (e.g. proteases, phospholipases and
haemolysins).
Infection models of candidosis in animals suggest that
C. albicans is the most pathogenic species (Samaranayake &
Samaranayake, 2001), and in vitro investigations indicate
that it also expresses higher levels of putative virulence
factors compared with other species (Jayatilake et al.,
2006). Furthermore, it is important to emphasize that these
yeasts are not usual pathogens of these animals and therefore
such studies do not necessarily reflect the reality of patho-
genicity of Candida species. Moreover, Candida species can
colonize and cause disease at several anatomically distinct
sites including the skin, oral cavity, gastrointestinal tract,
vagina and vascular system. In order to establish infection,
opportunistic pathogens have to evade the immune system,
survive, reproduce in the host environment, and in the case
of systemic infection, disseminate to new tissues and organs.
Adhesion and biofilm formation
The primary event in Candida infection is adherence to host
surfaces, which is required for initial colonization. Adher-
ence contributes to persistence of the organism within the
host, and is considered essential in the establishment of
disease. Furthermore, Candida species can also adhere to the
surfaces of medical devices and form biofilms. Several
factors have been implicated in influencing adhesion,
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
294 S. Silva et al.

including the profile of cell wall proteins (Chaffin, 2008) and
cell surface physicochemical properties (Anil et al., 2001;
Henriques et al., 2002).
Candida cell surface proteins that are involved in specific
adherence are described as adhesins. In C. glabrata, a major
group of adhesins is encoded for by the EPA (epithelial
adhesin) gene family (De las Penas et al., 2003). The overall
structure of Epa proteins is similar to that of the ALS
(agglutinin-like sequence) proteins of C. albicans. Although
there are few studies concerning C. glabrata Epa proteins, it
is known that EPA1p is a calcium-dependent lectin that
binds to N-acetyl lactosamine-containing glycol conjugates
(Cormack et al., 1999). Furthermore, despite the large
number of EPA genes, it has been shown that deletion of
merely Epa1p reduces adherence in vitro (De las Penas et al.,
2003). In addition, although EPA6 is not expressed in vitro,
its expression increases during in vivo urinary tract infec-
tion, suggesting that C. glabrata is capable of adapting to
different environmental conditions (Domergue et al., 2005).
Furthermore, a bioinformatic search of pathogen-specific
gene families of Candida species revealed a number of genes
for putative cell wall adhesins-like-proteins in C. parapsilo-
sis. This study included genes for five Als proteins and six for
Pga 30 (predicted glycosylphosphatidylinositol-anchored
protein 30) (Butler et al., 2009). Unfortunately, there has
been no further work in studying the role that these
proteins play in C. parapsilosis adhesion. Concerning,
proteins from the C. tropicalis cell wall, at least three Alsp
have been identified through Western blot analysis with
anti-Als antibodies (Hoyer et al., 2001); however, to the
authors’ knowledge, no further work has been undertaken in
this area.
The fungal cell surface is the site of physical–chemical
interactions with host tissues or medical devices leading to
its adherence (Cannon & Chaffin, 1999). Previous studies of
the cell wall of Candida have suggested a relationship
between cell surface hydrophobicity and adherence (Pana-
goda et al., 2001). In a study with a limited number of C.
glabrata isolates, this species was found to exhibit a degree of
hydrophobicity similar to C. albicans (Hazen et al., 1986).
Interestingly, while the hydrophobicity of C. albicans was
extremely sensitive to specific growth conditions, numerous
isolates of C. glabrata were relatively insensitive to the same
growth conditions (Kikutani & Makino, 1992). In addition,
Camacho et al. (2007) did not find a correlation between the
hydrophobicity and adherence for Candida cells on silico-
nized latex catheters, demonstrating that cell hydrophobi-
city alone was not a predictor for adhesion levels. As
reported for C. glabrata, Panagoda et al. (2001) showed that
the initial adhesion of C. parapsilosis and C. tropicalis cells
was associated with surface hydrophobicity.
Initial attachment of Candida to host or/and medical
devices is followed by cell division, proliferation and sub-
sequent biofilm development (Ramage et al., 2006). Biofilms
are described as surface-associated communities of micro-
organisms embedded within an extracellular matrix. It is
now considered that biofilms represent the most prevalent
growth form of microorganisms (Al-Fattani & Douglas,
2006; Silva et al., 2009a). Biofilm formation is an important
virulence factor for a number of Candida species, as it
confers significant resistance to antifungal therapy by limit-
ing the penetration of substances through the matrix and
protecting cells from host immune responses (Donlan &
Costerton, 2002; Mukherjee & Chandra, 2004). Moreover,
biofilms formed by C. albicans, C. parapsilosis, C. tropicalis
and C. glabrata isolates have been associated with higher
morbidity and mortality rates compared with isolates un-
able to form biofilms (Kumamoto, 2002). It is assumed that
the formation of mature biofilms and subsequent produc-
tion of extracellular matrix is strongly dependent on species,
strain and environmental conditions (pH, medium compo-
sition, oxygen) (Ramage et al., 2006; Jain et al., 2007).
Recently, Silva et al. (2010b) showed that C. glabrata
produced a higher biofilm biomass on silicone surfaces in
the presence of urine, compared with C. parapsilosis and C.
tropicalis. The opposite was found for biofilms formed in
Sabouraud dextrose broth (Silva et al., 2009a). These results
are in accordance with Shin et al. (2002) who reported that
biofilm formation by C. glabrata was lower compared with
other NCAC species, when grown in rich culture media.
Candida tropicalis clinical isolates have been classified as
being extensive biofilm formers on silicone and latex
catheter (Fig. 3) (Redding et al., 2002; Silva et al., 2009a;
Negri et al., 2010b). Biofilms are readily formed by C.
parapsilosis cells grown in media containing high glucose
and lipid concentrations, and can be associated with the
increased prevalence of this organism in bloodstream infec-
tions of patients receiving parenteral nutrition (Nosek et al.,
2009). The selective preference of this species for plastic
medical devices is of particular interest, as biofilm formation
enhances the capacity of the organism to colonize catheters
and intravascular lines (Weems, 1992; Trofa et al., 2008). In
contrast to C. albicans, C. parapsilosis biofilms are thinner,
less structured and consist exclusively of aggregated blastos-
pores (Kuhn et al., 2002). These biofilm features are in
accordance with those recently reported by Silva et al.
(2009a). Lattif et al. (2009) demonstrated that, like C.
parapsilosis, the two newly identified Candida species (C.
orthopsilosis and C. metapsilosis) were also able to form
biofilms.
Little is known about the matrix composition of NCAC
species biofilms, but according to Baillie & Douglas (2000),
C. albicans biofilm matrix is mainly composed of carbohy-
drates, proteins, phosphorus and hexosamines. Silva et al.
(2009a) reported that the extracellular matrices of C. para-
psilosis biofilms contained large amounts of carbohydrates
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 295

with correspondingly low levels of proteins. In the same
study, C. glabrata biofilm matrices were found to have high
levels of both proteins and carbohydrates, while C. tropicalis
biofilm matrices had low levels of carbohydrates and pro-
teins compared with the other NCAC species. Interestingly,
the biofilm matrix composition is highly strain dependent, a
phenomenon that has not been observed in related yeasts
(Silva et al., 2009a). Furthermore, Al-Fattani & Douglas
(2006) showed that matrix material extracted from biofilms
of C. tropicalis and C. albicans contained carbohydrates,
proteins, hexosamine, phosphorus and uronic acid. How-
ever, the major component in C. tropicalis biofilm matrices
was hexosamine (27%). The same authors also reported that
these biofilms partially detached after treatment with lipase
type VII and chitinase, which is in contrast to biofilms
of C. albicans that detached only after treatment with
proteinase K, chitinase, DNase I or b-N-aceytyglucosami-
dase. DNA has been described as a component of the
extracellular matrix in bacteria biofilms (Allesen-Holm
et al., 2006; Vilain et al., 2009). In Candida species, there is
scarce knowledge concerning the contribution of extracel-
lular DNA to biofilm matrix and overall structure. Recently,
Martins et al. (2010a, b) highlighted the importance of DNA
in C. albicans biofilm formation, integrity and structure.
