Embed Size (px)
Citation preview

COEXISTENCE SHAPES ANTIPREDATOR DEFENSES
3Present address: European Commission, Joint Research Centre, Institute for Environment and
Sustainability, Water Resources Unit, Ispra, Italy. E-mail: [email protected]
Rapid evolution of constitutive and inducible defenses against an invasive predator 1
ANA L. NUNES1,2,3
, GERMÁN ORIZAOLA2, ANSSI LAURILA
2, AND RUI REBELO
1 2
3
1Centro de Biologia Ambiental, D.B.A., Faculdade de Ciências da Universidade de Lisboa, 4
Lisboa, Portugal 5
2Animal Ecology/Department of Ecology and Genetics, Evolutionary Biology Centre, Uppsala 6
University, Uppsala, Sweden7

NUNES ET AL.
2
Abstract 8
Invasive alien predators can impose strong selection on native prey populations and induce rapid 9
evolutionary change in the invaded communities. However, studies on evolutionary responses to 10
invasive predators are often complicated by the lack of replicate populations differing in 11
coexistence time with the predator, which would allow determining how prey traits change during 12
the invasion. The red swamp crayfish Procambarus clarkii has invaded many freshwater areas 13
worldwide, with negative impacts for native fauna. Here, we examined how coexistence time 14
shapes antipredator responses of the Iberian waterfrog (Pelophylax perezi) to the invasive crayfish 15
by raising tadpoles from five populations differing in historical exposure to P. clarkii (30 years, 20 16
years or no coexistence). Tadpoles from non-invaded populations responded to the presence of P. 17
clarkii with behavioral plasticity (reduced activity), whereas long-term invaded populations 18
showed canalized antipredator behavior (constant low activity level). Tadpoles from one of the 19
long-term invaded populations responded to the crayfish with inducible morphological defenses 20
(deeper tails), reflecting the use of both constitutive and inducible antipredator defenses against the 21
exotic predator by this population. Our results suggest that, while naive P. perezi populations 22
responded behaviorally to P. clarkii, the strong predation pressure imposed by the crayfish has 23
induced the evolution of qualitatively different antipredator defenses in populations with longer 24
coexistence time. These responses suggest that strong selection by invasive predators may drive 25
rapid evolutionary change in invaded communities. Examining responses of prey species to 26
biological invasions using multiple populations will help us better forecast the impact of invasive 27
predators in natural communities. 28
Keywords: coexistence time, invasive predator, plastic and constitutive defenses, population, 29
rapid evolution, predator recognition 30
31

NUNES ET AL.
3
Introduction 32
Predation is an important selective force acting on the fitness of prey organisms, and selection on 33
prey populations for improved ability to escape or deter predation is often strong (Lima and Dill 34
1990). Prey exhibit a wide variety of antipredator defenses involving modifications in behavior, 35
morphology and life-history that reduce their vulnerability to predators (Lima and Dill 1990, 36
Tollrian and Harvell 1999, Benard 2004). These defenses can be expressed constitutively, or be 37
inducible and only expressed when predators are present (Tollrian and Harvell 1999). Constitutive, 38
or canalized, defenses are common in habitats under constant and high predator pressure (Clark 39
and Harvell 1992, Moran 1992, Edgell et al. 2009). On the contrary, inducible, or plastic, defenses 40
evolve in habitats with spatially or temporally variable predation risk and are favored when 41
reliable cues from predation are available (Padilla and Adolph 1996). Inducible defenses have a 42
fitness advantage when predators are present, but incur costs in the absence of predators, 43
preventing them from becoming constitutive (Clark and Harvell 1992, Tollrian and Harvell 1999). 44
While constitutive and plastic defenses are often considered as independent alternatives, selection 45
for both types of defenses can act simultaneously on different traits, and the expression and 46
integration of the different defenses may vary among populations (DeWitt et al. 1999, Poitrineau 47
et al. 2004, Bourdeau 2012). 48
Successful predator recognition and effective avoidance strategies typically occur when predator 49
and prey share a common evolutionary history (Strauss et al. 2006, Sih et al. 2010). Invasive alien 50
predators are spreading over new environments at an unprecedented rate, resulting in new 51
predator-prey interactions between organisms sharing no evolutionary history (Thompson 1998, 52
Strauss et al. 2006). Invaded prey populations often suffer dramatic declines and even extinctions 53
due to their inability to respond with appropriate antipredator defenses (Cox and Lima 2006, 54

NUNES ET AL.
4
Strauss et al. 2006, Salo et al. 2007, Sih et al. 2010). However, by imposing strong selection 55
pressure upon native organisms, invasive predators may also induce rapid evolutionary changes, 56
through which prey may evolve the ability to recognize and respond to them (Schlaepfer et al. 57
2005, Strauss et al. 2006, Sih et al. 2010). For rapid evolution to occur, selection pressure imposed 58
by the exotic predator has to be strong and consistent, and native populations have to be 59
genetically variable in susceptibility to the predator (Strauss et al. 2006). Indeed, rapid adaptive 60
evolution in native prey species as a response to introduced predators has been reported in several 61
taxa including mollusks, crustaceans, fish, amphibians and reptiles (Kiesecker and Blaustein 1997, 62
O’Steen et al. 2002, Losos et al. 2004, Freeman and Byers 2006, Fisk et al. 2007). 63
However, some prey are able to innately detect and elicit adaptive responses to stimuli of novel 64
predators in the absence of a common evolutionary history (Epp and Gabor 2008, Peluc et al. 65
2008, Sündermann et al. 2008, Rehage et al. 2009). This ability may be due to the latter’s 66
phylogenetic or phenotypic similarity to native predators co-occurring with the prey (Epp and 67
Gabor 2008, Rehage et al. 2009). Prey may also exhibit generalized antipredator responses, 68
whereby exposure to predation risk induces the expression of defenses, regardless of previous 69
direct experience with a particular predator (e.g. response to a general fish odor or to a large 70
moving object; Magurran 1990, Gherardi et al. 2011). This can be facilitated if detection of 71
predators relies on general predation cues, such as cues associated with the predator’s diet or alarm 72
cues released by startled or attacked prey (Sündermann et al. 2008, Ferrari et al. 2010b). 73
Alternatively to innate responses, prey may learn to identify and respond to alien predators 74
through experience by pairing novel predator scents with cues released by attacked or consumed 75
conspecifics and hence learn to ‘label’ exotic predators as a threat (reviewed in Ferrari et al. 76
2010b). 77

