Embed Size (px)
DESCRIPTION
El mejoramiento del valor nutricional de las plantas mediante ingenieria genetica, ha demostrado que muchas deficiencias de estas pueden ser implantadas para su mejora nutricional y nutraceutica
Citation preview

167
Critical Reviews in Plant Sciences, 21(3):167–204 (2002)
0735-2689/02/$.50© 2002 by CRC Press LLC
Genetic, Molecular, and Genomic Approaches toImprove the Value of Plant Foods and Feeds
Gad Galili,1* Shmuel Galili,2 Efraim Lewinsohn,2 and Yaakov Tadmor 2
1Department of Plant Sciences, The Weizmann Institute of Science, Rehovot 76100 Israel, and 2Institute ofField and Garden Crops, Agricultural Research Organization, PO Box 6, Bet Dagan 50250 Israel
Referee: Dr. T.J. Higgins, Chief Research Scientist, CSIRO, Divistion of Plant Industry, Clunies Ross Street, Box 1600, Canberr a,
2601, Australia
* This review was written as an equal contribution of four scientists studying different disciplines of food and feed quality. Theseinclude amino acid and storage protein metabolism (GG, Email: [email protected]); ruminant and non-ruminant animalsfeeding (SG, Email: [email protected]); secondary metabolism (EL, Email: [email protected]); lipids and minerals(YT, Email: [email protected]).
ABSTRACT: Recent advances in gene isolation, plant transformation, and genetic engineering are being usedextensively to alter metabolic pathways in plants by tailormade modifications to single or multiple genes. Manyof these modifications are directed toward increasing the nutritional value of plant-derived foods and feeds. Theseapproaches are based on rapidly growing basic knowledge, understanding, and predictions of metabolic fluxes andnetworks. Some of the predictions appear to be accurate, while others are not, reflecting the fact that plantmetabolism is more complex than we presently understand. Tailor-made modifications of plant metabolism hasso far been directed into improving the levels of primary metabolites that are essential for growth and developmentof humans and their livestock. Yet, the list of improved metabolites is expected to grow tremendously after newdiscoveries in nutritional, medical, and health sciences. Despite our extensive knowledge of metabolic networks,many of the genes encoding enzymes, particularly those involved in secondary metabolism, are still unknown.These genes are being discovered at an accelerated rate by recent advances in genetic and genomics approaches.In the present review, we discuss examples in which the nutritional and health values of plant-derived foods andfeeds were improved by metabolic engineering. These include modifications of the levels of several essentialamino acids, lipids, fatty acids, minerals, nutraceuticals, antinutritional compounds, and aromas.
I. INTRODUCTION
The classic role of agricultural crops as themajor food supplier to feed a growing and hungrypopulation is still substantial, but today there isalso a great demand to increase the nutritionalquality of this food. Improved nutritional qualitymay help solve problems encountered in caseswere plant foods are the major or sole source offood, that is, plant foods in many developingcountries as well as plant feeds for livestock,which are consumed as human foods. The de-mands for these sectors focus particularly on im-proving the levels of essential amino acids, min-erals, and basic vitamins to allow the healthygrowth of humans and livestock. In addition, withthe developed awareness of human health, there
is also a great demand by the public in Westerncountries for fortified plant foods to improve hu-man health and life expectancy. The demands forthis category include a number of exotic healthcompounds such as essential oils and exotic anti-oxidants, which may improve life, especially dur-ing the elder life stages. Besides the nutritionalaspect, the value of plant foods depends also onits taste and structure. Thus, improving the tasteand aroma of foods are also important areas incrop breeding.
As is the case with other agronomic traits, themajor approach to improving the value of plantfoods and feeds is by classic genetics and breeding.This approach has been tremendously successful;however, it is a relatively slow process and de-pends on the relatively narrow genetic variability

168
within many crop plants. During the last 2 or sodecades, other approaches for tailormade improve-ments of food and feed quality have developed andare being incorporated into plant breeding togetherwith classic genetics. These include DNA-marker-assisted breeding, direct gene transfer, and, morerecently, the use of genomics.
The strategies for enhancing the value of plantfoods and feeds include altering metabolic pathwaysusing genes for enzymes responsible for the biosyn-thesis of specific phytochemicals. Such studies de-pend on existing metabolic knowledge, which is notalways accurate. So-called tailormade perturbationof metabolic pathways does not always result in theexpected changes. Thus, an essential part of a genetechnology based approach to enhanced value ofplant foods and feeds depends on the full knowledgeof the relevant biochemical pathway.
Many aspects of the nutritive value of plantsand how to improve it have already been dis-cussed (for examples see Abelson and Hines, 1999;Agarwal and Rao, 2000; Baucher et al., 1998;Brink and Beynen, 1992; Dillard and German,2000; Dixon and Steele, 1999; Dunwell, 1999;Dunwell, 2000; Galili, 1995; Gaskell et al., 1999;Giddings et al., 2000; Grima-Pettenati and Goffner,1999; Grusak, 1999; Grusak and DellaPenna,1999; Hefford, 1997; Mandal and Mandal, 2000;Marriott, 2000; Merchen and Trigemeyer, 1992;Miflin et al., 1999; Ohlrogge and Benning, 2000;Serageldin, 1999; Shotwell and Larkins, 1989;Teferedegne, 2000; Van Duyn and Pivonka, 2000;Williamson et al., 1999). Here we focus and at-tempt to provide a critical opinion on selectedimprovements to the nutritional value of plantsfor foods and feeds. These include essential aminoacids, fatty acids and lipids, minerals, vitaminsand health products, antinutritional factors, as wellas aroma and flavor. We discuss new genetic andgenomic approaches that are promising for thegenetic introgression of foreign genes into culti-vated species as well as the identification of novelgenes regulating plant metabolism.
II. IMPROVING THE LEVELS OFESSENTIAL AMINO ACIDS
Non-ruminant animals (such as humans, poul-try, and swine) cannot synthesize 10 out of the 20
protein amino acids and therefore have to obtainthese “essential” amino acids in their diet. Al-though ruminant animals (such as cattle and sheep)also cannot synthesize essential amino acids, theyhave special microbial flora in their rumen, whichmetabolize nonessential amino acids into essen-tial amino acids and incorporate them into micro-bial proteins that later become available for nutri-tion. However, these microbial proteins, althoughof better nutritional quality than plant proteins,provide only ~65% of the total protein requiredfor intensive milk production (Leng, 1990). Hence,ruminant animals also suffer from limitations inessential amino acids. Moreover, attempts to im-prove the nutritional quality of foods for ruminantanimals require specific considerations, based onthe nature of their rumen microbial flora (seeSection III.B.5).
Lysine and methionine are the most impor-tant essential amino acids because they are presentin limiting levels in the major feed and food crops.Cereal grains generally contain low levels of lysine(Shotwell and Larkins, 1989), while legumes aregenerally deficient in methionine (Duke, 1981).Due to the vital nutritional significance of lysineand methionine, most efforts have focused onenhancing the levels of these two essential aminoacids in their free or protein-bound forms.
A. Breeding of High-Lysine Cereals: TheStory of “Quality Protein Maize”
Cereal grains represent the main dietary sourceof protein for human and livestock worldwide.Maize is one of the most important cereal crops,providing between 50 and 70% of the dietaryproteins for humans, depending on geographicaldistribution. It is also one of the major crops usedfor livestock feeding. Because maize is very lowin lysine, a large effort was made to identify high-lysine corn varieties. These efforts resulted in thediscovery of the high-lysine opaque2 mutants(Mertz, 1997; Mertz et al., 1964). These lines arecharacterized by low levels of lysine-poor seedstorage proteins (called zeins), and by a compen-satory increase in lysine-rich, non-zein, seed pro-teins, as well as free lysine. Rat feeding trialsshowed that opaque2 flour, together with miner-als and vitamins, promoted a fourfold increase in

169
growth rate over that in rats grown on normalmaize flour (Mertz et al., 1964). Moreover, theopaque2 flour was found to have 90% the valueof milk protein when fed to Guatemalan children(Bressani, 1966). A diet based solely on opaque2flour was also later shown to cure children whosuffered from the protein deficiency disease,kwashiorkor (Harpstead, 1971). Success withopaque2 maize stimulated extensive research toidentify similar mutants in other cereals. Similarhigh-lysine mutants were found in barley (Doll etal., 1974; Munck et al., 1970), and sorghum (Singhand Axtell, 1973).
However, despite the initial optimism, subse-quent detailed field analyses showed that the high-lysine mutations in these cereals were associatedwith inferior agronomic traits that could not beeasily overcome. The undesirable traits includedreduced yield and protein content as well as softendosperm that caused disease and insect suscep-tibility, kernel breakage, and poor food process-ing (Glover, 1992; Munck, 1992). Commercialutilization of the opaque2 mutants seemed un-likely until 1992, when researchers at the Maizeand Wheat Improvement Center (CIMMYT) inMexico and the University of Natal in South Af-rica (Geevers and Lake, 1992; Glover, 1992) couldgenetically separate the inferior agronomic traitsfrom the benefits of the opaque2 mutation. Thisresulted in high-lysine “Quality Protein Maize”(QPM) lines with normal kernel properties. Sincethis important discovery, high-lysine QPM culti-vars have been used extensively in Brazil(Magnavaca et al., 1993), and interest in suchcultivars is increasing in North America (Bockholtand Rooney, 1992). A recent analysis of a numberof QPM cultivars, adapted to the conditions ofCanada (Zarkadas et al., 2000), looks quite prom-ising. Total grain protein in these cultivars rangedfrom 8.0 to 10.2%, which is similar to the 7.9 to10.3% range in leading non-QPM cultivars. Lysinecontent in the QPM lines ranged between 4.43and 4.58 g lysine/100 g protein, which is signifi-cantly higher than that the levels in the non-QPMcultivars (between 3.43 and 4.21 g of lysine/100g protein). The lysine levels in these QPM lineswas calculated to supply around 70% of the opti-mal human protein requirement, whereas the bestnon-QPM cultivars supply less than 50% of the
requirement. The molecular mechanism underly-ing the QPM genotype has been studied in detail,particularly by Larkins and associates (Burnettand Larkins, 1999; Lopes et al., 1995; Or et al.,1993; Sun et al., 1997), and is not discussed here.The production of one of the high-lysine proteinsin the maize kernel, the translation factor EF-1α,is positively correlated with lysine levels in dif-ferent QPM lines (Habben et al., 1995). This canbe used to rapidly screen and select outstandingQPM lines adapted to various growth conditionsand geographical locations.
B. Improving Free Amino AcidSynthesis and Accumulation
1. Regulation of Lysine and MethionineSynthesis
The essential amino acids lysine, methionine,isoleucine, and threonine are synthesized fromaspartate by several different branches of the as-partate-family pathway (Figure 1) (Galili, 1995).Methionine receives its sulfur moiety from cys-teine (Figure 1) (Ravanel et al., 1998). Lysineregulates its own synthesis mostly by feedbackinhibiting the activity of dihydrodipicolinate syn-thase (DHPS); threonine synthesis is primarilyregulated by the sensitivity of aspartate kinase tofeedback inhibition by lysine and threonine (Galili,1995); methionine synthesis is subjected to a morecomplex control, but it has been suggested that amajor point of regulation of methionine produc-tion occurs by the competition between cystathion-ine γ-synthase and threonine synthase for theircommon substrate, phosphohomoserine (Figure 1)(see Ravanel et al., 1998 for detailed discussionof this competition).
2. Improving the Level of Free Lysine inGrain Crops
Conceptually, the synthesis of lysine can beenhanced by reducing the sensitivity of DHPS tofeedback inhibition by lysine (Figure 1). This hasbeen proven in studies where recombinant genesencoding bacterial feedback-insensitive DHPS
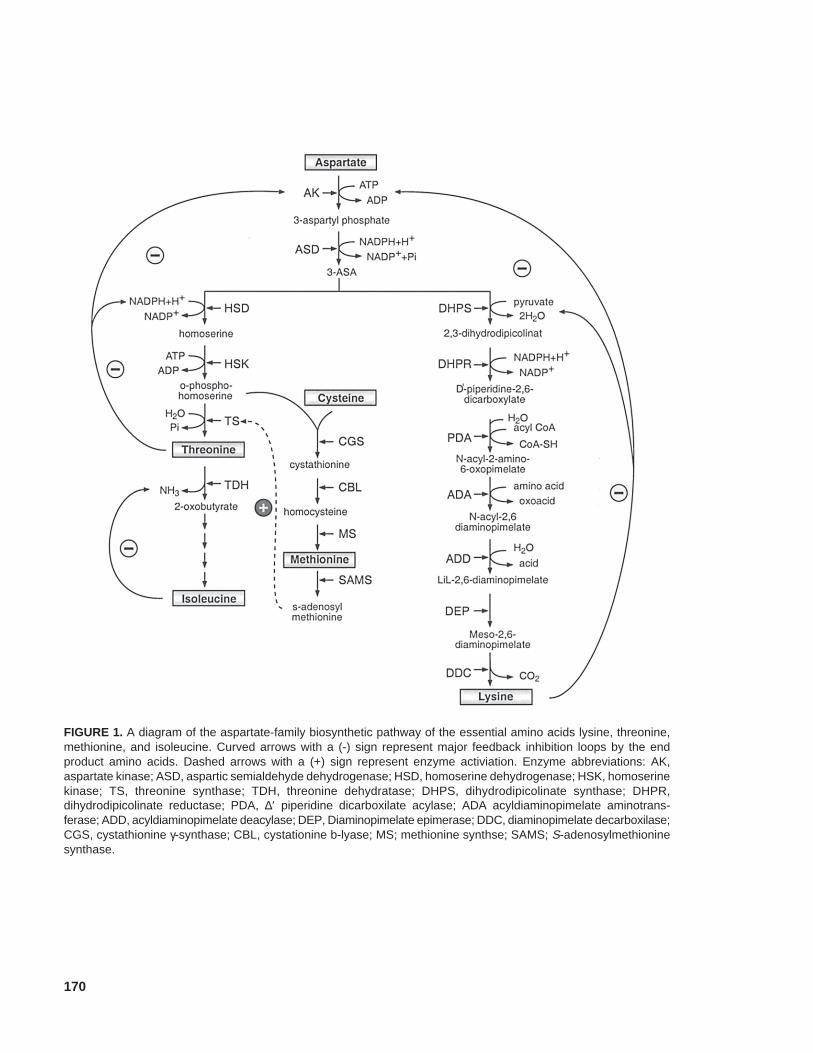
170
FIGURE 1. A diagram of the aspartate-family biosynthetic pathway of the essential amino acids lysine, threonine,methionine, and isoleucine. Curved arrows with a (-) sign represent major feedback inhibition loops by the endproduct amino acids. Dashed arrows with a (+) sign represent enzyme activiation. Enzyme abbreviations: AK,aspartate kinase; ASD, aspartic semialdehyde dehydrogenase; HSD, homoserine dehydrogenase; HSK, homoserinekinase; TS, threonine synthase; TDH, threonine dehydratase; DHPS, dihydrodipicolinate synthase; DHPR,dihydrodipicolinate reductase; PDA, ∆′ piperidine dicarboxilate acylase; ADA acyldiaminopimelate aminotrans-ferase; ADD, acyldiaminopimelate deacylase; DEP, Diaminopimelate epimerase; DDC, diaminopimelate decarboxilase;CGS, cystathionine γ-synthase; CBL, cystationine b-lyase; MS; methionine synthse; SAMS; S-adenosylmethioninesynthase.

171
enzymes were expressed in transgenic dicot andmonocot plants (Galili, 1995; Brinch-Pedersen etal., 1996; Mazur et al., 1999). Constitutive ex-pression of genes for bacterial enzymes in plantsresulted in amino acid overproduction, but in manycases this expression was also associated withabnormal phenotypes and partial sterility (BenTzvi-Tzchori et al., 1996; Frankard et al., 1992;Shaul and Galili, 1992; Shaul and Galili, 1993).To overcome these problems, the bacterial en-zymes were produced in a seed-specific manner,using seed storage protein promoters to controlexpression of the genes. Seed-specific expressionof the bacterial DHPS gene was first performed intobacco plants, using the bean phaseolin promoter(Karchi et al., 1994). Lysine synthesis was en-hanced specifically in the developing seeds ofthese transgenic plants, but its level in matureseeds was not higher than in nontransgenic plants.Seed-specific expression of the bacterial DHPSgene was correlated with a significant elevation inthe activity of lysine ketoglutarate reductase(LKR), the first enzyme in the α-amino adipicacid pathway, which catabolizes lysine inglutamate, α-amino adipic acid, and acetyl CoA(Arruda et al., 2000; Galili et al., 2001). Theseresults were the first indirect and unexpected evi-dence that lysine catabolism may be an importantfactor regulating free lysine accumulation in seeds.Moreover, it also suggested that lysine autoregu-lates its own catabolism, at least in seeds, bystimulating LKR activity (Arruda et al., 2000;Galili et al., 2001; Karchi et al., 1995; Karchi etal., 1994). In subsequent studies, bacterial DHPSshave been expressed in a seed-specific manner ina number of transgenic crop plants, includingsoybean, rapeseed, maize, and narbon beans (Falcoet al., 1995; Mazur et al., 1999; M. Meixner,S. Gillandt, K. Waigand, G. Galili, and T. Pickardt,unpublished). In contrast to tobacco, all of thesetransgenic crops showed significant elevation offree lysine levels, and nearly doubled total seedlysine in soybean and rapeseed. Lysine overpro-duction in these plants was also associated withincreased levels of various catabolic products,showing that most if not all seeds possess anactive process of lysine catabolism. The negativeeffects of lysine catabolism are twofold: first itreduces the extent of free lysine accumulation;
second, some of the catabolic products of lysine,such as glutamate and the products of γ-aminobutyric acid and α-amino adipic acid act as neu-rotransmitters in animals and may be toxic at highlevels (Bonaventure et al., 1985; Karlsen et al.,1982; Reichenbach and Wohlrab, 1985; Welinderet al., 1982). Therefore, the reduction of lysinecatabolism may be an important trait to be consid-ered in breeding for high-lysine crops. Lysinecatabolism can be reduced by antisense, co-sup-pression, or knockout of genes encoding enzymesin this part of the pathway.
The use of seed storage protein gene promot-ers for the expression of the bacterial DHPS geneis based on the assumption that amino acid syn-thesis and storage protein production are subjectto concerted spatial and temporal regulation ofexpression during seed development. This is ap-parently true for dicot plants in which seed stor-age protein genes are expressed in the developingembryo and utilization of such promoters to ex-press the bacterial DHPS results in lysine over-production. However, whether this is also true formonocot plants, in which storage protein synthe-sis occurs mainly in the endosperm, is still debat-able. Falco and associates (Mazur et al., 1999)have expressed a bacterial feedback-insensitiveDHPS gene in transgenic maize, using either en-dosperm or embryo-specific promoters. Increasedfree lysine levels were detected only when DHPSproduction was controlled by the embryo-specificpromoter. This study raises an important funda-mental issue. If amino acid biosynthesis in mono-cot seeds occurs mostly in the developing em-bryo, a mechanism should exist for rapid deliveryof the amino acids into the endosperm to supportthe massive process of storage protein synthesis.If amino acids are synthesized in the endospermtissues, then lysine level may be regulated byadditional factors, such as lysine catabolism. Thesignificance of lysine catabolism in dicot andmonocot seeds was discovered as a consequenceof this transgenic approach (see previous para-graph). However, this still does not provide a fullexplanation for the results of Falco and associates(Mazur et al., 1999), because embryo-specific,but not endosperm-specific, expression of the bac-terial DHPS was accompanied by increased lev-els of lysine catabolic products.

