Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARANÁ
SETOR DE CIÊNCIAS DA TERRA
DEPARTAMENTO DE GEOLOGIA
VICTOR AMIR CARDOSO DORNELES
CARACTERIZAÇÃO GEOQUÍMICA E GEOMICROBIOLÓGICA DE
MICROBIALITO DA LAGOA SALGADA, ESTADO DO RIO DE JANEIRO
CURITIBA
2018

i
VICTOR AMIR CARDOSO DORNELES
CARACTERIZAÇÃO GEOQUÍMICA E GEOMICROBIOLÓGICA DE MICROBIALITO
DA LAGOA SALGADA, ESTADO DO RIO DE JANEIRO
Trabalho de Conclusão de Curso apresentado ao
Curso de Geologia da Universidade Federal do
Paraná como requisito parcial à obtenção do
grau de Bacharel em Geologia.
Orientadora: Profª Dra. Anelize Manuela Bahniuk
Rumbelsperger
CURITIBA
2018

ii
“Sei que minha escalada nunca terá fim, sei que minha busca será constante,
pois dentro de mim existe um universo infinito.”
Waldemar Niclevicz - Everest: O diário de uma Vitória

iii
AGRADECIMENTOS
À Universidade Federal do Paraná pela oportunidade de obter uma formação de qualidade
em uma universidade pública e gratuita.
Ao Departamento de Geologia e ao grupo PET por todo o conhecimento e experiências
transmitidos.
À minha professora orientadora Dra. Anelize Bahniuk pelo incentivo e motivação de fazer
ciência e por me mostrar o admirável e encantador mundo dos microbialitos. Obrigado pela
paciência e por toda a troca de experiência nesses cinco anos.
À quase-geóloga Kimberlym Tábata e aos professores doutores Ricardo Jahnert e Robson
Bolzon pelas dicas e valiosa ajuda nesta pesquisa.
Ao Laboratório de Análise de Minerais e Rochas (LAMIR) pela infraestrutura e
equipamentos disponibilizados, à equipe técnica pela prestatividade e auxílio nas análises, e
também ao Projeto Microbial (FUNPAR-UFPR/LAMIR/PETROBRÁS) pelo apoio financeiro da
pesquisa.
À minha querida supervisora de estágio Claudia Parellada, e ao amigo e companheiro de
pesquisas Thiago Gomes da Silva, por acreditarem no meu potencial e me motivarem tanto a ser
cientista.
֍֍֍
“Eu poderia suportar, embora não sem dor, que tivessem morrido todos os meus amores, mas
enlouqueceria se morressem todos os meus amigos! ” (Paulo Sant’Ana)
Agradeço imensamente aos amigos e colegas da turma de Geologia 2014, com os quais eu
tive o prazer de passar os cinco melhores anos da minha vida. Anderson, Arthur, Felipe, Gabriela,
Geovanne, Gustavo, Hugo, Joana, Kimberlym, Laís, Leticia, Luísa, Luís, Marcelo, Matheus, Rafaella
e Renata, obrigado por todos os momentos que vivemos juntos e que ainda viveremos. Eu não
tenho dúvidas de que fui agraciado com a melhor turma de faculdade que eu poderia ter tido. Vocês
todos, de alguma forma, marcaram minha vida e me ajudaram a evoluir!
Aos demais amigos da Geologia, obrigado pelas trocas de experiências, conselhos e
momentos de descontrações tão necessários para aguentar firme na graduação. Destes, quero citar
Giovana, Mayara, Gabriel e Veleda por sempre acreditarem no meu trabalho e torcerem por mim.
Aos amigos de outros cursos e de fora da universidade, em especial Wagner, Polyana,
Janaina e Kelly, obrigado pela compreensão de minha ausência nestes anos e por estarem comigo
desde o início dessa jornada.
Agradeço mais que tudo à minha querida e maravilhosa mãe. Marli, sem a sua ajuda eu não
teria conquistado nada disso e nem faria sentido lutar por uma vida melhor se não fosse para
retribuir tudo que você já fez por mim. Obrigado pelo seu exemplo de ser humano, seus
ensinamentos e pelo seu amor. Você me fez o homem que eu sou e eu serei eternamente grato a ti.
Aos demais familiares, especialmente Cleonice e Bira, que direta ou indiretamente
contribuíram para a realização desse sonho que é a formação acadêmica, e por compreenderem
minha ausência em diversos momentos ao longo desses anos.

iv
RESUMO
O objetivo deste trabalho foi caracterizar as diferentes assinaturas geoquímicas dos estratos de estromatólitos da Lagoa Salgada, estado do Rio de Janeiro, e correlacioná-las com as distintas tramas encontradas, buscando contribuir para o refinamento do conhecimento acerca da evolução paleoambiental da lagoa. Realizou-se a descrição macroscópica da amostra e análises de Espectrometria de Massa por Plasma Acoplado Indutivamente (ICP-MS), Difratometria de Raios X (DRX) e Isótopos Estáveis de C & O. O estromatólito foi dividido em cinco fácies, da base para o topo: esteira microbiana, trombólito, estromatólito vugular, estromatólito digitado e estromatólito trombolítico. Na fácies trombólito ocorrem carapaças de microgastrópodes da espécie Heleobia australis. Os diferentes tamanhos das valvas indicam que não foram transportados e trapeados pelo estromatólito, mas viveram na estrutura estromatolítica durante o desenvolvimento desta. A composição mineralógica e química do estromatólito não é homogênea, mas varia ao longo da amostra. É evidente uma variação composicional de elementos traços no estromatólito, bem como para os teores de terras raras. Em relação aos isótopos, os valores de δ 13C variam da base para o topo de 2 a 12,78 ‰. Enquanto que os valores de δ 18O variam de 0,92 até -1,65 ‰. A variação na composição química e na trama ao longo das fácies implica em possíveis variações ambientais e das comunidades microbianas responsáveis pela formação deste estromatólito. A presença de dolomita na base de estromatólito indica processos biológicos mais ativos nessas fases de crescimento, ao passo que a notável presença de Mg-calcita nos estratos mais acima indica que processos abióticos devem ser considerados. A calcita com alto teor de Mg no topo da amostra sugere que a dolomita encontrada na base seja, provavelmente, resultado do processo diagenético precoce, responsável por disponibilizar íons de magnésio para a geração dessas. Teores elevados de elementos traços, tais como Rb, Sr, Zr, Ba e elementos terras raras foram encontrados ao longo da amostra, estando provavelmente relacionados à presença dos microrganismos formadores do estromatólito, os quais contribuem para a captação de íons e concentração de elementos menores em sedimentos e rochas carbonáticas. O enriquecimento de δ 13C, pode ser resultado de processos de precipitação mineral influenciados pela metanogênese microbiana, ao longo da formação dessa bioestrutura. Os sinais isotópicos de δ 18O na base do estromatólito, apontam para uma influência marinha nesse sistema, todavia a presença de microgastrópodes pastadores no estrato dois indica um ambiente com influência fluvial. Sugere-se, então, uma fase de deposição em um contexto transicional já na base do estromatólito, durante a formação da fácies trombólito, seguindo até a fácies estromatólito vugular a partir da qual, houve a instalação de um ambiente lacustre restrito até o os dias atuais.
Palavras chave: fácies, paleoambiente, microbialitos, Heleobia australis

v
ABSTRACT
The main goal of this research was to characterize the different geochemical signatures of the stromatolite from Lagoa Salgada, Rio de Janeiro State, and to correlate them with the different fabrics, contributing to the refinement of the knowledge about the paleoenvironmental evolution of the lagoon. The methodology followed in this work, were macroscopic description of the sample and analyses of Inductively Coupled Plasma Mass Spectrometry (ICP-MS), X-ray Diffractometry (XRD) and Stable C & O Isotopes. The stromatolite has divided into five facies, from the base to top: microbial mat, thrombolite, vugular stromatolite, typed stromatolite and thrombolytic stromatolite. In the facies thrombolite occurs carapaces of micro gastropods of the species Heleobia australis. The different sizes of the leaflets indicate that they have not transported and trapped by the stromatolite, but have lived in the stromatolitic structure during the stromatolite development. Chemical and mineralogical studies showed that the composition of the stromatolite is not homogeneous, but varies throughout the sample. It is evident a compositional variation of trace elements in the stromatolite, as well as for the contents of ETRs. Values of δ13C vary from base to top from 2.0 to 12.78 ‰. While the δ 18O values range from 0.92 to -1.65 ‰. The variation in the chemical composition and the fabric along the facies implies possible environmental variations and the microbial communities responsible for the formation of this stromatolite. The presence of dolomite in the stromatolite base implies more active biological processes in these growth stages, while the remarkable presence of Mg-calcite in the above strata indicates that abiotic processes must be considered. Calcite with high Mg content at the top of the sample suggests that the dolomite found in the base is probably the result of the early diagenetic process, responsible for making available magnesium ions for the generation of these. High trace element contents such as Rb, Sr, Zr, Ba and rare earth elements have found throughout the sample, probably related to the presence of stromatolite-forming microorganisms, which contribute to ion uptake and concentration of minor elements in sediments and carbonate rocks. The enrichment of δ 13C may be the result of mineral precipitation processes influenced by microbial methanogenesis, during the formation of this biostructure. The isotopic signals of δ 18O at the stromatolite base. However, the presence of grazing micro gastropods in stratum two indicates an environment with fluvial influence. A phase of deposition have suggested in a transitional context already in the base of the stromatolite, during the formation of the thrombolite facies, following to the facies vugular stromatolite from which, there was the installation of a restricted lacustrine environment until now.
Keywords: facies, paleoenvironment, microbialite, Heleobia australis

vi
LISTA DE FIGURAS
Figura 1: Imagem esquemática da formação de estromatólitos.. ................................... 9
Figura 2: Mapa de localização da Lagoa Salgada, localizada no estado do Rio de
Janeiro (Fonte: Bahniuk, 2013). .................................................................................... 11
Figura 3: Lagoa Salgada, Rio de Janeiro, onde ocorrem estromatólitos modernos em
desenvolvimento atualmente. (Fonte: Adaptado de Bahniuk, 2013) ............................. 12
Figura 4: Estruturas estromatolíticas de acordo com a classificação de Aitken (1967).
(Fonte: Aitken, 1967). .................................................................................................... 14
Figura 5: Exemplos de estruturas organossedimentares que compõe o grupo dos
microbialitos. .................................................................................................................. 16
Figura 6: Classificação de microbialitos segundo sua trama, proposta por Riding
(2011). (Fonte: Riding, 2011)......................................................................................... 18
Figura 7: Classificação de Jahnert & Collins (2011) para estruturas estromatolíticas de
Hamellin Pool, Shark Bay, de acordo com suas profundidades de ocorrência nas zonas
inframaré, intermaré e supramaré. (Fonte: Jahnert & Collins, 2011) ............................. 19
Figura 8: Amostra do estromatólito proveniente da Lagoa Salgada para uso neste
trabalho. O traçado em amarelo mostra o local de seccionamento da amostra, que após
lavada apresentou um realce de suas feições internas, com destaque dos dez estratos
de amostragem definidos para coleta de material. ........................................................ 21
Figura 9: Tipos de porosidades em rochas carbonáticas proposto por Choquette & Pray
(1970). (Fonte: Adaptado de Choquette & Pray, 1970) ................................................. 22
Figura 10: Tabela periódica mostrando (em azul) os elementos químicos detectados
pela técnica de ICP-MS. (Fonte: Modificado de Bureau Veritas Minerals, 2018). ......... 24
Figura 11: Fácies definidas neste trabalho para amostra de estromatólito domal da
Lagoa Salgada, usando como critérios a textura e trama. ............................................ 26
Figura 12: Detalhes das porosidades e tramas das fácies analisadas.. ....................... 29
Figura 13: Conchas de microgastrópodes Heleobia australis encontrados no
estromatólito da Lagoa Salgada.. .................................................................................. 30
Figura 14: Ciclo de vida da população de H. australis sugerida para os exemplares
coletados no estromatólito da Lagoa Salgada. Os tamanhos das valvas (conchas)
foram medidos pelo comprimento e pelo diâmetro, em milímetros. .............................. 31
Figura 15: Dados de composição mineralógica ao longo da amostra de estromatólito, a
partir da Difratometria de Raios X. ................................................................................ 32