NUNES ET AL.
5
The potential for the evolution of antipredator responses may vary among populations due to 78
factors such as the intensity of selection, the amount of genetic variation within prey populations 79
or the genetic architecture of the traits (Strauss et al. 2006). Studies using single or few invaded 80
populations may, thus, fail to fully show the complexity of the evolutionary responses of natives to 81
introduced predators. Therefore, designing studies using populations with differing co-82
evolutionary history with the invasive predator may be crucial for better forecasting the long-term 83
impacts of biological invasions (see e.g. Kiesecker and Blaustein 1997, Trussell and Smith 2000, 84
Edgell et al. 2009). 85
Freshwater environments, which have been widely invaded by exotic species, are particularly 86
vulnerable to introduced predators (Cox and Lima 2006). One of the most conspicuous non-native 87
predator species introduced worldwide is the red swamp crayfish (Procambarus clarkii), native to 88
southern USA and northern Mexico (Gherardi 2006). This invasive species was first recorded in 89
Europe in the Iberian Peninsula in 1973 and has rapidly spread throughout the area (Holdich et al. 90
2009). P. clarkii is an extraordinarily successful invader, often altering the dynamics of the 91
ecosystems it colonizes (e.g. Geiger et al. 2005). Crayfish are efficient predators of larvae of many 92
amphibian species (e.g. Cruz and Rebelo 2005, Cruz et al. 2008). 93
Tadpoles of the Iberian waterfrog (Pelophylax perezi) exhibit typical plastic behavioral (reduced 94
activity level) and morphological (shorter and deeper headbodies and deeper tails) antipredator 95
responses when exposed to chemical stimuli of native aquatic predators (Richter-Boix et al. 2007, 96
Polo-Cavia et al. 2010, Gómez-Mestre and Díaz-Paniagua 2011, Nunes 2012, Nunes et al. 2013). 97
Although previous studies have not detected behavioral or morphological responses of P. perezi to 98
P. clarkii (Gómez-Mestre and Díaz-Paniagua 2011, Nunes 2012, Nunes et al. 2013), Gómez-99
Mestre and Díaz-Paniagua (2011) found that induced morphological defenses elicited by P. perezi 100

NUNES ET AL.
6
in the presence of a native larval dragonfly predator (a deeper and more pigmented tail) increased 101
tadpole survival in the presence of P. clarkii. 102
In this study, we investigated how coexistence time with P. clarkii influences the ability of P. 103
perezi to respond to this invasive predator. We compared behavioral and morphological responses 104
of five P. perezi populations characterized by different exposure times to P. clarkii: long (ca. 30 105
yr), medium (ca. 20 yr) or no coexistence time with the predator. Assuming that P. clarkii imposes 106
strong selection pressure upon P. perezi populations (Cruz and Rebelo 2005, Cruz et al. 2008), we 107
expect that tadpoles from long-term invaded populations will show antipredator defenses towards 108
the crayfish (such as alterations in morphology and/or a reduction in activity rates), while 109
individuals from non-invaded populations will show no responses to this predator. The medium-110
term invaded population should present intermediate responses, or at least respond differently 111
from the non-invaded populations. 112
113
Methods 114
Study species and populations 115
The Iberian waterfrog (P. perezi, see PLATE 1) is the most abundant and widely distributed 116
anuran in Portugal (Loureiro et al. 2008). Although inhabiting a wide variety of aquatic habitats, it 117
reproduces preferably in permanent water bodies (Almeida et al. 2001). We selected five 118
populations of P. perezi from southern Portugal, a region where P. clarkii is currently widespread 119
and where there are no present or historical records of native crayfishes (Almaça 1991). Two 120
populations were collected in the area of the Caia River, the region where P. clarkii was first 121
recorded in Portugal in 1979 (Ramos and Pereira 1981; Table 1, Fig. 1), and they constitute the 122
populations with longer coexistence time with P. clarkii in this study (10-15 P. perezi generations, 123

NUNES ET AL.
7
hereafter LC populations). A population with shorter coexistence time (hereafter SC) was 124
collected at the Melides River, an area that was invaded by the crayfish ca. 1990 (R. Rebelo pers. 125
obs.; Table 1, Fig. 1). Finally, we collected two P. perezi populations in the Monchique region, 126
which have never been in contact with P. clarkii (No Coexistence, hereafter NC; Table 1, Fig. 1). 127
The Monchique region is characterized by numerous natural springs, streams and rivers, where 128
topographic barriers and the distance to the crayfish introduction source may have prevented the 129
invasion by P. clarkii until now (Kerby et al. 2005, Cruz and Rebelo 2007). Repeated surveys 130
using baited traps in 2009 proved this area devoid of crayfish (A.L. Nunes and R. Rebelo, 131
unpubl.). Trapping and visual observations confirmed that dense P. clarkii populations coexisted 132
with P. perezi in LC and SC populations. All study populations were collected from slow-flowing 133
sections of streams harboring a rich fauna of native predators (insects, fish and snakes) or from 134
rice fields with immediate connection to the river. Although the Caia river harbors a more diverse 135
fish community (9 species) than the other two areas (5 species; Ribeiro et al. 2007), the former 136
community is composed of mostly detritivorous and insectivorous species not known to actively 137
prey on tadpoles. Overall, apart from the differences in crayfish invasion history, localities were 138
selected to be as identical as possible on other ecological parameters that could possibly influence 139
the traits studied here (e.g. vegetation structure, water temperature, shading, etc.). 140
Experimental procedure 141
During June and July 2009, we collected 8-12 freshly laid egg clutches of P. perezi at each of the 142
study populations (Table 1). The eggs were taken to the laboratory in the field station of Centro de 143
Biologia Ambiental in Grândola (38º06’N, 8º34’W), where the experiment took place. Each clutch 144
was divided in two and kept in 1L containers filled with well water until the start of the 145
experiment. Adult crayfish were caught in streams close to the field station using baited funnel 146

NUNES ET AL.
8
traps, kept in 28 × 20 × 14 cm containers and fed commercial fish food ad libitum until the start of 147
the experiment. 148
The experiment was a 5 x 2 factorial design with population origin crossed with two predator 149
treatments. When most of the larvae reached Gosner developmental stage 25 (complete absorption 150
of gills and active feeding; Gosner 1960), we divided them haphazardly into groups of 10 151
individuals, which were then allocated to the experimental containers (39 × 28 × 28 cm) filled with 152
10L of well water (day 1 of the experiment), where they remained throughout the whole 153
experiment. Containers were provided with an opaque suspended predator cage (diameter 85 mm, 154
length 16 cm) with top and bottom covered with fine mesh netting (1.5 mm), allowing the tadpoles 155
to receive chemical, but no visual or tactile cues from the crayfish. Half of the cages received one 156
crayfish (predator treatment), whereas the other half were left empty (no-predator treatment). Each 157
population/predator treatment combination was replicated once in each of five randomized blocks. 158
Crayfish were fed inside the predator cages with three P. perezi tadpoles every other day, and 159
tadpoles were fed commercial fish food and boiled lettuce ad libitum every three days. Water in 160
the containers was changed once a week. Water temperature was kept at 21.0 ºC (± 0.1) and the 161
photoperiod was 12L:12D. 162
Response variables 163
We recorded tadpole activity level at early (day 10 of the experiment) and mid (day 60 of the 164
experiment) larval development. Larval developmental stage of tadpoles at day 10 was considered 165
identical for all treatment combinations, and it did not differ among populations (P= 0.75) or 166
predation treatments (P= 0.66) at day 60 (Gosner range 33-36). The number of active tadpoles in 167
each tank, during a 10-s observation interval, was recorded five times between 9 and 12 a.m., the 168