172
Another strategy for the production of high-lysine plants is to introduce genes encoding lysine-rich proteins. These proteins will serve as a lysinesink and may reduce the problem of lysine ca-tabolism. In maize, a variety of genes for natural,modified, and synthetic proteins were tested, andthe most successful encoded hordothionine (HT12)and barley high lysine protein 8 (BHL8), contain-ing 28 and 24% lysine, respectively. These pro-teins accumulated to between 3 to 6% of totalgrain proteins, and when introduced together witha bacterial DHPS resulted in a marked elevationof total lysine to over 0.7% of seed dry weight(Jung and Falco, 2000), compared with around0.2% in wild-type maize. If this dramatic eleva-tion of lysine levels does not interfere with yieldand other grain quality factors, then the commer-cial application of transgenic maize expressingthese high-lysine proteins (either alone or togetherwith a bacterial feedback-insensitive DHPS) forfeeding human and nonruminant livestock looksvery promising. The suitability of such transgenicplants for ruminant feeding depends on whetherthe high-lysine proteins are stable inside the ru-men.
3. Improving Methionine Levels in GrainCrops
Methionine synthesis is far more complicatedthan that of lysine. Methionine receives its carbonskeleton from the aspartate family pathway, whileits sulfur moiety is derived from cysteine, whosesynthesis is also subject to a complex regulation.The regulation of sulfate uptake and incorpora-tion into cysteine and other sulfur compounds hasbeen reviewed recently (Bick and Leustek, 1998;Saito, 2000) and is not discussed here. Rather, wefocus on efforts to manipulate the carbon flux intomethionine, as well as on attempts to expressmethionine-rich proteins in transgenic plants.Because methionine synthesis diverges from thethreonine branch of the aspartate-family pathway(Figure 1), it is expected that plants possessing afeedback-insensitive aspartate kinase will alsooverproduce methionine because they possessincreased flux toward threonine (Galili, 1995).Yet, such an approach has resulted in the produc-
tion of relatively minor increases in methioninelevels over that in nontransgenic plants (Galili,1995; Karchi et al., 1993), suggesting that otherregulatory factors exist. Methionine synthesis wasthought to be regulated by competition betweencystathionine γ-synthase and threonine synthasefor their common substrate phosphohomoserine(Figure 1) (Ravanel et al., 1998). Threonine syn-thase activity is also negatively regulated byS-adenosyl methionine (SAM), a direct productof methionine (Figure 1), further implicating au-toregulation of methionine synthesis by modulat-ing metabolite flux via the threonine synthase/cystathionine γ-synthase branch point (Ravanel etal., 1998). Yet, despite extensive studies, the regu-lation of metabolite flux via this branch point isstill unclear. Constitutive overexpression of cys-tathionine γ-synthase in transgenic Arabidopsiscaused a severalfold increase in free methioninein rosette leaves (Gakiere et al., 2000; Tarczynskiet al., 2001). A more dramatic ~40-fold elevationin free methionine was reported in rosette leavesof the Arabidopsis mto1 mutant, which possessesa point mutation in the coding region of the cys-tathionine γ-synthase gene (Chiba et al., 1999;Inba et al., 1994). Yet, in both the transgenic andmutant Arabidopsis plants, no overproduction ofmethionine was observed in mature plants(Chiba et al., 1999; Gakiere et al., 2000; Inbaet al., 1994). This suggests that methioninesynthesis is differentially regulated duringplant development. Inhibition of threoninesynthase activity by an antisense approachresulted in a huge overaccumulation of freemethionine both in Arabidopsis and potato(Batlem et al., 2000; Zeh et al., 2001). More-over, the increase in methionine was muchhigher than the decrease in threonine, suggest-ing that the reduction in threonine synthaseactivity somehow triggers the channeling ofmore aspartate into methionine, despite thefeedback sensitivity of AK. Although the com-plexity of methionine synthesis is not under-stood, these results are promising from a nu-tritional point of view, suggesting that it ispossible to manipulate methionine levels inplants.
Attempts to increase methionine levels intransgenic plants by expressing genes encod-

173
ing methionine-rich proteins were performedin a number of plant species, using a variety ofgenes. These attempts have been discussed indetail in several reviews (see, for example,Muntz et al., 1998; Tabe and Higgins, 1998).In most cases, genes for methionine-rich 2Sstorage proteins were used. In soybean, whosegrain methionine level amounts more than halfof the FAO standard for nutritionally balancedfood protein, expression of the gene for Brazilnut 2S albumin raised seed methionine con-tent to 100% of the FAO standard. Expressionof the same gene in transgenic narbon beans,whose seed methionine level is only 40% ofthe FAO standard, doubled seed methioninecontent (Saalbach et al., 1995a; Saalbach etal., 1995b). Unfortunately, this Brazil nut pro-tein was subsequently found to be allergenicin some people, reducing the usefulness ofthis protein as a target for increasing plantnutritional quality. In another study, a differ-ent 2S albumin, derived from sunflower, wasused to significantly increase seed methioninecontent in transgenic lupin, an important graincrop for animal feeding in Australia that con-tains less than half of the methionine recom-mended by FAO. Expression of the sunfloweralbumin doubled seed methionine contentreaching 80% of the FAO standard (Molvig etal., 1997). Notably, rat feeding experimentswith these transgenic lupin grains showed notonly an increased of methionine availability,but also an increased in their general dietaryvalue (Molvig et al., 1997).
Although expression of genes for methion-ine-rich proteins seems to be a promising ap-proach to increasing overall methionine avail-ability in foods and feeds, it is still not enough toincrease methionine content to 100% of the FAOrecommendation. Müntz and associates(D. Demidov, C. Horstmann, M. Meixner,T. Pickardt, I. Saalbach, G. Galili, and K. Müntz,unpublished) have therefore combined the ex-pression of a Brazil nut protein together with abacterial feedback-insensitive aspartate kinasein narbon bean seeds, which controls the carbonflux for free methionine synthesis (Galili, 1995).This combined approach raised methionine con-tent in the seeds to 100% of the FAO standard.
4. Improving Lysine and MethionineLevels in Forage Crops
In forage crops the main consumed part is thevegetative tissue, and therefore efforts to increasethe essential amino acid content in vegetativetissues were mainly conducted by constitutiveexpression of recombinant constructs expressingseed vacuolar storage proteins, fused to the 35Spromoter. These storage proteins which stablyaccumulate in seeds vacuoles, failed to accumu-late in the protease-rich vegetative vacuoles, dueto their efficient degradation (Saalbach et al.,1994). Preventing the trafficking of the seed stor-age proteins from the endoplasmic reticulum (ER)to the vegetative vacuole by engineering of an ERretention signal (KDEL) into the C-terminus ofthese proteins only partially solved their stabilityproblems (Khan et al., 1996; Tabe et al., 1995;Wandelt et al., 1992). More successful resultswere obtained by using two methionine-rich seedstorage proteins of maize, namely, the 15-kDaβ-zein and the 10-kDa δ-zein, which naturallyaccumulate in ER-derived protein bodies (Shotwelland Larkins, 1989), Maize β-zein and δ-zein genes,constitutively expressed alone in transgenic to-bacco plants, accumulated in novel ER-derivedprotein bodies and were moderately stable (Baggaet al., 1995). Co-expression of the two proteinstogether significantly increased their stability(Bagga et al., 1997). Stability problems associ-ated with the expression of seed storage proteinsin vegetative tissues suggest that expression ofgenes for other types of nutritionally balancedproteins should also be tried. Inasmuch as a num-ber of plants also naturally accumulate vegetativestorage proteins (VSPs) to high levels inside veg-etative vacuoles (Staswick, 1994), such proteinsmay be better targets for nutritional improvementof forage crops than seed storage proteins. VSPsmay also have additional beneficial effects, suchas enhancement of shoot regrowth after cutting offorage crops (Avice et al., 1997; Corbel et al.,1999). Galili and associates (Guenoune et al.,1999) overexpressed the soybean VSPα gene,fused to the Cauliflower mosaic virus (CaMV)35S promoter, in transgenic tobacco plants. Thisprotein was highly stable in vacuoles of bothvegetative and seed tissues. The level of the soy-

174
bean VSPα ranged between 2 and 6% of thesoluble proteins in leaves of the transgenic plants,causing a significant increase of total soluble lysineby about 15%. This suggests that VSPs can serveas excellent protein sources for improving thenutritional quality of forage crops.
5. Improving Protein Quality forRuminants Feeding
Specific approaches to increase the content ofessential amino acids in transgenic plants shouldtake into consideration the target uses of theseplants. Nonruminant animals depend absolutelyon the dietary essential amino acids, but can effi-ciently absorb both free and protein-bound aminoacids. The situation with ruminant animals is morecomplex due to the special microbial flora in theirrumen. Although the rumen micro-flora can pro-duce essential amino acids, it can also degradeintake proteins and convert their amino acids intoother nitrogenous compounds. Thus, when feed-ing ruminants with dietary proteins either bal-anced or enriched for essential amino acids, it isimportant to minimize their degradation by therumen micro-flora. As much as 40% of the di-etary protein may be lost from the rumen of ani-mals grazing on temperate legumes due to micro-bial degradation (Ulyatt et al., 1988). Thisphenomenon can also limit the availability oflysine and methionine for young ruminants(Merchen and Trigemeyer, 1992) and for lactat-ing dairy cows (Rulquin and Verite, 1993). Thus,proteins with optimal lysine and methionine con-tent for ruminant nutrition should be highly resis-tant to degradation in the rumen.
Analyzing various proteins by SDS PAGEafter in vitro or in situ rumen digestions, Spenceret al. (Spencer et al., 1988) showed that someplant and animal proteins, such as bovine serumalbumin (BSA) and pea albumins, are highly stableto rumen proteolysis. In contrast, other proteinssuch as casein and vicilin were rapidly degraded(McNabb et al., 1994; Spencer et al., 1988; Tabeet al., 1993). Recent studies (Hancock et al., 1994;McNabb et al., 1994; Tabe et al., 1993) usedWestern blot analysis to follow more accuratelythe in vitro and in situ rumen degradation rates of
specific proteins. Barry and associates (McNabbet al., 1994) found that the degradation of vicilinand Rubisco small subunit occurred in singlephase, whereas the degradation of the Rubiscolarge subunit, ovalbumin and sunflower albumin8 was biphasic. The half-life inside the rumenfluid varied between 10 min for vicilin to 69.3 hfor the second component of the sunflower albu-min 8. Comparing the in situ degradation rates ofseveral proteins having different proportions ofsulfur-containing amino acids, White and associ-ates (Hancock et al., 1994) concluded that thestability of a protein to rumen degradation posi-tively correlates with the degree of cross-linkingby disulfide bonds.
Searches for stable proteins as targets forexpression in transgenic plants for ruminant feed-ing assume that the stability of a given proteinwill be similar when produced in different plantspecies. This may, however, not be always thecase. Galili and Guenoune (Guenoune et al., 2002)studied the rumen stability of either VSPs fromsoybean or recombinant genes for soybean VSPs,expressed in transgenic tobacco plants. The soy-bean-derived proteins were much more stable torumen proteolysis than those produced in thetransgenic tobacco.
In the rumen, protein stability can be increasedby moderate amounts of condensed tannins (CT),which are produced by some forage crops. CTform reversible associations with proteins; theformation of these protein-tannin complexes mak-ing protein unavailable for ruminal microbialdeamination (for review see Aerts et al., 1999).However, excess CT, as it occurs in several tem-perate and tropical forages, can be detrimental tothe overall nutritive value of the crop because itprevents forage intake and digestion by the ani-mal. The amount of CT necessary to prevent pro-tein degradation, but not to reduce intake, must beestablished for each forage crop species, and spe-cies containing optimal CT levels may be excel-lent targets for transformation with genes encod-ing proteins rich in essential amino acids.Alternatively, it may be possible to modify CTstructure and concentration in forage crops bymolecular approaches. This research is underwaybut is still at a very early stage (Robbins et al.,1998).

175
Because animal feeds undergo different typesof processing, stability of the transgenic proteinsunder these conditions should also be considered.Galili and associates (Galili et al., 1999) showedthat both leaf and seed storage proteins of wheatwere completely degraded during ensiling. Com-plete degradation was also shown for a Bacillusthuringiensis toxin in transgenic corn plants (Fear-ing et al., 1997). Degradation of the transgenicproteins may be less problematic when the feedsare supplied as hay (Khoudi et al., 1999).
III. IMPROVING LIPIDS AND FATTY ACIDCOMPOSITION AND CONTENT
Lipids are an important class of natural prod-ucts, which includes fat-soluble steroids, prostag-landins, triglycerides, waxes, and long-chain fattyacids. Many of these compounds are essential tothe human diet (e.g., essential fatty acids, fat-soluble vitamins) and therefore are of great inter-est to the food industry. Fatty acids are an impor-tant component of lipids in plants, animals, andmicroorganisms. They are composed of long, even-numbered carbon chains with a carboxylic groupat one end of the chain and a methyl group at theother. Saturated fatty acids with 16 (palmitic acid)and 18 (stearic acid) carbon atoms are the mostcommon in nature.
Higher plants produce more than 200 differ-ent fatty acids. There are many questions aboutthe nature of the enzymes involved in their syn-thesis (Somerville et al., 2000). The synthesis offatty acids in plants takes place in various or-ganelles and in some cases involves the move-ment of lipids from one cellular compartment toanother (Ohlrogge and Browse, 1995; Ohlroggeand Jaworski, 1997; Padley et al., 1994). Althoughfatty acid metabolism in plants has many featuresin common with other organisms, the plant path-ways are complex and not well understood.
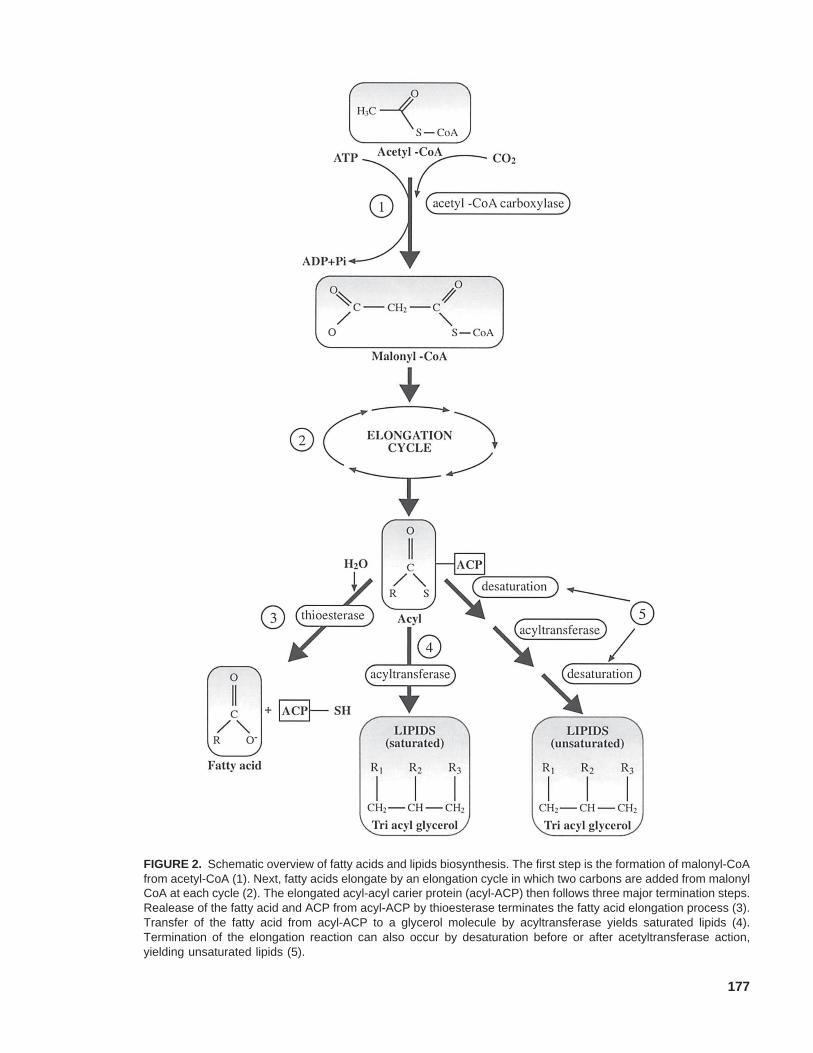
Fats are broadly divided into saturated andpolyunsaturated classes. Within the polyunsatu-rated fatty acids are two families of essential fattyacids (EFAs). They are termed essential becauseour bodies need them but cannot manufacturethem (Simopolus, 1999). Major EFAs are theomega-6 linoleic acid, its omega-6 derivative
γ-linolenic acid (abundant in Evening Primrose,Oenothera biennis), as well as the omega-3α-linolenic acid, largely present in linseed, Linumusitatissimum (Table 1). EFAs function mainly ascomponents of cellular membranes, and as pre-cursors to eicosanoids, including prostaglandinsand leukotrienes (Newton, 1998).
The incidences of chronic degenerative dis-eases such as coronary diseases and cancer havebeen increasing in developed countries. Thesediseases were very rare in developing countriesand unknown among traditional Eskimos. Therate of increase of these diseases in the latter twosocieties is associated with adaptation to a mod-ern diet. Deficiency and unbalanced EFAs in thebody cause many of the diseases. Thus, searchingfor ways to increase the content of specific EFAsin human diets is of high nutritional priority.
A. Synthesis of Lipids
Significant progress has been made in the last3 decades on the genetics and physiology of lipidsmetabolism. These have been reviewed in detailby several authors (Harwood, 1996; Harwood,1997; Ohlrogge and Browse, 1995; Ohlrogge andJaworski, 1997; Weselake and Taylor, 1999).Genes encoding key enzymes of fatty acid andlipid biosynthesis have been cloned and charac-terized from a number of plants (Murphy, 1999;Napier et al., 1999), where lipid synthesis is ini-tiated in the plastids. The first step is the carboxy-lation of acetyl-CoA to malonyl-CoA (Figure 2,reaction 1), catalyzed by the enzyme, acetyl-CoAcarboxylase (ACCase). Next, an elongation cycle,catalyzed by several enzymes, attaches a series oftwo carbon additions to the growing chain (Fig-ure 2, reaction 2). Typically, the elongation endsby production of saturated fatty acids of 16 or 18carbons. Among the most common terminatingreactions are hydrolysis of the acyl moiety fromacyl carrier protein by a thioesterase (Figure 2,reaction 3), transfer of the acyl moiety from acylcarrier protein directly onto a glycerolipid by anacyl-transferase (Figure 2, reaction 4), or doublebond formation on the acyl moiety by an acyl-ACP desaturase (Figure 2, reaction 5) (Somervilleet al., 2000).
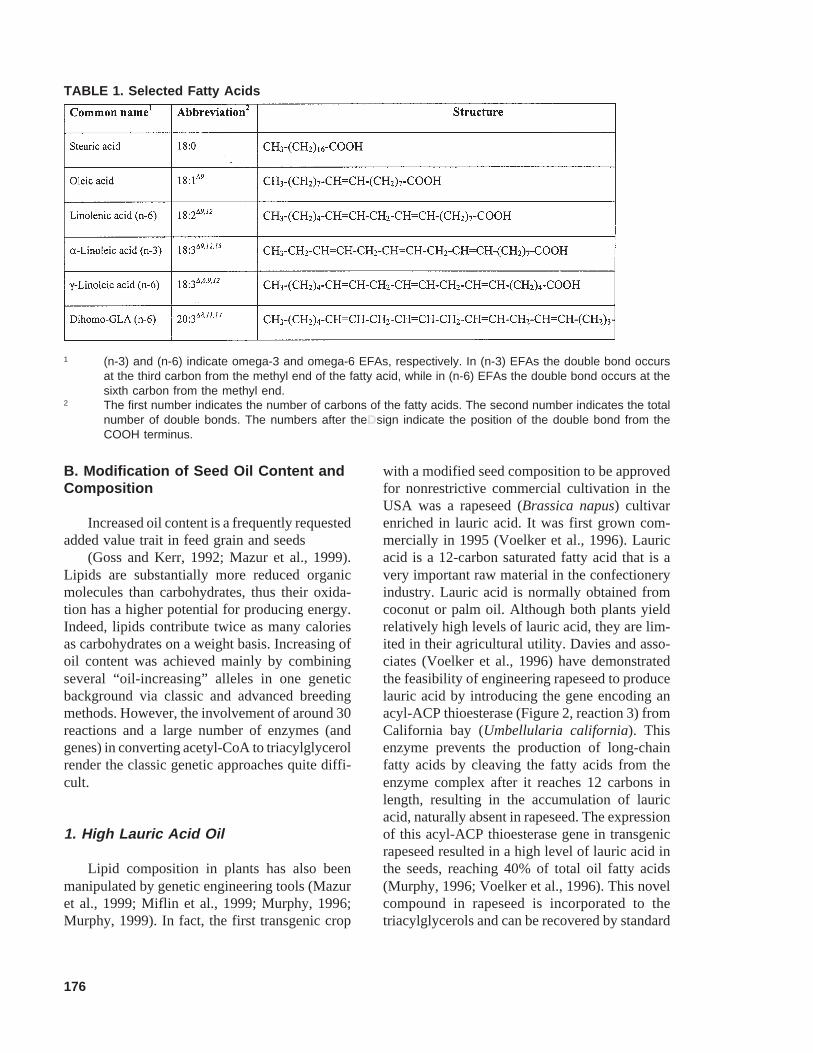
176
B. Modification of Seed Oil Content andComposition
Increased oil content is a frequently requestedadded value trait in feed grain and seeds
(Goss and Kerr, 1992; Mazur et al., 1999).Lipids are substantially more reduced organicmolecules than carbohydrates, thus their oxida-tion has a higher potential for producing energy.Indeed, lipids contribute twice as many caloriesas carbohydrates on a weight basis. Increasing ofoil content was achieved mainly by combiningseveral “oil-increasing” alleles in one geneticbackground via classic and advanced breedingmethods. However, the involvement of around 30reactions and a large number of enzymes (andgenes) in converting acetyl-CoA to triacylglycerolrender the classic genetic approaches quite diffi-cult.
1. High Lauric Acid Oil
Lipid composition in plants has also beenmanipulated by genetic engineering tools (Mazuret al., 1999; Miflin et al., 1999; Murphy, 1996;Murphy, 1999). In fact, the first transgenic crop
with a modified seed composition to be approvedfor nonrestrictive commercial cultivation in theUSA was a rapeseed (Brassica napus) cultivarenriched in lauric acid. It was first grown com-mercially in 1995 (Voelker et al., 1996). Lauricacid is a 12-carbon saturated fatty acid that is avery important raw material in the confectioneryindustry. Lauric acid is normally obtained fromcoconut or palm oil. Although both plants yieldrelatively high levels of lauric acid, they are lim-ited in their agricultural utility. Davies and asso-ciates (Voelker et al., 1996) have demonstratedthe feasibility of engineering rapeseed to producelauric acid by introducing the gene encoding anacyl-ACP thioesterase (Figure 2, reaction 3) fromCalifornia bay (Umbellularia california). Thisenzyme prevents the production of long-chainfatty acids by cleaving the fatty acids from theenzyme complex after it reaches 12 carbons inlength, resulting in the accumulation of lauricacid, naturally absent in rapeseed. The expressionof this acyl-ACP thioesterase gene in transgenicrapeseed resulted in a high level of lauric acid inthe seeds, reaching 40% of total oil fatty acids(Murphy, 1996; Voelker et al., 1996). This novelcompound in rapeseed is incorporated to thetriacylglycerols and can be recovered by standard
1 (n-3) and (n-6) indicate omega-3 and omega-6 EFAs, respectively. In (n-3) EFAs the double bond occursat the third carbon from the methyl end of the fatty acid, while in (n-6) EFAs the double bond occurs at thesixth carbon from the methyl end.
2 The first number indicates the number of carbons of the fatty acids. The second number indicates the totalnumber of double bonds. The numbers after theDsign indicate the position of the double bond from theCOOH terminus.
TABLE 1. Selected Fatty Acids
177
FIGURE 2. Schematic overview of fatty acids and lipids biosynthesis. The first step is the formation of malonyl-CoAfrom acetyl-CoA (1). Next, fatty acids elongate by an elongation cycle in which two carbons are added from malonylCoA at each cycle (2). The elongated acyl-acyl carier protein (acyl-ACP) then follows three major termination steps.Realease of the fatty acid and ACP from acyl-ACP by thioesterase terminates the fatty acid elongation process (3).Transfer of the fatty acid from acyl-ACP to a glycerol molecule by acyltransferase yields saturated lipids (4).Termination of the elongation reaction can also occur by desaturation before or after acetyltransferase action,yielding unsaturated lipids (5).

