Embed Size (px)
Citation preview

1
Novel insights into the role of IL-27 and IL-23 in human malignant and normal plasma cells
Nicola Giuliani1 and Irma Airoldi2
1Hematology and BMT Center, University of Parma, Parma, Italy
2AIRC Laboratory of Immunology and Tumors, Department of Experimental and Laboratory
Medicine, G. Gaslini Institute, Genova, Italy
Corresponding Author
Irma Airoldi, Ph.D.,
A.I.R.C. Laboratory of Immunology and Tumors
Department of Experimental and Laboratory Medicine
G. Gaslini Institute, 16148 Genova, Italy
Phone: +390105636342, Fax: +39010377982
E-mail: [email protected]
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

2
ABSTRACT
Multiple myeloma (MM) is a monoclonal post-germinal center tumor that has phenotypic features
of plasmablasts/plasma cells and usually localizes at multiple sites in the bone marrow.
Pathogenesis of MM is complex and dependent on the interactions between tumor cells and their
microenvironment. Different cytokines, chemokines and pro-angiogenic factors released in the
tumor microenvironment are known to promote MM cell growth. Here we report recent advances
on the role of two stricktly related immunomodulatory cytokines, i.e. IL-27 and IL-23, in human
normal and neoplastic plasma cells highlighting their ability to i) act directly against MM cells, ii)
influence MM microenvironment by targeting osteoclast and osteoblast cells and iii) modulate
normal plasma cell function. Finally, the therapeutic implication of these studies will be discussed.
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

3
BACKGROUND
Interleukin (IL)-27 and IL-23 are immunomodulatory cytokines belonging to the IL-12 superfamily
(1), composed of structurally and functionally related heterodimeric cytokines including IL-12, IL-
23, IL-27 and IL-35 (1). These cytokines are predominantly produced by antigen presenting cells in
response to microbial or host immune stimuli and are involved in the regulation of immune
responses against infections and tumor development (2). Recent pre-clinical studies demonstrated
the direct anti-tumor activities of IL-12 family cytokines in different human hematological
malignancies including pediatric acute leukemias and multiple myeloma, as well as in solid tumors
(3-7). Here, we reported and discuss recent advances on the role of IL-27 and IL-23 in multiple
myeloma (MM) cells and its microenvironment, as well as in human normal plasma cells.
Interleukin-27
IL-27 is composed of the EBI3 and p35 subunits and activates both signal transducer and activator
of transcription (STAT)1 and STAT3 through distinct IL-27 receptor (R) that consists of the unique
receptor subunit WSX-1 paired with the gp130 chain (8). In T lymphocytes, the cytokine induces
STAT1 and STAT3 activation thus resulting in enhancement of naïve cluster of differentiation
(CD)4+ T cell proliferation, promotion of the early T helper (Th)1 differentiation and suppression of
the differentiation of Th2 and Th17 cells. In addition IL-27 plays a key role in generating IL-10
producing regulatory T cells (1, 9).
In human tonsils, the different B cell subsets show variable expression of IL-27R (10). In this study
it has been demonstrated that naïve and memory B lymphocytes express constitutively both chains
of IL-27R whereas germinal center B cells exhibited only barely detectable levels of the WSX-1
chain. However, the WSX-1 was found to be strongly up-regulated during germinal center to
memory B cell transition (10). In terms of signal transduction, IL-27 induces in naïve B cells a
strong phosphorylation of both STAT1 and STAT3, whereas a moderate STAT1 and a low
activation of STAT3 were observed in memory B lymphocytes (10). In addition, human plasma
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

4
cells (PC) express constitutively WSX-1 and gp130 and IL-27 specifically induces STAT-1
phosphorylation while unaffecting STAT-3 or STAT-5 activation (5). Differences in the STAT
activation driven by IL-27 may reflect the different ability of human B cell subsets to respond to IL-
27. For example, IL-27 enhances proliferation in naïve and activated germinal center B cells (10),
induces surface expression of CD54, CD95 and CD86 in stimulated memory B cells (10) and exerts
chemotactic activity on PC (5).
Anti-tumor activity of IL-27 against myeloma cells
IL-27, similarly to IL-12, shows anti-tumor activity in different cancers through indirect
mechanisms such as induction of natural killer (NK) and cytotoxic T lymphocyte (CTL) response or
inhibition of angiogenesis primarily due to induction of CXCL10 and CXCL9 (3, 11-14). In
addition, it has been reported that IL-27 inhibits directly the growth of melanoma cell lines (7) and
B-acute lymphoblastic leukemia cells (12) expressing complete IL-27R. In multiple myeloma (MM)
it has been reported that both chains of IL-27 receptor are expressed in CD138+ MM cells from
patients (5, 11). Although IL-27 does not modulate MM cell proliferation and apoptosis, a strong
reduction of the angiogenic potential of MM cells was clearly documented (5). It is of note that BM
neo-vascularization contributes to the biology of several hematologic malignancies (15) and MM
neo-angiogenesis may be induced by angiogenic cytokines produced by tumor and
microenvironmental cells (16). Cytokines such as basic fibroblast growth factor (bFGF), vascular
endothelial growth factor (VEGF) and angiopoietins (ANGPT) play a leading role in new vessel
formation as well as in tumor angiogenesis (17-19). However, other known and unknown factors
may be also involved and are under investigation. We reported that IL-27 down-regulates in vitro of
a wide panel of pro-angiogenic genes including VEGF-A, -C and –D, angiopoietins, and matrix
metalloproteinases and up-regulates the anti-angiogenic chemokines CXCL9 and CXCL10 (Figure
1) (11). Accordingly, pre-clinical studies using highly immunodeficient SCID/NOD mice injected
with MM cells revealed that IL-27 inhibits the MM cell growth through inhibition of angiogenesis,
that caused ischemic necrosis in the tumors. In this model, the main pro-angiogenic molecules and
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