However, there is a lack of knowledge concerning NCAC
species and the role of extracellular DNA on biofilm
composition.
While extensive work has been performed on the
C. albicans genes involved in adhesion/colonization and
biofilm formation, little is known about equivalent control-
ling genes in C. glabrata, C. parapsilosis and C. tropicalis.
However, two recent studies involving the study of
C. parapsilosis lipase knockout mutants found that these
had a decreased ability to form biofilms. The C. parapsilosis
mutants produced significantly less biofilm than the wild-
type strain (Gacser et al., 2007a). Moreover, the BCR
(biofilm and cell wall regulator) gene was also deemed
necessary for proper biofilm formation (Gacser et al.,
2007b). Notably, the biofilm-deficient C. parapsilosis lipase
mutants were less virulent in tissue culture infection models
and in mice (Gacser et al., 2007b). As documented above,
the cell wall seems to play a crucial role in colonization and
infection, and therefore, elucidation of its structure and
composition may lead to a better understanding of NCAC
pathogenicity, and also aid in identifying therapeutic targets.
Hydrolytic enzymes
Destruction of host tissues by Candida species may be
facilitated by the release of hydrolytic enzymes into the local
environment. Secreted aspartyl proteinases (Saps), phospho-
lipases, lipases (LIPs) and haemolysins are the enzymes most
frequently implicated in Candida species pathogenicity.
Saps facilitate invasion and colonization of host tissues by
disruption of the host mucosal membranes (Ruchel, 1999)
and by degrading important immunological and structural
defence proteins (Pichova et al., 2001). In the case of
C. glabrata, only one study has shown that this species is
capable of proteinase production, but the type of proteinase
was not specified (Chakrabarti et al., 1991). For C. para-
psilosis, three SAP genes have been identified (SAPP1-3), two
of which remain largely uncharacterized (Merkerova et al.,
2006). The Sapp1p isoenzyme has, however, been biochemi-
cally characterized (Fusek et al., 1994; Pichova et al., 2001;
Dostal et al., 2005), and SAPP2 encodes a functional
proteinase that constitutes only 20% of the Saps isolated
from a culture supernatant (Fusek et al., 1994). It has been
reported (Silva et al., 2009b) that SAPP1-3 genes expression
varies with different clinical isolates of C. parapsilosis when
grown in contact with an oral epithelium and even in
planktonic growth forms. However, there is a trend relating
Sap production and site of strain isolation, because both
vaginal and skin isolates of C. parapsilosis exhibit higher
in vitro Sap activity than blood isolates (Cassone et al., 1995;
Dagdeviren et al., 2005). Candida parapsilosis has been
shown to be poorly invasive of an oral epithelium, but can
nevertheless induce significant damage, which was related to
specific SAP gene expression (Silva et al., 2009b). As with C.
albicans, in vitro studies reveal that C. tropicalis secretes high
levels of Saps in a medium containing bovine serum
albumin as the sole source of nitrogen (Zaugg et al., 2001;
Negri et al., 2010b). Furthermore, C. tropicalis possesses at
least four genes encoding Saps, designated as SAPT1 to
SAPT4 (Togni et al., 1991; Zaugg et al., 2001). To date,
Sapt1p is the only enzyme that has been purified from
culture supernatant, biochemically characterized and
(a) (b)
Fig. 3. Scanning electron microscopy images of
Candida tropicalis biofilms formed on (a) silicone
and (b) latex catheter in artificial urine at 24 h.
Scale bar = 20 mm.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
296 S. Silva et al.

crystallized (Togni et al., 1991; Symersky et al., 1997). The
presence of Saps secreted by C. tropicalis has also been
reported on the surface of fungal elements penetrating
tissues during disseminated infection and evading macro-
phages after phagocytosis of yeast cells (Borg & Ruchel,
1990; Ruchel et al., 1992). Recently, Silva et al. (2010b)
demonstrated that, like C. albicans (Lermann & Morsch-
hauser, 2008; Naglik et al., 2008), Sap expression during
C. tropicalis colonization of an oral epithelium was not
associated with invasion and tissue damage.
In addition to Saps, enzymes categorized as phospholi-
pases are often considered to be involved in Candida
pathogenicity. Phospholipases are enzymes that hydrolyze
phospholipids into fatty acids. The production of all classes
of phospholipases have been described for Candida species
and are suggested to contribute to host cell membrane
damage, which could also expose receptors to facilitate
adherence (Ghannoum, 2000; Kantarciolu & Yucel, 2002).
The most widely used diagnostic method for phospholipase
determination is based on yeast growth in an egg yolk agar
media. Several studies indicate that NCAC species produce
extracellular phospholipases (Furlaneto-Maia et al., 2007;
Cafarchia et al., 2008; Galan-Ladero et al., 2010), but at
significantly lower levels compared with C. albicans (Ghan-
noum, 2000). There have been contradictory findings, with
some investigators reporting phospholipase activity in 51%
of the strains assayed (Ghannoum, 2000) and others finding
no phospholipase activity in the examined strains (Kantar-
ciolu & Yucel, 2002). According to recent studies, while
C. tropicalis appears to have a reduced ability to produce
extracellular phospholipases, production is highly strain
dependent (Furlaneto-Maia et al., 2007; Cafarchia et al.,
2008; Galan-Ladero et al., 2010; Negri et al., 2010b). Further-
more, contrary to the few studies on C. tropicalis and
C. parapsilosis (Kumar et al., 2009), no studies have been
reported concerning C. glabrata phospholipase production.
Lipases are involved in the hydrolysis of triacylglycerols.
In C. albicans, 10 genes encoding for lipases have been
identified and it has been shown that C. albicans CaLIP8
mutants were significantly less virulent in a murine intrave-
nous infection model (Gacser et al., 2007b). For C. para-
psilosis, CpLIP1 and CpLIP2 have been reported, with the
latter known to encode for an active protein (Neugnot et al.,
2002). Recently, Gacser et al. (2007a) demonstrated that a
lipase inhibitor significantly reduced tissue damage during C.
parapsilosis infection of a reconstituted human tissue, and
that CpLIP1/CpLIP2 mutants formed thinner and less com-
plex biofilms. Sequences similar to C. albicans (LIP1-10) were
also detected in C. tropicalis, but not in C. glabrata. However,
no studies have been performed to investigate the role of these
genes in the virulence of C. tropicalis (Fu et al., 1997).
Pathogenic microorganisms can grow in the host using
haemin or haemoglobin as a source of iron. Haemolysins are
used by Candida species to degrade haemoglobin and
facilitate recovery of the elemental iron from host cells.
Thus, haemolysins are considered key virulence factors
enabling pathogen survival and persistence in the host
(Manns et al., 1994; Watanabe et al., 1999; Luo et al., 2004).
Furthermore, it is known that C. albicans has the ability to
utilize iron to produce a factor that can release haemoglobin
by lysing erythrocytes (Manns et al., 1994; Watanabe et al.,
1999). Production of this haemolytic factor may be regu-
lated by the presence of glucose in the growth medium.
Candida glabrata, C. parapsilosis and C. tropicalis are all able
to produce haemolysins in vitro, inducing partial or total
erythrocyte lyses, although the extent of this is both strain
and species dependent (Luo et al., 2004). Other authors
(Furlaneto-Maia et al., 2007; Kumar et al., 2009; Negri et al.,
2010b) only observed production of haemolysins by
C. albicans. Although haemolysins are known to be putative
virulence factors contributing to pathogenicity in Candida
species, the genetic expression of haemolytic activity of
Candida is poorly understood at present, but a study
conducted by Luo et al. (2004) showed that a haemolysin-
like protein (HLP) gene was associated with the haemolytic
activity of C. glabrata.