NUNES ET AL.
9
observations being separated by at least 30 minutes. Tadpoles were considered active if they were 169
swimming, foraging or moving their tails. 170
We used geometric morphometrics tools to examine variation in tadpole morphology (Rohlf and 171
Marcus 1993, Zelditch et al. 2004). Morphology was recorded at day 60 of the experiment in five 172
haphazardly-selected individuals per container, by placing each tadpole in a small Plexiglas box 173
with a grid background and taking a side-view digital photograph. To minimize the effect of non-174
parallel tail positions, all the images were straightened with the straighten plugin in ImageJ 175
(http://rsb.info.nih.gov/ij, see Johansson et al. 2010 for details) and then loaded into MakeFan7 176
(Sheets 2009) to create a standardized template for digitizing the landmarks. For each tadpole, 19 177
landmarks representing an adequate coverage of the tadpole body shape were marked using the 178
image analysis software tpsDig2 (Rohlf 2008; Appendix 1; see e.g. Orizaola et al. 2012, 2013, for 179
a similar approach). In order to extract tadpole shape variables, landmark coordinates were 180
imported into tpsRelw (Rohlf 2007) and a Generalized least squares Procrustes Analysis was 181
performed in order to standardize the size, translate and rotate the configurations of landmark 182
coordinates (Rohlf and Slice 1990). TpsRelw allowed us to generate a mean (consensus) landmark 183
configuration for each experimental container, described by several components (relative warps), 184
which were later used as shape variables in the statistical analysis. The two first relative warps 185
(RW1 and RW2) explained most of the initial morphological variance (see Results) and were used 186
as dependent variables in the analyses. 187
Statistical analysis 188
We examined variation in antipredator responses for activity rates using a Generalized Linear 189
Mixed Model (GLMM) with binomial error distribution and a logit link function. We used time 190
period and predator treatment as fixed factors, time of coexistence with crayfish as a continuous 191

NUNES ET AL.
10
fixed factor and population as a random factor. We tested for all the possible interactions between 192
terms, but the 3-way interaction between time period, predator treatment and coexistence time was 193
removed from the final model because it was not significant (F1,89= 1.185, P= 0.279). Tank was 194
used as an additional random effect in order to account for potential overdispersion in the data. 195
The proportion of active tadpoles per container, estimated as the number of tadpoles active divided 196
by their total number in a container, was used as the dependent variable. This value was the 197
average of the five measurements recorded per container in each time period. LSD pairwise 198
comparisons were performed to inspect differences among populations. 199
We examined the effect of predator treatment on tadpole morphology across the different 200
populations using general linear model (GLM) analyses followed by Tukey HSD post-hoc tests. 201
Predator treatment was included as a fixed factor, time of coexistence as a fixed continuous 202
variable, and population origin as a random effect to account for the replication of related 203
genotypes. Since we found a nearly significant population × treatment interaction (see Results), 204
GLMs were performed for each population separately. Experimental block was initially used as a 205
random factor, but since this factor was never significant (P > 0.1) it was removed from the final 206
models. REML was used as estimation procedure. All analyses were performed using the software 207
package SPSS (version 21.0). 208
209
Results 210
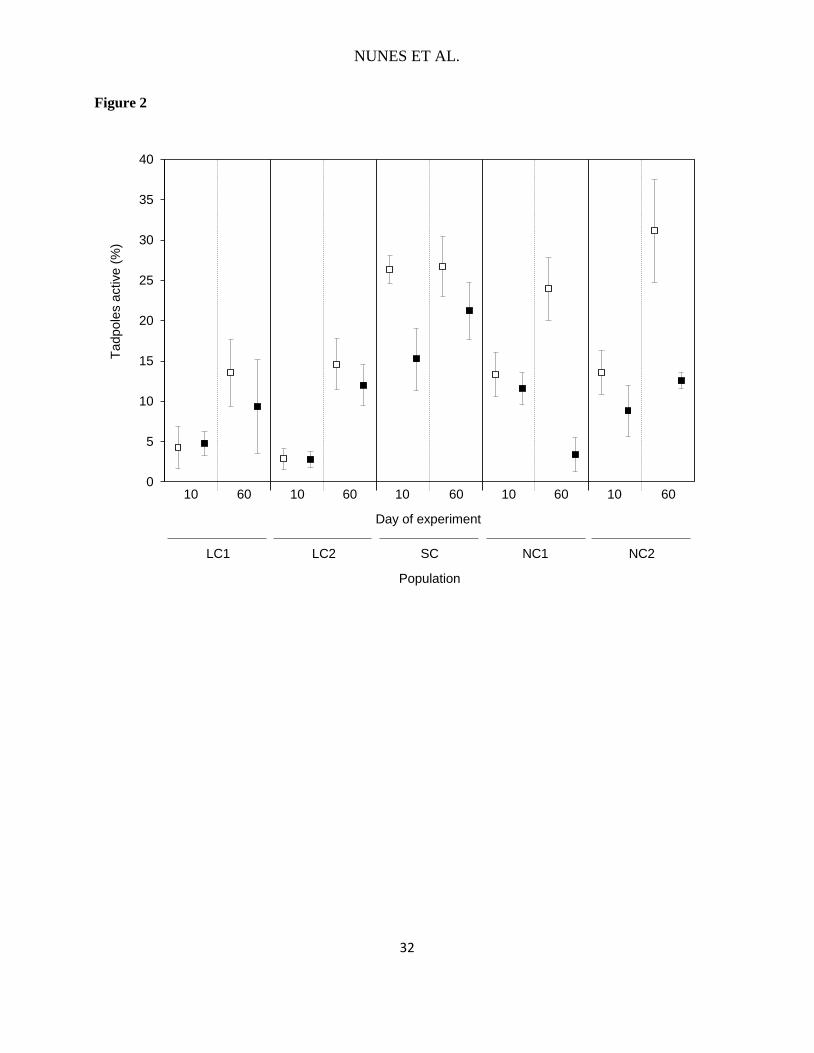
A significant interaction between time period and predator treatment (F1,90= 13.3, P= 0.020) 211
indicated higher activity in the absence of crayfish as larval development proceeded (day 60, Fig. 212
2). We also found a significant interaction between time period and coexistence time with crayfish 213
(F1,90= 10.7, P< 0.001) reflecting that, in general, activity differs among populations with different 214

