Embed Size (px)
Citation preview

Mukul Dev Yadav 1

INTRODUCTION
STRUCTURE OF HELICASE
CLASSIFICATION OF HELICASE
MECHANISM OF HELICASE ACTION
HELICASE AND HUMAN DISEASE
CONCLUSION
Mukul Dev Yadav 2

The family of nucleic acid strand separation enzyme known as Helicase.
This enzyme found in all organisms and participate in wide variety of cellular
processes.
The number of helicase expressed in higher organisms is strikingly high, with
approximately 1% of the genes in many eukaryotic genomes apparently encoding
RNA or DNA helicase.
Helicases are involved in virtually all aspects of nucleic acid metabolism,
including replication, repair, recombination, transcription, chromosome
segregation, and telomere maintenance .
Traditionally, helicases are known to unwind double-stranded DNA or RNA in an
ATP-dependent manner.
Mukul Dev Yadav 3

All helicases share a RecA fold.
Each helicase molecule contains a single NTP binding site and a distinct
polynucleotide binding site.
These sites are allosterically linked, since the NTPase activity modulates nucleic
acid binding affinity, and vice-versa.
Common Features of Helicase Structure
Sub-domains 1A and 2A contain RecA-like folds. Residues from conserved
helicase motifs line the interface of 1A and 2A and bind NTP.
Single-stranded nuleic acid binds in a groove that is formed by 1A and 2A sub-
domains.
Mukul Dev Yadav 4

Conserved helicase motifs and NTP binding site are at subunit interface.
A critical arginine residue from a neighboring subunit is within hydrogen-
bonding distance of the gamma phosphate of NTP bound at the interface, and is
implicated in transducing conformational changes between subunits of the
hexamer.
The central channel of the ring is large enough to accommodate a single strand of
DNA or RNA.
Rings appear to adopt multiple asymmetric conformations in response to ligand
binding.
Mukul Dev Yadav 5
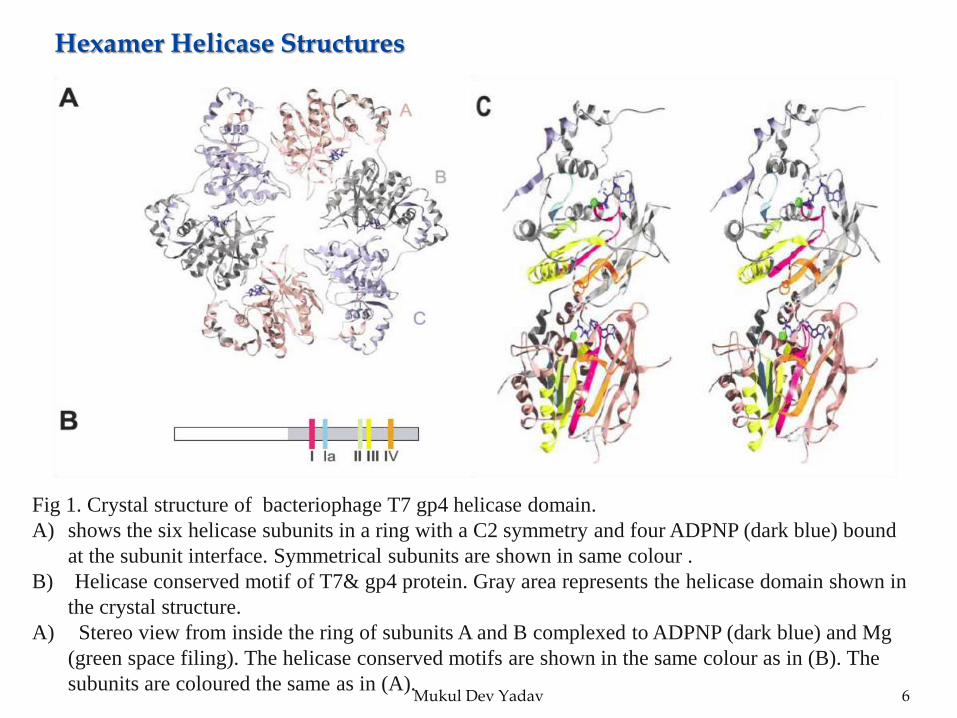
Mukul Dev Yadav 6
Fig 1. Crystal structure of bacteriophage T7 gp4 helicase domain.
A) shows the six helicase subunits in a ring with a C2 symmetry and four ADPNP (dark blue) bound
at the subunit interface. Symmetrical subunits are shown in same colour .
B) Helicase conserved motif of T7& gp4 protein. Gray area represents the helicase domain shown in
the crystal structure.
A) Stereo view from inside the ring of subunits A and B complexed to ADPNP (dark blue) and Mg
(green space filing). The helicase conserved motifs are shown in the same colour as in (B). The
subunits are coloured the same as in (A).
Hexamer Helicase Structures