5
MM growth factors down-regulated by IL-27 were IL-6, VEGF-D and CCL2 that serve also as
autocrine and/or paracrine growth factors (20, 21). Thus, we can envisage that IL-27 may interrupt
or reduce the “growth loop” (Figure 1) in which cytokines and growth factors released by MM cells
induce paracrine stimulation of stromal cells that, in turn, secrete additional growth factors such as
IL-6 and VEGF that induce new vessel formation and support MM cell proliferation, progression
and migration. In addition, MM cells and BM stromal cells produce matrix metalloproteinases
(MMP) that degrade collagens and fibronectin, thus rendering MM cells capable of invading both
stroma and the subendothelial basement membrane (Figure 1) (22-25). The enhanced invasive
ability of MM cells along with their increased angiogenic capacity may explain the frequent
intramedullary and extramedullary dissemination observed in MM pathogenesis. Pre-clinical studies
showed that IL-27 down-regulates MMP9, thus suggesting that IL-27 may also reduced the MM
cells invasion (Figure 1) (11).
Interleukin-23
IL-23 is a heterodimeric cytokine composed of the promiscuous IL-12p40 and the exclusive p19
subunits(26). The IL-23 receptor (R) is composed of the IL-12Rβ1 chain and the unique IL-23R
chain, that associated with Jak2 and STAT3 (26). In T lymphocytes and leukemic T cells, IL-23
induces activation of STAT family members, i.e. STAT1, 3, 4 and STAT5 (26). IL-23 is produced
predominantly by myeloid dendritic cells (DC) activated by Toll like receptor-2, -4 and -8 ligands
and by type 1 macrophages (27, 28). This cytokine is now considered the master switch in several T
cell mediated inflammatory disorders whereas its anti-tumor activity of IL-23 remains still
controversial. In this context, it has been reported that pro-inflammatory cytokines including IL-
17A, IL-6, and IL-23 can impair CD8+ T lymphocytes mediated immune surveillance, and promote
de novo carcinogenesis and tumor neo-vascularization (29-31). In contrast, other groups have shown
that IL-23 exerts anti-tumor activity through stimulation of T and NK cells (32-37).
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

6
Actually any information is not available on IL-23R expression and function in normal human B
cells with the only exception for plasma cells and early B lymphocytes (4, 5). In this context, it has
been demonstrated that human plasma cells express complete IL-23 receptors and that IL-23 may
modulate normal plasma cell functions (5). However the signal transduction activated by IL-23
treatment has never been investigated in these cells. On the contrary, it has been reported that in
human normal early B cells IL-23 induces a specific activation of the STAT3 molecule through
phosphorylation of serine727 and tyrosine 701 residues, while unaffecting the activation of STAT1,
4, 5 and 6 (4).
Anti-tumor activity of IL-23 against myeloma and B-acute lymphoblastic leukemia (ALL).
Although complete IL-23 receptor is expressed in MM cells ((3) and Airoldi I. personal
communication), IL-23 is ineffective in terms of modulation of MM cell proliferation and
angiogenesis, induction of apoptosis and chemotaxis, regulation of chemokine receptor expression
and cytokine production. The mechanisms underlying the lack of function of IL-23R in these cells
remain to be established.
A different scenario has been envisaged using IL-23 as potential anti-tumor agent against (4)
pediatric B-acute lymphoblastic leukemia (ALL). In this context, it has been demonstrated that IL-
23 dampens B-ALL cell growth in vitro and in pre-clinical models by inhibiting proliferation and
inducing apoptosis. Such effects were related to IL-23 induced up-regulation of miR-15a in
leukemic cells and to the consequent down-regulation of the target proteins BCL-2 and Cyclin D1
(38). These two molecules function as oncogenes, play an important role in the regulation of
apoptosis and proliferation respectively and were found to be over-expressed in different tumors.
The proof of evidence that mir-15a was sufficient and necessary for IL-23 anti-tumor activity
against pediatric B-ALL cells has been obtained using two different and complementary
approaches. Thus, it has been investigated whether i) silencing of mir-15a in B-ALL cells abolished
the effects driven by IL-23 and ii) forced expression of miR15a had a similar anti-tumor activity to
the treatment of IL-23. In vitro and in vivo experiments allowed to demonstrate unambiguously that
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

7
the anti-leukemic function of IL-23 was strictly associated to the up-regulation of miR-15a and that
enforced expression of miR-15a had a similar anti-tumor activity to the treatment of IL-23 causing a
clear inhibition of leukemic cell growth in vivo (4). Although the molecular mechanisms underlying
regulation of miR-15a mediated by IL-23 were not elucidated, it was hypothesized that, similarly to
other cytokines, components of IL-23 signaling pathway may interfere in the biogenesis or
transcription of miR-15a. This pre-clinical study provided the first evidence that cytokines-mediated
regulation of miRNA expression pattern play a key role in the control of B-ALL cell growth.
Finally, studies on the mechanisms underlying the pro-apoptotic effect of IL-23 showed that the
cytokine not only down-regulated the anti-apoptotic BCL-2 but also induced PARP cleavage in the
absence of caspase activation (4).
IL-23 and Th17 function in MM
IL-23 is a well known Th17 differentiating cytokine (39). Recent data highlighted that Th17 play an
important role in MM pathobiology and may be an important therapeutic target for anti-MM
activity and to improve immune function (40). In this view, a number of Th17-associated cytokines,
including IL-23, IL-21, IL-22 and IL-17, were found to be significantly elevated in MM compared
with healthy donors and elevated IL-17 serum levels have been correlated with stage II and III MM
(41). In addition, IL-23 was associated with reduced CD8+ T-cell infiltration in the tumor micro-
environment (30) and a combination of Th17-associated pro-inflammatory cytokines may suppress
T-cell responses. This recent evidence suggests a potential role of IL-23 in Th17-mediated
promotion of MM cell growth and inhibition of immune function. Finally, an imbalance between
Treg/Th17 in favor of Th17, that are not adequately suppressed by T regulatory (reg) cells, may be
suggested since has been reported that Treg cells are decreased in number and dysfunctional in MM
patients (42).
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