NUNES ET AL.
11
coexistence times as larval development advances, with activity increasing with lower coexistence 215
time with crayfish in day 60 of the experiment (Fig. 2). The evolutionary history of crayfish 216
contact affected the tadpoles' behavioral response to the presence of a fed crayfish (coexistence 217
time × predator treatment interaction: F1,90= 34.8, P< 0.001; Fig. 2). Overall, tadpoles from 218
populations with longer coexistence time with crayfish (LC1 and LC2) had very low activity 219
levels, and did not modify their activity in response to crayfish presence (pairwise comparisons, 220
P< 0.05; Fig. 2). When exposed to fed crayfish, tadpoles from populations without crayfish (NC1 221
and NC2) and populations coexisting for a short time with crayfish (SC) showed, overall, lower 222
activity rates than control tadpoles from the same populations (pairwise comparisons, P< 0.05; 223
Fig. 2). When looking at each population separately, only tadpoles from SC population had lower 224
activity when in the presence of fed crayfish early in larval development (day 10 of the 225
experiment, P= 0.003). As larval development advanced (day 60 of the experiment), tadpoles from 226
the two NC populations exposed to crayfish became less active than those from the respective 227
control treatments (P< 0.001), whereas activity of tadpoles from LC and SC populations was not 228
affected by treatment (Fig. 2). 229
Morphometric analyses on tadpole morphology yielded 34 relative warps, of which the first two 230
(RW1 and RW2) explained 54.78% of the total shape variation. RW1 accounted mainly for 231
constitutive differences between populations (P= 0.09), with populations coexisting with crayfish 232
(both LC and SC) having a more bulky body, with longer and deeper headbodies, expanded 233
particularly around the gut area (negative values of RW1; post-hoc tests, P< 0.05; Fig. 3a). For 234
RW1, no significant differences between predator treatments or a population × predator treatment 235
interaction were found (P> 0.523; Fig. 3a). For the second relative warp (RW2), there was a non-236
significant tendency towards a population × predator interaction (F4,36 = 2.1, P= 0.096). Additional 237

NUNES ET AL.
12
analyses showed that, in the presence of fed crayfish, tadpoles from population LC1 expanded 238
their tail region, developing deeper tail muscles and tail fins (F1,6 = 10.5, P= 0.018; Fig. 3b, 239
positive values of RW2), indicating morphological plasticity in response to crayfish in this 240
population. No evidence for alterations in morphology in response to crayfish presence was found 241
in any of the other populations (P≥ 0.1; Fig. 3b). 242
243
Discussion 244
We found that P. perezi populations differing in coexistence time with P. clarkii showed variation 245
in their antipredator responses to this exotic crayfish. Specifically, populations with a long 246
coexistence with P. clarkii (LC) exhibited a constitutive behavioral defense (maintenance of very 247
low activity levels even in the absence of predators) and LC1 population showed adaptive 248
morphological plasticity in the presence of fed crayfish. On the contrary, populations with shorter 249
(SC) or no (NC) coexistence time with crayfish showed plastic antipredator behavior when 250
exposed to the fed crayfish and lacked plastic morphological responses. These results highlight 251
that i) invasive predators can promote rapid evolution of adaptive antipredator responses in native 252
prey, ii) the responses in individual traits can be either constitutive or plastic, and iii) even naive 253
populations can respond adaptively to exotic predators. The considerable variation detected among 254
the P. perezi populations reflects the complexity of antipredator responses to P. clarkii, and 255
perhaps to invasive predators in general, and may partly explain why previous studies using either 256
a single population or two geographically close populations of P. perezi have failed to detect 257
antipredator responses in this predator-prey system (Gómez-Mestre and Díaz-Paniagua 2011, 258
Nunes 2012, Nunes et al. 2013). 259

NUNES ET AL.
13
Contrary to our predictions, populations of P. perezi with longer coexistence history with P. 260
clarkii did not alter their activity level when exposed to fed crayfish. Instead, tadpoles in these 261
populations maintained very low activity levels irrespective of predator treatment, suggesting the 262
evolution of a constitutive (canalized) behavioral defense. In general, constitutive defenses are 263
expected to evolve under strong uniform and prolonged predation risk (Moran 1992, Edgell et al. 264
2009). In our case, strong directional selection by P. clarkii towards low activity appears to have 265
promoted the loss of behavioral plasticity in LC populations, leading to permanently defended, 266
inactive tadpoles. Similar results were reported by Edgell et al. (2009), who showed that snails 267
from the native area of a crab predator show constitutive behavioral defenses against the predator, 268
whereas conspecific snails from areas recently invaded by this predator show behavioral plasticity. 269
A longer coexistence time or a more intense selection pressure may be required for this 270
constitutive response to evolve in SC populations. It is important to mention, though, that the 271
amphibian eggs were collected in the field shortly after laying and the early embryonic stages in 272
LC and SC populations may have been exposed to crayfish cues for a short time. While embryonic 273
learning of predators has been demonstrated in amphibians (Ferrari et al. 2010a), we find it 274
unlikely that this short early exposure would have affected antipredator responses during the larval 275
stage. Transgenerational effects, with mothers pre-conditioning larval strategies as a response to 276
predator presence (e.g. McGhee et al. 2012), may be another way of responding to invasive 277
predators, although this option remains to be explored. 278
Interestingly, our results suggest that behavioral plasticity may be the ancestral state of 279
antipredator responses of P. perezi to P. clarkii cues. NC populations reduced their activity levels 280
in the presence of fed crayfish at day 60, indicating that P. perezi tadpoles can elicit plastic 281
behavioral defenses towards a predator with which they have had no previous contact. Indeed, 282

NUNES ET AL.
14
reduced activity is a common adaptive defense, which reduces prey detectability and encounter 283
rates with predators (Lima and Dill 1990, Kats and Dill 1998). High activity levels in presence of 284
predators have been linked with lower survival in many tadpole species (e.g. Lawler 1989, Skelly 285
1994), including in P. perezi (Gómez-Mestre and Díaz-Paniagua 2011).While innate recognition 286
of chemical stimuli from native predators is common (reviewed in Wisenden 2003), there are only 287
few such examples involving exotic predators (e.g. Epp and Gabor 2008, Rehage et al. 2009, 288
Bourdeau et al. 2013). Naive populations responding to unknown predators often share an 289
evolutionary history with native predators closely related to the novel predator (Epp and Gabor 290
2008, Rehage et al. 2009). The original geographical range of the white-clawed crayfish 291
Austropotamobius pallipes, the only native crayfish species in Portugal, did not reach Portugal 292
south of the river Tejo basin (Almaça 1991), suggesting that the responses of NC populations are 293
not likely due to phylogenetic similarity to a native predator in the community (Strauss et al. 294
2006). Instead, responses may have been facilitated by the detection of cues from predation events, 295
resulting from injured or digested conspecifics (Ferrari et al. 2010b) or by an innate generalized 296
antipredator response towards predator signals (Gherardi et al. 2011). Native prey can also 297
increase their chances of survival when exposed to new predators by using phenotypically plastic 298
neophobia as an adaptive antipredator strategy (Brown et al. 2013). 299
We also found inducible morphological defenses in response to fed crayfish in one of the LC 300
populations. When exposed to fed crayfish, LC1 tadpoles developed deeper tail fins and muscles, 301
which are common adaptive morphological defenses of larval anurans to predators (e.g. McCollum 302
and Van Buskirk 1996, Relyea 2001, Van Buskirk 2009). Recently, Gómez-Mestre and Díaz-303
Paniagua (2011) showed that similar morphological alterations (deeper tails) - induced by 304
dragonflies - are efficient in decreasing P. perezi mortality imposed by P. clarkii. Predation trials 305