vii
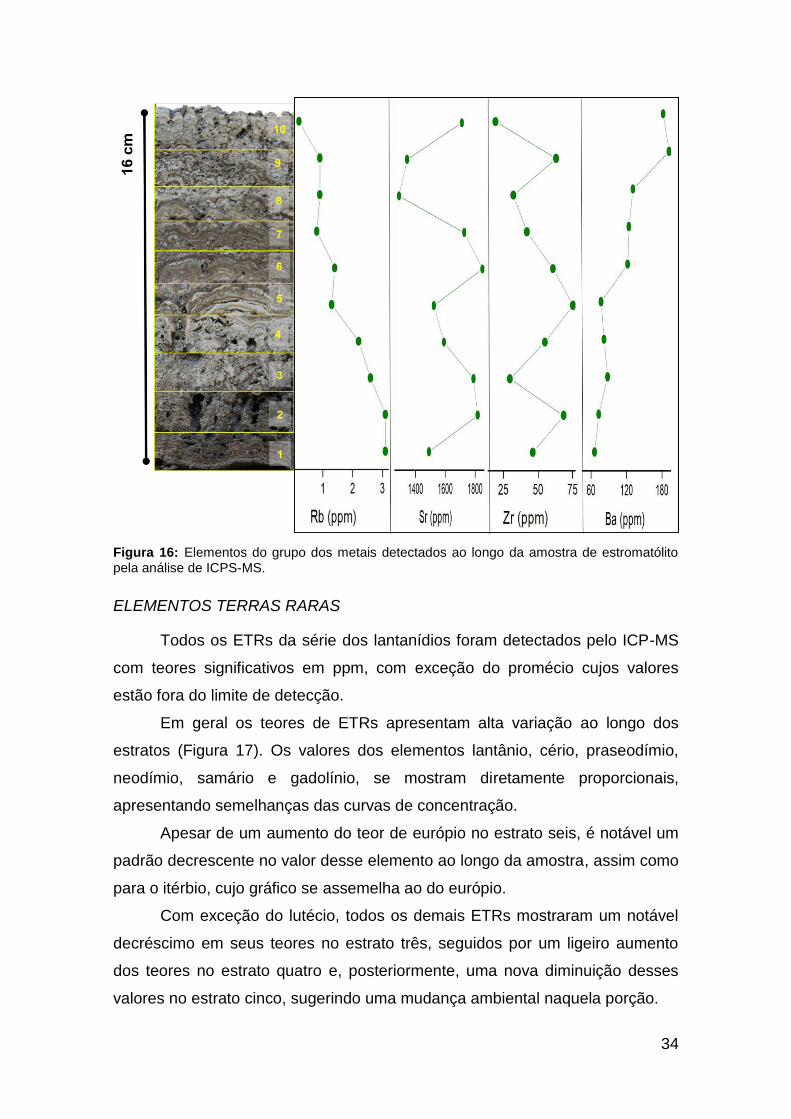
Figura 16: Elementos do grupo dos metais detectados ao longo da amostra de
estromatólito pela análise de ICPS-MS. ........................................................................ 34
Figura 17: Padrão de Elementos Terras Raras detectados por ICP-MS em cada um
dos estratos da amostra de estromatólito (à esquerda).. .............................................. 35
Figura 18: Amostras de estromatólito utilizadas no trabalho de Bahniuk (2013) e no
presente trabalho, com definição das fácies (em branco). ............................................ 37
Figura 19: Comparação dos teores de Sr encontrados no estromatólito da Lagoa
Salgada por Bahniuk (2013) e neste trabalho. .............................................................. 41
Figura 20: Diagrama de teores de ETRs na amostra de estromatólito da Lagoa
Salgada. ........................................................................................................................ 43
Figura 21: Comparação dos dados da análise de isótopos estáveis de C & O, para este
trabalho e Bahniuk (2013). ............................................................................................ 44
Figura 22: Amostras usados no trabalho de Bahniuk (2013) e neste trabalho,
mostrando as fácies e seus possíveis ambientes de formação associados. ................. 46
LISTA DE TABELAS
Tabela 1: Resultados da análise de Isótopos Estáveis de 13C e 18O. ......................... 36
Tabela 2: Comparação dos resultados mineralógicos de Bahniuk (2013) com o
presente trabalho.. ......................................................................................................... 38

viii
SUMÁRIO
1. INTRODUÇÃO ........................................................................................................ 9
1.1 ÁREA DE ESTUDO...................................................................................... 10
1.2 JUSTIFICATIVA ........................................................................................... 12
1.3 OBJETIVOS ................................................................................................. 12
2. ESTADO DA ARTE .............................................................................................. 13
3. MATERIAIS E MÉTODOS .................................................................................... 20
2.1 DESCRIÇÃO MACROSCÓPICA ..................................................................... 21
2.2 DIFRATOMETRIA DE RAIOS X ...................................................................... 22
2.3 ESPECTROMETRIA DE MASSA POR PLASMA ACOPLADO
INDUTIVAMENTE .................................................................................................. 23
2.4 ISÓTOPOS ESTÁVEIS DE C & O ................................................................... 24
4. RESULTADOS ...................................................................................................... 25
4.1. DESCRIÇÃO MACROSCÓPICA .................................................................... 25
4.2. ESTUDOS MINERALÓGICOS........................................................................ 31
4.3. ESTUDOS QUÍMICOS .................................................................................... 33
4.4. ESTUDOS ISOTÓPICOS ............................................................................... 36
5. DISCUSSÃO ......................................................................................................... 36
5.1 FORMAÇÃO DAS TRAMAS DO ESTROMATÓLITO ...................................... 37
5.2 EVOLUÇÃO PALEOAMBIENTAL .................................................................... 43
6. CONCLUSÕES ..................................................................................................... 46
REFERÊNCIAS ......................................................................................................... 49
APÊNDICE I – Tabela de Dados Químicos a partir da Análise de ICP-MS.......... 55

9
1. INTRODUÇÃO
Burne & Moore (1987), em seu trabalho clássico, definiram microbialitos
como estruturas organossedimentares que se formam por consequência da
interação de comunidades de microorganismos bentônicos que aprisionam
sedimentos detríticos e/ou precipitam sedimentos químicos (Figura 1). Brehm
et al. (2004) englobaram no grupo de microbialitos os estromatólitos,
trombólitos, esteiras microbianas, oncoides e oóides. Então, Riding (2011)
atualizou a definição de microbialitos para sedimentos bentônicos produzidos
pela indução de microorganismos, e englobou nesse grupo além dos
estromatólitos, trombólitos e dendrólitos, todas as estruturas similares que
ocorrem como domos e colunas em ambientes de águas rasas e são formadas
pelo mesmo processo.
Figura 1: Imagem esquemática da formação de estromatólitos. a) amostra de estromatólito recente de Shark Bay, Austrália, mostrando a estruturação interna laminada; b) interação de microrganismos e sedimentos durante a formação da estrutura estromatolítica (Fonte: Adaptado de Grotzinger & Jordan, 2013).
Estromatólitos podem ser distinguidos dos demais microbialitos com
base na sua estruturação interna, sendo aqueles depósitos microbiais que
apresentam uma laminação (Riding, 1991). Conforme o mesmo autor,
estromatólitos são formados através de processos distintos (aprisionamento,

10
biomineralização ou mineralização) que agiram no passado geológico em
determinados ambientes que hoje são raros.
O arcabouço dos estromatólitos pode apresentar estruturas internas
(trama) distintas entre si, as quais são resultantes de processos microbianos de
captura e ligação, precipitação induzida por carbonato, teor de matéria
orgânica, quantidade de entrada de sedimentos, presença de vazios, presença
de esqueletos, bioturbação e orientação macroporosa (Burns et al., 2004).
Essa variação interna também está relacionada ao local de deposição, por
exemplo, da zona supramaré até a inframaré, pois o substrato é colonizado por
diferentes comunidades bacterianas, expressadas em morfologias superficiais
características para os microbialitos.
Estromatólitos recentes podem ser encontrados em desenvolvimento em
poucos lugares do planeta, tais como Shark Bay (Austrália), Solar Lake (Israel),
Salt Lake (Estados Unidos), Ilha de Hai-Nan (China) e na Lagoa Salgada
(Brasil), permitindo que estas ocorrências modernas sejam estudadas como
análogas aos estromatólitos antigos (Bahniuk, 2013). A reconstrução
paleoambiental do registro fossilífero estromatolítico indicando condições
ambientais que prevaleciam no passado representa a relevância geológica dos
estromatólitos (Silva e Silva et al., 2004).
1.1 ÁREA DE ESTUDO
Para o presente trabalho foram utilizadas amostras de estromatólitos
atuais coletadas da Lagoa Salgada, localizada no sudeste do Brasil, na região
litorânea nordeste do estado do Rio de Janeiro, próximo ao município Campo
dos Goytacazes, cujas coordenadas são 41º00’30’’W e 21º54’10’’S (Figura 2).
As vias de acesso à lagoa são a rodovia BR-101 a partir da cidade do
Rio de Janeiro, sentido município de Campos, por cerca de 290 km, e então
pela rodovia RJ-216 sentido a Cabo de São Tomé, por 50 km. O restante do
trajeto é feito por uma estrada secundária de terra, por 20 km, atravessando os
vilarejos de Santa Rosa e Quixaba, sentido Barra do Açu (Iespa, 2010).

11
Figura 2: Mapa de localização da Lagoa Salgada, localizada no estado do Rio de Janeiro (Fonte: Bahniuk, 2013).
A Lagoa Salgada tem uma área de 16 km², comprimento de 8,6 km e
largura máxima de 1,9 km com o eixo principal NW-SE (Lemos et al., 1994).
Esta laguna integra o Complexo Deltaico do Rio Paraíba do Sul, estando
situada em uma planície arenosa cuja origem está associada às oscilações do
nível do mar durante o Holoceno (Martin et al, 1993). Segundo Srivastava
(1999), essa lagoa hospeda as únicas ocorrências bem desenvolvidas de
estromatólitos carbonáticos colunares, domais e estratiformes de idade
holocênica no Brasil. A ocorrência destas no estado do Rio de Janeiro é
favorecida diretamente pela presença de cianobactérias e condições físico-
químicas extremas (hipersalinas) associadas à sedimentação carbonática
(Damazio, 2004).
Na margem e no interior da Lagoa Salgada ocorrem estromatólitos
estratiformes e domais, oncoides, trombólitos e esteiras microbianas (Iespa et
al., 2012). Devido a ocorrência dessas estruturas, essa lagoa se assemelha
àquelas do sistema lagunar de Araruama, Rio de Janeiro, como proposto por
Iespa (2010). Bahniuk (2013), identificou exemplares modernos, ocorrentes
principalmente na borda sudoeste da lagoa, como domais e do tipo couve-flor
(Figura 3).

12
Figura 3: Lagoa Salgada, Rio de Janeiro, onde ocorrem estromatólitos modernos em desenvolvimento atualmente. a) visão geral da lagoa; b) ocorrência de estromatólito tipo domal. (Fonte: Adaptado de Bahniuk, 2013)
1.2 JUSTIFICATIVA
O estudo de estromatólitos se mostra muito importante, pois tais
estruturas são consideradas o registro mais antigo de vida na Terra, chegando
a idades de até 3,5 bilhões de anos (Awramik, 1992; McNamara & Awramik,
1992; Hofmann et al., 1999). Esse estudo pode ser aplicado a diversas áreas,
uma vez que auxilia nas interpretações de paleoambientes, paleosalinidades,
paleolatitudes, correlações bioestratigráficas, datação de rochas,
micropaleontologia, direções de paleocorrentes e determinação de topo e base
de sequências deformadas (Hofmann, 1973). Portanto, estromatólitos são
potenciais estruturas do registro de processos geoquímicos, ambientais e
atmosféricos em suas tramas, e revelam-se importantes bioassinaturas de
paleoambiente. Dessa forma o estudo de sua estruturação interna conciliado
com parâmetros geoquímicos, configura uma forma de entender as diferentes
condições de formação dessas bioestruturas.
1.3 OBJETIVOS
Este trabalho objetivou caracterizar as diferentes assinaturas
geoquímicas dos estratos do estromatólito e correlacioná-las com as distintas
tramas encontradas, buscando contribuir para o refinamento do conhecimento
da evolução paleoambiental da Lagoa Salgada, em especial na porção superior
onde existe potencial influência antrópica.