Filamentous growth
Hyphae are believed to play an important role in tissue
invasion, and in vitro research has shown that C. albicans
lacking hyphal formation exhibited lower ability to invade
tissue compared with wild-type C. albicans strains (Jayati-
lake et al., 2006). Furthermore, filamentous forms (hyphae
and/or pseudohyphae) of Candida species also demonstrate
increased resistance to phagocytosis compared with yeast
(Gow et al., 2002). The morphological forms exhibited by
C. tropicalis are similar to those shown by C. albicans, but
despite this few studies have explored the importance of
C. tropicalis morphology on virulence. However, Silva et al.
(2010a) demonstrated recently that only filamentous forms
of C. tropicalis were able to invade an oral epithelium. In the
case of C. parapsilosis, it has been found that hyphal
transition occurs in a strain-dependent manner (Enger
et al., 2001), and contrary to C. albicans and C. tropicalis,
the ability of C. parapsilosis to invade an oral epithelium did
not correlate with pseudohyphal production (Silva et al.,
2009b).
Antifungal therapies and mechanisms ofresistance of NCAC species
Compared with antibiotics, the development of antifungal
agents has been relatively limited. This can be attributed to
several factors including inherent problems in the identifi-
cation of an effective agent that acts on eukaryotic fungal
cell type without being toxic to host cells. Resistance to
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 297

antifungal drugs is an increasingly recognized phenomenon
and can be defined clinically as the persistence of signs and
symptoms of the infection despite the presence of a tolerable
level of the drug. Depending on the drug and the Candida
species, the mechanism of antifungal resistance can either be
inherent (present without previous exposure to the anti-
fungal) or acquired, where resistance develops in a pre-
viously susceptible organism after a period of exposure to
the agent.
The classification of these drugs is currently based on
their target of activity (Table 3). Polyene antifungals, such as
amphotericin B, are fungicidal due to their ability to interact
with the ergosterol component within the cell membrane to
generate pores, causing cell membrane leakage leading to
loss of cytoplasmic content. Azoles are another class of
antifungal agents that inhibit the biosynthesis of ergosterol
by interfering with the fungal enzyme, lanosterol demethy-
lase. A key function of this enzyme is to convert lanosterol to
ergosterol and the inhibition of this leads to depletion of the
sterol in the fungal cell membrane. Azole antifungals have a
fungistatic or fungicidal activity against Candida species and
the most frequently used azole antifungals are fluconazole
and itraconazole.
5-Flucytosine is an antifungal that enters the fungal cell
through a cytosine permease and is then converted by the
fungus into 5-fluorouracil. This nucleoside analogue gets
incorporated into RNA molecules and subsequently inter-
feres with the synthesis of proteins within the fungal cell.
Several antifungals have been developed that target cell
wall components, for example b-1,3-D-glucan, a key com-
ponent of the fungal cell wall is not present within mamma-
lian cells. Interference with the enzyme b-1,3-D-glucan
synthetase can inhibit the synthesis of b-1,3-D-glucan. A
group of antifungals demonstrating this mode of action are
the echinocandins, and three drugs (caspofungin, micofun-
gin and anidulafungin) have shown antifungal activity
against Candida species.
Amphotericin B is generally regarded to have the broadest
spectrum of antifungal activity and is used in cases of serious
and invasive Candida infections, such as in the treatment of
systemic infection in hospitalized patients. Resistance to
amphotericin B remains uncommon during treatment, but
reports of isolates exhibiting elevated minimum inhibitory
concentration (MIC) have become more frequent (Pfaller &
Diekema, 2007). Resistance to polyenes is believed to result
from the alteration of sterol content or composition in the
cell membrane (Ghannoum & Rice, 1999). Lupetti et al.
(2002) described that among Candida species, polyene
resistance was usually due to defective ergosterol biosynth-
esis, and most likely resulted from mutation in the ERG3
gene that produces altered d5,6-sterol desaturase activity. In
addition to ERG3 gene, mutation in ERG11 (the gene that
produces lanosterol 14a-demethylase) and in ERG6 (a gene
that is required for normal membrane function, but is not
essential for sterol biosynthesis) may generate polyene
resistance. Importantly, C. glabrata isolates have been iden-
tified with mutations in the ERG6 gene (Vandeputte et al.,
2007).
The development of the azole antifungals enhanced the
treatment options for fungal infections and their reduced
host toxicity has led to their widespread use. Consequently,
with this extensive use, it is perhaps not surprising that
resistance to these agents, particularly fluconazole, has been
encountered (Rex et al., 1995; Pfaller & Diekema, 2007).
Resistance to the azoles can result from quantitative or
qualitative modifications of target enzymes, reduced access
of the drug to the target or a combination of these mechan-
isms. Qualitative modifications in target enzymes result
from point mutations in ERG11, the gene responsible for
production of 14a-demethylase, which is the principal
target of the azoles. The other primary mechanism by which
Candida species resist the effects of azole antifungals in-
volves the active efflux of the drug out of the cell via the
activation of two kinds of efflux transport proteins encoded
by either MDR or CDR genes (Lupetti et al., 2002; Ghan-
noum & Rice, 1999; Sanglard & Odds, 2002). Candida
glabrata may be intermediately resistant to all azoles and
about 20% of strains develop resistance during therapy and
prophylaxis with fluconazole (Pfaller & Diekema, 2007).
Susceptibility testing has shown that fluconazole is active
against several Candida species, including C. albicans, C.
parapsilosis and C. tropicalis (Pfaller & Diekema, 2007).
Itraconazole is moderately active against most medically
important fluconazole-susceptible and -resistant Candida
species, with the exception of C. glabrata (Pfaller et al.,
2005). Voriconazole exerts fungicidal activity against most
Table 3. Common antifungal agents used in the management of
candidosis, mode of action and susceptibilities of Candida species
Antifungal
Susceptibility of NCAC species
C. albicans C. parapsilosis C. tropicalis C. glabrata
Polyenes Disruption of fungal cell membrane
Amphotericin B S S S S to I
Azoles Inhibition of ergosterol synthesis
Fluconazole S S S SDD to R
Itraconazole S S S SDD to R
Voriconazole S S S S
Posoconazole S S S S
Ravuconazole S S S S
5-Flucytosine Inhibition of DNA and protein synthesis
S S S S
Echinocandins Inhibition of b1,3-D-glucan synthesis
Caspofungin S S S S
Adapted from references Rex et al., 1995, 1997, 2000; Diekema et al.,
2002; Roling et al., 2002; Eggimann et al., 2003; S-DD, susceptible-dose
dependent; I, intermediate; R, resistant.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
298 S. Silva et al.

yeasts and certain opportunistic fungi, specifically against
some NCAC species (Groll et al., 2001). This agent is
generally active against Candida species including flucona-
zole-resistant C. albicans and C. glabrata (Pfaller & Dieke-
ma, 2007). With the exception of C. tropicalis, voriconazole
is more active than fluconazole against medically important
Candida species (Pfaller & Diekema, 2007). Posaconazole
exerts fungistatic activity against some NCAC species,
including C. glabrata, C. tropicalis and C. parapsilosis (Scozo
et al., 2007).
Flucytosine has a narrow spectrum of activity, and several
mechanisms of resistance are possible due to the multiple
intracellular enzymatic steps required for its action. These
include alterations in the target enzymes UMP pyropho-
sphorylase, cytosine permease and cytosine deaminase, or
increased production of pyrimidines (Atkinson & Israel,
1973). The antifungal spectrum of flucytosine is extremely
narrow: Candida species, Cryptococcus species and Aspergil-
lus species (Polak et al., 1982; Vermes et al., 2000). Further-
more, due to the multiple steps in its mode of action,
including transport into the cell and deamination of the
active compound, flucytosine is normally used only in
combination with other agents, including amphotericin B
and fluconazole (Vermes et al., 2000).