178
processing methods. Transgenic rapeseed lauricoil is now marketed for use in confectionery inNorth America under the ‘Laurical’ trademark.This work demonstrates the feasibility of produc-ing large amounts of transgene-modified plantoils to supplement or replace existing sources.
Lipids are usually stored as triacylglycerols,three fatty acids esterified to glycerol (Padley etal., 1994). Seed oils predominantly contain C18unsaturated fatty acids (Hilditch and Williams,1964). Saturated fatty acids are normally foundonly in the sn-1 and sn-3 positions (sn-1 = R1, sn-3= R3; figure 2) of the triacylglycerol (Frentzen,1998; Padley et al., 1994). Even in oilseed speciesthat produce triacylglycerol with elevated levelsof saturated fatty acids, the saturated fatty acidsare mainly located in position sn-1 and sn-3.Analysis of the ‘high-laurate’ rapeseed oil showedthat lauric acid was found almost exclusively atthe sn-1 and sn-3 positions (Voelker et al., 1996).Coconut (Cocos nucifera) oil, for example, con-tains mostly tri-saturated lipids (Padley et al.,1994). Knutzon et al. (Knutzon et al., 1995) iso-lated the saturated fatty acid acyltransferase genefrom coconut. Co-expression of the genes of thecoconut acyltransferase and California bay acyl-ACP thioesterase in rapeseed facilitated efficientlauric acid deposition at the sn-2 position, result-ing in the accumulation of tri-laurin, and furtherincreased total lauric acid levels above 50%(Knutzon et al., 1999). This demonstrates thatonce a limiting factor in metabolism is known andunderstood it can be overcome by a rational ex-perimental design. The content of lauric acid inthe laurate rapeseed could not be increased be-cause the lauric acid was positioned exclusivelyin sn-1 and sn-3 and not in sn-2. Introducing thegene for the coconut enzyme into the high-lauraterapeseed enabled the incorporation of more laurateinto the triacylglycerol fraction leading to higherincrease in lauric acid.
2. Overexpression of ACCase
Another attempt to alter plant oil content andcomposition was by increasing the activity ofACCase (Figure 2, reaction 1) (Roesler et al.,1997). ACCase appears in two forms in the cell.
The plastidic form that is heteromeric, and thecytosolic form that is homomeric (HO). The ex-pression of the plastidic ACCase is strongly nega-tively auto-regulated (Somerville et al., 2000).Thus, Roesler et al. (Roesler et al., 1997) ex-pressed an Arabidopsis thaliana gene encodingthe HO cytosolic ACCase in seeds of transgenicrapeseed, and targeted the enzyme to the plastids,using a transit peptide from the small subunit ofRubisco. The plastid-localized HO-ACCase wasbiotinylated at a level comparable to cytosolicHO-ACCase and its activity in mature seeds was10- to 20-fold higher than the endogenous ACCaseactivity. ACCase overexpression altered rapeseedfatty acid composition, mostly increasing oleicacid. However, total seed oil content was increasedby only about 5% above to the control plants.This study indicates that ACCase by itself is notthe rate-limiting factor in oil accumulation. Thereason for the increase in oleic acid in thesetransgenic plants is unclear, highlighting the com-plexity of lipid synthesis and the intricate regula-tion of this process in plants. In order to increaseoil content utilizing genetic engineering tools weneed to learn more about the rate limiting factorsthat dictate its accumulation (see below).
3. Expression of sn-2 Acyl-Transferase
Zou et al. (Zou et al., 1997) attempted toincrease oil content in transgenic plants by ma-nipulating the level of sn-2 acyl-transferase (Fig-ure 2, reaction 4), an advanced step intriacylglycerol biosynthesis. Constitutive ex-pression of a yeast sn-2 acyl-transferase intransgenic Arabidopsis and rapeseed resultedin a substantial increase (8 to 48%) in total seedoil content (Zou et al., 1997) as well as in-creases in both proportions and amounts of verylong-chain fatty acids in seed triacylglycerols.Furthermore, the transgenic plants exhibitedelevated activities of lysophosphatidic acidacyltransferase in developing seeds and in-creased proportions of very long-chain fattyacids in the sn-2 position of triacylglycerols.These results illustrate the potential of geneticengineering to increase oil content and compo-sition in plants.

179
C. Essential Fatty Acids
Unsaturated fatty acids, especially the EFAs,are widely marketed as health food supplements.EFAs accumulate in a limited number of plants,including seeds of borage (starflower; Boragoofficinalis L.) and evening primrose. Borage seedscontain 20 to 25% γ-linolenic acid (GLA; Table1), a very uncommon n-6 fatty acid (Gibson et al.,1992), but borage produces low oil yields (aboutone-tenth of the yield of rapeseed). GLA is notproduced by the major oil seed crops; however,many of these crop plants produce significantamounts of the related fatty acid, linoleic acid(Table 1). Desaturation of linoleic acid to GLA iscatalyzed by ∆6 desaturase, which is not presentin major oil crops. The first attempt to increaseGLA involved the constitutive expression ofcyanobacterial ∆6 fatty acid desaturase intransgenic tobacco as a model system (Reddy andThomas, 1996). This study demonstrated the fea-sibility of engineering the production of ‘novel’polyunsaturated fatty acids in transgenic plants,although the GLA yield was poor. A more suc-cessful attempt followed the cloning of the geneencoding ∆6 fatty acid desaturase from borage,and its constitutive expression in transgenic to-bacco (Sayanova et al., 1997). This resulted in theaccumulation of GLA up to 13% of total leaflipids. Seed-specific expression of ∆6 fatty aciddesaturase could cause an increase in GLA pro-duction in oil seed crops, and sunflower, whichcontains 50 to 70% linoleic acid, is a suitablecandidate.
Very long-chain unsaturated EFAs, such asarachidonic acid (AA C20:4 n-6), eicosapentenoicacid (EPA C20:5 n-3), and docosahexenoic acid(DHA C22:5 n-3), are also considered to be nutri-tionally beneficial because of their function ascholesterol-lowering agents (Newton, 1998). EPAis naturally present in fish oils and other marineorganisms (Padley et al., 1994). AA is found insignificant amounts in animal liver and adrenalglands and is also produced by the filamentousfungus Mortierelaa alpina (Padley et al., 1994).The gene responsible for the ∆5 desaturation ofdihomo-GLA (Table 1) to AA (Table 1) was iso-lated from a M. alpina cDNA library, using PCRwith primers from the conserved histidine box
region of microsomal desaturases (Michaelson etal., 1998a), and from Caenorhabditis elegans uti-lizing a similar procedure (Michaelson et al.,1998b). The functionality of the cloned genes hadbeen confirmed by the ability of their products todesaturate dihomo-GLA to AA in yeast. In a morerecent study (Parker-Barnes et al., 2000), a cDNAlibrary, made from the fungus M. alpina, wasexpressed in yeast and screened for the ability toelongate n-6 and n-3 polyunsaturated fatty acids.The protein product of one clone could convertGLA (n-6) to dihomo-GLA (C20:3 n-6), andstearidonic acid (C18:4 n-3) to eicosatetraeonicacid (C20:4 n-3). Yeast cells, co-transformed withthe elongase and the ∆5 desaturase from M. alpina(Michaelson et al., 1998a), produced the expectedAA (C20:4 n-6) and the C20:5 n-3 EPA (Parker-Barnes et al., 2000). The availability of a recom-binant polyunsaturated fatty acid specific elonga-tion enzyme, as well as desaturases, offers excitingpossibilities for producing a wide range of EFA inoil seed crops. Such transgenic crops should helpsatisfy the demands of the nutraceutical and phar-maceutical industries in the near future. To achievethe efficient production of novel EFA by transgenicplants, we need more efforts on the identificationand characterization of key genes involved in thebiosynthesis of EFA, and the targeting of thetransgene products into the storage oil fractionand not to the membrane fraction, where theymight affect membrane properties or signalingfunctions.
D. Increasing the Cooking and FryingQuality of Oil
Plant oils are not only important as directfood components. The quality of oil for cookingand frying is also dependent on the characteristicsof the oils used. Oleic acid (Table 1) is morestable for frying and cooking than are the poly-unsaturated forms, linoleic and linolenic acids(Table 1) (Kinney, 1996; Mazur et al., 1999).Chemical hydrogenation is widely used in thefood industry to raise the oleic acid concentra-tion; however, the chemical hydrogenation alsoincreases the levels of undesired trans-fatty acids.To enhance the production of oleic acid in soy-

180
bean seeds, the soybean FAD2 gene, encoding aspecific enzyme that inserts a second double bondat ∆12 into oleic acid (18:1 ∆9), has been silenced(Mazur et al., 1999). This increased oleic acidfrom 25% of total seed oil in the wild type to 85%in the transgenic plants. The transgenic linesshowed unchanged agronomic properties and arean example of the power of genetic engineering toalter seed oil content and composition withoutaltering plant performance in the field.
IV. IMPROVING THE CONTENT ANDAVAILABILITY OF ESSENTIALMINERALS
A. Phosphorous and Complexed Metals
A large proportion of the nutritionally impor-tant minerals in seeds are associated with thechelating agent phytic acid, which renders themunavailable to humans and livestock due to thelow solubility of the complexes. The minerals arezinc, iron, magnesium, and possibly calcium, aswell as the phosphorous, which is an integral partof the phytic acid molecule. The most criticalantinutritional effect of phytic acid is related tophosphorous availability. As much as 50 to 80%of the total seed phosphorous is not utilized andexcreted in the manure (Reddy et al., 1989). Fur-thermore, it has been shown that phytic acid mayalso interfere with zinc absorption in humans,weanling swine and rats (Couzy et al., 1993; Leiet al., 1993; Zhou et al., 1992), as well as iron andmagnesium absorption in humans (Brink andBeynen, 1992; Hurrell et al., 1992). The possiblechelating effect of phytic acid on the mineralcation, calcium, is less conclusive (Mitchell andEdwards, 1996), although at least one study(Jongbloed and Kemme, 1994) suggests phyticacid may increase retention of dietary calcium inswine.
1. Low Phytic Acid Plant Mutants
An alternative approach to improve phospho-rous availability is by classic genetic means. Sev-eral low phytic acid (lpa) mutants have been iso-
lated in cereals and legumes (Ertl et al., 1998;Larson et al., 2000; Larson et al., 1998; Raboy,1998; Raboy et al., 2000; Raboy et al., 1998;Wilcox et al., 2000). Seeds produced by “lowphytic acid” crops have total phosphorous levelssimilar to standard crops, but greatly reduced lev-els of phytic acid phosphorous (Raboy, 1997;Raboy, 1998). When monogastric animals con-sume “low phytic acid” grain, they absorb a muchlarger fraction of grain phosphorous than whenthey consume standard grain, and excrete propor-tionally less phosphorous (Ertl et al., 1998; Huffet al., 1998; Sugiura et al., 1999).
Utilizing paper electrophoresis and colorimet-ric screening methods, chemical mutagenesis of amaize synthetic Flint/Dent population, referred toas “Early ACR”, gave rise to two phenotypicclasses of mutants (Raboy, 1997; Raboy andGerbasi, 1996; Raboy et al., 2000). In low phyticacid 1 (lpa-1) mutants, the decrease in phytic acidphosphorous is matched by an increase in inor-ganic P, the sum of which remains constant. Noother large or obvious change in seed phospho-rous chemistry is observed. In low phytic acid 2(lpa-2) mutants, however, only ~75% of the re-duction in phytic acid phosphorous is matched byan increase in inorganic P. The remainder of theP is present in intermediate inositol phosphates.The molecular nature of the lpa-1 and lpa-2 mu-tants in maize is still unknown. However, it istempting to hypothesize that lpa-1 mutants areperturbed in myo-inositol metabolism (an earlystep in the pathway to phytic acid), while the lpa-2 mutants are perturbed in myo-inositol phos-phate metabolism (later steps in the pathway).
The lpa-1 mutant has now been crossed to alarge number of standard inbred lines and anincreasing numbers of hybrids have been synthe-sized. To date, lpa-1’s effect on seed phospho-rous fractions appears stable across genetic back-grounds. In field trials of the first 14 near-isogenichybrid pairs, each pair consisting of homozygouswild-type and homozygous lpa-1 iso-hybrids, aneffect on yield similar in extent was observed forseed dry weight as mentioned above (Ertl et al.,1998). A significant component of the yield lossappears to be an effect of homozygosity for lpa-1 on seed dry weight. The immediate issue with“low phytic acid/high available P” maize is lower

181
yield, but stress response, disease susceptibility,and storage problems still need to be addressed.
Mutants of both the lpa-1 and lpa-2 pheno-typic classes have also been isolated and mappedin barley (Larson et al., 1998), an lpa-1-like mu-tant has been isolated and mapped in rice (Larsonet al., 2000), and an lpa mutant was identifiedrecently in soybean (Wilcox et al., 2000). Thegenes encoding the enzyme D-myo-inositol3-monophosphate synthase (MIPS) in maize, bar-ley, and rice have been cloned (Larson and Raboy,1999; Larson et al., 2000). The isolation of simi-lar mutants in these three cereals, and the isola-tion of MIPS genes, represent the first phase in acomparative genomics approach. MIPS catalyzesthe conversion of glucose 6-P to L-myo-inositol1-P, is the only known source of the inositol ring(Loewus, 1990), and is a critical step in pathwaysbeginning with inositol. In maize there are mul-tiple MIPS sequences and one maps to the samesite on chromosome 1S as lpa-1. It is assumedthat in maize, lpa-1 is a MIPS mutant. MIPS is asingle-copy gene in barley and rice and maps tosites not linked to barley and rice lpa-1 loci (Larsonet al., 2000; Larson et al., 1998), so the nature ofthis mutation in these crops is still unclear.
The lpa mutations in a number of crops arebeing tested in animal feeding trials. These trialswill evaluate whether the lpa types will save phos-phorous supplements, reduce manure phospho-rous contamination, and increase mineral absorp-tion, especially in human societies where maizeserves as the staple food. Organoleptic analysis oflpa-1 sweet corn had shown no significant effectof lpa-1 on flavor (Tadmor et al., 2001). Feedtrials had been conducted with poultry (Douglaset al., 2000; Ertl et al., 1998; Li et al., 2000;Waldroup et al., 2000; Yan et al., 2000), swine(Spencer et al., 2000), and rainbow trout (Sugiuraet al., 1999). These studies demonstrated that theapparent availability of phosphorous in lpa grainswas higher than that in ordinary grains and thatthe fecal phosphorous content was significantlydecreased. In a study conducted at the MontanaState University, heifers were fed with hay pre-pared from normal and three lpa barley varieties.The average daily gain of the heifers fed with thelpa barley was 20% higher (p<0.001) than that ofthose fed with normal barley (V. Raboy, unpub-
lished results). These preliminary results indicatethat lpa grains may benefit ruminant animals too.
Mineral (iron, zinc, and calcium) absorptionalso increased when animals were fed with lpagrains (Li et al., 2000; Sugiura et al., 1999). Brownand associates (Mendoza et al., 1998) measurediron absorption from tortillas prepared with lpacorn vs. wild-type corn. They concluded that theconsumption of lpa strains of maize might im-prove iron absorption in human populations thatconsume maize-based diets. In a recent pilot studythat analyzed the effect of lpa-1 corn on zincabsorption, a comparison of the fractional absorp-tion of Zn (FAZ) between individuals consumingnormal and lpa-1 corn was conducted(M. Hambridge, unpublished results). FAZ wasconsistently and significantly greater on the lpacorn diet. An average FAZ from polenta preparedfrom lpa-1 corn was 78% greater than polentafrom normal corn. This increase in FAZ is ofsufficient magnitude to suggest that substitutionof lpa-1 in diets in which corn is a major staplewill have a beneficial impact on Zn bioavailability.
The next generation of lpa types will have afurther reduction in phytic acid and a yield similarto that of the normal varieties. This will beachieved by selecting for better mutants or bygenetic engineering of the seed phytic acid me-tabolism.
2. Utilization of Phytase to BreakdownPhytic Acid
In contrast to the situation in vertebrate mono-gastric metabolism, phytic acid complexes can bebiodegraded by a number of bacteria and fungi. Thisdegradation is catalyzed by an enzyme termedphytase. Indeed, the utilization of natural or recom-binant phytases has provided an important solutionto the antinutritional characteristics of phytic acid.Supplementation of animal diets with industriallyproduced phytase, extracted mainly from fungi(Shmeleva et al., 2000), or recombinant phytasesproduced in bacteria (Sunitha et al., 2000; Yo et al.,1999), was shown to increase animals phosphorousuptake by up to 42% (Lei and Stahl, 2000) (http://www.dfrc.ars.usda.gov/Research_Summaries/RS98_pdfs/wwwpp19-20.pdf).

182
An additional way to increase phosphorousavailability is by overexpressing phytase genes intransgenic plants. Pen and associates (Pen et al.,1993) overexpressed the Aspergillus niger phytasegenes, fused to a signal peptide of tobacco PR-Sprotein (in order to direct it to the apoplasm forincreased protein stability) under the control ofCaMV 35S promoter, in transgenic tobacco. TheA. niger phytase was stable and accumulated upto 1% of the total soluble protein in seeds. Phytaseactivity in the transgenic plants was found to bestable for up to 1 year of storage. In vitro experi-ments, which simulate the digestive tract of poul-try, showed that the addition of milled transgenicphytase seeds resulted in release of inorganicphosphate. Furthermore, the feeding of youngchickens showed that addition of either milledtransgenic seeds, or industrially produced A. nigerphytase, or inorganic phosphate had a compa-rable effects on growth rate of the animals (Pen etal., 1993). The transgenic phytase was extremelystable in the transgenic tobacco leaves and accu-mulated, in the extracellular fluid, at up to 14.4%of total soluble proteins in mature leaves(Verwoerd et al., 1995). The gene for A. nigerphytase was also introduced into transgenic al-falfa plants (http://www.dfrc.ars.usda.gov/Research_Summaries/RS97_pdfs/FH3.pdf).Phytase concentration in the best performingtransgenic lines ranged from 0.85 to 1.8% of totalsoluble protein. The transgenic alfalfa plants werevegetatively propagated to produce about 7500plants for a field test (http://www.dfrc.ars.usda.gov/Research_Summaries/RS98_pdfs/wwwpp19-20.pdf). The results indicated that economicallysignificant bioavailable phosphorous was presentin the transgenic alfalfa in its second year in fieldplots. Feeding trials with chickens and swine in-dicates that phytase-overexpressing transgenicalfalfa does not require inorganic phosphorous supple-mentation in the feeds (http://www.dfrc.ars.usda.gov/Research_Summaries/RS98_pdfs/wwwpp21-22.pdf).Similar results in improving phosphorous utiliza-tion were also reported in feeding experimentsutilizing soybean seeds transformed with theA. niger gene (Denbow et al., 1998). In addition,A. niger phytase was found to be stable in soy-bean cell-suspension culture (Li et al., 1997).Because some plant feeds for livestock are pro-
cessed at high temperatures, the stability of therecombinant phytase to such processing tempera-tures is extremely important. Indeed, the recom-binant A. niger phytase appears not to be stableenough to withstand the elevated temperaturesinvolved in soybean processing. This might besolved either by using a yeast phytase gene that isstable at 80oC (Nakamura et al., 2000) or bytransforming the A. niger phytase gene into lowtrypsin inhibitor lines that can be used withoutheat processing (Clarke and Wiseman, 2000).
Expression of phytase genes has not beenrestricted to dicot plants. Two A. niger phytasegene constructs were introduced into transgenicwheat under the control of the constitutiveubiquitin-1 promoter (Brinch-Pedersen et al.,2000). To ensure protein stability, the phytasegene, in one construct, was fused to an α-amy-lase signal peptide. The second construct wassimilar to that of first, but lacked the signalpeptide. An immunoreacting polypeptide of thesize expected for the A. niger phytase was de-tected in both seed and leaf tissues, but not inthose of the embryo. The heterologous phytasewas exclusively present in the pericarp-seed coat-aleurone fraction up to 25 days after pollination,and thereafter it accumulated in the endosperm.The secreted and nonsecreted phytases providedaround 4-fold and 1.6-fold increase phytase ac-tivity, compared with control nontransformedplants. The authors concluded that a functionalA. niger phytase can be produced in significantamounts in wheat grains that could be used toimprove the nutritional quality of monogastricanimal diets.
Animal feeding trials were conducted tocompare the efficacy of genetically engineeredmicrobial and plant phytases for enhancing theutilization of phytic acid-bound phosphorous incorn-soybean meal-based diets fed to youngbroilers (Zhang et al., 2000). The addition ofboth sources of phytase resulted in similar in-creases (P < 0.05) of body weight gain; feedintake; gain:feed; apparent retention of drymatter, phosphorous, and calcium; and toe ashmeasurements. Phosphorous excretion de-creased as phytase addition increased. No sig-nificant abnormalities were seen in any of the40 broilers necropsies.

