Embed Size (px)
Citation preview

Camp. Biochem. Physiol. Vol. 92A, No. I, pp. 61-63, 1989 0300-9629/89 $3.00 + 0.00 Printed in Great Britain 0 1989 Pergamon Press plc
VOLTAGE-DEPENDENT ANION-SELECTIVE CHANNELS IN CULTURED SMOOTH MUSCLE
CELLS OF THE RAT AORTA
JUN-ICHI SADOSHIMA,*~ NORIO AKAIKE,f HITONOBU TOMOIKE,* HIDEO KANAIDE* and
MOT~~MI NAKAMURA*
*Research Institute of Angiocardiology and Cardiovascular Clinic and tDepartment of Physiology, Faculty of Medicine Kyushu University, Fukuoka 812, Japan. Telephone: (092) 641-l 151
(Receiued 6 June 1988)
Abstract-l. Properties of the voltage-dependent anion-selective channel in cultured smooth muscle cells of the rat aorta were studied using the patch-clamp technique.
2. The channel had a single channel conductance of 346 & 4 pS (n = 43, mean + SEM) with symmetrical 142 mM-Cl- solution in inside-out patch configurations.
3. The channel was activated spontaneously at a potential range - 20 _ +20 mV and inactivated more rapidly with increases to more positive or negative potentials.
4. The channel was selective for anions and the permeability ratio for monovalent anion was Br- :Cl- : HCOO- :CH,COO-:propionate- :aspartate- = 1.1: 1:0.7:0.4: ~0.02: ~0.02.
5. The openings of the channels were observed more frequently in inside-out membrane patches than in cell-attached ones, and were independent of intracellular free Ca concentrations.
6. The density of this channel was estimated to be 1.3/pmZ. 7. Physiological roles of the channel were discussed.
INTRODUCTION
Chloride conductance in smooth muscle cells has
heen considered to play an important role in gener- ation of the resting membrane potential (Burnstock and Straub, 1958), slow wave (El-Sharkawy & Dan- iel, 1975), negative after-potential (Ohashi, 1970) and agonist-induced depolarization (Bullock and Kirkpa- trick, 1975; Byrne and Large, 1987). However, direct evidence for the Cl- currents in smooth muscle cells has been reported only partially (Shoemaker et al., 1985; Kokubun and Soejima, 1987). In ion flux studies of the rat aorta, a relatively high permeability for Cl- was noted (Jones, 1980). In the present work, we examined properties of the single voltage- dependent Cl- channels in primary cultured smooth muscle cells of adult rat aorta, using the patch-clamp technique (Hamill et al., 1981).
MATERIALS AND METHODS
Smooth muscle cells of the aortic media of Wistar rats (20&25Og) were dispersed enzymatically and cultured for 4-7 days, as described (Yamamoto et al., 1981). Single- channel currents were measured in inside-out configurations (Hamill et al., 1981) using a patch-clamn amnlifier (List. EPC-7) and stored dn a P?M hata recorder (NF, RPl880j for later analysis. Tip resistance of the patch electrode filled with test solutions ranged between 5 and 10 MR. The bath connected to the ground with an agar bridge in series with a reference electrode. The shifts of the junction potential between the ground and the bath during the change of the bathing solution were corrected. The patch pipette solution was a normal external solution (NES) containing (mM): NaCl 136, KC1 6, glucose 12, EGTA 1.0, HEPESTris OH 10 @H 7.3). The bathing medium was a 142mM KCl-
IAuthor to whom all correspondence should be addressed.
internal solution (IS) containing (mM): KC1 142, glucose 12, EGTA 1, HEPES-Tris OH 10 @H 7.2). For experiments on ionic selectivity, KC1 in a 142 mM KCl-IS was replaced with equimolar CsCl, NaCl, TrisCl, KBr, HCOOK, CH,COOK, CH,CH,COOK (K-propionate) or K-aspartate. Some- times, appropriate amounts of CaCl, were added to a 142 mM KCl-IS to make a free Ca concentration. All test solutions were continuously gassed with 99.9% oxygen. Experiments were carried out at room temperature (20-24°C).