As a class, the echinocandins are the most recent addition
to the antifungal arsenal, and to date their use has been very
limited to assess whether significant resistance will develop
to these agents. Microorganisms that demonstrate inherent
resistance to echinocandins either generate insufficient
target enzyme b-1,3-D-glucan synthase or produce an alter-
nate form of the enzyme with reduced echinocandin bind-
ing. All echinocandins exert fungicidal activity against
Candida species. The echinocandins are highly active against
C. albicans, C. glabrata and C. tropicalis both in vitro and in
vivo (Pfaller et al., 2003, 2005; Bayegan et al., 2010;
Kucharınova et al., 2010). It is important is to emphasize
that the MIC values for echinocandins tend to be higher for
C. parapsilosis than for most other common Candida
species, particularly C. albicans (Walsh, 2002).
From the clinical perspective, the most important feature
of Candida biofilms is their role in increasing tolerance to
conventional antifungal therapy. The reduced susceptibility
of C. albicans biofilms to antifungal agents was first reported
in 1995 (Hawser et al., 1995). Furthermore, several groups
have demonstrated that biofilm cells drastically increase
their tolerance to the most commonly used antifungal
agents (fluconazole and amphotericin B) (Ramage et al.,
2001). Biofilms of NCAC species, such as C. tropicalis,
C. parapsilosis and C. glabrata, have also been shown to
exhibit reduced antifungal susceptibility (Hawser &
Douglas, 1994, 1995). Although the mechanisms of biofilm
drug resistance are not fully understood, the current con-
sensus is that biofilm tolerance is a complex multifactorial
phenomenon involving different molecular mechanisms,
restricted penetration of the drug through the matrix and
the presence of so-called ‘persister’ cells within the biofilm,
which survive exposure to the agent (Lewis, 2001; Donlan &
Costerton, 2002; Douglas, 2003).
Concluding remarks
Changes in the host are generally required for opportunistic
yeast to alter from harmless commensal microorganisms to
potentially life-threatening human pathogens. Management
of candidosis involves the identification and control of host
factors that may predispose one to infection. Furthermore,
Candida species can exhibit several virulence factors such as
adherence, biofilm formation and secretion of hydrolytic
enzymes that both increase their persistence within the host
as well as cause host cell damage. Therefore, the increase in
the incidence and antifungal resistance of NCAC species,
specifically C. glabrata, C. parapsilosis and C. tropicalis, and
the unacceptably high morbidity and mortality associated
with these species, make it essential to further enhance our
knowledge on the virulence and resistance mechanisms
associated with these species. Studies in this area will con-
tribute towards the identification of new targets for novel
therapeutics against these recently emerged pathogens.
Acknowledgements
The authors acknowledge FCT, Portugal, for supporting
S.S.’s work through grant SFRH/BD/28341/2006 and
CAPES, Brazil, for supporting M.N.’s work through grant
BEX-4642/06-6. We would like to thank Designer Fabio
Grassi for helping in the improvement of the images.
References
Abi-Said D, Anaissie E, Uzun O, Raad I, Pinzcowski H &
Vartivarian S (1997) The epidemiology of hematogenous
candidiasis caused by different Candida species. Clin Infect Dis
24: 1122–1128.
Alexander BD, Ashley ED, Reller LB & Reed SD (2006) Cost
savings with implementation of PNA FISH testing for
identification of Candida albicans in blood cultures. Diagn
Micr Infec Dis 54: 277–282.
Al-Fattani MA & Douglas LJ (2006) Biofilm matrix of Candida
albicans and Candida tropicalis: chemical composition and
role in drug resistance. J Med Microbiol 55: 999–1008.
Allesen-Holm M, Barken KB, Yang L et al. (2006) A
characterization of DNA release in Pseudomonas aeruginosa
cultures and biofilms. Mol Microbiol 59: 1114–1128.
Almirante B, Rodriguez D, Cuenca-Estrella M et al. (2006)
Epidemiology, risk factors, and prognosis of Candida
parapsilosis bloodstream infections: case-control population-
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 299

based surveillance study of patients in Barcelona, Spain, from
2002 to 2003. J Clin Microbiol 44: 1681–1685.
Alvarez-Lerma F, Nolla-Salas J, Leon C, Palomar M, Jorda R,
Carrasco N & Bobillo F (2003) Candiduria in critically ill
patients admitted to intensive care medical units. Intens Care
Med 29: 1069–1076.
Anil S, Ellepola AN & Samaranayake LP (2001) The impact of
chlorhexidine gluconate on the relative cell surface
hydrophobicity of oral Candida albicans. Oral Dis 7: 119–122.
Atkinson GW & Israel HL (1973) 5-Fluorocytosine treatment of
meningeal and pulmonary aspergillosis. Am J Med 55: 496.
Baillie GS & Douglas LJ (2000) Matrix polymers of Candida
biofilms and their possible role in biofilm resistance to
antifungal agents. J Antimicrob Chemoth 46: 397–403.
Bassetti M, Righi E, Costa A, Fasce R, Molinari M, Rosso R &
Viscoli C (2006) Epidemiological trends in nosocomial
candidemia in intensive care. BMC Infect Dis 6: 21.
Bayegan S, Majoros L, Kardos L, Kemeny-Beke A, Miszti C,
Kovacs R & Gesztelyi R (2010) In vivo studies with a Candida
tropicalis isolate exhibiting paradoxical growth in vitro in the
presence of high concentration of caspofungin. J Microbiol 48:
170–173.
Benjamin DK, Ross K, McKinney RE, Benjamin DK, Auten R &
Fisher RG (2000) When to suspect fungal infection in
neonates: a clinical comparison of Candida albicans and
Candida parapsilosis fungemia with coagulase-negative
staphylococcal bacteremia. Pediatrics 106: 712–718.
Binelli CA, Moretti ML, Assis RS et al. (2006) Investigation of the
possible association between nosocomial candiduria and
candidaemia. Clin Microbiol Infec 12: 538–543.
Bonassoli LA, Bertoli M & Svidzinski TIE (2005) High frequency
of Candida parapsilosis on the hands of healthy hosts. J Hosp
Infect 59: 159–162.
Borg M & Ruchel R (1990) Demonstration of fungal proteinase
during phagocytosis of Candida albicans and Candida
tropicalis. J Med Vet Mycol 28: 3–14.
Brito LR, Guimaraes T, Nucci M, Rosas RC, Almeida PL, Matta
DA & Colombo AL (2006) Clinical and microbiological
aspects of candidemia due to Candida parapsilosis in Brazilian
tertiary care hospitals. Med Mycol 44: 261–266.
Butler G, Rasmussen MD, Lin MF et al. (2009) Evolution of
pathogenicity and sexual reproduction in eight Candida
genomes. Nature 459: 657–662.
Cafarchia C, Romito D, Coccioli C, Camarda A & Otranto D
(2008) Phospholipase activity of yeasts from wild birds and
possible implications for human disease. Med Mycol 46:
429–434.
Calderone RA (2002) Introduction and historical perspectives.
Candida and Candidiasis (Calderone R., ed), pp. 15–25. ASM
Press, Washington, DC.
Camacho D, Gasparetto A & Svidzinski T (2007) The effect of
chlorhexidine and gentian violet on the adherence of Candida
spp. to urinary catheters. Mycopathology 163: 261–266.
Cannon RD & Chaffin WL (1999) Oral colonization by Candida
albicans. Crit Rev Oral Biol M 10: 359–383.
Carvalho A, Costa-De-Oliveira S, Martins ML, Pina-Vaz C,
Rodrigues AG, Ludovico P & Rodrigues F (2007) Multiplex
PCR identification of eight clinically relevant Candida species.
Med Mycol 45: 619–627.
Cassone A, De Bernardis F, Pontieri E, Carruba G, Girmenia C,
Martino P, Fernandez-Rodriguez M, Quindos G & Ponton J
(1995) Biotype diversity of Candida parapsilosis and its
relationship to the clinical source and experimental
pathogenicity. J Infect Dis 171: 967–975.