183
B. Iron
Nearly 30% of the world population suffersfrom iron deficiency (WHO, 1992) and it is moreprevalent in developing countries where plant-based diets are common (Craig, 1994). Iron con-tent is limited in most major crops. Moreover,even in crops that are rich in iron, such as spinachand legumes, iron is complexed with phytic andoxalic acids and therefore is inefficiently absorbedby humans. However, phytic and oxalic acids arenot the only storage forms of iron. In animals,plants, and bacteria, iron is also stored in ferritins,a family of iron storage proteins (Theil, 1987).The bioavailability of iron to mammals appears tobe efficient when it is provided as an iron-ferritincomplex (Beard et al., 1996; Theil et al., 1997).
In attempts to increase iron availability inplant-based diets, Goto and associates (Goto etal., 1999) transformed rice with a soybean ferritingene, under the control of an endosperm-specificgebe promoter. This resulted in the stable accu-mulation of the soybean ferritin in seeds of thetransgenic rice and up to a threefold increase inseed iron content. A meal-size portion of such aferritin-fortified rice is predicted to provide 30 to50% of the daily adult iron requirement (Goto etal., 1999). Expression of a recombinant ferritingene may be only a partial solution for iron for-tification of plant foods because other factors,such as iron transport efficiency to plant seedsand its association with phytic acid complexesmay limit level and availability. Indeed, Potrykusand associates have transformed rice with threegenes encoding a French bean ferritin, a fungalheat-stable phytase and a rice metallothionin-likeprotein (a protein that helps iron absorption in thehuman digestive tract) (Gura, 1999). Suchtransgenic plants could help solve iron deficiencyin humans. Unfortunately, because commerciallygrown indica rice strains are very difficult totransform, most of the studies on rice used thejaponica strains. Thus, these genes will have to betransferred from the “japonica” into the “indica”strains, which can be performed by classic breed-ing.
Although the utilization of ferritin to produceiron-fortified plant foods looks promising, there areseveral physiological and safety issues to be consid-
ered. Iron availability depends not only on its stor-age, but also on its absorption from the soil andtransport within the plant (Grusak and DellaPenna,1999). Moreover, iron uptake in many plants occursvia transporters that may not be entirely iron-spe-cific. Briat and associates (Van Wuytswinkel et al.,1999) constitutively overexpressed a soybean fer-ritin gene in transgenic tobacco plants using theCaMV 35S promoter. The expression of the ferritingene not only increased leaf iron content, but alsoactivated iron transport systems as indicated by anincrease in root ferric reductase activity. The expres-sion of ferritin genes could result in accumulation oftoxic metals in plants (Briat, 1999). Thus, the min-eral content of transgenic plants, expressing a re-combinant ferritin gene, should be thoroughly ex-amined for mineral toxicity before they are released.The expression of the transgenic ferritin gene spe-cifically in the seed, as was reported by Gotto et al.(1999), may overcome problems of excess accumu-lation of toxic metals.
V. PLANT PRODUCTS WITH IMPROVEDQUALITY OBTAINED BY REDIRECTINGSECONDARY METABOLISM
A. Vitamins and Neutraceuticals
1. The Terpenoid Pathway, Carotenoids,Vitamins A and E
Many important metabolites are synthesized inplants at least partially via the terpenoid pathway.They include the phytol chain found in chlorophyll,as well as plant growth regulators such as gibberel-lins, abscisic acid, and cytokinins; accessory photo-synthetic pigments, chromophores, and vitamins suchas carotenoids, and tocopherols (Figure 3) (Croteauet al., 2000). Using metabolic engineering, the caro-tenoid pathway has been modified not only to pro-duce valuable compounds and pigments (Hirschberg,1999; Mann et al., 2000), but also to enhance thenutritional value and quality of foods. Using geneshuffling and recombinant genes, novel carotenoidshave been produced in bacteria, illustrating the po-tential of engineering the terpenoid pathway to pro-duce unique carotenoids (Albrecht et al., 2000;Schmidt-Dannert et al., 2000). Nonetheless, direct
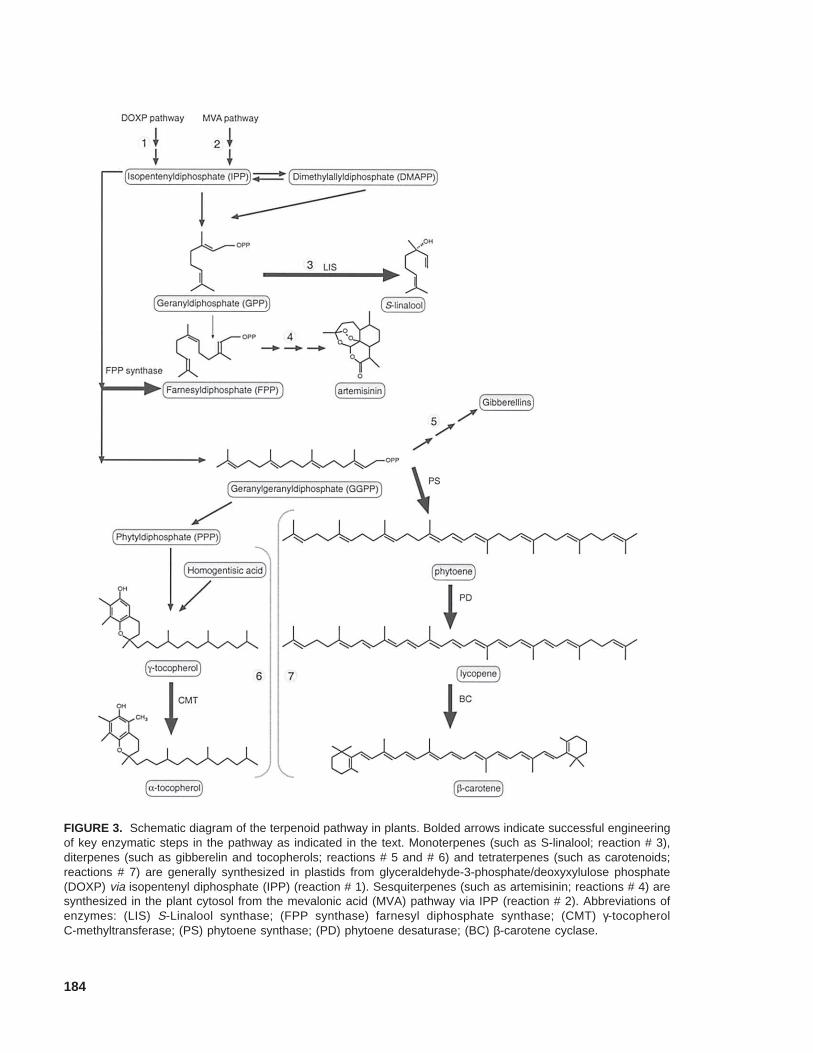
184
FIGURE 3. Schematic diagram of the terpenoid pathway in plants. Bolded arrows indicate successful engineeringof key enzymatic steps in the pathway as indicated in the text. Monoterpenes (such as S-linalool; reaction # 3),diterpenes (such as gibberelin and tocopherols; reactions # 5 and # 6) and tetraterpenes (such as carotenoids;reactions # 7) are generally synthesized in plastids from glyceraldehyde-3-phosphate/deoxyxylulose phosphate(DOXP) via isopentenyl diphosphate (IPP) (reaction # 1). Sesquiterpenes (such as artemisinin; reactions # 4) aresynthesized in the plant cytosol from the mevalonic acid (MVA) pathway via IPP (reaction # 2). Abbreviations ofenzymes: (LIS) S-Linalool synthase; (FPP synthase) farnesyl diphosphate synthase; (CMT) γ-tocopherolC-methyltransferase; (PS) phytoene synthase; (PD) phytoene desaturase; (BC) β-carotene cyclase.

185
commercial application of these results is not trivial.Constitutive manipulation of the terpenoid pathwayin plants might have undesired results. Attempts toincrease carotenoid levels by overexpressingphytoene-synthase in tomato plants, utilizing theCaMV 35S promoter, resulted in dwarf phenotypesdue to a reduction of key diterpene derivatives suchas gibberellic acid, and an accompanied reduction ofchlorophyll levels, presumably due to lack of phytol(Figure 3, reaction # 5) (Fray, 1995). The carefulselection of terpenoid pathway genes and specificpromoters were more successful (see below), show-ing that it should be possible to manipulate terpe-noids to improve the nutritional quality of foods.
a. Carotenoids and Vitamin A
Carotenoids are tetraterpene pigments, essen-tial in photosynthesis, but often accumulating innonphotosynthetic tissues at high levels, impart-ing color and antioxidant properties to fruits. Pro-vitamin A (β-carotene and other cyclic caro-tenoids) is converted into retinol (vitamin A) inhumans. Vitamin A deficiency is one of the lead-ing causes of night blindness in humans and hasalso been correlated with increased occurrence ofseveral diseases such as diarrhea, respiratory ail-ments, tuberculosis, malaria, and ear infections.According to the World Health Organization(WHO), around 2.8 million children under 5 yearsof age currently exhibit a severe clinical manifes-tation of vitamin A deficiency known as xe-rophthalmia (Humphrey et al., 1992).
Our knowledge in the biosynthesis of terpe-noids has been applied to the production of plantfoods rich in lycopene and provitamin A. Lyco-pene is a noncyclic tetraterpene, and normally aprecursor of other carotenoids such as the carotenesand xanthophylls (Figure 3), but often accumulat-ing in fruits such as tomatoes, papayas, and wa-termelons (Van den Berg et al., 2000). Besidesthe utilization of genes introgressed from a wildrelative, marker-assisted breeding has been usedto obtain high-lycopene tomatoes (Chen et al.,1999). Additionally, using genetic engineering,the manipulation of the carotenoid pathway hasresulted in plant products enriched in provitaminA at the expense of the pigment lycopene (Romer,
2000). Conversely, the inhibition of the phytoenesynthase gene in tomato has resulted in decreasedcarotene and xanthophyll levels (Fraser et al.,1995).
The manipulation of the carotenoid pathwayhas also been accomplished in rice, a major worldfood source. Rice contains poor levels of β-caro-tene in the endosperm, which is the major tissueconsumed as food after mechanical processing ofthe grain. Immature rice endosperm is able tosynthesize the carotenoid precursor geranylgeranyldiphosphate (Figure 3), but normally lacks caro-tenoids. The accumulation of the noncolored caro-tenoid precursor phytoene (Figure 3) occurs intransgenic rice plants expressing a daffodil (Nar-cissus pseudonarcissus) phytoene synthase geneunder the control of an endosperm-specific pro-moter (Burkhardt et al., 1997). By combining thisgene with genes encoding a bacterial phytoenedesaturase and daffodil lycopene β-cyclase,β-carotene was produced in the endosperm, yield-ing the so-called “Golden Rice” (Figure 3, reac-tions #7) (Ye, 2000). Some transgenic rice geno-types also accumulated substantial levels ofxanthophylls, such as lutein and zeaxanthin, prob-ably due to endogenous activities of cyclases andhydroxylases in the endosperm tissue (Ye, 2000).As for ferritin (see Section V.B), the rice speciesjaponica was used for this proof of concept ex-periment. The next step will be to transfer thesegenes into the indica rice varieties, the speciesgrown in Asia.
Because the precursor of carotenoids,geranylgeranyl diphosphate, is ubiquitous andoften abundant in many plant tissues, this genetechnology is promising for carotenoid-rich plantproducts with enhanced or modified color andnutritional value. For example, transgenic rape-seed producing high α- and β-carotene levels havebeen produced using a bacterial phytoene syn-thase gene fused to a seed-specific promoter(Shewmaker et al., 1999).
b. Vitamin E
Vitamin E was discovered 75 years ago as afat-soluble dietary factor effective in preventingfetal death (Combs, 1992). Although vitamin E is

186
the generic descriptor for all tocopherols that quali-tatively exhibit the biological activity of α-toco-pherol, other tocopherols also have vitamin Eactivity. Still, the most active tocopherol isα-tocopherol (Combs, 1992; Lambert, 1994;Traber and Sies, 1996), produced only by plantsand is most concentrated in plant oils, especiallywheat germ oil (Combs, 1992). Vitamin E defi-ciency not only causes fetal death, but also sev-eral other disorders, such as mammalian infertil-ity, kidney and liver damage, cardiovasculardiseases, and cancer (Combs, 1992; Dowd andZheng, 1995; Sies and Stahl, 1995; Stampfer etal., 1993). Tocopherols are antioxidants that pre-vent the autooxidation of highly unsaturated fattyacids mediated by molecular oxygen. Thus, oneof the roles of vitamin E in humans may be thepreservation of membranes from oxidative dam-age (Burton and Ignold, 1981; Combs, 1992; Erinet al., 1985).
Seeds normally contain γ-tocopherol, but notα-tocopherol. The gene encoding the γ-toco-pherol specific C-methyltransferase, an enzymethat converts γ-tocopherol to α-tocopherol by asingle methylation, is not highly expressed inseeds. High α-tocopherol crop plants have beenproduced by classic breeding programs (Galliheret al., 1985; Hallauer and Miranda, 1988). Im-proving α-tocopherol production in seeds bygenetic engineering was achieved when theArabidopsis gene encoding γ-tocopherol specificC-methyltransferase was cloned in an elegantseries of experiments (Shintani and DellaPenna,1998). This genomics-based approach is de-scribed in detail later (see Section VII.B).Constitutive overexpression of the γ-tocopherolC-methyltransferase gene in transgenicArabidopsis caused a significant conversion ofγ-tocopherol to α-tocopherol in the seeds (Fig-ure 3, reactions # 6) (Shintani and DellaPenna,1998). It is highly likely that this transgenicapproach will be applicable to many other seeds.This approach, however, does not cause an in-crease in the levels of total tocopherols, but onlyconverts most of the γ-tocopherol already presentinto α-tocopherol. Nevertheless, with the adventof genes that control total tocopherol content, itmay be possible to obtain food products withincreased total tocopherol levels.
B. Phenolic Compounds, Stilbenes andPhytoestrogens
1. Resveratrol
Several plants, including grapevine, pine, andpeanuts, produce the stilbene-type phytoalexinresveratrol when attacked by pathogens. Thiscompound appears to be one of the health-pro-moting factors of grapevine that are associatedwith reduced risk of heart diseases (popularlyknown as “The French Paradox”) and long rec-ognized by folklore medicine. Clinical studieshave demonstrated the beneficial effect ofresveratrol, isolated from red wine, on cardio-vascular disease and confirmed the involvementof resveratrol in fighting arteriosclerosis andvascular tissue diseases (Das et al., 1999;Pendurthi et al., 1999; Zou et al., 1999).Resveratrol has also been shown to inhibit cellu-lar processes associated with tumor initiation,promotion, and progression (Mgbonyebi et al.,1998; Park et al., 2001).
Resveratrol is synthesized from the ubiqui-tous precursors malonyl CoA and coumaryl CoAby stilbene synthase. The gene encoding thisenzyme was cloned from grapevine and intro-duced into tobacco (Hain et al., 1993). Due tothe availability of malonyl CoA and coumaryl-CoA, resveratrol was readily accumulated afterinduction of the transgenic tissues, rendering thetransgenic plants more resistant to fungal attackthan the nontransgenic controls (Hain et al.,1993). Thus, the overexpression of only one geneled to the diversion of the existing metabolicflow to the production of a novel metabolite(Gustine, 1995; Hain et al., 1993). Using thisapproach, the stilbene synthase gene thereforecould be used to produce resveratrol in foodsalready associated with anticancer properties, orto create “functional foods” with health benefits.The metabolic engineering for the production ofa phytoalexin, originally intended to introducefungal resistance into plants, could lead to theproduction of functional foods. Moreover, be-cause resveratrol can be generated in grape cellsuspension cultures, it may also be possible toproduce resveratrol to be marketed as a foodsupplement.

187
2. Flavonoids
Many members of the Fabaceae accumulate anumber isoflavonoid compounds, such as theisoflavones genistein and daidzein, as well as theirglycosides that exist in soybeans (Jung et al., 2000).Several health benefits have been assigned to thesecompounds, at times referred to as phytoestrogens.Phytoestrogens are associated with relief of meno-pausal symptoms, reduction of osteoporosis, im-provement of blood cholesterol levels, and lower-ing the risk of certain hormone-related cancers,and coronary heart disease (see Dixon and Steele,1999). The biochemical basis of these effects hasnot been fully established, but the weak estrogenicactivity of isoflavones may be a factor conferringthese properties. The potential for metabolic engi-neering of the isoflavonoid pathway has been rec-ognized (Dixon and Steele, 1999).
Isoflavones are synthesized by a branch of thephenylpropanoid pathway and normally play a rolein plant defense against fungal attacks (Dixon andPaiva, 1995). They also help to establish a symbioticassociation between legumes and nitrogen fixingrhizobial bacteria (Dixon and Paiva, 1995). Thebranching of the flavonoid metabolic pathway toisoflavones occurs by the action of the enzymeisoflavone synthase. Isoflavone synthase, a memberof the cytochrome P450 family, oxidizes the fla-vanone intermediates naringenin and liquiritigenininto genistein and daidzein, respectively (Jung et al.,2000). Naringenin is synthesized by most plants asan intermediate to other flavonoids, such as thecommon anthocyanin pigments (Croteau et al., 2000).Overexpression of the soybean isoflavone synthasegene in transgenic Arabidopsis, tobacco, and maizeplants, which naturally do not produce isoflavones,resulted in the production of genistein and its deriva-tives, possibly through the conversion of endogenousnaringenin (Yu et al., 2000). These results prove thatby metabolic engineering, it is possible to producehealth-associated isoflavones in nonlegume plants.
C. Improving the Flavor and Aroma ofPlant Foods
The aromas of fruits, vegetables, and otherfoods are due to the mixtures of volatile metabo-
lites. The different proportions of the volatile com-ponents, their thresholds for perception by human’snose, and the presence or absence of trace compo-nents often determine aroma properties (Thomson,1987). Breeding programs of fruits and vegetableshave been focused traditionally on desirableagronomical characteristics, such as yield andresistance to environmental stresses, pests, andpathogens (Stevens and Rick, 1986). Breeding forimproved fruit flavor was mainly directed towardcontrolling sugar/acid ratios and improving tex-ture and storage characteristics (Jones and Scott,1983; Stevens and Rick, 1986). Conventionalbreeding to improve the aromas of agriculturalproducts is often impeded by the large number ofgenes involved, the significant environmental anddevelopmental effects, and the lack of simple andcheap methodologies to probe both aroma prefer-ences of the public and the complex chemistry.
1. Modification of the Early Steps of theTerpenoid Pathway
Monoterpenes are key determinants of thearomas of many aromatic plants, vegetables, andfruits. Therefore, the potential of genetic engi-neering to modify the early steps of the terpe-noid pathway in order to modify aroma has beennoted (Haudenschild and Croteau, 1998;Lewinsohn, 1996). Linalool is an acyclic monot-erpene alcohol that imparts an aroma with asweet floral alcoholic note. Linalool is a majorcomponent of the scent of many flowers (Dob-son, 1993; Knudsen et al., 1993) and is alsopresent in many edible fruits, such as guava,peach, plum, pineapple, and passionfruit(Bernreuther and Schreier, 1991). Linalool is achiral compound, naturally appearing in twoforms (S- and R-linalool) that differ in theiraroma. The enzyme that catalyzes the formationof S-linalool from the ubiquitous precursorgeranyl diphosphate (Figure 3, reaction # 3) hasbeen purified (Pichersky et al., 1995), and itsgene (LIS) cloned from the flowers of a smallCalifornian annual plant Clarkia breweri(Dudareva et al., 1996). This gene is a promisingcandidate for future attempts to manipulatemonoterpene metabolism in transgenic plants.