NUNES ET AL.
15
conducted with other anuran species have shown that the tadpoles surviving predation have deeper 306
tails than those sampled before the trials (e.g. Van Buskirk et al. 1997, Van Buskirk and Relyea 307
1998, Teplitsky et al. 2005, Álvarez and Nicieza 2006). These results strongly suggest that the 308
morphological response to P. clarkii in LC1 population is adaptive, indicating that this population 309
has evolved plastic morphological defenses against the invasive crayfish. Previous studies have 310
found high heritability of antipredator responses in tadpoles (Relyea 2005), in particular for 311
activity and tail depth induced by the presence of predators, suggesting that rapid evolutionary 312
response is possible when selection pressure is high. Taken together, our results agree with the 313
idea that invasive species can be a strong selective force generating local adaptation and rapid 314
evolution (Strauss et al. 2006, Sih et al. 2010). While our study reports the fastest evolution of 315
antipredator responses documented for an amphibian (ca. 30 years, 10-15 P. perezi generations), 316
there are several studies reporting rapid evolution in response to selection by invasive predators 317
(Schlaepfer et al. 2005, Freeman and Byers 2006, Strauss et al. 2006, Bourdeau et al. 2013), 318
including in an amphibian (Kiesecker and Blaustein 1997). 319
Our results indicate that LC1 population has developed both constitutive and inducible defenses 320
against P. clarkii. This suggests rapid evolution of a functional integration of constitutive and 321
inducible defenses, promoting a highly effective defensive phenotype, improving adaptation to 322
changing local predation risk and enhancing prey survival (DeWitt et al. 1999, Bourdeau 2012). 323
Theory predicts that high constitutive defenses and moderate/low inducibility - which seems to be 324
the case here - evolve when the frequency of predator encounter rates is relatively high (Poitrineau 325
et al. 2004). Population LC1 is located in a rice field close to the Guadiana River, the river through 326
which P. clarkii likely expanded into Portugal (Ramos and Pereira 1981). Rice fields are one of 327
the preferred habitats of the crayfish in the Iberian Peninsula, where they can attain very high 328

NUNES ET AL.
16
densities (Anastácio et al. 1995). This suggests that population LC1 may have coexisted for a 329
slightly longer time with more dense populations of P. clarkii than, for example, population LC2. 330
In a previous study, Gómez-Mestre and Díaz-Paniagua (2011) did not find morphological 331
responses of P. perezi to P. clarkii in a population from Doñana National Park, where P. clarkii 332
was first found in 1974. While we cannot exclude population differences in intensity of selection 333
or in P. perezi’s potential to evolve antipredator defenses against P. clarkii, the discrepancy 334
between those and the present results could potentially be also explained by the different approach 335
used to examine tadpole morphology in the present study (i.e. geometric morphometric 336
techniques). In any case, studies using multiple populations from a wider geographical area could 337
shed additional light into the role of rapid evolution of antipredator defenses in populations 338
invaded by P. clarkii. 339
We found significant phenotypic divergence in larval morphology among LC/SC and NC 340
populations, the former having deeper headbodies (expanded especially around the gut area) than 341
the latter. While the functional significance of the deeper headbody is unclear at the moment, this 342
result may suggest the existence of a constitutive morphological adaptation in populations 343
coexisting with crayfish, expressed in the form of a larger gut area. Larger guts may allow for a 344
more efficient digestion and rapid growth (Relyea and Auld 2004, Lindgren and Laurila 2005; but 345
see Steiner 2007), which can reduce the time tadpoles are exposed to predators when searching for 346
food, likely reducing the risk of being captured. Actually, a separate experiment showed that P. 347
perezi tadpoles’ tended to grow faster in the presence of fed crayfish than in non-predator 348
environments (Nunes 2012). While this hypothesis agrees well with the generally low activity 349
levels found in LC populations, activity was considerably higher in population SC, suggesting that 350
behavior and gut area were uncoupled in these populations. Alternatively, wider body dimensions 351

NUNES ET AL.
17
may provide protection from gape-limited predators (Kishida and Nishimura 2004; Urban 2007). 352
Although crayfish are not strictly gape-limited predators (Nilsson et al. 2000), they are probably 353
less able to capture and handle prey above a certain size (Renai and Gherardi 2004). Finally, while 354
the populations chosen for this study have similar habitats and experience similar native predator 355
regimes (Ribeiro et al. 2007), we cannot fully exclude the possibility that these differences may 356
result from adaptation to other local habitat features such as competitors or food availability (e.g. 357
Relyea 2002, Van Buskirk and Arioli 2005, Richter-Boix et al. 2010), or from environmental 358
maternal effects (Räsänen and Kruuk 2007). 359
Taking into account the enormous invasion potential of P. clarkii (Capinha and Anastácio 2011), 360
this species is likely to colonize many crayfish-free regions in the future. Given the strong 361
selection pressure imposed by this predator (Cruz and Rebelo 2005, Cruz et al. 2008, Nunes et al. 362
2010, Ficetola et al. 2011), some populations or species may not be able to evolve defensive 363
adaptations rapidly enough to hinder strong declines or even extinctions. Overall, our results 364
suggest that invasive predators may drive rapid evolutionary change in invaded communities. 365
Consequently, examining possible evolutionary responses of prey to invasive predators in invaded 366
communities is crucial for understanding the impact of these on natural populations. However, the 367
alterations in prey traits can be complex, as prey can evolve both constitutive and plastic responses 368
to invasive predators. Considering the variability of the responses and the current rates of spread of 369
exotic predators, our study reinforces the need of multi-population approaches, covering a range of 370
coexistence times, in order to better forecast the long-term impact of biological invasions. 371
372
Acknowledgments 373

NUNES ET AL.
18
We thank Bruno Carreira, Rosário Sumares, João Nunes and Sónia Longueira for their help in the 374
laboratory. We also thank Rui Braz for designing the map of P. perezi populations. Mark Urban 375
and two anonymous reviewers made valuable comments on a previous version of the manuscript. 376
Ángel Ruiz Elizalde kindly provided the P. perezi photograph. The study was conducted in 377
accordance with Portuguese legislation; the collection permit was provided by the Portuguese 378
Instituto da Conservação da Natureza e da Biodiversidade (Licence n.º228/2009/CAPT/TRANS). 379
This research was funded by the FCT Project POCI/BIA-BDE/56100/2004 (to RR), FCT grant 380
SFRH/BD/29068/2006 (ALN) and by Stiftelsen för Zoologisk Forskning (ALN). 381
382
Literature Cited 383
Almaça, C. 1991. L’ecrevisse a pieds blancs, Astacus pallipes Lereboullet 1858, au Portugal. 384
L’Astaciculteur de France 28:11-16. 385
Almeida, N. F., P. Almeida, H, Gonçalves, F. Sequeira, J. Teixeira, and F. Almeida. 2001. Guias 386
FAPAS: Anfíbios e Répteis de Portugal. FAPAS and Câmara Municipal do Porto, Porto, 387
Portugal. 388
Álvarez, D., and A.G. Nicieza. 2006. Factors determining tadpole vulnerability to predators: can 389
prior experience compensate for a suboptimal shape? Evolutionary Ecology 20:523-534. 390
Anastácio, P. M., S. N. Nielsen, J. C. Marques, and S. E. Jorgensen. 1995. Integrated production of 391
crayfish and rice: a management model. Ecological Engineering 4:199-210. 392
Benard, M. F. 2004. Predator-induced phenotypic plasticity in organisms with complex life cycles. 393
Annual Review of Ecology, Evolution and Systematics 35:651-673. 394