On the basis of Directionality
Type A- These type helicases unwind the nucleic acid strand in 3’-5’direction.
These helicases requires a 3’tail on the substrate duplex.
Type B- These type of helicases unwind the nucleic acid strand in 5’-3’direction.
These helicases require a 5’tail on the substrate duplex.
Fig 2. Schematic of an unwinding reaction catalyzed by 3’-5’Helicase.
Some helicases have ability to initiate unwinding from a blunt ended duplex.
E.g. – Rec BCD system in E.coli
Mukul Dev Yadav 7

DNA Helicases
It translocate on DNA lattices.
Unwind duplex DNA to form ssDNA intermediates required for DNA replication,
recombination, repair.
Process, translocate branched DNA structures-- Holliday junctions, D-loops, etc.
RNA Helicases
It translocate on RNA lattices.
Destabilize RNA secondary structure; promote ribosome assembly, translation,
RNA splicing, editing, transport, & degradation.
DNA/RNA Helicases
Unwind RNA/DNA hybrids; transcription termination, regulate DNA replication
initiation, etc.
Mukul Dev Yadav 8

On the basis of sequence level helicases divided into Six main group i.e., super
families one to six. This grouping based on their shared sequence motif.
Super Family-1 (SF-1)
Super family -1 can be further divided in to two classes
SF-1A helicases- These helicases have 3’-5’ polarity.
SF-1B helicases- These helicases have 5’-3’ polarity.
But maximum member of SF-1 have 3’-5’ directionality.
It contain seven so called helicase motif I, Ia, II, III, IV,V,VI.
Most known helicases of
SF-1A- Rep & Uvr D in gram negative bacteria.
Pcr A in gram positive bacteria.
SF-1B- Rec D & Dda helicase.
Mukul Dev Yadav 9

This is the largest group of helicase.
It is characterised by the presence of nine conserved motifs like Q, I, Ia, Ib, II, III, IV, V,
VI.
They possess mainly 3’-5’ directionality i.e. type A helicases. But there are some
exceptions like XPD family which is has a polarity of 5’-3’ i.e. type B helicases.
Motif I & II are same as SF-1, but other motif differ in conserved sequences .
Motif I, II, Q, VI all needed for ATP binding & hydrolysis. Motif Ia, Ib, III, IV, V may be
involved in intra molecular rearrangement and RNA interactions.
E.g. Dead box RNA Helicases. XPD (Xeroderma Pigmentosum factor-D) family.
Super Family-3(SF-3)
SF-3 consist of helicase encoded mainly by small DNA viruses and large nucleocytoplasmic DNA
viruses. These are mainly of type A helicases i.e. have 3’-5’ polarity.
Mukul Dev Yadav 10

SF-3 family contain three conserved motifs as motif I, II, III.
E.g.- Papilloma virus have E-1 helicase.
Super Family-4(SF-4)
All SF-4 family helicase have type-B i.e. have 5’-3’ polarity.
These enzyme present in ring structure, mainly present in hexameric ring
structure.
The most studied SF-4 helicases is gp4 (gene 4 helicase-primase) from
bacteriophage T7.
gp4 helicase is member of ring-shaped family of helicases.
Super Family-5(SF-5)
SF-5 is a small family have enzyme such as bacterial transcription factor Rho.
Rho is an essential transcription protein in prokaryotes.
It function by wrapping nucleic acid around a single cleft extending around the
entire hexamer.
Mukul Dev Yadav 11

In recent works SF-6 also called AAA+ protein (ATPase associated with diverse
cellular activities).
It has some enzyme such as RUB branch migration enzyme and MCM proteins
( minichromosome protein complex).
MCM is a eukaryotic DNA helicase complex required for the process DNA
replication, specifically formation and elongation of replication fork. It is
hexamer of six related polypeptide that forms a ring structure.
MCM is also a component of the pre replication complex that form on eukaryotic
ori.
Mukul Dev Yadav 12

Helicases couple the chemical energy of NTP binding and hydrolysis to separate
the complementary strands of double-stranded nucleic acids, remove nucleic acid
associated proteins, or catalyze homologous DNA recombination.
The helicase function is required for efficient and accurate replication, repair, and
recombination of the genome. Similarly, helicase functions facilitate RNA
metabolic processes such as transcription, ribosome biogenesis, translation, RNA
splicing, RNA editing, RNA transport, and RNA degradation.
Here, we focus on the strategies that helicases use to translocate and catalyze
strand separation coupled to NTP binding and hydrolysis.
Mukul Dev Yadav 13