8
Bone remodeling in MM
Bone destruction is a hallmark of MM (43) and approximately 85-90% of MM patients present lytic
bone lesions frequently associated with severe bone pains and fractures. Osteolytic lesions result
from increased bone resorption due to stimulation of osteoclast formation and a severely
unbalanced bone remodelling decreased or absent bone formation activity that occur close to MM
cells (44, 45).
The critical role of the receptor activator of nuclear factor kappaB ligand (RANKL), in MM-
induced osteoclastogenesis (44) has been demonstrated. RANKL is over-expressed in the BM in
relationship with MM cell infiltration whereas its decoy receptor osteoprotegerin (OPG) is down-
regulated (44). Other cytokines produced by MM cells such as Macrophage Inhibitory Protein
(MIP)-1α (namely CCL3) or induced in the microenvironment by the cell contact are responsible
for enhancement of osteoclast differentiation and activity as well as for inhibition of osteoblast
functions (46). Interestingly, the growth factors released in the course of increased bone resorptive
process as well as activated osteoclasts in turn support MM cell growth and survival, thus inducing
a “vicious cycle”(47-49). Consistently it has been reported that anti-osteoclastic agents may exert
anti-MM effect both in vitro and in vivo (50).
Effects of IL-27 and IL-23 on osteoblastic and osteoclastic cells and bone remodelling
Recent evidences suggest that both IL-27 and IL-23 may affect in a different manner the bone
remodeling process. Osteoclast progenitors from MM patients and controls (5) express functional
IL-27R and IL-27 dampens directly osteoclast differentiation and activity (11). It has been also
reported that IL-27 induced proliferation in osteoblastic cells while unaffecting osteoblast formation
(11, 51), indicating that IL-27 amplifies the mature osteoblastic compartment without modulating
its functionality. Thus, IL-27 may counteract the unbalanced osteoclast and osteoblast activity
induced by MM cells mainly exerting an inhibitory effect on osteoclastic cells.
The effect of IL-23 on osteoclast and bone resorption has been evaluated showing conflicting
results. Yago et al. (52) demonstrated that in human peripheral blood mononuclear cells, IL-23
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

9
induced osteoclastogenesis in the absence of exogenous soluble RANKL. In this study, IL-17
induced by IL-23 from human activated T cells was considered the pivotal cytokine for
osteoclastogenesis through the involvement of the RANK–RANKL system and tumor necrosis
factor (TNF)-α. Thus, IL-17 acted on monocytes and induced osteoclast differentiation and TNF-α
production by cooperating with RANKL. Similarly, it was reported that IL-23 stimulate RANKL
expression on CD4+ T cells and contribute to osteoclastogenesis in an arthritis mouse model (53). In
contrast, Quinn et al. reported that IL-23 inhibited osteoclastogenesis indirectly thought CD4+ T
cells and that IL-23p19-/- had reduced bone mass (54). Kamiya and coworkers (51) showed that IL-
23 was ineffective on RANKL expression and that osteoclastogenesis induced by soluble RANKL
was partially inhibited by IL-23, while unaffecting proliferation of osteoclast progenitors. Any
significant effect of IL-23 was reported on osteoblastic cells and on osteoblast formation (51).
Overall these evidences suggest that under physiological condition IL-23 favors high bone mass by
limiting bone resorption whereas in pathophisiological conditions IL-23 share a stimulatory effect
on osteoclast formation mainly through the induction of RANKL by T cells and the differentiation
of Th17 subset and IL-17 production. The role and the involvement of Th17 and IL-17 in osteoclast
activation in MM has been recently demonstrated (55). Because IL-23 is a well known cytokine
involved in Th17 differentiation and expansion, it is possible that the high BM levels of IL-23
observed in MM patients (40) contribute to the increased osteoclastogenesis through the expansion
of Th17 and consequently the overproduction of IL-17.
Human normal plasma cells
In humans, three major subsets of plasma cells have been isolated and are supposed to represent a
gradual increase in PC maturation: early PC in tonsils, transitional PC in peripheral blood and
mature PC in the BM (56). However, PC are rare cells in vivo representing only 1-2% of tonsil
mononuclear cells and less than 0.5% of BM cells in healthy individuals. Thus, it has been
developed an in vitro reproducible model of peripheral blood B cell differentiation into polyclonal
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

10
plasmablastic cells (PPC) that have morphological and phenotypic features of PC (57). In addition,
this model allowed to obtain a homogenous population of normal plasmablastic cells that provided a
powerful tool to identify genes specifically involved in normal plasma cell differentiation, to further
understand the biology of MM and to improve therapeutic developments..
Expression and role of I-L27 and IL-23 receptor in normal PC
The expression of both chains of the IL-27R and of IL-23R was demonstrated in tonsil and BM PC
as well as in PPC generated in vitro and the functional role of IL-27 and IL-23 was recently
elucidated (5). Since production of immunoglobulin (Ig) represents the main PC function and a key
step that occurs during antigen-specific immune response, we asked whether IL-27 and IL-23 may
modulate Ig secretion in these cells. These experiments showed that stimulation of PPC with IL-2 in
association with IL-27 or IL-23 caused significant up-regulation of IgM secretion paralleled by
inhibition of IgG secretion and unaltered IgA production. However, no effects were reported in
terms of modulation of IgG class switching (5, 10).
Thus, it has been hypothesized that these cytokines are effective on PPC that have not yet
undergone terminal differentiation into antibody-secreting cells since such effects were reverted
when IL-6 was present for the last days of PPC culture. This latter consideration was strongly
supported by the finding that IL-23 and IL-27 had no effect on modulation of Ig secretion in
CD138+ tonsil PC.
Another novel finding was the first demonstration that IL-27 possesses chemotactic properties
versus PC and PPC (5) conceivably related to dependent to STAT-1 activation, as recently
demonstrated for dendritic cells in response to cytokines, such as IFN-β, was (58).
Finally, IL-27 up-regulated in PC and PPC the expression and production of chemokines such as
CXCL9 that induce migration of activated T and NK cells and possess potent anti-angiogenic
activity through interferon (IFN)-γ dependent/independent pathways (11, 59, 60). However, in PPC
and PC, CXCL9 and 10 modulated expressions by IL-27 was not accompanied by induction of IFN-
γ production.
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