RESULTS
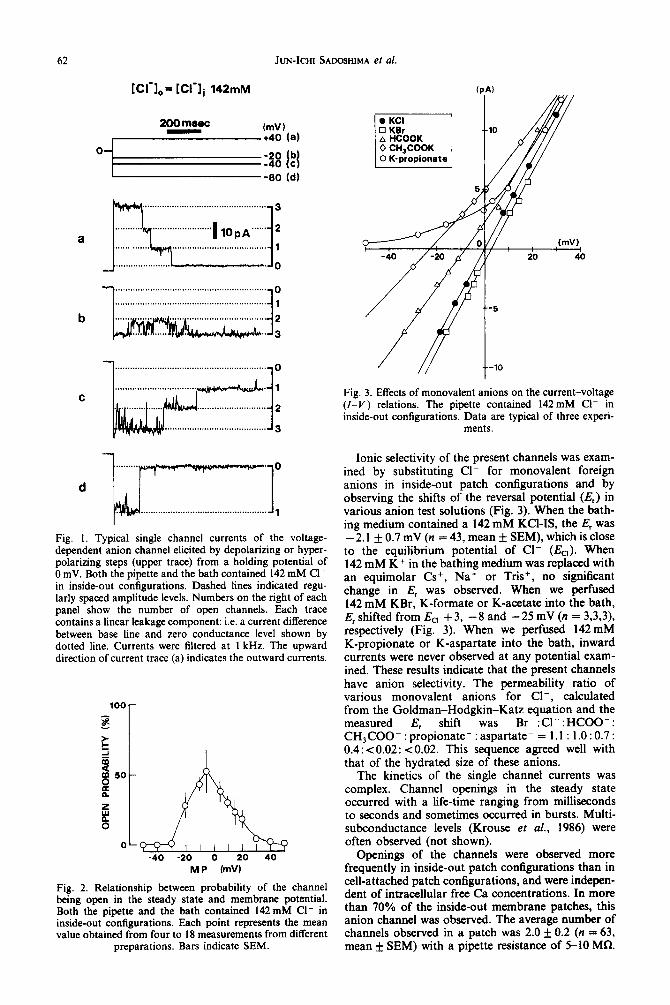
In inside-out membrane patches from cultured smooth muscle cells of the rat aorta, we observed single channel currents characterized by voltage de- pendent openings and a large conductance. The channels showed a high probability of openings in a narrow potential range around 0 mV. However, when membrane potential was stepped to more positive or negative ones from this voltage range, the channels rapidly closed. At further step up of the potential, inactivation of the channels occurred more rapidly and the spontaneous openings were observed less frequently (Fig. 1). Figure 2 shows a plot of steady state open probability as a function of membrane potential. The open probability was determined by analysing continuous recording for more than 30 set at each membrane potential. The channel openings in the steady state were observed in the membrane potential range between -20 and +20 mV and open probability became maximum around OmV. When the pipette contained a NES (142 mM Cl-) and the bath contained a 142mM KCl-IS in inside-out configurations, the channel had a linear single- channel current-voltage (I-V) relationship over a potential range - 60 _ + 50 mV, and a single channel conductance of 346 f 4 pS (n = 43, mean f SEM).
61
62 JIJN-ICHI SADOSHIMA et al.
[Cl-I,= [Cl’]i 142mM
-t . . . . . . . . . . . . . . . . . . . . l--.-10
7 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . +)
b
.............................................................
/l 0
................................. . 1 C . . ........ ...... ............................ * ......................................... 3
d
Fig. I. Typical single channel currents of the voltage- dependent anion channel elicited by depolarizing or hyper- polarizing steps (upper trace) from a holding potential of 0 mV. Both the pipette and the bath contained 142 mM Cl- in inside-out configurations. Dashed lines indicated regu- larly spaced amplitude levels. Numbers on the right of each panel show the number of open channels. Each trace contains a linear leakage component: i.e. a current difference between base line and zero conductance level shown by dotted line. Currents were filtered at 1 kHz. The upward direction of current trace (a) indicates the outward currents.
MP ImV)
Fig. 2. Relationship between probability of the channel being open in the steady state and membrane potential. Both the pipette and the bath contained 142mM Cl- in inside-out configurations. Each point represents the mean value obtained from four to 18 measurements from different
preparations. Bars indicate SEM.