Chaffin WL (2008) Candida albicans cell wall proteins. Microbiol
Mol Biol R 72: 495–544.
Chakrabarti A, Nayak N & Talwar P (1991) In vitro proteinase
production by Candida species. Mycopathology 114: 163–168.
Chakrabarti A, Chatterjee SS, Rao KLN, Zameer MM,
Shivaprakash MR, Singhi S, Singh R & Varma SC (2009)
Recent experience with fungaemia: change in species
distribution and azole resistance. Scand J Infect Dis 41:
275–284.
Chandra J, Kuhn DM, Mukherjee PK, Hoyer LL, McCormick T &
Ghannoum MA (2001) Biofilm formation by the fungal
pathogen Candida albicans: development, architecture, and
drug resistance. J Bacteriol 183: 5385–5394.
Chen S, Tong Z, Lee O, Halliday C, Playford E, Widmer F, Kong F,
Wu C & Sorrell T (2008) Clinician response to Candida
organisms in the urine of patients attending hospital. Eur J
Clin Microbiol 27: 201–208.
Chen YC, Eisner JD, Kattar MM, Rassoulian-Barrett SL, LaFe K,
Yarfitz SL, Limaye AP & Cookson BT (2000) Identification of
medically important yeasts using PCR-based detection of
DNA sequence polymorphisms in the internal transcribed
spacer 3 region of the rRNA genes. J Clin Microbiol 38:
2302–2310.
Colombo AL, Perfect J, DiNubile M, Bartizal K, Motyl M, Hicks P,
Lupinacci R, Sable C & Kartsonis N (2003) Global distribution
and outcomes for Candida species causing invasive
candidiasis: results from an international randomized double-
blind study of caspofungin versus amphotericin B for the
treatment of invasive candidiasis. Eur J Clin Microbiol 22:
470–474.
Colombo AL, Guimaraes T, Silva LRBF, Monfardini LPDA,
Cunha AKB, Rady P, Alves T & Rosas RC (2007) Prospective
Observational Study of Candidemia in Sao Paulo, Brazil:
Incidence Rate, Epidemiology, and Predictors of Mortality.
Infect Cont Hosp Ep 28: 570–576.
Contreras I, Ponton J & Quindos G (1994) Prevalence of Candida
parapsilosis in the oral cavities of infants in Spain. Clin Infect
Dis 18: 480–481.
Cormack BP, Ghori N & Falkow S (1999) An adhesin of the yeast
pathogen Candida glabrata mediating adherence to human
epithelial cells. Science 285: 578–582.
Costa-de-Oliveira S, Pina-Vaz C, Mendonca D & Rodrigues AG
(2008) A first Portuguese epidemiological survey of fungaemia
in a university hospital. Eur J Clin Microbiol 27: 365–374.
Dagdeviren M, Cerikcioglu N & Karavus M (2005) Acid
proteinase, phospholipase and adherence properties of
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
300 S. Silva et al.

Candida parapsilosis strains isolated from clinical specimens of
hospitalised patients. Mycoses 48: 321–326.
De Las Penas A, Pan SJ, Castano I, Alder J, Cregg R & Cormack BP
(2003) Virulence-related surface glycoproteins in the yeast
pathogen Candida glabrata are encoded in subtelomeric
clusters and subject to RAP1- and SIR-dependent
transcriptional silencing. Genes Dev 17: 2245–2258.
Diekema DJ, Messer SA, Brueggemann AB et al. (2002)
Epidemiology of candidemia: 3-year results from the
Emerging Infections and the Epidemiology of Iowa Organisms
Study. J Clin Microbiol 40: 1298–1302.
Domergue R, Castano I, De Las Penas A, Zupancic M, Lockatell
V, Hebel JR, Johnson D & Cormack BP (2005) Nicotinic acid
limitation regulates silencing of Candida adhesins during UTI.
Science 308: 866–870.
Donlan RM & Costerton JW (2002) Biofilms: survival
mechanisms of clinically relevant microorganisms. Clin
Microbiol Rev 15: 167–193.
Dorko E, Pilipcinec E & Tkacikova L (2002) Candidal urinary
tract infections caused by non-albicans Candida species. Folia
Microbiol 47: 182–184.
Dostal J, Dlouha H, Malon P, Pichova I & Hruskova-
Heidingsfeldova O (2005) The precursor of secreted aspartic
proteinase Sapp1p from Candida parapsilosis can be activated
both autocatalytically and by a membrane-bound processing
proteinase. Biol Chem 386: 791–799.
Douglas LJ (2003) Candida biofilms and their role in infection.
Trends Microbiol 11: 30–36.
Eggimann P, Garbino J & Pittet D (2003) Epidemiology of
Candida species infections in critically ill non-
immunosuppressed patients. Lancet Infect Dis 3: 685–702.
Ellepola ANB & Morrison CJ (2005) Laboratory diagnosis of
invasive Candidiasis. J Microbiol 43: 65–84.
Enger L, Joly S, Pulol C, Simonson M, Pfaller MA & Soll R (2001)
Cloning and characterization of a complex DNA
fingerprinting probe for Candida parapsilosis. J Clin Microbiol
39: 658–669.
Fell JW, Statzell-Tallman A, Lutz MJ & Kurtzman (1992) Partial
rRNA sequences in marine yeasts; a model for identification of
marine eukaryotes. Mol Mar Biol Biotech 1: 175–186.
Fidel PL, Vazquez JA & Sobel JD (1999) Candida glabrata: review
of epidemiology, pathogenesis, and clinical disease with
comparison to C. albicans. Clin Microbiol Rev 12: 80–96.
Fu Y, Ibrahim AS, Fonzi W, Zhou X, Ramos CF & Ghannoum MA
(1997) Cloning and characterization of a gene (LIPI) which
encodes a lipase from the pathogenic yeast Candida albicans.
Microbiology 143: 331–340.
Furlaneto-Maia L, Specian A, Bizerra F, Oliveira M & Furlaneto
M (2007) In vitro evaluation of putative virulence attributes of
oral isolates of Candida spp. obtained from elderly healthy
individuals. Mycopathologia 166: 209–217.
Fusek M, Smith EA, Monod M, Dunn BM & Foundling SI (1994)
Extracellular aspartic proteinases from Candida albicans,
Candida tropicalis, and Candida parapsilosis yeasts differ
substantially in their specificities. Biochemistry 33: 9791–9799.
Gacser A, Schafer W, Nosanchuk JS, Salomon S & Nosanchuk JD
(2007a) Virulence of Candida parapsilosis, Candida
orthopsilosis, and Candida metapsilosis in reconstituted human
tissue models. Fungal Genet Biol 44: 1336–1341.
Gacser A, Trofa D, Schafer W & Nosanchuk JD (2007b) Targeted
gene deletion in Candida parapsilosis demonstrates the role of
secreted lipase in virulence. J Clin Invest 117: 3049–3058.
Galan-Ladero MA, Blanco MT, Sacristan B, Fernandez-Calderan
MC, Perez-Giraldo C & Gomez-Garcia AC (2010) Enzymatic
activities of Candida tropicalis isolated from hospitalized
patients. Med Mycol 48: 207–210.
Ghannoum MA (2000) Potential role of phospholipases in
virulence and fungal pathogenesis. Clin Microbiol Rev 13:
122–143.
Ghannoum MA & Rice LB (1999) Antifungal agents: mode of
action, mechanisms of resistance, and correlation of these
mechanisms with bacterial resistance. Clin Microbiol Rev 12:
501–517.
Gonzalez GM, Elizondo M & Ayala J (2008) Trends in species
distribution and susceptibility of bloodstream isolates of
Candida collected in Monterrey, Mexico, to seven antifungal
agents: results of a 3-year (2004 to 2007) surveillance study. J
Clin Microbiol 46: 2902–2905.