11
Taken together, these findings support the concept that IL-27 and more marginally IL-23 plays an
important role in the course of immunological response to pathogens by acting at different levels.
First, IL-23 and IL-27 produced by antigen-presenting cells stimulated by Toll-like receptor signals
may induce short-lived PC to produce low affinity IgM providing a rapid initial response to
pathogens. Thus, IL-27 may attract plasmablast/short-lived PC that up-regulate CXCL9, but not
CCL chemokines, thus creating chemokine gradients that recruit to inflamed tissue activated T,
memory T and NK cells but not granulocytes and monocytes. Finally, CXCL9 and 10 may limit
tissue damage provoked by persistent activation of inflammatory cells in view of their well known
anti-angiogenic activities. These findings may be also relevant in the MM pathogenesis since
normal PC take part in the MM microenvironment. Thus, the anti-angiogenic CXCL9 up-regulated
in normal PC may amplify the inhibition of angiogenesis caused by the direct activity of IL-27 on
MM cells.
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

12
CLINICAL-TRANSLATIONAL ADVANCES
Cytokines of the IL-12 family: potential clinical implications
The potential translational implication of the new evidences on the role of IL-23 and/or IL-27 in the
biology of normal and malignant PC is related to the ability of these cytokines to stimulate immune
responses, target directly tumor cells expressing the corresponding receptors and, at least for IL-27,
to inhibit tumor angiogenesis. Either the modulation of IL-12 family cytokines including IL-23 and
IL-27 or the direct use of these cytokines could be supposed in the future in clinical trials.
In the case of IL-12, pre-clinical studies demonstrated that this cytokine can target directly MM
cells and cause a strong reduction of MM pro-angiogenic potential, with marginal effects on MM
cell proliferation and survival (3). However a particular note of caution regarding the potential
therapeutic implication of IL-12, comes from the side effects that have been reported in clinical
trials addressing the anti-tumor activity of IL-12 against different malignancies.
The initial clinical studies have been performed in phase I trials with patients affected by advanced
malignancies (renal cancer, colon carcinoma and melanoma) to determine the optimal protocol of
treatment; the parameters analyzed were toxicity, maximum tolerated dose (MTD), biological and
anti-tumor activities, and route of administration. Both intravenous (i.v) (61-63) and sub-cutaneous
(s.c) (64, 65) injection schedules have been tested in patients at different times and doses. The IL-12
MTD was different in i.v. and s.c. route of injection, 500 ng/kg and 1000-1500 ng/kg respectively,
and s.c administration resulted in fewer and milder toxic effects than i.v. treatment. Although
preliminary studies suggested that IL-12 might be efficacious, subsequent trials did not confirm the
first encouraging results. Thus, despite an enhancement of immune response, the clinical efficacy
was minimal. Moreover, escalating doses of IL-12 induced serious side effects in patients. Good
therapeutic results have been obtained in patients affected by cutaneous T cell lymphoma, AIDS-
related Kaposi sarcoma and non-Hodgkin’s lymphoma which showed a clear increase in circulating
and tumor infiltrating T cells with objective responses of 56%, 50%, 43%, respectively (66-68). The
favourable results obtained in immunodeficient AIDS patients suffering from Kaposi’s sarcoma was
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

13
not surprising since this tumor is strongly angiogenic and angiogenesis inhibition is one of the most
prominent IL-12 activities. Thus, it was speculated that the anti-neoplastic activity of IL-12 was
largely dependent on its ability to inhibit angiogenesis.
In order to get over or reduce systemic toxicity, novel forms of IL-12 non-viral delivery such as
alum, liposome and polymer-based delivery, have shown in pre-clinical models promising effects
(69). A further refinement of tumor targeted IL-12 therapy would be the use of targeted
formulations of the cytokine such as that developed by Halin and coworkers (70), who have
generated a fusion protein containing an IL-12 moiety fused to a single chain monoclonal antibody
specific for tenascin-C, an isoform of this extacellular matrix protein expressed predominantly in
the tumor stroma. This fusion protein, however, has not yet been tested in clinical trials. More
recently, the effectiveness of electrogene therapy with IL-12 has been evaluated in preclinical
studies (71) and the anti-tumor efficacy was tested in different types of primary tumors, distantly
growing tumors and induced metastases. Thus, it has been shown that intra-tumoral IL-12
electrogene therapy has been proved to be very effective in local tumor control, having also a
systemic effect, whereas intramuscular and peri-tumoral IL-12 electrogene therapy had also a
pronounced systemic effect. The anti-tumor effectiveness of IL-12 electrogene therapy was due to
the induction of adaptive immunity and innate resistance and anti-angiogenic action.
A further promising area of investigation, with potentially relevant clinical applications, stems from
the emerging knowledge on both IL-23 and IL-27 have been used to effectively treat tumors in
murine models. Systemic administration of IL-23 has been demonstrated to be effective in mouse
tumor system including melanoma and fibrosarcoma (32, 72). In addition preclinical evidences
suggest that IL-23 is a promising candidate as a tumor vaccine adjuvant (37). Interestingly adverse
toxic effect of systemic treatment with IL-23 appear to be much less than those observed for IL-12
although some authors reported weight loss and inflammation of the intestinal tract on the basis of
IL-23 schedule treatment (37). On the other hand, mice injected intra-peritoneally with recombinant
IL-23 did not show splenomegaly and liver inflammation as compared to those treated with IL-12
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