13
2. ESTADO DA ARTE
Ernst Kalkowsky, em 1908, cunhou os termos estromatoide e
estromatólito, usados em paralelo com ooide e oólito, mas não deu uma
definição abrangente sobre estromatólitos (Riding, 1999). Entretanto, em seu
trabalho pioneiro, Kalkowsky enfatizou dois aspectos dos estromatólitos: sua
estrutura laminada e a natureza orgânica. De acordo com Riding (1999) é
razoável concluir que Kalkowsky considerava os estromatólitos essencialmente
como estruturas microbianas laminadas e tinha uma visão semelhante da
origem dos ooides.
Durante um longo período de tempo não se teve trabalhos sobre o novo
termo criado em 1908. Foi na década de sessenta que surgiram então novos
debates a respeito do termo estromatólito, quando o termo foi de fato aceito
(Riding, 1999). Hofmann (1969) atualizou o termo estromatólito para rochas
estruturadas por uma sucessão de lâminas, as quais representam intervalos de
acumulação de sedimentos finos em superfícies supostamente povoadas por
uma comunidade de microorganismos. O autor ainda propõe que material
sedimentar seria acumulado por aprisionamento ou aglutinação no plexo
orgânico, ou por precipitação não-esquelética resultante da atividade
metabólica dos microorganismos.
Logan (1964) sugere que o substrato de formação de microbialitos,
normalmente ambiente litorâneo protegido ou lagos continentais, estaria sujeito
a fatores físico-químicos que seriam responsáveis por moldar e modificar
lateralmente as estruturas estromatolíticos. Tais fatores seriam, entre outros:
episódios periódicos de seca e inundações, devido às flutuações das águas de
marés; prolongados períodos de dessecação durante marés baixas em
ambientes marinhos, ou evaporação de lagos rasos e salinos; limpeza e
fragmentação mecânica por ondas de tempestade; lavagem e canalização por
escoamento de água das marés; enterro pelo influxo de sedimentos no terreno
da esteira; e atividade biótica.
Aitken (1967) sugeriu classificações para as estruturas estromatolíticas
conforme sua morfologia externa: estromatólitos colunares, são
hemisferoides ou paraboloides, ramificados ou não, empilhados verticalmente e

14
arqueados, de projeção constante ou variável, formando colunas cilíndricas,
podendo ser agrupados em estromatólitos compostos isolados em forma de
torre com paredes verticais (Figura 4a); domais, corpos simples ou compostos,
em forma de colmeia, geralmente de baixo relevo, raramente excedendo 25
centímetros de altura, sendo o contorno no plano comumente circular para
elíptico, além de apresentarem lâminas regulares muito finas, de superfícies
lisas e simples ou lâminas irregulares, com superfícies irregulares ou
onduladas (Figura 4b); digitados, são compostos de colunas ou dedos de
hemisferoides de baixo ângulo, diferenciando-os dos colunares (Figura 4c); e
poligonais, possuem orientação convexa para baixo das lâminas
hemisferoidais proeminentes e muito regulares, sendo esses hemisférios
empilhados em colunas curtas, raramente mais do que alguns centímetros de
altura (Figura 4d).
Figura 4: Estruturas estromatolíticas de acordo com a classificação de Aitken (1967). a) estromatólitos colunares em calcários pré-cambrianos do Glacier National Park, Estados Unidos (Fonte: Commons Wikimedia); b) Estromatólito domal de idade holocênica da Lagoa Salgada, Brasil (Fonte: Srivastava, 1991); c) Estromatólito digitado da Formação Oneota, em Tomah, Estados Unidos (Fonte: Aitken, 1967); d) estromatólito digitado em dolomitos, Formação Lyell, Pico Nestor, Canadá (Fonte: Aitken, 1967).

15
Awramik & Margulis (1974) deram um passo significativo por sugerir que
os microrganismos envolvidos na formação de estromatólitos fossem
principalmente algas verde e azuis. Porém, os autores abandonaram a
laminação como um caráter essencial dos estromatólitos, pois sua pesquisa
revelou que alguns depósitos microbianos não eram laminados ou apenas
forma grosseira, por exemplo, os depósitos de Shark Bay, que haviam sido
descritos por Logan em 1961.
Ocorreu então uma abrangência do termo estromatólito para qualquer
estrutura organomineral microbiana, independentemente da estrutura interna,
permitindo que Burne & Moore (1987), criassem o termo microbialito. Segundo
os autores microbialitos são estruturas organossedimentares que se
acumularam como resultado da interação de comunidades microbianas
bentônicas, as quais aprisionam sedimentos detríticos e/ou induzem a
precipitação de sedimentos químicos. Brehm et al. (2004), por fim, englobaram
no grupo de microbialitos os estromatólitos, trombólitos, esteiras microbianas,
oncoides e oóides.
As esteiras microbianas são um grupo de estruturas
organossedimentares laminadas e constituídas principalmente por
cianobactérias (Figura 5a). A microbiota construtora das esteiras tem a
capacidade de migrar verticalmente para escapar ao enterro pelos sedimentos
e recolonizar a superfície recém-depositada, podendo atingir vários centímetros
até decímetros de espessura (Gerdes et al., 1993). Considera-se que estas
esteiras formam o primeiro estágio de desenvolvimento dos estromatólitos,
sendo o primeiro substrato fixo necessário para o crescimento estromatolítico
(Iespa, 2010).
Segundo Aitken (1967), os trombólitos são estruturas
organossedimentares derivadas do aprisionamento de sedimentos por
cianobactérias, sem haver a formação de uma estrutura laminada, mas sim
coagulada ou esponjosa (Figura 5b). Podem interagir com os estromatólitos à
medida que os coágulos individuais se tornam distintamente laminados,
gerando formas intermediárias designadas de trombolíticas-estromatolíticas.
Além de ser recorrente a ocorrência de trombólitos graduados para cima
encerrados por estromatólitos domais. Além disso, tem-se o estromatólito

16
trombolítico, tipificado pelo mesmo autor como fracamente coagulado e
grosseiramente laminado.
Figura 5: Exemplos de estruturas organossedimentares que compõe o grupo dos microbialitos. a) esteira microbiana laminada mostrando uma variação de cor, resultado dos diferentes microrganismos formadores (Fonte: Grotzinger & Jordan, 2013); b) trombólito gigante do Lago Sarmiento, Parque Nacional Torres Del Paine, Chile. (Foto: Veleda Astarte)
Hofmann (1973) propõe alguns requisitos necessários para o
desenvolvimento estromatolítico: presença de um substrato no qual os
estromatólitos podem crescer; um sistema aberto contendo água, servindo
como meio de crescimento; presença de compostos que satisfazem exigências
metabólicas de elementos de uma microbiota; fonte de energia para continuar a
atividade metabólica, como luz solar; população de algas ou outros
microrganismos bentônicos; matéria mineral de uma determinada faixa de
tamanho pequeno que pode ser retida e ligada e/ou precipitada nos plexos; um
ritmo, periódico ou episódico, que prevê a descontinuidade dos processos de
acresção resultando no desenvolvimento da estrutura laminada; e litificação e
enterramento, permitindo a preservação do registro.
Estabeleceu-se que estruturas estromatolíticas se formam em ambientes
marinhos e não marinhos (McNamara & Awramik, 1992). Entretanto, ambientes
carbonáticos hipersalinos de águas rasas são os locais mais propícios ao seu
desenvolvimento (Awramik, 1992), pois são ricos em cálcio e bicarbonato,
permitindo a formação do carbonato de cálcio como matéria prima para o
desenvolvimento estromatolítico (McNamara & Awramik, 1992).
Os exemplares vivos de estromatólitos se desenvolvem em áreas com
uma redução de atividade faunística, e sem a ocorrência de macroalgas e
plantas (McNamara & Awramik, 1992). De acordo com estes autores, essas

17
áreas apresentam águas com altas salinidade e alcalinidade, baixas taxas de
nutrientes, temperaturas elevadas, dessecação periódica e outras mudanças
sazonais que favorecem a precipitação mineral durante o crescimento do
estromatólito.
Os microrganismos formadores dos microbialitos são
predominantemente cianobactérias, bactérias sulfato redutores, metano
oxidantes e pequenas algas, as quais ocupam uma variedade de ambientes,
incluindo águas de composição química muito diferentes, e seu envolvimento
na sedimentação é igualmente variada. (Riding, 2011). Segundo o mesmo
autor, do ponto de vista da biocalcificação, os carbonatos de microbialitos são
bioinduzidos em águas menos saturadas para minerais, enquanto que em
ambientes onde a precipitação inorgânica é favorecida, a contribuição
sedimentar geral das bactérias acaba sendo mais na trama do que o processo
de deposição.
Riding (2011) apresentou um modelo de diferentes microbialitos
definidos com base na sua trama (Figura 6): estromatólito (laminado),
trómbolito (coagulado), dendrólito (dendrítico) e leilólito (afanítico). Os
microrganismos estariam associados diretamente a estas tramas. Entretanto,
uma complicação é que as tramas de leilólitos e estromatólitos são definidas
por características internas do carbonato microbiano, tal como laminação. Em
contraste, as tramas de trombólitos e dendrólitos são definidas pela forma
externa de massas individuais de carbonato microbiano, tais como coágulos ou
pequenas massas semelhantes a arbustos.

18
Figura 6: Classificação de microbialitos segundo sua trama, proposta por Riding (2011).
(Fonte: Riding, 2011)
Trabalhos mais recentes, como de Jahnert & Collins (2011) trouxeram
novos modelos de estromatólitos, baseado nas ocorrências modernas de Shark
Bay, Austrália, e sua distribuição específica em relação à profundidade da água
(Figura 7). Os autores referidos mostraram que diferentes aspectos externos e
características internas dos estromatólitos estão associadas à atividade distinta
da comunidade microbiana e à disponibilidade de sedimentos finos nos
distintos nível de ocorrência. As classificações de Jahnert & Collins (2011), da
ocorrência mais rasa para a mais profunda na lâmina de água, nas zonas de
intermaré e inframaré, são: trombólito pustular (coagulado), estromatólito liso
(laminado) ou coloforme (ondulado), cerebroides (não laminado) e tapetes
microbianos (sub-laminado irregular).

19
Figura 7: Classificação de Jahnert & Collins (2011) para estruturas estromatolíticas de Hamellin Pool, Shark Bay, de acordo com suas profundidades de ocorrência nas zonas inframaré, intermaré e supramaré. (Fonte: Jahnert & Collins, 2011)
Diversos trabalhos foram feitos a respeito dos estromatólitos recentes da
Lagoa Salgada a partir do ano de 1999, quando Srivastava publicou seu
trabalho registrando a lagoa em questão como um sítio paleoambiental que
abriga estruturas estromatolíticas atuais. Surgiram trabalhos cujo foco foi a
caracterização e classificação das estruturas organo-sedimentares encontradas
na lagoa: Silva e Silva et al (2005; 2007; 2008; 2013); Iespa, et al (2008);
Cataldo (2011). Gomes (2011); Silva e Silva et al. (2013) identificaram ainda a
composição bacteriana formadora das estruturas e relacionaram as fácies de
estromatólitos com trombólitos e oncoides recentes, também ocorrentes na
Lagoa Salgada. A partir de 2012 alguns trabalhos (Servidoni, 2012; Rezende et
al., 2013; Hayashi, 2014 e Pacheco, 2014) deram uma nova aplicação ao
estudo dos estromatólitos modernos da Lagoa Salgada, sendo estes usados
como análogos às rochas de reservatórios microbiais carbonáticos de
hidrocarbonetos.
Bahniuk (2013) utilizou métodos geoquímicos de isótopos estáveis e de
cumpled para investigar a formação e evolução dos estromatólitos da lagoa,
além de determinar taxas de crescimento, com base em datação de
radiocarbono, associadas ao paleoambiente. No referido trabalho foi proposto
uma taxa de crescimento para os estromatólitos daquele sítio como sendo de
0,1 mm/ano. Além disso, foram definidas três principais fases de formação das
lâminas estromatolíticas: a fase inicial há cerca de 2300 anos BP, em um corpo

20
de água aberto ao oceano com uma influência terrestre; uma fase transicional
entre 1980 e 1130 anos BP; e a fase final, datada entre 592 e 200 anos BP,
representando um sistema restrito, totalmente microbiano.
Outros trabalhos (Iespa, et al., 2012 e Pereira, 2014) também visaram
compreender a evolução paleoambiental da Lagoa Salgada utilizando as fácies
estromatóliticas. Esses exemplares vivos brasileiros continuam sendo
importantes objetos de estudo dentro da geologia. Trabalhos mais recentes
estão inovando nos estudos, como visto em Birgel, et al (2015), que propõe
uma indução abiótica para a formação daqueles; Callefo (2018) que estudou
biominerais como forma de entender as bioassinaturas microbianas; e Silva et
al. (2018), que mapearam os tipos de microbialites, identificando danos e
ameaças e fornecendo sugestões de geoconservação.
3. MATERIAIS E MÉTODOS
Foram coletadas pelos professores doutores Anelize Manuela Bahniuk
Rumbelsperger e Crisogono Vasconcelos, amostras de estromatólitos recentes
da Lagoa Salgada, durante campanha de campo realizada no ano de 2011. As
amostras foram enviadas ao Instituto Federal de Tecnologia de Zurique
(ETHZ), Suíça, e ao Laboratório de Análise de Minerais e Rochas (LAMIR),
para uso no trabalho de doutorado de Bahniuk (2013), desenvolvido no ETHZ e
no Instituto de Tecnologia da Califórnia (Caltech), Estados Unidos.
Uma das amostras, cujas dimensões são de 16 cm de altura por 20 cm
de diâmetro, foi selecionada para a realização da descrição macroscópica e de
análises químicas necessárias para este trabalho (Figura 8). O preparo inicial
desta se deu por seccionamento vertical e lavagem com água para melhor
expressão de suas feições estruturais e texturais, auxiliando na descrição do
estromatólito. Posteriormente, cerca de 4,0 g de material pulverizado foi
amostrado de dez estratos do estromatólito para a realização das análises de
Espectrometria de Massa por Plasma Acoplado Indutivamente (ICP-MS),
Difratometria de Raios X (DRX) e Isótopos Estáveis de C & O.