Gonzalez Gravina H, Gonzalez de Moran E, Zambrano O, Lozano
Chourio M, Rodriguez de Valero S, Robertis S & Mesa L
(2007) Oral candidiasis in children and adolescents with
cancer. Identification of Candida spp. Med Oral Patol Oral Cir
Bucal 12: 419–423.
Gow NAR, Brown AJP & Odds FC (2002) Fungal morphogenesis
and host invasion. Curr Opin Microbiol 5: 366–371.
Groll AH, Piscitelli SC & Walsh TJ (2001) Antifungal
pharmacodynamics: concentration–effect relationship in vitro
and in vivo. Pharmacotherapy 21: 133S–148S.
Hagerty JA, Ortiz J, Reich D & Manzarbeitia C (2003) Fungal
infections in solid organ transplant patients. Surg Infect
(Larchmt) 4: 263–271.
Hajjeh RA, Sofair AN, Harrison LH et al. (2004) Incidence of
bloodstream infections due to Candida species and in vitro
susceptibilities of isolates collected from 1998 to 2000 in a
population-based active surveillance program. J Clin Microbiol
42: 1519–1527.
Hasan F, Xess I, Wang X, Jain N & Fries BC (2009) Biofilm
formation in clinical Candida isolates and its association with
virulence. Microbes Infect 11: 753–761.
Hawser SP & Douglas LJ (1994) Biofilm Formation by Candida
species on the surface of catheter materials in vitro. Infect
Immun 62: 915–921.
Hawser SP & Douglas LJ (1995) Resistance of Candida albicans
biofilms to antifungal agents in vitro. Antimicrob Agents Ch 39:
2128–2131.
Haynes KA (2001) Virulence in Candida species. Trends Microbiol
9: 591–596.
Haynes KA & Westerneng TJ (1996) Rapid identification of
Candida albicans, C. glabrata, C. parapsilosis and C. krusei by
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 301

species-specific PCR of large subunit ribosomal DNA. J Med
Microbiol 44: 390–396.
Hazen KC, Plotkin BJ & Klimas DM (1986) Influence of growth
conditions on cell surface hydrophobicity of Candida albicans
and Candida glabrata. Infect Immun 54: 269–271.
Henriques M, Gasparetto K, Azeredo J & Oliveira R (2002)
Experimental methodology to quantify Candida albicans cell
surface hydrophobicity. Biotechnol Lett 24: 1111–1115.
Hoyer LL, Fundyga R, Hecht JE, Kapteyn JC, Klis FM & Arnold J
(2001) Characterization of agglutinin-like sequence genes
from non-albicans Candida and phylogenetic analysis of the
ALS family. Genetics 157: 1555–1567.
Isenberg HD, Tucci V, Cintron F, Singer C, Weinstein GS & Tyras
DH (1989) Single-source outbreak of Candida tropicalis
complicating coronary bypass surgery. J Clin Microbiol 27:
2426–2428.
Jain N, Kohli R, Cook E, Gialanella P, Chang T & Fries BC (2007)
Biofilm formation by and antifungal susceptibility of Candida
isolates from urine. Appl Environ Microb 73: 1697–1703.
Jayatilake JA, Samaranayake YH, Cheung LK & Samaranayake LP
(2006) Quantitative evaluation of tissue invasion by wild type,
hyphal and SAP mutants of Candida albicans, and non-
albicans Candida species in reconstituted human oral
epithelium. J Oral Pathol Med 35: 481–491.
Joachim H & Polayes S (1940) Subculture endocarditis and
systemic mycosis (monilia). JAMA 115: 205–208.
Kantarciolu AS & Yucel A (2002) Phospholipase and protease
activities in clinical Candida isolates with reference to the
sources of strains. Mycoses 45: 160–165.
Kauffman CA, Vazquez JA, Sobel JD et al. (2000) Prospective
multicenter surveillance study of funguria in hospitalized
patients. Clin Infect Dis 30: 14–18.
Kiehn TE, Edwards FF & Armstrong D (1980) The prevalence of
yeasts in clinical specimens from cancer patients. Am J Clin
Pathol 73: 518–521.
Kikutani H & Makino S (1992) The murine autoimmune
diabetes model: NOD and related strains. Adv Immunol 51:
285–322.
Kobayashi C, Fernandes ODFL, Miranda K, Sousa E & Silva MRR
(2004) Candiduria in hospital patients: a study prospective.
Mycopathologia 158: 49–52.
Kojic EM & Darouiche RO (2004) Candida infections of medical
devices. Clin Microbiol Rev 17: 255–267.
Kontoyiannis DP, Vaziri I, Hanna HA, Boktour M, Thornby J,
Hachem R, Bodey GP & Raad II (2001) Risk Factors for
Candida tropicalis fungemia in patients with cancer. Clin Infect
Dis 33: 1676–1681.
Kossoff EH, Buescher ES & Karlowicz MG (1998) Candidemia in
a neonatal intensive care unit: trends during fifteen years and
clinical features of 111 cases. Pediatr Infect Dis J 17: 504–508.
Krcmery V (1999a) Torulopsis glabrata an emerging yeast
pathogen in cancer patients. Int J Antimicrob Ag 11: 1–6.
Krcmery V & Barnes AJ (2002) Non-albicans Candida spp.
causing fungaemia: pathogenicity and antifungal resistance.
J Hosp Infect 50: 243–260.
Krcmery VJ (1999b) Candidemia in cancer patients: risk factors
and outcome in 140 episodes from a single cancer institution.
Acta Chemoth 5: 133–145.
Kucharınova S, Tournu H, Holtappels M, Van Dijck P & Lagrou K
(2010) In vivo efficacy of anidulafungin against mature
Candida albicans biofilms in a novel rat model of catheter-
associated candidiasis. Antimicrob Agents Ch 54: 4474–4475.
Kuhn DM, Chandra J, Mukherjee PK & Ghannoum MA (2002)
Comparison of biofilms formed by Candida albicans and
Candida parapsilosis on bioprosthetic surfaces. Infect Immun
70: 878–888.
Kumamoto CA (2002) Candida biofilms. Curr Opin Microbiol 5:
608–611.
Kumar VG, Latha R, Vedhagiri K, Sathiamoorthi T, Jayarani G,
Sasikala R, Selvin J & Natarajaseenivasan K (2009)
Phospholipase C, proteinase and hemolytic activities of
Candida spp. isolated from pulmonary tuberculosis patients.
J Mycol Med 19: 3–10.
Larone D (2002) Medically Important Fungi; A Guide to
Identification. 4th edn. ASM Press, Washington.
Lass-Florl C (2009) The changing face of epidemiology of
invasive fungal disease in Europe. Mycoses 52: 197–205.
Lattif AA, Mukherjee PK, Chandra J, Swindell K, Lockhart SR,
Diekema DJ, Pfaller MA & Ghannoum MA (2009)
Characterization of biofilms formed by Candida parapsilosis,
C. metapsilosis, and C. orthopsilosis. Int J Med Microbiol 300:
265–270.
Lermann U & Morschhauser J (2008) Secreted aspartic proteases
are not required for invasion of reconstituted human epithelia
by Candida albicans. Microbiology 154: 3281–3295.
Lewis K (2001) Ridlle of biofilm resistance. Antimicrob Agent Ch
45: 999–1007.
Liguori G, Onofrio V, Lucariello A, Galle F, Signoriello G, Colella
G, D’Amora M & Rossano F (2009) Oral candidiasis: a
comparison between conventional methods and multiplex
polymerase chain reaction for species identification. Oral
Microbiol Immun 24: 76–78.
Luo G, Samaranayake LP, Cheung BPK & Tang G (2004) Reverse
transcriptase polymerase chain reaction (RT-PCR) detection
of HLP gene expression in Candida glabrata and its possible
role in vitro haemolysin production. APMIS 112: 283–290.
Lupetti A, Danesi R, Campa M et al. (2002) Molecular basis of
resistance to azole antifungals. Trends Mol Med 8: 76–81.