14
(72). Therefore in a clinical perspective, the route of administration for IL-23 should be adequately
designed to avoid its adverse effects and to exert efficient anti-tumor effect.
Despite the several evidences describing the anti-tumoral effect of IL-23 in tumoral models, caution
in its clinical application would be justified due to either the role of IL-23 in inflammatory diseases
or the possible role of this cytokine in tumoral progression (30, 36, 73, 74). In fact in mice lacking
IL-23p19 or IL-23 receptor, IL-23 showed a tumoral promoting effect in contrast to its potent anti-
tumor effect. The reasons for this discrepancy are not known, however the potential clinical use of
this cytokines should be carefully evaluated. In addition, the IL-23 seems to be ineffective against
MM cells and its role in the control of MM microenvironment has not been clearly elucidated.
By contrast, we could envisage combinatorial approaches using IL-12 and IL-27 to promote T and
NK cell responses against MM in addition to their direct anti-tumor activity against MM and to the
IL-27 ability to stimulate the osteoblast proliferation and to inhibit osteoclast differentiation and
function. In this view, exciting results have been reported against melanoma and breast cancer using
IL-12 and IL-27 sequential gene therapy through intramuscular electroporation (75). More recently,
it has been reported the efficacy of IL-27 as anti-tumor agent against prostate cancer using a new
mode of gene delivery named 'sonoporation' that involved low-frequency ultrasound irradiation
(76). In this study, three models of immune-competent prostate adenocarcinoma have been used and
detailed characterizations of the tumor growth reduction, gene profile expression, and effector
cellular profiles have been reported. Results from this investigation suggested that IL-27 can be
effective in reducing tumor growth and can help enhance accumulation of effector cells in prostate
tumors in vivo (76).
In conclusion, pre-clinical studies support the concept that IL-27 may represent a novel, promising
therapeutic agent for patients affected by different oncologic diseases including potentially MM,
based upon its documented multifunctional activity. Before this step, the development of future
clinical trials to evaluate the toxicity and efficacy of IL-27 will be mandatory. An additional
argument in favor of this proposal is the low toxicity shown by IL-27 in animal models, likely in
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

15
relation to the low induction of IFN-γ in vivo (32). Clearly, dedicated studies are required to define
the clinical utility of IL-27 as anti-angiogenic and anti-tumoral agent in patients with solid cancer
and MM.
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

16
Acknowledgments
This work was supported by grants from Associazione Italiana Ricerca sul Cancro (A.I.R.C.)
Milano, Italy (grant number 4014 to I.A. and 8530 to N.G.), Italian Ministry of Health (RF, RC,
5/1000, Progetto Strategico Oncologico 2006 rif070701), International Myeloma Foundation (to
N.G.), Italian Ministry of Health-Progetti Regione Emilia Romagna and the and Special Program
Molecular Clinical Oncology 5 per mille (n. 9965).
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

17
References
1. Trinchieri G, Pflanz S and Kastelein RA. The IL-12 family of heterodimeric cytokines: new
players in the regulation of T cell responses. Immunity 2003;19: 641-4
2. Hunter CA. New IL-12-family members: IL-23 and IL-27, cytokines with divergent functions.
Nat Rev Immunol 2005;5: 521-31
3. Airoldi I, Cocco C, Giuliani N, Ferrarini M, Colla S, Ognio E, et al. Constitutive expression of
IL-12R beta 2 on human multiple myeloma cells delineates a novel therapeutic target. Blood
2008;112: 750-9
4. Cocco C, Canale S, Frasson C, Di Carlo E, Ognio E, Ribatti D, et al. Interleukin-23 acts as
antitumor agent on childhood B-acute lymphoblastic leukemia cells. Blood 2010;116: 3887-98
5. Cocco C, Morandi F and Airoldi I. Interleukin-27 and interleukin-23 modulate human plasmacell
functions. J Leukoc Biol 2011; 89:729-34
6. Ferretti E, Di Carlo E, Cocco C, Ribatti D, Sorrentino C, Ognio E, et al. Direct inhibition of
human acute myeloid leukemia cell growth by IL-12. Immunol Lett 2010;133: 99-105
7. Yoshimoto T, Morishima N, Mizoguchi I, Shimizu M, Nagai H, Oniki S, et al. Antiproliferative
activity of IL-27 on melanoma. J Immunol 2008;180: 6527-35
8. Pflanz S, Hibbert L, Mattson J, Rosales R, Vaisberg E, Bazan JF, et al. WSX-1 and glycoprotein
130 constitute a signal-transducing receptor for IL-27. J Immunol 2004;172: 2225-31
9. Xu M, Mizoguchi I, Morishima N, Chiba Y, Mizuguchi J and Yoshimoto T. Regulation of
antitumor immune responses by the IL-12 family cytokines, IL-12, IL-23, and IL-27. Clin Dev
Immunol 2010: 2010. pii: 832454
10. Larousserie F, Charlot P, Bardel E, Froger J, Kastelein RA and Devergne O. Differential effects
of IL-27 on human B cell subsets. J Immunol 2006;176: 5890-7
11. Airoldi I, Di Carlo E, Banelli B, Moserle L, Cocco C, Pezzolo A, et al. The IL-12Rbeta2 gene
functions as a tumor suppressor in human B cell malignancies. J Clin Invest 2004;113: 1651-9
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