21
Figura 8: Amostra do estromatólito proveniente da Lagoa Salgada para uso neste trabalho. O traçado em amarelo mostra o local de seccionamento da amostra, que após lavada apresentou um realce de suas feições internas, com destaque dos dez estratos de amostragem definidos para coleta de material.
2.1 DESCRIÇÃO MACROSCÓPICA
Com base nas características texturais, na trama e na porosidade das
camadas do estromatólito, dividiu-se a amostra, da base para o topo, em cinco
fácies que foram descritas macroscopicamente e com auxílio de
Estereomicrocópio Zeiss, modelo V8, no LAMIR.
Para a classificação de porosidade, foi utilizado neste trabalho os tipos
de porosidades estabelecidos por Choquette & Pray (1970), para rochas
carbonáticas, classificadas dentro de tramas seletivas e não-seletivas de
acordo com a gênese dos poros (Figura 9). De acordo com os autores, a
porosidade é seletiva quando existe uma relação específica entre os poros e os
elementos da trama (poros intrapartículas, entrepartículas, do tipo fenestral,
shelter, em crescimento, entrecristais e móldicos) e não-seletiva quando não
existe essa relação (poros de fratura, em canais, vugulares e em cavernas).

22
Figura 9: Tipos de porosidades em rochas carbonáticas proposto por Choquette & Pray (1970).
(Fonte: Adaptado de Choquette & Pray, 1970)
Na descrição macroscópica encontrou-se bioclastos associados a uma
das fácies, os quais foram identificados como carapaças de microgastrópodes.
Estes organismos foram separados da amostra estromatolítica para evitar
interferência nos resultados das análises químicas. Os bioclastos foram
descritos por estereomicroscopia e contabilizados conforme o seu possível
estágio de vida.
2.2 DIFRATOMETRIA DE RAIOS X
A identificação da composição mineralógica das amostras aqui
estudadas se deu através da técnica de DRX. Trata-se do processo pelo qual
os raios X são dispersos pelos elétrons dos átomos dos cristais, sem que haja
uma mudança do comprimento de onda (dispersão coerente), quando as
condições geométricas são satisfeitas (Gomes, 1984). A difração de um cristal
corresponde a uma propriedade física fundamental da substância, uma vez que
compreende a posição e intensidade das linhas de difração desse cristal, por

23
isso acaba servindo para identificação e também estudo da estrutura (Gomes,
1984).
Esta análise foi realizada no LAMIR com a utilização do difratômetro
PANalytical e modelo EMPERYAN. Os difratogramas foram interpretados com
base na porcentagem semi-quantitativa dos minerais pelo método RIR
(Reference Intensity Ratio – Razões das Intensidades de Interferência), com
auxílio do software X’Pert High Score Plus Data Collector, com banco de dados
PDF-2.
2.3 ESPECTROMETRIA DE MASSA POR PLASMA ACOPLADO
INDUTIVAMENTE
A técnica analítica de ICP-MS foi usada com o intuito de caracterizar a
composição química das amostras. Tal análise é bastante utilizada em estudos
geoquímicos devido a sua capacidade multielementar, seu baixo limite de
detecção possível, ampla faixa dinâmica linear e a rápida determinação das
razões isotópicas utilizando uma pequena quantidade de amostra (Houk et al.,
1992). Um dos principais benefícios do IPC-MS está no fato desta técnica
atingir baixos limites de detecção em soluções aquosas, uma vez que
aproximadamente 90% dos elementos químicos podem ser determinados em
um intervalo de 0,1 a 0,001 ng/mL (Veiga, 2000).
A análise de ICP-MS aqui apresentada foi realizada no laboratório
Bureau Veritas Minerals, em Vancouver, Canadá. Foram necessárias
quantidades de amostras de 1,0 g, que passaram por um preparo de digestão
ácida por água régia modificada (1:1:1, HNO3:HCl:H2O), reduzindo a amostra
em 0,5 g para a leitura química do material. O ICP-MS é capaz de detectar
diversos elementos da tabela periódica (Figura 10), entretanto, no presente
trabalho identificou-se teores significativos de metais de transição e ETRs.

24
Figura 10: Tabela periódica mostrando (em azul) os elementos químicos detectados pela
técnica de ICP-MS. (Fonte: Modificado de Bureau Veritas Minerals, 2018).
2.4 ISÓTOPOS ESTÁVEIS DE C & O
A análise de isótopos estáveis de C & O em rochas carbonáticas é feita
pela variação das relações isotópicas 18O/16O e 13C/12C da amostra em relação
a um padrão internacional (Neumann et al., 2010). Os padrões mais aceitos
para carbonatos são o PDB (Belemnites da Formação Pee Dee de idade
Cretácea, nos Estados Unidos) usado para o oxigênio e o carbono, e o SMOW
(“Standard Mean Ocean Water”, Craig 1961) para o oxigênio. Portanto os
valores δ analisados pode ser positivo, quando a amostra é mais rica em
isótopos mais pesados que a amostra padrão; zero, quando a amostra
apresenta um valor igual ao padrão; e negativos, quando a amostra está
enriquecida em isótopos mais leves que na amostra padrão (Neumann et al.,
2010).
A análise isotópica foi realizada no LAMIR, através do equipamento
Espectrômetro de Massa Thermo-Scientifique, modelo Delta V Advantage
Isotope Ratio MS, acoplado a um sistema Gas Bench 2, sendo a leitura dos
dados feita a partir do software Isodat. Para esta análise foram necessárias
aproximadamente 5,0 mg de amostra pulverizada, após secagem em estufa à
70º C, por 24 horas.

25
4. RESULTADOS
Neste capítulo será apresentado os dados e resultados obtidos a partir
da descrição e análises da amostra, dessa forma as discussões acerca destes
resultados serão realizadas somente no capítulo 5.
A morfologia externa do estromatólito usado neste trabalho foi
classificada de acordo com Aitken (1967), como sendo um estromatólito do tipo
domal (Ver Capítulo 2). Em termos descritivos a amostra foi dividida em cinco
fácies, da base para o topo, levando em conta suas características texturais e
tramas. Para as análises químicas, foram retiradas amostras de dez estratos
de amostragem cujos resultados serão apresentados conforme a disposição
vertical desses, sendo o mais antigo (estrato um) o mais basal e o mais recente
(estrato dez) o mais superior.
4.1. DESCRIÇÃO MACROSCÓPICA
Os critérios para a distinção das cinco fácies foram as características
texturais e as tramas: micro e macrolaminações, bioerosão, cavidades
vugulares e fenestras, que estão por sua vez associados a porosidade dos
estratos.
Considerando as classificações para microbialitos estabelecidas na
literatura, as cinco fácies definidas neste trabalho foram: esteira microbiana,
trombólito, estromatólito vugular, estromatólito digitado e estromatólito
trombolítico (Figura 11), as quais serão descritos a seguir. As classificações
adotadas neste trabalho foram aquelas propostas por Gerdes et al. (1993),
para esteiras microbianas, e por Riding (2011), para estromatólitos e
trombólitos.

26
Figura 11: Fácies definidas neste trabalho para amostra de estromatólito domal da Lagoa Salgada, usando como critérios a textura e trama. Foram consideradas as classificações de Gerdes et al. (1993), para esteiras microbianas, e Riding (2011), para estromatólitos e trombólitos
FÁCIES ESTEIRA MICROBIANA
A primeira fácies corresponde ao primeiro estrato de amostragem, o qual
encontra-se na base do estromatólito e tem espessura de 3,0 cm. Apresenta
uma estruturação estratiforme, marcada por uma laminação contínua e
ondulada, evidenciada pela alternância de cores castanho claro, composta por
carbonato, e castanho avermelhada, provável devido à alta concentração de
matéria orgânica. Ocorre porosidade do tipo fenestral e, localmente, do tipo
caverna (Figura 12a). A passagem da fácies basal, esteira microbiana, para a
seguinte se deu de forma abrupta uma vez que a trama dessas são bastante
distintas.
FÁCIES TROMBÓLITO
A segunda fácies corresponde ao estrato de amostragem dois, cuja
espessura é de 4,5 cm, sendo o mais espesso da amostra. A cor em geral é
castanho levemente avermelhado. A trama não apresenta laminação, mas um

27
desarranjo na estrutura, formando coágulos e cavidades que se irterligam, além
de ter um aspecto friável. Está fácies constitui a porção mais porosa da
amostra, prevalecendo a porosidade do tipo caverna (Figura 12a). Além disso é
recorrente a presença de bioclastos preenchendo as cavidades, os quais foram
identificados como valvas de microgastrópodes. A ocorrência destes evidencia
uma bioerosão bastante significativa no estrato, gerando uma estruturação
maciça. A espécie dos microgastrópodes foi identificada como Heleobia
australis e será descrita posteriormente à descrição das fácies, neste capítulo.
FÁCIES ESTROMATÓLITO VUGULAR
Está fácies compreende o estrato de amostragem três e quatro,
apresentando 2,0 cm de espessura. A coloração é castanho claro e a estrutura
é em geral maciça, mas uma laminação descontínua ocorre de forma incipiente
no estrato. Não há presença de bioclastos e a porosidade é do tipo vugular e
localmente fenestral (Figura 12a).
FÁCIES ESTROMATÓLITO DIGITADO
Está fácies compreende os estratos de amostragem 5, 6, 7, 8 e sua
espessura é de 3,0 cm. Apresenta uma intercalação de cores castanho claro e
castanho avermelhado. A trama é composta dor microlaminações onduladas e
descontínuas, formando colunas ou formas de dedos, sugerindo sua
classificação digitada (Figura 12a). Essas colunas apresentam larguras de 0,5
até 3,2 cm, variando em tamanhos, da base para o topo da fácies, de forma
crescente. A porosidade encontrada na base da fácies é fenestral, e se
desenvolve acompanhando o topo das formas colunares. Entretanto, nos
estratos acima quase não há porosidade visível macroscopicamente, em geral
a fácies é bastante cimentada. Além disso, os espaços entre as colunas das
laminações estão preenchidos majoritariamente por sedimentos detríticos
cimentados possivelmente por carbonato.
FÁCIES ESTROMATÓLITO TROMBOLÍTICO
A fácies do topo do estromatólito constitui-se dos estratos de
amostragem nove e dez, cuja espessura varia de 3,5 cm, no centro, e 2,0 cm,
na margem da amostra. Há uma variação também na coloração da fácies, de

28
castanho claro a castanho avermelhado. A estrutura é difusa, sem um arranjo
definido da trama. Contudo, ocorrem porções levemente laminadas e
onduladas, sendo estas descontínuas e formando, localmente, micro colunas
espaçadas nas lâminas, semelhante a fácies de estromatólito digitado (Figura
12a). A porosidade móldica é bastante evidente na margem dos estratos,
sendo possível reconhecer moldes de valvas dos microgastrópodes
encontrados na fácies trombólito (Figura 12b).