Fig. 3. Effects of monovalent anions on the current-voltage (I-V) relations. The pipette contained 142 mM Cl- in inside-out configurations. Data are typical of three experi-
ments
Ionic selectivity of the present channels was exam- ined by substituting Cl- for monovalent foreign anions in inside-out patch configurations and by observing the shifts of the reversal potential (E,) in various anion test solutions (Fig. 3). When the bath- ing medium contained a 142 mM KCl-IS, the E, was - 2.1 + 0.7 mV (n = 43, mean + SEM), which is close to the equilibrium potential of Cl- (Ea). When 142 mM K+ in the bathing medium was replaced with an equimolar Cs+, Na+ or Tris+, no significant change in E, was observed. When we perfused 142 mM KBr, K-formate or K-acetate into the bath, E, shifted from EC, + 3, - 8 and - 25 mV (n = 3,3,3), respectively (Fig. 3). When we perfused 142mM K-propionate or K-aspartate into the bath, inward currents were never observed at any potential exam- ined. These results indicate that the present channels have anion selectivity. The permeability ratio of various monovalent anions for Cl-, calculated from the Goldman-Hodgkin-Katz equation and the measured E, shift was Br-:Cl-:HCOO-: CH3COO- : propionate- : aspartate- = 1 .l : 1 .O : 0.7 : 0.4: x0.02: ~0.02. This sequence agreed well with that of the hydrated size of these anions.
The kinetics of the single channel currents was complex. Channel openings in the steady state occurred with a life-time ranging from milliseconds to seconds and sometimes occurred in bursts. Multi- subconductance levels (Krouse et al., 1986) were often observed (not shown).
Openings of the channels were observed more frequently in inside-out patch configurations than in cell-attached patch configurations, and were indepen- dent of intracellular free Ca concentrations. In more than 70% of the inside-out membrane patches, this anion channel was observed. The average number of channels observed in a patch was 2.0 f 0.2 (n = 63, mean + SEM) with a pipette resistance of 5-10 MR.

DISCUSSION the cultures. This research was supported in part by Grants-
We obtained evidence for the existence of voltage- in-Aid (Nos 61132008, 61232616, 61440042, 61480109,
dependent, large conductance anion-selective chan- 61870037 and 62222021) from the Ministrv of Education.
nels in cultured smooth muscle cells of the rat aorta. Science and Culture and (6OC-1 and 6iA-1) from the Ministry of Health and Welfare, Japan.
Anion-selective channels which have similar voltage dependency and conductance were also noted in REFERENCES cultured rat skeletal muscle (Blatz and Magleby, 1983), Schwann cells (Gray et al., 1984) and cultured Blatz A. L. and Magleby K. L. (1983) Single voltage-
mouse alveolar cells (Krouse et al., 1986). dependent chloride-selective channels of large conduc-
An average of at least two channels are observed tance in cultured rat muscle. Biophys. J. 43,237-241.
in a patch. Assuming that a membrane patch forms Bormann J.. Hamill 0. P. and Sakmann B. (1987) Mech-
a hemispheric shape with a diameter of 1.0 pm, the anism of ‘anion permeation through channels gated by
channel density would be l.3/pm2, which is greater glycine and y-aminobutyric acid in mouse cultured spinal neurones. J. Physiol., Lond. 385, 243-286.
than that of large conductance anion channels in Bullock C. G. and Kirkpatrick C. T. (1976) Effects of other preparations. When a single channel conduc- histamine on ionic distribution in bovine tracheal smooth tance is 350 pS, using the channel density calculated muscle. Abstract, J. Physiol., Land. 254, 63P-64P.
above, the maximum conductance of this anion chan- Burnstock G. and Straub R. W. (1958) A method for
nel is estimated to be 455 mS/cm2, a value very high studying the effects of ions and drugs of the resting and
compared to even that of the K+ conductance noted action potentials in smooth muscle with external elec-
in cultured rat aortic cells (Toro and Stefani, 1987). trodes. J. Physiol., Lond. 140, 156167.