188
Many modern tomato varieties have impairedaromas, as they lack many of the commonvolatiles, such as linalool, present in the oldertomato varieties. Metabolic engineering to modifythe aroma of tomato fruits has been describedrecently (Lewinsohn et al., 2001). The ClarkiaLIS gene under the control of the late-ripeningspecific promoter E8 has been transformed intotomatoes and this has resulted in fruits that pro-duce S-linalool (Figure 3, reaction #3). Unexpect-edly, the expression of LIS also caused the accu-mulation of 8-hydroxylinalool, a linaloolderivative possibly produced by allylic hydroxy-lation of the linalool via an unknown endogenousenzyme.
Notably, although only a small fraction of themetabolic flow through the terpenoid pathwaywas diverted into linalool in these transgenicplants, this was sufficient to change the aroma,because the threshold levels for the perception oflinalool are very low (6 ppb) (Buttery et al., 1971).In fact, many other volatiles also have very lowthresholds of detection (Buttery et al., 1971), andtherefore aroma enhancement may be relativelyeasily achieved by diverting only a small fractionof the metabolic flow to their production, withnegligible perturbation of the general metabolismof the plant. The potential of genetic engineeringfor the improvement of aroma and taste proper-ties of agricultural products is just beginning to beinvestigated. With the discovery of other genesencoding key enzymes involved in the productionof volatile aroma chemicals, the potential to uti-lize genetic engineering for the manipulation ofcrops is very promising.
2. Modification of Lipid-Derived Volatiles
Many of the volatiles that affect the aroma offresh produce are formed by degradation of lipids(Croteau and Karp, 1991). Therefore, anotherapproach to improve tomato fruit aroma was tomodify the oxidation pattern of the lipids that arenaturally degraded into aroma compounds.Overexpressing a yeast gene encoding ∆9
desaturase in transgenic tomato plants elevatedthe levels of saturated and unsaturated fatty acidsin the fruits. These changes were associated with
the increased production of flavor compoundsderived from the degradation of fatty acids, suchas cis-3-hexenol, 1-hexanol, hexanal, and cis-3-hexenal. These compounds impart a fresh aromasensation (Wang et al., 1996). In another study,the levels and ratios of short-chain aldehydes andalcohols were modified by the respective repres-sion and overexpression of a tomato alcohol de-hydrogenase gene in transgenic tomato fruits(Prestage et al., 1999; Speirs, 1998). As a result,minute changes in aroma were detected by tastepanelists.
D. Antinutritional Compounds
Many natural products are induced in plants asa result of fungal attacks and are often consideredimportant in plant protection. Some of these de-fense metabolites have beneficial human healthproperties (see Section VI.B.1), while others havetoxic effects. It is usually recommended that in-fected plant products not be consumed. This is notonly due to the presence of mycotoxins producedby pathogens, but also to the possible presence ofplant-derived compounds that are toxic to humans(Kuc, 1995). Plants bred for pest resistance, byincorporating genes from wild relatives, must betested carefully to avoid the inclusion of toxic traits(Kuc, 1995). Another important antinutritional agentis lignin, which is not discussed in the presentreview because it has been discussed in a numberof recent reviews (Baucher et al., 1998; Grima-Pettenati and Goffner, 1999).
1. Furanocoumarins
The furanocoumarins psoralen, bergapten, andxanthotoxin are found in many food crops of theApiaceae, Rutaceae, and Moraceae, including cel-ery, parsnip, parsley, citrus, and figs (Beier andNigg, 1992). These metabolites have antimicrobialand insecticidal properties but are also potent pho-tosensitizing toxins for humans. They cause severedermatitis, blistering, and other serious damage tothe skin in the presence of UV light or solar radia-tion (Beier and Nigg, 1992). At low doses,furanocoumarins can have therapeutical and cos-

189
metic value, and they are sometimes added to sunlotions as photosensitizers. Furanocoumarins arealso used to treat psoriasis, vitiligo, and other skindisorders.
Fungal infections and other stress conditionsduring the growing season can markedly increaseplant furanocoumarin levels. Because of theirantimicrobial and insecticidal activities, plantbreeders developed lines of celery with highpsoralen levels (Kuc, 1995). These lines had to beremoved from the market when toxicological dataconcerning psoralens became widely available andaccepted. Well-intentioned efforts to protect plantsby increasing their natural defense compoundshad serious health repercussions.
2. Nitrogen Compounds: Alkaloids andGlucosinolates
a. Caffeine
Caffeine (trimethylxanthine) is an odorless,bitter purine alkaloid. The most common caffeinecontaining plants are tea (Thea sinensisfam. Theaceae) and coffee (Coffea spp. Fam.Rubiaceae), but it also occurs in other unrelatedfamilies (Samuelsson, 1992). The average cup ofcoffee contains about 100 mg of caffeine. Caf-feine has a stimulating effect on the central ner-vous system, heart, blood vessels, and kidneys.Its potent stimulatory action makes it a valuableantidote to respiratory depression induced by drugoverdose (e.g., from morphine or barbiturates).People who use caffeine can show improved motorperformance, decreased fatigue, enhanced sen-sory activity, and increased alertness. This maypartly explain the compulsion of many adults toconsume coffee or other caffeine-containing bev-erages as part of their “morning ritual”. Caffeineintake can also cause irritability, nervousness oranxiety, jitteriness, headaches, and insomnia. Bythe mid-1980s decaffeinated coffee and soft drinkshad become widely available, giving consumersthe choice of regulating their caffeine intake.Caffeine is removed from the coffee beans. Caf-feine is synthesized from the pool of free purinesavailable for nucleic acid biosynthesis throughthe action of a 7-N-methyltransferase that con-
verts xanthosine to form 7-methyl xanthosine.After hydrolysis of the ribose moiety, 7-methylxanthine is formed, which after two se-quential N-methylations give rise to theobromine(the major alkaloid of cacao beans), and caffeine(Waldhauser et al., 1997). The gene for xanthosine-N7-methyltransferase, the first enzyme in this path-way, was silenced by antisense, resulting in virtu-ally caffeine-free transgenic coffee plants (http://www.nbiap.vt.edu/biomon/datacat.htm). These“naturally” caffeine-free products do not requiredecaffeination. Field experiments designed to testthe performance of these plants are currently un-der way. One could envision similar experimentsto lower caffeine content in other plants.
b. Glucosinolates
Glucosinolates, also called mustard oil glyco-sides, are anionic thioglucosides normally associ-ated with plant defense due to their insecticidal andantibacterial properties. They occur in many plantfamilies, but they are particularly common in theBrassicaceae. Plants that contain glucosinolates alsocontain thioglucosidase (myrosinase) enzymes thatcleave the sugar moiety and release the free agly-cones. The free aglycones are nonstable and de-compose into sulfur-containing volatiles that im-part a typical pungent odor and biting taste. The hotflavor of mustard seed and horseradish, as well asthe more subtle flavors of rutabaga and cauliflower,are consequences of the presence of such com-pounds.
By the 1960s, rapeseed varieties were bred tocontain oil low in erucic acid, which is associatedwith myocardial toxicity. Rapeseed meal proteinhas a favorable balance of amino acids, but its usein rations was limited by its glucosinolate con-tent. Canadian breeders developed “double low”rapeseed: (low erucic acid, low glucosinolate con-tent), which has become a major oil crop.
Glucosinolates are derived from amino acidsvia aldoxime and thiohydroximic acid intermediates(Figure 4) (Luckner, 1990). Indole glucosinolatesstill present in rapeseed are derived from tryptophan.They adversely affect the quality of rapeseed oil andmeal. The pathway from tryptophan to glucosinolateshas been diverted in transgenic rapeseed by

190
overexpressing a gene encoding tryptophan decar-boxylase of Catharanthus roseus (Chavadej et al.,1994). This enzyme converts tryptophan totryptamine on the route to the synthesis of indolealkaloids in C. roseus (Figure 4). By reducing thepool of tryptophan available for the production ofglucosinolates, the metabolically engineered rape-seed seed accumulate only 3% of their original lev-els of indole glucosinolates (Figure 4) (Chavadej etal., 1994). Interestingly, the transgenic seeds alsodid not accumulate tryptamine, probably due to itsmobilization or further metabolism, also observed intransgenic tobacco overexpressing tryptophan de-carboxylase (Guillet et al., 2000). Additional ex-amples of modifications to the glucosinolate path-way through metabolic engineering are provided inthe following references: Bak et al., 2000; Bak et al.,1999; Wittstock and Halkier, 2000.
c. Steroid Glycoalkaloids
Steroid glycoalkaloids found in varioustissues of tomatoes and potatoes and are in-hibitors of choline esterases and can be fatalto humans and livestock (Beier and Nigg,1992; Kuc, 1995). Such compounds are par-ticularly enriched in potato vines, sprouts,
peels, or green and cull tubers, and severehuman illness and fatalities have resulted fromthe consumption of greened or sprouted pota-toes (Kuc, 1995). People generally believethat most of the potato’s nutrients are in thepeel, but they are unaware or ignore the factthat the peel also contains toxic steroidglycoalkaloids.
Breeding for pest-resistant potato plantsresulted in increased levels of steroidglycoalkaloids, later found to be toxic to hu-mans (Kuc, 1995). These resistant cultivarswere withdrawn from cultivation. The levelsof steroidal glucoalkaloids were reducedby expressing an antisense constructof a glucoyltransferase gene in transgenic po-tatoes (http://www.nbiap.vt.edu/biomon/datacat.htm). Because many of the toxic com-pounds in plants may protect them againstpathogens, lowering the toxins to improve foodand feed may also reduce the plants’ ability totolerate biotic stresses. The solution may be acompromise that biotechnological tools canprovide: careful selection of tissue-specificgene promoters may direct the metabolicchanges only to tissues that are consumed asfoods with minimal effects on the rest of theplant.
FIGURE 4. Diversion of the synthesis of undesired tryptophan-derived glucosinolates in Rapeseed. The enzymetryptophan decarboxylase (TDC), which was introduced from Catharanthus roseus to Rapeseed (see text) isillustrated in a bolded arrow.

191
E. Metabolic Engineering of MedicinalAgents in Plants
Some of the terpenoid pathway metabolites,particularly the sesqui- and diterpenes, possessimportant medicinal properties. Novel sesquiter-penes have been produced in transgenic tobaccoplants and cell cultures, as well as in bacteria byoverexpressing heterologous sesquiterpene syn-thase genes. This illustrates the potential for themanipulation of the terpenoid pathway to im-prove the medicinal value of plant foods (Hohnand Ohrogge, 1991; Hohn and Plattner, 1989;Zook et al., 1996).
1. Artemisinin
Wormwood (Artemisia annua L., Asteraceae),known in China as qinghao, is an annual herb thatcontains artemisinin, a sesquiterpene lactone withan endoperoxide bridge (Figure 3). Artemisinin isavailable commercially in China and Vietnam asan antimalarial drug efficacious against drug-re-sistant strains of Plasmodium, the malarial para-site. Malaria is one of the most devastating dis-eases and affects over 200 million people annually.Artemisinin production by A. annua is usually inthe range of 0.01 to 0.4%, but some varieties canproduce over 1% DW (Bruneton, 1995).Artemisinin is formed by a series of steps initi-ated by cyclization of farnesyl diphosphate.Overexpression of a cotton gene encoding farnesyldiphosphate synthase (the enzyme catalyzingfarnesyl diphosphate synthesis) in transgenicwormwood plants resulted in the increased pro-duction of artemisinin, probably due to an in-crease in the pool of available farnesyl diphos-phate (Figure 3, reactions #4) (Chen et al., 2000).
2. Tropane Alkaloids
Wild-type belladonna (or deadly nightshade,Atropa belladonna) accumulates the tropane al-kaloid hyoscyamine, which together with its race-mic form atropine acts as a nerve anticholinergicagent. Hyoscyamine and atropine are used as aremedy against spasms in skeletal muscles, and as
an antidote against poisonous cholinesterase in-hibitors (Samuelsson, 1992). Belladonna extractswere used in Medieval Europe to dilate pupils,and hence the name belladonna, “beautifulwoman” in Italian (Kutchan, 1995). Theepoxidated-derivative scopolamine is also an an-ticholinergic agent, but, unlike hyoscyamine, it isprimarily a depressant of the central nervous sys-tem (Samuelsson, 1992). Scopolamine is usedprophylactically against motion sickness and, of-ten, in combination with morphine, for premedi-cation before surgery. Scopolamine is synthesizedfrom hyoscyamine by the action of an hyos-cyamine 6-β-hydroxylase present in Hyoscyamusniger, a producer of both hyoscyamine and sco-polamine. The enzyme, a 2-oxoglutarate-depen-dent dioxygenase, has been characterized, and itsgene was cloned from cultured roots of H. niger(Matsuda et al., 1991). The gene was transferredto transgenic belladonna under the control of theCaMV 35S promoter. The transgenic belladonna,accumulated almost exclusively scopolamine intheir aerial parts (Yun et al., 1992), exemplifyingthe vast potential prominent in the further imple-mentation of recombinant DNA technology intoother medicinal crops.
VI. ADVANCED GENETIC ANDGENOMIC APPROACHES
A. Advanced Breeding Approaches
The rapid development of molecular tools isexpected to speed up classic breeding. A richgene pool exists in the wild relatives of the culti-vated crops. However, the assessment of the po-tential of the wild gene pools is not easy becauseit cannot be easily predicted how these genes willbehave when introgressed into the cultivated spe-cies. An elegant solution to this problem is theproduction of pools of near isogenic cultivatedlines (NILs) of a given plant species, each con-taining a small introgressed chromosomal seg-ment from wild relatives of that species. In theseNILs, the wild genes function in a genetic back-ground that is mostly derived from the cultivatedplants, and hence the agronomic quality of spe-cific wild genes can be assessed quite accurately.

192
The potential of this approach has been demon-strated recently in two populations of NILs of thetomato (Lycopersicon esculentum) cultivars, con-taining segments derived from the wild relativesL. pennellii and L. hirsutum (Eshed and Zamir,1995; Monforte and Tanksley, 2000). In the NILscontaining the L. pennellii, two important traitloci related to food value were identified, andtheir genes isolated by map based cloning. One ofthese wild trait loci encodes an invertase allelethat significantly increases the total soluble solids(TSS or brix; mainly sugars and acids) (Fridmanet al., 2000). The second wild trait locus controlsa novel fruit and flower-specific lycopene β-cy-clase, which provides an alternative pathway forthe synthesis of β-carotene (provitamin A) (Ronenet al., 2000). This gene, once identified, can bymanipulated to increase β-carotene content in fruitsor to increase the level of lycopene (another im-portant carotenoid) by eliminating its activity intransgenic tomato (Ronen et al., 2000). Becausethese wild trait loci were identified genetically,their introduction into leading commercial tomatovarieties should be straightforward.
The production of NIL is a long and difficultprocess. Tanksley and Nelson (Tanksley andNelson, 1996) suggested an alternative to thisprocedure termed “advanced backcross qualitytrait locus analysis” (AB-QTL). This method hasbeen used extensively to locate QTLs in tomatoand other crops. For example, alleles were trans-ferred from L. hirsutum to cultivated tomato re-sulting in a significant (15%) increase the TSSvalue (Bernacchi et al., 1998). AB-QTL was usedto transfer alleles from the wild species teosinte toelite cultivated maize, for increased kernel oil andprotein content, without compromising other ag-ronomic traits (Y. Tadmor and associates, unpub-lished).
B. Utilization of Genomic Approaches toImprove the Value of Plant Food andFeed
Although classic genetic approaches are andwill remain dominant in the future, it is expectedthat molecular methods of engineering metabolicpathways will become more common in breeding
programs aimed at increasing the value of foodsand feeds in the future. These approaches have sofar been mostly limited by the availability of thenecessary genes. This limitation has been over-come recently by the immense information on newgenes from the plant expressed sequence tag (EST)databases and genome sequencing programs. Ge-nomic approaches to improve the nutritional as-pects of plants (called “Nutritional Genomics”)have been reviewed before and several selectedpublications are cited here (Allona et al., 1998;DellaPenna, 1999b; Grusak, 1999; Lange et al.,2000; Murphy, 1999; Ohlrogge and Benning, 2000;Somerville and Somerville, 1999; Wang et al.,2000). Based on DNA and amino acid sequencehomologies and similarities (bioinformatics), andpatterns of expression of individual ESTs, predic-tions can be made about the biochemical roles ofnewly identified genes. New genes with predictedfunctions, derived from these genomic programs,are expected to be progressively incorporated intofuture plant breeding programs where they willhave a significant impact on increasing the value offoods and feeds.
The genome of Arabidopsis has already beensequenced, and the rice genome is nearly com-plete. In addition, EST (cDNA) databases areavailable from various tissues of many plants aswell as from different tissues subjected to varioushormone and stress treatments. Conserved genesand cDNA sequences from these databases, forwhich clear functional annotation can be predictedby the current bioinformatic tools, are easily iden-tified and their potential biotechnological valuecan be tested. However, although many genes areconserved between plant species, other genes existonly in one or a few species. These species-spe-cific genes are primarily involved in the synthesisof more than 100,000 different secondary me-tabolites, such as unusual fatty or amino acids andterpenoids, many of which possess nutritional andmedical benefits. Such species-specific genes areunlikely to be in the current genomic and ESTdatabases. The way to identify species-specificgenes whose function can be hinted at by thecurrently available bioinformatic tools is by gen-erating EST databases from selected tissues orplants where these genes are abundantly expressed.This approach has been used successfully for the

193
cloning and manipulation of novel genes regulat-ing the synthesis of unusual fatty acids (Cahoonet al., 1999; van de Loo et al., 1995), as well asgenes regulating terpenoid biosynthesis from pep-permint (Lange et al., 2000; Ohlrogge andBenning, 2000), and phenylpropanoids from basil(Gang et al., 2001).
It is not only plant genomic and EST data-bases that can aid with the identification of novelgenes for improving plant value. Many plant genespossess homologues in microbial species, whosegenomes have been entirely sequenced. In somecases, such bacterial homologues can be moreeasily identified, using either special selectionprocedures, or taking into account that in prokary-otes genes with related functions are often linkedtogether on a single operon. An elegant exampleof utilizing microbes to discover new plant genesis phytoene desaturase, a key enzyme in the caro-tenoid biosynthetic pathway. Hirschberg and as-sociates (Chamovitz et al., 1993) took advantageon the fact that the activity of this enzyme isstrongly inhibited by the herbicides norflurazonand fluridone and isolated herbicide-resistantmutants in cyanobacteria. Many of the mutantsnested in a single gene and showed the alteredactivity of phytoene desaturase as well as alteredproduction of colored carotenoids. This impliedthat the mutated gene encodes phytoene desaturase(Chamovitz et al., 1993). The information fromthe cyanobacteria was later used to clone thephytoene desaturase gene from tomato (Mann etal., 1994).
Another exquisite example of utilizing infor-mation on microbial genes to improve the nutri-tional value of plants is the production α-toco-pherol (vitamin E). Plant oils typically contain highlevels of a much less effective antioxidant, γ-toco-pherol, which is the immediate precursor ofα-tocopherol (see detailed discussion in SectionVI.A.1.b). The enzyme γ-tocopherol methyl trans-ferase, which converts γ- to α-tocopherol, belongsto a large gene family of methyl transferases, whichpossess a general signature, and therefore it is verydifficult to annotate a specific function to theirmembers based on the currently availablebioinformatic tools. Thus, Shintani and DellaPenna(Shintani and DellaPenna, 1998) took a very el-egant approach to identify this gene. Taking into
account: (1) the presence of operons in prokary-otes, and (2) that other genes in the biosyntheticpathway of tocopherol are easier to identify bycurrent bioinformatic tools, these authors searcheda cyanobacterial genomic database for a homo-logue of an Arabidopsis p-hydroxyphenylpyruvatedioxygenase, another enzyme in the tocopherolbiosynthesis pathway. A cyanobacterial homologuewas identified within an operon of around 10 genes.Notably, one of the other genes in this operon wasa predicted methyl transferase, and likely to be thebacterial homologue of γ-tocopherol methyl trans-ferase. This identity was confirmed by mutant analy-sis and expression of a recombinant protein inEscherichia coli (Shintani and DellaPenna, 1998).The cyanobacterial γ-tocopherol methyl transferasewas later used to search for an homologue fromArabidopsis. One EST candidate was found and shownto encode the same activity on its expression in E.coli. Overexpression of this Arabidopsis gene intransgenic Arabidopsis resulted in an enhanced con-version of γ-tocopherol to α-tocopherol (Shintani andDellaPenna, 1998). Increasing the α-tocopherol lev-els in oilseed crops by overexpressing the Arabidopsisγ-tocopherol methyl transferase is now feasible, as-suming that γ-tocopherol methyl transferase activityis also limiting in these crops.
In addition, the genomics-based discovery ofnovel genes related to food and feed value cannotonly be performed by creating EST libraries andsearching for potential candidates. The genes ofinterest may also be identified by their specificexpression pattern in different tissues. For instance,genes controlling fruit aroma or flower color areexpected to be highly regulated in the fruit andflower tissues, respectively. Genomic searches forgenes that are based on expression pattern requiresspecific tools to test the activity of many genes.This can be performed by two different techniques:(1) analysis of “DNA microarrays”, in which manyESTs are blotted on microchips and hybridizedwith RNAs derived from different tissues or differ-ent growth conditions; and (2) searches of ESTsthat are based on their relative frequencies in cDNAlibraries derived from different tissues or differentgrowth conditions (Ohlrogge and Benning, 2000).An elegant “nutritional genomics” approach tosearch for novel genes related to food value, usinga combination of expression profiling and