18
12. Airoldi I, Di Carlo E, Cocco C, Taverniti G, D'Antuono T, Ognio E, et al. Endogenous IL-12
triggers an antiangiogenic program in melanoma cells. Proc Natl Acad Sci U S A 2007;104: 3996-
4001
13. Hisada M, Kamiya S, Fujita K, Belladonna ML, Aoki T, Koyanagi Y, et al. Potent antitumor
activity of interleukin-27. Cancer Res 2004;64: 1152-6
14. Shimizu M, Shimamura M, Owaki T, Asakawa M, Fujita K, Kudo M, et al. Antiangiogenic and
antitumor activities of IL-27. J Immunol 2006;176: 7317-24
15. Rajkumar SV, Leong T, Roche PC, Fonseca R, Dispenzieri A, Lacy MQ, et al. Prognostic value
of bone marrow angiogenesis in multiple myeloma. Clin Cancer Res 2000;6: 3111-6
16. Vacca A and Ribatti D. Bone marrow angiogenesis in multiple myeloma. Leukemia 2006;20:
193-9
17. Giuliani N, Colla S, Lazzaretti M, Sala R, Roti G, Mancini C, et al. Proangiogenic properties of
human myeloma cells: production of angiopoietin-1 and its potential relationship to myeloma-
induced angiogenesis. Blood 2003;102: 638-45
18. Holash J, Maisonpierre PC, Compton D, Boland P, Alexander CR, Zagzag D, et al. Vessel
cooption, regression, and growth in tumors mediated by angiopoietins and VEGF. Science
1999;284: 1994-8
19. Uneda S, Matsuno F, Sonoki T, Tniguchi I, Kawano F and Hata H. Expressions of vascular
endothelial growth factor and angiopoietin-2 in myeloma cells. Haematologica 2003;88: 113-5
20. Podar K, Tai YT, Davies FE, Lentzsch S, Sattler M, Hideshima T, et al. Vascular endothelial
growth factor triggers signaling cascades mediating multiple myeloma cell growth and migration.
Blood 2001;98: 428-35
21. Vincent L, Jin DK, Karajannis MA, Shido K, Hooper AT, Rashbaum WK, et al. Fetal stromal-
dependent paracrine and intracrine vascular endothelial growth factor-a/vascular endothelial growth
factor receptor-1 signaling promotes proliferation and motility of human primary myeloma cells.
Cancer Res 2005;65: 3185-92
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

19
22. Barille S, Akhoundi C, Collette M, Mellerin MP, Rapp MJ, Harousseau JL, et al.
Metalloproteinases in multiple myeloma: production of matrix metalloproteinase-9 (MMP-9),
activation of proMMP-2, and induction of MMP-1 by myeloma cells. Blood 1997;90: 1649-55
23. Di Raimondo F, Azzaro MP, Palumbo G, Bagnato S, Giustolisi G, Floridia P, et al. Angiogenic
factors in multiple myeloma: higher levels in bone marrow than in peripheral blood. Haematologica
2000;85: 800-5
24. Vacca A, Ribatti D, Presta M, Minischetti M, Iurlaro M, Ria R, et al. Bone marrow
neovascularization, plasma cell angiogenic potential, and matrix metalloproteinase-2 secretion
parallel progression of human multiple myeloma. Blood 1999;93: 3064-73
25. Vacca A, Ribatti D, Ria R, Pellegrino A, Bruno M, Merchionne F, et al. Proteolytic activity of
human lymphoid tumor cells. Correlation with tumor progression. Dev Immunol 2000;7: 77-88
26. Parham C, Chirica M, Timans J, Vaisberg E, Travis M, Cheung J, et al. A receptor for the
heterodimeric cytokine IL-23 is composed of IL-12Rbeta1 and a novel cytokine receptor subunit,
IL-23R. J Immunol 2002;168: 5699-708
27. Carmody RJ, Ruan Q, Liou HC and Chen YH. Essential roles of c-Rel in TLR-induced IL-23
p19 gene expression in dendritic cells. J Immunol 2007;178: 186-91
28. Verreck FA, de Boer T, Langenberg DM, Hoeve MA, Kramer M, Vaisberg E, et al. Human IL-
23-producing type 1 macrophages promote but IL-10-producing type 2 macrophages subvert
immunity to (myco)bacteria. Proc Natl Acad Sci U S A 2004;101: 4560-5
29. Langowski JL, Kastelein RA and Oft M. Swords into plowshares: IL-23 repurposes tumor
immune surveillance. Trends Immunol 2007;28: 207-12
30. Langowski JL, Zhang X, Wu L, Mattson JD, Chen T, Smith K, et al. IL-23 promotes tumour
incidence and growth. Nature 2006;442: 461-5
31. Tartour E, Fossiez F, Joyeux I, Galinha A, Gey A, Claret E, et al. Interleukin 17, a T-cell-
derived cytokine, promotes tumorigenicity of human cervical tumors in nude mice. Cancer Res
1999;59: 3698-704
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

20
32. Oniki S, Nagai H, Horikawa T, Furukawa J, Belladonna ML, Yoshimoto T, et al. Interleukin-23
and interleukin-27 exert quite different antitumor and vaccine effects on poorly immunogenic
melanoma. Cancer Res 2006;66: 6395-404
33. Ugai S, Shimozato O, Yu L, Wang YQ, Kawamura K, Yamamoto H, et al. Transduction of the
IL-21 and IL-23 genes in human pancreatic carcinoma cells produces natural killer cell-dependent
and -independent antitumor effects. Cancer Gene Ther 2003;10: 771-8
34. Yuan X, Hu J, Belladonna ML, Black KL and Yu JS. Interleukin-23-expressing bone marrow-
derived neural stem-like cells exhibit antitumor activity against intracranial glioma. Cancer Res
2006;66: 2630-8
35. Hu J, Yuan X, Belladonna ML, Ong JM, Wachsmann-Hogiu S, Farkas DL, et al. Induction of
potent antitumor immunity by intratumoral injection of interleukin 23-transduced dendritic cells.
Cancer Res 2006;66: 8887-96
36. Kaiga T, Sato M, Kaneda H, Iwakura Y, Takayama T and Tahara H. Systemic administration of
IL-23 induces potent antitumor immunity primarily mediated through Th1-type response in
association with the endogenously expressed IL-12. J Immunol 2007;178: 7571-80
37. Overwijk WW, de Visser KE, Tirion FH, de Jong LA, Pols TW, van der Velden YU, et al.
Immunological and antitumor effects of IL-23 as a cancer vaccine adjuvant. J Immunol 2006;176:
5213-22
38. Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin M, Shimizu M, et al. miR-15 and miR-16
induce apoptosis by targeting BCL2. Proc Natl Acad Sci U S A 2005;102: 13944-9
39. Korn T, Bettelli E, Oukka M and Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol
2009;27: 485-517
40. Prabhala RH, Pelluru D, Fulciniti M, Prabhala HK, Nanjappa P, Song W, et al. Elevated IL-17
produced by TH17 cells promotes myeloma cell growth and inhibits immune function in multiple
myeloma. Blood 2010;115: 5385-92
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