NUNES ET AL.
19
Bourdeau, P. E. 2012. Intraspecific trait cospecialization of constitutive and inducible 395
morphological defences in a marine snail from habitats with different predation risk. Journal 396
of Animal Ecology 81:849-858. 397
Bourdeau, P. E., K. L. Pangle, E. M. Reed, and S. D. Peacor. 2013. Finely-tuned response of 398
native prey to an invasive predator in a freshwater system. Ecology 94:1449-1455. 399
Brown, G. E., M. C. O. Ferrari, C. K. Elvidge, I. Ramnarine, and D. P. Chivers. 2013. 400
Phenotypically plastic neophobia: a response to variable predation risk. Proceedings of the 401
Royal Society of London B 280:2012-2712. 402
Capinha, C., and P. Anastácio. 2011. Assessing the environmental requirements of invaders using 403
ensembles of distribution models. Diversity and Distributions 17:13-24. 404
Clark, C. W., and C. D. Harvell. 1992. Inducible defences and the allocation of resources: a 405
minimalist model. The American Naturalist 139:521-539. 406
Cox, J. G., and S. L. Lima. 2006. Naiveté and an aquatic-terrestrial dichotomy in the effects of 407
introduced predators. Trends in Ecology and Evolution 21:674-680. 408
Cruz, M. J., and R. Rebelo. 2005. Vulnerability of southwest Iberian amphibians to an introduced 409
crayfish, Procambarus clarkii. Amphibia-Reptilia 26:293-303. 410
Cruz, M. J., and R. Rebelo. 2007. Colonization of freshwater habitats by an introduced crayfish, 411
Procambarus clarkii, in Southwest Iberian Peninsula. Hydrobiologia 575:191-201. 412
Cruz, M. J., P. Segurado, M. Sousa, and R. Rebelo. 2008. Collapse of the amphibian community 413
of the Paul do Boquilobo Natural Reserve (central Portugal) after the arrival of the exotic 414
American crayfish Procambarus clarkii. Herpetological Journal 18:197-204. 415
DeWitt, T. J., A. Sih, and J. A. Hucko. 1999. Trait compensation and cospecialization in a 416
freshwater snail: size, shape and antipredator behaviour. Animal Behaviour 58:397-407. 417

NUNES ET AL.
20
Edgell, T. C., B. R. Lynch, G. C. Trussell, and A. R. Palmer. 2009 Experimental evidence for the 418
rapid evolution of behavioral canalization in natural populations. American Naturalist 419
174:434-440. 420
Epp, K. J., and C. R. Gabor. 2008. Innate and learned predator recognition mediated by chemical 421
signals in Eurycea nana. Ethology 114:607-615. 422
Ferrari, M. C. O., A. K. Manek, and D. P. Chivers. 2010a. Temporal learning of predation risk by 423
embryonic amphibians. Biology Letters 6:308-310. 424
Ferrari, M. C. O., B. D. Wisenden, and D. P. Chivers. 2010b. Chemical ecology of predator-prey 425
interactions in aquatic ecosystems: a review and prospectus. Canadian Journal of Zoology 426
88:698-724. 427
Ficetola, F. G., M. E. Siesa, R. Manenti, L. Bottoni, F. De Bernardi, and E. Padoa-Schioppa. 2011. 428
Early assessment of the impact of alien species: differential consequences of an invasive 429
crayfish on adult and larval amphibians. Diversity and Distributions 17:1141-1151. 430
Fisk, D. F., L. C. Latta, R. A. Knapp, and M. E. Pfrender. 2007. Rapid evolution in response to 431
introduced predators I: rates and patterns of morphological and life-history trait divergence. 432
BMC Evolutionary Biology 7:22. 433
Freeman, A.S., and J. E. Byers. 2006. Divergent induced responses to an invasive predator in 434
marine mussel populations. Science 313:831-833. 435
Geiger, W., P. Alcorlo, A. Baltanás, and C. Montes. 2005. Impact of an introduced crustacean on 436
the trophic webs of Mediterranean wetlands. Biological Invasions 7:49-73. 437
Gherardi, F. 2006. Crayfish invading Europe: the case study of Procambarus clarkii. Marine and 438
Freshwater Behaviour and Physiology 39:175-191. 439

NUNES ET AL.
21
Gherardi, F., K. M. Mavuti, N. Pacini, E. Tricarico, and D. M. Harper. 2011. The smell of danger: 440
chemical recognition of fish predators by the invasive crayfish Procambarus clarkii. 441
Freshwater Biology 56:1567-1578. 442
Gómez-Mestre, I., and C. Díaz-Paniagua. 2011. Invasive predatory crayfish do not trigger 443
inducible defences in tadpoles. Proceedings of the Royal Society of London B 278:3364-444
3370. 445
Gosner, K. L. 1960. A simplified table for staging anuran embryos and larvae with notes on 446
identification. Herpetologica 16:183-190. 447
Holdich, D. M., J. D. Reynolds, C. Souty-Grosset, and P. J. Sibley. 2009. A review of the ever 448
increasing threat to European crayfish from non-indigenous crayfish species. Knowledge and 449
Management of Aquatic Ecosystems 394-395:11. 450
Johansson, F., B. Lederer, and M. Lind. 2010. Trait performance correlations across life stages 451
under environmental stress conditions in the common frog, Rana temporaria. PLoS One 452
5:e11680. 453
Kats, L. B., and L. M. Dill. 1998. The scent of death: chemosensory assessment of predation risk 454
by prey animals. Ecoscience 5:361-394. 455
Kerby, J. L., S. P. D. Riley, L. B. Kats, and P. Wilson. 2005. Barriers and flow as limiting factors 456
in the spread of an invasive crayfish (Procambarus clarkii) in southern California streams. 457
Biological Conservation 126:402-409. 458
Kiesecker, D. G., and A. R. Blaustein. 1997. Population differences in responses of red-legged 459
frogs (Rana aurora) to introduced bullfrogs (Rana catesbeiana). Ecology 78:1752-1760. 460
Kishida, O., and K. Nishimura. 2004. Bulgy tadpoles: inducible defense morph. Oecologia 461
140:414-421. 462