194
bioinformatics, has been proposed recently byDellaPenna (DellaPenna, 1999a). Moreover, ex-pression profiling, using DNA microarrays, hasalready been used to identify a novel alcohol fruitacetyltransferase that plays a major role in theproduction of the strawberry flavor (Aharoni et al.,2000). Because the development of the fruit coloris closely associated with the ripening process,Aharoni and associates searched for ESTs with anacetyltransferase sequence signature and a similarexpression pattern to genes encoding fruit color. Apromising candidate gene was identified and theenzyme has been produced in bacteria. The recom-binant enzyme could use various alcohols as sub-strates together with acetyl-CoA and producedvolatile esters similar to those produced by straw-berry fruits.
VII. CONCLUSIONS AND FUTUREPROSPECTS
There has been a tremendous increase in theeffort to improve the nutritional and technologi-cal value of plant foods and feeds. This is a resultof advances in various disciplines, such as foodand health sciences, analytical chemistry, plantand animal biochemistry, molecular biology,genomics, bioinformatics, plant transformation,and biotechnology. Because many plant metabo-lites have profound effects on human and live-stock health and well-being, they also play crucialroles in the value of plant foods and feeds. Theidentification of suitable targets for improvementare key issues. Examples of important targets in-clude provitamin A, iron, and phosphorous inplant seeds, which are key to alleviating severemalnutrition problems. In many cases, a givennutritional trait is the outcome of the action of anumber of genes. For example, by introducing thegene that encodes the γ-tocopherol C-methyl trans-ferase to enrich for α-tocopherol content in seeds,the ratio γ-/α-tocopherol clearly changed, but inorder to increase the total tocopherol levels theexpression of more genes is needed.
Because production of a given metabolite isgenerally controlled by multiple enzymes, eachencoded by a different gene, the choice of themost appropriate genes and the procedures to clone
and manipulate them is also important. In manycases, predictions of the effect caused by theoverexpression or inhibition of important gene(s)to alter a given metabolic flux are realized. Inother cases, although the basic predictions appearto be accurate, the level of the altered metaboliteis not as high as expected, apparently due to aninsufficient level of precursors. In yet other cases,results are unexpected due to differential specific-ity of various enzymes for various substrates invivo, or due to differential regulation of a givenmetabolic pathway in different tissues of a par-ticular plant or between different plant species.Moreover, at times, unknown or silent pathwaysare uncovered that redivert the metabolic flow tototally unexpected metabolites. In these cases,additional studies are necessary to unravel thecomplex networks and the compartmentalizationof the cellular, molecular, and biochemical pro-cesses of plant metabolism in order to achieve thegoal of the metabolic engineering project.
Progress on the identification of desirablegenes and loci also plays an important role inspeeding up modern plant breeding. Improvedgenetic methodologies allow more efficient iden-tification of QTLs affecting the value of plantfoods and feeds. Moreover, the new genetic ap-proaches, described briefly in our review (seeSection VII.A), have enabled the cloning of aQTL (Fridman et al., 2000). This will undoubt-edly increase our understanding of the geneticbasis of quantitative traits and eventually increaseour abilities to actually modify complex traits,such as yield.
More knowledge is also needed in order toimprove the production of desired metaboliteswithout any negative side effects. This stronglydepends on careful design of metabolic modifica-tions so that the engineering processes will notaffect other primary metabolic processes that areessential for plant growth and development. Fur-thermore, many natural products, including thoseof commercial or health importance to man, arenaturally produced in specialized organs or cells(such as glandular trichomes, resin cells, andducts), which are compartmentalized from therest of the cells either due to their own phytotox-icity and insolubility in water. A potential prob-lem could arise if such compounds are constitu-

195
tively produced in other plant tissues becausethey might be toxic or have other nondesirableside effects. This problem can be minimized by acareful choice of promoters that could direct thesynthesis of the phytotoxic compounds to appro-priate anatomical specialized structures in the tar-get plants, or restrict their synthesis to followspecific signals such as wounding or the applica-tion of a chemical trigger close to harvesting.
Despite the massive number of genes that havebeen become available recently by genomic andother approaches, many genes encoding key en-zymes in plant metabolism, particularly secondarymetabolism, are still unknown. This is largely dueto limitations of the current bioinformatic tools toassign specific functions to nearly 50% of the genesin the plant genomes. The development of newbioinformatic tools will be pivotal in the futureidentification of new genes, particularly those en-coding protein families with closely related func-tion, such as methyl transferases and terpenesynthases. Because bioinformatic and genomic toolsare also limited, classic procedures for gene clon-ing and identification will also be important infuture studies. These approaches include purifica-tion of enzymes, identification of functions byexpression of genes in heterologous plant and non-plant systems, as well as by gene knockout ap-proaches.
A better understanding of metabolic fluxes andthe key genes that control them is not sufficient forimproving the values of plant foods and feeds. Trans-fer of desired genes to crop plants depends on effi-cient transfer systems, including plant transforma-tion and wide crosses, which are still not available orpublicly accessible for a number of staple crops.Future efforts should also be directed to increase thenumber of crop species to which genes can be readilyintroduced and remain genetically stable. In addi-tion, in some cases such as high methionine proteinsused for improving plant nutritional quality, we willalso need to understand factors that affect accumu-lation and stability of the transgenic proteins in tar-get tissues of the heterologous crop plants. Themodified crop species should then be tested in fieldtrials to ensure that they can produce sufficient tar-get metabolites without a significant loss of plantperformance and yield under a range of field condi-tions in different geographical locations.
The potential of modern genetic molecularand genomics approaches to improve the value ofplant foods and feeds is beyond the proof of con-cept stage. However, many legal issues concern-ing the regulation and marketing of geneticallyengineered plants and plant products still need tobe addressed, and, ultimately, public acceptanceof transgenic products in the form of foods, fra-grances, and industrial materials will determinetheir availability.
REFERENCES
Abelson, P. H. and Hines, P. J. 1999. The plant revolution.Science 285: 367–368.
Aerts, R. J., Barry, T. N., and McNabb, W. C. 1999. Polyphenolsand agriculture: beneficial effects of proanthocyanidins inforages. Agric Ecosyst Environ 75: 1–12.
Agarwal, S. and Rao, A. V. 2000. Carotenoids and chronicdiseases. Drug Metabol Drug Interact 17: 189–210.
Aharoni, A., Keizer, L. C., Bouwmeester, H. J., Sun, Z.,Alvarez-Huerta, M., Verhoeven, H. A., Blaas, J., vanHouwelingen, A. M., De Vos, R. C., van der Voet, H.,Jansen, R. C., Guis, M., Mol, J., Davis, R. W., Schena,M., van Tunen, A. J., and O’Connell, A. P. 2000.Identification of the SAAT gene involved in straw-berry flavor biogenesis by use of DNA microarrays.Plant Cell 12: 647–662.
Albrecht, M., Takaichi, S., Steiger, S., Wang, Z. Y., andSandmann, G. 2000. Novel hydroxycarotenoids withimproved antioxidative properties produced by genecombination in Escherichia coli. Nat Biotechnol 18:843–846.
Allona, I., Quinn, M., Shoop, E., Swope, K., Cyr, S. S.,Carlis, J., Ridel, J., Retzel, E., Campbell, M. M.,Sederoff, R., and Whetten, R. W. 1998. Analysis ofxylem formation in pine by cDNA sequencing. ProcNatl Acad Sci USA 95: 9693–9698.
Arruda, P., Kemper, E. L., Papes, F., and Leite, A. 2000.Regulation of lysine catabolism in higher plants. TrendsPlant Sci 5: 324–330.
Avice, J. C., Ourry, A., Lemaire, G., Volenec, J. J., andBoucaud, J. 1997. Root protein and vegetative storageprotein are key organic nutrients for alfalfa shootregrowth. Crop Sci 37: 1187–1193.
Bagga, S., Adams, H., Kemp, J. D., and Sengupta-Gopalan,C. 1995. Accumulation of 15–kilodalton zein in novelprotein bodies in transgenic tobacco. Plant Physiol107: 11–23.
Bagga, S., Adams, H. P., Rodriguez, F. D., Kemp, J. D., andSengupta-Gopalan, C. 1997. Coexpression of the maizeδ-zein and β-zein genes results in stable accumulationof δ-zein in endoplasmic reticulum derived proteinbodies formed by β-zein. Plant Cell 9: 1683–1696.

196
Bak, S., Olsen, C. E., Halkier, B. A., and Moller, B. L. 2000.Transgenic tobacco and Arabidopsis plants express-ing the two multifunctional sorghum cytochrome P450enzymes, CYP79A1 and CYP71E1, are cyanogenicand accumulate metabolites derived from intermedi-ates in dhurrin biosynthesis. Plant Physiol 123: 1437–1448.
Bak, S., Olsen, C. E., Petersen, B. L., Moller, B. L., andHalkier, B. A. 1999. Metabolic engineering of p-hydroxybenzylglucosinolate in Arabidopsis by expres-sion of the cyanogenic CYP79A1 from Sorghum bi-color. Plant J 20: 663–671.
Bartlem, D., Lambein, I., Okamoto, T., Itaya, A., Uda, Y.,Kijima, F., Tamaki, Y., Nambara, E., and Naito, S.2000. Mutation in the threonine synthase gene resultsin an overaccumulation of soluble methionine inArabidopsis. Plant Physiol 123: 101–110.
Baucher, M., Monties, B., Van Montagu, M., and Boerjan,W. 1998. Biosynthesis and genetic engineering oflignin. Crit Rev Plant Sci 17: 125–197.
Beard, J. L., Burton, J. W., and Theil, E. C. 1996. Purifiedferritin and soybean meal can be sources of iron fortreating iron deficiency in rats. J Nutr 126: 154–160.
Beier, R. C. and Nigg, H. N. 1992. Natural Toxicants inFoods. In: Phytochemical Resources for Medicineand Agriculture. pp. 247–367. Nigg, H. N. and Seigler,D., Eds., Plenum Press, New York.
Ben Tzvi-Tzchori, I., Perl, A., and Galili, G. 1996. Lysineand threonine metabolism are subject to complex pat-terns of regulation in Arabidopsis. Plant Mol Biol 32:727–734.
Bernacchi, D., Beck-Bunn, T., Eshed, Y., Lopez, J., Petiard,V., Uhlig, J., Zamir, D., and Tanksley, S. D. 1998.Advanced backcross QTL analysis in tomato. I. Iden-tification of QTLs for traits of agronomic importancefrom Lycopersicon hirsutum. Theor Appl Genet 97:381–397.
Bernreuther, A. and Schreier, P. 1991. Multidimensional gaschromatography/mass spectrometry: a powerful toolfo the direct chiral evaluation of aroma compounds inplant tissues. II. Linalool in essential oils and fruits.Phytochem Anal 2: 167–170.
Bick, J. A. and Leustek, T. 1998. Plant sulfur metabolism—the reduction of sulfate to sulfite. Curr Opin PlantBiol 1: 240–244.
Bockholt, A. J. and Rooney, L. W. 1992. QPM hybrids in theUnited States. In: Quality Protein Maize. Mertz, E.T., Ed., St. Paul MN, pp. 111–121.
Bonaventure, N., Wioland, N., and Roussel, G. 1985. Ste-reospecific effects of the alpha-aminoadipic acid onthe retina: a morphological and electrophysiologicalstudy. Doc Ophthalmol 61: 71–77.
Bressani, R. 1966. Protein quality of opaque2 maize in chil-dren. Paper presented at: High Lysine Corn pp. 34–39. Washington, DC, Corn Industries Research Foun-dation.
Briat, J. F. 1999. Plant ferritin and human iron deficiency.Nat Biotechnol 17: 621.
Brinch-Pedersen, H., Galili, G., Knudsen, S., and Holm,P. B. 1996. Engineering of the aspartate familybiosynthetic pathway in barley (Hordeum vulgareL.) by transformation with heterolgogous genesencoding feed-back-insensitive aspartate kinase anddihydrodipicolinate synthase. Plant Mol Biol 32:611–620.
Brinch-Pedersen, H., Olesen, A., Rasmussen, S. K., andHolm, P. B. 2000. Generation of transgenic wheat(Triticum aestivum L.) for constitutive accumulationof an Aspergillus phytase. Mol Breed 6: 195–206.
Brink, E. J. and Beynen, A. C. 1992. Nutrition and magne-sium absorption: a review. Prog Food Nutr Sci 16:124–162.
Bruneton, J. 1995. Pharmacognosy, Phytochemistry, Me-dicinal Plants, Paris, Lavoisier Publishing.
Burkhardt, P. K., Beyer, P., Wunn, J., Kloti, A., Armstrong,G. A., Schledz, M., von Lintig, J., and Potrykus, I.1997. Transgenic rice (Oryza sativa) endosperm ex-pressing daffodil (Narcissus pseudonarcissus)phytoene synthase accumulates phytoene, a key inter-mediate of provitamin A biosynthesis. Plant J 11:1071–1078.
Burnett, R. J. and Larkins, B. A. 1999. Opaque2 modifiersalter transcription of the 27–kDa gamma-zein genesin maize. Mol Gen Genet 261: 908–916.
Burton, G. W. and Ignold, K. U. 1981. Auto-oxidation ofbiological molecules. The antioxidant activity of vita-min E and related chain-breaking phenolic antioxi-dants in vitro. J Am Chem Soc 103: 6472–6476.
Buttery, R. G., Siefert, R. M., Guadagni, D. G., and Ling, D.G. 1971. Characterization of additional volatile com-ponents of tomato. J Agric Food Chem 19: 524–529.
Cahoon, E. B., Carlson, T. J., Ripp, K. G., Schweiger, B. J.,Cook, G. A., Hall, S. E., and Kinney, A. J. 1999.Biosynthetic origin of conjugated double bonds: pro-duction of fatty acid components of high-value dryingoils in transgenic soybean embryos. Proc Natl AcadSci USA 96: 12935–12940.
Chamovitz, D., Sandmann, G., and Hirschberg, J. 1993.Molecular and biochemical characterization of herbi-cide-resistant mutants of cyanobacteria reveals thatphytoene desaturation is a rate-limiting step in caro-tenoid biosynthesis. J Biol Chem 268: 17348–17353.
Chavadej, S., Brisson, N., McNeil, J. N., and De Luca, V.1994. Redirection of tryptophan leads to productionof low indole glucosinolate canola. Proc Natl AcadSci USA 91: 2166–2170.
Chen, D. H., Ye, H. C., and Li, G. F. 2000. Expression of achimeric farnesyl diphosphate synthase gene in Arte-misia annua L. transgenic plants via Agrobacteriumtumefaciens-mediated transformation. Plant Sci 155:179–185.
Chen, F. Q., Foolad, M. R., Hyman, J., St Clair, D. A., andBeelaman, R. B. 1999. Mapping QTLs for lycopeneand other fruit traits in a Lycopersicon esculentum x Lpimpinellifolium cross and comparison of QTLs acrosstomato species. Mol Breed 5: 283–299.

197
Chiba, Y., Ishikawa, M., Kijima, F., Tyson, R. H., Kim, J.,Yamamoto, A., Nambara, E., Leustek, T., Wallsgrove,R. M., and Naito, S. 1999. Evidence for autoregula-tion of cystathionine gamma-synthase mRNA stabil-ity in Arabidopsis. Science 286: 1371–1374.
Clarke, E. J. and Wiseman, J. 2000. Developments in plantbreeding for improved nutritional quality of soya beansII. Anti-nutritional factors. J Agric Sci 134: 125–136.
Combs, G. F. 1992. The Vitamins. Academic Press, SanDiego, CA.
Corbel, G., Robin, C., Frankow-Lindberg, B. E., Ourry, A.,and Guckert, A. 1999. Regrowth of white clover afterchilling: assimilate partitioning and vegetative stor-age proteins. Crop Sci 39: 1756–1761.
Couzy, F., Kastenmayer, P., Mansourian, R., Guinchard, S.,Munoz-Box, R., and Dirren, H. 1993. Zinc absorptionin healthy elderly humans and the effect of diet. Am JClin Nutr 58: 690–694.
Craig, W. J. 1994. Iron status of vegetarians. Am J Clin Nutr59: 1233S-1237S.
Croteau, R., and Karp, F. 1991. Origin of natural odorants.In: Perfumes. Art, Science and Technology. Muller,P. M., and Lamparsky, D., Eds., Elsevier AppliedScience, London, pp. 101–126.
Croteau, R., Kutchan, T. M., and Lewis, N. G. 2000. Naturalproducts (secondary metabolites). In: Biochemistryand Molecular Biology of Plants. Buchanan, B.,Gruissem, W., and Jones, R., Eds., American Societyof Plant Physiologists, Rockville, MD, pp. 1250–1318.
Das, D. K., Sato, M., Ray, P. S., Maulik, G., Engelman, R. M.,Bertelli, A. A., and Bertelli, A. 1999. Cardioprotectionof red wine: role of polyphenolic antioxidants. DrugsExp Clin Res 25: 115–120.
DellaPenna, D. 1999a. Nutritional genomics: manipulatingplant micronutrients to improve human health. Sci-ence 285: 375–379.
DellaPenna, D. 1999b. Nutritional genomics: manipulationplant micronutrients to improve human health. Sci-ence 285: 375–379.
Denbow, M. D., Grabau, E. A., Lacy, G. H., Kornegay, E. T.,Russell, D. R., and Umbeck, P. F. 1998. Soybeanstransformed with a fungal phytase gene improve phos-phorus availability for broilers. Poult Sci 77: 875–881.
Dillard, C. J. and German, J. B. 2000. Phytochemicals:nutraceuticals and human health. J Sci Food Agric 80:1744–1756.
Dixon, R. A. and Paiva, N. L. 1995. Stress-inducedphenylpropanoid metabolism. Plant Cell 7: 1085–1097.
Dixon, R. A. and Steele, C. L. 1999. Flavonoids andisoflavonoids - a gold mine for genetic engineering.Trends Plant Sci 4: 394–400.
Dobson, H. E. M. 1993. Floral volatiles in insect biology. In:Insect-Plant Interactions. Bernays, E., Ed., CRC Press,Boca Raton FL, pp. 47–81.
Doll, H., Koie, B., and Eggum, B. O. 1974. Induced highlysine mutants in barley. Radiation Bot 14: 73–80.
Douglas, M. W., Boling, P. S. D., Parsons, C. M., and Baker,D. H. 2000. Nutritional evaluation of low phytate andhigh protein corns. Poult Sci 79: 1586–1591.
Dowd, P. and Zheng, Z. B. 1995. On the mechanism ofanticlotting action of vitamin E quinone. Proc NatlAcad Sci USA 92: 8171–8179.
Dudareva, N., Cseke, L., Blanc, V. D., and Pichersky, E.1996. Evolution of floral scent in Clarkia: Novel pat-terns of S-linalool synthase gene expression in the C.breweri flower. Plant Cell 8: 1137–1148.
Duke, J. A. 1981. Handbook of Legumes of World Eco-nomic Importance.Plenum Press, New York.
Dunwell, J. M. 1999. Transgenic crops: the next generation,or an example of 2020 vision. Annal Bot 84: 269–277.
Dunwell, J. M. 2000. Transgenic approaches to crop im-provement. J Exp Bot 51: 487–496.
Erin, A. N., Skrypin, V. V., and Kagan, V. E. 1985. Forma-tion of alpha-tocopherol complexes with fatty acids.Nature of complexes. Biochim Biophys Acta 14: 209–214.
Ertl, D., Young, K. A., and Raboy, V. 1998. Plant geneticapproaches to phosphorus management in agriculturalproduction. J Environ Qual 27: 299–304.
Eshed, Y. and Zamir, D. 1995. An introgression line popu-lation of Lycopersicon pennellii in the cultivated to-mato enables the identification and fine mapping ofyield-associated QTL. Genetics 141: 1147–1162.
Falco, S. C., Guida, T., Locke, M., Mauvais, J., Sandres, C.,Ward, R. T., and Webber, P. 1995. Transgenic canolaand soybean seeds with increased lysine. Bio/Technol13: 577–582.
Fearing, P. L., Brown, D., Vlachos, D., Meghji, M., andPrivalle, L. 1997. Quantitative analysis of CryIA (b)expression in Bt maize plants, tissues, and silage andstability of expression over successive generations.Mol Breed 3: 169–176.
Frankard, V., Ghislain, M., and Jacobs, M. 1992. Two feed-back-insensitive enzymes of the aspartate pathway inNicotiana sylvestris. Plant Physiol 99: 1285–1293.
Fraser, P. D., Hedden, P., Cooke, D. T., Bird, C. R., Schuch,W., and Bramley, P. M. 1995. The effect of reducedactivity of phytoene synthase on isoprenoid levels intomato pericarp during fruit development and ripen-ing. Planta 196: 321–326.
Fray, R. G. 1995. Constitutive expression of a fruit phytoenesynthase gene in transgenic tomatoes causes dwarf-ism by redirecting metabolites from the gibberellinpathway. Plant J 8: 693–701.
Frentzen, M. 1998. Acyltransferases from basic science tomodified seed oils. Fett Lipid 100: 161–166.
Fridman, E., Pleban, T., and Zamir, D. 2000. A recombina-tion hotspot delimits a wild-species quantitative traitlocus for tomato sugar content to 484 bp within aninvertase gene. Proc Natl Acad Sci USA 97: 4718–4723.
Gakiere, B., Denis, L., Droux, M., Ravanel, S., and Job, D.2000. Methionine synthesis in higher plants: Sensestrategy applied to cystathionine γ-synthase and cys-