21
41. Alexandrakis MG, Pappa CA, Miyakis S, Sfiridaki A, Kafousi M, Alegakis A, et al. Serum
interleukin-17 and its relationship to angiogenic factors in multiple myeloma. Eur J Intern Med
2006;17: 412-6
42. Prabhala RH, Neri P, Bae JE, Tassone P, Shammas MA, Allam CK, et al. Dysfunctional T
regulatory cells in multiple myeloma. Blood 2006;107: 301-4
43. Kyle RA and Rajkumar SV. Multiple myeloma. N Engl J Med 2004;351: 1860-73
44. Giuliani N, Colla S and Rizzoli V. New insight in the mechanism of osteoclast activation and
formation in multiple myeloma: focus on the receptor activator of NF-kappaB ligand (RANKL).
Exp Hematol 2004;32: 685-91
45. Roodman GD. Pathogenesis of myeloma bone disease. Blood Cells Mol Dis 2004;32: 290-2
46. Giuliani N and Rizzoli V. Myeloma cells and bone marrow osteoblast interactions: role in the
development of osteolytic lesions in multiple myeloma. Leuk Lymphoma 2007;48: 2323-9
47. Giuliani N, Mangoni M and Rizzoli V. Osteogenic differentiation of mesenchymal stem cells in
multiple myeloma: identification of potential therapeutic targets. Exp Hematol 2009;37: 879-86
48. Giuliani N, Rizzoli V and Roodman GD. Multiple myeloma bone disease: Pathophysiology of
osteoblast inhibition. Blood 2006;108: 3992-6
49. Roodman GD. Pathogenesis of myeloma bone disease. Leukemia 2009;23: 435-41
50. Fowler JA, Edwards CM and Croucher PI. Tumor-host cell interactions in the bone disease of
myeloma. Bone 48: 121-8
51. Kamiya S, Nakamura C, Fukawa T, Ono K, Ohwaki T, Yoshimoto T, et al. Effects of IL-23 and
IL-27 on osteoblasts and osteoclasts: inhibitory effects on osteoclast differentiation. J Bone Miner
Metab 2007;25: 277-85
52. Yago T, Nanke Y, Kawamoto M, Furuya T, Kobashigawa T, Kamatani N, et al. IL-23 induces
human osteoclastogenesis via IL-17 in vitro, and anti-IL-23 antibody attenuates collagen-induced
arthritis in rats. Arthritis Res Ther 2007;9: R96
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

22
53. Ju JH, Cho ML, Moon YM, Oh HJ, Park JS, Jhun JY, et al. IL-23 induces receptor activator of
NF-kappaB ligand expression on CD4+ T cells and promotes osteoclastogenesis in an autoimmune
arthritis model. J Immunol 2008;181: 1507-18
54. Quinn JM, Sims NA, Saleh H, Mirosa D, Thompson K, Bouralexis S, et al. IL-23 inhibits
osteoclastogenesis indirectly through lymphocytes and is required for the maintenance of bone mass
in mice. J Immunol 2008;181: 5720-9
55. Noonan K, Marchionni L, Anderson J, Pardoll D, Roodman GD and Borrello I. A novel role of
IL-17-producing lymphocytes in mediating lytic bone disease in multiple myeloma. Blood 116:
3554-63
56. Medina F, Segundo C, Campos-Caro A, Gonzalez-Garcia I and Brieva JA. The heterogeneity
shown by human plasma cells from tonsil, blood, and bone marrow reveals graded stages of
increasing maturity, but local profiles of adhesion molecule expression. Blood 2002;99: 2154-61
57. Tarte K, De Vos J, Thykjaer T, Zhan F, Fiol G, Costes V, et al. Generation of polyclonal
plasmablasts from peripheral blood B cells: a normal counterpart of malignant plasmablasts. Blood
2002;100: 1113-22
58. Yen JH, Kong W and Ganea D. IFN-beta inhibits dendritic cell migration through STAT-1-
mediated transcriptional suppression of CCR7 and matrix metalloproteinase 9. J Immunol 184:
3478-86
59. Kanegane C, Sgadari C, Kanegane H, Teruya-Feldstein J, Yao L, Gupta G, et al. Contribution of
the CXC chemokines IP-10 and Mig to the antitumor effects of IL-12. J Leukoc Biol 1998;64: 384-
92
60. Mahalingam S, Chaudhri G, Tan CL, John A, Foster PS and Karupiah G. Transcription of the
interferon gamma (IFN-gamma )-inducible chemokine Mig in IFN-gamma-deficient mice. J Biol
Chem 2001;276: 7568-74
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