29
Figura 12: Detalhes das porosidades e tramas das fácies analisadas. a) na esteira microbiana predomina porosidade fenestral (em branco) e a trama é laminada e ondulada; no trombólito não há um arranjo na estrutura, mas sim cavidades e coágulos, sendo a porosidade do tipo caverna (em azul); no estromatólito vugular, ocorrem microlaminações descontínuas e a porosidade é vugular (em amarelo); no estromatólito digitado ocorre uma trama de colunas espaçadas, com porosidade fenestral na porção inferior e baixa porosidade macroscópica acima; no estromatólito trombolítico ocorre um desarranjo na trama, com localizadas colunas, e a porosidade que predomina é a móldica (área destacada em verde); b1, b2, b3) fotos destaques da margem superior da amostra, correspondente à fácies estromatólito trombolítico cuja porosidade de ocorrência é a móldica. É possível reconhecer os moldes das valvas dos mesmos microgastrópodes que ocorrem na fácies trombólito, Heleobia australis.
BIOCLASTOS DE MICROGASTRÓPODES
Os bioclastos encontrados nas cavidades da fácies trombólito (Figura
13a) e também formando a porosidade móldica da fácies estromatólito
trombolítico correspondem a carapaças de microgastrópodes da espécie
Heleobia australis (d’Orbigny, 1835). Trata-se de um molusco cujo habitat é a
zona entremarés, pois vivem em ambientes protegidos da ação direta do mar,
como estuários e lagoas costeiras (Fiori & Carcedo, 2011). Os H. australis tem
geralmente comprimento total <7,0 mm e constituem um dos principais
componentes bióticos da fauna em estuários, além de possuir uma distribuição
geográfica ampla, do sudeste brasileiro até o sul da Argentina (Gaillard &
Castellanos, 1976, apud Francesco & Isla, 2004).
Foram coletados 500 exemplares de conchas de H. australis (Figura
13b), as quais apresentam cor branca, com exemplares translúcidos, e brilho
perolado (Figura 13c). Não apresentam sinais de tração ou arrasto,
encontrando-se majoritariamente inteiras. Algumas carapaças apresentam
perfurações concêntricas, possíveis evidências de bioerosão. As conchas
variam em tamanhos de 1,0 mm até 4,0 mm de comprimento com diâmetros
variando de 1,0 mm até 2,0 mm (Figura 13d). Os exemplares de tamanhos
diferentes ocorrem juntos nas cavidades dos estratos.

30
Figura 13: Conchas de microgastrópodes Heleobia australis encontrados no estromatólito da Lagoa Salgada. a) concha de H. australis encrustada nas cavidades da fácies trombólito; b) amostras coletadas para descrição e identificação; c) exemplar de H. australis translúcido, com brilho sedoso; d) variação dos tamanhos das conchas, mostrando que ocorrem exemplares de idades diferentes ocorrem juntos.
Os ciclos de vida dos H. australis são estimados por meio de análise da
distribuição tamanho-frequência (Figueiredo-Barros et al., 2006). Tais ciclos
podem ser influenciados por variações na salinidade e na temperatura, e
também na produção de organismos primários.
A Figura 14 mostra a distribuição em tamanho pela frequência das
conchas de H. australis encontradas na amostra do presente trabalho. O valor
amostral da análise é de 500 exemplares. Os tamanhos de conchas
encontrados variaram de 1,0 x 1,0 mm (comprimento por diâmetro) até 4,0 x
2,0 mm. A curva de frequência no gráfico (em vermelho) mostra uma
população jovem com um provável ciclo de vida curto durante o
desenvolvimento estromatolítico.

31
Figura 14: Ciclo de vida da população de H. australis sugerida para os exemplares coletados no estromatólito da Lagoa Salgada. Os tamanhos das valvas (conchas) foram medidos pelo comprimento e pelo diâmetro, em milímetros.
4.2. ESTUDOS MINERALÓGICOS
A análise de DRX mostrou que a composição mineralógica do
estromatólito não é homogênea, mas varia ao longo da amostra. A calcita
magnesiana (Mg-Calcita) é o mineral mais abundante (valores médios de
66,9%), seguido do quartzo e da dolomita.
É possível observar que os estratos basais apresentam uma mistura de
minerais de quartzo, Mg-calcita e dolomita. Enquanto que os estratos mais ao
topo do estromatólito são exclusivamente formados por Mg-calcita, com
presença de quartzo, além de ocorrer uma variação desses dois minerais ao
longo da amostra (Figura 15).

32
Figura 15: Dados de composição mineralógica ao longo da amostra de estromatólito, a partir da Difratometria de Raios X.
Apesar do quartzo ter sido detectado em todos os estratos amostrados,
suas porcentagens não são maiores que aquelas encontradas para a Mg-
calcita, que por sua vez, ocorre em nove dos dez estratos. Somente o primeiro
estrato (mais basal) não aponta a presença de Mg-calcita.
Os teores mais elevados de quartzo aparecem nos estratos dois, três e
quatro, sendo respectivamente, 31, 39,4 e 34%. Em contraste com a Mg-calcita
que mostra seus menores valores nesses mesmos estratos de abundância
quartzosa: 19, 19,2 e 66%. Nos estratos do topo da amostra (nova e dez)
ocorrem os maiores teores de Mg-calcita (91 e 90%).

33
A dolomita ocorre somente nos três primeiros estratos basais, mas com
teores elevados (69, 50 e 41,4%), promovendo a diminuição nos teores de Mg-
calcita, e elevação dos teores de quartzo (estratos 2 e 3).
4.3. ESTUDOS QUÍMICOS
A análise de composição química detectou a presença de diversos
elementos do grupo dos metais e dos elementos terras raras (ETRs), os quais
são apresentados na íntegra no Apêndice I. Neste trabalho foram selecionados
os elementos químicos que ocorrem com teores mais significativos na amostra.
ELEMENTOS TRAÇO
Dentro do grupo dos metais os elementos que ocorrem com maiores
teores, em ppm, são o bário, rubídio, estrôncio e zircônio. Em geral, é evidente
uma variação composicional desses elementos da base para o topo do
estromatólito (Figura 16). Entretanto, elementos como o estrôncio e zircônio
apresentam uma alta variação de teores dentro dos estratos. O estrôncio é o
elemento cujos valores são os mais altos detectados, variando entre 1289 e
1848 ppm.
Os teores de bário formam uma curva ascendente da base para o topo
da amostra, enquanto que os valores do rubídio mostram uma curva
decrescente, portanto, a concentração desses dois metais é inversamente
proporcional ao longo dos estratos.
34
Figura 16: Elementos do grupo dos metais detectados ao longo da amostra de estromatólito pela análise de ICPS-MS.
ELEMENTOS TERRAS RARAS
Todos os ETRs da série dos lantanídios foram detectados pelo ICP-MS
com teores significativos em ppm, com exceção do promécio cujos valores
estão fora do limite de detecção.
Em geral os teores de ETRs apresentam alta variação ao longo dos
estratos (Figura 17). Os valores dos elementos lantânio, cério, praseodímio,
neodímio, samário e gadolínio, se mostram diretamente proporcionais,
apresentando semelhanças das curvas de concentração.
Apesar de um aumento do teor de európio no estrato seis, é notável um
padrão decrescente no valor desse elemento ao longo da amostra, assim como
para o itérbio, cujo gráfico se assemelha ao do európio.
Com exceção do lutécio, todos os demais ETRs mostraram um notável
decréscimo em seus teores no estrato três, seguidos por um ligeiro aumento
dos teores no estrato quatro e, posteriormente, uma nova diminuição desses
valores no estrato cinco, sugerindo uma mudança ambiental naquela porção.

35
Figura 17: Padrão de Elementos Terras Raras detectados por ICP-MS em cada um dos estratos da amostra de estromatólito (à esquerda). Na porção em destaque vermelho fica evidente uma diminuição dos valores dos ETRs no estrato três, com um aumento significativo dos teores no estrato imediatamente acima e, posteriormente, uma nova diminuição dos teores no estrato cinco.

36
4.4. ESTUDOS ISOTÓPICOS
A análise de isótopos estáveis também mostrou uma variação nas assinaturas
isotópicas do δ 18O e δ 13C ao longo da amostra. Os resultados da isotopia de 18O
e 13C estão apresentados na Tabela 1.
Tabela 1: Resultados da análise de Isótopos Estáveis de 13C e 18O.
Amostras δ ¹³C/¹²C
(‰ VPDB) δ ¹⁸O/¹⁶O
(‰ VPDB)
10 12,78 -1,65
9 12,85 -1,79
8 13,20 -1,80
7 18,15 -1,65
6 19,24 -0,88
5 15,65 -1,64
4 14,91 -1,19
3 14,61 0,14
2 12,11 0,75
1 2,42 0,92
Os valores de δ 13C variam bastante da base para o topo da amostra, indo de
2 até 12,78 ‰. Isso mostra um enriquecimento de isótopos pesados de carbono na
amostra como um todo. Somente no primeiro estrato, mais basal, ocorre um valor
muito baixo de δ 13C: 2,42‰. Já nos estratos sete e seis, ocorrem os maiores
valores isotópicos: 19,24 e 18,15 ‰.
Quanto aos isótopos de oxigênio é possível observar que há um decréscimo
nos valores de δ 18O da base para o topo da amostra, indo de 0,92 até -1,65 ‰. O
estromatólito, no geral, apresenta um enriquecimento de isótopos leves de oxigênio.
Porém, no estrato de amostragem seis ocorre um pico de aumento do valor de δ
18O, mesmo que não muito significativo.
5. DISCUSSÃO
Uma observação direta de estromatólitos vivos é essencial para a
interpretação de paleoambiente relacionados a distintos tipos de texturas

37
microbianas, além de inferir processos metabólicos associados aos ecossistemas
dinâmicos (Des Marais, 1990). Como forma de discutir os resultados obtidos neste
trabalho serão considerados os dados da descrição macroscópica integrados com
os resultados da análise química de ICP-MS e isótopos estáveis. Uma primeira
abordagem será dada à relação da trama do estromatólito com a sua composição
química, e em seguida, será discutido uma possível interpretação da evolução
paleoambiental da Lagoa Salgada, usando como base os dados geoquímicos.
Uma contribuição para a discussão será concedida a partir do trabalho de
Bahniuk (2013), como uma forma de comparação de resultados e possível
refinamento dos conhecimentos acerca dos estromatólitos da Lagoa Salgada (Figura
18).
Figura 18: Amostras de estromatólito utilizadas no trabalho de Bahniuk (2013) e no presente
trabalho, com definição das fácies (em branco).
5.1 FORMAÇÃO DAS TRAMAS DO ESTROMATÓLITO
A amostra de estromatólito analisada apresenta uma variação na composição
química e na trama ao longo das fácies, o que implica em possíveis variações
ambientais e das comunidades microbianas responsáveis pela formação deste
estromatólito.