198
tathionine β-lyase in Arabidopsis thaliana. Paper pre-sented at: Sulfur Nutrition and Sulfur Assimilation inHigher Plants. Paul Haupt Publishers, Bern, Switzer-land.
Galili, G. 1995. Regulation of lysine and threonine synthesis.Plant Cell 7: 899–906.
Galili, G., Tang, G., Zhu, X., and Gakiere, B. 2001. Lysinecatabolism: a stress and development super-regulatedmetabolic pathway. Curr Opin Plant Biol 4: 261–266.
Galili, S., Weinberg, Z., and Leshem, Y. 1999. The use ofSDS polyacrylamide gel electrophoresis for detectionof protein degradation during silage fermentation. Is-rael J Plant Sci 48: 97–98.
Galliher, H. L., Alexander, D. E., and Weber, E. J. 1985.Genetic variability of alpha-tocopherol and gammatocopherol in corn embryos. Crop Sci 25: 547–549.
Gang, D. R., Wang, J., Dudareva, N., Nam, K. H., Simon, J.E., Lewinsohn, E., and Pichersky, E. 2001. An investi-gation of the storage and biosynthesis of phenylpropenesin sweet basil. Plant Physiol 125: 539–555.
Gaskell, G., Bauer, M. W., Durant, J., and Allum, N. C.1999. World apart? the reception of genetically modi-fied foods in Europe and U.S. Science 285: 384–387.
Geevers, H. O. and Lake, J. K. 1992. Development of modi-fied opaque2 maize in South Africa. In: Quality Pro-tein Maize. Mertz, E. T., Ed., American Associationof Cereal Chemists, St. Paul, MN, pp. 49–78.
Gibson, R. A., Lines, D. R., and Neumann, M. A. 1992.Gamma linolenic acid (GLA) content of encapsulatedevening primrose oil products. Lipids 27: 82–84.
Giddings, G., Allison, G., Brooks, D., and Carter, A. 2000.Transgenic plants as factories for biopharmaceuticals.Nat Biotechnol 18: 1151–1155.
Glover, D. V. 1992. Corn protein-genetics, breeding, andvalue in foods and feeds. In: Quality Protein Maize.Mertz, E. T., Ed., American Association of CerealChemists, St. Paul, MN, pp. 49–78.
Goss, J. R. and Kerr, P. S. 1992. Challenges and opportuni-ties for identity preserved varieties. Paper presentedat: 47th Annual Corn and Sorghum Research confer-ence. American Seed Trade Association, Chicago.
Goto, F., Yoshihara, T., Shigemoto, N., Toki, S., and Takaiwa,F. 1999. Iron fortification of rice seed by the soybeanferritin gene. Nat Biotechnol 17: 282–286.
Grima-Pettenati, J. and Goffner, D. 1999. Lignin geneticengineering revisited. Plant Sci 145: 51–65.
Grusak, M. A. 1999. Genomics-assisted plant improvementto benefit human nutrition and health. Trends PlantSci 4: 164–166.
Grusak, M. A. and DellaPenna, D. 1999. Improving thenutrient composition of plants to enhance human nu-trition and health. Ann Rev Plant Physiol Mol Biol 50:133–161.
Guenoune, D., Amir, R., Ben-Dor, B., Wolf, S., and Galili,S. 1999. A soybean vegetative storage protein accu-mulates to high levels in various organs of transgenictobacco plants. Plant Sci 145: 93–98.
Guillet, G., Poupart, J., Basurco, J., and De Luca, V. 2000.Expression of tryptophan decarboxylase and tyrosinedecarboxylase genes in tobacco results in altered bio-chemical and physiological phenotypes. Plant Physiol122: 933–943.
Gura, T. 1999. New genes boost rice nutrition. Science 285:994–995.
Gustine, D. L. 1995. Enhancement of plant disease resis-tance through expression of foreign phytoalexins. In:Phytochemicals and Health. Gustine, D. L. and Flores,H. E., Eds., American Society of Plant Physiologists,Rockville, MD.
Habben, J. E., Moro, G. L., Hunter, B. G., Hamaker, B. R.,and Larkins, B. A. 1995. Elongation factor 1 alphaconcentration is highly correlated with the lysine con-tent of maize endosperm. Proc Natl Acad Sci USA 92:8640–8644.
Hain, R., Reif, H. J., Krause, E., Langebartels, R., Kindl, H.,Vornam, B., Wiese, W., Schmelzer, E., Schreier, P.H., Stocker, R. H., and Stenzel, K., 1993. Diseaseresistance results from foreign phytoalexin expressionin a novel plant. Nature 361: 153–156.
Hallauer, A. R. and Miranda, F. J. B. 1988. Quantitative Genet-ics in Maize Breeding, IA State University Press, IA.
Hancock, K. R., Ealing, P., and White, D. W. R. 1994.Identification of sulphur-rich proteins which resistrumen degradation and are hydrolysed rapidly by in-testinal proteases. Br J Nutr 72: 855–863.
Harpstead, D. D. 1971. High lysne corn. Sci Am 225: 32–34.Harwood, J. 1996. Recent advances in the biosynthesis of
plant fatty-acids. Biochem Biophys Acta 301.Harwood, J. L. 1997. Plant Lipid Metabolism. In: Plant
Biochemistry. Dey, P. M. and J.B., H., Eds., Aca-demic Press, San Diego, pp. 237–272.
Haudenschild, C. D. and Croteau, R. B. 1998. Molecularengineering of monoterpene production. Genet Engi-neer 20: 267–280.
Hefford, M. A. 1997. Engineering nutritious proteins.Biotechnol Gen Engin Rev 14: 191–210.
Hilditch, T. P. and Williams, P. N. 1964. The ChemicalConstitution of Natural Fats. Chapman and Hall,London.
Hirschberg, J. 1999. Production of high-value compounds:carotenoids and vitamin E. Curr Opin Biotechnol 10:186–191.
Hohn, T. M. and Ohrogge, J. B. 1991. Expression of a fungalsesquiterpene cyclase gene in transgenic tobacco. PlantPhysiol 97: 460–462.
Hohn, T. M. and Plattner, R. D. 1989. Expression of thetrichodiene synthase gene of Fusarium sporotrichioidesin Escherichia coli results in sesquiterpene production.Arch Biochem Biophys 275: 92–97.
Huff, W. E., Moore, P. A., Waldroup, P. W., Waldroup, A.L., Balog, J. M., Huff, G. R., Rath, N. C., Daniel, T.C., and Raboy, V. 1998. Effect of dietary phytase andhigh available phosphorus corn on broiler chickenperformance. Poult Sci 77: 1899–1904.

199
Humphrey, J. H., West, K. P., and Sommer, A., Jr. 1992.Vitamin A deficiency and attributable mortality amongunder-5–year-olds. Bull World Health Organization70: 225–232.
Hurrell, R. F., Juillerate, M. A., Reddy, M. B., Lynch, S. R.,Dassenke, S. A., and Cook, J. D. 1992. Soy protein,phytate, and iron absorption in humans. Am J ClinNutr 56: 573–578.
Inba, K., Fujiwara, T., Hayashi, H., Chino, M., Komeda, Y.,and Naito, S. 1994. Isolation of an Arabidopsis thalianamutant, mto1, that overaccumulates soluble methion-ine. Temporal and spatial patterns of soluble methion-ine accumulation. Plant Physiol 104: 881–887.
Jones, R. A. and Scott, S. J. 1983. Improvement of tomatoflavor by genetically increasing sugar and acid con-tents. Euphytica 32: 845–855.
Jongbloed, A. W. and Kemme, P. A. 1994. Apparent digest-ibility and retention of nutrients bound to phytatecomplexes as influenced by microbial phytase andfeeding regimen in pigs. J Anim Sci 72: 126–132.
Jung, R. and Falco, S. C. 2000. Transgenic corn with animproved amino acid composition. Paper presentedat: 8th International Symposium on Plant seeds.Gatersleben, Germany.
Jung, W., Yu, O., Lau, S. M. C., O’Keefe, D. P., Odell, J.,Fader, G., and McGonigle, B. 2000. Identification andexpression of isoflavone synthase, the key enzyme forbiosynthesis of isoflavones in legumes. NatureBiotechnol 18: 208–212.
Karchi, H., Miron, D., Ben-Yaacov, S., and Galili, G. 1995.The lysine-dependent stimulation of lysine catabo-lism in tobacco seeds requires calcium and proteinphosphorylation. Plant Cell 7: 1963–1970.
Karchi, H., Shaul, O., and Galili, G. 1993. Seed specificexpression of a bacterial desensitized aspartate kinaseincreases the production of seed threonine and me-thionine in transgenic tobacco. Plant J 3: 721–727.
Karchi, H., Shaul, O., and Galili, G. 1994. Lysine synthesisand catabolism are coordinately regulated during to-bacco seed development. Proc Natl Acad Sci USA 91:2577–2581.
Karlsen, R. L., Pedersen, O. O., Schousboe, A., andLangeland, A. 1982. Toxic effects of DL-alpha-aminoadipic acid on muller cells from rats in vivo andcultured cerebral astrocytes. Exp Eye Res 35: 305–311.
Khan, M. R. I., Ceriotti, A., Tabe, L., Aryan, A., McNabb,W., Moore, A., Craig, S., D., S., and Higgins, T. J. V.1996. Accumulation of a sulfur-rich seed albuminfrom sunflower in the leaves of transgenic subterra-nean clover (Trifolium subterraneum L). TransgenicRes 5: 179–185.
Khoudi, H., Laberge, S., Ferullo, J. M., Bazin, R., Darveau,A., Castonguay, Y., Allard, G., Lemieux, R., andVezina, L. P. 1999. Production of a diagnostic mono-clonal antibody in perennial alfalfa plants. BiotechnolBioeng 64: 135–143.
Kinney, A. J. 1996. Development of genetically engineeredsoybean oils for food applications. J Food Lipids 3:273–292.
Knudsen, J. T., Tollsten, L., and Bergstrom, G. 1993. Floralscents — a check list of volatile compounds isolatedby headspace techniques. Phytochemistry 33: 253–280.
Knutzon, D. S., Hayes, T. R., Wyrick, A., Xiong, H. , Davies,H. M., and Voelker, T. A. 1999. Lysophosphatidicacid acyltransferase from coconut endosperm medi-ates the insertion of laurate at the sn-2 position oftriacylglycerols in lauric rapeseed oil and can increasetotal laurate levels. Plant Physiol 120: 739–746.
Knutzon, D. S., Lardizabal, K. D., Nelsen, J. S., Bleibaum,J. L., H.M., D., and Metz, J. G. 1995. Cloning of acoconut endosperm cDNA encoding a 1–acyl-sn-glyc-erol-3–phosphate acyltransferase that accepts medium-chain-length substrates. Plant Physiol 109: 999–1006.
Kuc, J. 1995. Plant defense compounds and human health.In: Phytochemicals and Health. Gustine, D. L. andFlores, H. E., Eds., American Society of Plant Physi-ologists, Rockville MD, pp. 69–77.
Kutchan, T. M. 1995. Alkaloid biosynthesis — the basis formetabolic engineering of medicinal plants. Plant Cell7: 1059–1070.
Lambert, J. L. 1994. High oil corn hybrids. In: SpecialtyCorns. Hallauer, A. R., Ed., CRC Press, Boca Raton..
Lange, B. M., Wildung, M. R., Stauber, E. J., Sanchez, C.,Pouchnik, D., and Croteau, R. 2000. Probing essentialoil biosynthesis and secretion by functional evalua-tion of expressed sequence tags from mint glandulartrichomes. Proc Natl Acad Sci USA 97: 2934–2939.
Larson, S. R. and Raboy, V. 1999. Linkage Mapping maizeand barley myo-inositol 1–phosphate synthase DNAsequences: correspondence with a low phytic acidmutation. Theor Appl Genetics 99:27–36.
Larson, S. R., Rutger, J. N., Young, K. A., and Raboy, V.2000. Isolation and genetic mapping of non-lethal rice(Oryza sativa L.) low phytic acid 1 mutation. Crop Sci40: 1397–1405.
Larson, S. R., Young, K. A., Cook, A., Blake, T. K., andRaboy, V. 1998. Linkage mapping of two mutationsthat reduce phytic acid content of barley grain. TheorAppl Genet 97: 141–146.
Lei, X. G., Ku, P. K., Miller, E. R., Ullrey, D. E., andYokoyama, M. T. 1993. Supplemental microbialphytase improves bioavailability of dietary zinc toweanling pigs. J Nutr 123: 1117–1123.
Lei, X. G. and Stahl, C. H. 2000. Nutritional benefits ofphytase and dietary determinants of its efficacy. JAppl Anim Res 17: 97–112.
Leng, R. A. 1990. Factors affecting the utilization of ‘poorquality’ forages by ruminants particularly under tropi-cal conditions. Nutr Res Rev 3: 277–303.
Lewinsohn, E. 1996. Molecular biology for the improvementof medicinal and aromatic plants. Paper presented at:Proceedings of the International Symposium on Me-

200
dicinal and Aromatic Plants. Amherst, MA, ActaHorticulturae, pp. 443–466.
Lewinsohn, E., Schalechet, F., Wilkinson, J, Matsui, K.,Tadmor, Y., Nam, K.H., Amar, O., Lastochkin, E.,Larkov, O., Ravid, U., Hiatt, W., Gepstein, S., andPichersky, E. 2001. Enhanced levels of the aroma andflavor compound S- linalool by metabolic engineeringof the terpenoid pathway in tomato fruits. Plant Physiol127: 1256–1257.
Li, J., Hegeman, C. E., Hanlon, R. W., Lacy, G. H., Denbow,D. M., and Grabau, E. A. 1997. Secretion of activerecombinant phytase from soybean cell-suspensioncultures. Plant Physiol 114: 1103–1111.
Li, Y. C., Ledoux, D. R., Veum, T. L., Raboy, V., and Ertl,D. S. 2000. Effects of low phytic acid corn on phos-phorus utilization, performance, and bone mineraliza-tion in broiler chicks. Poult Sci 79: 1444–1450.
Loewus, F. A. 1990. Inositol biosynthesis. In: Inositol Me-tabolism in Plants. Morre, D. J., Boss, W. F., andLoewus, F. A., Eds., Wiley-Liss, New York, pp. 13–19.
Lopes, M. A., Takasaki, K., Bostwick, D. E., Helentjaris, T.,and Larkins, B. A. 1995. Identification of two opaque2modifier loci in quality protein maize. Mol Gen Genet247: 603–613.
Luckner, M. 1990. Secondary Metabolism in Microorgan-isms, Plants and Animals. 3 ed., Springer Verlag,Berlin.
Magnavaca, R., Larkins, B. A., Schaffert, R. E., and Lopes,M. A. 1993. Improving protein quality in maize andsorghum. In: International Crop Science. Buxton, D.R., Shibles, R., Forsberg, R. A., Blad, B. L., Asay, K.H., Paulsen, G. M., and Wilson, R. F., Eds., CropScience Society of America, Madison WI., Chap. 85.
Mandal, S. and Mandal, R. K. 2000. Seed storage proteinsand approaches for improvement of their nutritionalquality by genetic engineering. Curr Sci 79: 576–589.
Mann, V., Harker, M., Pecker, I., and Hirschberg, J. 2000.Metabolic engineering of astaxanthin production intobacco flowers. Nat Biotechnol 18: 888–892.
Mann, V., Pecker, I., and Hirschberg, J. 1994. Cloning andcharacterization of the gene for phytoene desaturase(Pds) from tomato (Lycopersicon esculentum). PlantMol Biol 24: 429–434.
Marriott, B. M. 2000. Functional foods: an ecologic perspec-tive. Am J Clin Nutr 71: 1728–1734.
Matsuda, J., Okabe, S., Hashimoto, T., and Yamada, Y.1991. Molecular cloning of hyoscyamine 6 beta-hy-droxylase, a 2–oxoglutarate-dependent dioxygenase,from cultured roots of Hyoscyamus niger. J Biol Chem266: 9460–9464.
Mazur, B., Krebbers, E., and Tingey, S. 1999. Gene discov-ery and product development for grain quality traits.Science 285: 372–375.
McNabb, W. C., Spencer, D., Higgins, T. J., and Barry, T. N.1994. In vitro rates of rumen proteolysis of ribulose-1,5–bisphosphate carboxylase (Rubisco) from lucerneleaves, and of ovalbumin, vicilin and sunflower albu-min 8 storage proteins. J Sci Food Agric 64: 53–61.
Mendoza, C., Viteri, E. F., Lonnerdal, B., Young, K. A.,Raboy, V., and Brown, K. H. 1998. Effect of geneti-cally modified, low-phytic acid maize on absorptionof iron from tortillas. Am J Clin Nutr 68: 1123–1127.
Merchen, N. R. and Trigemeyer, E. C. 1992. Manipulation ofamino acid supply to the growing ruminant. J AnimSci 70: 3238–3247.
Mertz, E. T. 1997. Thirty years of opaque2 maize. In: Qual-ity Protein Maize: 1964–1994. Larkins, B. A. andMertz, E. T., Eds. Purdue University Press, WestLafayette, IN, pp. 25–37.
Mertz, E. T., Bates, L. S., and Nelson, O. E. 1964. Mutantgene that changes protein composition and increaseslysine content of maize endosperm. Science 145: 279–280.
Mgbonyebi, O. P., Russo, J., and Russo, I. H. 1998.Antiproliferative effect of synthetic resveratrol onhuman breast epithelial cells. Int J Oncol 12: 865–869.
Michaelson, L. V., Lazarus, C. M., Griffiths, G., Napier, J.A., and Stobart, A. K. 1998a. Isolation of a Delta5–fatty acid desaturase gene from Mortierella alpina. JBiol Chem 273: 19055–19059.
Michaelson, L. V., Napier, J. A., Lewis, M., Griffiths, G.,Lazarus, C. M., and Stobart, A. K. 1998b. Functionalidentification of a fatty acid delta5 desaturase genefrom Caenorhabditis elegans. FEBS Lett 439: 215–218.
Miflin, B., Napier, J., and Shery, P. 1999. Improving plantproduct quality. Nat Biotechnol 17: 13–14.
Mitchell, R. D. and Edwards, J. H. M. 1996. Effects ofphytase and 1,25–dihydroxycholecalciferol on phytateutilization and the quantitative requirement for cal-cium and phosphorous in young broiler chickens. PoultSci 75: 95–110.
Molvig, L., Tabe, L. M., Eggum, B. O., Moore, A. E., Craig,S., Spencer, D., and Higgins, T. J.V. 1997. Enhancedmethionine levels and increased nutritive value ofseeds of transgenic lupins (Lupinus angustifolius L.)expressing a sunflower seed albumin gene. Proc NatlAcad Sci USA 94: 8393–8398.
Monforte, A. J., and Tanksley, S. D. 2000. Development ofa set of near isogenic and backcross recombinant in-bred lines containing most of the Lycopersiconhirsutum genome in a L. esculentum genetic back-ground: a tool for gene mapping and gene discovery.Genome 43: 803–813.
Munck, L. 1992. The case of high-lysine barley breeding.In: Barley: Genetics, Biochemistry, Molecular Biol-ogy and Biotechnology. Shewry, P. R., Ed., CABInternational, Wallingford, U.K., pp. 573–601.
Munck, L., Karlsson, K. E., Hagberg, A., and Eggum, B. O.1970. Gene for improved nutritional value in barleyseed protein. Science 168: 985–987.
Muntz, K., Christov, V., Saalbach, G., Saalbach, I., Waddell,D., Pickardt, T., Schieder, O., and Wustenhagen, T.1998. Genetic engineering for high methionine grainlegumes. Nahrung 42: 125–127.