NUNES ET AL.
22
Lawler, S. P. 1989. Behavioural responses to predators and predation risk in four species of larval 463
anurans. Animal Behaviour 38:1039-1047. 464
Lima, S. L., and L. M. Dill. 1990. Behavioral decisions made under the risk of predation: A review 465
and prospectus. Canadian Journal of Zoology 68:619-640. 466
Lindgren, B., and A. Laurila. 2005. Proximate causes of adaptive growth rates: growth efficiency 467
variation among latitudinal populations of Rana temporaria. Journal of Evolutionary 468
Biology 18:820-828. 469
Losos, J. B., T. W. Schoener, and D. A. Spiller. 2004. Predator-induced behaviour shifts and 470
natural selection in field-experimental lizard populations. Nature 432:505-508. 471
Loureiro, A., N. Ferrand de Almeida, M. A. Carretero, and O. S. Paulo (eds.) 2008. Atlas dos 472
Anfíbios e Répteis de Portugal. Instituto da Conservação da Natureza e da Biodiversidade, 473
Lisboa, Portugal. 474
Magurran, A. E. 1990. The adaptive significance of schooling as an anti-predator defence in fish. 475
Annales Zoologici Fennici 27:3-18. 476
McCollum, S. A., and J. Van Buskirk. 1996. Costs and benefits of a predator-induced polyphenism 477
in the gray treefrog Hyla chrysoscelis. Evolution 50:583-593. 478
McGhee, K. E., L. M. Pintor , E. L.Suhr, and A. M. Bell. 2008. Maternal exposure to predation 479
risk decreases offspring antipredator behavior and survival in threespined stickleback. 480
Functional Ecology 26:932-940. 481
Moran, N. A. 1992. The evolutionary maintenance of alternative phenotypes. American Naturalist 482
139:971-989. 483

NUNES ET AL.
23
Nilsson, P. A., K. Nilsson, and P. Nyström. 2000. Does risk of intraspecific interactions induce 484
shifts in prey-size preference in aquatic predators? Behavioral Ecology and Sociobiology 485
48:268-275. 486
Nunes, A. L. 2012. Predator-prey interactions between an invasive crayfish predator and native 487
anurans: ecological and evolutionary outcomes. PhD thesis, University of Lisbon. 488
Nunes, A. L., M. J. Cruz, M. Tejedo, A. Laurila, and R. Rebelo. 2010. Nonlethal injury caused by 489
an invasive alien predator and its consequences for an anuran tadpole. Basic and Applied 490
Ecology 11:645-654. 491
Nunes, A. L., A. Richter-Boix, A. Laurila, and R. Rebelo. 2013. Do anuran larvae respond 492
behaviourally to chemical cues from an invasive crayfish predator? A community-wide 493
study. Oecologia 171:115-127. 494
Orizaola, G., E. Dahl, and A. Laurila. 2012. Reversibility of predator-induced plasticity and its 495
effect at a life-history switch point. Oikos 121:44-52. 496
Orizaola, G., E. Dahl, A. G. Nicieza, and A. Laurila. 2013. Larval life history and anti-predator 497
strategies are affected by breeding phenology in an amphibian. Oecologia 171:873-881. 498
O'Steen, S., A. J. Cullum, and A. F. Bennet. 2002. Rapid evolution of escape ability in Trinidadian 499
guppies (Poecilia reticulata). Evolution 56:776-784. 500
Padilla, D. K., and S. C. Adolph. 1996. Plastic inducible morphologies are not always adaptive: 501
The importance of time delays in a stochastic environment. Evolutionary Ecology 10:105-502
117. 503
Peluc, S. I., T. S. Sillet, J. T. Rotenberry, and C. K. Ghalambor. 2008. Adaptive phenotypic 504
plasticity in an island songbird exposed to a novel predation risk. Behavioral Ecology 505
19:830-835. 506

NUNES ET AL.
24
Poitrineau, K., S. P. Brown, and M. E. Hochberg. 2004. The joint evolution of defence and 507
inducibility against natural enemies. Journal of Theoretical Biology 231:389-396. 508
Polo-Cavia, N., A. Gonzalo, P. Lopez, and J. Martin. 2010. Predator recognition of native but not 509
invasive turtle predators by naive anuran tadpoles. Animal Behaviour 80:461-466. 510
Ramos, M. A., and T. M. Pereira. 1981. Um novo Astacidae para a fauna portuguesa: 511
Procambarus clarkii (Girard, 1852). Boletim do Instituto Nacional de Investigação das 512
Pescas, Lisboa 6:37-47. 513
Räsänen, K., and L. E. B. Kruuk. 2007. Maternal effects and evolution at ecological time scales. 514
Functional Ecology 21:408-421. 515
Rehage, J. S., K. L. Dunlop, and W. F. Loftu. 2009. Antipredator responses by native mosquitofish 516
to non-native cichlids: an examination of the role of prey naiveté. Ethology 115:1-11. 517
Relyea, R. A. 2001. Morphological and behavioral plasticity of larval anurans in response to 518
different predators. Ecology 82:523-540. 519
Relyea, R. A. 2002. Local population differences in phenotypic plasticity: Predator-induced 520
changes in wood frog tadpoles. Ecological Monographs 72:77-93. 521
Relyea, R. A. 2005. The heritability of inducible defenses in tadpoles. Journal of Evolutionary 522
Biology 18:856-866. 523
Relyea, R. A., and J. R. Auld. 2004. Having the guts to compete: how intestinal plasticity explains 524
costs of inducible defenses. Ecology Letters 7:869-875. 525
Renai, B., and F. Gherardi. 2004. Predatory efficiency of crayfish: comparison between indigenous 526
and non-indigenous species. Biological Invasions 6:89-99. 527
Ribeiro, F., R. Beldade, M. Dix, and J. Bochechas. 2007. Carta Piscícola Nacional Direcção Geral 528
dos Recursos Florestais-Fluviatilis, Lda. Publicação Electrónica (versão 01/2007). 529

NUNES ET AL.
25
Richter-Boix, A., G. A. Llorente, and A. Montori. 2007. A comparative study of predator-induced 530
phenotype in tadpoles across a pond permanency gradient. Hydrobiologia 583:43-56. 531
Richter-Boix, A., C. Teplitsky, B. Rogell, and A. Laurila. 2010. Local selection modifies 532
phenotypic divergence among Rana temporaria populations in the presence of gene flow. 533
Molecular Ecology 19:716-731. 534
Rohlf, F. J. 2007. tpsRelw. Department of Ecology and Evolution. State Univ. New York, Stony 535
Brook. New York, USA. 536
Rohlf, F. J. 2008. tpsDig2. Department of Ecology and Evolution. State Univ. New York, Stony 537
Brook. New York, USA. 538
Rohlf, F. J., and L. F. Marcus. 1993. A revolution in morphometrics. Trends in Ecology and 539
Evolution 8:129-132. 540
Rohlf, F. J., and D. E. Slice. 1990. Extensions of the Procrustes method for the optimal 541
superimposition of landmarks. Systematics Zoology 39:40-59. 542
Salo, P., E. Korpimäki, P. B. Banks, M. Nordström, and C. R. Dickman. 2007. Alien predators are 543
more dangerous than native predators to prey populations. Proceedings of the Royal Society 544
B 274:1237-1243. 545
Schlaepfer, M. A., P. W. Sherman, B. Blossey, and M. C. Runge. 2005. Introduced species as 546
evolutionary traps. Ecology Letters 8:241-246. 547
Sheets, H. D. 2009. MakeFan7. Department of Physics, Canisius College, Buffalo, USA. 548
Sih, A., D. I. Bolnick, B. Luttbeg, J. L. Orrock, S. D. Peacor, L. M. Pintor, E. Preisser, J. S. 549
Rehage, and J. R. Vonesh. 2010. Predator-prey naiveté, antipredator behavior, and the 550
ecology of predator invasions. Oikos 119:610-621. 551