Luque AG, Biasoli MS, Tosello ME, Binolfi A, Lupo S & Magaro
HM (2009) Oral yeast carriage in HIV-infected and non-
infected populations in Rosario, Argentina. Mycoses 52: 53–59.
Manns JM, Mosser DM & Buckley HR (1994) Production of a
hemolytic factor by Candida albicans. Infect Immun 62:
5154–5156.
Manzano-Gayosso P, Hernandez-Hernandez F, Zavala-Velasquez
N, Mendez-Tovar LJ, Naquid-Narvaez JM, Torres-Rodriguez
JM & Lopez-Martinez R (2008) Candiduria in type 2 diabetes
mellitus patients and its clinical significance. Candida spp.
antifungal susceptibility. Rev Med Inst Mex Seguro Soc 46:
603–610.
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
302 S. Silva et al.

Martin MV (1979) Germ-tube formation by oral strains of
Candida tropicalis. J Med Microbiol 12: 187–194.
Martins M, Henriques M, Ribeiro AP, Fernandes R, Goncalves V,
Seabra A, Azeredo J & Oliveira R (2010a) Oral Candida
carriage of patients attending a dental clinic in Braga, Portugal.
Rev Iberoam Micol 27: 119–124.
Martins M, Uppuluri P, Thems DD, Cleary IA, Henriques M,
Lopez-Ribot LJ & Oliveira R (2010b) Presence of extracellular
DNA in the Candida albicans matrix and its contribution to
biofilms. Mycopathologia 5: 323–331.
Merkerova M, Dostal J, Hradilek M, Pichova I & Hruskova-
Heidingsfeldova O (2006) Cloning and characterization of
Sapp2p, the second aspartic proteinase isoenzyme from
Candida parapsilosis. FEMS Yeast Res 6: 1018–1026.
Messer SA, Jones RN & Fritsche TR (2006) International
surveillance of Candida spp. and Aspergillus spp.: report from
the SENTRY Antimicrobial Surveillance Program (2003). J
Clin Microbiol 44: 1782–1787.
Miranda LN, van der Heijden IM, Costa SF, Sousa AP, Sienra RA,
Gobara S, Santos CR, Lobo RD, Pessoa VP & Levin AS (2009)
Candida colonisation as a source for candidaemia. J Hosp
Infect 72: 9–16.
Mukherjee PK & Chandra J (2004) Candida biofilm resistance.
Drug Resist Update 7: 301–309.
Naglik JR, Moyes D, Makwana J et al. (2008) Quantitative
expression of the Candida albicans secreted aspartyl proteinase
gene family in human oral and vaginal candidiasis.
Microbiology 154: 3266–3280.
Negri M, Goncalves V, Silva S, Henriques M, Azeredo J & Oliveira
R (2010a) Crystal violet staining to quantify Candida adhesion
to epithelial cells. Brit J Biomed Sci 67: 120–125.
Negri M, Martins M, Henriques M, Svidzinski T, Azeredo J &
Oliveira R (2010b) Examination of potential virulence factors
of Candida tropicalis clinical isolates from hospitalized
patients. Mycopathologia 169: 175–182.
Neugnot V, Moulin G, Dubreucq E & Bigey F (2002) The lipase/
acyltransferase from Candida parapsilosis: molecular cloning
and characterization of purified recombinant enzymes. Eur J
Biochem 269: 1734–1745.
Nosek J, Holesova Z, Kosa P, Gacser A & Tomaska L (2009)
Biology and genetics of the pathogenic yeast Candida
parapsilosis. Curr Genet 55: 497–509.
Nucci M & Colombo AL (2007) Candidemia due to Candida
tropicalis: clinical, epidemiologic, and microbiologic
characteristics of 188 episodes occurring in tertiary care
hospitals. Diagn Micr Infec Dis 58: 77–82.
Odds F (1988) Candida and Candidiasis. 2nd edn. Bailliere
Tindall, London.
Orazio R, Scordino F, Pernice I, Passo CL & Criseo G (2009) A
multiplex PCR protocol for rapid identification of Candida
glabrata and its phylogenetically related species Candida
nivariensis and Candida bracarensis. J Microbiol Meth 79:
117–120.
Panagoda GJ, Ellepola ANB & Samaranayake LP (2001) Adhesion
of Candida parapsilosis to epithelial and acrylic surfaces
correlates with cell surface hydrophobicity. Mycoses 44: 29–35.
Passos XS, Sales WS, Maciel PJ, Costa CR, Miranda KC, Lemos
JA, Batista MA & Silva MRR (2005) Candida colonization in
intensive care unit patients’ urine. Mem I Oswaldo Cruz 100:
925–928.
Pfaller MA (1992) Laboratory aids in the diagnosis of invasive
candidiasis. Mycopathology 120: 65–72.
Pfaller MA & Diekema DJ (2007) Epidemiology of invasive
Candidiasis: a persistent public health problem. Clin Microbiol
Rev 20: 133–163.
Pfaller MA, Jones RN, Messer SA, Edmond M & Wenzel RP
(1998) National surveillance of nosocomial blood stream
infection due to Candida albicans: frequency of occurrence
and antifungal susceptibility in the SCOPE Program. Diagn
Micr Infec Dis 31: 327–332.
Pfaller MA, Boyken L, Hollis RJ et al. (2005) In vitro
susceptibilities of clinical isolates of Candida species,
Cryptococcus neoformans, and Aspergillus species to
itraconazole: global survey of 9,359 isolates tested by clinical
and laboratory standards institute broth microdilution
methods. J Clin Microbiol 43: 3807–3810.
Pfaller MA, Castanheira M, Messer AS, Moet GJ & Jones RN
(2010) Variation in Candida spp. distribution and antifungal
resistance rates among bloodstream infection isolates by
patient age: report from the SENTRY Antimicrobial
Surveillance Program (2008–2009). Diagn Micr Infec Dis 68:
278–283.
Pichova I, Pavlickova L, Dostal J, Dolejsi E, Hruskova-
Heidingsfeldova O, Weber J, Ruml T & Soucek M (2001)
Secreted aspartic proteases of Candida albicans, Candida
tropicalis, Candida parapsilosis and Candida lusitaniae:
inhibition with peptidomimetic inhibitors. Eur J Biochem 268:
2669–2677.
Polak AM, Scholer HJ & Wall M (1982) Combination of action of
experimental candidiasis, cryptococcosis, and aspergilosis in
mice. Chemotherapy 28: 461–479.
Ramage G, Wickes BL & Lopez-Ribot JL (2001) Biofilms of
Candida albicans and their associated resistance antifungal
agents. Antimicrob Agents Ch 20: 43–44.
Ramage G, Martinez JP & Lopez-Ribot JL (2006) Candida
biofilms on implanted biomaterials: a clinically significant
problem. FEMS Yeast Res 6: 979–986.
Redding SW, Kirkpatrick WR, Coco BJ, Sadkowski L, Fothergill
AW, Rinaldi MG, Eng TY & Patterson TF (2002) Candida
glabrata oropharyngeal candidiasis in patients receiving
radiation treatment for head and neck cancer. J Clin Microbiol
40: 1879–1881.
Rex JH, Rinaldi MG, Galgiani JN & Pfaller MA (1995) Resistance
of Candida species to fluconazole. Antimicrob Agents Ch 39:
1–8.
Rho J, Shin J, Song J, Park M, Kee S, Jang S, Park Y, Suh S & Ryang
D (2004) Molecular investigation of two consecutive
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 303

nosocomial clusters of Candida tropicalis candiduria using
pulsed-field gel electrophoresis. J Microbiol 42: 80–86.
Rigby S, Procop GW, Haase G et al. (2002) Fluorescence In Situ
hybridization with peptide nucleic acid probes for rapid
identification of Candida albicans directly from blood culture
bottles. J Clin Microbiol 40: 2182–2186.