Active State of Helicases
Monomers of ring-shaped helicases are not active in catalyzing NTPase or
unwinding reaction, and hexamer formation is essential.
The ring-shaped structure is stabilized by the binding of NTP, a metal ion, or
both, and by the nucleic acid substrate .
The enclosure of the nucleic acid by the protein subunits decreases the
probability of helicase falling off, thus increasing the ability of the helicase to
stay on track.
Another advantage of this arrangement is the coupling of NTPase cycles
between the hexameric subunits that can increase the efficiency of the
NTPase cycles in promoting translocation.
Oligomerization is an important strategy for non-ring-shaped SF1 and SF2
helicases as well.
Mukul Dev Yadav 14

Some helicases utilize a structural interaction whereas others rely on a functional
interaction. Structural interaction results in the formation of a homodimer or
heterodimer that converts the helicase into a more effective enzyme.
Many helicases show functional cooperativity and enhanced processivity when
multiple molecules of helicases are loaded on the tracking strand .
Yet their activity is enhanced when multiple helicases are loaded on the tracking
strand, which is attributed either to prevention of backward helicase slips or
simply the availability of additional helicase molecules when one falls of the
track.
Mukul Dev Yadav 15

Mukul Dev Yadav 16
Fig 3. Functional oligomeric states of helicases on nucleic acids.
a, ring-shaped hexamer.
b, helicase monomer.
c, helicase dimer.
d, heterodimer.
e, higher oligomer .

Most helicases need a single-stranded nucleic acid region to bind and to initiate
their action of strand separation. Once loaded on the strand, they show a
directional bias and translocate either 5′–3′ or 3′–5′.
Ring-shaped helicases require Y-shaped nucleic acid structures with a loading
strand and a noncomplementary strand of an optimum length to initiate
unwinding.
Helicases show different degrees of tolerance to changes in the chemical nature
of the loading strand while translocating. Some are sensitive to breaks to a basic
sites or to electrostatic disruptions.
While unwinding , certain helicases show no sensitivity to the nature of the
displaced strand. On the other hand, the nature of the displaced strand appears to
influence the activity of some helicase.
Mukul Dev Yadav 17

Mukul Dev Yadav 18
Fig 4. Modes of interacting with the nucleic acid substrate .
a, helicase interacts with one of the single strands of the nucleic acid near the
unwinding junction.
b, helicase interacts with one of the single strands and the duplex region near the
unwinding junction.
c, helicase interacts with both strands of the nucleic acid.
d, helicase interacts with the duplex region.
e, helicase interacts with both the duplex and the two strands of the nucleic acid.

All of the mechanisms involve NTPase coupled nucleic acid affinity changes and
a conformational change (power stroke or ratchet) to explain biased movement
that results in base pair separation or translocation.
The differences in the proposed mechanisms reflect the diverse biochemical
properties including the oligomeric state of the helicase, its mode of binding the
nucleic acid at the unwinding junction, and the effect of the NTP ligation state on
nucleic acid binding properties.
Stepping Mechanisms
In the stepping models, the helicase is always bound to the nucleic acid via two
nucleic acid binding site.
Mukul Dev Yadav 19

In an inchworm type stepping model for a monomeric helicase, a cycle of
nucleic acid binding, release, and translocation events begins with one
helicase site bound tightly to the nucleic acid and the second helicase site
bound weakly to the nucleic acid .
The weak site dissociates from the nucleic acid and in a power stroke motion
moves away from the tight site to bind at a position ahead.
After the weak site has moved and made tight interactions ahead, the original
tight site becomes weak, and as it dissociates from the nucleic acid, in a
power stroke motion it moves forward to get close in distance to the site
ahead.
One cycle in an inchworm stepping mechanism is completed in six
conformational changes.
Mukul Dev Yadav 20

An alternative stepping mechanism (rolling model) for a dimeric helicase has
been proposed for DNA unwinding .
In this model, each of the two subunits of the helicase alternate their binding to
single-stranded and duplex DNA as they change their NTP ligation states.
In contrast to the inchworm model, where the subunits maintain their relative
positioning along the DNA, the subunits in the rolling model take turn in being
the trailing or the leading subunit.
Brownian Motor Mechanism
This model invokes Brownian motion and power stroke, and it is based on two
conformational states of the helicase.
Structural and biochemical studies have identified two distinct conformational
states of helicase with weak and tight nucleic acid binding modes resulting from
the different NTP ligation states.
Mukul Dev Yadav 21