NUNES ET AL.
26
Skelly, D. K. 1994. Activity level and the susceptibility of anuran larvae to predation. Animal 552
Behaviour 47:465-468. 553
Steiner, U. K. 2007. Linking antipredator behaviour, ingestion, gut evacuation and costs of 554
predator-induced responses in tadpoles. Animal Behaviour 74:1473-1479. 555
Strauss, S. Y., J. A. Lau, and S. P. Carroll. 2006. Evolutionary responses of natives to introduced 556
species: what do introductions tell us about natural communities? Ecology Letters 9:354-557
371. 558
Sündermann, D., M. Scheumann, and E. Zimmermann. 2008. Olfactory predator recognition in 559
predator-naive gray mouse lemurs (Microcebus murinus). Journal of Comparative 560
Psychology 122:146-155. 561
Teplitsky, C., S. Plenet, J. - P. Léna, N. Mermet, E. Malet, and P. Joly. 2005. Escape behaviour 562
and ultimate causes of specific induced defences in an anuran tadpole. Journal of 563
Evolutionary Biology 18:180-190. 564
Thompson, J. N. 1998. Rapid evolution as an ecological process. Trends in Ecology and Evolution 565
13:329-332. 566
Tollrian, R., and C. D. Harvell (eds.). 1999. The ecology and evolution of inducible defenses. 567
Princeton University Press, Princeton, USA. 568
Trussell, G. C., and L. D. Smith. 2000. Induced defenses in response to an invading crab predator: 569
An explanation of historical and geographic phenotypic change. Proceedings of the National 570
Academy of Sciences USA 97: 2123-2127. 571
Urban, M. C. 2007. The growth–predation risk trade-off under a growing gape-limited predation 572
threat. Ecology 88:2587-2597. 573

NUNES ET AL.
27
Van Buskirk, J. 2009. Natural variation in morphology of larval amphibians: Phenotypic plasticity 574
in nature? Ecological Monographs 79:681-705. 575
Van Buskirk, J., and R. A. Relyea. 1998. Selection for phenotypic plasticity in Rana sylvatica 576
tadpoles. Biological Journal of the Linnean Society 65:301-328. 577
Van Buskirk, J., and M. Arioli. 2005. Habitat specialization and adaptive phenotypic divergence of 578
anuran populations. Journal of Evolutionary Biology 18:596-608. 579
Wisenden, B. D. 2003. Chemically mediated strategies to counter predation. Pages 236-251 in: S. 580
P. Collin, and N. J. Marshall, editors. Sensory Processing in Aquatic Environments. 581
Springer-Verlag, New York, USA. 582
Zelditch, M. L., D. L. Swiderski, H. D. Sheets, and W. L. Fink. 2004. Geometric morphometrics 583
for biologists: a primer. Academic Press, San Diego, USA. 584

NUNES ET AL.
28
Table 1. Area, coordinates, elevation above sea level of collection sites, type of water body,
approximate coexistence time between Procambarus clarkii and Pelophylax perezi and dates of
collection and of the start of the experiment for each of the five studied populations of P. perezi
Population LC1 LC2 SC NC1 NC2
Area Caia Caia Melides Monchique Monchique
Type of water body
Rice field on
the bank of the
Caia river
Caia river
Melides
stream and
rice fields
Monchique
river
Affluent of
the Seixe
river
Coordinates 38º52’N
7º03’W
39º07’N
7º17’W
38º08’N
8º44’W
37º17’N
8º33’W
37º22’N
8º38’W
Elevation a.s.l 170 m 270 m 25 m 200 m 100 m
Coexistence time 29 years 29 years 19 years 0 years 0 years
Date of collection 10-June 16-June 08-July 18-June 18-June
Number of clutches 8 10 10 9 12
Start of experiment 02-July 02-July 17-August 04-August 05-August

NUNES ET AL.
29
Figure legends
FIG. 1. Map showing the location of the studied Pelophylax perezi populations (LC – Longer
Coexistence, SC – Shorter Coexistence and NC – No Coexistence populations). The black
triangle indicates the location where P. clarkii was first introduced in Spain and the white triangle
indicates the site where crayfish presence was first registered in Portugal. Grey areas indicate
regions >700 m a.s.l.
FIG. 2. Activity rate of Pelophylax perezi tadpoles (mean ± SE) of the five studied populations
raised either in the presence (black square) or absence (white square) of crayfish, measured on
days 10 and 60 of the experiment. LC - Longer Coexistence, SC - Shorter Coexistence and NC -
No Coexistence populations.
FIG. 3. Morphology of Pelophylax perezi tadpoles of the five studied populations raised either in
the presence (black square) or absence (white square) of crayfish, measured after 60 days of the
experiment. LC - Longer Coexistence, SC - Shorter Coexistence and NC - No Coexistence
populations. Values are mean relative warps (RW) scores (± SE) for the first two relative warps
(RW1 and RW2). The figures on the right (grid plots) are thin-plate spline visualizations obtained
from tpsRelw (Rohlf 2007) and illustrate shape variation along the axes, representing the
(positive and negative) extremes of the range of phenotypes found for each RW axis (indicated
by the black arrows).

NUNES ET AL.
30
PLATE 1. Iberian waterfrog (Pelophylax perezi) at a breeding pond. Photo credit: Ángel Ruiz
Elizalde.

NUNES ET AL.
31
Figure 1
NUNES ET AL.
32
Figure 2
0
5
10
15
20
25
30
35
40
Ta
dp
ole
s a
ctive
(%
)
Day of experiment
Population
LC1 LC2 SC NC1 NC2
10 10 10 10 1060 60 60 60 60

NUNES ET AL.
33
Figure 3
No predator Crayfish
Population
-0.04
-0.03
-0.02
-0.01
0.00
0.01
0.02
0.03
0.04
RW
1 =
39
.74
%
LC1 LC2 SC NC1 NC2
a)
-0.02
-0.01
0.00
0.01
0.02
0.03
RW
2 =
15.0
4%
b)
Population
LC1 LC2 SC NC1 NC2

NUNES ET AL.
34
PLATE 1.





