38
A composição mineralógica encontrada no estromatólito está de acordo com
aquela esperada para sistemas de precipitação carbonática química e bioinduzida.
Entretanto, há ocorrência do mineral quartzo em toda a estrutura, o que deve
corresponder a uma contribuição sedimentar alóctone. Observou-se que os estratos
mais porosos da amostra, fácies trombólito e estromatólito vugular, são aqueles com
quantidades maiores do mineral de quartzo, concentrando-se na porção inferior do
estromatólito. É nessa porção mais basal que ocorre também o mineral dolomita,
somente nos três primeiros estratos. Já na porção superior, fácies estromatólito
digitado e trombolítico, predomina o mineral Mg-calcita, além da amostra apresentar
uma maior cimentação, com a ligeira diminuição de porosidade.
Resultados mineralógicos semelhantes foram relatados por Bahniuk (2013),
para amostra da mesma lagoa, contudo foi registrado a presença de calcita
altamente magnesiana, em vez de Mg-calcita. Os valores de quartzo do referido
trabalho são maiores na base do estromatólito, onde há ocorrência de dolomita,
enquanto que na porção superior diminui-se a quantidade quartzo e não ocorre a
dolomita, aumentando expressivamente o valor de calcita altamente magnesiana
(Tabela 2).
Tabela 2: Comparação dos resultados mineralógicos de Bahniuk (2013) com o presente trabalho. Em ambos os trabalhos é notável que o mineral dolomita ocorre somente na base do estromatólito (destaque em azul), onde há um expressivo valor de quartzo (destaque em amarelo), sendo a porção superior das amostras compostas predominantemente por calcita altamente magnesiana e Mg-calcita (destaque em verde).
Mineralogia (%)
Bahniuk (2013) Este trabalho
Estratos Calcita Alto Mg
Dolomita Quartzo Estratos Mg-
calcita Quartzo Dolomita
LS-6 81 - 13 10 90 10 -
LS-5 65 - 22 9 91 9 -
LS-4 21 - 7 8 75 25 -
LS-3 31 37 32 7 75 25 -
LS-2 37 19 23 6 84 16 -
LS-1 52 25 23 5 83 17 -
4 66 34 -
3 19,2 39,4 41,4
2 19 31 50
1 - 31 69

39
Segundo Bahniuk (2013), a presença de dolomita na base de estromatólito
indica processos biológicos mais ativos nessas fases de crescimento, ao passo que
a notável presença de Mg-calcita nos estratos mais acima indica que processos
abióticos devem ser considerados. A presença de tramas distintas de trombólitos e
estromatólitos na amostra permite sugerir um sintrofismo influenciado por mudanças
nas condições ambientais. Ainda segundo a autora, o mineral de calcita com alto
teor de Mg no topo da amostra sugere que a dolomita encontrada na base seja,
provavelmente, resultado do processo diagenético precoce, responsável por
disponibilizar íons de magnésio para a geração dessas.
Os microgastrópodes cujas conchas foram encontradas na fácies trombólito
não foram transportados e trapeados pelo estromatólito, mas viveram na estrutura
estromatolítica durante o desenvolvimento desta. Essa afirmação é evidenciada com
base na variação dos tamanhos das conchas encontradas, que representam
diferentes estágios de vida dos microgastrópodes.
Figueiredo-Barros et al. (2006) analisaram carapaças de H. australis da Lagoa
Imboassica, Rio de Janeiro, e encontraram tamanhos variando de 1,4 mm a 6,5 mm,
identificando um ciclo de vida mais curto devido as altas taxas de crescimento
promovidas pela elevada temperatura. Conde e Senra (2007), encontraram
tamanhos entre 0,3 mm e 5,3 mm para conchas de H. australis da Lagoa Salgada.
Visto que a presença das carapaças de microgastrópodes da espécie
Heleobia australis se deu de forma in situ, pode-se inferir que esses organismos
foram, provavelmente, responsáveis pela intensa bioerosão presente nessa amostra,
culminando com o aumento da porosidade daquele estrato, pois trata-se de
metazoários pastadores, que se alimentam diretamente de microalgas (Conde &
Senra, 2007).
Este pequeno gastrópode detrívoro é amplamente distribuído em ambientes
estuarinos, habitando fundos lamosos e com alto conteúdo de matéria orgânica
(Conde & Senra, 2007). Essa espécie é bem-sucedida na zona estuarina porque a
ação contínua da maré promove uma temperatura relativamente estável, sendo o
período reprodutivo e de crescimento influenciados por mudanças de salinidade e
exposição à maré, fatores que também afetam sua ecologia distribucional
(Francesco & Isla, 2004).

40
As carapaças dos microgastrópodes são os bioclastos carbonáticos mais
abundantes na Lagoa Salgada e apresentam intensa microbioerosão, gerada por
cianobactérias (Conde & Senra, 2007). A presença desses organismos na lagoa
está ligada à alta produtividade primária devido a produção de estromatólitos
recentes.
Enquanto que a presença de H. australis gerou a bioturbação bastante
pervasiva na fácies trombólito, estes organismos podem ter contribuído com a
bioconstrução da fácies seguinte, posto que as conchas de microgastrópodes
começam a ser dissolvidas logo após a morte dos moluscos (Hecth, 1933, apud
Morse, 2003). A atividade do íon carbonato diminui rapidamente à medida que
ocorre a entrada de CO2 no sistema, derivado da decomposição da matéria
orgânica, levando a uma diminuição no estado de saturação da água (Morse, 2003).
Os trabalhos de Hutchings (1986) e Garcia-Pichel et al. (2004) tem mostrado que
parte do carbonato de cálcio que é gerado em ambientes carbonáticos é derivado da
interação entre moluscos e cianobactérias, sugerindo que o mecanismo básico de
destruição de carbonatos marinhos seja a dissolução bioquímica ocasionada pela
atividade microbiana. Os microgastrópodes H. australis foram apontados por Senra
et al. (2006) como fonte de carbonato na Lagoa Salgada, uma vez que apresentam
microbioerosões causadas pelas cianobactérias e sulfobactérias formadoras das
esteiras microbianas presentes na lagoa.
Quanto a composição química das fácies, foi possível notar um alto teor de Sr
em todos os estratos, sendo os maiores valores encontrados na base do
estromatólito, assim como para os valores de Rb. Ambos os elementos compõem o
grupo dos oito solutos mais abundantes na água do mar (Railsback, 2003). O
elemento Sr, por sua vez, apresenta características químicas muito semelhantes ao
Ca, por isso sua distribuição em minerais de carbonato, como aragonita e calcita, é
bastante marcante devido a substituição catiônica existente entre os dois íons na
estrutura cristalina (ambos possuem raio iônico = 2Å), sobretudo em ambientes
marinhos (Baker et al., 1982). Além do Sr, o Mg é outro cátion que substitui o Ca nos
minerais carbonatados marinhos, segundo os mesmos autores. Bahiunk (2013)
encontrou valores de Sr para as fácies de estromatólito da Lagoa Salgada bastante
semelhantes às encontradas neste trabalho (Figura 19).

41
Figura 19: Comparação dos teores de Sr encontrados no estromatólito da Lagoa Salgada por
Bahniuk (2013) e neste trabalho.
O elemento Ba está no segundo grupo dos elementos mais abundantes na
água do mar (Railsback, 2003), pois assim como o Sr e o Rb, apresenta íons
grandes que formam fluoretos simples. Os cátions de Rb, Ba e Sr, são capazes de
se ligar fortemente com o oxigênio presente nos grupos carboxílicos orgânicos, por
isso são solúveis em solução aquosa, abundantes em águas naturais e capazes de
entrar em membranas celulares funcionando como nutrientes (Souza & Railsback,
2012). O Ba geralmente mostra uma gama incomumente ampla de concentrações
em comparação com intervalos para outros elementos, sendo que os teores de Ba
esperados para rochas carbonáticas são de 150 ppm, segundo Graf (1960) e de 20
a 200 ppm, de acordo com Krauskopf (1955, apud Graf, 1960).
A concentração de Zr varia bastante ao longo da amostra, sendo seu maior
teor no estrato cinco, correspondente à fácies estromatólito digitado. O elemento Zr

42
possui íons comumente enriquecidos em sedimentos residuais na crosta e em
nódulos de Fe-Mn em mar profundo (Railsback, 2003). Adams & Weaver (1958)
analisaram o teor de Zr em calcários e dolomitos e chegaram a valores entre 20 e
340 ppm, cuja média é de 145 ppm. Todo o conteúdo de Zr das rochas carbonáticas
é geralmente encontrado em resíduos insolúveis (Graf, 1960), enquanto que a
quantidade de Zr nos organismos marinhos é relacionada à quantidade de Fe que
eles contêm (Degenhardt, 1957 apud, Graf, 1960). Mesmo que os teores detectados
no estromatólito sejam bastante expressivos, variando de 25 a 75 ppm, um maior
número de dados é necessário para afirmar uma possível fonte desse elemento.
Contudo, não deve ser descartado a possível relação com uma entrada fluvial que
tenha carreado o Zr para o sistema.
Além dos processos citados acima, os elevadores teores de elementos
encontrados na amostra estão provavelmente relacionados a presença dos
microrganismos formadores do estromatólito, uma vez que de acordo com Graf
(1960) organismos vivos tem uma importância potencial em contribuir com
concentração de elementos menores para rochas e sedimentos carbonáticos.
Compreender a absorção de elementos traços por microbialitos é
especialmente importante no que diz respeito aos elementos terras raras, uma vez
que os carbonatos microbianos podem fornecer proxies para avaliar a química da
água de paleomares (Webb & Kamber, 2000). Os mesmos autores relataram que
especificamente os microbialitos holocênicos da Grande Barreira de Corais,
Austrália, parecem incorporar ETRs da água do mar a concentrações
substancialmente mais altas do que outros precipitados biogeoquímicos comumente
empregados, e com pouco fracionamento através da série de ETR.
No presente trabalho foram detectados teores significativos desses elementos
ao longo de toda a amostra, em concordância com a hipótese de que a comunidade
microbiana presente no estromatólito seja responsável pela captação dos íons de
ETRs. Entretanto, os ETRs com teores mais elevados são aqueles do grupo do
cério, ETRs leves, em relação aos elementos do grupo do ítrio, ETRs pesados
(Figura 20).

43
Figura 20: Diagrama de teores de ETRs na amostra de estromatólito da Lagoa Salgada. Os ETRs
leves estão destacados em verde, enquanto que os ETRs pesados em vermelho.
Johannesson et al. (2014) analisaram os teores de ETRs em estromatólitos
recentes de Cuatro Ciénegas (México) e verificaram que os maiores teores
detectados eram de ETRs pesados. Takahashi et al. (2005) demonstraram que os
grupos funcionais de superfície nas paredes celulares bacterianas preferencialmente
absorvem os ETRs pesados em relação aos ETRs leves, pois estas superfícies
provavelmente excedem os relativamente fracos complexos de sulfato, bicarbonato
e cloreto dos ETRs, levando ao enriquecimento de ETRs pesados nos
estromatólitos. Deste modo, os altos teores de ETRs leves apresentados neste
trabalho sugerem uma possível absorção inorgânica desses elementos no
estromatólito.
5.2 EVOLUÇÃO PALEOAMBIENTAL

44
Martin et al. (1993) sugere que a formação da Lagoa Salgada se deu devido à
subida de nível relativo do mar, após os deslocamentos da barreira arenosa
holocênica da planície litorânea do Rio Paraíba do Sul, rumo ao continente, nas
condições hidrodinâmicas geradas por ondas. Tal deslocamento da barreira arenosa
só poderia ser resultado da última elevação de nível do mar entre 3600 - 2800 anos
B.P.
Ao longo dos últimos 2300 anos diferentes eventos de mudanças físico-
químicas, metabólicas e sedimentares ocorreram na Lagoa Salgada, de forma que
tais registros se preservaram nas texturas, estruturas e fácies dos estromatólitos
(Bahniuk, 2013). A análise isotópica mostrou uma variação ao longo da amostra
para os valores de isótopos de carbono e oxigênio. Da base para o topo há um
aumento da razão de δ ¹³C, cuja variação é de 2,42 até 19,24 ‰, enquanto que para
a razão de δ 18O há uma variação de -1,80 até 0,92 ‰ (Figura 21), valores muito
próximos dos encontrados por Bahniuk (2013).
Figura 21: Comparação dos dados da análise de isótopos estáveis de C & O, para este trabalho e o
de Bahniuk (2013).