201
Murphy, D. J. 1996. Engineering oil production in rapeseedand other oil crops. Trends Biotechnol 14: 206–213.
Murphy, D. J. 1999. Production of novel oils in plants. CurrOpin Biotechnol 10: 175–180.
Nakamura, Y., Fukuhara, H., and Sano, K. 2000. Secretedphytase activities of yeasts. Biosci Biotechnol Biochem64: 841–844.
Napier, J. A., Michaelson, L. V., and Stobart, A. K. 1999.Plant desaturases: harvesting the fat of the land. CurrOpin Plant Biol 2: 123–127.
Newton, I. S. 1998. Long-chain polyunsaturated fatty acids— the new frontier in nutrition. Lipid Technol 10: 77–81.
Ohlrogge, J. and Benning, C. 2000. Unraveling plant me-tabolism by EST analysis. Curr Opin Plant Biol 3:224–228.
Ohlrogge, J. and Browse, J. 1995. Lipids biosynthesis. PlantCell 7: 957–970.
Ohlrogge, J. and Jaworski, J. 1997. Regulation of plant fattyacid biosynthesis. Ann Rev Plant Physiol Plant MolBiol 48: 109–136.
Or, E., Boyer, S. K., and Larkins, B. A. 1993. opaque2modifiers act post-transcriptionally and in a polarmanner on gamma-zein gene expression in maizeendosperm. Plant Cell 5: 1599–1609.
Padley, F. B., Gunstone, F. D., and Harwood, J. L. 1994.Occurrence and characteristics of oil and fats. In: TheLipid Handbook. Gunstone, F. D., Harwood, J. L., andPadley, F. B., Eds., Chapman and Hall, London, pp.49–170.
Park, J., Choi, Y., Jang, M., Lee, Y., Jun, D. Y., Suh, S.,Baek, W., Suh, M., Jin, I., and Kwon, T. K. 2001.Chemopreventive agent resveratrol, a natural productderived from grapes, reversibly inhibits progressionthrough S and G2 phases of the cell cycle in U937cells. Cancer Lett 163: 43–49.
Parker-Barnes, J. M., Das, T., Bobik, E., Leonard, A. E.,Thurmond, J. M., Chaung, L., Huang, Y., and Mukerji,P. 2000. Identification and characterization of an en-zyme involved in the elongation of n-6 and n-3 poly-unsaturated fatty acids. Proc Natl Acad Sci USA 97:8284–8289.
Pen, J., Verwoerd, T. C., van Paridon, P. A., Beudeker, R. F.,van den Elzen, P. J. M., Geerse, K., van der Klis, J. D.,Versteegh, H. A. J., van Ooyen, A. J. J., and Hoekema,A. 1993. Phytase-containing transgenic seeds as anovel feed additive for improved phosphorus utiliza-tion. Bio/Technol 11: 811–814.
Pendurthi, U. R., Williams, J. T., and Rao, L. V. 1999.Resveratrol, a polyphenolic compound found in wine,inhibits tissue factor expression in vascular cells: apossible mechanism for the cardiovascular benefitsassociated with moderate consumption of wine.Arterioscler Thromb Vasc Biol 19: 419–426.
Pichersky, E., Lewinsohn, E., and Croteau, R. 1995. Purifica-tion and characterization of S-linalool synthase, anenzyme involved in the production of floral scent inClarkia breweri. Arch Biochem Biophys 316: 803–807.
Prestage, S., Linforth, R. S. T., Taylor, A. J., Lee, E., Speirs,J., and Schuch, W. 1999. Volatile production in to-mato fruit with modified alcohol dehydrogenase ac-tivity. J Sci Food Agric 79: 131–136.
Raboy, V. 1997. Accumulation and storage of phosphate andminerals. In: Cellular and Molecular Biology of PlantSeed Development. pp. 441–477. Larkins, B. A., andVasil, I. K., eds. Kluwer Academic Publishers,Dordrecht, Netherlands.
Raboy, V. 1998. The genetics of seed storage phosphoruspathways. In: Current Topics in Plant Physiology. J.P. Lynch and J. Deikman, Eds., American Society ofPlant Physiologists Publication, Rockville, Maryland,pp. 133–144.
Raboy, V., and Gerbasi, P. 1996. Genetics of myo-inositolphosphate synthesis and accumulation. In: In myo-Inositol Phosphates, Phosphoinositides, and SignalTransduction. Biswas, B. B. and Biswas, S., Eds.,Plenum Press, New York.
Raboy, V., Gerbasi, P., Young, K. A., Stoneberg, S. D.,Pickett, S. G., Bauman, A. T., Murthy, P. P. N.,Sheridan, W. F., and Ertl, D. 2000. Origin and seedphenotype of maize low phytic acid 1–1 and lowphytic acid 2–1. Plant Physiol 124: 355–368.
Raboy, V., Young, K. A., and Ertl, D. S. 1998. Breeding cornfor improved nutritional value and reduced environ-mental impact. Paper presented at: Proceedings, 52ndAnnual Corn and Sorghum Research Conference,American Seed Trade Association Chicago IL, pp.271–282.
Ravanel, S., Gakiere, B., Job, D., and Douce, R. 1998. Thespecific features of methionine biosynthesis and me-tabolism in plants. Proc Natl Acad Sci USA 95: 7805–7812.
Reddy, N. R., Pierson, M. D., Sathe, S. K., and Salunkhe, D.K. 1989. Phytates in Cereals and Legumes. CRCPress, Boca Raton, FL.
Reddy, T. S. and Thomas, T. L. 1996. Expression of acyanobacterial delta 6–desaturase gene results ingamma-linolenic acid production in transgenic plants.Nat Biotechnol 14: 639–642.
Reichenbach, A. and Wohlrab, F. 1985. Effects of alpha-aminoadipic acid on the glutamate-isolated P III of therabbit electroretinogram. Doc Ophthalmol 59: 359–364.
Robbins, M. P., Bavage, A. D., Strudwicke, C., and Morris,P. 1998. Genetic manipulation of condensed tanninsin higher plants. II. Analysis of birdsfoot trefoil plantsharboring antisense dihydroflavonol reductase con-structs. Plant Physiol 116: 1133–1144.
Roesler, K., Shintani, D., Savage, L., Boddupalli, S., andOhlrogge, J. 1997. Targeting of the Arabidopsishomomeric acetyl-coenzyme A carboxylase to plas-tids of rapeseeds. Plant Physiol 113: 75–81.
Romer, S. 2000. Elevation of the provitamin A content of transgenictomato plants. Nature Biotechnol 18: 666–669.
Ronen, G., Carmel-Goren, L., Zamir, D., and Hirschberg, J.2000. An alternative pathway to beta -carotene forma-

202
tion in plant chromoplasts discovered by map-basedcloning of beta and old-gold color mutations in to-mato. Proc Natl Acad Sci USA 97: 11102–11107.
Rulquin, H. and Verite, R. 1993. Amino acid nutrition ofdairy cows: productive effects and animal require-ments. In: Recent Advances in Animal Nutrition.Garnworthy, P. C. and Cole, D. J. A., Eds., NottinghamUniversity Press., Nottingham, pp. 57–77.
Saalbach, I., Pickardt, T., Machemehl, F., Saalbach, G.,Schieder, O., and Muntz, K. 1994. A chimeric geneencoding the methionine-rich 2S albumin of the Bra-zil nut (Bertholletia excelsa H.B.K.) is stably expressedand inherited in transgenic grain legumes. Mol GenGenet 242: 226–236.
Saalbach, I., Pickardt, T., Waddell, D. R., Hillmer, S.,Schieder, O., and Muntz, K. 1995a. The sulfur-richBrazil nut 2S albumin is specifically formed intransgenic seeds of the grain legume vicia-narnonensis.Euphytica 85: 181–192.
Saalbach, I., Waddell, D., Pickardt, T., Schieder, O., andMuntz, K. 1995b. Stable expression of the sulfur-rich2S albumin gene in trangsenic vicia-narbonensis in-creases the methionine content of seeds. J Plant Physiol145: 674–681.
Saito, K. 2000. Regulation of sulfate transport and synthesisof sulfur-containing amino acids. Curr Opin PlantBiol 3: 188–195.
Samuelsson, G. 1992. Drugs of Natural Origin. SwedishPharmaceutical Press, Stockholm.
Sayanova, O., Smith, M. A., Lapinskas, P., Stobart, A. K.,Dobson, G., Christie, W. W., Shewry, P. R., andNapier, J. A. 1997. Expression of a borage desaturasecDNA containing an N-terminal cytochrome b5 do-main results in the accumulation of high levels ofdelta6–desaturated fatty acids in transgenic tobacco.Proc Natl Acad Sci USA 94: 4211–4216.
Schmidt-Dannert, C., Umeno, D., and Arnold, F. H. 2000.Molecular breeding of carotenoid biosynthetic path-ways. Nat Biotechnol 18: 750–753.
Serageldin, I. 1999. Biotechnology and food security in the21st century. Science 285: 387–389.
Shaul, O. and Galili, G. 1992. Threonine overproduction intransgenic tobacco plants expressing a mutant desen-sitized aspartate kinase from Escherichia coli. PlantPhysiol 100: 1157–1163.
Shaul, O. and Galili, G. 1993. Concerted regulation of lysine andthreonine synthesis in tobacco plants expressing bacterialfeedback-insensitive aspartate kinase and dihydrodipicolinatesynthase. Plant Mol Biol 23: 759–768.
Shewmaker, C. K., Sheehy, J. A., Daley, M., Colburn, S.,and Ke, D. Y. 1999. Seed-specific overexpression ofphytoene synthase: increase in carotenoids and othermetabolic effects. Plant J 20: 401–412.
Shintani, D. and DellaPenna, D. 1998. Elevating the vitaminE content of plants through metabolic engineering.Science 282: 2098–100.
Shmeleva, V. G., Lissitskaya, T. B., Vardoian, G. S., andYakovlev, V. I. 2000. Extracellular phytase synthesis
by fungus Aspergillus niger. Mikol Fitopatol 34: 30–35.
Shotwell, M. A. and Larkins, B. A. 1989. The biochemistryand molecular biology of seed storage proteins. In:The Biochemistry of Plants. Marcus, A., Ed., Aca-demic Press, San Diego, CA, pp. 297–345.
Sies, H. and Stahl, W. 1995. Vitamins E and C, beta-caro-tene, and other carotenoids as antioxidants. Am J ClinNutr 62: 1315S-1321S.
Simopolus, A. P. 1999. Essential fatty acids in health andchronic disease. Am J Clinic Nutr 70: 560S-569S.
Singh, R. and Axtell, J. D. 1973. High lysine mutant gene(hl) that improves protein quality and biological valueof grain sorghum. Crop Sci 13: 535–539.
Somerville, C., Browse, J., Jaworski, J. G., and Ohrlogge, J.B. 2000. Lipids. In: Biochemistry and Molecular Bi-ology of Plants. Buchanan, B., Gruissem, W., andJones, R., Eds., American Society of Plant Physiolo-gists, Rockville, MD, pp. 456–527.
Somerville, C. and Somerville, S. 1999. Plant functionalgenomics. Science 285: 380–383.
Speirs, J., Lee, E., Holt, K., Yong-Duk, K., Steele Scott, N.,Loveys, B., Schuch, W. 1998. Genetic manipulationof alcohol dehydrogenase levels in ripening tomatofruit affects the balance of some flavor aldehydes andalcohols. Plant Physiol 117: 1047–1058.
Spencer, D., Higgins, T. J. V., Freer, M., Dove, H., andCoombe, J. B. 1988. Monitoring the fate of dietaryproteins in rumen fluid using gel electrophoresis. Br JNutr 60: 241–247.
Spencer, J. D., Allee, G. L., and Sauber, T. E. 2000. Phos-phorus bioavailability and digestibility of normal andgenetically modified low-phytate corn for pigs. J AnimSci 78: 675–681.
Stampfer, M. J., Hennekens, C. H., Munson, J. E., Colditz,G. A., Rosner, B., and Willett, W. C. 1993. VitaminE consumption and the risk of coronary disease inwomen. New Engl J Med 328: 1444–1449.
Staswick, P. E. 1994. Storage proteins of vegetative planttissues. Anual Rev Plant Physiol Plant Mol Biol 45:303–322.
Stevens, M. A. and Rick, C. M. 1986. Genetics and breeding.In: The Tomato Crop. Atherton, J. G. and Rudich, J.,Eds., Chapman and Hall, New York.
Sugiura, S. H., Raboy, V., Young, K. A., Dong, F. M., andHardy, R. W. 1999. Availability of phosphorus andtrace elements in low-phytate varieties of barley andcorn for rainbow trout (Oncorhynchus mykiss). Aquac-ulture 170: 285–296.
Sun, Y., Carneiro, N., Clore, A. M., Moro, G. L., Habben, J.E., and Larkins, B. A. 1997. Characterization of maizeelongation factor 1A and its relationship to proteinquality in the endosperm. Plant Physiol 115: 1101–1107.
Sunitha, K., Yo, K., Lee, J.K., and T.K., O. 2000. Statisticaloptimization of seed and induction conditions to en-hance phytase production by recombinant Escheri-chia coli. Biochem Eng J 5: 51–56.

203
Tabe, L. and Higgins, T. J. V. 1998. Engineering plantprotein composition for improved nutrition. TrendsPlant Sci 3: 282–286.
Tabe, L. M., Higgins, C. M., McNabb, W. C., and Higgins,T. J. V. 1993. Genetic engineering of grain and pas-ture legumes for improved nutritive value. Genetica90: 181–200.
Tabe, L. M., Wardley-Richardson, T., Ceriotti, A., Aryan,A., McNabb, W., Moore, A., and Higgins, T. J. V.1995. A biotechnological approach to improving thenutritive value of alfalfa. J Anim Sci 73: 2752–2759.
Tadmor, Y., Tracy, W. F., Yousef, G., Juvik, J. A., andRaboy, V. 2001. Low phytic acid1–1 does not affectsugar metabolism in sugary-1 kernels. MAYDICA 46:11–19.
Tanksley, S. D. and Nelson, J. C. 1996. Advanced backcrossQTL analysis: a method for the simultaneous discov-ery and transfer of valuable QTLs from unadaptedgermplasm into elite breeding lines. Theor Appl Genet92: 191–203.
Tarczynski, M. C., Bo, S., Changjiang, L., Leustek, T., Falco,C., and Allen, W. 2001. Control and Manipulation ofSulfur Amino Acid Metabolism in plants. In: PlantFoods for Human Health: Manipulating Plant Me-tabolism to Enhance Nutritional Quality. KeystoneSympos, Breckenridge, Colorado, 6.4.-11.4.
Teferedegne, B. 2000. New perspectives on the use of tropi-cal plants to improve ruminant nutrition. Proc NutrSoc 59: 209–214.
Theil, E. C. 1987. Ferritin: structure, gene regulation, andcellular function in animals, plants, and microorgan-isms. Ann Rev Biochem 56: 289–315.
Theil, E. C., Burton, J. W., and Beard, J. L. 1997. A sustain-able solution for dietary iron deficiency through plantbiotechnology and breeding to increase seed ferritincontrol. Eur J Clin Nutr 51: S28–31.
Thomson, D. M. H. 1987. The meaning of flavour. In:Developments in Food Flavours. Birch, G. G. andLindley, N. G., Eds., Elsevier, London, pp. 1–21.
Traber, M. G., and Sies, H. 1996. Vitamin E in humans:demand and delivery. Ann Rev Nutr 16: 321–347.
Ulyatt, M. J., Thomson, D. J., Beever, D. E., Evans, R. T.,and Haines, M. J. 1988. The digestion of perennialryegrass (Lolium perenne cv. Melle) and white clover(Trifolium repens cv Blanca) by grazing cattle. Br JNutr 60: 137–149.
van de Loo, F. J., Broun, P., Turner, S., and Somerville, C.1995. An oleate 12–hydroxylase from Ricinus com-munis L. is a fatty acyl desaturase homolog. Proc NatlAcad Sci USA 92: 6743–6747.
Van den Berg, H., Faulks, R., Fernando Granado, H.,Hirschberg, J., Olmedilla, B., Sandmann, G., Southon,S., and Stahl, W. 2000. The potential for the improve-ment of carotenoid levels in foods and their likelysystemic effects. J Sci Food Agric 80: 880–912.
Van Duyn, M. A. and Pivonka, E. 2000. Overview of thehealth benefits of fruit and vegetable consumption for
the dietetics professional: selected literature. J AmDiet Assoc 100: 1511–1521.
Van Wuytswinkel, O., Vansuyt, G., Grignon, N., Fourcroy,P., and Briat, J. F. 1999. Iron homeostasis alteration intransgenic tobacco overexpressing ferritin. Plant J17: 93–97.
Verwoerd, T. C., van Paridon, P. A., van Ooyen, A. J. J., vanLent, J. W. M., Hoekema, A., and Pen, J. 1995. Stableaccumulation of Aspergillus niger phytase in transgenictobacco leaves. Plant Physiol 109: 1199–1205.
Voelker, T. A., Hayes, T. R., Cranmer, A. C., and Davies, H.M. 1996. Genetic engineering of a quantitative trait:metabolic and genetic parameters influencing the ac-cumulation of laurate in rapeseed. Plant J 9: 229–241.
Waldhauser, S. S. M., Kretschmar, J. A., and Baumann, T.W. 1997. N-Methyltransferase activities in caffeinebiosynthesis: biochemical characterization and timecourse during leaf development of Coffea arabica.Phytochemistry 44: 853–859.
Waldroup, P. W., Kersey, J. H., Saleh, E. A., Fritts, C. A.,Yan, F., Stilborn, H. L., Crum, R. C., and Raboy, V.2000. Nonphytate phosphorus excretion of broilerchicks fed diets composed of normal or high availablephosphate corn with and without microbial phytase.Poult Sci 79: 1451–1459.
Wandelt, C. I., Khan, M. R. I., Craig, S., Schroeder, H. E.,Spencer, D., and Higgins, T. J. V. 1992. Vicilin withcarboxy-terminal KDEL is retained in the endoplas-mic reticulum and accumulates to high levels in theleaves of transgenic plants. Plant J 2: 181–192.
Wang, C., Chin, C. K., Ho, C. T., Hwang, C. F., Polashock,J. J., and Martin, C. E. 1996. Changes of fatty acidsand fatty acid-derived flavor compounds by express-ing the yeast D-9 desaturase gene in tomato. J AgricFood Chem 44: 3399–3402.
Wang, J., Sheehan, M., Brookman, H., and Timko, M. P.2000. Characterization of cDNA differentially ex-pressed in roots of tobacco (Nicotiana tabacum cvBurley 21) during the early stages of alkaloid biosyn-thesis. Plant Sci 158: 19–32.
Welinder, E., Textorius, O., and Nilsson, S. E. 1982. Effectsof intravitreally injected DL-alpha-aminoadipic acidon the c-wave of the D.C.-recorded electroretinogramin albino rabbits. Invest Ophthalmol Vis Sci 23: 240–245.
Weselake, R. J. and Taylor, D. C. 1999. The study of storagelipid biosynthesis using microspore-derived culturesof oilseed rape. Prog Lipid Res 38: 401–460.
(WHO), W. H. O. 1992. National strategies for overcomingmicronutrients. In: Document A45/3 WHO, Geneva,Switzerland.
Wilcox, J. R., Premchndra, G. S., Young, K. A., and Raboy,V. 2000. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci 40: 1601–1605.
Williamson, G., Rhodes, M. J. C., and Parr, A. J. 1999.Disease prevention and plant dietary substances. In:Chemicals from Plants, Perspectives on Plant Sec-

204
ondary Products. Walton, N. J. and Brown, D. E.,Eds., Imperial College Press, London, pp. 251–276.
Wittstock, U., and Halkier, B. A. 2000. Cytochrome P450CYP79A2 from Arabidopsis thaliana L. catalyzes theconversion of L-phenylalanine to phenylacetaldoximein the biosynthesis of benzylglucosinolate. J Biol Chem275: 14659–14666.
Yan, F., Kersey, J. H., Fritts, C. A., Waldroup, P. W., Stilborn,H. L., Crum, R. C., Rice, D. W., and Raboy, V. 2000.Evaluation of normal yellow dent corn and high avail-able phosphorus corn in combination with reduceddietary phosphorus and phytase supplementation forbroilers grown to marker weights in litter pens. PoultSci 79: 1282–1289.
Ye, X., Al-Babili, S., Kloti, A., Zhang, J., Lucca, P., Beyer,P., Potrykus, I. Related Articles 2000. Engineeringthe provitamin A (β-carotene) biosynthetic pathwayinto (carotenoid-free) rice endosperm. Science 287:303–305.
Yo, K., Lee, J. K., Oh, B. C., and Oh, T. K. 1999. High-levelexpression of a recombinant thermostable phytase inBacillus subtilis. Biosci Biotechnol Biochem 63: 2205–2207.
Yu, O., Jung, W., Shi, J., Croes, R. A., Fader, G. M.,McGonigle, B., and Odell, J. T. 2000. Production ofthe isoflavones genistein and daidzein in non-legumedicot and monocot tissues. Plant Physiol 124: 781–793.
Yun, D. J., Hashimoto, T., and Yamada, Y. 1992. Metabolicengineering of medicinal plants: transgenic Atropabelladonna with an improved alkaloid composition.Proc Natl Acad Sci USA 89: 11799–11803.
Zarkadas, C. G., Hamilton, R. I., Yu, Z. R., Choi, V. K.,Khanizadeh, S., Rose, N. G., and Pattison, P. L. 2000.Assessment of the protein quality of 15 new northernadapted cultivars of quality protein maize using aminoacid analysis. J Agric Food Chem 48: 5351–5361.
Zhang, Z. B., Kornegay, E. T., Radcliffe, J. S., Denbow, D.M., Veit, H. P., and Larsen, C. T. 2000. Comparisonof genetically engineered microbial and plant phytasefor young broilers. Poult Sci 79: 709–717.
Zeh, M. Casazza, A. P., Kreft, O. Roessner, U. Bieberich, K.,Willmitzer, L., Hoefgen, R. and Hesse. H. 2001Antisense inhibition of threonine synthase leads tohigh methionine content in transgenic potato plants.Plant Physiol. 127: 792–802
Zhou, J. R., Fordyce, E. J., Raboy, V., Dickinson, D. B.,Wong, M. S., Burns, R. A., and Erdman, J. W. 1992.Reduction of phytic acid in soybean products improvezinc bioavailability in rats. J Nutr 122: 1466–2473.
Zook, M., Hohn, T., Bonnen, A., Tsuji, J., and Hammerschmidt,R. 1996. Characterization of novel sesquiterpenoid bio-synthesis in tobacco expressing a fungal sesquiterpenesynthase. Plant Physiol 112: 311–318.
Zou, J. G., Huang, Y. Z., Chen, Q., Wei, E. H., Hsieh, T. C.,and Wu, J. L. 1999. Resveratrol inhibits copper-ion-induced and azo-compound-initiated oxidative modi-fication of human low density lipoprotein. BiochemMol Biol Int 47: 1089–1096.
Zou, J. T., Katavic, V., Giblin, E. M., Barton, D. L., MacKenzie,S. L., Keller, W. A., Hu, X., and Taylor, D. C. 1997.Modification of seed oil content and the acyl composi-tion in the Brassicaceae by expression of a yeast sn-2acyltransferase gene. Plant Cell 9: 909–923.






