Roling EE, Klepser ME, Wasson A, Lewis RE, Ernest EJ & Pfaller
MA (2002) Antifungal activities of fluconazole, caspofungin
(MK0991), and anidulafungin (LY303366) alone and in
combination against Candida spp and Cryptococcus
neoformans via time kill methods. Diagn Micr Infec Dis 2:
73–85.
Ruan S & Hsueh P (2009) Invasive candidiasis: an overview from
Taiwan. J Med Assoc 108: 443–451.
Ruchel R (1999) Proteinases of pathogenic fungi. Mycoses 42:
48–52.
Ruchel R, de Bernardis F, Ray TL, Sullivan PA & Cole GT (1992)
Candida acid proteinases. J Med Vet Mycol 30: 123–132.
Ruping MJ, Vehreschild JJ & Cornely OA (2008) Patients at high
risk of invasive fungal infections: when and how to treat. Drugs
68: 1941–1962.
Saiman L, Ludington E, Dawson JD et al. (2001) Risk factors for
Candida species colonization of neonatal intensive care unit
patients. Pediatr Infect Dis J 20: 1119–1124.
Samaranayake LP, Fidel PL, Naglik JR, Sweet SP, Teanpaisan R,
Coogan MM, Blignaut E & Wanzala P (2002) Fungal infections
associated with HIV infection. Oral Dis 8: 151–160.
Samaranayake YH & Samaranayake LP (2001) Experimental oral
candidiasis in animal models. Clin Microbial Rev 14: 389–429.
Sampaio P, Gusmao L, Correia A, Alves C, Rodrigues AG, Pina-
Vaz C, Amorim A & Pais C (2005) New microsatellite
multiplex PCR for Candida albicans strain typing reveals
microevolutionary changes. J Clin Microbiol 43: 3869–3876.
Sanglard D & Odds FC (2002) Resistance of Candida species to
antifungal agents: molecular mechanisms and clinical
consequences. Lancet Infect Dis 2: 73–85.
Shin JH, Kee SJ, Shin MG, Kim SH, Shin DH, Lee SK, Suh SP &
Ryang DW (2002) Biofilm production by isolates of Candida
species recovered from nonneutropenic patients: comparison
of bloodstream isolates with isolates from other sources. J Clin
Microbiol 40: 1244–1248.
Silva S, Henriques M, Martins A, Oliveira R, Williams D &
Azeredo J (2009a) Biofilms of non-Candida albicans Candida
species: quantification, structure and matrix composition.
Med Mycol 47: 681–689.
Silva S, Henriques M, Oliveira R, Azeredo J, Malic S, Hooper SJ &
Williams DW (2009b) Characterization of Candida
parapsilosis infection of an in vitro reconstituted human oral
epithelium. Eur J Oral Sci 117: 669–675.
Silva S, Hooper SJ, Henriques M, Oliveira R, Azeredo J &
Williams DW (2010a) The role of secreted aspartyl proteinases
in Candida tropicalis invasion and damage of oral mucosa.
Clin Microbiol Infec 17: 264–272.
Silva S, Negri M, Henriques M, Oliveira R, Williams D & Azeredo
J (2010b) Silicone colonisation by non-Candida albicans
Candida species in the presence of urine. J Med Microbiol 59:
747–754.
Solomon SL, Khabbaz RF, Parker RH, Anderson RL, Geraghty
MA, Furman RM & Martone WJ (1984) An outbreak of
Candida parapsilosis bloodstream infections in patients
receiving parenteral nutrition. J Infect Dis 149: 98–102.
Scozo G, Kardos G, McNicholas PM et al. (2007) Correlation of
posaconazole minimum fungicidal concentration and time kill
test against nine Candida species. J Antimicrob Chemoth 60:
1004–1009.
Sullivan DJ, Westerneng TJ, Haynes KA, Bennett DE & Coleman
DC (1995) Candida dubliniensis sp. nov: phenotypic and
molecular characterization of a novel species associated with
oral candidosis in HIV-infected individuals. Microbiology 141:
1507–1521.
Symersky J, Monod M & Foundling SI (1997) High-resolution
structure of the extracellular aspartic proteinase from Candida
tropicalis yeast. Biochemistry 36: 12700–12710.
Tavanti A, Davidson AD, Gow NAR et al. (2005) Candida
ortopsilosis and Candida metapsilosis spp. nov to replace
Candida parapsilosis groups II and III. J Clin Microbiol 43:
284–292.
Togni G, Sanglard D, Falchetto R & Monod M (1991) Isolation
and nucleotide sequence of the extracellular acid protease gene
(ACP) from the yeast Candida tropicalis. FEBS Lett 286:
181–185.
Tortorano AM, Kibbler C, Peman J, Bernhardt H, Klingspor L &
Grillot R (2006) Candidaemia in Europe: epidemiology and
resistance. Int J Antimicrob Ag 27: 359–366.
Trick WE, Fridkin SK, Edwards JR, Hajjeh RA & Gaynes RP
(2002) Secular trend of hospital-acquired candidemia among
intensive care unit patients in the United States during
1989–1999. Clin Infect Dis 35: 627–630.
Trofa D, Gacser A & Nosanchuk JD (2008) Candida parapsilosis,
an emerging fungal pathogen. Clin Microbiol Rev 21: 606–625.
Vandeputte P, Tronchin C, Berges T et al. (2007) Reduced
susceptibility to polyenes associated with a missense mutation
in the ERG6 gene in a clinical isolate of Candida glabrata with
pseudohyphal growth. Antimicrob Agents Ch 51: 982–990.
Vazquez JA, Sanchez V, Dmuchowski C, Dembry LM, Sobel JD &
Zervos MJ (1993) Nosocomial acquisition of Candida albicans:
an epidemiologic study. J Infect Dis 168: 195–201.
Vermes A, Guchelaar HJ & Dankert J (2000) Flucytosine: a review
of its pharmacology, clinical indications, pharmacokinetics,
toxicity and drug interactions. J Antimicrob Chemoth 46:
171–179.
Verweij PE, Breuker IM, Rijs AJ & Meis JF (1999) Comparative
study of seven commercial yeast identification systems. J Clin
Pathol 52: 271–273.
Vilain S, Pretorius JM, Theron J & Brozel VS (2009) DNA as an
adhesin: Bacillus cereus requires extracellular DNA to form
biofilm. Appl Environ Microb 75: 2861–2868.
Voss A, Hollis RJ, Pfaller MA, Wenzel RP & Doebbeling BN
(1994) Investigation of the sequence of colonization and
ª 2011 Federation of European Microbiological Societies FEMS Microbiol Rev 36 (2012) 288–305Published by Blackwell Publishing Ltd. All rights reserved
304 S. Silva et al.

candidemia in nonneutropenic patients. J Clin Microbiol 32:
975–980.
Walsh TJ (2002) Echinocandins – an advance in the primary
treatment of invasive candidiasis. New Engl J Med 347:
2070–2072.
Watanabe T, Takano M, Murakami M, Tanaka H, Matsuhisa A,
Nakao N, Mikami T, Suzuki M & Matsumoto T (1999)
Characterization of a haemolytic factor from Candida albicans.
Microbiology 145: 689–694.
Weems JJ (1992) Candida parapsilosis: epidemiology,
pathogenicity, clinical manifestations, and antimicrobial
susceptibility. Clin Infect Dis 14: 756–766.
Williams DW, Wilson MJ, Lewis MA & Potts (1995)
Identification of Candida species by PCR and restriction
fragment polymorphism analysis of intergenic spacer regions
of ribosomal DNA. J Clin Microbiol 33: 2476–2479.
Yoshio O & Kouji G (2006) Antigenicity of Candida tropicalis
strain cells cultured at 27 and 37 1C. FEMS Immunol Med Mic
46: 438–443.
Zaugg C, Borg-von Zepelin M, Reichard U, Sanglard D & Monod
M (2001) Secreted aspartic proteinase family of Candida
tropicalis. Infect Immun 69: 405–412.
FEMS Microbiol Rev 36 (2012) 288–305 ª 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Non-Candida albicans Candida species pathogenicity 305