45
Os três estratos mais basais apresentam valores de δ 18O muito próximo
àqueles esperados para o padrão da água do mar, 0 ‰, assumindo que o carbonato
preserva os sinais isotópicos estáveis originais (Bahniuk, 2013), infere-se que as
composições isotópicas de ¹⁸O da porção inferior do estromatólito (0,92, 0,14 e 0,75
‰) correspondem a uma configuração marinha. Uma variação crescente dos valores
em direção ao topo da amostra revela uma transição para um sistema lagunar mais
fechado (Bahniuk, 2013), uma vez que os valores de ¹⁸O se tornam mais negativos (-
0,88 até -1,80), inferindo-se uma influência da água doce.
As assinaturas isotópicas do ¹³C para a Lagoa Salgada indicam variações no
metabolismo microbiano específico envolvido na precipitação de carbonatos,
podendo assim sugerir informações sobre a evolução deste sistema (Bahniuk,
2013). A variação dos valores de ¹³C ao longo da amostra sugere uma diversificação
de microrganismos atuantes de acordo com as novas condições do ambiente. Na no
estrato mais basal, fácies esteira microbiana, o valor de ¹³C é de 2,42, o menor
registrado, sendo consistente com a precipitação de carbonato de águas marinhas
não restritas, cuja composição isotópica não é afetada pelo metabolismo microbiano.
As demais fácies apresentam valores fortemente enriquecidos ¹³C, podendo-
se interpretar como resultado de processos de precipitação mineral influenciados
pela metanogênese microbiana. De acordo com Bahniuk (2013), os altos valores de
¹³C encontrados indicam que o metano produzido pelo metabolismo microbiano é
liberado, em vez de ser utilizado pela atividade metanotrófica e, como consequência,
deixa uma assinatura isotópica de ¹³C pesada que é incorporada na fase
carbonatada. A metanogênese é energeticamente o modo menos favorável de
respiração, porém, se o suprimento de água do mar diminuir, os metabolismos da
metanogênese predominam no ambiente. Esta afirmação apoiou a hipótese do
trabalho de Bahniuk (2013) de que a Lagoa Salgada sofreu considerável variação na
fonte de água durante o crescimento dos estromatólitos.
Levando em consideração todos os fatores aqui levantados: a presença de
minerais com valores elevados de Mg; o alto teor de Sr na base da estrutura
estromatolítica; e as razões isotópicas próximas do padrão da água do mar, pode-se
inferir uma influência marinha no ambiente de formação desses. Entretanto, a partir
da segunda fácies mais basal, trombólito, tem-se a ocorrência das valvas dos
microgastópodes pastadores, os quais são restritos a ambientes lacustrinos,

46
sugerindo que não se tratava completamente de um sistema marinho aberto, mas
sim transicional, já durante a formação da base do estromatólito. Portanto, como
forma de refinar o modelo de evolução da lagoa proposto por Bahniuk (2013),
sugere-se que a ligação com o mar aberto, registrado nos estromatólitos da Lagoa
Salgada, foi interrompida antes do que se propôs, evoluindo para um ambiente
transicional com contribuição fluvial continental.
Bahniuk (2013) sugere a ligação com o mar aberto entre 2222 e 1982 anos
B.P. Entretanto a presença de microgastrópodes na fácies trombólito definida neste
trabalho, sugere um início anterior para a evolução do ambiente transicional. Este
sistema sem a influência total de mar aberto, mas também contribuição fluvial e
desenvolvimento de um ambiente estuarino pode ter surgido por volta de 2306 anos
B.P (Figura 22).
Figura 22: Amostras usados no trabalho de Bahniuk (2013) e neste trabalho, mostrando as fácies e
seus possíveis ambientes de formação associados.
6. CONCLUSÕES
O estromatólito da Lagoa Salgada analisado foi dividido em cinco fácies, da
base para o topo: esteira microbiana, trombólito, estromatólito vugular,
estromatólito digitado e estromatólito trombolítico.

47
As diferentes fácies apresentam diferentes tramas e composições químicas,
as quais sugerem diferentes fases de evolução da lagoa.
A presença de diferentes tamanhos nas valvas dos microgastrópodes da
espécie Heleobia australis, representando diferentes estágios de vida, revela
que esses moluscos habitavam aquele ecossistema, sugerindo uma predação
dos microrganismos formadores do estromatólito.
A amostra estromatolítica apresentou uma variação na composição química e
mineralógica ao longo das fácies. Na porção basal tem-se a predominância
de minerais de quartzo e dolomita, enquanto que no topo prevalece a Mg-
calcita com pouco quartzo, e não ocorre dolomita.
A presença de dolomita na base de estromatólito implica processos biológicos
mais ativos, entretanto pode também ser resultado do processo diagenético
precoce, responsável por disponibilizar íons de magnésio da calcita com alto
teor de Mg no topo da amostra.
Teores elevados de elementos traços, tais como Rb, Sr, Zr, Ba e ETRs foram
encontrados ao longo da amostra, estando provavelmente relacionados à
presença dos microrganismos formadores do estromatólito, os quais
contribuem para a captação de íons e concentração de elementos menores
em sedimentos e rochas carbonáticas.
Os teores elevados de Sr nas camadas 2/3 e 7/8 são possíveis indicadores
de influência de águas marinhas, ao passo que Ba elevado no topo provém
de fontes fluviais.
As fácies do estromatólito estão enriquecidas de δ 13C, podendo-se
interpretar como resultado de processos de precipitação mineral influenciados
pela metanogênese microbiana, ao longo da formação dessa bioestrutura.
Ressalta-se que as bactérias utilizam o δ 12C o tempo todo na fotossíntese e
outros processos, por isso o enriquecimento da água e depósitos
carbonáticos em δ 13C, logo, não é somente metanogênese que pode ser
considerado.
Em lagunas e estuários próximos ao mar a presença de musgos e outros
produtos atrai organismos como gastrópodes pastadores.
Os sinais isotópicos de δ 18O na base do estromatólito, mostra uma
influência marinha nesse sistema, todavia a presença de microgastrópodes

48
pastadores no estrato dois indica um ambiente com influência fluvial ou uma
restrição ao mar aberto. Sugere-se, então, uma fase de deposição em um
contexto transicional já na base do estromatólito, durante a formação da
fácies trombólito, seguindo até a fácies estromatólito vugular a partir da qual,
houve a instalação de um ambiente lacustre restrito até o estágio final de
formação da amostra.

49
REFERÊNCIAS
Adams, J. A., & Weaver, C. E. Thorium-to-uranium ratios as indicators of sedimentary processes: example of concept of geochemical facies. AAPG Bulletin, 42(2), 387-430. 1958.
Aitken, J.D. Classification and environmental significance of cryptalgal limestones and dolomites with illustrations from the Cambrian and Ordovician of southwestern Alberta. Journal of Sedimentary Petrology 37, 1163–1178. 1967.
Awramik S.M. The History and Significance of Stromatolites. In: Schidlowski M., Golubic S., Kimberley M.M., McKirdy D.M., Trudinger P.A. (Eds) Early Organic Evolution. Springer, Berlin, Heidelberg, p. 435-449. 1992.
Awraniik, S.M. & Margulis, L. Strornatolitc Newsletter 2, 5. 1974.
Bahniuk, A.M. Coupling organic and inorganic methods to study growth and diagenesis of modern microbial carbonates, Rio de Janeiro State, Brazil: implications for interpreting ancient microbialite facies development. Tese de Doutorado. ETHZ, Zurich. 169 p. 2013.
Baker, P. A., Gieskes, J. M., & Elderfield, H. Diagenesis of carbonates in deep-sea sediments; evidence from Sr/Ca ratios and interstitial dissolved Sr (super 2+) data. Journal of Sedimentary Research, 52(1), 71-82. 1982.
Barbosa, J.A.; Fambrini, G.L.; Carvalho, R.R. Microfácies carbonáticas e comportamento isotópico de C e O nos calcários laminados aptianos lacustres da Serra Negra, Bacia do Jatobá, Nordeste do Brasil. Estudos Geológicos v. 20 (1), 2010.
Birgel, D., Meister, P., Lundberg, R., Horath, T.D., Bontognali, T.R.R, Bahniuk, A.M., Rezende, C.E., Vasconcelos, C., Mckenzie, J.A. Methanogenesis produces strong 13C enrichment in stromatolites of Lagoa Salgada, Brazil: a modern analogue for Palaeo-Neoproterozoic stromatolites? Geobiology, 13:245–266. 2015.
Brehm, U.; Palinska, K.A.; Krumbein, W.E. Laboratory cultures of calcifying biomicrospheres generate ooids – A contribution to the origin of oolites. Carnets de Géologie/Notebooks on Geology, Maintenon, Letter v. 3, p. 6, 2004.
Bureau Veritas Minerals. Schedule of Services & Fees 2018. USD. 48p. 2018.
Burne, R.V., Moore, L.S. Microbialites: organosedimentary deposits of benthic microbial communities. Palaios, 2:241-254. 1987.
Burns, B. P., Goh, F., Allen, M., & Neilan, B. A. Microbial diversity of extant stromatolites in the hypersaline marine environment of Shark Bay, Australia. Environmental Microbiology, 6(10), 1096-1101. 2004.
Callefo, F. A utilização de biominerais como possíveis bioassinaturas microbianas. Tese de Doutorado. Instituto de Geociências da Universidade Estadual de Campinas, Campinas, 145 p. 2018.

50
Cataldo, R.A. Análise dos Estromatólitos e sedimentos associados – Lagoa Salgada/RJ. Trabalho de Conclusão de Curso. Universidade Estadual de Campinas, Instituto de Geociências, Campinas, 61 p. 2011.
Choquette, P. W., & Pray, L. C. Geologic nomenclature and classification of porosity in sedimentary carbonates. AAPG bulletin, 54(2), 207-250. 1970.
Collins, L.B., Zhao, J.-X., Freeman, H. A high-precision record of mid-late Holocene sea-level events from emergent coral pavements in the Houtman Abrolhos Islands, southwest Australia. Quaternary International 145–146, 78–85. 2006.
Commons Wikimedia. Stromatolites in limestone. Disponível em <https://bit.ly/2OOh5KA>. Acessado em 16/11/2018.
Conde, J. N., & Senra, M. C. E. Marcas de predação em microgastrópodos da Lagoa Salgada, RJ, Brasil. Anuário do Instituto de Geociências, 30(1), 226-226. 2007.
Damazio, C.M. Tipificação e bioestratificação cianobacteriana das esteiras microbianas da borda noroeste da lagoa Pitanguinha, Holoceno do Rio de Janeiro, Brasil. Monografia (Bacharelado em Ciências Biológicas) – Universidade Federal do Estado do Rio de Janeiro. Rio de Janeiro. 171 p. 2004.
De Francesco, C. G., & Isla, F. I. The life cycle and growth of Heleobia australis (D'Orbigny, 1835) and H. conexa (Gaillard, 1974)(Gastropoda: Rissooidea) in Mar Chiquita coastal lagoon (Argentina). Journal of Molluscan Studies, 70(2), 173-178. 2004.
Des Marais, D.J. Microbial mats and the early evolution of life. Trends in ecology & evolution, v. 5, n. 5, p. 140-144, 1990.
Elderfield, Henry. The oceanic chemistry of the rare-earth elements. Phil. Trans. R. Soc. Lond. A, v. 325, n. 1583, p. 105-126, 1988.
Figueiredo-Barros, M. P., Leal, J. J., Esteves, F. D. A., Rocha, A. D. M., & Bozelli, R. L. Life cycle, secondary production and nutrient stock in Heleobia australis (d'Orbigny 1835)(Gastropoda: Hydrobiidae) in a tropical coastal lagoon. Estuarine, Coastal and Shelf Science, 69(1-2), 87-95. 2006.
Fiori, S. M., & Carcedo, M. C. Estado actual del conocimiento sobre Heleobia australis y perspectivas futuras. In: Amici Molluscarum. Número especial, año 2011. Sociedad Malacológica de Chile. 2011.
Fossil Purchase Information. Broadly Branched Digitate Stromatolites from Ordovician Wisconsin. Disponível em <https://bit.ly/2Dohf9E>. Acessado em 16/11/2018.
Garcia‐Pichel, F., Al‐Horani, F. A., Farmer, J. D., Ludwig, R., & Wade, B. D. Balance between microbial calcification and metazoan bioerosion in modern stromatolitic oncolites. Geobiology, 2(1), 49-57. 2004.