To translocate the helicase needs to loosen its interactions with the nucleic acid,
and this happens when the helicase changes its NTP ligation state.
In the weak state, the helicase-nucleic acid energy profile is shallow and
symmetric, and the helicase can move in either direction (Brownian motion) or
completely dissociate from the nucleic acid.
The short lifetime of the weak state keeps the helicase close to the starting
position. When the helicase resumes the tight state, it makes a step forward
(power stroke).
Those molecules that have fluctuated in the forward direction move ahead and
those that have fluctuated in the opposite direction return to the original position.
Repetition of these steps leads to net forward movement of the helicase along the
nucleic acid.
Mukul Dev Yadav 22

Mukul Dev Yadav 23
Fig 5. Proposed mechanisms of translocation(A, stepping inchworm mechanism. A helicase monomer with a
tight (closed hand) and a weak (open hand) nucleic acid binding site is shown to undergo steps of helicase
movement (power stroke) and nucleic acid affinity changes (tight to weak transitions).
B, Brownian motor mechanism. On the right, the helicase is shown to undergo nucleic acid affinity changes
(tight to weak). In the weak state (2), the helicase fluctuates in either direction. Upon resuming the tight state
(3), some helicase molecules move forward (3) and some return to their original position (1). On the left, the free
energy of the helicase-nucleic acid complex is shown along the nucleic acid length. In the tight state (1), the
helicase is trapped in a deep energy well unable to move. In the weak state (2), thermal fluctuations allow the
helicase to fluctuate in either direction or to completely dissociate from the nucleic acid (4). Upon resuming the
tight state, the deep energy proነle is restored and some helicases move forward (3).

Base pair separation occurs at the junction of single-stranded and duplex regions.
Helicases unwind long stretches of duplex nucleic acids by coupling base pair
separation to translocation.
Depending on how the base pairs are separated, the base pair separation
mechanisms are classified as active or passive .
If the helicase needs to move and bind more than one base at a time, it would
employ some type of an active mechanism to bring about strand separation in an
efficient manner.
In a passive mechanism, the helicase waits for the base pairs to open
spontaneously by thermal fluctuations before it moves and binds the newly
opened bases. Because the terminal base pair at the junction opens and closes at a
very fast rate , this type of a mechanism is attractive for helicases that can move
and occupy one base at a time.
Mukul Dev Yadav 24

Mukul Dev Yadav 25
Fig 6. Proposed mechanisms of strand separation.
A, helicase separates the base pairs at the junction by translocating along one strand of the nucleic acid while
displacing the other using strand exclusion, wire stripper, or a wedge mechanism..
B, helicase destabilizes the duplex region by direct interactions (helix destabilizing mechanism).
C–F illustrate proposed mechanisms for ring-shaped helicases catalyzing bidirectional replication fork
movement. C, helicase rings encircle one of the DNA strands and move in opposite directions to unwind DNA by
strand exclusion mechanism. D, helicase rings encircle duplex DNA and unwind DNA by a
torsional mechanism. E, helicase rings encircle and pump duplex DNA and separate the strands by threading
them through side channels. F, helicase rings encircle and pump duplex DNA and separate the strands by a
Ploughshare mechanism (blue triangle).

Translocation and base pair separation activities of helicases are driven by NTP
binding and hydrolysis.
During each NTPase cycle, the helicase goes through defined NTP ligation states
including empty, NTP, NDP*Pi, and NDP. One or more of the NTP ligation states
causes changes in the affinity of the helicase for the nucleic acid and brings about
a power stroke that leads to translocation and/or strand separation.
Power stroke and nucleic acid affinity modulation steps are energetic events;
therefore, only those changes in the NTPase cycle that are associated with an
observable energy change are likely to drive these events at the nucleic acid
binding site.
Ring-shaped helicases potentially can bind and hydrolyze six NTPs, and many
show co-operativity in NTP binding and hydrolysis.
Mukul Dev Yadav 26

Mukul Dev Yadav 27

Mukul Dev Yadav 28

Helicases are required for the efficient catalysis of most DNA and RNA
metabolic processes where they perform diverse functions.
All helicases share a RecA fold. Each helicase molecule contains a single NTP
binding site and a distinct polynucleotide binding site.
Most helicases need a single-stranded nucleic acid region to bind and to initiate
their action of strand separation.
Base pair separation occurs at the junction of single-stranded and duplex regions.
The basic activity of helicases is to couple NTP binding and hydrolysis to
conformational changes that bring about separation of base pairs or translocation
along nucleic acid.
Mukul Dev Yadav 29

Mukul Dev Yadav 30





