23
61. Atkins MB, Robertson MJ, Gordon M, Lotze MT, DeCoste M, DuBois JS, et al. Phase I
evaluation of intravenous recombinant human interleukin 12 in patients with advanced
malignancies. Clin Cancer Res 1997;3: 409-17
62. Gollob JA, Mier JW, Veenstra K, McDermott DF, Clancy D, Clancy M, et al. Phase I trial of
twice-weekly intravenous interleukin 12 in patients with metastatic renal cell cancer or malignant
melanoma: ability to maintain IFN-gamma induction is associated with clinical response. Clin
Cancer Res 2000;6: 1678-92
63. Robertson MJ, Cameron C, Atkins MB, Gordon MS, Lotze MT, Sherman ML, et al.
Immunological effects of interleukin 12 administered by bolus intravenous injection to patients with
cancer. Clin Cancer Res 1999;5: 9-16
64. Motzer RJ, Rakhit A, Thompson JA, Nemunaitis J, Murphy BA, Ellerhorst J, et al. Randomized
multicenter phase II trial of subcutaneous recombinant human interleukin-12 versus interferon-
alpha 2a for patients with advanced renal cell carcinoma. J Interferon Cytokine Res 2001;21: 257-
63
65. Portielje JE, Kruit WH, Schuler M, Beck J, Lamers CH, Stoter G, et al. Phase I study of
subcutaneously administered recombinant human interleukin 12 in patients with advanced renal cell
cancer. Clin Cancer Res 1999;5: 3983-9
66. Geller AC, Swetter SM, Brooks K, Demierre MF and Yaroch AL. Screening, early detection,
and trends for melanoma: current status (2000-2006) and future directions. J Am Acad Dermatol
2007;57: 555-72; quiz 573-6
67. Little RF, Pluda JM, Wyvill KM, Rodriguez-Chavez IR, Tosato G, Catanzaro AT, et al. Activity
of subcutaneous interleukin-12 in AIDS-related Kaposi sarcoma. Blood 2006;107: 4650-7
68. Younes A, Pro B, Robertson MJ, Flinn IW, Romaguera JE, Hagemeister F, et al. Phase II
clinical trial of interleukin-12 in patients with relapsed and refractory non-Hodgkin's lymphoma and
Hodgkin's disease. Clin Cancer Res 2004;10: 5432-8
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

24
69. Salem ML, Gillanders WE, Kadima AN, El-Naggar S, Rubinstein MP, Demcheva M, et al.
Review: novel nonviral delivery approaches for interleukin-12 protein and gene systems: curbing
toxicity and enhancing adjuvant activity. J Interferon Cytokine Res 2006;26: 593-608
70. Halin C, Rondini S, Nilsson F, Berndt A, Kosmehl H, Zardi L, et al. Enhancement of the
antitumor activity of interleukin-12 by targeted delivery to neovasculature. Nat Biotechnol 2002;20:
264-9
71. Cemazar M, Jarm T and Sersa G. Cancer electrogene therapy with interleukin-12. Curr Gene
Ther 10: 300-11
72. Nagai H, Oniki S, Fujiwara S, Yoshimoto T and Nishigori C. Antimelanoma immunotherapy:
clinical and preclinical applications of IL-12 family members. Immunotherapy 2010;2: 697-709
73. Hao JS and Shan BE. Immune enhancement and anti-tumour activity of IL-23. Cancer Immunol
Immunother 2006;55: 1426-31
74. Lo CH, Lee SC, Wu PY, Pan WY, Su J, Cheng CW, et al. Antitumor and antimetastatic activity
of IL-23. J Immunol 2003;171: 600-7
75. Zhu S, Lee DA and Li S. IL-12 and IL-27 sequential gene therapy via intramuscular
electroporation delivery for eliminating distal aggressive tumors. J Immunol 2010; 184: 2348-54
76. Zolochevska O, Xia X, Williams J, Ramsay A, Li S and Figueiredo ML. Sonoporation delivery
of Interleukin 27 gene therapy efficiently reduces prostate tumor cell growth in vivo. Hum Gene
Ther 2011 Jul 29. [Epub ahead of print]
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

25
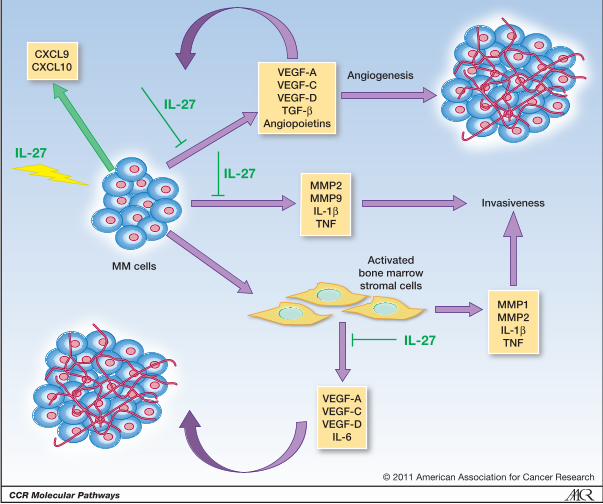
Legend to Figure 1
IL-27 activity against MM cells and the “growth paracrine loop”
IL-27 acts directly on MM cells that express complete IL-27R. IL-27 inhibits the angiogenic
potential of MM cells and down-regulates different angiogenic factors serve also as autocrine
and/or paracrine growth factors. Thus, IL-27 may interrupt or reduce the “growth loop” in which
cytokines and growth factors released by MM cells induce paracrine stimulation of stromal cells
that, in turn, secrete additional growth factors that stimulate new vessel formation and support MM
cell proliferation, progression and migration.
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724
© 2011 American Association for Cancer Research
MMP2MMP9IL-1βTNF
CXCL9CXCL10
MMP1MMP2IL-1βTNF
VEGF-AVEGF-CVEGF-DTGF-β
Angiopoietins
VEGF-AVEGF-CVEGF-D
IL-6
Activatedbone marrowstromal cells
Invasiveness
Angiogenesis
IL-27
IL-27
IL-27
IL-27
MM cells
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724

Published OnlineFirst August 31, 2011.Clin Cancer Res Nicola Giuliani and Irma Airoldi malignant and normal plasma cellsNovel insights into the role of IL-27 and IL-23 in human
Updated version
10.1158/1078-0432.CCR-11-1724doi:
Access the most recent version of this article at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/early/2011/08/31/1078-0432.CCR-11-1724To request permission to re-use all or part of this article, use this link
Research. on December 21, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2011; DOI: 10.1158/1078-0432.CCR-11-1724





