51
Gerdes, G., Claes, M., Dunajtschik-Piewak, K., Riege, H., Krumbein, W. E., & Reineck, H. E. Contribution of microbial mats to sedimentary surface structures. Facies, 29(1), 61. 1993.
Gomes, Celso B.; Formoso, M. L. L.; Trescases, J J; Dutra, C V; Gomes, Celso B. Técnicas analíticas instrumentais aplicadas à geologia. [S.l: s.n.], 1984.
Gomes, S. Composição Geomicrobiológica e Orgânica das Esteiras Microbianas de Três Lagoas Costeiras do Sudeste do Rio de Janeiro, Brasil. Diss. Tese de doutoramento, Universidade Federal do Rio de Janeiro, 190 (não publicada), 2011.
Graf, D.L. Geochemistry of Carbonate Sediments and Sedimentary Carbonate Rocks. Part III Minor Element Distribution. Illinois State Geological Survey Circular 301. 71 p., 3 figs., 38 tables. 1960.
Grotzinger, John; Jordan, Tom. Para entender a Terra. 6. ed Porto Alegre: Bookman. 738 p. ISBN 9788565837774 (broch.). 2013.
Hayashi, F.Y.D. Avaliação do Volume Representativo Elementar (REV) em estromatólito de Lagoa Salgada. Trabalhao Final de Curso (Bacharelado em Geologia) - Universidade Federal do Rio de Janeiro. Rio de Janeiro. 90p. 2014.
Hofmann, H. J., Grey, K., Hickman, A. H., & Thorpe, R. I. Origin of 3.45 Ga coniform stromatolites in Warrawoona group, Western Australia. Geological Society of America Bulletin, 111(8), 1256-1262. 1999.
Hofmann, H.J. Stromatolites from the Proterozoic Animikie and Sibley Groups. Geol. Surv. Can. Pap., 68-69:77 pp. 1969.
Hofmann, H.J. Stromatolites: characteristics and utility. Earth-Sci Rev. 9:339-373. 1973.
Houk, R.S.; Jarvis, K.E.; Gray, A.L. Handbook of Inductively Coupled Plasma Mass Spectrometry, Chapman and Hall, New York: 1992.
Hutchings, P. A. Biological destruction of coral reefs. Coral reefs, 4(4), 239-252. 1986.
Iespa, A.A.C. O Plexo Estromatólito–Trombólito–Oncoide, Lagoa Salgada, RJ, Brasil. Rio de Janeiro. Tese (Doutorado em Geologia) – Universidade Federal do Rio de Janeiro. Rio de Janeiro. 105 p. 2010.
Iespa, A.A.C., Iespa, C.M.D., Borghi, L. Evolucão paleoambiental da lagoa salgada utilizando microbialitos, com ênfase em microfácies carbonáticas. Geociências, São Paulo, UNESP, v. 31, 3:71-380. 2012.
Iespa, A.A.C., Iespa, C.M.D., Borghi, L. Microestratigrafia do complexo estromatólito, trombólito e oncoide holocênico da Lagoa Salgada, Estado do Rio de Janeiro, Brasil. Revista de Geologia, 22:7-14. 2008.
Jahnert, R. J., & Collins, L. B. Significance of subtidal microbial deposits in Shark Bay, Australia. Marine Geology, 286(1-4), 106-111. 2011.

52
Johannesson, K. H., Telfeyan, K., Chevis, D. A., Rosenheim, B. E., & Leybourne, M. I. Rare earth elements in stromatolites—1. Evidence that modern terrestrial stromatolites fractionate rare earth elements during incorporation from ambient waters. In Evolution of Archean Crust and Early Life (pp. 385-411). Springer, Dordrecht. 2014
Lemos,R.M.T.; Silva,C.G.; Spadini,A.R. Estratigrafia e estromatólitos recentes da Lagoa Salgada - RJ. In: Congresso Brasileiro de Geologia, 38, Camboriú/SC,1994, SBG. Anais,3: 258- 260. 1994.
Lith, Y.van; Vasconcelos, C.; Warthmann, R.; Martines, J.C.F. & Mckenzie, J.A. Bacterial sulfate reduction and salinity: two controls on dolomite precipitation in Lagoa Vermelha and Brejo do Espinho (Brazil). Hydrobiologia, 485: 35-49. 2002.
Logan, B. W., Rezak, R., & Ginsburg, R. N. Classification and environmental significance of algal stromatolites. The Journal of Geology, 72(1), 68-83. 1964.
Martin, L., Suguio, K., & Flexor, J. M. As flutuações de nível do mar durante o quaternário superior e a evolução geológica de" deltas" brasileiros. Boletim IG-USP. Publicação Especial, (15), 01-186. 1993.
McNamara, K.J., Awramik, S.M. Stromatolites: a key to understanding the early evolution of life. Science Progress, 76:345-364. 1992.
Morse, J. W. Formation and diagenesis of carbonate sediments. Treatise on Geochemistry, 7, 407. 2003.
Neumann, V. H., Rocha, D.E.G.A., Moraes, A. S., Sial, A. N., Castro, M. T., Barbosa, J. A., Fambrini, G.L. & Carvalho, R. R. Microfácies Carbonáticas e Comportamento Isotópico de CeO nos Calcários Laminados Aptianos Lacustres da Serra Negra, Bacia do Jatobá, Nordeste do Brasil. Estudos Geológicos, 20(1), 89. 2010.
Pacheco, R.C. Avaliação Da Interação Físico-Química De Componentes De Um Petróleo Com Estromatólitos Da Lagoa Salgada, RJ. Dissertação de Mestrado. Centro de Ciência e Tecnologia da Universidade Estadual do Norte Fluminense. Universidade Estadual do Norte Fluminense Darcy Ribeiro – UENF. 111 p. 2014.
Pereira, L. D. D. O. Fácies sedimentares e evolução paleoambiental da Lagoa Salgada (Litoral norte do Rio de Janeiro). Trabalho de Conclusão de Curso. Instituto de Geociências, Universidade Federal do Rio de Janeiro – UFRJ. 87p. 2014.
Railsback, L. B. An earth scientist's periodic table of the elements and their ions. Geology, 31(9), 737-740. 2003.
Reid, P.R., James, N.P., Macintyre, I.G., Dupraz, C.P., Burne, R.V. Shark Bay stromatolites: microfabrics and reinterpretation of origins. Facies 49, 299–324. 2003.
Rezende, M.F., Tonietto, S.N. and Pope, M.C. Three-dimensional pore connectivity evaluation in a Holocene and Jurassic microbialite buildupThree-Dimensional Pore Connectivity Evaluation in Microbialite Buildup. AAPG bulletin, 97(11), pp.2085-2101. 2013.

53
Riding, R. Classification of microbial carbonates. In: Riding, R. (Ed.) Calcareous Algae and Stromatolites. Springer-Verlag, New York. p. 21-51. 1991.
Riding, R. Microbialites, stromatolites, and thrombolites. In: Reitner, J., Thiel, V. (Eds.) Encyclopedia of Geobiology. Encyclopedia of Earth Science Series, Springer, Heidelberg, 635-654. 2011.
Riding, R. The term stromatolite: towards an essential definition. Lethaia, 32(4), 321-330. 1999.
Senra, M. C. E., da Silva, L. H., Conde, J. N., & Iespa, A. A. C. Microbioerosão em conchas de Heleobia australis (Gastropoda: Rissooidea) da lagoa Salgada, Rio de Janeiro, Brasil. Anuário do Instituto de Geociências, 29(2), 199-209. 2006.
Servidoni, A.P.S. Análise de porosidade e o potencial como rocha reservatório em estromatólito do recente da lagoa salgada, litoral fluminense – RJ. Dissertação de Mestrado. Programa de Pós-Graduação em Geologia, Setor de Ciências da Terra, Universidade Federal do Paraná. 142 p. 2012.
Silva e Silva L.H., Iespa A.A.C., Damazio-Iespa C. M. Composição dos Estromatólitos Estratiformes da Lagoa Salgada, Rio de Janeiro, Brasil. Anuário do Instituto de Geociências, 31(2), 42-49. 2008.
Silva e Silva L.H., Iespa A.A.C., Damazio-Iespa C. M., Carvalhal S.B.V., Alves S.A.P. Confronto entre estruturas estromatolíticas domais (composição cianobacteriana) das lagoas Pernambuco e Salgada, Brasil. Revista de Geologia, Fortaleza, 18(2), 159-165. 2005.
Silva e Silva, L.H. Contribuição ao conhecimento da composição microbiana e química das estruturas estromatolíticas da Lagoa Salgada, Quaternário do Rio de Janeiro, Brasil. Programa de Pós-graduação em Geologia, Universidade Federal do Rio de Janeiro, Tese de Doutorado, 176p. 2002.
Silva e Silva, L.H.; Alves, S.A.P.M.N., Magina, F.C., Gomes, S.B.V.C. Composição cianobacteriana e química dos estromatólitos da Lagoa Salgada, Neógeno do estado do Rio de Janeiro, Brasil. Geol. USP, Sér. Cient. São Paulo, v. 13, 1:9-106. 2013.
Silva e Silva, L.H.; Iespa, A.A.C., Iespa, C.M.D. Considerações sobre Estromatólito do Tipo Domal da Lagoa Salgada, Estado do Rio de Janeiro, Brasil. Anuário do Instituto de Geociências - UFRJ, Vol. 30, 1:50-57. 2007.
Silva e Silva, L.H.; Senra, M.C.E.; Faroulo, T.C.L.M.; Carvalhal, S.B.V.; Alves, S.A.P.M.N.; Damazio, C.M.; Shimizu, V.T.A.; Santos, R.C. & Iespa, A.A.C. Composição paleobiológica e tipos morfológicos das construções estromatolíticas da Lagoa Vermelha, RJ, Brasil. Revista Brasileira de Paleontologia, 7(2), 193-198. 2004.
Silva, D.R., Mansur, K.L. & Borghi, L. Evaluation of the scientific value of Lagoa Salgada (Rio de Janeiro, Brazil): characterization as geological heritage, threats and strategies for geoconservation. Journal of the Geological Survey of Brazil vol 1, no 2, 69-80, August, 2018.

54
Souza, R. F., & Railsback, L. B. (2012). Uma tabela periódica dos elementos e seus íons para cientistas da terra. Terrae Didática, 8(2), 73-82. 2012.
Srivastava, N.K. Lagoa Salgada, RJ - Estromatólitos recentes. In: Schobbenhaus, C., Campos, D.A., Queiroz, E.T., Winge, M., Berbert-Born, M.L.C (Eds.) Sítios Geológicos e Paleontológicos do Brasil. DNPM/CPRM - Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP) – SIGEP 041.1999.
Takahashi, Y., Châtellier, X., Hattori, K. H., Kato, K., & Fortin, D. Adsorption of rare earth elements onto bacterial cell walls and its implication for REE sorption onto natural microbial mats. Chemical Geology, 219(1-4), 53-67. 2005.
Vasconcelos, C.; Warthmann, R.; McKenzie, J.A.; Visscher, P.T.; Bittermann, A.G.; Lith, Y. Lithifying microbial mats in Lagoa Vermelha, Brazil: Modern Precambrian relics? Sedimentary Geology. 185p. 175–183. 2006.
Veiga, M.A.M.S. Determinação de elementos traço por AAS e ICP-MS após extração no ponto nuvem, usando dietilditiofosfato como agente quelante. (Tese de Doutorado) Programa de Pós-Graduação em Química da Universidade Federal de Santa Catarina, 2000.
Webb G.E. & Kamber B.S. Rare earth elements in Holocene reefal microbialites: a new shallow water proxy. Geochim Cosmochim Acta 64:1557–1565. 2000.

La Ce Pr Nd Sm Eu Gd Tb Dy Ho Er Tm Yb Lu
Amostras
245/18-10 3,5 6,9 71 2,5 36 2 29 3 10 <0.02 5 <0.01 <0.05 <0.01
245/18-09 1,8 3,6 35 1,5 23 3 21 2 12 2 7 <0.01 8 2
245/18-08 1,3 2,1 22 0,9 13 2 15 2 12 <0.02 5 <0.01 7 1
245/18-07 2,2 4,1 43 1,6 30 3 18 3 18 3 11 1 9 1
245/18-06 3,2 5,6 59 2,3 34 6 43 5 24 5 11 2 14 2
245/18-05 5,5 5,3 46 1,6 27 4 26 4 18 4 12 2 11 2
245/18-04 7,6 14,5 141 5,5 96 5 80 9 42 6 13 2 10 1
245/18-03 2 5,5 37 1,3 18 4 15 3 19 3 7 1 9 2
245/18-02 4,7 12,1 98 4,1 61 6 48 6 32 6 17 2 14 2
245/18-01 3,8 10,3 77 2,6 37 6 34 4 22 4 13 2 12 2
Ba Rb Sr Zr
Amostras
245/18-10 182 0,2 1711,1 19,2
245/18-09 192 0,9 1343,9 63,1
245/18-08 131 0,9 1289,7 32,2
245/18-07 124 0,8 1727,2 42
245/18-06 122 1,4 1848,2 60,8
245/18-05 77 1,3 1523,1 75,3
245/18-04 82 2,2 1591,6 55
245/18-03 88 2,6 1789 29,7
245/18-02 73 3,1 1817,3 68,6
245/18-01 66 3,1 1490,7 46,3
54
ppm
Metais
ppm
ETRs
APÊNDICE I - Tabela de Dados Químicos a partir da Análise de ICP-MS





