This evidence may support the result of ion flux Byrne N. G. and Large W. A. (1987) Action of nor-
studies which have showed a relatively high perme- adrenaline on single smooth muscle cells freshly dispersed
ability for Cl- of the rat aorta (Jones, 1980). from the rat anococcygeus muscle. J. Physiol. Lond. 389, 513-525.
Some large polyatomic anions, such as formate or El-Sharkawy T. Y. and Daniel E. E. (1975) Ionic mech- acetate, were permeant in this channel; however, anisms of intestinal electrical control activity. Am. J.
propionate was not measurably permeant. This evi- Physiol. 229, 1287-1298.
dence provides an estimate of the minimum pore Gray P. T. A., Bevan S. and Ritchie J. M. (1984) Hiah
diameter of this channel of 5.2 A, a diameter similar conductance anion-selective channels in rat cultured Sch-
to glycine receptor channels (Bormann et al., 1987). wann cells. Proc. R. Sot. Lond. B221. 395-409.
This channel was not permeable to small cations, a Jones A. W. (1980) Content and fluxes’of electrolvtes. In
finding different from large conductance anion chan- Handbook of Physiology : The Cardiovascular -System (Edited by Bohr, D. F.. Somlvo A. P. and Somlvo A. V-1.
nels in rat cultured Schwann cells (Gray et al., 1984). pp. 253%?00. Am. Physiol. S&. Bethesda. The resting membrane potential of the cultured rat Kokubun S. and Soejima M. (1987) Single voltage-
aortic cells is around - 55 mV (Harder and Sper- dependent chloride channel and its ion selectivity in
elakis, 1979). Since this anion channel is almost cultured smooth muscle cells of rat aorta (Abstract).
inactivated at this potential, it does not seem to play J. Physiol. Sot. Jpn 49, 482.
a role in generation of the resting potentials. The Krouse M. E., Schneider G. T. and Gage P. W. (1986)
intracellular Cl- content of this preparation is esti- A large anion-selective channel has seven conductance
mated to be around 56 mM (Jones, 1980). Assuming levels. Nature 319, 58-60.
that the extracellular Cl- concentration is 142 mM, Hamill 0. P., Marty A., Neher E., Sakmann B. and
the equilibrium potential of the Cl- calculated from Sigworth F. J. (1981) Improved patch-clamp techniques
the Nernst equation is -23 mV. As this anion chan- for high-resolution current recordings from cells and cell-free membrane patches. PflGgers Arch. 391, 85-100.
nel has a high open probability between -20 and Harder D. R. and Sperelakis N. (1979) Action potential +20 mV, when depolarization is induced in this generation in reaggregates of rat aortic smooth muscle
preparation, this channel may play a role in the cells in primary culture. Blood Vessels 16, 186201.
repolarization or generation of the negative after- Mayer M. L. (1985) A calcium-activated chloride current
potential (Ohashi, 1970). The contribution of Cl- to generates the after-depolarization of rat sensory neurones
the after-potential has also been noted in rat sensory in culture. J. Physiol., Land. 364, 217-239.
neurones (Mayer, 1985). Ohashi H. (1970) Effects of changes of ionic environment on
Openings of this channel were observed more the negative after-potential of the spike in rat uterine muscle. J. Physiol., Lond. 210. 785-797.
frequently in inside-out patch than in cell-attached Shoemaker R., Naftel J. and Fariey J. (1985) Measurement patch preparations, indicating that some of the intra- of K+ and Cl- channels in rat cultured vascular smooth
cellular substances suppressed the channel activities. muscle cells (Abstract). Biophys. J. 47, 46Sa.
Further studies are needed to clarify the effects of the Toro L. and Stefani E. (1987) Ca2+ and K+ current in
intracellular messengers on this large conductance cultured vascular smooth muscle cells from rat aorta.
anion channel. Pfliiaers Arch. 408. 417419.
Yamamoto H., Kanaide H. and Nakamura M. (1983) Metabolism of glycosaminoglycans of cultured aortic
Acknowledgements-We thank N. Tokutomi for technical assistance and M. Hasegawa and H. Marui for preparing
smooth muscle cells altered during subculture. Br. J. exp. Parh. 64, lSG-165.
Voltage dependent Cl channel of vascular SMC 63